Aus DEM DEPARTMENT FüR PATHOBIOLOGIE DER VETERINäRMEDIZINISCHEN UNIVERSITäT WIEN DEPARTMENTSPRECHERIN: UNIV.PROF. DR.MED.VET. TZT. ANJA JOACHIM FACH: PARASITOLOGIE ENDOPARASITENBELASTUNG UND -MANAGEMENT BEI MILCHZIEGEN IN DER BIOLOGISCHEN LANDWIRTSCHAFT UNTER BERüCKSICHTIGUNG VON HALTUNG UND FüTTERUNG DIPLOMARBEIT DER VETERINäRMEDIZINISCHEN UNIVERSITäT WIEN VORGELEGT VON BARBARA PATTISS-KLINGEN WffiN, SEPTEMBER 2008

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Aus DEM
DEPARTMENT FüR PATHOBIOLOGIE
DER
VETERINäRMEDIZINISCHEN UNIVERSITäT WIEN
DEPARTMENTSPRECHERIN: UNIV.PROF. DR.MED.VET. TZT. ANJA JOACHIM
FACH: PARASITOLOGIE
ENDOPARASITENBELASTUNG UND -MANAGEMENT BEI MILCHZIEGEN IN DER BIOLOGISCHEN LANDWIRTSCHAFT
UNTER BERüCKSICHTIGUNG VON HALTUNG UND FüTTERUNG
DIPLOMARBEIT
DER
VETERINäRMEDIZINISCHEN UNIVERSITäT WIEN
VORGELEGT VON
BARBARA PATTISS-KLINGEN
WffiN, SEPTEMBER 2008
ubmeisch
Dissertation

BETREUER UND ERSTBEGUTACHTER:
UNIV.PROF. DR.MED.VET. HEINRICH PROSL
ZWEITBEGUTACHTERIN:
UNIV.PROF. DR.MED.VET. TZT. ANJA JOACHIM

INHALTSVERZEICHNIS
1. EINLEITUNG UND FRAGESTELLUNG 1
2. LITERATüRüBERSICHT 2
2.1 Biologische Landwirtschaft und Ziegenhaltung als Wirtschaftsfaktor 2
2.2 Häufige Parasitosen bei Ziegen 4
2.2.1 Helminthosen , 4
2.2.2 Protozoonosen 16
2.3 Mögliche Folgen einer Endoparasitose 18
2.4 Gesetzliche Richtlinien der biologischen Viehwirtschaft bzgl. Haltung und Medikamenteneinsatz 19
2.4.1 Ausläufe und Haltungsgebäude 19
2.4.2 Krankheitsvorsorge und tierärztliche Behandlung 19
2.4.3 Erlaubter Einsatz synthetischer Anthelminthika in der biologischen Landwirtschaft 20
2.5 Anthelminthika-Einsatz und mögliche Komplikationen 22
2.5.1 Anthelminthika-Resistenzen 22
2.5.2 Selektiver Anthelminthika-Einsatz als möglicher Lösungsansatz... 28
2.6 Alternative Behandlungsmethoden 31
2.6.1 Homöopathie und Phytotherapie 31
2.6.1.1 Phytotherapie 32 2.6.1.2 Homöopathische Therapieformen 33
2.6.2 Kupfer 33
2.6.3 Biologische Kontrollverfahren 33
2.7 Optimierung von Managementfaktoren und präventive Maßnahmen 35
2.7.1 Weidemanagement 35
2.7.2 Fütterung 39
2.7.2.1 Proteine 40 2.7.2.2 Energie 41 2.7.2.3 Kondensierte Tannine 42 2.7.2.4 Weitere Sekundärstoffe 44
2.7.3 Prävention durch Vakzinierung 44
2.7.4 Prävention durch Zucht und Rasseauswahl 45
2.7.4.1 Rasse 45 2.7.4.2 Zucht 46

3. MATERIAL UND METHODE 47
3.1 Klimadaten der untersuchten Region 47
3.1.1 Niederschlagsmenge 47
3.1.2 Temperatur 47
3.2 Betriebe 47
3.3 Befragung der Landwirte zu den Betriebsabläufen 47
3.4 Tierauswahl 48
3.5 Klinische Untersuchung 48
3.6 Erhebung des Body Condition Score (BCS) 48
3.7 Probenentnahme 49
3.8 Probenaufarbeitung 49
3.8.1 Kombiniertes Sedimentations-Flotations-Verfahren 50
3.8.2 Modifiziertes McMaster-Verfahren 50
3.8.3 Kotkultur (nach Roberts und O'Sullivan) und Auswander- verfahren (nach Baermann-Wetzel) 51
3.9 Statistische Auswertung 52
4. ERGEBNISSE 52
4.1 Betriebscharakteristik und Befragungsergebnisse 52
4.1.1 Betrieb 1 mit Auslauf und Heufütterung 52
4.1.2 Betrieb 2 mit Auslauf und Grünfütterung 54
4.1.3 Betrieb 3 mit ganzjähriger Weidehaltung 57
4.2 Ergebnisse der klinischen Untersuchung 59
4.3 Ergebnisse des Body Condition Scorings 59
4.4 Ergebnisse der parasitologischen Untersuchungen 61
4.4.1 Kombiniertes Sedimentations-Flotations-Verfahren 61
4.4.1.1 Betriebliche Unterschiede 61 4.4.1.2 Einfluss des Alters auf die Ausscheidungsraten 64 4.4.1.3 Saisonale Variation der Eiausscheidung 67
4.4.2 Modifiziertes McMaster-Verfahren 70
4.4.2.1 Betriebsabhängige Unterschiede von epg bzw. opg 70 4.4.2.2 Einfluss des Alters auf die Ei- bzw. Oozystenausscheidung 72 4.4.2.3 Saisonale Variation des epg bzw. opg 74
4.4.3 Kotkultur (nach Roberts und O'Sullivan) und Auswander- verfahren (nach Baermann-Wetzel) 75
4.4.3.1 Betriebsabhängige Unterschiede der Larvenprävalenz 76 4.4.3.2 Einfluss des Alters auf die Larvenprävalenz 78 4.4.3.3 Saisonale Variation der Larvenprävalenz 79

5. DISKUSSION 82
5.1 Methodenkritik 83
5.2 Haltungs- und Fütterungspraxis der Betriebe 84
5.3 Betriebliche EndoparasitenkontroUe und mögliche Optimierung 86
5.4 BCS und dessen Korrelation mit der Endoparasitenbelastung 89
5.5 Einfluss des Haltungssystems 92
5.5.1 Endoparasitenspektrum 93
5.5.2 Befallsintensität 94
5.6 Einfluss des Alters 97
5.7 Einfluss von Jahreszeit und Klima 99
5.7.1 Allgemeine klimatische Faktoren 100
5.7.2 Niederschlagsmenge 100
5.7.3 Temperatur 101
5.8 Die Bedeutung immunologischer Faktoren 101
5.8.1 Aggregierte Parasitenverteilung 103
5.8.2 „Resilience" oder die immunologische Belastbarkeit 104
5.8.3 Immunität in Laktation und Trächtigkeit 104
5.8.4 Strategien der Ziege zum Schutz vor Endoparasiten 106
5.9 Frühdiagnostik 107
6. SCHLUSSFOLGERUNG = 109
7. ZUSAMMENFASSUNG 109
8. SUMMARY 110
9. LITERATURVERZEICHNIS 113
10. ANHANG 123

1. EINLEITUNG UND FRAGESTELLUNG
Der Sektor der biologischen Landwirtschaft und der damit verbundene Wirtschafts-
zweig konnten in den letzten Jahren starken Zuwachs verzeichnen. Der Begriff der
biologischen Landwirtschaft steht dabei für Bestrebungen nach dem Erhalt fruchtba-
rer Böden, Umweltschutz, Tiergesundheit sowie der Produktion qualitativ hochwer-
tiger (Tier-)Produkte ohne den prophylaktischen Einsatz von Chemotherapeutika zur
Krankheitsprävention.
Unabhängig von den Zuwächsen innerhalb des biologischen Produktionssektors ver-
doppelte sich in den letzten 30 Jahren auch der weltweite Ziegenbestand (RINALDI et
al., 2007) und damit die wirtschaftliche Bedeutung der Ziege. Infolgedessen gewin-
nen Faktoren, die die Gesundheit und Produktivität der Ziegen negativ beeinflussen
können, immer mehr an Bedeutung. Endoparasiten spielen dabei als schwerwiegen-
des Gesundheitsrisiko insbesondere beim kleinen Wiederkäuer eine bedeutende Rol-
le (PAPADOPOULOS et al., 2003; KNOX et al., 2006), können sie doch massive öko-
nomische (Produktions-)Verluste bis hin zu Todesfällen verursachen (WALLER,
1999; KooPMANN, 2005; PAPADOPOULOS et al., 2003; HOSTE et al., 2008).
Allerdings unterliegt die biologische Viehwirtschaft restriktiven Beschränkungen
hinsichtlich eines prophylaktischen Medikamenteneinsatzes, so dass medikamenteile
Behandlungen ausschließlich auf kurativer Ebene gestattet sind. Darin inkludiert ist
auch der Einsatz jeglicher Antiparasitika. Nicht zuletzt darauf sind höhere Parasi-
tenprävalenzen in biologisch wirtschaftenden Betrieben im Vergleich zu konventio-
neller Viehwirtschaft zurückzuführen, während sonstige Aspekte von Gesundheit
und Wohlbefinden der Tiere als gleichwertig oder sogar besser eingestuft werden
können (LuND u. ALGERS, 2003).
Dementsprechend gewinnt ein nachhaltiges Endoparasitenmanagement, das nicht nur
auf der Verabreichung chemisch-synthetischer Anthelminthika basiert, in biologi-
schen - aufgrund der eskalierenden Anthelminthika-Resistenzen jedoch auch in kon-
ventionellen - Tierhaltungssystemen immer mehr an Bedeutung, um die Verluste
weiterhin so gering wie möglich zu halten (WALLER, 1999). Der Begriff der Nach-
haltigkeit umfasst in diesem Zusammenhang eine Kombination aus biologischen,
biotechnologischen und chemischen Kontrollmechanismen in der Tierhaltung sowie
auch in der Tierzucht mit dem Ziel, die Verwendung chemischer Substanzen zur Pa-
rasitenkontrolle auf ein Minimum reduzieren zu können (THAMSBORG et al., 1999),

und impliziert zusätzlich die immense Bedeutung der Prophylaxe durch Optimierung
der Rahmenbedingungen (HOSTE et al., 2005).
Zahlreiche Forschungsprojekte der vergangenen Jahre widmeten sich bereits der Un-
tersuchung des Einflusses von Managementfaktoren wie Haltung und Fütterung der
Tiere auf die Endoparasitenbelastung, um daraus Methoden zur Prophylaxe ableiten
zu können, und beschäftigten sich mit der Suche nach alternativen Behandlungsmög-
lichkeiten klinisch manifester Parasitosen. Themen wie Herden- und Gesundheits-
management rücken im Zuge dessen immer mehr in den Mittelpunkt (KOOPMANN,
2005), wobei das Wissen um das bei Ziegen qualitativ und quantitativ vorkommende
Endoparasitenspektrum die Grundlage zur wirksamen Vorbeugung bzw. erfolgrei-
chen Therapie darstellen muss (REHBEIN et al., 1998).
Vor diesem Hintergrund war es das primäre Anliegen dieser Arbeit, das Ausmaß
einer Endoparasitosenproblematik anhand der Eiausscheidungsrate in drei biologisch
wirtschaftenden Milchziegenbetrieben mit unterschiedlichen Haltungsformen im
Sommer und Spätherbst zu ermitteln. Des Weiteren sollte das Parasitenausmaß zu
Body Condition Score (BCS), Haltungsform, Alter der Tiere und Jahreszeit in Bezie-
hung gesetzt werden, um etwaige vorhandene Korrelationen aufzuzeigen und aus
diesen Vorschläge zur Optimierung des Endoparasitenmanagements zu entwickeln.
Insbesondere wurde die Validität des BCS als Indikator für das Ausmaß vorliegender
Parasiteninfektionen geprüft, um daraus eine mögliche Hilfestellung für den Land-
wirt zur Erkennung erkrankter und behandlungswürdiger Tiere ableiten zu können.
2. LITERATURüBERSICHT
2.1 Biologische Landwirtschaft und Ziegenhaltung als Wirt- schaftsfaktor
Der prozentuale Anteil der biologisch bewirtschafteten Flächen innerhalb der EU
wuchs im Zeitraum von 1985 bis 1998 um mehr als das Doppelte an (von 1 % auf
2,1 %; LAMPKIN, 2000, zitiert in CABARET et al., 2002). Als Gründe für diesen An-
stieg können ein gesteigertes Emährungsbewusstsein (weniger Medikamenten- und
Pestizidrückstände in tierischen und pflanzlichen Produkten) und die damit einherge-
hende erhöhte Nachfrage nach „sauberen", ökologisch produzierten Lebensmitteln
vermutet werden (THAMSBORG et al., 1999; CABARET et al.; 2002; WALLER, 2006a).

Der Verdopplung des Ziegenbestandes liegt nicht zuletzt die Überlegenheit der Zie-
gen gegenüber anderen Nutztieren zu Grunde, die auf anspruchslosen Futterbedürf-
nisse und der Widerstandsfähigkeit gegenüber unwirtlichen Umweltbedingungen bei
einer gleichzeitig breit gefächerten Produktpalette von Milch über Fleisch, Leder
und, je nach Rasse, auch Wolle beruht (BALICKA-RAMISZ, 1999). In Industrieländern
steht vor allem die Milchproduktion im Vordergrund, die Fleischgewinnung stellt
einen Nebenzweig dar (REHBEIN et al., 1998; RINALDI et al., 2007). Allein Europa
produziert mit seiner nur 2,5 % des Weltziegenbestandes umfassenden Ziegenpopu-
lation 18 % der weltweit erwirtschafteten Ziegenmilch (BOYAZOGLU et al., 2005).
Die im Rahmen der biologischen Bewirtschaftung vollzogene extensive Viehwirt-
schaft mit Freiland- und Weidehaltung stellt einen der Hauptunterschiede zu intensi-
ven Haltungssystemen dar. Im Zuge dessen steigt mit längeren Weideperioden je-
doch gleichzeitig die Exposition gegenüber parasitären Weideinfektionen (WALLER,
2006a), und häufig können eine hohe Befallsintensität, ein starker Infektionsdruck
und ein breiteres Parasitenspektrum als in konventionellen Betrieben beobachtet
werden (CABARET et al., 2002). Gastrointestinale Nematoden, Leberegel und Band-
würmer (Moniezia) sind die von biologisch arbeitenden Landwirten am häufigsten
genannten parasitär-bedingten Probleme in der biologischen Wiederkäuerhaltung in
Großbritannien (RODERICK et al., 1999, zitiert in CABARET et al., 2002).
Die größten wirtschaftlichen Einbußen durch Parasiten stellen subklinische Produk-
tionsverluste dar (PERRY U. RANDOLPH, 1999, zitiert in WALLER, 2006a). Daraus er-
geben sich neben dem ökonomischen auch neue bedeutsame Aspekte des Tierschut-
zes, wonach unkontrollierte Parasiteninfektionen die Tiere massiv belasten können
(WALLER, 2006a). Die in biologischer Viehwirtschaft besondere Relevanz von Para-
sitosen ist nicht zuletzt Folge des bereits erwähnten, ausschließlich restriktiv und
kurativ erlaubten Medikamenteneinsatzes, dem prophylaktische Maßnahmen, geeig-
nete und Erfolg versprechende homöopathische und/oder phytotherapeutische Be-
handlungsformen in jedem Fall vorzuziehen sind, so dass auf chemisch-synthetische
Allopathika erst in zweiter Instanz oder bei ausbleibendem Behandlungserfolg und
generell so selten wie möglich zurückgegriffen werden soll (EU-Verordnung
2092/91,2007).
Im Folgenden soll nun auf bedeutsame Aspekte, Zusammenhänge, Probleme und
Möglichkeiten hinsichtlich der Endoparasitosenkontrolle bei Ziegen im Einzelnen
eingegangen werden.

