Isolation and characterisation of four distinct cytotoxic factors of Salmonella Weltevreden B R Singh & V.D. Sharma Department of Veterinary Microbiology College of Veterinary Sciences G B Pant University of Agriculture & Technology Pantnagar-263 145, India Four distinct cytotoxins with different biological, physico-chemical and antigenic characteristics were isolated from a single Salmonella Weltevreden strain recovered from buffalo meat. The toxins were purified through salt precipitation, dialysis, gel filtration and ion-exchange chromatographic methods. Cytotoxin I was dermonecrotic, verocytotoxic and lethal to mouse with 120 μg LD 50 . It induced accumulation of serosanguinous fluid in rabbit ligated ileal loop (RLIL) and mucinous fluid in the stomach. It was active within a narrow pH range (5.0 - 8.0) and lost its activity on autoclaving for 1 min. Cytotoxin II was verocytotoxic, enterotoxic and lethal to mouse (LD 50 , 1 mg). It induced delayed vasopermeability in rabbit skin, was active between pH 5.6 and 8.4 only and heat- sensitive (100°C, 30 min). Cytotoxin III was neither dermatotoxic nor enterotoxic. It induced vacuolation, multinucleation and formation of syncytia in vero cells. It was sensitive to pH beyond the range of 4.8 to 8.2. It was completely inactivated on boiling for 30 min. Cytotoxin IV was intensely necrotizing to rabbit skin within 6 h of inoculation and lysed Vero cells. It also possessed haemolytic and lecithinase/phospholipase-C activities. The cytotoxin lost its activity on heating at 90°C (30 min) but remained active between pH 2.5 and 7.5. The PIs of the cytotoxins were estimated to be 9.0, 7.0, 5.6 and 3.0, respectively. All the four cytotoxins were immunogenic in rabbits but antigenically unrelated as the anticytotoxin neutralized only the homologous cytotoxin and did not cross react with heterologous cytotoxins. * Present address: Scientist, National Salmonella Centre (Vet), Division of Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar-243 122 (UP), India. PDF created with pdfFactory Pro trial version www.pdffactory.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Isolation and characterisation of four distinct cytotoxic factors of
Salmonella Weltevreden
B R Singh & V.D. Sharma
Department of Veterinary Microbiology
College of Veterinary Sciences
G B Pant University of Agriculture & Technology
Pantnagar-263 145, India
Four distinct cytotoxins with different biological, physico-chemical and antigenic
characteristics were isolated from a single Salmonella Weltevreden strain recovered from buffalo meat.
The toxins were purified through salt precipitation, dialysis, gel filtration and ion-exchange
chromatographic methods. Cytotoxin I was dermonecrotic, verocytotoxic and lethal to mouse with 120
µg LD50. It induced accumulation of serosanguinous fluid in rabbit ligated ileal loop (RLIL) and
mucinous fluid in the stomach. It was active within a narrow pH range (5.0 - 8.0) and lost its activity on
autoclaving for 1 min. Cytotoxin II was verocytotoxic, enterotoxic and lethal to mouse (LD50, 1 mg). It
induced delayed vasopermeability in rabbit skin, was active between pH 5.6 and 8.4 only and heat-
sensitive (100°C, 30 min). Cytotoxin III was neither dermatotoxic nor enterotoxic. It induced
vacuolation, multinucleation and formation of syncytia in vero cells. It was sensitive to pH beyond the
range of 4.8 to 8.2. It was completely inactivated on boiling for 30 min. Cytotoxin IV was intensely
necrotizing to rabbit skin within 6 h of inoculation and lysed Vero cells. It also possessed haemolytic and
lecithinase/phospholipase-C activities. The cytotoxin lost its activity on heating at 90°C (30 min) but
remained active between pH 2.5 and 7.5. The PIs of the cytotoxins were estimated to be 9.0, 7.0, 5.6 and
3.0, respectively. All the four cytotoxins were immunogenic in rabbits but antigenically unrelated as the
anticytotoxin neutralized only the homologous cytotoxin and did not cross react with heterologous
cytotoxins.
* Present address: Scientist, National Salmonella Centre (Vet), Division of Bacteriology and Mycology, Indian
Veterinary Research Institute, Izatnagar-243 122 (UP), India.
PDF created with pdfFactory Pro trial version www.pdffactory.com

Introduction
Salmonella, still remains to be one of the most important causes of
foodborne outbreaks. It leads to gastro-intestinal disorders frequentlyresulting to
systemic salmonellosis. Although Salmonella has been most thoroughly investigated and
more than 2400 serovars are known, its viruelence factors and pathogenesis are yet to be
fully delineated11,27 . Invasiveness of Salmonella is considered to be an important
attribute of virulence2, 4. However, all invasive Salmonella do not induce gastroenteritis.
Enterotoxins have been shown to play significant role in enteric diseases and as many as
54-98% of Salmonella strains have been found to be enterotoxigenic1,2,18. Of the two
types of enterotoxins, i.e. cytotonic and cytotoxic, the former one, which is commonly
referred to as enterotoxin, has inconsistent production and is considered to be less
significant than the cytotoxic enterotoxins (cytotoxins)1,24. Cytotoxigenicity of
Salmonella strains has been shown to be more closely related to their pathogenic potential
in experimental studies1,3,13,22. Partially purified cytotoxin is found to produce typhoid
like lesions, viz., enlargement of spleen, pin point haemorrhages in liver, heart, lungs,
kidneys and brain, congestion in thoracic and peritoneal cavites, focal necrosis in liver
and blood tinged contents in small intestine in mice22. Salmonella cytotoxin has been
purified to a variable extent 1,13,22. This paper reports the isolation of four distinct
cytotoxic moieties from a single strain of S. Weltevreden and their physico-chemical,
biological and immunological characterization.
Materials and Methods
A strain of S. Weltevreden (BM 1643) isolated from buffalo meat and identified
as potent cytotoxin producer22 was procured from the Department of Veterinary
Microbiology, College of Veterinary Sciences, Pantnagar. Escherichia coli K12 (J-53)
strain obtained from the National Salmonella Centre (Veterinary), Izatnagar, was used as
non-toxigenic reference strain.
PDF created with pdfFactory Pro trial version www.pdffactory.com

