ORIGINAL RESEARCH ARTICLE Determination of the status of the etiological agent of American foulbrood, Paenibacillus larvae, in Swaziland Dermot Cassidy 1* , Teresa Goszczynska 2 , John Burnet 3 , Ulrike Hirschauer 4 , Solomon Gebeyehu 1 , Gudeta W Sileshi 5 and Lise Korsten 6 1 USAID, P.O. Box 11218, Silver Lakes, Pretoria 0054, South Africa. 2 Agricultural Research Council, Plant Protection Research Institute, Private Bag X 134, 0121 Queenswood, Pretoria, South Africa. 3 Eswatini Swazi Kitchen Honey, P.O. Box 1137, Manzini, Swaziland. 4 TechnoServe, Inc., P.O. Box 663, Ezulwini, Swaziland. 5 World Agroforestry Centre (ICRAF), SADC-ICRAF Agroforestry Programme, Chitedze Agricultural Research Station, P.O. Box 30798, Lilongwe, Malawi. 6 Department of Microbiology and Plant Pathology, University of Pretoria, 0002, Pretoria, South Africa. Received 28 January 2010, accepted subject to revision 1 June 2011, accepted for publication 7 July 2011. *Corresponding author: Email: [email protected] Summary American foulbrood (AFB) is a cosmopolitan disease affecting both larval and pupal stages of honey bees. There are considerable doubts about the true status of AFB in Africa and there is, indeed, some evidence that sub-Saharan Africa, until recently, was largely free of AFB. Requirements for honey imports into South Africa are governed by a concern for the potential introduction of AFB. The study describes a cost effective and simple methodology for science-based trade in honey from Swaziland into South Africa that complies with the guidelines developed by the World Organisation for Animal Health (OIE). Determinación del estado del agente etiológico de la loque americana, Paenibacillus larvae, en Suazilandia Resumen La loque americana (LA) es una enfermedad cosmopolita que afecta a los estadios de larva y pupa de la abeja de la miel. Hay dudas considerables acerca del estado real de la LA en África y hay, de hecho, alguna evidencia de que África sub-sahariana estaba, hasta hace poco, libre de LA. Los requisitos para la importación de miel a Sudáfrica están regidos por una cierta preocupación por la potencial introducción de LA. Este estudio describe una metodología simple y efectiva para el comercio basado en la ciencia de miel desde Suazilandia hasta Sudáfrica que cumple con los preceptos desarrollados por la Organización de Salud Animal. Keywords: American foulbrood, honey, survey design, market access, sub-Saharan Africa Journal of Apicultural Research 50(4): 284-291 (2011) © IBRA 2011 DOI 10.3896/IBRA.1.50.4.05 Introduction American foulbrood (AFB), caused by the Gram positive sporulating bacterium Paenibacillus larvae, is an infectious, highly contagious, cosmopolitan disease affecting the larval and pupal stages of the honey bee Apis mellifera and other Apis spp., occurring in most countries where bees are kept (World Organization for Animal Health, 2009). Taxonomic studies of the two subspecies of Paenibacillus larvae, P. larvae subsp. larvae and P. larvae subsp. pulvifaciens, have led to the reclassification of these subspecies into a single species, P. larvae (Kilwinski et al., 2004; Genersch et al., 2006; OIE, 2009). In this paper the designation AFB specifically refers to the ERIC I (aβ, ab, and Ab) genotypes of Paenibacillus larvae and excludes ERIC II (AB), III and IV genotypes (Kilwinski et al., 2004; Genersch et al., 2006; OIE, 2009). From a scientific and phytosanitary perspective there are doubts about the true status of AFB in Africa, and there is some evidence that sub-Saharan Africa, until recently, was considered largely AFB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL RESEARCH ARTICLE Determination of the status of the etiological agent of
American foulbrood, Paenibacillus larvae, in Swaziland
Dermot Cassidy1*, Teresa Goszczynska2, John Burnet3, Ulrike Hirschauer4, Solomon Gebeyehu1, Gudeta W Sileshi5 and Lise Korsten6 1USAID, P.O. Box 11218, Silver Lakes, Pretoria 0054, South Africa. 2Agricultural Research Council, Plant Protection Research Institute, Private Bag X 134, 0121 Queenswood, Pretoria, South Africa. 3Eswatini Swazi Kitchen Honey, P.O. Box 1137, Manzini, Swaziland. 4TechnoServe, Inc., P.O. Box 663, Ezulwini, Swaziland. 5World Agroforestry Centre (ICRAF), SADC-ICRAF Agroforestry Programme, Chitedze Agricultural Research Station, P.O. Box 30798, Lilongwe, Malawi. 6Department of Microbiology and Plant Pathology, University of Pretoria, 0002, Pretoria, South Africa. Received 28 January 2010, accepted subject to revision 1 June 2011, accepted for publication 7 July 2011. *Corresponding author: Email: [email protected]
Summary American foulbrood (AFB) is a cosmopolitan disease affecting both larval and pupal stages of honey bees. There are considerable doubts
about the true status of AFB in Africa and there is, indeed, some evidence that sub-Saharan Africa, until recently, was largely free of AFB.