2.2 Häufige Parasitosen bei Ziegen
Im Folgenden werden zunächst in Form eines kurzen Überblicks jene Endoparasiten
vorgestellt, die aufgrund ihres gehäuften Auftretens - allgemein und insbesondere in
den hier untersuchten Betrieben - von besonderer Relevanz sind (nach SMITH U.
SHERMAN, 1994; ECKERT et al., 2005a; SCHNIEDER, 2006).
2.2.1 Helminthosen (nach SCHNffiDER, 2006)
• Familie der Tnchostrongylidae und ihre Verwandten
Zu dieser weltweit verbreiteten Familie zählen Haemonchus, Teladorsagia fsyn.
Ostertagia; SCHNIEDER, 2006J, Trichostrongylus, Cooperia und - inzwischen der
eng verwandten Familie der Molineidae zugehörig - Nematodirus. Geringe Wurm-
bürden beeinträchtigen das Wirtstier für gewöhnlich kaum. Mit steigender Belastung
können subklinische Symptome wie reduzierte Gewichtszunahme und Appetitlosig-
keit auftreten. Klinische Symptome treten erst im Falle eines massiveren Befalls mit
Trichostrongylidae auf und äußern sich in Gewichtsabnahme, Diarrhö, Anämie und
Kehlgangsödemen. Alle genannten Arten zeigen einen direkten Entwicklungszyklus,
d.h. sie brauchen keinen Zwischenwirt. Adulte, im Gastro-Intestinaltrakt (GI) des
Wirtes lebende Würmer legen Eier, die vom Wirt mit dem Kot ausgeschieden wer-
den. Außerhalb des Tieres erfolgt die Embryonalentwicklung zu den ersten Larven in
den Eiern (L I), die sodann aus der Eihülle schlüpfen, sich häuten und so zu Zweit-
larven (L II) werden. Die L II leben weiterhin im Kot, nehmen bedeutend an Größe
zu und durchleben die Weiterentwicklung zur infektiösen Drittlarve (L III), wobei
jedoch die vollständige Häutung zur L III erst nach erfolgter Ingestion durch den
neuen Wirt erfolgt. Die Entwicklung zur infektiösen Drittlarve umfasst bei idealen
Umweltbedingungen (Temperatur um 20-25° C und ausreichend Feuchtigkeit) eine
Zeitspanne von sieben bis zehn Tagen. Nachdem diese optimalen Bedingungen je-
doch in Mitteleuropa nur für kurze Zeit und nur während des Hochsommers vorlie-
gen, kann die Entwicklung unter weniger günstigen Umständen bis zu Monate dau-
ern. Eine Ausnahme stellt hier Nematodirus spp. dar, dessen Entwicklung zur infek-
tiösen Drittlarve noch im Ei stattfindet. Bei Nematodirus battus erfolgt das Schlüpfen
der Larve erst nach längerer Kälteeinwirkung. Das Risiko der Infektion mit den be-
reits während der Weideperiode des Vorjahres ausgeschiedenen Eiern entsteht somit
verzögert in der Weidesaison des Folgejahres.

Eine Besonderheit der Trichostrongylidae ist die Fähigkeit, ihren Entwicklungszyk-
lus innerhalb des Wirtstieres unterbrechen zu können. Diese Phase der metabolischen
Inaktivität wird Hypobiose genannt und kann vom Immunstatus des Wirtes, jedoch
primär von Klima- und jahreszeitlichen Faktoren nachhaltig beeinflusst werden, so
dass die Einwirkung tiefer Temperaturen im Herbst vor der Ingestion der Parasiten-
stadien die Wahrscheinlichkeit der Hypobiose wesentlich erhöhen (EYSKER, 1997,
zitiert in ZAJAC, 2006). In Regionen, wo hypobiotische Larvenstadien auftreten, sind
in Spätwinter und Frühjahr erhöhte Eiausscheidungsraten durch die plötzliche Wie-
deraufnahme der Larvenentwicklung zu beobachten, wovon in erster Linie Tiere mit
vorübergehend herabgesetzter Immunreaktion, insbesondere Tiere vor und nach der
Geburt, betroffen sind (GroBS, 1986, zitiert in ZAJAC, 2006).
Bei Trichostrongylidosen handelt es sich primär um Weideinfektionen. Infektiöse
Stadien aller Spezies können auf der Weide überwintern und sterben erst mit zuneh-
menden Temperaturen und geringerer Feuchtigkeit im Lauf des Sommers ab. Die
Infektion erfolgt meist unmittelbar nach dem Austrieb durch Aufnahme kontaminier-
ten Futters oder Wassers durch den Wirt, was durch die Wanderung der aktiv beweg-
lichen Larven vor allem in den Morgen- und Abendstunden erleichtert wird. Die
Larven gelangen nach erfolgter Ingestion an den je nach Art unterschiedlichen Ziel-
ort des GI-Trakts des Wirtes, vollenden dort die Häutung zur parasitischen L III und
dringen sodann in die Krypten und Drüsen der Mucosa ein, wo sie sich innerhalb von
ein bis zwei Tagen zu L FV häuten. Die vierten Larven erlangen nun, nach erneuter
Häutung, auf der Mucosaoberfläche das Stadium des adulten, fortpflanzungsfähigen
Nematoden. Nach drei Wochen scheidet der neu infizierte Wirt bereits Eier aus, aus
denen wiederum - je nach Witterung - ab Mitte Juli bzw. August/September eine
erheblich Anzahl von infektiösen L III hervorgeht, die bei nicht immunogenen Tie-
ren zu klinischen Erscheinungen (Sommerteladorsagiose) führen können. Die durch-
schnittliche Präpatenzzeit beträgt je nach Spezies zwischen zwei und vier Wochen.
Viele der noch im Herbst aufgenommenen L III überwintern im Wirtstier, d.h. sie
verfallen vorübergehend bis Ende des Winters in ein Ruhestadium (Hypobiose) und
vollenden ihre Entwicklung erst vier bis sechs Monate nach der Aufnahme.
Haemonchus contortus, Teladorsagia circumcinta, Teladorsagia daytiani sowie Tri-
chostrongylus axei leben im Abomasum, wovon der blutsaugende H. contortus in-
folge des entstehenden, mitunter massiven Blutverlustes als pathogenster Nematode
der kleinen Wiederkäuer angesehen werden kann (K00PMA^fN et al., 2006). Übrige

Trichostrongylus spp. sind ebenso wie Cooperia curticei und Nematodirus spp. im
Dünndarm anzutreffen.
Haemonchose
Während der histotropen Phase der Entwicklung von H. contortus, dem gedrehten
Magenwurm, sind meist nur unspezifische Verdauungsstörungen zu verzeichnen.
Zehn Tage post infectionem, nach Abschluss der histotropen Phase, ist eine normo-
zytäre, hypochrome Anämie mit gesteigerter Erythropoese zu beobachten. Der durch
das Saugen der Würmer verursachte Blutverlust und die dadurch entstehenden Läsi-
onen an der Labmagenmucosa begründen die auftretende Leukopenie, wobei es sich
vorrangig um eine Lymphopenie handelt. Gleichzeitig tritt infolge des Blutverlustes
eine Hypoproteinämie, vor allem eine Hypalbuminämie, auf. Klinisch präsentiert
sich die Haemonchose vorrangig bei Kitzen bis zum 6. Lebensmonat. Mit fortschrei-
tendem Alter ist zwar eine rasseabhängige Immunitätsbildung zu beobachten, jedoch
kann es auch bei laktierenden Muttertieren zu chronischer Anämie, Inappetenz,
Kümmern und einer durch die Hypalbuminämie bedingte Ödembildung an Triel und
Unterbrust kommen. Durchfall tritt hingegen nur sehr selten auf. Bei ausbleibender
Behandlung endet der Krankheitsverlauf, der sich mitunter über Monate hinziehen
kann, tödlich.
Teladorsagiose
Bei Schaf und Ziege ist vor allem die Sommerteladorsagiose, hervorgerufen durch
die Infektion mit T. circumcinta, von Bedeutung. Die Parasitose tritt witterungsab-
hängig ab Mitte Juli, nach Aufnahme vieler Larven und sich daraus zur gleichen Zeit
und in großer Anzahl entwickelnden Würmem, auf. Auch hier können sich noch im
Spätherbst aufgenommene Larven in eine Hypobiose begeben, woraufhin diese ihre
Entwicklung erst im späten Winter oder Frühling vollziehen, was als Wintertelador-
sagiose in Erscheinung tritt. Diese Form der Teladorsagiose wird jedoch bei den
kleinen Wiederkäuern seltener beobachtet.
Die Pathogenese der Teladorsagiose kann in drei Phasen unterteilt werden: die
histotrope, die Luminal- und die Reparationsphase. In der histotropen Phase nisten
sich die parasitären L III im Lumen der Labmagendrüsen ein, was zur Abnahme der
Belegzellen führt, die für die HCl-Produktion zuständig sind. Stattdessen werden die
Belegzellen durch undifferenzierte, nicht funktionstüchtige Zellen ersetzt. Dies be-

dingt sowohl eine Aufweichung der Zellverbände und damit eine Vergrößerung des
Interzellularspalts als auch einen Anstieg des pH-Wertes auf bis zu 7 (physiologisch
ist ein pH-Wert von zwei). Das dadurch entstandene neutrale Milieu ist für das Über-
leben der Parasiten essentiell, fördert jedoch gleichzeitig die bakterielle Vermehrung
und beeinträchtigt infolge verringerter Umwandlung von Pepsinogen in Pepsin die
Proteinverdauung, wodurch es zu einer Hypalbuminämie kommt. Außerdem resul-
tiert die durchlässig gewordene Schleimhautbarriere in zusätzlichem Plasmaprotein-
verlust ins Darmlumen. Im Blut ist - aufgrund der bereits erwähnten verringerten
Umwandlung des Pepsinogens in Pepsin - ein erhöhter Pepsinogengehalt nachweis-
bar.
Das zunehmende Larvenwachstum beeinträchtigt in der Folge auch benachbarte Drü-
sen und resultiert dort ebenfalls in einer Reduktion der Belegzellen, wodurch es zu
einer Ausbreitung der Schädigung kommt. Bei der Sommerteladorsagiose, die bei
kleinen Wiederkäuern bevorzugt auftritt, beginnt die Luminalphase ca. zehn Tage
post infectionem infolge der Auswanderung der Würmer aus den Drüsen in das
Labmagenlumen (bei den überwinternden Stadien erfolgt dies erst nach Ende der
Hypobiose vier bis sechs Monate nach der Infektion). Die Luminalphase wird von
einer hyperplastischen Gastritis und pathologischen Labmagenveränderungen (Epi-
thelverlusten) begleitet, was sich klinisch in Durchfall äußern kann. Die Phase der
Reparation, die von der Rückbildung der pathologischen Veränderung an der Lab-
magenschleimhaut gekennzeichnet ist, beginnt nach der spontanen Beseitigung der
adulten Nematoden.
Die Sommerteladorsagiose ist vor allem bei Jungtieren zwischen Juli und September
zu beobachten und kann sich klinisch in intermittierender, wässriger Diarrhöe (kot-
verschmutzte Tiere), Mattigkeit, Inappetenz und Abmagerung zeigen. Häufig tritt
jedoch der subklinische Verlauf auf, der sich in verringerter Körperkondition zeigt.
Ebenso kann reduziertes Knochenwachstum und verminderte Mineralisierung der
Knochen beobachtet werden. Es erfolgt eine Immunitätsbildung, die jedoch erst mit
der zweiten Weidesaison voll ausgeprägt ist, so dass adulte Tiere selten erkranken.
Trichostrongylose
Bei der Ziege kommen vor allem T. colubriformis, T. capricola und T. vitrinus, die
den oralen Anteil des Dünndarms besiedeln, wie auch T. axei, der sowohl im Dünn-
darm wie auch im Labmagen gefunden werden kann, vor. Die Entwicklung der Lar-

8
ven erfolgt in der Magenschleimhaut, womit nur geringe Schäden der Mucosa ein-
hergehen. Im Verlauf bilden sich daraus flache Ulzera. Die adulten Nematoden leben
mit ihrem vorderen Anteil intraepithelial, während sich deren Hinterteil frei im Kryp-
ten- oder Drüsenlumen befindet. Dies verursacht weit reichende Zellnekrosen und
führt damit zum Untergang von Haupt- und Belegzellen, die durch undifferenzierte
Vorläuferzellen ersetzt werden, was wiederum einen Anstieg des pH-Wertes im
Labmagen bewirkt. Infolge der Besiedelung ist die Mucosa verdickt, wobei jedoch
die Dünndarmzotten verkürzt bzw. atrophiert vorliegen. Die vermehrte Zellabschilfe-
rung an Zottenspitze bzw. an der Mucosaoberfläche begünstigt die Entstehung von
kleinen Erosionen.
Bereits bei subklinischem Befall ist die Darmmotilität verringert, die Passage somit
verlangsamt. Im Blut ist neben einer Hypalbuminämie auch ein steigender Pepsino-
gengehalt nachweisbar, bei durchfallbedingten Flüssigkeitsverlusten kann es zum
Anstieg des Hämatokrit kommen. Besonders der Stickstoffverlust in das Darmlumen
in Form von austretenden Plasmaproteinen, verstärkter Schleimbildung und vermehr-
ter Abschilferung ist Ursache für die pathophysiologischen Veränderungen, die sich
an verminderter Gewichtszunahme und schlechterem Fleischansatz zeigen. Das Vor-
liegen einer Hypophosphorämie kann auch ein verringertes Knochenwachstum bzw.
eine verminderte Mineralisation verursachen. Klinisch kann sich eine Infektion in
Inappetenz, Kümmern, starkem Durchfall zeigen und auch zum Tod führen (ZAJAC,
2006). Sehr häufig bleibt der Verlauf jedoch subklinisch. Eine Immunitätsbildung
erfolgt innerhalb der ersten sechs bis neun Monate, bei Frühjahrskitzen demnach im
Herbst. Da ist jedoch von vielen Faktoren abhängig ist, variiert sie von Tier zu Tier.
Cooperiose
Die Cooperiose ist meist mit einer Teladorsagiose assoziiert, was die Pathogenität
der jeweiligen Parasitose verstärkt. Vor allem C. curticei ist hier zu nennen, die v.a.
den oralen Dünndarm, insbesondere die Schleimhautkrypten besiedeln. Die Dünn-
darmzotten sind auch hier verkürzt und die Schleimbildung durch vermehrt vorlie-
gende Becherzellen erhöht. Durch Invasion von Leukozyten und Bildung von Öde-
men ist die Dünndarmmucosa verdickt und es kommt zur protein loosing entero-
pathy, d.h. einem Proteinverlust durch reduzierte Resorptionsfähigkeit, was wieder-
um zu verminderter Gewichtszunahme führt.

