Journal of Experimental Botany, Page 1 of 12 doi:10.1093/jxb/erm149 This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details) RESEARCH PAPER Depression of sink activity precedes the inhibition of biomass production in tomato plants subjected to potassium deficiency stress S. Kanai 1 , K. Ohkura 2 , J. J. Adu-Gyamfi 3 , P. K. Mohapatra 4 , N. T. Nguyen 1 , H. Saneoka 1 and K. Fujita 1, * 1 Graduate School of Biosphere Science, Hiroshima University, 1-4-4 Kagamiyama, Higashi-hiroshima, 739-8528, Japan 2 Faculty of Environmental and Information Sciences, Yokkaichi University, 1200 Kayoucho, Yokkaichi, 512-8512, Japan 3 International Atomic Energy Agency (IAEA), Wagramer Strasse 5, A-400, Vienna, Austria 4 School of Life Science, Sambalpur University, Jyoti vihar, Sambalpur 768019, India Received 20 February 2007; Revised 30 May 2007; Accepted 1 June 2007 Abstract Tomato [Solanum lycopersicum (formerly Lycopersi- con esculentum) L. cv. Momotarou] plants were grown hydroponically inside the greenhouse of Hiroshima University, Japan. The adverse effects of potassium (K) deficiency stress on the source–sink relationship during the early reproductive period was examined by withdrawing K from the rooting medium for a period of 21 d. Fruits and stem were the major sink organs for the carbon assimilates from the source. A simple non- destructive micro-morphometric technique was used to measure growth of these organs. The effect of K deficiency was studied on the apparent photosynthe- sis (source activity), leaf area, partitioning 13 C, sugar concentration, K content, and fruit and stem diameters of the plant. Compared with the control, K deficiency treatment severely decreased biomass of all organs. The treatment also depressed leaf photosynthesis and transport of 13 C assimilates, but the impact of stress on these activities became evident only after fruit and stem diameter expansions were down-regulated. These results suggested that K deficiency diminished sink activity in tomato plants prior to its effect on the source activity because of a direct effect on the water status of the former. The lack of demand in growth led to the accumulation of sugars in leaves and concom- itant fall in photosynthetic activity. Since accumulation of K and sugars in the fruit was not affected, low K levels of the growing medium might not have affected the fruit quality. The micro-morphometric technique can be used as a reliable tool for monitoring K defi- ciency during fruiting of tomato. K deficiency directly hindered assimilate partitioning, and the symptoms were considered more detrimental compared with P deficiency. Key words: Fruit and stem diameter, partitioning, potassium, tomato, micro-morphometry. Introduction The potassium (K) requirement of greenhouse tomatoes is high for vegetative growth (Wall, 1940; Lucas, 1968), fruit production (Besford and Maw, 1975), and fruit quality (Winsor, 1968; Trudel and Ozbun, 1971). Low K levels of the nutrient medium limit assimilation of the element into plant parts and retard plant growth, flower development, and fruit set (Besford and Maw, 1975). There is evidence that K exerts a direct effect on the par- titioning of dry matter to the fruits and roots, and the growth of these organs is inhibited at low K (Haeder and Mengel, 1972). Also, the quality of the fruit changes according to the availability of K in the growth medium (Davies and Winsor, 1967; Winsor, 1968). During the reproductive phase of growth, fruits are the strongest sink for both carbon assimilates and K. In some determinate tomatoes, the demand for K during rapid fruit growth is * To whom correspondence should be addressed. E-mail: [email protected] ª 2007 The Author(s). This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. Journal of Experimental Botany Advance Access published July 13, 2007 by guest on April 30, 2015 http://jxb.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Experimental Botany, Page 1 of 12
doi:10.1093/jxb/erm149This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details)
RESEARCH PAPER
Depression of sink activity precedes the inhibitionof biomass production in tomato plants subjected topotassium deficiency stress
S. Kanai1, K. Ohkura2, J. J. Adu-Gyamfi3, P. K. Mohapatra4, N. T. Nguyen1, H. Saneoka1 and K. Fujita1,*
1 Graduate School of Biosphere Science, Hiroshima University, 1-4-4 Kagamiyama, Higashi-hiroshima,739-8528, Japan2 Faculty of Environmental and Information Sciences, Yokkaichi University, 1200 Kayoucho, Yokkaichi,512-8512, Japan3 International Atomic Energy Agency (IAEA), Wagramer Strasse 5, A-400, Vienna, Austria4 School of Life Science, Sambalpur University, Jyoti vihar, Sambalpur 768019, India
Received 20 February 2007; Revised 30 May 2007; Accepted 1 June 2007
Abstract
Tomato [Solanum lycopersicum (formerly Lycopersi-
con esculentum) L. cv. Momotarou] plants were grown
hydroponically inside the greenhouse of Hiroshima
University, Japan. The adverse effects of potassium
(K) deficiency stress on the source–sink relationship
during the early reproductive period was examined by
withdrawing K from the rooting medium for a period of
21 d. Fruits and stem were the major sink organs for
the carbon assimilates from the source. A simple non-
destructive micro-morphometric technique was used
to measure growth of these organs. The effect of K
deficiency was studied on the apparent photosynthe-
sis (source activity), leaf area, partitioning 13C, sugar
concentration, K content, and fruit and stem diameters
of the plant. Compared with the control, K deficiency
treatment severely decreased biomass of all organs.
The treatment also depressed leaf photosynthesis and
transport of 13C assimilates, but the impact of stress
on these activities became evident only after fruit
and stem diameter expansions were down-regulated.
These results suggested that K deficiency diminished
sink activity in tomato plants prior to its effect on the
source activity because of a direct effect on the water
status of the former. The lack of demand in growth led
to the accumulation of sugars in leaves and concom-
itant fall in photosynthetic activity. Since accumulation
of K and sugars in the fruit was not affected, low K
levels of the growing medium might not have affected
the fruit quality. The micro-morphometric technique
can be used as a reliable tool for monitoring K defi-
ciency during fruiting of tomato. K deficiency directly
hindered assimilate partitioning, and the symptoms
were considered more detrimental compared with P
deficiency.
Key words: Fruit and stem diameter, partitioning, potassium,
tomato, micro-morphometry.
Introduction
The potassium (K) requirement of greenhouse tomatoes ishigh for vegetative growth (Wall, 1940; Lucas, 1968),fruit production (Besford and Maw, 1975), and fruitquality (Winsor, 1968; Trudel and Ozbun, 1971). Low Klevels of the nutrient medium limit assimilation of theelement into plant parts and retard plant growth, flowerdevelopment, and fruit set (Besford and Maw, 1975).There is evidence that K exerts a direct effect on the par-titioning of dry matter to the fruits and roots, and thegrowth of these organs is inhibited at low K (Haeder andMengel, 1972). Also, the quality of the fruit changesaccording to the availability of K in the growth medium(Davies and Winsor, 1967; Winsor, 1968). During thereproductive phase of growth, fruits are the strongest sinkfor both carbon assimilates and K. In some determinatetomatoes, the demand for K during rapid fruit growth is
* To whom correspondence should be addressed. E-mail: [email protected]
ª 2007 The Author(s).This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) whichpermits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Experimental Botany Advance Access published July 13, 2007 by guest on A
pril 30, 2015http://jxb.oxfordjournals.org/
Dow
nloaded from

above the uptake capacity such that leaf K is remobilized,resulting in foliar deficiency of the element (Widders andLorenz, 1979). Because of remobilization and recyclingfrom old parts to new organs (Pujos and Morard, 1997;Peuke et al., 2002), visible symptoms of injury do notappear on the growing sinks immediately in K-deficientnutrient medium (Mengel and Kirkby, 1987). Visualsymptoms such as leaf necrosis and chlorosis appearmuch later when plants are exposed to a long period of thestress (Besford, 1978a, b; Pujos and Morard, 1997). It isimportant that farmers recognize the incidence of Kdeficiency during the early production stage and restorenormal growth of the crop by supplementation of K in theculture medium.Plant cells have a very high requirement for K for
photosynthesis, enzyme activation, protein synthesis, cellturgor, and ion homeostasis (Marschner, 1995). Low Klevels in the growing medium can disturb these processesand destabilize the source–sink relationship. In tomato, theadverse effect of K deficiency was documented on leafphotosynthesis (Zhao et al., 2001), but the effect was notdue to impairment of turgor-induced regulation of stoma-tal conductance (Behboudian and Anderson, 1990). Someother authors have shown that mild K deficiency sup-presses assimilate translocation without affecting photo-synthesis at the source (Hart, 1969, 1970; Mengel andViro, 1974). On the other hand, the effects of K def-iciency on the sink activity are mired in controversies. It isreported that K deficiency reduces photosynthate trans-location (Huber, 1984) without affecting the metabolismat the sink side (Mengel, 1980; Beringer and Haeder,1981). Hart (1969) observed that the reduction in assim-ilate translocation by K deficiency in sugarcane is causedneither by death of the phloem nor by diminished sinkgrowth. Conversely, Tsuno and Fujise (1965) found thatK deficiency affects photosynthate translocation throughreduced growth of storage roots, which is a predominantsink in sweet potato. Geiger and Conti (1983) reportedthat translocation and partitioning of dry matter remainindependent over a wide range of K supply in the growthmedium. These contradictions led to the suggestion thatthe effect of K deficiency on plant growth remains elusive,and it is necessary to re-examine the source–sink relation-ship of the plant under the influence of the stress (Roitsch,1999). Biomass production is reduced due to impairmentof sink activity by phosphorus (P) deficiency in tomato(Fujita et al., 2003b) and by salt stress in Japanesepersimmon (Fujita et al., 2003a) and tobacco (Moghaiebet al., 2006). Also, there are examples where sourceactivity is depressed earlier than that of the sink under theinfluence of stresses such as drought (Berman and Dejong,1996; Sowder et al., 1997; Van den Boogaard et al.,1997; Escobar-Gutierrez et al., 1998) and nitrogen de-ficiency (Fujita et al., 2004). However, the effects of Kdeficiency on the source–sink relationship of tomato plants
have not been studied so far. Additionally, the incidenceof K deficiency at the early production stage in tomato isdetrimental to fruit growth (Pujos and Morard, 1997).Therefore, a reliable technique is necessary to assess theability of the plant to encounter nutritional disorders andreplenish the stock solution. In the present experiment, theobjective was to monitor the effect of K deficiency on thediurnal changes in stem and fruit diameters of tomatoplants while simultaneously recording the impact of thestress on activities such as leaf photosynthesis and trans-port of 13C assimilates, and the carbohydrate status of theplant. It is necessary to identify the resource managementtechniques on the part of the plant at suboptimal K levelsfor the benefit of farmers.