Requirements for honey imports into South Africa are governed by a concern for the potential introduction of AFB. The study describes a cost
effective and simple methodology for science-based trade in honey from Swaziland into South Africa that complies with the guidelines
developed by the World Organisation for Animal Health (OIE).
Determinación del estado del agente etiológico de la loque
americana, Paenibacillus larvae, en Suazilandia Resumen
La loque americana (LA) es una enfermedad cosmopolita que afecta a los estadios de larva y pupa de la abeja de la miel. Hay dudas
considerables acerca del estado real de la LA en África y hay, de hecho, alguna evidencia de que África sub-sahariana estaba, hasta hace
poco, libre de LA. Los requisitos para la importación de miel a Sudáfrica están regidos por una cierta preocupación por la potencial
introducción de LA. Este estudio describe una metodología simple y efectiva para el comercio basado en la ciencia de miel desde Suazilandia
hasta Sudáfrica que cumple con los preceptos desarrollados por la Organización de Salud Animal.
Keywords: American foulbrood, honey, survey design, market access, sub-Saharan Africa
Journal of Apicultural Research 50(4): 284-291 (2011) © IBRA 2011 DOI 10.3896/IBRA.1.50.4.05
Introduction
American foulbrood (AFB), caused by the Gram positive sporulating
bacterium Paenibacillus larvae, is an infectious, highly contagious,
cosmopolitan disease affecting the larval and pupal stages of the
honey bee Apis mellifera and other Apis spp., occurring in most
countries where bees are kept (World Organization for Animal Health,
2009). Taxonomic studies of the two subspecies of Paenibacillus
larvae, P. larvae subsp. larvae and P. larvae subsp. pulvifaciens, have
led to the reclassification of these subspecies into a single species, P.
larvae (Kilwinski et al., 2004; Genersch et al., 2006; OIE, 2009). In
this paper the designation AFB specifically refers to the ERIC I (aβ,
ab, and Ab) genotypes of Paenibacillus larvae and excludes ERIC II
(AB), III and IV genotypes (Kilwinski et al., 2004; Genersch et al.,
2006; OIE, 2009).
From a scientific and phytosanitary perspective there are doubts
about the true status of AFB in Africa, and there is some evidence
that sub-Saharan Africa, until recently, was considered largely AFB

free (Matheson, 1996; Fries and Raina, 2003). Paenibacillus larvae
subsp. pulvifaciens, i.e. the now reclassified ERIC III and IV
genotypes of P. larvae, has previously been isolated in South Africa
and Zambia (Ash et al., 1993; WTO 2008). Hansen et al. (2003)
suggested that retail honey purchased in South Africa and Guinea
Bissau was contaminated with P. l. larvae spores, but other surveys
conducted for the detection of AFB in honey samples from Kenya,
Senegal, South Africa, Tanzania, Uganda, Zambia, and Zimbabwe
found no contamination with AFB spores in the honey, or clinical
symptoms of the disease in bee colonies (Fries and Raina, 2003).
Since 2000, South Africa has been extensively surveyed for the
presence of AFB and after initial negative results, its presence has
now been confirmed in the Western Cape Province of the country
(Baxter, 2009).
International trade in honey and honey bee products is regulated
by the World Trade Organization (WTO), which sets the rules of trade
between nations at a global level. The Agreement on the Application
of Sanitary and Phytosanitary Measures (SPS Agreement) sets
constraints on members' trade policies relating to a number of areas
including animal health. Under the SPS agreement, trade restrictions
are required to be science based and in the case of animals and
animal products, these are administered in detail by the World
Organisation for Animal Health (OIE) based in Paris. The OIE
publishes scientifically based rules and regulations on trade in animals
and animal products which are referred to as the Terrestrial Animal
Health Code (TAHC) for any given year. The TAHC used as the basis
of this study is that of 2009. The current requirements for honey imports into South Africa from
any part of the world, including sub-Saharan Africa, are irradiation of
10kGy for honey, or other bee products containing any of these
ingredients (NDA, 2008). Irradiation is required due to a current non-
declaration of freedom from AFB status in any part of sub-Saharan
Africa, and is used as a mitigation measure to prevent the spread of
AFB in honey sourced from potentially infected areas. Honey
subjected to this treatment loses its organic properties, has a reduced
shelf life, and often turns black. Under the rules laid out in the OIE-
TAHC, 2009 it is therefore necessary for Swaziland to develop and
implement an effective and efficient honey bee pest surveillance
programme as a necessary component of the science based system
governing trade in honey with South Africa. The OIE-TAHC, 2009 requirements for declaration of freedom
from AFB can follow two paths, either with the declaration of a
historically free status in compliance with the provisions of Chapter
1.4 of the OIE-TAHC 2009, or as the result of an eradication program.
Given negative results of a risk assessment within the provisions of
Chapter 1.4 of the OIE-TAHC 2009 there is technically no requirement
for any specific type of survey or surveillance programme to
determine presence or absence of AFB. Surveillance to demonstrate
freedom from disease or infection (Article 1.4.6.1 subsection ‘a’
Historically free) requires, however, that infection is not known to be
established in the wild and that annual surveys have been carried out.
In the case of Swaziland, there is no historical or scientific
evidence to show the status of AFB in the country. The objectives of
this study therefore were: 1. to develop a scientific methodology for
market access for honey without irradiation that complies with
Chapter 9.2.3 paragraph 1 of the OIE-TAHC 2009; and 2. to develop a
methodology as a model for honey market access and bee health
studies in other parts of sub Saharan Africa.