Klinisch kann außerdem weicherer Kot bis zu wässrigem Durchfall, begleitet von
Inappetenz, beobachtet werden. Eine außergewöhnlich massive Primärinfektion as-
soziiert mit hochgradigen Durchfällen kann schwerwiegende bis letale Folgen haben.
• Familie der Strongyloididae
Der Zwergfadenwurm Strongyloides papillosus ist der einzig bedeutsame Vertreter
dieser Familie und kommt weltweit vor. Sein Entwicklungszyklus zeichnet sich
durch einen Generationswechsel (Heterogonie) aus, wobei ausschließlich die Weib-
chen parasitieren. Sie leben in der Dünndarmmucosa, wo sie ihre Eier ablegen, die
bereits fortgeschritten embryoniert durch den Wirt ausgeschieden werden. Der Groß-
teil davon entwickelt sich bei > 10°C und genügend Feuchtigkeit innerhalb weniger
Tage zur L II, aus denen sich dann die infektiösen L III differenzieren. Aus dem Rest
der Eier entsteht bereits nach fünf Tagen eine frei lebende Population von Weibchen
und Männchen. Aus den von dieser Population produzierten Eiern schlüpfen wieder-
um L I, die sich zweimal häuten und sich so zu infektiösen L III weiterentwickeln,
die in feuchter Umgebung bis zu vier Monate lebensfähig sind.
Die Infektion kann perkutan oder galaktogen erfolgen und ist somit als eine Erkran-
kung der Neugeborenen und Jungtiere zu betrachten. Bei der perkutanen Infektion
dringen die infektiösen Larven an Stellen dünner Haut (Klauensaum, Bauch, Ingui-
nalgegend) ein und vollziehen eine Körperwanderung über Lymphe und Blut zu
Herz, Lunge, Trachea, Larynx und darüber in den Darm. Die Larven erreichen auf
diesem Weg bereits nach drei Tagen Duodenum und Jejunum und damit ihre Zielor-
te, in deren Schleimhaut sie sich bis zum fünften Stadium weiter differenzieren. Die
Präpatenz umfasst 9-14 Tage. Bei bereits immunen Tieren gelangen die Larven di-
rekt über den großen Körperkreislauf in verschiedene Organe, ohne sich in die Lunge
auszubohren, und besiedeln bevorzugt Muskulatur. Durch vor der Geburt ins Euter
gewanderte L III kann die Infektion somit auch galaktogen über Kolostrum bzw.
Milch erfolgen. Bei optimalen Umweltbedingungen können auf perkutanem Infekti-
onsweg schnell Superinfektionen entstehen, die einen dramatischen Verlauf nehmen
können.
Pathologische Veränderungen finden sich der Entwicklung des Parasiten entspre-
chend an Haut, Lunge und Dünndarm. An der Haut handelt es sich dabei um bei der
Invasion der Larven entstehende Erytheme. In der Lunge kommt es drei Tage post
infectionem durch Auswanderung der Larven aus dem Blut in die Lungenalveolen zu

10
Blutungen, woraus eine interstitielle Pneumonic entstehen kann. Pathologische Ver-
änderungen am Dünndarm finden ihre Ursache in den im Epithel wandernden und
Eier ablegenden Weibchen und können sich als Epithelverluste, Petechien und Ek-
chymosen, ödemisierter Mucosa oder auch als katarrhalische Duodenitis und Jejuni-
tis 9-14 Tage post infectionem manifestieren, woraus Malabsorption resultiert.
Entsprechend der Pathogenese finden sich Symptome an der Haut in Form von Der-
matitiden, Quaddeln oder Erythemen an dünnhäutigen Stellen wie Bauch, Schenke-
linnenflächen oder am Klauensaum. Symptome der Atemwege sind trockener Husten
und/oder beschleunigte Atem- und Herzfrequenz. Intermittierende, wässrige Diar-
rhöe und damit assoziierte Hypalbuminämie, Inappetenz, rascher Gewichtsverlust
und Anämie gehen mit der Darmbesiedelung durch die adulten Weibchen einher.
Selten kann die Infektion auch letal enden. Jungtiere etablieren rasch eine Immunität,
sofern die Infektion nicht zu massiv ist und zum frühzeitigen Zusammenbruch der
Körperabwehr führt.
• Familie der Chabertiidae
Innerhalb dieser Familie ist bei der Ziege vor allem die Art Chabertia ovina von Be-
deutung, die weltweit verbreitet ist. In den meisten Fällen liegt eine Weideinfektion
vor, eine Infektion im Stall ist jedoch gelegentlich zu beobachten. Die Entwicklung
der L III umfasst im Freien bei 25°C eine Zeitspanne von 5-7 Tagen. Die Drittlarven
können im Sommer nur 6-8 Wochen überleben und nur in Einzelfällen überwintern,
weshalb für die Kontamination der Weiden im Frühjahr ältere Ausscheider von be-
sonderer Bedeutung sind, bei denen es zu peripartaler erhöhter Eiausscheidung
kommt.
In ihrem Entwicklungszyklus durchlaufen die L III eine einwöchige histotrope Phase
in der Mucosa bzw. Submucosa des Dünndarms, wo sie sich zur L IV häuten. Nur
bei starkem Befall kommt es während dieser Phase zu Epithelschäden der Dünn-
darmmucosa. Nach dieser dritten Häutung breiten sich die L IV nun auch auf den
caudalen Dünndarm und bevorzugt auf das Caecum aus. Ab dem 24. Tag post infec-
tionem erfolgt die Häutung zum fünften Stadium, das daraufhin die Schleimhaut des
proximalen, bei starkem Befall aber auch des distalen Colon bis zum Rektum besie-
delt. Die Präpatenz beträgt sechs bis sieben Wochen.
Schäden entstehen vor allem durch L V und Adulte im aufgerollten Teil des Colons,
wo die L V große Mucosapfröpfe einsaugen und enzymatisch verdauen, wodurch je

11
nach Ausmaß lokale Läsionen bis zu großflächige Erosionen entstehen. Die Mucosa
ödemisiert, wird von Entzündungszellen infiltriert und produziert vermehrt Schleim.
Gleichzeitig bringt diese Phase massive Proteinverluste ins Darmlumen, insbesonde-
re von Albumin, mit sich. Mit dem Ortswechsel der fünften Stadien entstehen lokale
Blutungen. Zur Entwicklung Eier legender adulter Nematoden kommt es nur bei
Vorliegen einer schwacher Infektion (wenige hundert Larven), bei starken Infektio-
nen (mehrere tausend Larven) unterbleibt eine Eiausscheidung, nachdem die (Prä)-
Adulten im zweiten Monat nach erfolgter Infektion abgehen.
Sehr häufig liegen Mischinfektionen mit Trichostrongyliden vor. Reine Infektionen
mit Knötchenwürmem führen nur selten zur Erkrankung infizierter Tiere. Wenn es
jedoch zur Erkrankung kommt, so kann eine akute, durch histotrope Larven hervor-
gerufene, und eine chronische, durch lumenbewohnende Stadien verursachte Phase
unterschieden werden. Die akute Phase tritt im Herbst nach erfolgter Primärinfektion
auf und kann mit Fieber, Inappetenz und profusen, schleimigen bis wässrigen, später
blutigen Durchfällen vergesellschaftet sein. In der chronischen Phase kommt es zum
Absetzen schwarzen oder roten Kots und verminderte Darmperistaltik, Anämie, Hy-
palbuminämie, Hypoglucosämie, Hypocalc- und Hypophosphorämie sowie Leis-
tungseinbußen können beobachtet werden.
Ebenso können auch in Hypobiose eingetretene, inhibierte L FV, die sich bereits ab
Winter weiterentwickeln, zu profusen Durchfällen führen, in denen nicht selten
Würmer gefunden werden. Die Folgen sind Hypalbuminämie, Anämie, Inappetenz
und Gewichtsabnahme. Mit wiederholten, leichten Chabertia-Infekiionen nimmt die
Adultenbürde deutlich ab, was auf immunologische Auseinandersetzung des Wirtes
mit der Infektion schließen lässt.
• Familie der Trichuridae
Bei kleinen Wiederkäuern kommen vor allem die Peitschenwürmer Trichuris ovis, T.
capreoli und T. skrjabini vor, die alle einen direkten Entwicklungszyklus durchlau-
fen. Die Drittlarven von Trichuris spp. entwickeln sich im Ei und schlüpfen erst nach
erfolgter Ingestion des Eis durch den Wirt, wobei die Weiterentwicklung Temperatu-
ren > 14°C und genügend Feuchtigkeit voraussetzt. Das infektiöse Stadium auf der
Weide wird erst nach drei bis vier Monaten erreicht. Eine Überwinterung und sogar
jahrelanges Überleben sind bei guten Bedingungen möglich. Nach Ingestion infekti-
öser Eier schlüpfen die Larven im aboralen Teil des Dünndarms und penetrieren die

12
Schleimhaut des Caecums und proximalen Colons, wo sich nach zwei Wochen zur L
in, nach einem Monat zu L IV und nach sieben Wochen zu Präadulten häuten. Die
Präpatenz umfasst sechs bis acht Wochen. Infolge eines Befalls kann es zu entzünd-
lichen Erscheinungen in Caecum und Colon und damit verbunden zu Störungen des
Flüssigkeitshaushaltes kommen, wobei jedoch die meisten Infektionen subklinisch
und harmlos verlaufen oder nur von unspezifischen Symptomen wie Inappetenz,
verminderte Darmmotorik, struppiges Haarkleid und Minderzunahme begleitet wer-
den. Spezifischere Symptome können wässrige bis zu blutige Durchfälle mit Hy-
poproteinämie, Anämie infolge des Blutsaugens des Parasiten und Ödeme am Hals
sein. Der Verlauf kann v.a. bei Jungtieren aber auch schwerwiegend vorliegen und
von Festliegen bis zu letalem Ausgang führen.
• Familie der Oxyuridae
Aus der Familie der Pfriemenschwänze kommt bei Ziegen die Art Skrjabinema ovis
vor, die im Dickdarm leben. Die Eiablage erfolgt im Bereich des Perineums. Dort
können die eine Larve enthaltenden Eier mit einem Klebestreifen nachgewiesen wer-
den. Nach der oralen Aufnahme der Eier schlüpfen die Larven bereits im Dünndarm,
die zwei Häutungen erfolgen jedoch im Dickdarm. Die Präpatenz beträgt 25-48 Ta-
ge. Eine Infektion bleibt normalerweise symptomlos und ist in den meisten Fällen ein
Zufallsbefund.
• Familie der Protostrongylidae
Protostrongyliden sind unter den kleinen Wiederkäuern zwar weltweit verbreitet,
jedoch nur begrenzt pathogen und deshalb von geringerer wirtschaftlicher Relevanz.
In Mitteleuropa sind fünf Arten der kleinen Lungenwürmer bei Schaf und Ziege an-
zutreffen, die häufig als Mischinfektionen vorliegen. Dazu gehören Protostrongylus
rufescens, P. brevispiculum (nur beim Schaf), Muellerius capillaris, Cystocaulus
ocreatus und Neostrongylus linearis.
Die Entwicklung erfolgt über Zwischenwirte (heteroxen). Als solche fungieren am
Land lebende Nackt- oder Gehäuseschnecken. Die Weibchen leben in Brutknoten in
der Lunge und legen hier ihre Eier ab, in denen die Erstlarven heranwachsen. Diese
bohren sich in die Alveolen aus und erreichen über das Flimmerepithel die Trachea
und den Pharynx, wo sie abgeschluckt und via naturalis mit dem Kot ausgeschieden
werden. Die Erstlarven der Kleinen Lungenwürmer sind allgemein außerordentlich

13
widerstandsfähig und überleben daher bei feuchten Bedingungen monatelang. Auch
Kälte und Frost überstehen sie unbeschadet. Längere Trockenheit, wechselnde Witte-
rungsverhältnisse sowie lange, trockene Kälteperioden reduzieren jedoch Lebens-
dauer und Anzahl der Larven. Sobald Erstlarven mit geeigneten Zwischenwirten
konfrontiert sind, dringen sie innerhalb weniger Minuten in den Schneckenfuß ein,
wo sie sich zweimal häuten und zu bescheideten Drittlarven heranwachsen. Die Ent-
wicklung zur L III dauert im Hochsommer zwei Wochen, kann bei ungünstigen Be-
dingungen jedoch einige Monate betragen. Im Schneckenfuß können die Larven je-
doch auch wochenlangen Frost unbeschadet überstehen und lange überleben. Das
Risiko der Infektion ist im Frühjahr und Herbst besonders groß, wobei je nach Jah-
reszeit unterschiedliche Schneckenarten als Zwischenwirte fungieren.
Der Endwirt infiziert sich über die orale Aufnahme von Drittlarven entweder samt
Schnecke oder durch das Freiwerden der Larven nach dem Tod des Zwischenwirtes.
Die aufgenommenen L III invadieren im Bereich des Dickdarms die Darmwand und
wandern zu den Mesenteriallymphknoten, in denen sie sich zur L IV häuten. Über
Ductus thoracicus, Herz und Pulmonalarterie erreichen sie sodann die Lunge. Dort
angekommen, bohren sie sich in ventilierte Anteile der Lunge aus und erreichen nach
der letzten Häutung in Brutknoten lebend, je nach Art innerhalb vier bis neun Wo-
chen, das Stadium der Geschlechtsreife. Die Patenz kann mehrere Jahre umfassen,
wobei Unterbrechungen in der Larvenausscheidung zu beobachten sind. Die Exkreti-
on der Larven scheint peripartal und in jedem Fall mit Verlauf der Weidesaison an-
zusteigen, wobei Jährlinge und Muttertiere bzw. alte Ziegen häufig stärker befallen
sind als Jungtiere. Das Ausmaß der Infektion ist jedoch nicht nur alters- sondern
auch witterungsabhängig und sinkt mit steigenden Temperaturen.
Schäden verursachen die Brutherde und Wurmknoten in der Lunge. Brutherde sind
entweder lobulär-emphysematöse (Protostrongylus-Arten) oder inflammatorische
und zum Teil auch atelektatische Bereiche, die - je nach Lungenwurmspezies - ein
beträchtliches Ausmaß annehmen können. In den Brutknoten sind zumeist mehrere
fortpflanzungsfähige Weibchen, vereinzelte Männchen, jedoch unzählige Eier und
Erstlarven anzutreffen. Wurmknoten hingegen stellen sich als kleine, kugelförmige,
infertile Protostrongyliden enthaltende Veränderungen dar und sind je nach Art un-
terschiedlich lokalisiert. Erst nach der Begattung beginnt die Eiausscheidung und es
entstehen aus Wurmknoten Brutknoten.