Materials and methods
Experiment 1
Plant material and culture: Tomato (Solanum lycopersicum (for-merly Lycopersicon esculentum) L. cv. Momotarou] plants weregrown in pots inside the greenhouse of the Graduate School ofBiosphere Science, Hiroshima University, Japan. Each pot (70.0 l)was filled with nutrient solutions, consisting of N [Ca(NO3)2.4H2O]3.57 mM, P (NaH2PO4.2H2O) 0.32 mM, K (K2SO4/KCl 1:1)1.02 mM, Ca (CaCl2.H2O) 0.75 mM, Mg (MgSO4.7H2O)0.82 mM, Fe (Fe+3-EDTA) 0.02 mM, Mn (MnSO4.4H2O)3.64 lM, B (H3BO3) 0.05 mM, Zn (ZnSO4.7H2O) 0.15 lM, Cu(CuSO4.5H2O) 0.16 lM, Mo (NaMoO4.2H2O) 0.1 lM, Co(CoSO4.7H2O) 0.17 lM. There were seven pots; each pot containedsix plants. At the fruiting stage (74 d old), K2SO4/KCl was totallyomitted from the nutrient medium in three pots, and this treatmentwas continued for 21 d. The plants in the control conditionreceived full nutrition. The plants were grown under a daily lightperiod of 14 h (05.00 h to 19.00 h) and maximum irradiance of800 lmol m�2 h�1. The maximum and minimum temperatureswere 32 �C and 20 �C, respectively.
Measurement of biomass production: Plants from both control andK deficiency conditions were harvested at 0, 7, 14, and 21 d aftertreatment (DAT) in three replicates. The plant was separated intoleaves, stem, fruits, and roots. The plant parts were dried in an air-forced oven at 70 �C for 7 d for the estimation of dry weight. Thedry plant parts were ground into a powder with a vibrating samplemill (Model T1-100, Heiko Co. Ltd, Fukushima, Japan) andaliquots were taken for analysis of K, sugar concentration, and 13C.
Measurement of minerals: An aliquot of the plant material wasdigested with an acid mixture (HNO3:HClO4, 3:1, v/v). The Kconcentration in the digest was determined by flame photometry(ANA-135, Tokyo, Japan).
Measurement of photosynthesis, transpiration, and stomatalconductance: Photosynthetic rate, transpiration, and stomatal con-ductance of the leaves just below the first fruiting truss weremeasured with a portable infrared gas analyser (Model L1-6400, Li-Cor, Lincoln, NE, USA). The leaf chamber was the open type andmeasurements were made at 11:00 h each day in both control and Kdeficiency treatment plants. The photosynthetically active radiationduring measurement was >1000 lmol m�2 s�1, and observationswere recorded after the plant reached steady-state photosynthesis.
2 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

All measurements were recorded three times and averaged on eachoccasion of sampling.
Measurement of stem and fruit diameter: Changes in stem and fruitdiameter were continuously recorded in both control andK-deficient plants during the period of treatment with a shrinkage-type micro-displacement detector (Fujita et al., 2003b). The sensorswere fastened to the stem at 15 cm above the basal end of the stemor a growing fruit, and connected to the power system and datalogger. The sensors were connected to a computerized dataacquisition system (NEC, Sanei Kogyo Co. Ltd, Tokyo, Japan). Allmeasurements were recorded twice, and the pattern of response wasfound to be similar in all.
13CO2 feeding: 13CO2 feedings were given to the leaf immediatelybelow the first fruiting node on days 7 and 14 after K deficiencytreatment in both control and treated conditions. The leaf wasenclosed in a transparent plastic bag containing barium carbonate(Ba13CO3), packed in a plastic cylinder. The bag was filled with13CO2 from barium carbonate. 13CO2 was generated by addition of40% lactic acid (v/v). The leaf was allowed to assimilate 13CO2 for1.5 h on days 7 and 14. The plants were harvested 36 h after feedingby separating into 13CO2-fed leaf, other leaves, fruits (first, second,and third truss), stem, and roots. The freeze-dried plant parts wereground into a powder for measurement of 13C abundance.
13C analyses: The 13C abundance in the powdered plant samplewas determined with a mass spectrometer (model Delta plus,Finnigan Co., San Jose, CA, USA) (Nobuyasu et al., 2003). The13C atom% excess in the plant sample was calculated as thedifference in 13C atom% between 13C-fed and unfed samples.The amount of labelled C in the plant sample was calculated usingthe equation shown below.
13C amount in the plant part =13C atom % excess in the sample
13C atom % of 13CO2 administered3
amount of C in the plant parts
The element analyser facilitated in the mass spectrometerdetermined the concentration of C. The concentration obtained wasmultiplied by the weight of the plant part to determine the amountof total C in the sample. The partitioning of isotope was calculatedas the percentage of the total label in a plant part relative to that inthe plant part of the fed leaf.
13C partitioning rate ð%Þ =13C amount in the plant part
13C amount in the whole plant3 100
The export rate of the isotope was calculated by the equationgiven below.
13C export rate ð%Þ =13C amount in the whole plant� 13 C amount in the fed fix leaf
13C amount in the whole plant3 100
Measurement of sugar content: Aliquots of the powdered plantparts were boiled with 80% (v/v) aqueous ethanol three times forthe extraction of sugars. The extracts were pooled in a volumetricflask and the flask was filled up to the mark with distilled water.The sugar content in the ethanol-soluble extract was determinedusing the anthrone reagent, according to the method of Yemm andWillis (1954).
Experiment 2
Plant material and biomass production: Tomato (S. lycopersicumL. cv. Momotarou) plants were grown according to Experiment 1.At the flowering stage (53 d), K was withdrawn from the nutrientmedium, and this treatment was continued for 9 d. The plants in thecontrol received full nutrition. The plants were grown under anatural light period of 14 h (05.00 h to 19.00 h) and maximumirradiance of 800 lmol m�2 h�1. The maximum and minimumtemperatures were 30 �C and 18 �C, respectively. Biomass pro-duction of plants was determined according to Experiment 1.Tomato was harvested at 0, 3, and 7 DAT in eight replicates.
Measurement of stem diameter: Changes in stem diameter werecontinuously recorded in both control and K-deficient plants duringthe period of treatment according to Experiment 1. All measure-ments were recorded four times, and the pattern of response wasfound to be similar in all.