Materials and methods Field survey and sampling
A stratified sampling frame was constructed to ensure that the survey
covered all three geographical (Murdoch, 1968) and five ecological
(Acocks, 1988) zones within Swaziland. In addition, it was necessary
to account for the high prevalence of hives in the main highveld
production areas comprising the eucalyptus forests between Sandlane
and Nhlangano in roughly the area designated as Piet Retief sourveld,
as well as the eucalyptus forests in an area roughly centred on Piggs
Peak in the Piet Retief sourveld transition area. The sampling frame
was therefore a conventional stratified four-stage cluster design in
which the first stage consisted of sampling clusters (honey producing
company), the second stage consisting of vegetation type, the third
stage of district or producers association and the fourth stage of
individual hives owned by a beekeeper (Table 1). The total number of
samples collected was estimated to represent just over 6% of the
total of 1500 commercial hives in Swaziland.
The sampling strategy was based on the honey robbing practices
of bees where it has been shown that colonies near to an AFB
collapsed colony have a high chance of rapidly contracting AFB, at
least sub-clinically. Studies show that 75% of colonies located 500m
or less from an AFB collapsed colony contracted AFB and died of the
disease, 50% of colonies 1km from the AFB collapsed colony died of
AFB disease (De Graaf et al., 2001; Pernal and Melanopoulos, 2006;
Lindstrom et al., 2008a). Approximately half the hives surveyed were of the straight sided
Tanzania Top Bar design and contained combs with mixed brood and
honey, the remainder being conventional Langstroth hives. Based on
the findings of Lindström et al. (2008a; 2008b) AFB, if present in the
swarm, would be also be detected in the honey. A further assumption
is based on recent work which indicates that where colonies are
known to be infected, analysis of honey samples may be of lesser
sensitivity as only 86% of such samples may be positive for AFB
(Gillarda et al., 2008). Brood comb and honey was sampled by
examining the brood comb for dead or discoloured brood and then
cutting a 20cm2 section of the brood comb for packing in paper
sample bags. About 60 to 80 g of honey was collected from each hive
American foulbrood in Swaziland 285
and placed in capped plastic bottles. The colour of the honey was
noted, ranging from light yellow to dark brown, and consistency from
liquid to solid. The survey was conducted in September to November
2008 during the main honey harvesting season.
Statistical analyses
The possible distribution of AFB was assumed to follow either a
binomial or negative binomial distribution, i.e. binomial distributions
when presence or absence was recorded (binomial sampling), and the
negative binomial when counts (integer values) are recorded (Sileshi,
2008). Therefore in order to determine the validity of the results,
probability mass functions (pmf) and cumulative distribution functions
(cdf) were calculated for both negative binomial and binomial
distributions. In terms of OIE standards, the required rate of detection
is at a 95% confidence rate given a 5% prevalence rate. The basic
assumption is that AFB occurs in 5% of the population of bee hives.
The 97 honey samples were picked at random using the sampling
frame in Table 1.
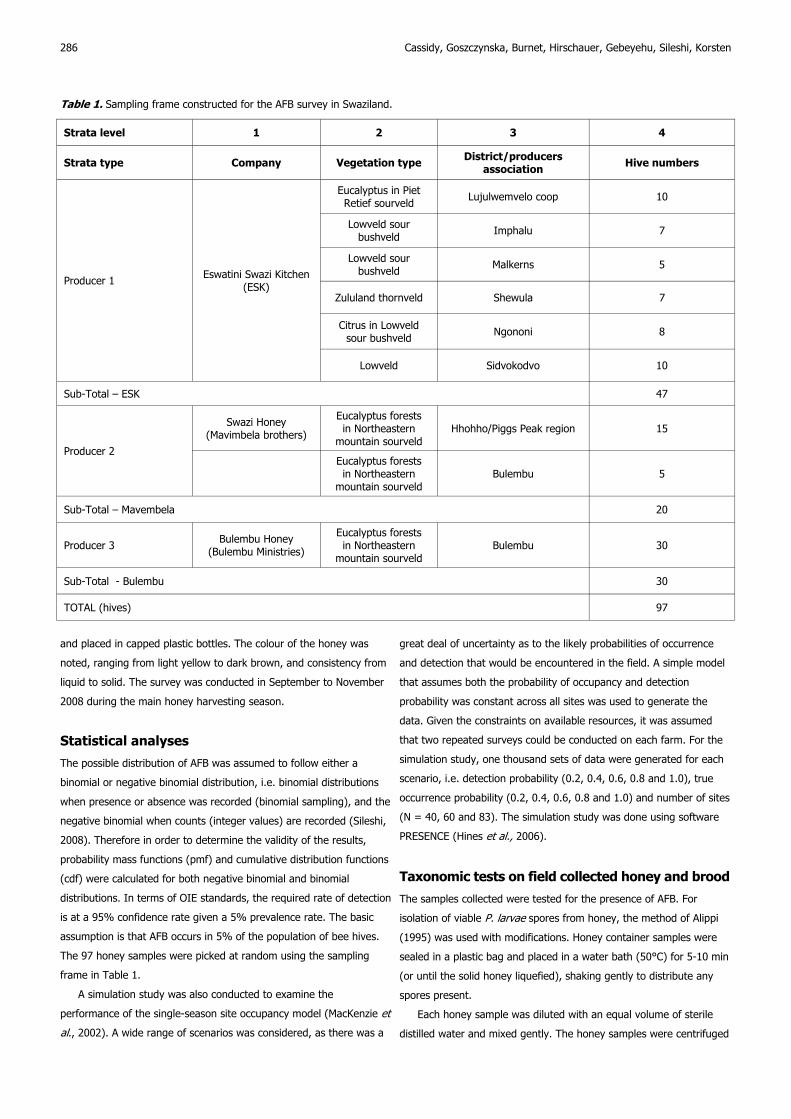
A simulation study was also conducted to examine the
performance of the single-season site occupancy model (MacKenzie et
al., 2002). A wide range of scenarios was considered, as there was a
286 Cassidy, Goszczynska, Burnet, Hirschauer, Gebeyehu, Sileshi, Korsten
great deal of uncertainty as to the likely probabilities of occurrence
and detection that would be encountered in the field. A simple model
that assumes both the probability of occupancy and detection