Experimental animals. Adult (3-month-old) New Zealand White rabbits and adult Swiss
albino mice were obtained from Laboratory Animal Section of the College of Veterinary
Sciences, Pantnagar. Animals were maintained on pathogen-free diet supplied from the
Laboratory Animal Section of the College.
Cytotoxin production. Cell-lysate (CL), cell-free culture filtrate (CFCF) and polymyxin B
extract (PBE) were prepared according to the method described by Malik et al.12,13. The
test organism was propagated in brain heart infusion broth (Hi Media, Mumbai) at 37°C
on a shaker (200 rpm) for 18-24 h. The growth was then centrifuged at 10,000 X g for 10
min. The supernatant was separated and filtered through a 0.45 µm membrane filter to
obtain CFCF. The pellet was subjected to sonication at 150 mA for 30 min and
centrifuged at 10,000 X g for 20 min; the supernatant, so collected, was designated as
CL. To prepare PBE, cell pellet from one litre culture material was suspended in 50 ml
PBS (pH 7.2, 0.15 M) and supplemented with 100 IU of polymyxin B/ml. The preparation
was incubated at 37°C on a shaker (200 rpm) for 4-6 h. The supernatant was collected
after centrifugation as described earlier. All preparations were stored at -20°C after
determining their protein concentration11 and adjusting it to 5 mg/ml.
Test for cytotoxicity. Cytotoxicity of different preparations was determined on African
green monkey kidney (Vero) cells and Madin Darby bovine kidney (MDBK) cells
according to the method of Spiers et al.23 in 96 well tissue culture plates. Stock cells
were trypsinized and grown in the minimum essential medium (MEM, Hi-Media,
Mumbai) supplemented with penicillin G (100 IU/ml), streptomycin (100 µg/ml) and
foetal calf serum (10%). Monolayers were exposed to 100 µl of serially diluted (in PBS,
pH 7.2, 0.15 M) test preparation for 30 min and then 200 µl of MEM supplemented as
above was added to each well. Plates were incubated at 37°C and examined up to 72 h at
6 h interval under inverted light microscope from cytopathic effects (CPE). Cytotoxic
units per mg of protein were determined by dilution method21. Minimum quantity of the
test preparation in terms of protein inducing appreciable CPE in about 50% of tissue
PDF created with pdfFactory Pro trial version www.pdffactory.com

culture cells within 30 h of exposure was designated as one cytotoxic unit. Some of the
tissue culture plates were stained with May Grhnwald Giemsa staining method14, using
absolute methanol as fixative and destainer instead of acetone, xylene and methanol
mixture.
Purification of cytotoxin. PBE was used for the purification of Salmonella cytotoxins as it
contained comparatively less impurities13,19. PBE obtained from 100 ml culture was salt
precipitated with ammonium sulphate at different saturation levels ranging from 30 to
80% at an intervals of 10%. Its pH was maintained near neutrality with help of
ammonium hydroxide. Prepcipitate was washed with corresponding ammonium sulphate
salt solution. The pellet was dissolved in distilled water and desalted on Sephadex G10
(Pharmacia) column equilibrated with Tris-HCl buffer (0.037 M, pH 7.2) and tested for
cytotoxicity. PBE from one litre culture was processed to get salt precipitated protein
(SPP) after determining the salt concentration that precipitated maximum amount of the
cytotoxin. After desalting, SPP was concentrated by dialysing the precipitate against
polyethylene glycol 20,000 to a concentration of 10 mg/ml.
For gel filtration, Seralose 6 B column (2 x 50 cm, SRL, Mumbai) was
equilibrated and eluted at a low rate 1 ml/min with Tris HCl buffer. Elution was
monitored on 280 nm with UV monitor (Pharmacia) and peak materials were collected,
tested for cytotoxicity and stored at -20°C. The cytotoxic peak materials were again
filtered through the same column and protein concentration was adjusted to 5 mg/ml
through dialysis as above. For further purification, 0.5 ml of the preparation was loaded
onto Mono-P column of FPLC Pharmacia equilibrated and eluted with Tris-HCl buffer
for 20 min at 0.75 ml/min speed. The salt gradient was raised by 0.5 M at every 20 min
interval up to 5 M. Finally, it was eluted with 0.025 M acetic acid. Eluent was monitored
at 280 nm and peak materials were collected separately, tested for cytotoxicity and stored
at -20°C. Cytotoxic peak materials collected from 20 runs of column P were concentrated
and passed again through the same column as above. For further purification of eluent
PDF created with pdfFactory Pro trial version www.pdffactory.com