14CO2 feeding and analyses: 14CO2 feedings were given to the fifthleaf from the bottom of a plant on days 3 and 7 after K deficiencytreatment in both control and treated conditions. The leaf wasenclosed in a transparent plastic bag with a plastic cylinder includingNaH14CO3 solution (185 kBq per treatment). 14CO2 was generatedby addition of 40% lactic acid (v/v). The leaf was allowed to assim-ilate 14CO2 for 1.5 h under natural light conditions. The plants wereharvested 24 h after feeding. These plants were separated into14CO2-fed leaf, other leaves, stem, roots, and shoot apex. The plantparts were placed in an air-forced drought oven at 70 �C for 7 d. Allsamples were ground into a powder by sample mill (model T1-100,Heiko Co. Ltd., Fukushima, Japan). The specific activity of 14C ineach plant part was determined using a liquid scintillation counter(model LSC-5100, Aloka Co., Tokyo, Japan).
Results
Experiment 1
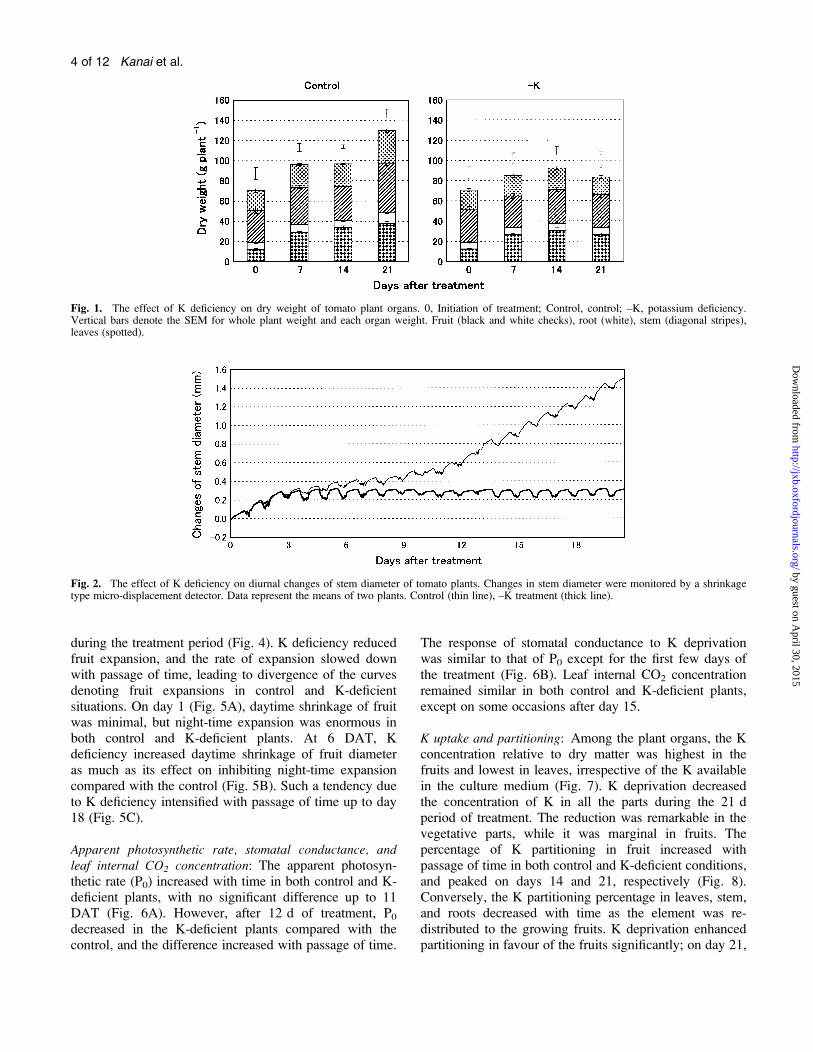
Dry mass accumulation: Careful examination of the dataseems to indicate that there may be a statistically signif-icant difference between the dry weight of the plants fromthe two treatments at either 7 d or 14 d (Fig. 1). TheK-deficient plants are lower in weight at these points andthey appeared to be statistically different. However, duringthe last 7 d, dry weights in the leaves, stem, and rootsdecreased to 53, 66, and 72%, respectively, relative to thecontrol.
Stem diameter: The diameter of the stem in the controlexhibited daytime shrinkage and night-time expansion, andincreased temporally up to 21 DAT (Fig. 2). Similar rhyth-mic shrinkage and expansion in the stem were observedin spite of K deficiency. K deficiency did not influencestem expansion during the first 3 d of treatment, but thestem did not expand in diameter thereafter (Fig. 2). Kdeprivation treatment initially increased daytime shrinkage.The effect on night-time expansion was also depressed byK deficiency at 15 DAT and subsequently (Fig. 3E).
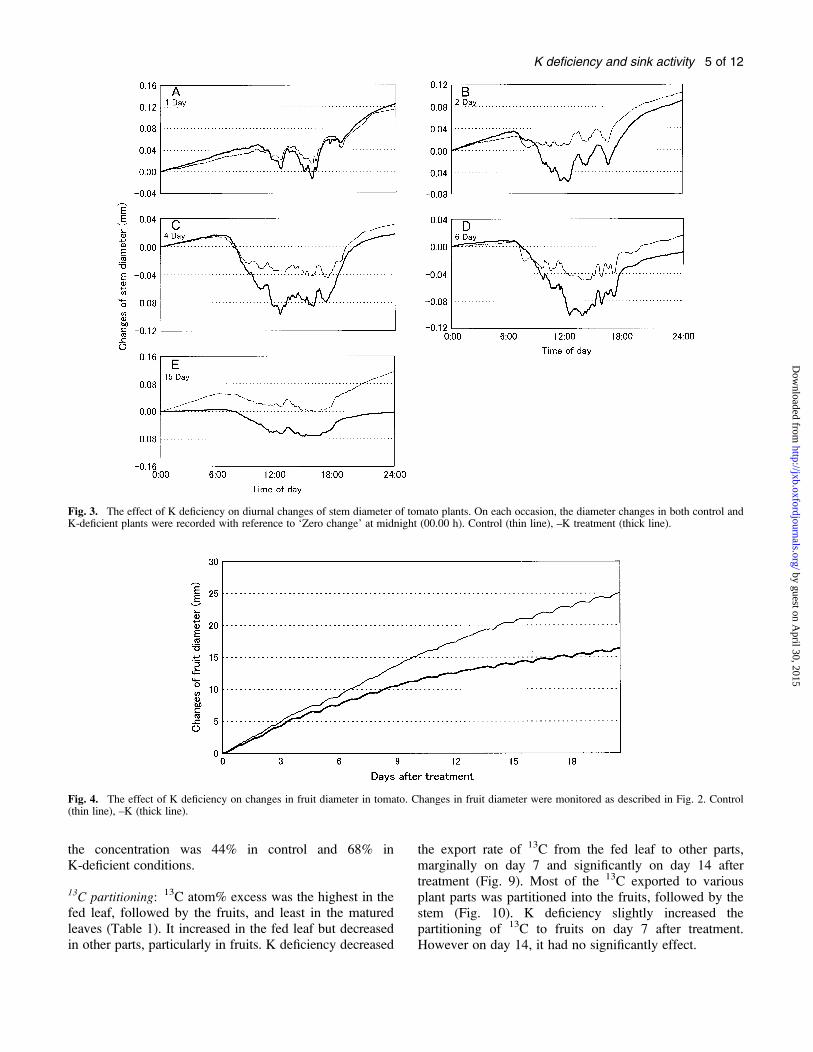
Fruit diameter: The diameter of fruit increased with thepassage of time in both control and K-deficient plants
K deficiency and sink activity 3 of 12
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
during the treatment period (Fig. 4). K deficiency reducedfruit expansion, and the rate of expansion slowed downwith passage of time, leading to divergence of the curvesdenoting fruit expansions in control and K-deficientsituations. On day 1 (Fig. 5A), daytime shrinkage of fruitwas minimal, but night-time expansion was enormous inboth control and K-deficient plants. At 6 DAT, Kdeficiency increased daytime shrinkage of fruit diameteras much as its effect on inhibiting night-time expansioncompared with the control (Fig. 5B). Such a tendency dueto K deficiency intensified with passage of time up to day18 (Fig. 5C).