14
M. capillaris ruft mit seinen wandernden L III punktförmige, hämorrhagische Ver-
änderungen in der Lunge hervor. Seine Brutknoten sind klein, rundlich, liegen an
oder zumindest dicht unter der Oberfläche und erscheinen bei Palpation schrotkugel-
ähnlich. In ihnen sind die Larven und Adulten enthalten. Der klinische Verlauf einer
Infektion verläuft zumeist symptomlos, es kann jedoch zu vereinzeltem trockenem
Husten kommen, der Anzeichen für eine chronisch vorliegende Bronchopneumonie
ist. Bei starkem Befall wird der Sauerstoffaustausch behindert, was sich klinisch in
reduziertem Sauers toffpartialdruck (pOa) äußert, sich aber nach erfolgter Behandlung
rasch wieder auf Normalwerte regeneriert.
Ziegen scheinen im Allgemeinen empfänglicher für die Pathogenität von Protostron-
gyliden zu sein als Schafe, wobei die Befallsraten bei Ziegen jedoch rasseabhängig
variieren können (SCHNIEDER, 2006). Die Tatsache, dass infizierte Tiere jahrelang
Ausscheider von Erstlarven sein können, deutet darauf hin, dass die Immunitätsbil-
dung gegen adulte Würmer nicht ausreichend ausgeprägt zu sein scheint.
• Familie der Anoplocephalidae
Monieziose
Bei kleinen Wiederkäuern wird häufiger die weltweit verbreitete, im Dünndarm le-
bende Art Moniezia expansa, seltener M. benedeni nachgewiesen. Die Artenbestim-
mung erfolgt anhand der adulten Würmer und deren Bandwurmglieder (Proglotti-
den). Als Zwischenwirte fungieren Moosmilben, die in humusreichen Böden leben
und dort über die Ingestion der vom Wirtstier mit dem Kot ausgeschiedenen Proglot-
tiden die darin enthaltenen Bandwurmeier aufnehmen. Bei optimalen Temperaturen
(28 °C) bilden sich im Zwischenwirt nach ca. vier Wochen infektiöse Cysticercoide
(Finnen). Nimmt der Endwirt - bevorzugt Jungtiere - nun über die Nahrung (vor
allem während des Weidegangs) die mit Bandwurmfinnen infizierten Moosmilben
auf, werden die Finnen im Duodenum des Wirtstieres freigesetzt und entwickeln sich
dort im Zeitraum von 30-52 Tagen zu geschlechtsreifen Adulten. Die Patenz beträgt
bei Schafen häufig nur drei Monate, kann jedoch bis zu acht Monate umfassen. Die
Bandwurmeier können selbst tiefe Temperaturen unbeschadet überstehen und somit
monatelang überleben.
Die Monieziose verläuft häufig subklinisch. Bei der chronischen Form können als
Folge einer Dysbakterie abwechselnd Durchfall und Verstopfung beobachtet werden,
die zu verminderter Gewichtszunahme führen können. Pathologisch kann infolge

15
einer Infektion eine exsudative, selten hämorrhagische Enteritis beobachtet werden.
Die Diagnose erfolgt über im Kot makroskopisch nachweisbare Proglottiden bzw.
mikroskopisch identifizierbare Eier.
• Familie der Fasciolidae
Fasciolose
Die Fasciolose ist eine weltweit verbreitete Erkrankung der Wiederkäuer mit meist
chronischem Verlauf. In gemäßigtem Klima ist vor allem der Große Leberegel, Fas-
ciola hepatica, weit verbreitet, während Fasciola gigantica hauptsächlich in subtro-
pischen und tropischen Klimazonen beheimatet ist.
Der Große Leberegel durchläuft eine heteroxene Entwicklung, wobei die Zwerg-
schlammschnecke, Lymnaea truncatula, in Mitteleuropa als Zwischenwirt fungiert.
Die Infektion des Endwirts erfolgt oral mit der Nahrung, woraufhin die jungen Egel
innerhalb weniger Tage die Dünndarmwand des Wirtstieres penetrieren und somit in
die Bauchhöhle gelangen. Im Zeitraum von ca. 90 Stunden invadieren sie die Leber,
wo sie für einige Wochen das Leberparenchym durchwandern, um schlussendlich als
fortpflanzungsfähige Egel die Gallengänge zu erreichen. Dort legen sie Eier, die mit
der Gallenflüssigkeit in den Darm abgegeben und innerhalb weniger Tage ausge-
schieden werden. Die Eier können im feuchten Kot einige Monate überleben, sind
jedoch gegenüber Austrocknung empfindlich. In den Eiern entwickeln sich bei opti-
malen Temperaturen (16-20°C) innerhalb von 10-20 Tagen Miracidien, die nur unter
idealen Bedingungen schlüpfen und innerhalb eines Tages einen Zwischenwirt errei-
chen müssen, da sie andernfalls absterben. In den Schnecken vollzieht sich sodann
die weitere Entwicklung zur Sporocyste, weiters zur Redie, Tochterredie bis zur Cer-
carie, die letztlich die Zwischenwirte aktiv verlassen, zu Metacercarien werden und
als solche die infektiösen Stadien in der Umwelt darstellen, die vom Wirt mit dem
Futter aufgenommen werden. Demzufolge ist die Fasciolose als typische Weidein-
fektion anzusehen.
Pathologische Veränderungen variieren einhergehend mit den Entwicklungsstadien
des Leberegels und reichen von meist kleinen Läsionen der Dünndarmwände durch
die penetrierenden Jungegel, Verletzungen der Leber beim Eindringen in das Leber-
parenchym, erheblichen Schäden des Leberparenchyms in Form von Bohrgängen,
die vernarben und so zu einer fortschreitenden Fibrosierung des Lebergewebes füh-

16
ren bis zu erheblichen Schleimhautverdickungen in den Gallengängen durch die a-
dulten Stadien.
Klinisch kann entweder die akute, die subakute oder auch die chronische Verlaufs-
form beobachtet werden. Die akute Erkrankung wird dabei von einer großen Anzahl
von durch das Leberparenchym wandernden Jungegeln ausgelöst, ist aber eher selte-
ner anzutreffen. Eier sind in diesem Stadium nicht im Kot nachweisbar. An der we-
sentlich häufiger auftretenden subakut verlaufenden Fasciolose sind sowohl Wander-
stadien als auch bereits adulte Egel beteiligt, weswegen auch Eier im Kot gefunden
werden können. Die ebenso häufige chronische Form wird fast ausschließlich durch
die in den Gallengängen lebenden adulten Egel verursacht, deren Eier im Kot nach-
gewiesen werden können. Die akute bzw. die häufiger auftretende subakute Fascio-
lose äußert sich in Symptomen wie raschem Verfall der Tiere, Inappetenz, Apathie,
Anämie und Ödembildung - mitunter auch Ikterus, Aszites und fibrinöser Peritonitis
- und führen zum Tod der Tiere. Beim chronischen Verlauf stehen Leistungsminde-
rungen aller Art im Vordergrund und werden häufig von Anämie und Hypalbuminä-
mie begleitet.
Die Anamnese und die im Herbst bzw. Winter auftretende Erkrankung geben Hin-
weis auf eine eventuell vorliegende Fasciolose. Die Verifizierung der Verdachtsdi-
agnose erfolgt über den Einachweis im Kot, welcher jedoch bei der akuten Form
negative Ergebnisse liefert. Zur Absicherung der Diagnose können deshalb zusätz-
lich labordiagnostische Verfahren zum Nachweis einer Anämie, einer Hypalbumi-
nämie und dem Vorliegen von erhöhten Leberenzymwerten hinzugezogen werden.
Immunologische Nachweisverfahren wie z.B. der ELISA gewinnen zunehmend an
Bedeutung. Die im Zuge der Infektion ablaufenden und mittels ELISA nachweisba-
ren Immunreaktionen führen jedoch nicht zum Aufbau einer ausreichend protektiven
Immunität, weswegen befallene Tiere ohne Behandlung bis zum natürlichen Abster-
ben der Egel infiziert bleiben.
2.2.2 Protozoonosen (nach TENTER, 2006)
Eimeriose (Kokzidiose)
Eimerien sind weltweit verbreitet. Jungtiere im Alter von vier bis zehn Wochen
durchleben unabhängig von ihrer Haltung eine Infektion. Ab dem sechsten Lebens-
monat nimmt die Befallsstärke jedoch stark ab. In Mitteleuropa kommen bei der Zie-
ge am häufigsten die Arten Eimeria arloingi, E. hirci, E. ninakohlyakimovae, E.

17
christenseni, E. caprina und E. alijevi vor. Bei Untersuchungen in Polen wurden am
häufigsten E. jolchijevi, E. ninakohlyakimovae und E. arloingi (BALICKA-RAMISZ,
1999) gefunden, während bei einer Studie in Dänemark E. ninakohlyakimovae, E.
arloingi und E. aspheronica am häufigsten vorzufinden war (BORGSTEEDE U.
DERCKSEN, 1996). In Tschechien konnte wiederum vor allem E. arloingi, gefolgt von
E. hirci und E. ninakohlyakimovae nachgewiesen werden (KOUDELA U. BOKOVä,
1998).
Haltungsunabhängig scheiden Jungtiere bedeutend höhere Mengen an Oozysten aus
als Jährlinge oder Muttertiere (siehe auch BoRGSTEEDE u. DERCKSEN, 1996; KOUDE-
LA u. BOKOVä, 1998), Neugeborene und sehr junge Kitze sind hingegen zumeist als
resistent zu betrachten. Klinisch äußert sich die Eimeriose im Alter von vier bis sie-
ben Wochen bzw. zwei bis vier Wochen nach Trennung von den Muttertieren (Kou-
DELA u. BOKOVä, 1998) wobei bei Weidetieren vor allem die Monate Mai und Juni
ein erhöhtes Risiko an Jungtierverlusten mit vermehrter Oozystenausscheidung mit
sich bringen. Bei Masttieren ist in den ersten sechs Wochen nach Aufstauung zur
Mast mit gefährlichen Coccidien-Infektionen zu rechnen, wohingegen bei der mut-
terlosen Aufzucht bereits am 13. Lebenstag bei Warm tränke- und am 20. Tag bei
Kalttränkeverfahren Eimerien-Oozysten im Kot nachgewiesen werden können. Die
Oozysten sind sehr unempfindlich gegenüber Umwelteinflüssen und können im spo-
rulierten Stadium - vermutlich auch im Freien - überwintern.
Die Arten unterscheiden sich hinsichtlich ihrer Pathogenität, wobei bei der Ziege E.
ninakohlyakimovae, E. arloingi, E. caprina (KoUDELA u. BOKOVÄ, 1998) aber auch
E. alijevi und E. christenseni als die pathogensten einzustufen sind. Das Ausmaß der
Symptomatik steht in direktem Zusammenhang mit der Anzahl der aufgenommenen
Oozysten. Faktoren wie Futterumstellung und Mangelernährung können prädisponie-
rend für klinisch apparente Eimeriosen sein.
Etwa zehn Tage nach oraler Infektion mit Eimerien-Oozysten bilden sich im Dünn-
darm Makromeronten, die die Infiltration des Darmabschnitts mit Leukozyten und
Makrophagen, eine Hyperplasie der Krypten und Epithelverlust verursachen. Klini-
sche Symptome sind in diesem Stadium jedoch noch nicht auffällig. Zu klinischen
Auffälligkeiten kommt es vor allem erst durch die Progamonten und Gamonten ins-
besondere im Caecum, seltener aber auch in Deum und Colon. Die Wand des Cae-
cums präsentiert sich entzündet, hyperämisch, ödemisiert und verdickt, während die
Mucosa hämorrhagisch sein und Epithelverlust auftreten kann. Bei der Infektion mit

18
weniger pathogenen Arten können katarrhalische Entzündungen des Dünn- und
Dickdarms beobachtet werden. Die sich entwickelnden Meronten zeigen sich als
über die Dünndarmmucosa verstreute, punktförmige Blutungen.
Klinisch zeigt sich ab dem 13.-15. Tag post infectionem für sieben bis zehn Tage
grünlicher bis dunkelbrauner, übelriechender, blutiger, auch schleimiger Durchfall,
Inappetenz bzw. Anorexic, Abmagerung und Exsikkose. Bei der Eimeriose mit we-
niger virulenten Arten wird zumindest eine reduzierte Gewichtszunahme verzeichnet.
Beim Auftreten klinisch apparenter Tiere muss sofort die gesamte Gruppe in die Be-
handlung miteinbezogen werden. Nachdem eine natürliche Infektion jedoch mit einer
guten Immunitätsbildung und damit dauerhaftem Schutz einhergeht, sollte der Ein-
satz von Antiparasitika überdacht werden und ist nur im dem Falle angebracht, so-
bald eine Infektion klinisch apparent verläuft und zu Produktionsverlusten führt
(BORGSTEEDE U. DERCKSEN, 1996).
2.3 Mögliche Folgen einer Endoparasitose (nach KNOX et al., 2006)
Endoparasitosen gehen sehr oft mit Produktionsverlusten einher und sind aus diesem
Grund von größter wirtschaftlicher Bedeutung. Diese Verluste resultieren zum einen
aus bis zu 50 % verringerter Futteraufnahme, die insbesondere den Proteinhaushalt
negativ beeinflusst, womit anabole Prozesse nur reduziert ablaufen können (SYKES U.
GREER, 2003). Das Ausmaß der Anorexic ist von Faktoren wie der Parasitenspezics,
Ort der Besiedelung und von Rasse, Alter und Immunstatus des Wirtstieres abhängig
(SYKES U. GREER, 2003). Zum anderen können endogene Proteinverluste in Form
von Blut, Plasma, geschädigten Epithelzellen und vermehrter Schleimbildung durch
deren ausbleibende Resorption und Ausscheidung über Faeces die Folge einer Infek-
tion sein. Des Weiteren kommt es zur verminderten Retention und Resorption essen-
tieller Mineralstoffe für Wachstum und Entwicklung und somit zur Beeinträchtigung
des Phosphor-, Calcium-, Magnesium- und Kupferhaushalts, was besonders bei
Jungtieren von großer Bedeutung sein kann (SvKES u. GREER, 2003). Lokale Gewe-
beschäden und Entzündungen können zwar die Resorption von Nährstoffen allge-
mein reduzieren, diese Verluste können jedoch im Allgemeinen durch vermehrte
Resorption distal der geschädigten Region kompensiert werden und sind demnach zu
vernachlässigen (HOSTE, 2001b).
Des Weiteren können sich Nematoden des Atmungs- oder Verdauungstraktes durch
Umverteilung der Nährstoffe von Produktion auf Reparation der durch Parasiten ent-

19
standenen Schäden negativ auf die Milchproduktion und Fruchtbarkeit auswirken,
was in der Milchproduktion verheerende Folgen haben kann (HOSTE et al. 2005).
2.4 Gesetzliche Richtlinien der biologischen Viehwirtschaft bzgl. Haltung und Medikamenteneinsatz
Die EU-Verordnung 2092/91 in der 61. ergänzten Auflage vom 15. Januar 2007 bil-
det die folgenden gesetzlichen Grundlagen.
2.4.1 Ausläufe und Haltungsgebäude
Die „(...) biologischen und ethologischen Bedürfnisse (...)" müssen in der Haltung
berücksichtigt werden, so dass eine „(...) artgerechte Unterbringung der Tiere ge-
währleistet ist (...)". Alle Säugetiere müssen „Weide- oder Freigeländezugang oder
Auslauf' zur Verfügung haben. Eine teilweise Überdachung der betreffenden Berei-
che ist erlaubt. Bei Pflanzenfressern soll die Nutzung der Weideflächen erfolgen,
sobald die Umstände dies zulassen. Im Winter kann von der Verpflichtung, Weide-
gang oder Auslauf zu gewähren, abgesehen werden, sofern den Tieren während der
Weidezeit Weidegang geboten wird und die Aufstauung während der Wintermonate
Bewegungsfreiheit gewährleistet. An Fläche müssen der adulten Ziege bis zum Jahr
2010 mindestens 1,5 m^/Tier Stall- und 2,5 m^/Tier Außenfläche zur Verfügung ste-
hen. Für Kitze sieht die Verordnung 0,35 m^/Tier Stall- und 0,5 m^/Tier Außenfläche
vor. Die Reinigung, Desinfektion und Schädlingsbekämpfung (Insekten und andere
Parasiten) in den Stallungen darf nur mit den in Anhang II der Verordnung genann-
ten Mitteln erfolgen.
2.4.2 Krankheitsvorsorge und tierärztliche Behandlung
Der Krankheitsvorsorge kommt innerhalb der biologischen Landwirtschaft eine vor-
rangige Bedeutung zu. Durch die Wahl geeigneter Rassen soll das Erkrankungsrisiko
so gering wie möglich gehalten werden. Die Haltung der Tiere muss tiergerecht und
an die Tierart angepasst erfolgen. Durch Förderung einer hohen Widerstandsfähig-
keit gegen Krankheiten soll Infektionen vorgebeugt werden. Eine hochwertige Fütte-
rung, regelmäßiger Auslauf und Weidezugang unterstützen die natürliche Immunität
der Tiere. Des Weiteren ist auf eine angemessene Besatzdichte zu achten, um so et-
waigen, mit Überbelegung assoziierten Gesundheitsrisiken entgegenzuwirken.