probability was constant across all sites was used to generate the
data. Given the constraints on available resources, it was assumed
that two repeated surveys could be conducted on each farm. For the
simulation study, one thousand sets of data were generated for each
scenario, i.e. detection probability (0.2, 0.4, 0.6, 0.8 and 1.0), true
occurrence probability (0.2, 0.4, 0.6, 0.8 and 1.0) and number of sites
(N = 40, 60 and 83). The simulation study was done using software
PRESENCE (Hines et al., 2006).
Taxonomic tests on field collected honey and brood
The samples collected were tested for the presence of AFB. For
isolation of viable P. larvae spores from honey, the method of Alippi
(1995) was used with modifications. Honey container samples were
sealed in a plastic bag and placed in a water bath (50°C) for 5-10 min
(or until the solid honey liquefied), shaking gently to distribute any
spores present.
Each honey sample was diluted with an equal volume of sterile
distilled water and mixed gently. The honey samples were centrifuged
Strata level 1 2 3 4
Strata type Company Vegetation type District/producers association Hive numbers
Producer 1 Eswatini Swazi Kitchen (ESK)
Eucalyptus in Piet Retief sourveld Lujulwemvelo coop 10
Lowveld sour bushveld Imphalu 7
Lowveld sour bushveld Malkerns 5
Zululand thornveld Shewula 7
Citrus in Lowveld sour bushveld Ngononi 8
Lowveld Sidvokodvo 10
Sub-Total – ESK 47
Producer 2
Swazi Honey (Mavimbela brothers)
Eucalyptus forests in Northeastern
mountain sourveld Hhohho/Piggs Peak region 15
Eucalyptus forests in Northeastern
mountain sourveld Bulembu 5
Sub-Total – Mavembela 20
Producer 3 Bulembu Honey (Bulembu Ministries)
Eucalyptus forests in Northeastern
mountain sourveld Bulembu 30
Sub-Total - Bulembu 30
TOTAL (hives) 97
Table 1. Sampling frame constructed for the AFB survey in Swaziland.

at 10,000 g for 25-30 minutes at 25°C. The supernatant was
discarded and the pellets were suspended in 0.5-1 ml of sterile
distilled water. The re-suspended pellets were transferred into
separate Eppendorf tubes and were placed in a heating block at 80°C
for 10 min in order to kill vegetative cells of bacteria and yeasts. The
pellet suspensions were plated on J-agar medium made in the
laboratory supplemented with 30µg/ml nalidixic acid and 20µg/ml
pipemedic acid (both obtained from Sigma Chemical Co.; St. Louis,
MO, USA) and 5% CO2 atmosphere which inhibits the growth of many
microorganisms present in honey (Alippi, 1995). A volume of 0.1 ml of
the suspension was streak-plated on the J-agar medium, so as the
total volume of suspensions was 1 to 1.5 ml, each suspension was
plated on 10 to 15 J-agar plates. The type strains of Ab, ab and aβ genotypes and AB genotype P.
larvae were plated on J-agar to serve as positive controls (P. larvae
LMG 9820T (synonym P. larvae subsp. P. larvae, type strain), P. larvae
LMG 15974T (synonym P. larvae subsp. pulvifaciens, type strain) and
P. alvei LMG 13253T, type strain. Inoculated plates were incubated in
5% CO2 atmosphere in the incubator at 37°C for 4-7days.
Representative colonies from each sample were selected for initial
identification assessing colony shape and margins, microscopic
characterization, and standard biochemical tests (Alippi, 1995).
Bacteria were isolated from all samples. Representative suspect
colonies grown on J-agar plates were examined for initial identification
assessing colony shape and margins, microscopic characterization and
standard biochemical tests, growth at 20°C, Gram stain, catalase test,
starch hydrolysis, milk hydrolysis, acid production from mannitol, D-
glucose and salicin (Alippi et al., 2002). Some non-target bacteria
may, however, still be recovered. For example P. larvae LMG 9820T
produces small, white, circular colonies on J-agar within 4-5 days.
Another genotype of P. larvae LMG 15974T also grows on J-agar.