Apparent photosynthetic rate, stomatal conductance, andleaf internal CO2 concentration: The apparent photosyn-thetic rate (P0) increased with time in both control and K-deficient plants, with no significant difference up to 11DAT (Fig. 6A). However, after 12 d of treatment, P0decreased in the K-deficient plants compared with thecontrol, and the difference increased with passage of time.
The response of stomatal conductance to K deprivationwas similar to that of P0 except for the first few days ofthe treatment (Fig. 6B). Leaf internal CO2 concentrationremained similar in both control and K-deficient plants,except on some occasions after day 15.
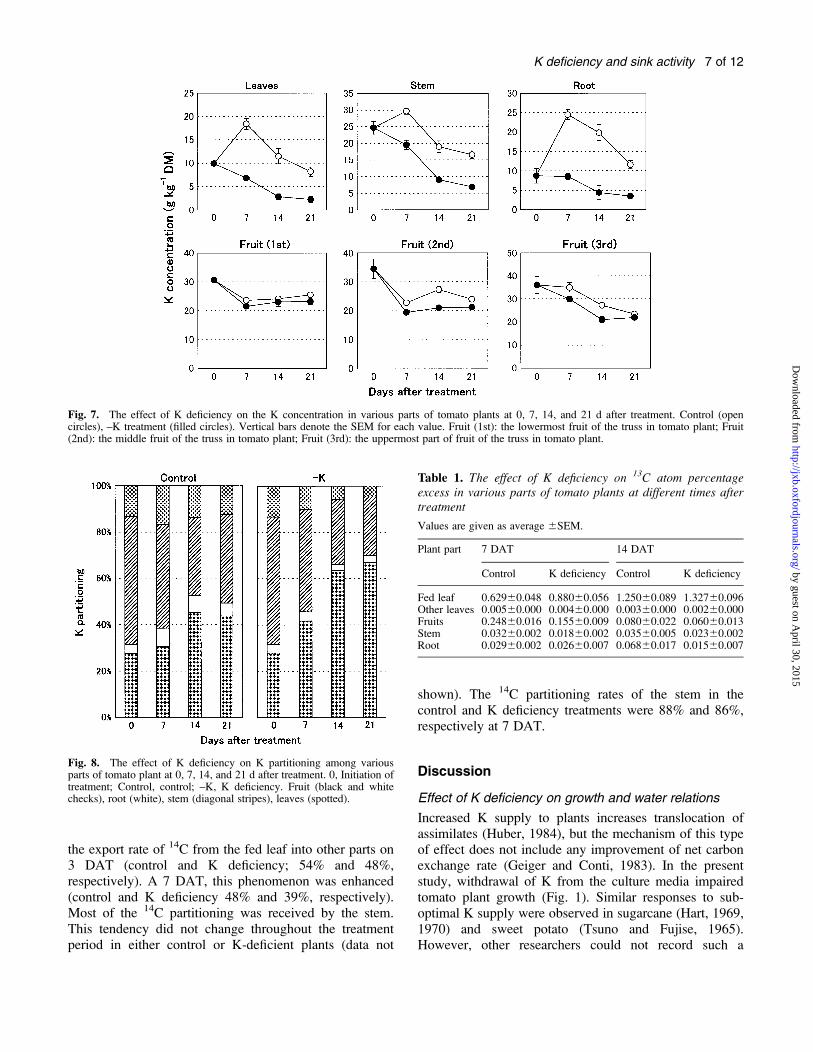
K uptake and partitioning: Among the plant organs, the Kconcentration relative to dry matter was highest in thefruits and lowest in leaves, irrespective of the K availablein the culture medium (Fig. 7). K deprivation decreasedthe concentration of K in all the parts during the 21 dperiod of treatment. The reduction was remarkable in thevegetative parts, while it was marginal in fruits. Thepercentage of K partitioning in fruit increased withpassage of time in both control and K-deficient conditions,and peaked on days 14 and 21, respectively (Fig. 8).Conversely, the K partitioning percentage in leaves, stem,and roots decreased with time as the element was re-distributed to the growing fruits. K deprivation enhancedpartitioning in favour of the fruits significantly; on day 21,
Fig. 1. The effect of K deficiency on dry weight of tomato plant organs. 0, Initiation of treatment; Control, control; –K, potassium deficiency.Vertical bars denote the SEM for whole plant weight and each organ weight. Fruit (black and white checks), root (white), stem (diagonal stripes),leaves (spotted).
Fig. 2. The effect of K deficiency on diurnal changes of stem diameter of tomato plants. Changes in stem diameter were monitored by a shrinkagetype micro-displacement detector. Data represent the means of two plants. Control (thin line), –K treatment (thick line).
4 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
the concentration was 44% in control and 68% inK-deficient conditions.
13C partitioning: 13C atom% excess was the highest in thefed leaf, followed by the fruits, and least in the maturedleaves (Table 1). It increased in the fed leaf but decreasedin other parts, particularly in fruits. K deficiency decreased
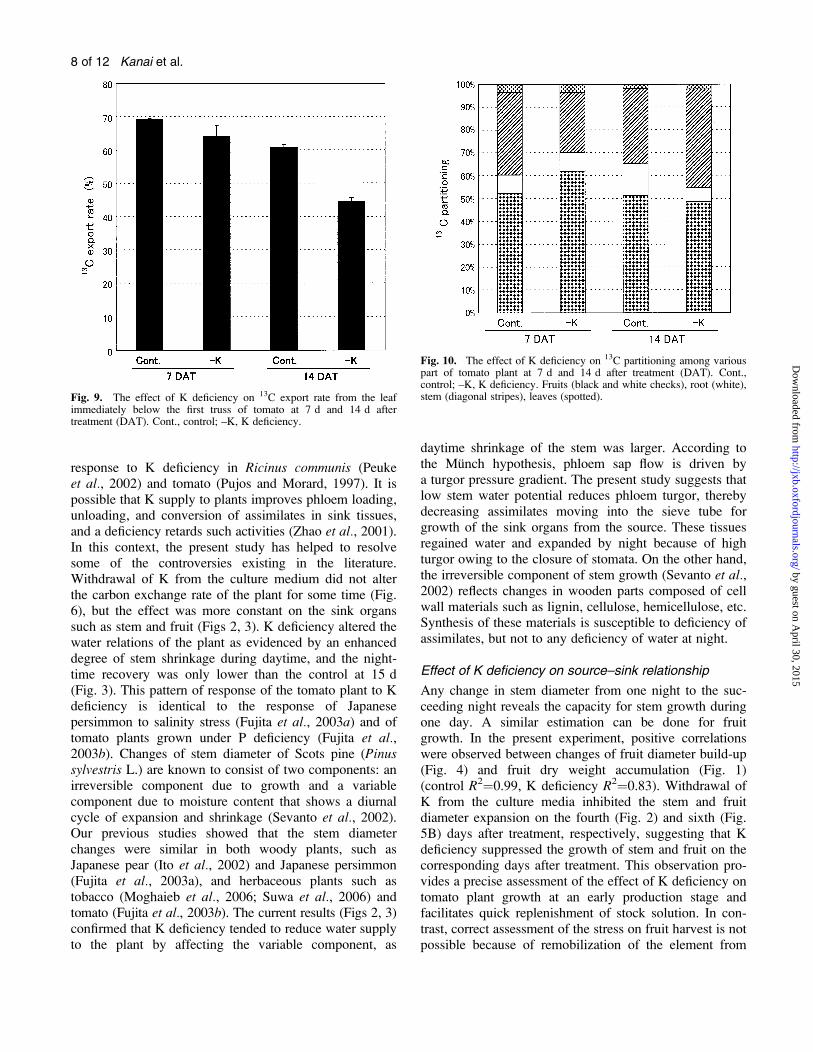
the export rate of 13C from the fed leaf to other parts,marginally on day 7 and significantly on day 14 aftertreatment (Fig. 9). Most of the 13C exported to variousplant parts was partitioned into the fruits, followed by thestem (Fig. 10). K deficiency slightly increased thepartitioning of 13C to fruits on day 7 after treatment.However on day 14, it had no significantly effect.
Fig. 3. The effect of K deficiency on diurnal changes of stem diameter of tomato plants. On each occasion, the diameter changes in both control andK-deficient plants were recorded with reference to ‘Zero change’ at midnight (00.00 h). Control (thin line), –K treatment (thick line).
Fig. 4. The effect of K deficiency on changes in fruit diameter in tomato. Changes in fruit diameter were monitored as described in Fig. 2. Control(thin line), –K (thick line).