20
Im Krankheitsfall müssen erkrankte oder verletzte Tiere sofort und nach folgenden
Grundsätzen, wenn nötig in separaten, zweckdienlichen Räumlichkeiten, behandelt
werden. Die Behandlung sollte primär mittels Phythotherapeutika, Homöopathika
oder in Anhang II genau definierten Spurenelementen erfolgen. Diese Behandlungs-
formen sind chemisch-synthetischen allopathischen Tierarzneimitteln generell vor-
zuziehen, sofern ein Behandlungserfolg zu erwarten ist. Falls eine derartige Therapie
jedoch als nicht zielführend erscheint bzw. tatsächlich nicht zu gewünschtem Thera-
pieerfolg führt, so darf auf chemisch-synthetische allopathische Tierarzneimittel oder
Antibiotika zurückgegriffen werden. Eine prophylaktische Gabe von chemisch-
synthetischen allopathischen Tierarzneimitteln oder Antibiotika ist jedoch untersagt.
Des Weiteren muss über Art des Präparats, exakte Diagnose, Dosierung, Art der
Verabreichung, Behandlungsdauer und gesetzliche Wartezeit Auskunft erteilt und die
Daten der zuständigen Kontrollbehörde oder -stelle mitgeteilt werden, bevor das Tier
oder dessen Erzeugnisse verwertet werden dürfen. Behandelte Tiere müssen außer-
dem eindeutig gekennzeichnet werden.
Die Wartezeit nach dem Einsatz allopathischer Tierarzneimittel muss bei Tieren bio-
logischer Produktionsform die doppelte der gesetzlich vorgeschriebenen Dauer
betragen und in jedem Fall 48 Stunden umfassen, sofern keine Wartezeit angegeben
ist. Ist innerhalb eines Jahres eine mehrmalige, höchstens dreimalige Therapie mit
chemisch-synthetischen allopathischen Tierarzneimitteln oder Antibiotika erforder-
lich, so dürfen betroffene Tiere oder Tiergruppen bzw. daraus hervorgehende Pro-
dukte nicht mehr als „aus biologischer Landwirtschaft stammend" vermarktet werden
bzw. müssen die dafür vorgesehenen Umstellungszeiträume unter Zustimmung der
Kontrollbehörde oder -stelle berücksichtigt werden. Dies gilt jedoch nicht für Imp-
fungen und Parasiten-Behandlungen.
2.4.3 Erlaubter Einsatz synthetischer Anthelminthika in der biologischen Landwirtschaft (nach CABARET et al., 2002)
Der Einsatz synthetischer Anthelminthika wird für EU-Staaten übergeordnet durch
die EU-VERORDNUNG 2092/91 und zusätzlich auf nationaler Ebene geregelt. Sehr
häufig geben die nationalen Richtlinien einen rigideren Rahmen vor, als es die Ver-
ordnung der EU verlangt.
Derzeit dürfen laut EU-VO adulte Ziegen zweimal pro Jahr antiparasitär behandelt
werden. Im Falle von Ektoparasitosen darf eine zusätzliche Ektoparasitenbehandlung

21
erfolgen. Bei Kitzen ist eine dreimalige Behandlung erlaubt. Diese Reglementierung
erschwert das auf den Einsatz synthetischer Antiparasitika basierende Management
im Falle einer gleichzeitig vorliegenden Helminthose und Kokzidiose, da diese mit
jeweils unterschiedlichen Wirkstoffen therapiert werden müssen.
Zur Zeit sind drei Wirkstoffgruppen von Anthelminthika zugelassen: 1. die Gruppe
der Benzimidazole (Albendazol, Fenbendazol und Flubendazol) und Probenzimida-
zole (Febantel und Netobimin), deren Wirkprinzip in der verhinderten Ausbildung
des Zytoskeletts der Parasiten liegt; 2. die Imidazothiazole mit dem Vertreter Leva-
misol, die eine Lähmung der Nematoden verursachen; und 3. die makrozyklischen
Laktone mit ihrem Hauptvertreter Ivemnectin, die auf die durch Chlorid-Kanäle ver-
mittelte Neurotransmission der Parasiten wirken (BOWMAN, 1999, zitiert in ZAJAC,
2006).
Die Auswahl des geeigneten Anthelminthikums muss individuell an Art und Ausmaß
der Endoparasitose angepasst werden, nachdem jeder Wirkstoff sein spezifisches
Wirkspektrum aufweist und nicht für jede Helminthose gleich gut geeignet ist. Rich-
tiger Behandlungszeitpunkt und Auswahl des passenden Anthelminthikums sind ei-
nerseits für die Effizienz der Parasitenbekämpfung ausschlaggebend, andererseits
bringt der Einsatz ungeeigneter Anthelminthika zum falschen Zeitpunkt aufgrund der
Ineffizienz erheblich höhere Behandlungskosten und auch die Gefahr der Selektion
resistenter Parasiten mit sich (MICHEL et al., 1981, zitiert in MAINGI et al., 1996a).
Erschwert wird die Wahl des passenden Mittels dann, wenn bereits Resistenzen ge-
gen die Gruppe der Benzimidazole vorliegen, was gerade bei kleinen Wiederkäuern
zunehmend auftritt, nachdem dies die einzig gleichzeitig wirksame Stoffgruppe ge-
gen Nematoden, Moniezia und Leberegel ist. Im Falle vorliegender Benzimidazol-
Resistenz muss demnach auf eine Kombination mehrerer Präparate zurückgegriffen
werden, d.h. im Falle einer Infektion mit Nematoden, Moniezia und Leberegeln wä-
ren drei verschiedene Behandlungen nötig. Das bedeutet, dass ein ausschließlich auf
synthetischen Antiparasitika basierendes Endoparasitenmanagement in der biologi-
schen Viehwirtschaft nicht möglich ist, weshalb alternative Kontrollmechanismen
zunehmend an Bedeutung gewinnen.
Anthelminthisches Potential wurde außerdem den Zystein-Proteinasen zugesprochen,
die in Pflanzen wie Papaya, Ananas und Feige enthalten sind. Angriffsort scheint die
Cuticula (= Schutzhülle) der Nematoden zu sein, die durch diese Enzyme geschädigt
wird (STEPEK et al., 2004, zitiert in WALLER U. THAMSBORG, 2004). Nachdem der

22
Wirkmechanismus jedoch eher unspezifisch und das therapeutische Fenster (maxi-
mal tolerierbare Dosis/empfohlene therapeutische Dosis) sehr eng zu sein scheint,
bleibt noch offen, ob diese Enzyme in Zukunft tatsächlich als eine weitere Anthel-
minthika-Klasse zum Einsatz kommen können (WALLER U. THAMSBORG, 2004). Hier
bedarf es noch etlicher Forschungsarbeit. In jüngsten Studien von KAMINSKY et al.
(2008) konnte die Wirksamkeit von „amino-acetonitrile derivatives (AADs)" an Ace-
tylcholin-Rezeptor-Untereinheiten von verschiedensten - bereits gegen andere Wirk-
stoffe resistenten - Nematoden nachgewiesen werden.
Neben der Auswahl des geeigneten Anthelminthikums ist vor allem der Zeitpunkt
der Durchführung für den Erfolg von Anthelminthika-Behandlungen ausschlagge-
bend (MAINGI et al., 1996a; PAPADOPOULOS et al., 2006). PAPADOPOULOS et al.
(2006) empfehlen hierfür z.B. die prophylaktische Behandlung gegen Ende der Win-
terperiode bzw. zu Frühlingsbeginn, d.h. vor dem ersten Höhepunkt der
Eiausscheidung. Nachdem jedoch in der biologischen Landwirtschaft präventive
Therapien mit chemisch-synthetischen Allopathika nicht zulässig sind, ist jegliche
systematisch-prophylaktische Vorgangsweise nicht umsetzbar, weshalb Methoden
der selektiven Behandlung bzw. Managementverbesserung (siehe Kapitel 2.5.2 sowie
2.7) zur Infektionsprävention größere Bedeutung zukommt.
2.5 Anthelminthika-Einsatz und mögliche Komplikationen
2.5.1 Anthelminthika-Resistenzen
Das seit rund fünf Jahrzehnten (HERTZBERG U. BAUER, 2000) auf den Einsatz synthe-
tischer Antiparasitika basierende Parasitenmanagement und deren großzügiger - zum
Teil auch unsachgemäßer - Einsatz führte in den vergangenen Jahren zu eskalieren-
dem Anstieg und schneller Verbreitung von Anthelminthika-Resistenzen gegen ein-
zelne oder auch mehrere Wirkstoffgruppen und stellt eine wachsende Bedrohung für
die Viehwirtschaft, insbesondere bei kleinen Wiederkäuern, dar (HERTZBERG u.
BAUER, 2000; SILVESTRE et al., 2000; KAPLAN, 2004). Die ausschließlich auf medi-
kamenteller Ebene vollzogene EndoparasitenkontroUe verliert infolgedessen an
Wirksamkeit, so dass es trotz des Anthelminthika-Einsatzes immer häufiger zu Leis-
tungseinbußen und/oder klinischen Endoparasitosen kommt. Alle Regionen der Welt,
in denen kleine Wiederkäuer gehalten werden, sind inzwischen von Anthelminthika-
Resistenzen betroffen, die eines der größten Probleme für die Betriebe darstellen

23
(KAPLAN, 2004). In Gebieten wie Australien und Südafrika ist die Situation derma-
ßen dramatisch, dass dazu übergegangen werden muss, mit hohen Wurmbürden zu
leben, um die Situation nicht noch mehr zu verschlechtem (WALLER, 2006).
Die Prävalenz von Anthelminthika-Resistenzen ist bei Ziegen, insbesondere bei
Milchziegen, im Vergleich zu Schafen deutlich höher (CABARET, 2000, zitiert in
HOSTE et al., 2002a, 2002c). Dies ist auf die bei Milchziegen aufgrund ihrer unzurei-
chenden Immunantwort deutlich häufiger nötigen Anthelminthika-Behandlungen
(HOSTE U. CHARTIER, 1998a) sowie auf die begrenzte Wirkstoffauswahl während der
Laktation angesichts der Gefahr von Lebensmittelrückständen und zusätzlich auf die
häufige Unterdosierung bei Ziegen aufgrund ihrer metabolischen Eigenheiten zu-
rückzuführen (CHARTffiR u. HOSTE, 1997, zitiert in HOSTE et al., 2002a).
Diese Tatsache macht die Suche nach alternativen bzw. komplementären Lösungen
bei Ziegen umso dringender und mündet in verschieden gearteten Versuchen, die
Immunantwort des Wirtstieres zu verbessern bzw. eine Kontamination der Umge-
bung zu vermeiden. Die Basis für die Immunmodulation bilden züchterische Bemü-
hungen hinsichtlich der Selektion resistenter Tiere sowie die Optimierung der Prote-
inzufuhr wie auch die Entwicklung von Vakzinen, während das Weidemanagement
und der Einsatz von nematophagen Pilzen Aspekte der Kontaminationsvermeidung
darstellen (WALLER, 1999, zitiert in HOSTE et al., 2002a).
In den untersuchten Betrieben ist derzeit zwar keine Resistenz-Problematik bekannt,
sie kann jedoch langfristig auch hier trotz des seltenen Einsatzes nicht ausgeschlos-
sen werden.
• Betroffene Wirkstoffgruppen
In Europa überwiegen zum jetzigen Zeitpunkt Resistenzen gegen Benzimidazole,
wobei diese vor allem innerhalb der letzten zehn Jahre deutlich gestiegen sind. Dies
ergibt sich aus dem weit verbreiteten Einsatz dieser Wirkstoffgruppe. Umfragen in
französischen Milchziegenbetrieben zufolge setzen 97 % der Betriebe, die syntheti-
sche Anthelminthika anwenden, Benzimidazole und Probenzimidazole ein (HOSTE et
al., 2000). Dem zufolge wurde in Frankreich von Resistenzprävalenzen gegen Ben-
zimidazole von nahezu 80 % berichtet (CHARTIER et al. 2001, zitiert in HOSTE et al.,
2002c), Resistenzen gegen eine oder mehrere Anthelminthika-Klassen von ebenfalls
80 % bei dänischen Ziegenbetrieben nachgewiesen werden konnte (MAINGI et al.,
1996). Vereinzelt können jedoch auch bereits levamisol-, und ivermectinresistente

24
Stämme nachgewiesen werden und auch hier ist in Folge des gehäuften Einsatzes
dieser Wirkstoffgruppen eine weitere Zunahme zu erwarten (HERTZBERG u. BAUER,
2000). Gehäuft können resistente Stämme bei H. contortus, T. circumcinta und T.
colubriformis gefunden werden (HERTZBERG U. BAUER, 2000).
• Resistenzmanifestation
Die Manifestation von Resistenzen bleibt oft lange unbemerkt und wird oft erst im
Zusammenhang mit erheblichen wirtschaftlichen Verlusten und/oder klinischer Er-
krankung evident, also zu einem Zeitpunkt, zu dem sie bereits ein erhebliches Aus-
maß erreicht haben (VAN WYK, 2001, zitiert in VAN WYK et al., 2006). Zusätzlich
kann ein Wirkstoffwechsel bzw. die Kombination mehrerer Anthelminthika-Familien
das Vorhandensein von Resistenzen maskieren, so dass sie lange unerkannt bleiben
(VAN WYK, 2001, zitiert in VAN WYK et al., 2006). Sind Resistenzen jedoch in einer
Wurmpopulation vertreten, so sind diese irreversibel und fortwährend verankert, so
dass die resistenten Würmer einen starken selektiven Vorteil gegenüber empfängli-
chen Parasiten haben, sobald es zu einer weiteren Konfrontation mit der entspre-
chenden Wirkstoffgruppe kommt (KAPLAN, 2004).
• Detektion von Anthelminthika-Resistenzen
Gemessen werden können Resistenzen anhand des Faecal egg count reduction test
(FECRT), bei dem zuerst die ursprüngliche Eiausscheidungsrate vor einer Anthel-
minthika-Behandlung ermittelt, daraufhin mit einer Anthelminthika-Klasse entwurmt
und letztendlich die Eiausscheidungsrate nach erfolgter Behandlung bestimmt wird.
Die Reduktion der Eiausscheidung sollte dabei zwischen 95-100% liegen, bei einer
Reduktion von nur 90-95% kann eine Resistenz vermutet werden. Liegt der Wert
jedoch unter 90%, so muss von vorhandenen Resistenzen gegen die jeweilig verwen-
dete Wirkstoff gruppe ausgegangen werden (MCKENNA, 1990, zitiert in MAINGI et
al., 1996; REQUEJO-FERNANDEZ et al., 1997).
• Die Resistenzentwicklung begünstigende Faktoren
Viele Faktoren begünstigen die Ausbreitung von Resistenzen. Die Grundlage dafür
stellt die Biologie und Epidemiologie des jeweiligen Parasiten und die Wirt-
Parasiten-Beziehung dar (KAPLAN, 2004).