passed unbound to the column, the preparation was again eluted with Tris-HCl buffer
(pH, 8.6) and salt gradient (0.1-0.5 M) as above through the same colum equilibrated
with Tris-HCl buffer (0.037 M, pH, 8.6). Concentrated peak materials were subjected to
polyacrylamide gel electrophoresis (PAGE) and sodium dodecyl sulphate PAGE16. For
PAGE and SDS-PAGE semipurified and purified preparations were loaded at the rate of
100 µg and 40 µg/ lane ,respectively. Gels were stained16 with coomassie blue and/or
periodic acid Schiff (PAS) stain to visualize the protein bands.
Preparation of immune serum. After collecting pre-immune serum, each of the three
rabbits, was inoculated intradermally in the abdominal region with 100 µg of purified
toxin without adjuvant1. Three booster doses, each of 100 µg, were administered at an
interval of 15 days. Serum was collected after 10 days of the last booster and stored at -
20°C.
Characterization of cytotoxin. To further define the purified cytotoxins, the preparations
were characterized physicochemically, biologically and immunologically as follows :
In order to evaluate heat sensitivity of the cytotoxin of the preparation, 0.5 ml of
the test preparation was heated at 60°C (30 min), 90°C (30 min), 100°C and 121°C (1, 5,
15 and 30 min).
To determine the effect of proteolytic enzymes, viz., trypsin, chymotrypsin,
pepsin, and bacterial proteases on cytotoxins, method of Jacks and Wu6 was followed.
The effect of pH on cytotoxicity was assessed by adjusting17 the pH of the toxin
in a range of 2.0 through 10.0 at an interval of 0.2 with 1N HCl or 1N NaOH. The test
material was incubated at 37°C for 4 h and finally the pH was readjusted to neutrality.
Cytotoxicity of treated preparations was determined as described earlier and compared
with that of untreated controls.
The effect of formalin on cytotoxicity was determined by the method of Lallier
and Lariviere9.
PDF created with pdfFactory Pro trial version www.pdffactory.com

Haemolytic potential of various purified cytotoxins was determined by using
freshly collected erythrocytes of rabbit, sheep, guinea pig and humans (O group) after
washing them with PBS (pH, 7.2, 0.15 M). 100 µl of toxin preparation was mixed with
equal volume of 2 % RBC (red blood cells) suspension (in PBS pH, 7.2, 0.15 M) in a 96-
well plate and then incubated at 37°C for 6 h. After recording the results, the plates were
kept at 4°C for overnight and read finally.
Lecithinase activity of different cytotoxins was assessed on egg yolk agar plates7.
Wells (6 mm dia) were cut, sealed and filled with 100 µl of test preparations. The plates
were incubated at 37°C for 18-24 h. Formation of a zone of opacity around wells was
considered as positive reaction.
Mouse foot pad test20 was performed by injecting 0.05 ml of toxic preparation
intradermally in one of the hind foot pads, while the other was kept as negative control
(0.05 ml PBS, pH 7.2, 0.15 M). Observations were recorded after 24 h by measuring
thickness of both the foot pads. An index of ≥ 125 was considered positive.
Dermatotoxicity of test preparations was assayed by injecting 0.1 ml of each
preparation intradermally on abdominal region of New Zealand white rabbits17. The
reaction in skin, if any, was recorded at 6, 18, 24, 48, 72 h and 7 days post-inoculation.
Rabbit ligated ileal loop (RLIL) technique17 was used to detect the enterotoxic
effect of crude and purified cytotoxins. A dilatation index of ≥ 0.5 was considered
positive. Cholera toxin (Sigma Chemical Co., USA) was used as positive control.
Mouse lethality assay22 was performed by injecting 0.1 ml of serially diluted
preparation (in PBS pH 7.2, 0.15 M) intraperitoneally (ip) in 6 mice for determining 50%
lethal dose (LD50) of the toxins. LD50 was considered to be equal to the amount capable
to causing mortality within 10 days of inoculation in 50% of the mice.
PDF created with pdfFactory Pro trial version www.pdffactory.com