K deficiency and sink activity 5 of 12
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
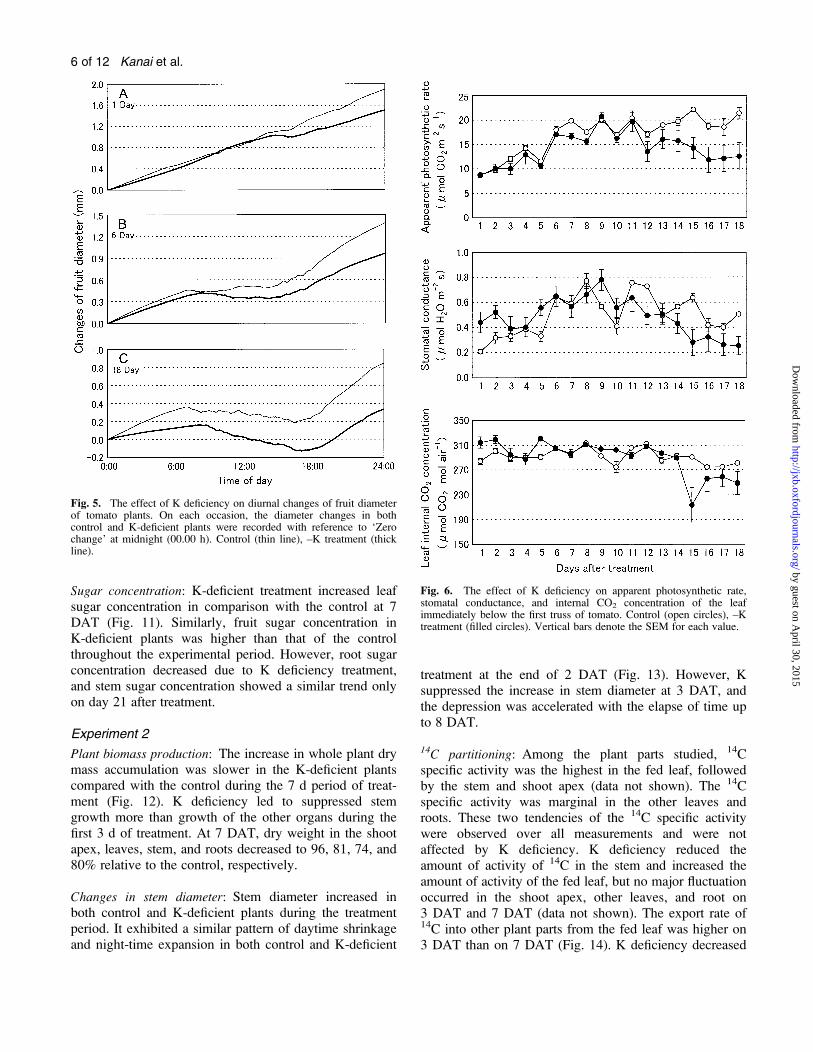
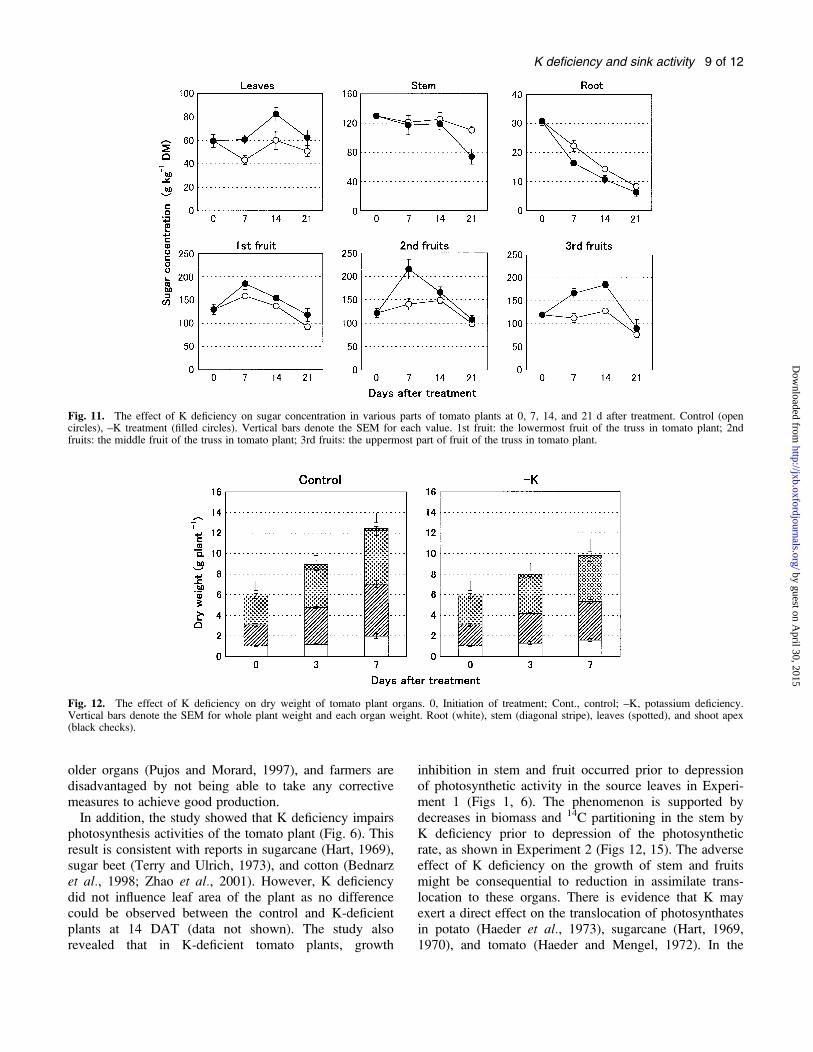
Sugar concentration: K-deficient treatment increased leafsugar concentration in comparison with the control at 7DAT (Fig. 11). Similarly, fruit sugar concentration inK-deficient plants was higher than that of the controlthroughout the experimental period. However, root sugarconcentration decreased due to K deficiency treatment,and stem sugar concentration showed a similar trend onlyon day 21 after treatment.
Experiment 2
Plant biomass production: The increase in whole plant drymass accumulation was slower in the K-deficient plantscompared with the control during the 7 d period of treat-ment (Fig. 12). K deficiency led to suppressed stemgrowth more than growth of the other organs during thefirst 3 d of treatment. At 7 DAT, dry weight in the shootapex, leaves, stem, and roots decreased to 96, 81, 74, and80% relative to the control, respectively.
Changes in stem diameter: Stem diameter increased inboth control and K-deficient plants during the treatmentperiod. It exhibited a similar pattern of daytime shrinkageand night-time expansion in both control and K-deficient
treatment at the end of 2 DAT (Fig. 13). However, Ksuppressed the increase in stem diameter at 3 DAT, andthe depression was accelerated with the elapse of time upto 8 DAT.
14C partitioning: Among the plant parts studied, 14Cspecific activity was the highest in the fed leaf, followedby the stem and shoot apex (data not shown). The 14Cspecific activity was marginal in the other leaves androots. These two tendencies of the 14C specific activitywere observed over all measurements and were notaffected by K deficiency. K deficiency reduced theamount of activity of 14C in the stem and increased theamount of activity of the fed leaf, but no major fluctuationoccurred in the shoot apex, other leaves, and root on3 DAT and 7 DAT (data not shown). The export rate of14C into other plant parts from the fed leaf was higher on3 DAT than on 7 DAT (Fig. 14). K deficiency decreased
Fig. 5. The effect of K deficiency on diurnal changes of fruit diameterof tomato plants. On each occasion, the diameter changes in bothcontrol and K-deficient plants were recorded with reference to ‘Zerochange’ at midnight (00.00 h). Control (thin line), –K treatment (thickline).
Fig. 6. The effect of K deficiency on apparent photosynthetic rate,stomatal conductance, and internal CO2 concentration of the leafimmediately below the first truss of tomato. Control (open circles), –Ktreatment (filled circles). Vertical bars denote the SEM for each value.
6 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
the export rate of 14C from the fed leaf into other parts on3 DAT (control and K deficiency; 54% and 48%,respectively). A 7 DAT, this phenomenon was enhanced(control and K deficiency 48% and 39%, respectively).Most of the 14C partitioning was received by the stem.This tendency did not change throughout the treatmentperiod in either control or K-deficient plants (data not
shown). The 14C partitioning rates of the stem in thecontrol and K deficiency treatments were 88% and 86%,respectively at 7 DAT.