25
Entwurmungspraxis
Die Entwurmungspraxis hat einen äußerst maßgeblichen Einfluss auf das Ausmaß
und das Fortschreiten der Resistenzbildung. Einerseits besteht ein unmittelbarer Zu-
sammenhang zwischen der Anzahl durchgeführter Anthelminthika-Behandlungen
und auftretenden Resistenzen, d.h. je häufiger Tiere bereits anthelminthisch behan-
delt wurden, desto häufiger kann das Auftreten resistenter Strongyliden-Stämme be-
obachtet werden (PRICHARD et al., 1980, zitiert in MAINGI et al.; 1996; BARGER,
1999). Um die Anzahl der nötigen Behandlungen reduzieren zu können, ist es unter
anderem nötig, die maximale Wirksamkeit der Anthelminthika zu erzielen. Dafür ist
der Futterentzug vor und nach der geplanten Behandlung (zwölf Stunden vor bis ei-
nige Stunden danach) ratsam, der eine verlangsamte Passage der oral verabreichten
Anthelminthika und damit eine Effizienzsteigerung mit sich bringen soll (HERTZ-
BERG u. BAUER, 2000). Der Umkehrschluss, wonach selten behandelte Tiere auch
geringere Resistenzbildung aufweisen, ist hingegen nicht zulässig - entscheidend ist
hier die Art der Anwendung (Dosis, Zeitpunkt der Applikation etc.).
Wie bereits erwähnt, ist der Entwurmungszeitpunkt entscheidend. So wären Ent-
wurmungen zum Zeitpunkt eines geringen Ausmaßes an auf der Weide frei lebenden
Parasitenstadien (z.B.: im Spätwinter vor dem Austrieb auf „saubere" Weiden) im
Hinblick auf eine erfolgreiche Parasitenbekämpfung zwar sehr effizient, seitens der
Resistenzentwicklung jedoch unterliegen die Parasiten hierbei einem starken Selekti-
onsdruck hinsichtlich ihrer Anthelminthika-Unempfindlichkeit. Es überleben nur
resistente Parasiten, die sich nun fortpflanzen und ihr genetisches Material an ihre
Nachkommen weitergeben, so dass diese Maßnahme stark zur Beschleunigung einer
Resistenzausbreitung beiträgt (BARGER, 1999). JACKSON u. COOP (2000) raten dazu,
Behandlungen im gleichen Intervall wie die Präpatenzzeiten der Parasiten zu ver-
meiden, um die Verstreuung anthelminthika-sensibler Wurmeier zu gewährleisten
und empfehlen außerdem die Behandlungen dann durchzuführen, wenn die Parasi-
tenbelastung der Weide höher ist, als die der Wirtstiere (HOSTE et al., 2002a). Je
kleiner das Refugium an unbehandelten Tieren bzw. an unselektierten freilebenden
Parasitenstadien, desto wahrscheinlicher ist eine Resistenzentwicklung. Sehr klein ist
das Refugium beim „dose and move"-System, bei dem alle Tiere anthelminthisch
behandelt und erst nach erfolgter Therapie auf saubere Weiden ausgetrieben werden.
Somit wird das neue Weideland ausschließlich mit Parasiten und deren Entwick-
lungsstadien besiedelt, die dem Anthelminthika-Selektionsdruck bereits ausgesetzt

26
waren, was die Resistenzentwicklung äußerst begünstigt (PRICHARD et al., 1980, zi-
tiert in MAINGI et al. 1996; HERTZBERG U. BAUER, 2000).
Nachdem also, wie bereits erwähnt, die Resistenzentwicklung nachweislich um so
schneller und massiver erfolgt, je mehr Tiere innerhalb einer Population anthel-
minthisch behandelt werden, d.h. je kleiner die Nischen (Refugien) sind, in denen
Nematoden keiner Anthelminthika-Selektion ausgesetzt sind (HERTZBERG U. BAUER,
2000), empfiehlt es sich, einen Teil der Herde unbehandelt zu lassen und nur beson-
ders empfängliche oder bereits erkrankte Tiere zu behandeln. Dieses Vorgehen for-
dert zwar eine besonders intensive Überwachung der Herdengesundheit, kann aber
die Resistenzentwicklung nachhaltig verzögern (HERTZBERG U. BAUER, 2000).
Weidemanagement
Die bereits erwähnten Refugien werden jedoch nicht nur durch das Behandlungsre-
gime sondern auch durch das Weidemanagement maßgeblich beeinflusst (HERTZ-
BERG u. BAUER, 2002). Neuere Studien zeigen, dass die bisher empfohlene Strategie
des „dose-and-move"-Systems (z.B.: die penpartale Anthelminthika-Behandlung der
Muttertiere während des Winters mit anschließendem Austrieb auf „saubere" Wei-
den) mit der bereits erwähnten resultierenden Verkleinerung der verbleibenden Re-
fugien für Anthelminthika-sensible Nematoden erheblich zur Resistenzentwicklung
beiträgt. Ursache dafür ist, dass die „sauberen" Weiden ausschließlich mit Strongyli-
den besiedelt werden, die bereits eine Anthelminthika-Behandlung überlebt haben.
Wirkstoffwechsel
Des Weiteren tragen Faktoren wie der nicht vollzogene jährliche bzw. mehrmals
jährliche Wirkstoffwechsel der eingesetzten Antiparasitika bedeutsam zur Resistenz-
ausbreitung bei. Bei einer Umfrage in Milchziegenbetrieben in Frankreich wechselte
keiner der befragten Landwirte den eingesetzten Wirkstoff innerhalb oder zumindest
zwischen den Behandlungsjahren (HOSTE et al., 2000). Abgesehen vom jährlichen
Wechsel der Anthelminthika-Gruppe wird auch die Kombination von Anthelminthi-
ka-Wirkstoffgruppen empfohlen (MCKENNA et al., 1996, zitiert in HERTZBERG u.
BAUER, 2000).

27
Unterdosierung
Unterdosierung stellt einen weiteren Risikofaktor in der Anthelminthika-Behandlung
dar. Dieser Gefahr liegt die im Vergleich zu Schafen bei Ziegen deutlich schneller
ablaufende Anthelminthika-Clearance zugrunde, weshalb unbedingt die doppelte für
Schafe empfohlene Dosis verabreicht werden muss (GILLHAM U. OBENDORF, 1985,
zitiert in MAINGI et al., 1996). Dies ist unter den Landwirten jedoch häufig nicht be-
kannt, so dass z.B. nur 55 % der Landwirte in Frankreich die empfohlene doppelte
Schafdosis von Benzimidazolen und Probenzimidazolen verabreichen und bei Leva-
misol, Pyrantel und Avermectin sogar generell nur die einfache Schafdosierung an-
gewandt wird (HOSTE et al., 2000). Unterdosierungen entstehen zusätzlich durch
häufig zu gering geschätzte Körpermasse der Tiere. Ratsam ist daher, nach dem
schwersten Tier in der Herde zu dosieren (WALLER et al., 1995, zitiert in HOSTE et
al., 2002).
Einsatz von Breitbandanthelminthika
Der vermehrte Einsatz von Breitbandantiparasitika beschleunigt nachhaltig die Re-
sistenzentwicklung, so dass dem Einsatz von Präparaten mit engem Wirkspektrum in
jedem Fall der Vorzug gegeben werden sollte (WALLER et al., 1995, zitiert in HOSTE
et al., 2002a).
Tierverkehr
Nicht vergessen werden darf der Faktor Tierverkehr, der gleichermaßen zur Ausbrei-
tung von Resistenzen beitragen kann (HERTZBERG U. BAUER, 2000), indem die neu in
den Betrieb eingebrachten Tiere resistente Wurmpopulationen aus dem Herkunftsbe-
trieb in der neuen Herde verbreiten können. Nach der Studie von CABARET U. GAS-
NIER (1994) könnte die Herdenbildung mit Tieren vieler verschiedener Herkunftsbe-
trieben die Wahrscheinlichkeit auftretender Resistenzen erhöhen, ein negativer Zu-
sammenhang konnte hingegen bezüglich des Herdenalters nachgewiesen werden
(SiLVESTRE et al., 2000). Insbesondere der Zukauf von Zuchtböcken, wie er auch in
den im Rahmen dieser Arbeit untersuchten Betrieben praktiziert wird, muss berück-
sichtigt werden. Vor der Integration eines zugekauften Tieres in die bestehende Her-
de wird deshalb die Quarantäne unter begleitender Durchführung von Kotuntersu-
chungen empfohlen. Wenn nötig, sollen Avermectin-Behandlungen vorgenommen
werden, bis negative Kotbefunde erreicht sind (HERTZBERG U. BAUER, 2000).

28
Intensive Haltungssysteme
Wie bereits erwähnt, ist ein gehäuftes Resistenzaufkommen in intensiveren Hal-
tungsformen zu beobachten (HERTZBERG U. BAUER, 2000). Ursache dafür könnte die
in intensiven Haltungsformen durch höhere Besatzdichte entstandene, höhere Parasi-
tenprävalenz (WALLER, 2006a) und daraus resultierende häufigere Anthelminthika-
Behandlungen sein (siehe auch im Kapitel 2.5.1).
In Anbetracht der in manchen Regionen bereits zum jetzigen Zeitpunkt prekären Re-
sistenzsituation muss es oberstes Ziel sein, die derzeitige Anthelminthika-
Abhängigkeit durch ein nachhaltigeres und umfassenderes Parasitenmanagement zu
ersetzen, d.h. unter Berücksichtigung der Parasitenepidemiologie durch biologische
Kontrollmechanismen, Entwicklung von Vakzinen und züchterische Maßnahmen
hinsichtlich resistenter Wirtstiere zu ergänzen (BARGER, 1999), um die Anzahl an
nötigen Behandlungen mit synthetischen Antiparasitika auf ein Minimum reduzieren
und die Resistenzentwicklung so verzögern zu können.
2.5.2 Selektiver Anthelminthika-Einsatz als möglicher Lösungsansatz
Dieses selektive Behandlungsmodell könnte vor allem in Zukunft eine viel verspre-
chende, kurzfristige Lösung des Endoparasitenproblems bei kleinen Wiederkäuern
durch die Kombination von Nematodenkontrolle mit Managementstrategien bzgl.
Anthelminthika-Resistenzen unter Aufrechterhaltung der Milchproduktion darstellen
(HOSTE et al., 2002c).
Die Basis für die Entwicklung der selektiven Anthelmintika-Behandlung, d.h. der
Behandlung ausschließlich besonders gefährdeter, stark verwurmter und/oder kli-
nisch durch schlechtere Körperkondition auffälliger Tiere, also den primären Risiko-
gruppen, bilden die bereits im Zusammenhang mit der Bedeutung des Alters und der
Milchleistung erwähnten Erkenntnisse, wonach die Empfänglichkeit gegenüber Ne-
matodeninfektionen bei Jungtieren, Erstlaktierenden und Hochleistungstieren als
besonders hoch einzuschätzen ist (HOSTE U. CHARTER, 1993, zitiert in HOSTE et al.,
2002a, 2002d). Ein Teil der Herde (ein Drittel bis die Hälfte) bleibt im Zuge dieses
gezielten Behandlungsregimes unbehandelt mit der Intention, die Sensibilität der
Endoparasiten auf genetischer Ebene zu erhalten und eventuell bereits vorhandene
Resistenzgene zu „verdünnen" (LEATHWICK et al., 1995, zitiert in HOSTE et al.,
2002a) bzw. deren Ausbreitung zu verzögern (VAN WYK et al., 2006). Wie groß der
Anteil unbehandelter Tiere sein muss, um das angestrebte Ziel des Gleichgewichts

29
aus wirksamer Parasitenkontrolle und gleichzeitiger Verzögerung der Resistenzent-
wicklung zu erhalten, kann nach derzeitigem Wissensstand noch nicht exakt angege-
ben werden, so dass es zusätzlicher Studien bezüglich idealer Proportionen innerhalb
einer Herde bedarf (HOSTE et al., 2002c). Die unbehandelten Tiere stellen ein „Refu-
gium für anthelminthika-sensible Nematodenstämme" (LEATHWICK et al., 1995; VAN
WYK, 2001, zitiert in KOOPMANN et al., 2006) dar, nachdem ein Teil der Endoparasi-
ten nicht dem Selektionsdruck einer Antiparasitika-Behandlung unterliegt. Diese
Maßnahme ist hinsichtlich einer angestrebten Verzögerung bzw. Vermeidung der
Weiterentwicklung von Anthelminthika-Resistenzen von größter Bedeutung, die
besonders bei kleinen Wiederkäuern in den letzten Jahren bereits bedrohliche Aus-
maße angenommen haben. Oberstes Ziel sollte deshalb sein, möglichst wenige Tiere
möglichst selten und mit maximaler Effizienz zu behandeln (HOSTE et al., 2002a).
Wie bereits im Punkt Befallsintensität erörtert, kann davon ausgegangen werden,
dass die Verteilung der Wurmbürden innerhalb eines Bestandes ungleich ist und es
einige Tiere gibt, die höhere Eiausscheidungsraten aufweisen als andere (HOSTE et
al., 2001a, 2002d). HOSTE et al. (2001a) wiesen nach, dass hier eine starke Wieder-
holbarkeit vorliegt bzw. innerhalb einer Herde immer dieselben Tiere sehr hohe
Wurmbürden tragen. So sind 10 % der Ziegen für 25 % der ausgeschiedenen Eier
verantwortlich (HOSTE et al., 2002a). Diese Erkenntnis ist sowohl aus genetischer
Sicht im Sinne der züchterischen Selektion resistenter Tiere als auch im Hinblick auf
die Auswahl der Tiere für selektive Anthelminthika-Behandlungen bedeutsam (BAR-
GER u. DASH, 1987, zitiert in HOSTE et al., 2001a). Als ökonomischer Nebeneffekt
kann die dadurch erzielte Reduktion der eingesetzten Medikamente zu nicht unerheb-
lichen Einsparungen beitragen und entspricht noch dazu den Grundsätzen des Öko-
landbaus, wonach Medikamente generell nur restriktiv einzusetzen sind.
Die Methodik der selektiven Behandlung wirkt sich laut HOSTE et al. (2002, 2002c,
2002d) in keiner Weise nachteilig auf die Milchleistung oder auf die Eiausschei-
dungsrate aus. In vorangegangenen Studien lieferte das selektive Behandlungsregime
sogar gleich effiziente Ergebnisse wie das systematische (HOSTE et al., 2002, 2002c)
und resultierte in deutlich reduziertem Selektionsdruck hinsichtlich der Entwicklung
von Anthelminthika-Resistenzen (HOSTE et al., 2002d). Allerdings muss es umso
intensiver von gutem Management und sorgfältiger Herdengesundheitsüberwachung
begleitet werden, um krankheitsbedingten Verlusten vorzubeugen (KAHN U. WAT-
SON, 2003).