Colonies are very similar to that of P. larvae LMG 9820T. Some strains
of P. larvae produce orange to brown colonies, but this is not
considered a diagnostic feature (Genersch et al., 2006).
Biochemical characterization of selected isolates
The isolated strains were tested with the API 50CHE system
(bioMérieux Clinical Diagnostics; La Balme les Grottes, Montalieu
Vercieu, France) using the procedure recommended by the
manufacturers. The results of the API 50CHE tests were recorded
after 72 h of incubation at 37oC.
Molecular detection and identification by PCR
The method of isolation of P. larvae from honey, primers and the PCR
conditions were performed as specified by Bakonyi et al. (2003).
Paenibacillus larvae specific confirmation of AB genotype suspected
colonies was carried out by PCR, using the primers AF6f 5’- GCA AGT
CGA GCG GAC CTT GT -3’ and AF7r 5’- GCA TCG TCG CCT TGG TAA
American foulbrood in Swaziland 287
GC -3’ which amplifies a fragment (237 bp) of the 16S rRNA gene of
the Ab, ab and aβ genotypes of Paenibacillus larvae. DNA was
extracted from the selected colonies as described by Bakonyi et al.
(2003) with the same reaction conditions.
BioPCR
The method used was that of Schaad et al. (1995) which enhances
the sensitivity of PCR reaction. Bio-PCR detects living cells of
pathogens, those that could cause a disease, as bacterial colonies are
washed up from agar plates preceding the PCR reaction. A honey
extract was plated on J-agar supplemented with 30µg/ml nalidixic acid
and 20µg/ml pipemedic acid (both obtained from Sigma Chemical Co.;
St. Louis, MO, USA). Plates were incubated for five to seven days at
37oC in 5% CO2 atmosphere which inhibits the growth of many
microorganisms present in honey (Alippi, 1995). After incubation the
bacterial growth was removed from the agar plates and suspended in
sterile distilled water. This bacterial suspension was used as a
template in the PCR with primers targeting P. larvae.
Extraction of DNA
Genomic DNA was extracted from pure cultures by using the GenElute
Bacterial Genomic DNA kit (Sigma). The concentration of DNA was
measured with the NanoDroP. Extracted DNA was stored at -20oC
until needed.
Electrophoresis of PCR products
The PCR products were separated in a 1% agarose gel with ethidium
bromide, in 1 x TAE buffer at 100 V for 45-50 min. The bands were
visualized under UV light and photographed with a Kodak DS
electrophoresis documentation and analysis system, using the Kodak
Digital Science ID software program (Fig. 2).
Tests using the VITA® AFB diagnostic kits
Confirmatory tests of suspect colonies were carried out using VITA®
AFB diagnostic test kit (Vita (Europe) Ltd., Basingstoke, UK). Orange
colonies and brood from these hives tentatively identified as Ab, ab
and aβ genotypes of P. larvae were tested using the methodology as
described in the VITA® test kit leaflet by substituting colonies of
suspected Ab, ab and aβ genotypes of Paenibacillus larvae instead of
larvae with suspicious symptoms. Test colonies were deposited into
the extraction bottle and shaken vigorously for about 20 seconds to
mix them into the buffer. A VITA® test device was removed from the
foil pack and a sample pipetted from the bottle immediately after
shaking. Two drops were placed onto the sample well of the device
and the device kept horizontal until the extract was absorbed (c. 30
seconds) and a blue dye appeared in the viewing window. With the
appearance of the control line after 1-3 minutes the result was
recorded.
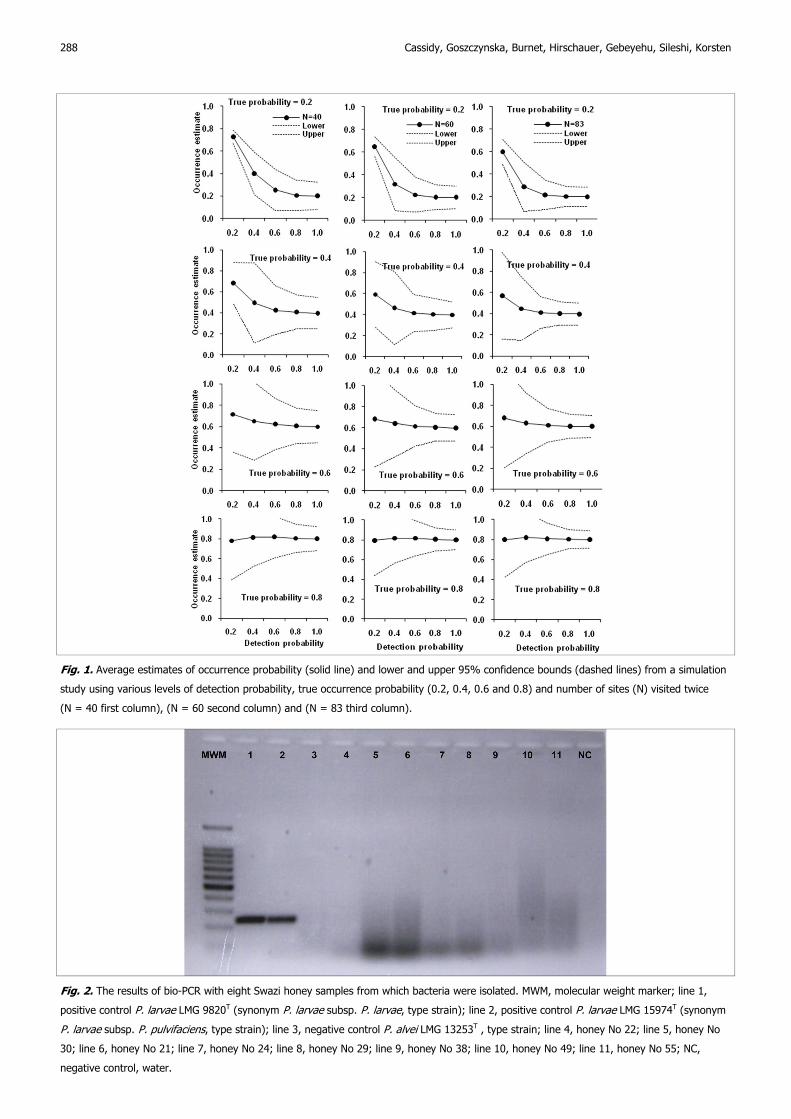
Cassidy, Goszczynska, Burnet, Hirschauer, Gebeyehu, Sileshi, Korsten
288
Fig. 2. The results of bio-PCR with eight Swazi honey samples from which bacteria were isolated. MWM, molecular weight marker; line 1,
positive control P. larvae LMG 9820T (synonym P. larvae subsp. P. larvae, type strain); line 2, positive control P. larvae LMG 15974T (synonym