Discussion
Effect of K deficiency on growth and water relations
Increased K supply to plants increases translocation ofassimilates (Huber, 1984), but the mechanism of this typeof effect does not include any improvement of net carbonexchange rate (Geiger and Conti, 1983). In the presentstudy, withdrawal of K from the culture media impairedtomato plant growth (Fig. 1). Similar responses to sub-optimal K supply were observed in sugarcane (Hart, 1969,1970) and sweet potato (Tsuno and Fujise, 1965).However, other researchers could not record such a
Fig. 7. The effect of K deficiency on the K concentration in various parts of tomato plants at 0, 7, 14, and 21 d after treatment. Control (opencircles), –K treatment (filled circles). Vertical bars denote the SEM for each value. Fruit (1st): the lowermost fruit of the truss in tomato plant; Fruit(2nd): the middle fruit of the truss in tomato plant; Fruit (3rd): the uppermost part of fruit of the truss in tomato plant.
Fig. 8. The effect of K deficiency on K partitioning among variousparts of tomato plant at 0, 7, 14, and 21 d after treatment. 0, Initiation oftreatment; Control, control; –K, K deficiency. Fruit (black and whitechecks), root (white), stem (diagonal stripes), leaves (spotted).
Table 1. The effect of K deficiency on 13C atom percentageexcess in various parts of tomato plants at different times aftertreatment
Values are given as average 6SEM.
Plant part 7 DAT 14 DAT
Control K deficiency Control K deficiency
Fed leaf 0.62960.048 0.88060.056 1.25060.089 1.32760.096Other leaves 0.00560.000 0.00460.000 0.00360.000 0.00260.000Fruits 0.24860.016 0.15560.009 0.08060.022 0.06060.013Stem 0.03260.002 0.01860.002 0.03560.005 0.02360.002Root 0.02960.002 0.02660.007 0.06860.017 0.01560.007
K deficiency and sink activity 7 of 12
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
response to K deficiency in Ricinus communis (Peukeet al., 2002) and tomato (Pujos and Morard, 1997). It ispossible that K supply to plants improves phloem loading,unloading, and conversion of assimilates in sink tissues,and a deficiency retards such activities (Zhao et al., 2001).In this context, the present study has helped to resolvesome of the controversies existing in the literature.Withdrawal of K from the culture medium did not alterthe carbon exchange rate of the plant for some time (Fig.6), but the effect was more constant on the sink organssuch as stem and fruit (Figs 2, 3). K deficiency altered thewater relations of the plant as evidenced by an enhanceddegree of stem shrinkage during daytime, and the night-time recovery was only lower than the control at 15 d(Fig. 3). This pattern of response of the tomato plant to Kdeficiency is identical to the response of Japanesepersimmon to salinity stress (Fujita et al., 2003a) and oftomato plants grown under P deficiency (Fujita et al.,2003b). Changes of stem diameter of Scots pine (Pinussylvestris L.) are known to consist of two components: anirreversible component due to growth and a variablecomponent due to moisture content that shows a diurnalcycle of expansion and shrinkage (Sevanto et al., 2002).Our previous studies showed that the stem diameterchanges were similar in both woody plants, such asJapanese pear (Ito et al., 2002) and Japanese persimmon(Fujita et al., 2003a), and herbaceous plants such astobacco (Moghaieb et al., 2006; Suwa et al., 2006) andtomato (Fujita et al., 2003b). The current results (Figs 2, 3)confirmed that K deficiency tended to reduce water supplyto the plant by affecting the variable component, as
daytime shrinkage of the stem was larger. According tothe Munch hypothesis, phloem sap flow is driven bya turgor pressure gradient. The present study suggests thatlow stem water potential reduces phloem turgor, therebydecreasing assimilates moving into the sieve tube forgrowth of the sink organs from the source. These tissuesregained water and expanded by night because of highturgor owing to the closure of stomata. On the other hand,the irreversible component of stem growth (Sevanto et al.,2002) reflects changes in wooden parts composed of cellwall materials such as lignin, cellulose, hemicellulose, etc.Synthesis of these materials is susceptible to deficiency ofassimilates, but not to any deficiency of water at night.
Effect of K deficiency on source–sink relationship
Any change in stem diameter from one night to the suc-ceeding night reveals the capacity for stem growth duringone day. A similar estimation can be done for fruitgrowth. In the present experiment, positive correlationswere observed between changes of fruit diameter build-up(Fig. 4) and fruit dry weight accumulation (Fig. 1)(control R2¼0.99, K deficiency R2¼0.83). Withdrawal ofK from the culture media inhibited the stem and fruitdiameter expansion on the fourth (Fig. 2) and sixth (Fig.5B) days after treatment, respectively, suggesting that Kdeficiency suppressed the growth of stem and fruit on thecorresponding days after treatment. This observation pro-vides a precise assessment of the effect of K deficiency ontomato plant growth at an early production stage andfacilitates quick replenishment of stock solution. In con-trast, correct assessment of the stress on fruit harvest is notpossible because of remobilization of the element from
Fig. 9. The effect of K deficiency on 13C export rate from the leafimmediately below the first truss of tomato at 7 d and 14 d aftertreatment (DAT). Cont., control; –K, K deficiency.
Fig. 10. The effect of K deficiency on 13C partitioning among variouspart of tomato plant at 7 d and 14 d after treatment (DAT). Cont.,control; –K, K deficiency. Fruits (black and white checks), root (white),stem (diagonal stripes), leaves (spotted).
8 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
older organs (Pujos and Morard, 1997), and farmers aredisadvantaged by not being able to take any correctivemeasures to achieve good production.In addition, the study showed that K deficiency impairs
photosynthesis activities of the tomato plant (Fig. 6). Thisresult is consistent with reports in sugarcane (Hart, 1969),sugar beet (Terry and Ulrich, 1973), and cotton (Bednarzet al., 1998; Zhao et al., 2001). However, K deficiencydid not influence leaf area of the plant as no differencecould be observed between the control and K-deficientplants at 14 DAT (data not shown). The study alsorevealed that in K-deficient tomato plants, growth
inhibition in stem and fruit occurred prior to depressionof photosynthetic activity in the source leaves in Experi-ment 1 (Figs 1, 6). The phenomenon is supported bydecreases in biomass and 14C partitioning in the stem byK deficiency prior to depression of the photosyntheticrate, as shown in Experiment 2 (Figs 12, 15). The adverseeffect of K deficiency on the growth of stem and fruitsmight be consequential to reduction in assimilate trans-location to these organs. There is evidence that K mayexert a direct effect on the translocation of photosynthatesin potato (Haeder et al., 1973), sugarcane (Hart, 1969,1970), and tomato (Haeder and Mengel, 1972). In the
Fig. 11. The effect of K deficiency on sugar concentration in various parts of tomato plants at 0, 7, 14, and 21 d after treatment. Control (opencircles), –K treatment (filled circles). Vertical bars denote the SEM for each value. 1st fruit: the lowermost fruit of the truss in tomato plant; 2ndfruits: the middle fruit of the truss in tomato plant; 3rd fruits: the uppermost part of fruit of the truss in tomato plant.
Fig. 12. The effect of K deficiency on dry weight of tomato plant organs. 0, Initiation of treatment; Cont., control; –K, potassium deficiency.Vertical bars denote the SEM for whole plant weight and each organ weight. Root (white), stem (diagonal stripe), leaves (spotted), and shoot apex(black checks).