To perform neutralisation test, serial dilution of Salmonella cytotoxins and their antisera
were mixed1 in 96 well tissue culture plates. The plates were incubated at 37°C for 4 h.
Contents of each of the wells was tested for cytotoxicity, dermatotoxicity, lecithinase
activity and mouse lethality following the procedures described earlier.
Agar gel precipitation test (AGPT) was performed5 to detect antigenic
relatedness between different S. Weltevreden toxins and their antisera. The effect of
various treatments on antigenicity of cytotoxins was also determined with this technique.
Besides, different purified preparations were also tested with AGPT against antisera to O
(3,10,15 ) and H (r, Z6) factors of Salmonella Weltevreden. To determine endotoxin
contamination in purified cytotoxic preparation, E. Toxate assay (Sigma Chemical Co.,
USA) was performed according to the instructions of manufacturer. Briefly, 0.1 ml of the
test solution (100 µg purified cytotoxins, 0.1 ml pyrogen-free water and 0.1 ml of E.
Toxate was mixed in endotoxin-free tubes, incubated at 37°C and examined after 30 and
60 min for gel formation). Positive control tubes contained 0.1 ml of S. Weltevreden LPS
(1 and 5 µg).
For performing agglutination test6, one drop of fresh 18 h culture was mixed with
a drop of antiserum and agglutination was observed within 5-10 min.
Results
Toxic activities of crude preparations. All the three preparations, i.e., the CFCS, CL and
PBE induced rounding, granulation, vacuolation, degenration, detachment and cell lysis
in the MDBK and Vero cell lines. On rabbit skin, crude cytotoxin caused central white
zone of necrosis surrounded by oedematous hot and red swelling. Necrosed area
sloughed off within 3 days leaving an ulcer behind. However, the CFCS did not cause
early necrosis but other lesions were similar to those induced by CL and PBE including
sloughing of skin. All the 3 preparations induced serosanguinous fluid accumulation in
RLIL loops and distension of the stomach. The mice dying with crude preparations
PDF created with pdfFactory Pro trial version www.pdffactory.com

experienced diarrhoea before death and had distended stomach and urinary bladder with
pin-point haemorrhages on lungs, liver, kidneys and heart.
The amount of cytotoxin varied greatly in different preparations (Table 1). The
PBE was found to be best source of cytotoxin.
Purification of Salmonella cytotoxins. Differential salt fractionation studies revealed that
protein precipitated up to 50% and below salt saturation level did not contain much
cytotoxicity, while precipitates at 60% and 70% salt saturation levels had cytotoxic
activity equivalent to 128 and 256 cytotoxic units/mg of protein, respectively. There was
no appreciable increase in cytotoxic activity on further raising the salt concentration.
Therefore, proteins that precipitated up to 50% salt saturation level were discarded and
then the salt concentration was raised to 70% saturation level to obtain the precipitate
for further purification.
On gel filtration through Seralose 6 B, two peaks (Fig. 1) were observed. The
first one was eluted in 33 to 43 ml and the second between 45 to 67 ml of eluent. The
first peak contained no significant cytotoxic activity, while the second peak contents
displayed marked cytotoxic effects on cell lines as well as in rabbit skin. The second
peak contents yielded more than 6 protein bands on native PAGE.
Concentrated second peak contents from Seralose 6 B column, when eluted
through Mono-P column yielded 6 major peaks (Fig. 2) of which 4 peaks (i.e. 1, 3, 4 and
6) were found to be cytotoxic to variable degrees on Vero and MDBK cells. Peak I was
eluted unbound to column P yielded one thick and two thin protein bands on PAGE and
SDS PAGE (Fig 3); when this peak contents were eluted through Mono-P column
equilibrated with Tris-HCl buffer (0.37 M, pH, 8.6), it yielded single vytotoxic peak at
0.1 M NaCl gradient, while other peaks at 0.0 and 0.5 M salt gradient were non-toxic.
Cytotoxic eluent yielded single protein band at about 67 kDa level in SDS PAGE (Fig 3).
This cytotoxic peak was considered as purified cytotoxin I. In addition to properties
mentioned in Table 2, it also induced hyperaemia, depilation and scab formation away
PDF created with pdfFactory Pro trial version www.pdffactory.com

from intradermal inoculation site in rabbits on either regions (Fig 4), while in mice, whole
of the hind quarters and tail developed areas of frank necrosis (Fig. 5) after 5-7 days of
cytotoxin inoculation. In RLIL loop assay, the toxin induced very little serosanguinous
fluid accumulation yielding RLIL indices ranging from 0.2 to 0.5. However, marked
distention of stomach and urinary bladder in rabbits and mice was invariably observed.
The toxin induced hyperthermia by 1.5 to 2.5°C after 4 to 6 h of intraperitoneal
inoculation. Dose-dependent loss of appetite was evident in all the animals experimented
with toxin. The dead mice and rabbits had extensive haemorrhage in lungs (Fig 6) and
kidneys. Besides, pin-point haemorrhages and small focal areas of necrosis were also
observed in the liver and spleen. Peritoneal cavity, thoracic cavity, meninges and brain
had pin-point to large areas of haemorrhages and extensive congestion. Small intestines
were full of blood tinged contents.
Native PAGE analysis of 3rd cytotoxic peak revealed it as single protein of about
65 kDa and was designated as cytotoxin II. The characteristics of cytotoxin II have been
depicted in Table 2. Apart from these, it also induced diarrhoea in mice after
intraperitoneal inoculation and positive RLIL response with indices ranging from 0.7 to
1.85, depending on dose. Fluid in RLIL was blood tinged (Fig 7). However, other
lesions, viz., accumulation of fluid in stomach, distension of urinary bladder and
haemorrhages in visceral organs, as observed with cytotoxin I were absent.
The 4th and 6th cytotoxic peaks also yielded single protein band both in PAGE
and SDS- PAGE at a level of about 55 and 72 kDa (Fig 3,8) and were designated as
cytotxin III and IV, respectively.
Besides, characteristics shown in Table 2, Type III cytotoxin appeared to be the
least toxic with no effect in RLIL, rabbit skin or mouse foot pads. It could not induce
lethality in mice even at 2 mg dose level. It induced distension in tissue culture cells,
often with massive vacuolation, granulation and multinucleation and eventual death (Fig.
9).
PDF created with pdfFactory Pro trial version www.pdffactory.com