P. larvae subsp. P. pulvifaciens, type strain); line 3, negative control P. alvei LMG 13253T , type strain; line 4, honey No 22; line 5, honey No
30; line 6, honey No 21; line 7, honey No 24; line 8, honey No 29; line 9, honey No 38; line 10, honey No 49; line 11, honey No 55; NC,
negative control, water.
Fig. 1. Average estimates of occurrence probability (solid line) and lower and upper 95% confidence bounds (dashed lines) from a simulation
study using various levels of detection probability, true occurrence probability (0.2, 0.4, 0.6 and 0.8) and number of sites (N) visited twice
(N = 40 first column), (N = 60 second column) and (N = 83 third column).

Results Detection simulations
The detection simulations were run using a residual valid sample
number corrected for this reduced sensitivity i.e. assuming that a total
of only 83 samples were collected. With the assumed prevalence rates
returns the required detection confidence rate of 95% at sample 58
for a negative binomial distribution.
The calculated confidence of detection at this prevalence rate is
thus well within the required OIE-TAHC 2009 norms. Alternatively
using the binomial equation based detection simulation software
inputted with the OIE-TAHC norms the following detection
probabilities were produced using PRESENCE2 software method as
described by Hines (Hines, 2006). A probability P =0.012 is returned
for 0 detections in 83 samples based on the assumption of an AFB
prevalence of 5%. A 95% confidence of detection rate requires 59
samples which are close to those calculated for the negative binomial
distribution above.
Fig. 1 presents results of the simulation study. These suggest that
when the true probabilities of occurrence is low (0.2) and detection
probability is also low (0.2-0.4), occurrence will be overestimated by
30-70%, even with large number of sites (83 sites). Similarly, when
the true probabilities of occurrence is 0.4 and detection probability is
also low (0.2-0.4), occurrence will be overestimated by 10-41%. At
these low levels of occurrence and detection probability, the standard
errors will also be large and erratic (Fig. 1). The simulation results
seem generally stable for true occurrence probabilities of 0.8 and
detection probability of 0.6 or more. When the true occurrence
probability is 0.6 or more, even with low detection probability levels
and fewer sites (as few as 40) can give reasonable estimates of the
average occurrence probability. These results highlight the fact that at
low levels of AFB prevalence (and if the method used cannot ensure
up to 60% detection), there will be huge false positive results. When
detection probability is high (1.0), there was also about 1.0% increase
in false negative values.
Taxonomic tests
All suspect colonies were purified on J-agar and identity confirmed
using biochemical tests. Bacterial growth in 39 samples was observed
on J-agar. From three samples, however, an almost pure culture of
yellow, round colonies was isolated, and nine colonies from these
samples were purified on J-agar for additional testing. None of these
bacterial colonies produced a biochemical profile similar to P. larvae
LMG 9820T. Three representative strains that were catalase negative
were, however, used in the API 50 CHE biochemical strips. The three
isolates produced biochemical profiles that did not resemble that of P.
larvae LMG 9820T. Bacterial growth of 39 samples from J-agar plates
showing presumptive pathogen growth was tested by bio-PCR with
negative results. Electrophoresis results of PCR products were
American foulbrood in Swaziland 289
negative for AFB (Fig. 2). Tests using the VITA® AFB diagnostic kits
were negative when tested with suspect colonies and gave weakly
positive, but definite reactions, to the type strains of AFB.