30
• Identifikation der behandlungswürdigen Tiere
Voraussetzung für die selektive Behandlung ist die Identifikation behandlungswürdi-
ger Einzeltiere oder auch Tiergruppen - ein Voraussetzung, die zugleich ein Problem
darstellt, gibt es doch bis zum heutigen Zeitpunkt noch keine praxisrelevante Mög-
lichkeit zur verlässlichen Detektion besonders infektionsgefährdeter bzw. erkrankter
Tiere, die gleichzeitig effizient, ohne größeren Zeitaufwand und vor allem kosten-
günstig durchführbar ist. Die Koproskopie (= Kotuntersuchung) ist bei kleinen Wie-
derkäuern aufgrund der großen Anzahl an zu untersuchenden Proben und damit ver-
bundenen Kosten nicht praxistauglich. Die FAMACHA®-Methode (d.h. die Detekti-
on behandlungswürdiger Tiere durch Beurteilung der Schleimhautfarbe bezüglich
einer klinisch apparenten Anämie; siehe Kapitel 5.9) ist zwar kostengünstig und des-
halb auch in größeren Herden durchführbar, jedoch nur zur Detektion blutsaugender
und in der Folge Anämie verursachender Parasiten wie H. contortus, weniger jedoch
zur Identifikation von Infektionen mit T. circumcinta und T. colubriformis geeignet,
die ihrerseits in Studien die am häufigsten gefundenen Parasiten in französischen
Milchziegenbetrieben darstellen (CABARET U. GASNffiR, 1994, zitiert in HOSTE et al.,
2002a).
Die Faktoren Alter und Laktationsstadium sind jedoch mögliche Parameter zur Ein-
grenzung von Risikogruppen, da - wie bereits erwähnt - vor allem Jungtiere,
Erstlaktierende und Hochleistungstiere als besonders empfänglich und gefährdet ein-
gestuft werden müssen. Eine gezielte Behandlung dieser Tiergruppen könnte die Ef-
fizienz eines Parasitenmanagements demnach deutlich erhöhen und ist hinsichtlich
ökonomischer Aspekte (geringere Einbußen in der Milchleistung) ebenso empfeh-
lenswert (HOSTE et al., 2002a, 2002b). Besonders zu beachten ist hierbei allerdings,
dass die Klassifizierung der Tiere nach ihrer Laktationsleistung zu einem Zeitpunkt
erfolgen muss, in dem die Befallsintensität möglichst niedrig ist, nachdem die Milch-
leistung der Hochleistungstiere besonders stark von Endoparasitosen beeinträchtigt
und reduziert wird und die Milchmenge somit leicht falsch interpretiert werden könn-
te (HOSTE et al., 2002a).
Wie im Kapitel 5.4 eingehend erörtert, können plötzlicher Gewichtsverluste und Va-
riationen bezüglich des BCS zusätzlich hilfreiche Indizien für einen vorliegenden
Parasitenbefall sein (VAN WYK et al., 2006), es muss jedoch berücksichtigt werden,
dass diese Parameter durch viele Faktoren beeinflusst werden können und deshalb im
Gesamtzusammenhang gesehen werden müssen bzw. isoliert betrachtet, wie auch in

31
den hier untersuchten Betrieben, keine verlässliche Aussage hinsichtlich eines Para-
sitenbefalls erlauben.
Zusammenfassend kann gesagt werden, dass das selektive Behandlungsmanagement
eine viel versprechende Option zur Parasitenkontrolle unter gleichzeitigem Resis-
tenzmanagement darstellt. Es muss allerdings mit anderen alternativen oder kom-
plementären Methoden wie Futterergänzung und Weidemanagement oder auch sys-
tematischer Parasitenbehandlung im Winter zum Zeitpunkt der Aufstauung kombi-
niert werden, um optimale Resultate zu erreichen (BARGER, 1999; HOSTE et al.,
2002c). Weitere diesbezügliche Forschungen sind in Anbetracht der bedrohlichen
Resistenzentwicklung, speziell bei den kleinen Wiederkäuern, dringend nötig.
2.6 Alternative Behandlungsmethoden
Der ökologisch sensibilisierte Konsument, Tierschutzaspekte, die Angst vor Medi-
kamentenrückständen in Lebensmitteln, die aus intensiven Haltungsformen resultie-
rende Mistentsorgungsproblematik, Treibhausgas-Emissionen, durch Bevölkerungs-
zunahme bedingter Platzmangel und die Bestrebungen zur Renaturierung ehemaliger
Weideflächen - all diese Aspekte bedingen ein - zum Teil bereits erfolgtes oder zu-
mindest in nächster Zukunft unumgängliches - Umdenken in der Tierproduktion.
Dieser Prozess beeinflusst nicht zuletzt auch die Vorgehensweise in Bezug auf die
Endoparasitenkontrolle, die in der Vergangenheit stark vom Einsatz chemischer An-
tiparasitika geprägt war (WALLER, 1999).
Die bereits im Kapitel 2.5.1 erörterte, weltweit zunehmende Bedrohung der Tierpro-
duktion, insbesondere der kleinen Wiederkäuer, durch Zunahme an Resistenzen ge-
gen die handelsüblichen Anthelminthika-Wirkstoffgruppen unterstreicht zusätzlich
die Notwendigkeit, alternative Methoden zur Endoparasitenkontrolle bzw. ein integ-
riertes Parasitenmanagement zu entwickeln (WALLER, 1999).
2.6.1 Homöopathie und Phytotherapie (nach CABARET et al., 2002)
Die Effizienz homöopathischer bzw. phytotherapeutischer Behandlungsansätze bei
Endoparasitosen konnte bisher noch nicht ausreichend wissenschaftlich belegt wer-
den. Nichtsdestotrotz wird vielen natürlichen Substanzen eine entweder direkte oder
- im Fall der später eingehender behandelten Homöopathika - indirekte anthel-
minthische Wirkung zugesprochen. ^

32
Nach heutigem Wissensstand können jedoch weder Phytotherapeutika noch Homöo-
pathika den Einsatz anderer Kontrollmechanismen ersetzen und sollten deshalb vor
allem unterstützend bzw. ergänzend eingesetzt werden. Erstaunlich ist, dass der Ein-
satz von Pflanzenextrakten bisher nicht den Regelungen bezüglich Medikamenten-
rückstände unterworfen ist, obwohl weithin bekannt ist, dass der Anteil des Wirkstof-
fes von Pflanze zu Pflanze variieren kann. So könnten etwa manche Wirkstoffe so-
wohl für das Tier als auch für den Menschen in Form von Lebensmittelrückständen
toxisch sein und die Qualität tierischer Produkte sogar verändern (z.B.: Trigonella
foenum-graecum in Milch und Fleisch) (CABARET, 1986). Weitere dahingehende
Untersuchungen sind auch hier geboten.
2.6.1.1 Phytotherapie
Phytotherapeutische Ansätze in Mexiko mit monatlicher Verabreichung von Cheno-
podium ambrosoides gegen Endoparasiten in Kombination mit einer Optimierung
des allgemeinen Managements bei Schafen (weniger Tiere nach Selektion der pro-
duktionsstärksten Tiere, verbesserte Fütterung mit lokal verfügbaren Futtermitteln,
bessere Überwachung zum Zeitpunkt des Ablammens) reduzierten die Sterblichkeit
insgesamt von 15 % auf 2 %, innerhalb der Muttertiere, von 18 % auf 8 % innerhalb
der Lämmer. Allerdings können die verbesserten Resultate durch die Kombination
verschiedener Maßnahmen nicht eindeutig auf den Einsatz eines Phytotherapeuti-
kums zurückgeführt werden.
Nach CABARET (1986) wird ebenso Nicotiana tabacum bei Schafen sowie Allium
sativum, dies jedoch nur eingeschränkt, eine anthelminthische Wirkung zugespro-
chen. In Wales und Irland wurde Dryopteris filix-mas gegen F. hepatica bei Wieder-
käuern eingesetzt. Bei Rindern wird des Weiteren noch Asarum europaeum gegen
Würmer allgemein und Pinus maritima gegen F. hepatica und Rundwürmer als
wirksam angegeben. Ein breit gefächertes antiparasitäres Wirkspektrum (gegen Ek-
toparasiten, Coccidien und gastrointestinale Strongyliden) zeigt auch Azadirachta
indica, der Neem-Baum (PIETROSEMOLI et al., 1999, zitiert in CABARET et al., 2002).
Problematisch ist häufig jedoch die potentielle Toxizität der erwähnten Pflanzen, so
dass weitere Untersuchungen bezüglich Wirksamkeit, Toxizität und Umweltverträg-
lichkeit unumgänglich sind (CABARET, 1986).

33
2.6.1.2 Homöopathische Therapieformen
Verschiedenste Homöopathika wurden bereits auf ihre direkte anthelminthische
Wirksamkeit untersucht, darunter Arremw/a cina, Filix mas, Allium sativum, Kamala,
Areca catechu und Solidago, eine nachweisliche Reduktion der Eiausscheidungsrate
konnte jedoch bei keiner der Substanzen verzeichnet werden (SCHüTTE, 1994).
Vielmehr scheint sich deren Wirksamkeit auf eine indirekte Komponente zu be-
schränken, wonach sie die „Resilience" des Wirtstieres, d.h. die Fähigkeit, die Infek-
tion zu kontrollieren, negative Auswirkungen zu minimieren bzw. die Produktions-
leistung aufrecht zu erhalten, steigern.
2.6.2 Kupfer
Die anthelminthischen Eigenschaften von Kupfer, insbesondere gegen im Aboma-
sum parasitierende Nematoden und hier besonders gegen H. contortus, sind bereits
seit der Antike bekannt und wurden bis zur Entwicklung erster modemer Anthel-
minthika um 1930 genützt (WALLER, 1999). Danach geriet Kupfer in Anbetracht der
wachsenden Vielfalt von Anthelminthika in Vergessenheit.
Mit zunehmender Resistenzproblematik erlebt der Einsatz von Kupfer in Form von
Kupferoxyd-Draht-Partikeln (copper-oxide wire particles) nun eine Renaissance.
Nach BANG et al. (1990) erfolgt die Verabreichung in Kapseln, die sich im Pansen
auflösen, woraufhin die Kupferpartikel weiter in das Abomasum transportiert wer-
den, sich dort in Schleimhautfalten anlagern und über bis zu drei Monate Kupfer-
Ionen freisetzen. Ergebnisse von Studien ergaben eine Reduktion der Infektionsrate
von 96 % bei H. contortus und 56 % bei T. circumcinta, zeigte jedoch keinerlei Ef-
fekt auf intestinale Parasitenformen. Aufgrund seiner offensichtlichen Effizienz, der
langen Wirkdauer, den geringen Kosten und der sicheren Anwendung scheint Kupfer
eine viel versprechende Alternative zu synthetischen Anthelminthika zu sein, die
jedoch noch weiterer Feldstudien bedarf (aus WALLER, 1999).
2.6.3 Biologische Kontrollverfahren
Ziel der biologischen Parasitenkontrolle ist es, die Menge frei lebender Parasitensta-
dien soweit zu reduzieren, dass sie einerseits keine gesundheitliche und/oder ökono-
mische Gefahr darstellen, andererseits ist es das Ziel, die Etablierung einer adäquaten
Immunität zu ermöglichen (THAMSBORG et al., 1999).

34
Zur biologischen Bekämpfung der frei lebenden Stadien auf den Weideflächen liefer-
te der Einsatz nematophager Pilze, besonders von Duddingtonia ßagrans, viel ver-
sprechende Ergebnisse. Drei besondere Eigenschaften zeichnen diesen Pilz aus: so
übersteht er zum einen die Darmpassage unbeschadet, zum zweiten weist er in fri-
schem Kot ein rasches Wachstum auf zum dritten zeichnet ihn ein breites Wirkspekt-
rum gegen verschiedenste Nematodenlarven aus (LARSEN, 1999, zitiert in WALLER U.
THAMSBORG, 2004). Nachteilige Wirkungen auf die Umwelt sind nach Langzeitstu-
dien außerdem keine bekannt (WALLER U. THAMSBORG, 2004). Der Einsatz nema-
tophager Pilze dient primär der Verhinderung von Reinfektionen im Verlauf der
Weidesaison, nicht jedoch der Unterbindung von Frühjahrsinfektionen durch über-
winterte Larven (HERTZBERG U. BAUER, 2000). Außerdem richtet sich der derzeit
untersuchte Einsatz nematophager Pilze ausschließlich gegen Larvenstadien im Kot,
hingegen nicht gegen infektiöse Eier (THAMSBORG et al., 1999).
Die zwei bis drei Wochen nach dem Weideaustrieb über einen Zeitraum von zwei bis
drei Monaten (also über den zu erwartenden Hochsommer-Peak hinaus) mit dem
Kraftfutter verfütterten Pilzsporen bilden unmittelbar nach dem Kotabsatz engma-
schige Netze im frischen Kot, in denen sich die Larven der Strongyliden und Lun-
genwürmer verfangen und daraufhin absterben (GR0NVOLD et al., 1993). Dies hat zur
Folge, dass die Wahrscheinlichkeit des Netzkontaktes bei mobileren Larven höher
ist. Weiters wird auch angenommen, dass die Netzbildung durch den Larvenkontakt
stimuliert wird, so dass das Ausmaß der Netzbildung maßgeblich von der Motilität
der Larven bestimmt wird und demnach je nach Larvenspezies differieren kann
(GR0NVOLD et al., 1996, zitiert in WAGHORN et al., 2003). Bereits 24 Stunden nach
Ausbringen der Sporen konnte eine signifikante Reduktion der sich entwickelnden
Larven im Kot nachgewiesen werden (WAGHORN et al., 2003). Dosisabhängig zeigte
sich dieser Effekt nur bei T. circumcinta, während H. contortus und T. colubriformis
nur gering von der Sporenmenge beeinflusst zu werden schienen (WAGHORN et al.,
2003).
Nematophage Pilze sind für die Zukunft als „Breitband-Anthelminthikum" sehr viel-
versprechend, da sie ubiquitär vorkommen (WALLER, 2003, zitiert in WALLER U.
THAMSBORG, 2004), effizient und gegen viele Spezies in gleichem Ausmaß wirksam
(WAGHORN et al., 2003) und gleichzeitig einfach kultivierbar und prinzipiell leicht
auszubringen sind, wobei die Applikationsart hinsichtlich ihrer Praxistauglichkeit
und ökonomischer Belange noch optimiert werden muss (THAMSBORG et al., 1999;