Type IV cytotoxin revealed phospholipase C activity on egg yolk agar and intense
necrotizing effect on rabbit skin (Fig. 10). In dead mice, except haemorrhagic fluid in
peritoneal cavity, no gross lesion was observed in brain, thoracic cavity or visceral
organs. It failed to induce fluid accumulation in RLIL but haemorrhages were present in
the inoculated loop. It invariably haemolysed the erythrocytes of sheep, rabbit, guinea pig
and human (O group) requiring 10, 8, 6 and 6 µg cytotoxin for complete haemolysis of
RBC (100 µl, 1 % v/v) within 6 h.
Serum Studies. Antisera to all the four cytotoxin formed precipitation lines in AGPT with
only homologous purified cytotoxin and crude preparations (PBE and CL). Also,
antisera to cytototxins- I, II and III produced line of precipitation with concentrated CFCF
(10 mg protein/ml). The antisera neutralized toxic effects of homologous toxin only and
no cross neutralization was observed in any of the bioassay models. The antisera except
antitoxin-IV, agglutinated the whole cell culture of S. Weltevreden.
The effect of heat, pH and proteolytic enzymes on cytotoxicity has been presented
in Table 2. These treatments had similar effect on antigenicity except that at pH 9.6
cytotoxicity was lost irreversibly while the treated preparations reacted with antitoxin
serum. None of the four purified cytotoxin preparations reacted with antiserum against
either O, H or common eneterobacteriaceae antigens. E Toxate analysis of purified
cytotoxic preparations revealed absence of endotoxin.
Discussion
Salmonella Weltevreden has frequently been isolated from ready-to-eat meat
products18,22 and has been shown to be a potent cytotoxin producer 12,22. In
the present investigation, crude preparations (CFCS, CL and PBE) of S. Weltevreden
induced similar types of CPE in Vero and MDBK cells and lesions in the skin and
internal organs of rabbit as reported earlier in case of cytotoxic preparations from a
number of other Salmonella serovars1,3,12,13,22..
PDF created with pdfFactory Pro trial version www.pdffactory.com

Partial purification of Salmonella cytotoxin was earlier achieved by means of gel
filtration13, membrane filtration and dialysis1,5,22. In this study, after partial
purification through salt precipitation, dialysis and gel filtration, further purification was
attempted by using ion-exchange chromatography. However, it did not yield promising
results as the proteins were either bound too tightly (DEAE-Sephadex) to be eluted with
buffer or eluted without any binding (CM-Sephadex) to the matrix. Therefore, partially
purified toxin was eluted through column P, which is a weak anion exchanger.
Interestingly, the preparation could be fractionated into 6 major peaks of which four
contained cytotoxic activity. This indicates that the cytotoxic proteins are highly charged.
The elution patterns of cytotoxins I, II, III and IV of S. Weltevreden through
Mono-P column and chromatofocusing of purified preprations indicated that their PI
values were approximately 9.0, 7.0, 5.6 and 3.0, respectively. SDS-PAGE profile
indicated their molecular weight to be approximately 67.65, 55 and 72 kDa, respectively.
Although these appears to be no report on PIs of Salmonella cytotoxin, closely related PIs
of shigalike toxin (SLT) I (7.0), SLT II (4.1-5.1) and SLT IIv (9.0) of E. coli26;
Klebsiella pneumoniae cytotoxin (KCT) II (6.8), KCT III (5.6) and KCT I (9.0)20 suggest
some structural similarity between Salmonella, E. coli and Klebsiella cytotoxins. It has
been claimed that antishiga toxin serum9, which neutralized SLT I of E. coli24 also
neutralized S. typhimurium cytotoxins but further studies did not confirm antigenic
relationship between Salmonella cytotoxins and other enterobacterial cytotoxins.
Formalin treatment of all 4 SCTs resulted in loss of their cytotoxic potential
similar to that observedduring earlier studies18,22 and can be ascribed to protein
denaturing activity of formalin.
Neither pepsin nor protease had any appreciable effect on any of the 4 SCTs but
it was observed that trypsin and chymotrypsin enhanced cytotoxicity of SCT I and II,
while these had adverse effect on SCT III and IV. Enhanced cytotoxicity of SCT I and II
in presence of enteric enzymes is significant as it might contribute to enteropathogenicity
PDF created with pdfFactory Pro trial version www.pdffactory.com