Discussion Despite the adoption and publication of AFB surveillance standards by
the OIE, there is no evidence in the literature that any current honey
bee disease surveillance programme in the world has ever been
designed to meet these standards. New Zealand finalized a Pest
Management Strategy (PMS) for AFB, and reported the results of the
national programme but the design does not explicitly follow OIE
guidelines (Anonymous, 2008). The Swaziland survey for AFB is
therefore the first such formal exercise ever conducted according to
OIE guidelines for the application of OIE-TAHC 2009 to establish a
path for future trade in honey, particularly within sub-Saharan Africa
where the current status of AFB is a concern.
The results of the survey reported here demonstrate that AB
genotype P. larvae is absent from Swaziland using the criteria
stipulated by the OIE-TAHC-2009. This confirms studies that adult
bees from wild colonies in areas without intensive beekeeping rarely
contain detectable spore levels (Lindström, 2006). Furthermore, until
the recently reported outbreak in the Western Cape of South Africa
(Baxter, 2009), clinical cases of AFB have never been found in wild
honey bees south of the Sahara (Fries and Raina, 2003).
The reasons for the freedom of sub-Saharan Africa from AFB are
the subject of some speculation. A significant part of the problem is
the lack of long term honey bee systematic surveillance programmes
on the continent outside South Africa. Even in South Africa, AFB
surveillance has been conducted only since 2000, and the results have
not been published in peer-reviewed journals. Given the lightly
regulated nature of African trade in honey, bees and used bee
keeping equipment, it is likely that AFB has been introduced many
times to the continent. The recent outbreak of AFB in the Western
Cape of South Africa shows, however, that the continent’s bees are
not resistant to this disease and that appropriate surveys followed by
ongoing surveillance based on Article 1.4 of the OIE-TAHC, 2009 are
urgently necessary, not only in Swaziland but in other parts of sub-
Saharan Africa where bees are economically important as well as
being a key part of the ecology. As follow up activities to these survey results, a full review of
existing Swazi legislation, statutory instruments and regulations as
they address bee matters and bee health is underway. In addition the
design and implementation of a survey and surveillance programme
using the guidelines in the OIE TAHC, 2009 coupled with an
appropriate AFB management strategy if and when the disease arrives
in Swaziland, has been devised. A survey of Swaziland’s potential for
bees and honey production (forage, habitat etc) is urgently needed.
This together with information on land use patterns, human attitudes

(security of hives), the existing use of moveable frame hives, and
deficiencies in apicultural management knowledge in the country is
hoped to form the foundation for the sustainable management of
honey bee pests and diseases and development of the apicultural
industry.
Tests of presumptive colonies of AFB using the VITA® AFB
diagnostic kits have been demonstrated as a quick method of
screening laboratory cultures to determine the presence of AFB.
Acknowledgements Funding and technical assistance for this study was made possible by
a grant from USAID under their SPS support programme for the
Southern African Development Community (SADC). This is through
their Participatory Agency Services Agreement (PASA) with the United
States Department of Agriculture - Foreign Agriculture Service (USDA-
FAS).
References ACOCKS, J P H (1988) Veld types of South Africa. Memoirs of the
Botanical Society of South Africa 57. ISBN 0621 11394 8.
ALIPPI, A M (1995). Detection of Bacillus larvae spores in Argentinean
honeys by using a semi-selective medium. Microbiologica SEM 11:
343–350.
ALIPPI, A M; LOPEZ, A C; AGUILAR, O M (2002) Differentiation of
Paenibacillus larvae subsp. P. larvae, the cause of American
foulbrood of honey bees, by using PCR and restriction fragment
analysis of genes encoding 16S rRNA. Applied and Environmental
Microbiology 68(7): 3655–3660.
ANONYMOUS (2006) Situation analysis of beekeeping industry. Total
Transformation Agribusiness (Pty) Ltd. http://
www.beekeeping.com/articles/us/beekeeping_regional_situational
-analysis.pdf (accessed on 04 August 2011).
ANONYMOUS (2008) Report on the American foulbrood National Pest
Management Strategy. MAF Biosecurity New Zealand Discussion
Paper No: 2008/07, 51pp ISBN 978-0-478-32183-8 ISSN 1176-
8398.
ANONYMOUS (2009a) The outbreak of American Foul Brood (AFB) in
the Republic of South Africa, 25 March 2009, http://
www.info.gov.za/speeches/2009/09040109451001.htm, (accessed
on 04 April 2009).
ANONYMOUS (2009b) Animal health monitoring and surveillance
status of reportable diseases in the United States. http://
www.aphis.usda.gov/vs/nahss/disease_status.htm (accessed on
03 February 2009).
290 Cassidy, Goszczynska, Burnet, Hirschauer, Gebeyehu, Sileshi, Korsten
ANONYMOUS (2009c) Development of the VITA ® AFB Diagnostic Kit,
http://www.vita-europe.com/Map_enscript/frmbuilder.php?
dateiname=/en/products/afb_dignosyic.htm (accessed on 02
February 2009).
ASH, C; PROEST, F; COLLINS, M (1993) Molecular identification of
rRNA group 3 bacilli using a PCR probe test. Antonie van
Leeuwenhoek 64: 253-260.
BAKONYI, T; DERAKHSHIFAR, I; GRABENSTEINER, E; NOVOTNY, N
(2003) Development and evaluation of PCR assays for the
detection of Paenibacillus larvae in honey samples: comparison
with isolation and biochemical characterization. Applied and
Environmental Microbiology 69: 1504-1510.