K deficiency and sink activity 9 of 12
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
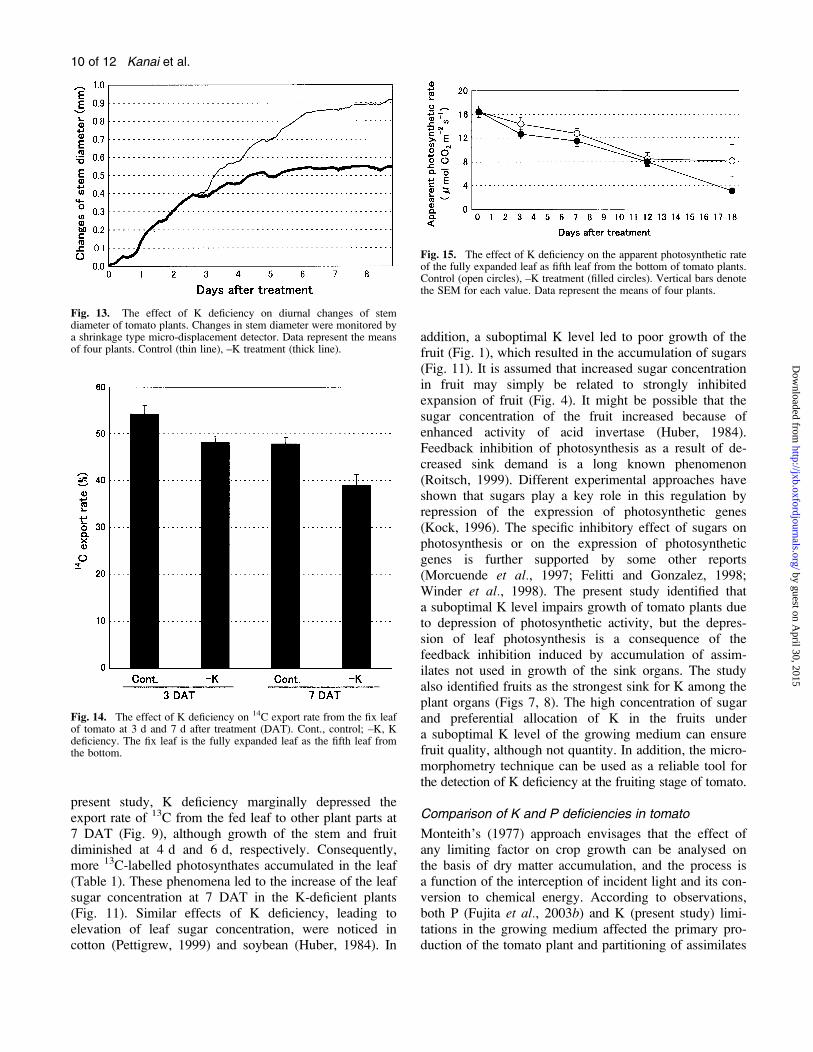
present study, K deficiency marginally depressed theexport rate of 13C from the fed leaf to other plant parts at7 DAT (Fig. 9), although growth of the stem and fruitdiminished at 4 d and 6 d, respectively. Consequently,more 13C-labelled photosynthates accumulated in the leaf(Table 1). These phenomena led to the increase of the leafsugar concentration at 7 DAT in the K-deficient plants(Fig. 11). Similar effects of K deficiency, leading toelevation of leaf sugar concentration, were noticed incotton (Pettigrew, 1999) and soybean (Huber, 1984). In
addition, a suboptimal K level led to poor growth of thefruit (Fig. 1), which resulted in the accumulation of sugars(Fig. 11). It is assumed that increased sugar concentrationin fruit may simply be related to strongly inhibitedexpansion of fruit (Fig. 4). It might be possible that thesugar concentration of the fruit increased because ofenhanced activity of acid invertase (Huber, 1984).Feedback inhibition of photosynthesis as a result of de-creased sink demand is a long known phenomenon(Roitsch, 1999). Different experimental approaches haveshown that sugars play a key role in this regulation byrepression of the expression of photosynthetic genes(Kock, 1996). The specific inhibitory effect of sugars onphotosynthesis or on the expression of photosyntheticgenes is further supported by some other reports(Morcuende et al., 1997; Felitti and Gonzalez, 1998;Winder et al., 1998). The present study identified thata suboptimal K level impairs growth of tomato plants dueto depression of photosynthetic activity, but the depres-sion of leaf photosynthesis is a consequence of thefeedback inhibition induced by accumulation of assim-ilates not used in growth of the sink organs. The studyalso identified fruits as the strongest sink for K among theplant organs (Figs 7, 8). The high concentration of sugarand preferential allocation of K in the fruits undera suboptimal K level of the growing medium can ensurefruit quality, although not quantity. In addition, the micro-morphometry technique can be used as a reliable tool forthe detection of K deficiency at the fruiting stage of tomato.
Comparison of K and P deficiencies in tomato
Monteith’s (1977) approach envisages that the effect ofany limiting factor on crop growth can be analysed onthe basis of dry matter accumulation, and the process isa function of the interception of incident light and its con-version to chemical energy. According to observations,both P (Fujita et al., 2003b) and K (present study) limi-tations in the growing medium affected the primary pro-duction of the tomato plant and partitioning of assimilates
Fig. 13. The effect of K deficiency on diurnal changes of stemdiameter of tomato plants. Changes in stem diameter were monitored bya shrinkage type micro-displacement detector. Data represent the meansof four plants. Control (thin line), –K treatment (thick line).
Fig. 14. The effect of K deficiency on 14C export rate from the fix leafof tomato at 3 d and 7 d after treatment (DAT). Cont., control; –K, Kdeficiency. The fix leaf is the fully expanded leaf as the fifth leaf fromthe bottom.
Fig. 15. The effect of K deficiency on the apparent photosynthetic rateof the fully expanded leaf as fifth leaf from the bottom of tomato plants.Control (open circles), –K treatment (filled circles). Vertical bars denotethe SEM for each value. Data represent the means of four plants.
10 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

to the sink, and a concurrent change in water status wasobserved. P deficiency lowered the water status of theplant, possibly by affecting the activity of water channelproteins (Clarkson et al., 2000; Steudle et al., 2000) andmaintenance of root hydraulic conductivity (Radin andEidenbock, 1984). In contrast, the present study suggeststhat water status may decline under the present conditions,because K nutrition is necessary for root hair elongation(Rigas et al., 2001) and maintenance of cell turgor (Taizand Zeiger, 2002). P deficiency did not affect leafphotosynthesis and diameters of stem and fruit comparedwith the control until a lag period of 12 d (Fujita et al.,2003b). However, under K deficiency, stem and fruitdiameter increase was reduced within the first few days oftreatment. Conversely, source activity (photosynthesis)declined at 12 DAT, possibly because of end-productinhibition. In the Munch pressure flow hypothesis,adequate phloem turgor is required for partitioning ofcarbon solutes from the source to the sink (Patrick, 1997).Both P and K deficiencies affected phloem turgor, leadingto shrinkage of stem diameter and reduction in partitioningof solutes. However, the effect of the former was lessimminent compared with the latter. Unlike P, K is a majorsolute for cell sap, and it can modulate cell water potentialdirectly and instantaneously. For the first time, the studiesreported here have identified that K nutrition is moreimportant for tomato plants than P nutrition.
References
Behboudian MH, Anderson DR. 1990. Effect of potassiumdeficiency on water relations and photosynthesis of tomato plants.Plant and Soil 127, 137–139.
Bendnarz CW, Oosterhuis DM, Evans RD. 1998. Leafphotosynthesis and carbon isotope discrimination of cotton inresponse to potassium deficiency. Environmental and Experimen-tal Botany 39, 131–139.
Beringer H, Haeder HE. 1981. Influence of potassium nutrition onstarch synthesis in barley grains. Zeitschrift fur Pflanzenernahringand Bodenkunde 144, 1–7.
Berman ME, Dejong TM. 1996. Water stress and crop load effectson fruit fresh and dry weights in peach (Prunus persica). TreePhysiology 16, 859–864.
Besford RT. 1978a. Effect of replacing nutrient potassium bysodium on uptake of and distribution of sodium in tomato plants.Plant Soil 50, 399–409.
Besford RT. 1978b. Effect of sodium in the nutrient medium on theincidence of potassium-deficiency symptoms in tomato plants.Plant and Soil 50, 427–432.
Besford RT, Maw GA. 1975. Effect of potassium nutrition ontomato plant growth and fruit development. Plant and Soil 42,395–412.
Clarkson DT, Carvajal M, Henzler T, Waterhouse RN,Smyth AJ, Cooke DT, Steudle E. 2000. Root hydraulicconductance: diurnal aquaporin expression and effects of nutrientstress. Journal of Experimental Botany 51, 61–70.
Davies JN, Winsor GW. 1967. Effect of nitrogen, phosphorus,potassium, magnesium and liming on the composition of tomatofruit. Journal of the Science of Food and Agriculture 18, 459–466.
Escobar-Gutierrez AJ, Zipperlin B, Carbonne F, Moing A,Gaudillere JP. 1998. Photosynthesis, carbon partitioning andmetabolite content during drought stress in peach seedlings.Australian Journal of Plant Physiology 25, 197–205.
Felitti SA, Gonzalez DH. 1998. Carbohydrates modulate theexpression of the sunflower cytochrome c gene at the mRNAlevel. Planta 206, 410–415.