of the organism. Similarly, augmentative effect of toxicity of different cytotoxic
preparations of Salmonella1,22 and Klebsiella19 and of other enteropathogens18 are on
record. This has been ascribed to the disintigration of cytotoxin molecule in smaller
biologically active subunits or hyperactivation of the less active cytotoxin18. Further
studies in this regard are, however, warranted for precise understanding of the actual
mechanism underlying it.
Isolation of 4 distinct cytotoxic moieties from a single Salmonella strain is
significant as there seems to be no previous report in this regard. Singh22 claimed
purification of a single cytotoxin of S. Weltevreden and Ashkenazi et al.1 predicted the
presence of more than one type of cytotoxin in different strains of Salmonella on the basis
of varied responses of partially purified Salmonella cytotoxins to heat, pH and
proteolytic enzymes. However, it appears that they did not investigate the subject further
to confirm it. The presence of four distinct cytotoxic factors with different biological
activities might be responsible for the varied toxic effects i.e., Verocytotoxic, enterotoxic,
dermatotoxic and neurotoxic effects of crude/partially purified preparations of different
Salmonella cystotoxins as reported earlier1,3,13,22. For example, cytotoxic preparations
from certain Salmonella strains failing to induce dermonecrosis in rabbit skin might be
lacking cytotoxin IV. Cytotoxin I, which caused haemmorhages in the intestine along
with fever might be responsible for dysentery like symptoms, while cytotoxin II had
diarrhogenic activity; these symptoms are commonly observed in enteric fever15.
All the four types of cytotoxins were found to be immunogenic in rabbits.
Formation of single precipitation bands in AGPT only between specific cytotoxin and its
homologous antiserum indicates that they do not share antigenic determinant. Negative
results in E-Toxate assay revealed absence of endotoxin in purified cytotoxins. It was also
substantiated by serum neutralization studies. AGPT results by using antisera to O and H
factors and common enterobacteriaceae antigen further revealed absence of contaminants
like LPS or flagellar proteins in purified preparations.
PDF created with pdfFactory Pro trial version www.pdffactory.com

In conclusion, isolation of four distinct cytotoxins from a single Salmonella strain
with varied biological activities, physicochemical characteristics and antigenicity
suggested multiplicity of cytotoxic moieties. This offers possible explanation for
discrepancies that exist in literature with regard to characteristics of crude/partially
purified cytotoxin of different Salmonella strains. Obviously, it will contribute
significantly to the understanding pathogenesis of salmonellosis.
Acknowledgement
The authors are thankful to Indian Council of Agricultural Research, New Delhi,
for providing funds under the National Fellow Scheme on ‘Salmonella enterotoxins and
Council of Scientific and Industrial Research, New Delhi, for awarding the Senior
Research Fellowship to the first author.
PDF created with pdfFactory Pro trial version www.pdffactory.com

References
1. Ashkenazi, S., T.G. Cleary, B.E. Murray, A. Wanger and L.K: Pickering.
Quantitative analysis and partial characterization of cytotoxin production by
Salmonella strains. Infect Immun. 56 (1988) 3089-3094
2. D'Aoust, J.Y. : Salmonella, In : Doyle M.D. (ed) Food borne Bacterial Pathogens,
Marul Dekker Ind; pp. 332-362 (1989)
3. Gonzalez, E.A., J. Blanco, M. Blanco, J.I. Garabal, and M.P. Alonso : Enterotoxic,
cytotoxic, necrotoxic and lethal activities in cell free extracts of Salmonella
strains isolated from humans. Zbl Bakt. 271 (1989) 281-292
4. Hsu, H.S. : Pathogenesis and immunity in murine salmonellosis. Microbiol. Rev.
53 (1989) 390-409
5. Hudson, L. and F.C. Hay : Practical Immunology, 2nd Edn. Blackwell, Oxford. pp.
117-120 (1980)
6. Jacks, T. M. and B. J. Wu : Biochemical properties of Escherichia coli low molecular
weight heat stable eneterotoxin. Infect Immun. 9 (1974) 324-347
7. Joshi, R.K. and V.D. Sharma : Detection of thermolabilie phospholipase-C in the cell
free supernatants of Salmonella. Indian J. Comp. Microbiol. Immunol. Infect.
Dis. 12 (1991) 153-157
8. Kety, I. : Toxins as virulence factors of bacterial enteric pathogens. Acta
Microbiologica Hungarica. 32 (1985) 279-304.
9.Lallier, R. and B. Lariviere : Effect of different treatment on the activity of heat labile
enterotoxin of Escherichia coli. Canadian J. Comp. Med. 42 (1978) 214-218
10. Lax, A.J., P.A. Barrow, P.W. Jones and T.S. Wallis : Current perspectives in
salmonellosis. British Vet. J. 151 (1995) 351-377
11. Lowry, O. H., N. J. Rosenbrough, A. L. Farr and R. J. Randalt: Protein measurement
with Folin phenol reagent. J. Biol. Chem. 13 91951) 265-377
PDF created with pdfFactory Pro trial version www.pdffactory.com