BAXTER, A (2009). American foulbrood (AFB) in the Western Cape:
advisory notice. South African Bee Journal 81(1): 8-9.
BRØDSGAARD, C J; HANSEN, H (2003). Tolerance mechanisms
against American Foulbrood in honey bee larvae and colonies.
Apiacta 38: 114–124.
DE GRAFF, D C; VANDEKERCHOVEA, D; DOBBELAERE, W; PEETERS,
J E; JACOBS, F J (2001) Influence of the proximity of American
foulbrood cases and apicultural management on the prevalence of
Paenibacillus larvae spores in Belgian honey. Apidologie 32: 587-599.
FRIES, R; RAINA, S (2003) American foulbrood and African honey
bees (Hymenoptera: Apidae) Journal of Economic Entomology 96
(6): 1641-1646.
GENERSCH, E; FORSGREN, E: PENTIKÄINEN, J; ASHIRALIEVA, A;
RAUCH, S; KILWINSKI, J; FRIES, I (2006) Reclassification of
Paenibacillus larvae subsp. P pulvifaciens and Paenibacillus larvae
subsp. P. larvae as Paenibacillus larvae without subspecies
differentiation. International Journal of Systematic Evolutionary
Microbiology 56: 501-511.
GILLARDA, M; CHARRIEREB, J D; BELLOY, L (2008) Distribution of
Paenibacillus larvae spores inside honey bee colonies and its
relevance for diagnosis. Journal of Invertebrate Pathology 99(1):
92-95.
GOVAN, V A; ALLSOPP, M H; DAVISON, S (1999) A PCR detection
method for rapid identification of Paenibacillus larvae. Applied and
Environmental Microbiology 65(5): 2243–2245.
HANSEN, H; BRØDSGAARD, C J; KRYGER, P; NICOLAISEN, M (2003)
A scientific note on the presence of Paenibacillus larvae larvae
spores in sub-Saharan African honey. Apidologie 34: 471–472.
HINES, J E; MACKENZIE, D I (2006) PRESENCE2 – Software to
estimate patch occupancy and related parameters. USGS-PWRC.
http://www.mbr-pwrc.usgs.gov/software/presence.html (accessed
19 February 2009)
KILWINSKI, J; PETERS, M; ASHIRALIEVA, A; GENERSCH, E (2004)
Proposal to reclassify Paenibacillus larvae subsp. P. pulvifaciens
DSM 3615 (ATCC 49843) as Paenibacillus larvae subsp. P. larvae.
Results of a comparative biochemical and genetic study.
Veterinary Microbiology 104: 1-2, 30, 31-42.

LINDSTRÖM, A; KORPALA, S; FRIES, I (2008a) Horizontal
transmission of Paenibacillus larvae spores between honey bee
(Apis mellifera) colonies through robbing. Apidologie 39: 515-521.
LINDSTRÖM, A; KORPALA, S; FRIES, I (2008b). The distribution of
Paenibacillus larvae spores in adult bees and honey and larval
mortality, following the addition of American foulbrood diseased
brood or spore-contaminated honey in honey bee (Apis mellifera)
colonies. Journal of Invertebrate Pathology 99: 82–86.
MACKENZIE, D I; NICHOLS, J D; LACHMAN, G B; DROEGE, S; ROYLE,
J A; LANGTIMM, C A (2002) Estimating site occupancy rates when
detection probabilities are less than one. Ecology 83: 2248–2255.
MATHESON, A (1996) World bee health update. Bee World 77: 45-51.
MURDOCH, G (1968) Soils and land capability in Swaziland. Swaziland
Ministry of Agriculture. 360pp.
NDA (2008), South African National Department of Agriculture, Honey
Import Permit, Additional declaration.
OIE (2009) Terrestrial Animal Health Code (TAHC) 2009. http://
www.oie.int/eng/normes/Mcode/en_sommaire.htm (accessed 02
November 2009).
PERNAL, S F; MELANOPOULOS, A P; (2006) Monitoring for American
foulbrood spores from honey and bee samples in Canada. Apiacta
41: 99-109.
American foulbrood in Swaziland 291
SCHAAD, N W; TAMAKI, S; HATZILOUKAS, E; PANOPUOLAS, N (1995)
A combined biological amplification (BIO-PCR) technique to detect
Pseudomonas syringae pv. phaseolicola in bean seed extracts.
Phytopathology 85: 243-248.
SILESHI, G (2008) The excess-zero problem in soil animal count data
and choice of appropriate models for statistical inference.
Pedobiologia 52: 1-17.
WAITE, R; THOMPSON, H; BROWN, M; WATKINS, M; BEW, M (2003)
Preliminary studies into novel detection methods for honey bee
pathogens. Proceedings of XXXVIIIth International Apicultural
Congress, Ljubljana, Slovenia.
WORLD ORGANIZATION FOR ANIMAL HEALTH (OIE) (2009) World
Animal Health Information Database (WAHID) - Version: 1.3
http://www.oie.int/wahis (accessed 05 March 2009).
WORLD TRADE ORGANIZATION (2008) Information on various SPS
matters, Communication from Zambia, G/SPS/GEN/836 27 March
2008 (08-1369), Committee on Sanitary and Phytosanitary
Measures.
Related Documents