Fujita K, Ito J, Mohapatra PK, Saneoka H, Lee K, Kurban H,Kawai K, Ohkura K. 2003a. Circadian rhythm of stem and fruitdiameter dynamics of Japanese persimmon (Diospyrus kakiThunb.) is affected by deficiency of water in saline environments.Functional Plant Biology 30, 747–754.
Fujita K, Lei K, Saneoka H, Moghaieb RE, Mohapatra PK.2004. Effect of N-deficiency on photoassimilate partitioningand rhythmic changes in fruit and stem diameter of tomato(Lycopersicon esculentum) during fruit growth. 4th InternationalCrop Science Congress. September 26–1 October Brisbane,Australia http://www.cropscience.org.au/icsc2004/poster/2/5/1/393_fujita.htm
Fujita K, Okada M, Lei J, Ito J, Ohkura K, Adu-Gyamfi JJ,Mohapatra PK. 2003b. Effect of P-deficiency on photo-assimilate partitioning and rhythmic changes in fruit and stemdiameter of tomato (Lycopersicon esculentum) during fruitgrowth. Journal of Experimental Botany 54, 2519–2528.
Geiger DR, Conti TR. 1983. Relation of increased potassiumnutrition to photosynthesis and translocation of carbon. PlantPhysiology 71, 141–144.
Haeder HE, Mengel K. 1972. Translocation and respiration ofassimilates in tomato plants as influenced by K nutrition.Zeitschrift fur Pflanzenernahrung und Bodenkunde 131, 139–147.
Haeder HE, Mengel K, Forster H. 1973. The effect of potassiumon translocation of photosynthates and yield pattern of potatoplants. Journal of the Science of Food and Agriculture 24, 1479–1487.
Hart CE. 1969. Effect of potassium deficiency upon translocationof 14C in attached blades and entire plants of sugarcane. PlantPhysiology 44, 1461–1469.
Hart CE. 1970. Effect of potassium deficiency upon translocationof 14C in detached blades of sugarcane. Plant Physiology 45,183–187.
Huber SC. 1984. Biochemical basis for effects of K-deficiency onassimilate export rate and accumulation of soluble sugars insoybean leaves. Plant Physiology 76, 424–430.
Ito J, Hasegawa S, Fujita K, Ogasawara S, Fujiwara T. 2002.Changes in water relations induced by CO2 enrichment governdiurnal stem and fruit diameters of Japanese pear. Plant Science163, 1169–1176.
Kock KE. 1996. Carbohydrate-modulated gene expression inplants. Annual Review of Plant Physiology and Plant MolecularBiology 47, 509–540.
Lucas RE. 1968. Potassium nutrition of vegetable crops. In:Kilmer VJ, Younts SE, Brady NC, eds. The role of potassiumin agriculture. American Society of Agronomy, Chapter 22.
Marschner H. 1995. Mineral nutrition of higher plants. London:Academic Press.
Mengel K. 1980. Effect of potassium on the assimilate conductionof storage tissue. Berichte der Deutschen Botanischen Gesell-schaft 93, 353–362.
Mengel K, Kirkby EA. 1987. Principles of plant nutrition. Bern:International Potash Institute.
Mengel K, Viro M. 1974. Effect of potassium supply on thetransport of photosynthates to the fruits of tomatoes (Lycopersi-con esculentum). Physiologia Plantarum 30, 295–300.
Moghaieb RE, Tanaka N, Saneoka H, Murooka Y, Ono H,Morikawa H, Nakamura A, Nguyen NT, Suwa R, Fujita K.
K deficiency and sink activity 11 of 12
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from

2006. Characterization of salt tolerance in ectoine-transformed
tobacco plants (Nicotiana tabaccum): photosynthesis, osmotic
adjustment, and nitrogen partitioning. Plant, Cell and Environ-ment 29, 173–182.
Monteith JL. 1977. Climate and the efficiency of crop production
in Britain. Philosophical Transactions of the Royal Society B:Biological Sciences 281, 277–294.
Morcuende R, Perez P, Martinez-Carrasco R. 1997. Short-termfeedback inhibition of photosynthesis in wheat leaves supplied
with sucrose and glycerol at two temperatures. Photosynthetica33, 179–188.
Nobuyasu H, Liu S, Adu-Gyamfi JJ, Mohapatra PK, Fujita K.2003. Variation in the export of 13C and 15N from soybean leaf:
the effects of nitrogen application and sink removal. Plant andSoil 253, 331–339.
Patrick JW. 1997. Phloem unloading and post-sieve element
transport. Annual Review of Plant Physiology and Plant Molec-ular Biology 48, 191–222.
Pettigrew WT. 1999. Potassium deficiency increases specific leaf
weights and leaf glucose levels in field-grown cotton. AgronomyJournal 91, 962–968.
Peuke AD, Jeschke DJ, Hartung W. 2002. Flows of element, ions
and abscisic acid in Ricinus communis and site of nitrate
reduction under potassium limitation. Journal of ExperimentalBotany 53, 241–250.
Pujos A, Morard P. 1997. Effect of potassium deficiency on
tomato growth and mineral nutrition at the early production stage.
Plant and Soil 189, 189–196.Radin JW, Eidenbock MP. 1984. Hydraulic conductance as
a factor limiting leaf expansion of phosphorus-deficient cotton
seedlings. Plant Physiology 75, 372–377.Rigas S, Debrosses G, Haralampidis K, Vicente-Agullo F,Feldmann KA, Grabov A, Dolan L, Hatzopoulos P. 2001.
TRH1 encodes a potassium transporter required for tip growth in
Arabidopsis root hairs. The Plant Cell 13, 139–151.Roitsch T. 1999. Source–sink regulation by sugar and stress.
Current Opinion in Plant Biology 2, 198–206.Sevanto S, Vesala T, Peramaki M, Nikinmaa E. 2002. Time lags
for xylem and stem diameter variations in a Scots pine tree. Plant,Cell and Environment 25, 1071–1077.
Sowder CM, Tarpley L, Vietor DM, Miller FR .1997. Leafphoto-assimilation and partitioning in stress-tolerant Sorghum.Crop Science 37, 833–838.
Steudle E. 2000. Water uptake by roots: effects of water deficit.Journal of Experimental Botany 51, 1531–1542.
Suwa R, Nguyen NT, Saneoka H, Moghaieb RE, Fujita K.2006. Effect of salinity stress on photosynthesis and vegetativesink in tobacco plants. Soil Science and Plant Nutrition 52,243–250.
Taiz L., Zeiger E. 2002. Plant physiology, 3rd edn. Sunderland,MA: Sinauer Associates, Inc.
Terry N, Ulrich A. 1973. Effect of potassium deficiency on thephotosynthesis and respiration of leaves of sugar beet. PlantPhysiology 51, 783–786.
Thuno Y, Fujise K. 1965. Studies on dry matter production ofsweet potato. Bulletin of the National Institute for AgriculturalScience, Series D, no. 13.
Trudel MJ, Ozbun JL. 1971. Influence of potassium on carotenoidcontent of tomato fruit. Journal of the American Society forHorticultural Science 96, 763–765.
Van den Boogaard R, Alewijns D, Veneklass EJ, Lambers H.1997. Growth and water-use efficiency of 10 Triticum aestivumcultivars at different water availability in relation to allocation ofbiomass. Plant, Cell and Environment 20, 200–210.
Wall ME. 1940. The role of potassium in plants. II. Effects ofvarying amounts of potassium on the growth, status and metab-olism of tomato plants. Soil Science 49, 315–331.
Widders IE, Lorenze OA. 1979. Tomato root development asrelated to potassium nutrition. Journal of the American Societyfor Horticultural Science 104, 216–220.
Winder TL, Jindong S, Okita TW, Edwards GE. 1998. Evidencefor the occurrence of feedback inhibition of photosynthesis inrice. Plant and Cell Physiology 39, 813–820.
Winsor GW. 1968. A long-term factorial study of the nutrition ofgreenhouse tomatoes. Proceedings of the 6th Colloquium,International Potash Institute, Florence, France.
Yemm EW, Willis AJ. 1954. The estimation of carbohydrates inplant extracts by anthrone. Biochemical Journal 57, 508–514.
Zhao D, Oosterhuis DM, Bednarz CW. 2001. Influence ofpotassium deficiency on photosynthesis, chlorophyll content, andchloroplast ultra-structure of cotton plants. Photosynthetica 39,103–109.
12 of 12 Kanai et al.
by guest on April 30, 2015
http://jxb.oxfordjournals.org/D
ownloaded from
Related Documents