12. Malik P., V.D. Sharma, and R. Chandra : Cytotoxigenicity in Salmonella Serovars.
Indian J. Exp. Biol. 33 (1995) 177-181
13. Malik, P., V. D. Sharma, and D. C. Thapliyal: Partial purification and
characterisation of Salmonella cytotoxin. Vet. Microbiol. 49 (1996) 11-19
14. Negroni, G. : Tissue culture techniques. In : Harris, R.J. (ed) Techniques in
Experimental Virology. Academic Press, London. pp 328-358 (1964)
15. Parker, M.T. : Enteric infections : typhoid and paratyphoid fever. In : Smith, G.R. and
Easman, C.S.F. (eds) Topley and Wilson's Principales of Bacteriology, Virology
and Immunity. 8th Ed. (3) Edwards and Arnold, London. pp 423-446 (1990)
16. Saoji, A.M. and P.M. Khare : Polyacrylamide gel electrophoresis (PAGE) : Disc
electrophoresis. Indian J. Pathol. Microbiol. 28 (1985) 179-186
17. Sharma, V.D., S.P. Singh and A. Taku : Purification of Salmonella stanley enterotoxin
and its immunology and dermatotoxicity. Indian J. Exp. Biol. 30 (1992) 23-25
18. Sharma, V.D., D.C. Thapliyal, S.P. Singh and P. Malik : Cytotonic and cytotoxic
enterotoxins of enterobacteria. Indian J. Microbiol. 32 (1992) 327-356
19. Singh, B.R. : Purification and molecular characterization of Klebsiella pneumoniae
cytotoxins. Ph.D. Thesis, G.B. Pant Univ. of Agric. & Tech., Pantnagar. pp 243
(1997)
20. Singh, B.R. and S.B. Kulshreshtha : Preliminary examinations on the
enterotixigenicity of isolates of Klebsiella pneumoniae from seafoods. Int. J.
Food Microbiol. 16 (1992) 349-352
21. Singh, B.R. and A.K. Tiwari : Toxigenicity of Edwardsiella tarda isolates of fish and
pig origin in experimental models. Indian J. Exp. Biol. 33 (1996) 1254-1256
22. Singh, Y. : Study on Salmonella cytotoxin. Ph.D. Thesis, G.B. Pant Univ. of Agric. &
Tech., Pantnagar. pp 173 (1995)
PDF created with pdfFactory Pro trial version www.pdffactory.com

23. Spiers, J.I., S. Stavric and J. Konowalchuk : Assay of Escherichia coli heat labile
enterotoxin with Vero cells. Infect. Immun. 16 (1977) 617-622
24. Strockbine, N.A., L.R.M. Marques, J.W. Newland, H.W.D. Smith, R.K. Holmes and
A.D.O'Brien : Two toxin converting phages from Excherichia coli 0157:H7
strain 933 encode antigenically distinct toxin with similar biological activities.
Infect. Immun. 53 (1986) 135-140
25. Wallis, T.S., W.G. Starky, J. Stephen, S.J. Hadden, M.P. Osborne, and D.C.A. Candy
: Enterotoxin production by Salmonella typhimurium strains of different
virulence. J. Med. Microbiol. 21 (1986) 19-23
26. Weinstein, D.L., M.P. Jackson, L.P. Parera and J.E. Samuel : In vivo formation of
hybrid toxin comprising shigatoxin and shigalike toxins and role of B subunits in
localization and cytotoxic activity. Infect. Immun. 57 (1989) 3743-3750
27. Wray, C. : Salmonellosis : A hundred years old and still going strong. British Vet. J.
151 (1995) 339-341
PDF created with pdfFactory Pro trial version www.pdffactory.com

Legends for Figures
1. Cytotoxin elution pattern of polymyxin B extract (PBE) of S. Weltevreden
through Seralose 6 B column (2 x 50 cm) eluted with Tris HCl buffer (pH 7.2,
0.037 M).
2. Cytotoxin elution pattern through Mono-P column (Pharmacia) loaded with 2.5
mg of second peak from Seralose 6 B column, eluted with Tris HCl (0.037 M, pH
7.2) NaCl gradient and finally with 0.025 M acetic acid.
3. Sodium dodecyl sulphate polyacrylamide gel electrophoresis profile of different
purified and semipurified preparations of S. Weltevreden cytotoxins.
Lane 1: Cytotoxic peak eluted through Seralose 6B column.
Lane 2: Molecular weight markers.
Lane 3: Cytotoxin I.
Lane 4: Cytotoxin I on re-purification at pH 8.6
Lane 5: Cytotoxin III.
4. Depilation of hair and scab formation at wither region after 5 days of intradermal
inoculation of cytotoxin I (100 µg) at andominal region.
5. Dermonecrosis on hind quarters and tail of mice (T) after 5 days of inoculation of
cytotoxin I (100 µg) compared with healthy control (C ).
6. Haemorrhage in lung of mice died after 18 h of intraperitoneal inoculation of
cytotoxin I (100 µg).
7. Accumulation of haemorrhagic fluid in rabbit ligated ileal loop1 inoculated with
100 µg of cytotoxin II, clear straw-coloured fluid in loop2 and 4 inoculated with
50 µg of Cholera toxin.
PDF created with pdfFactory Pro trial version www.pdffactory.com
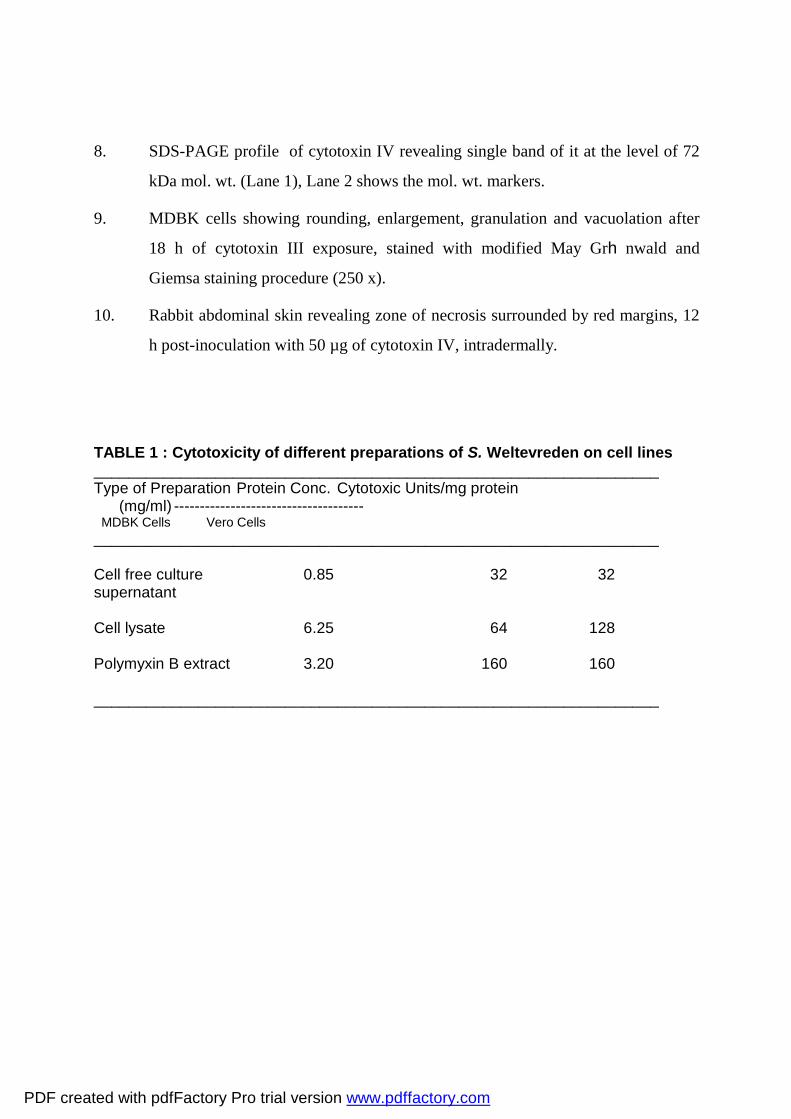
8. SDS-PAGE profile of cytotoxin IV revealing single band of it at the level of 72
kDa mol. wt. (Lane 1), Lane 2 shows the mol. wt. markers.
9. MDBK cells showing rounding, enlargement, granulation and vacuolation after
18 h of cytotoxin III exposure, stained with modified May Grhnwald and
Giemsa staining procedure (250 x).
10. Rabbit abdominal skin revealing zone of necrosis surrounded by red margins, 12
h post-inoculation with 50 µg of cytotoxin IV, intradermally.
TABLE 1 : Cytotoxicity of different preparations of S. Weltevreden on cell lines _________________________________________________________________ Type of Preparation Protein Conc. Cytotoxic Units/mg protein (mg/ml) ------------------------------------- MDBK Cells Vero Cells _________________________________________________________________ Cell free culture 0.85 32 32 supernatant Cell lysate 6.25 64 128 Polymyxin B extract 3.20 160 160 _________________________________________________________________
PDF created with pdfFactory Pro trial version www.pdffactory.com

TABLE 2 : Biological and physico-chemical characteristics of four S. Weltevreden cytotoxins ______________________________________________________________________________________________________________________________________________ Characteristics Type of Cytotoxin I II III IV Binding to column P at pH 7.2,0.037M No Yes Yes Yes Salt gradient needed for elution No 0.5M 1.5M >5.0M Effect on Vero/MDBK cells Rounding, detachment Rounding,degeneration, Vacuolation, syncytia- Rapid degradation and lysis detachment, lysis formation, multinucleation Vero cytotoxic units/mg of protein 640 320 1280 1280 Effect on rabbit skin Delayed (72h) hyperaemia, Hyperaemia and oedema No visible effect White zone of necrosis oedema and pus formation in 18 h and detachment with red margins of epidermis in 72 h Effect in mouse food pad Delayed oedema (72 h) Hyperaemia and oedema No effect Necrosis of foot pad Lethality to mice (LD50) Yes (120 µg) Yes (1000 µg) ND Yes (150 µg) Haemolytic action on rabbit, sheep and No No No Yes human erythrocytes PI values 9.0 7.0 5.6 3.0 Inactivation of cytotoxicity with heat treatment 121°C, 1 min 100°C, 30 min 100°C, 30 min 90°C, 30 min Activity in pH range of 5.0-8.0 5.6-8.4 4.8-8.2 2.5-7.5 Irrerversible inactivation at pH <4.6 and >9.6 <4.6 and >9.2 <4.0 and >9.2 <2.0 and >8.6 Effect of trypsin (0.1%, 6 h) 50% increase in 40% increase in 50% loss in activity 90% loss in cytotoxicity cytotoxicity cytotoxicity Effect of chymotrypsin (0.1%, 6 h) 20% increase in 10% increase in 50% loss in activity 76% loss of cytotoxicity cytotoxicity cytotoxicity Effect of 0.05% formalin (12 h) 100% loss of 100% loss of 100% loss of 100% loss of cytotoxicity cytotoxicity cytotoxicity cytotoxicity ______________________________________________________________________________________________________________________________________________ ND : could not be determined because mouse survived after inoculation of >1.0 mg cytotoxin/mouse. Note : Pepsin and protease had no effect on cytotoxicity of any of the 4 cytotoxins
PDF created with pdfFactory Pro trial version www.pdffactory.com


Related Documents