35
WAGHORN et al., 2003). Diesbezüglich bietet sich die Verabreichung als Futterergän-
zung an, wobei diese täglich erfolgen muss, um eine gleichmäßige Versorgung mit
Pilzsporen zu gewährleisten. Von praktischer Relevanz wäre deshalb die Entwick-
lung von effizienten Boluspräparaten, die eine kontinuierliche Sporenfreigabe über
mindestens zwei Monate aufrechterhalten (WALLER U. THAMSBORG, 2004). Derzeit
stehen jedoch noch keine derartigen Produkte zur Verfügung.
Beim Einsatz nematophager Pilze muss unbedingt von einer Kombination mit kon-
ventionellen Anthelminthika abgesehen werden, da das Verhältnis der anthelminthi-
ka-exponierten Parasitenstadien zu Ungunsten der frei in der Umwelt vorkommenden
Stadien erheblich ansteigt und damit auch der Selektionsdruck für resistente Parasi-
ten wächst. Dies könnte entscheidend zu einer beschleunigten Resistenzausbreitung
beitragen, weshalb nematophage Pilze anstelle synthetischer Anthelminthika einge-
setzt werden sollten (WAGHORN et al., 2003).
2.7 Optimierung von Managementfaktoren und präventive Maßnahmen
2.7.1 Weidemanagement
Die Umwelt, insbesondere die Weide, bildet das Bindeglied zwischen frei lebenden
und parasitierenden Parasitenstadien (WALLER, 2006a). Umso bedeutsamer ist dem-
nach das Weidemanagement, das im Grundgedanken der biologischen Landwirt-
schaft die Basis für eine nachhaltige Parasitenkontrolle bildet. Es kann in verschiene
Systeme eingeteilt werden: 1. Präventivmaßnahmen, bei denen z.B. nur uninfizierte
Tiere auf eine „saubere" Weide verbracht werden, dazu gehört jedoch auch späterer
Weidegang, Weidewechsel zwischen den Jahreszeiten und die Verwendung von
mehr Spätheu; 2. „evasive", also ausweichende Strategien, die nicht darauf abzielen,
die Weidekontamination von vornherein zu unterbinden, sondern die Infektionsge-
fahr durch einen Weidewechsel (mit oder ohne begleitender Anthelminthika-
Behandlung) innerhalb der Weidesaison auf eine „saubere" Weide noch vor Schlüp-
fen einer signifikanten Anzahl von Larven auf der ursprünglichen Weide zu reduzie-
ren versuchen; 3. „Verdünnungsstrategien", wobei entweder empfängliche (z.B.:
Erstlaktierende) mit resistenten Tieren (z.B. trockenstehende, adulte Tiere) der sel-
ben Tierspezies bewusst gemischt werden, die Weide gleichzeitig oder abwechselnd
mit anderen Spezies (z.B. Rinder, Pferde) beweidet, die Besatzdichte gesenkt

36
und/oder die Weideperiode verkürzt wird; und 4. Supplementierung der Fütterung im
Frühjahr (MICHEL, 1985, zitiert in CABARET et al., 2002; BARGER, 1997, zitiert in
THAMSBORG et al., 1999; WALLER, 2006a).
Damit eine Weide als „sauber" bezeichnet werden kann, muss sie eines der folgen-
den Kriterien erfüllen: sie muss 1. entweder bis dato unbeweidet gewesen sein; 2. im
Falle einer „Altweide" einer Wechselbeweidung mit anderen Tierspezies, die ein
anders Parasitenspektrum aufweisen, unterzogen worden sein; 3. für eine Zeit brach
liegen, wobei die dafür nötige Zeitspanne je nach Gegebenheiten variabel ist
(THAMSBORG et al., 1999).
• Populationsdvnamik der Parasiten
Die Grundlage guten Weidemanagements bildet das Wissen um die Populationsdy-
namik frei lebender infektiöser Parasitenstadien, insbesondere die Zeitspanne ab
Austrieb, in der mit der Etablierung eines gefährlichen Infektionsdrucks zu rechnen
ist bzw. die Zeitspanne nach dem Umtrieb, nach der die Weidefläche wieder als „si-
cher" angesehen werden kann. Dies unterliegt jedoch einer jahreszeitlichen Variati-
on, wonach Weideflächen im Mai und Juni nach drei Wochen Beweidung, von Juli
bis September bereits nach zwei Wochen als infiziert betrachtet werden müssen
(EYSKER et al., 2005). Besonders bedeutsam ist der massive Anstieg des Infektions-
drucks von Haemonchus, was die Etablierung hoher Wurmbürden innerhalb vier
Wochen zur Folge haben kann (EYSKER et al., 2005). Daraus resultiert die zwingende
Notwendigkeit, die Tiere zumindest in den Sommermonaten im Intervall von zwei,
sonst drei Wochen auf eine saubere Weidefläche umzutreiben, um eine Reinfektion
wirksam reduzieren zu können, was in vielen Betrieben jedoch aufgrund fehlender
Infrastruktur in dieser Form schwierig ist.
Des Weiteren spielt hier die frühzeitige Bekämpfung der im Kapitel 5.5.2 behandel-
ten saisonalen Peaks eine große Rolle, aus denen sich die tatsächliche Infektionsrate,
d.h. die tägliche Aufnahme infektiöser Parasitenstadien während des Grasens, ergibt.
Möglichkeiten der Nutzung der saisonalen Schwankungen in der Anthelminthika-
Therapie werden in Kapitel 5.7 behandelt.
• Umtrieb der Jungtiere auf eigene Weiden
Einen weiteren interessanten Aspekt ergaben Untersuchungen in französischen
Fleischschafbetrieben, wonach die Aufzuchterfolge bei Lämmern wesentlich damit

37
korrelieren, wie lange die Jungtiere zusammen mit den Muttertieren weiden bzw. zu
welchem Zeitpunkt sie von den Mutterschafen getrennt und auf saubere Weiden um-
getrieben werden. So zeigte das Umtreiben der Lämmer auf saubere Weiden mit Mit-
te Juni die besten Produktionserfolge, während die Trennung mit Anfang Juli
schlechte, mit Ende Juli/ Mitte August sehr schlechte Ergebnisse erbrachte, was nicht
zuletzt auf Strongyliden-Infektionen zurückgeführt werden konnte (MAGE et al.,
1998, zitiert in CABARET et al., 2002). Auch in dänischen Schafbetrieben konnten
mittels Entwöhnen der Lämmer mit Anfang Juli und Umgruppieren der Jungtiere
noch vor dem zu erwartenden Infektionsanstieg während des Hochsommers auf sau-
bere Weiden gute Produktionsresultate erzielt werden (GiTHiGiA, 2001, zitiert in CA-
BARET et al., 2002). Diese Maßnahme dient der aktiven Prävention parasitär-
bedingter Gastroenteritis und trägt auf diesem Weg - unabhängig von begleitenden
Anthelminthika-Anwendungen - zu verbesserten Produktionsergebnissen bei (CA-
BARET et al., 2002).
• Besatzdichte
Gute Ergebnisse können mittels Weidemanagement in kleinen Betrieben mit niedri-
ger Besatzdichte und in Kombination mit anderen Maßnahmen (z.B. selektive
Anthelminthika-Behandlung) in Form eines ganzheitlichen Parasitenmanagements
erzielt werden (EYSKER et al., 2005). In mehreren Studien konnte eine direkte Bezie-
hung zwischen steigender Besatzdichte und steigendem Parasitismus nachgewiesen
werden (WALLER, 2006a). Der positive Effekt einer niedrigen Besatzdichte beruht
neben der damit verbundenen niedrigeren Morbidität wohl nicht zuletzt darauf, dass
die dem Einzeltier in höherem Maße zur Verfügung stehende Futtermenge eine bes-
sere Nährstoffversorgung gewährleistet und sich so positiv auf die potentielle Wirts-
abwehr gegenüber Parasiteninfektionen auswirkt (THAMSBORG et al., 1999). Die ex-
akte Begründung für diesen Zusammenhang konnte jedoch noch nicht gefunden
werden.
• Weidebeschaffenheit und Graseverhalten
Weidebedingungen, die das arttypische Knabbern an Büschen und Bäumen erlauben,
wirken sich sehr positiv im Sinne einer geringeren Nematodenbelastung aus. Zum
einen fördern sie die Aufnahme zum Teil toxischer Sekundärstoffe (anthelminthisch
besonders bedeutsam sind kondensierte Tannine, auf die im Kapitel 2.7.2.3 näher

38
eingegangen werden soll), zum anderen gehen sie aufgrund der extensiven Haltungs-
form üblicherweise mit niedrigerer Besatzdichte und dadurch reduziertem Infektions-
risiko einher (HOSTE et al., 2001, 2005).
• Altemierende Beweidung
Eine alternierende Beweidung durch verschiedene Tierspezies, z.B. kleine Wieder-
käuer im Wechsel mit Rind oder auch Pferd, reduziert nachhaltig den Infektions-
druck, wobei man sich hierbei die Wirtsspezifität der Endoparasiten zu Nutze ma-
chen kann. Auch in neuseeländischen Schafbetrieben führte die alternierende Bewei-
dung mit Rindern zu guten Ergebnissen (NiEZEN et al., 1996). Dabei wurde die zur
Verfügung stehende Weidefläche in zwei Teile unterteilt. Die entwöhnten Lämmer
wurden gemeinsam mit den Rindern auf eine der beiden Weiden ausgetrieben, wäh-
rend die Muttertiere bis zum nächsten Ablammen die andere Weide beweideten. Der
für die Lämmer abgegrenzte Bereich auf der Rinderweide wurde im Abstand von ein
bis zwei Wochen gewechselt, so dass die begrasten Paddocks einer Ruhephase von
zumindest 60 Tagen unterlagen und in der Zwischenzeit von Rindern beweidet wur-
den. Zum Zeitpunkt des Ablammens wurden die Tiere auf die jeweils andere Weide
und danach monatlich umgetrieben (aus CABARET et al., 2002). Mit dieser Methode
konnte der Einsatz von Anthelminthika bei jungen Schafen von 12 bis 24 auf nur ein-
bis zweimalige Behandlungen pro Jahr limitiert werden (BARGER U. SOUTHCOTT,
1978, zitiert in WALLER, 1999). Bezüglich des optimalen Zeitpunktes des Wirts-
wechsels und der Länge der Beweidungsintervalle können jedoch keine pauschalen
Empfehlungen gegeben werden, nachdem die Vorgehensweise an lokale Begeben-
heiten (Parasitenspektrum und -Intensität, klimatische Bedingungen etc.) angepasst
werden muss (BARGER, 1999). BARGER U. SOUTHCOTT (1978) empfehlen jedoch ei-
nen Wirtswechsel im Intervall von zwei bis sechs Monaten (zitiert in WALLER,
2006a).
Nachteilig konnte beobachtet werden, dass sich manche zuvor wirtsspezifischen Pa-
rasiten infolge der Konfrontation mit einer anderen Spezies an diese anpassen und es
somit zur Infektion mit speziesunüblichen Parasiten kommen kann (BARGER, 1997,
zitiert in WALLER 2006a). Diesbezügliche Studien sind unbedingt nötig, um davon
ausgehende Gefahren abschätzen zu können.

39
• Rotationsbeweidung bzw. System der Wechselweide
Sofern die Infrastruktur eines Betriebes dies zulässt, kann auch das Prinzip des „Ro-
tational grazing" als ergänzende Maßnahme zur nachhaltigen Parasitenkontrolle ein-
gesetzt werden. Dabei wird die Gesamtweidefläche beispielsweise 15 Paddocks un-
terteilt, wobei immer ein Paddock für die Zeitspanne von nur einer Woche beweidet
wird. Danach werden die Tiere auf den nächsten Paddock umgetrieben, so dass der
jeweils beweidete Paddock insgesamt für 14 Wochen ruhen kann. Der Rotationszyk-
lus beträgt demnach 15 Wochen. Allerdings muss bedacht werden, dass eine Weide
in gemäßigten Klimazonen oft erst nach drei bis neun Monaten zu einer infektiösen
Weide wird und somit ein Intervall von 15 Wochen zu kurz wäre, um das daraus
entstehende Infektionsrisiko abzuwenden (BARGER, 1999), weshalb sich diese Me-
thode primär für tropische Klimazonen eignet (THAMSBORG et al., 1999), wo die In-
tervalle aufgrund der kürzeren Entwicklungszyklen der Parasiten auch verkürzt wer-
den können bzw. sogar müssen (drei bis vier Tage Beweidung mit darauffolgender
einmonatiger Ruhephase derselben Weide) (BARGER, 1999; WALLER U. THAMSBORG,
2004).
2.7.2 Fütterung
Ein wesentlicher Faktor zur Prävention gastrointestinaler Nematodeninfektionen ist
die Fütterung. So scheint eine direkte Korrelation zwischen dem Ausmaß der De-
ckung des Nährstoffbedarfs und der immunologischen Belastbarkeit bzw. Wider-
standsfähigkeit (siehe Kapitel 5.8) zu bestehen (HOSTE et al., 2008). Auch das Aus-
maß der negativen Auswirkungen einer Parasiteninfektion kann mittels Ernährung
moduliert werden, steht aber primär in direkter Relation mit der Anzahl im Wirtstier
vorhandener Parasiten (WALLER, 2006a). Die fütterungsassoziierte Endoparasiten-
kontrolle kann dabei drei mögliche Strategien verfolgen: 1. die Immunantwort des
Wirtstieres zu verbessern; 2. bei der Nahrungsaufnahme den Kontakt mit infektiösen
Parasitenstadien zu vermeiden; und 3. Pflanzen auszuwählen, die direkte anthel-
minthische Eigenschaften aufweisen (HOSTE et al., 2008).
Die Supplementierung der Nahrung, also die ergänzende Zufütterung gezielter Nähr-
stoffe, stellt einen wesentlichen Aspekt hinsichtlich der Resistenzsteigerung und der
Fähigkeit des Wirtstieres dar, eine Infektion abwehren oder zumindest deren nachtei-
ligen Effekt kontrollieren und minimieren zu können (KNOX et al, 2006; HOSTE et
al., 2008). Besonders relevant ist dies in Zeiten besonders großen Nährstoffbedarfs

40
und/oder Mangels an Proteinen, Energie, Mineralstoffen oder Spurenelementen, wo-
bei der Nährstoffbedarf besonders in den Wachstumsphasen, in der Trächtigkeit -
besonders vor und nach der Geburt - und in der Laktation immens erhöht ist. Zusätz-
lich ist in diesen Zeiten, wie schon im Kapitel 5.8 beschrieben, die Immunantwort
des Wirtstieres vorübergehend stark herabgesetzt, was bei Schafen als peri-
parturient relaxation of immunity (PPRI) bezeichnet wird und bereits in der Spätlak-
tation bzw. auch noch zu Laktationsbeginn beobachtet werden kann (TEMBELY et al.
1998, zitiert in HOSTE et al., 2008). Die Wachstumsphase stellt gleichermaßen hohe
Anforderungen an die Ernährung. So konnten bei der Supplementierung von Kitzen
messbar höhere Werte an peripheren eosinophilen Granulozyten und eine insgesamt
verbesserte Widerstandsfähigkeit gegen Parasiteninfektionen nachgewiesen werden
(TORRES-AcosTA et al., 2004, zitiert in HOSTE et al., 2008).
Somit ist die Unterstützung des Immunsystems im Falle nutritiver Engpässe zur
Vermeidung eines Peaks der Endoparasitenbelastung und einer damit verbundenen
massiven Kontamination der Umwelt bzw. zur Senkung der Eiausscheidungsraten
umso bedeutsamer. Diesbezüglich hat sich die Verbesserung der Wirtsemährung als
eine gute Option zur Verbesserung der Fähigkeit des Wirtes zur Parasitenkontrolle
erwiesen und auch, aber in geringerem Maße, der Resistenz erwiesen (HOSTE et al.,
2002a, 2005). Eine ganzjährige Zufütterung erbrachte dabei die besten Produktions-
leistungen. Die Menge der Zufütterung muss dabei an den Nährstoffbedarf der Tiere,
den Nährstoffgehalt des verfütterten Futters, das Ausmaß und die Art der Parasiten-
belastung sowie an Alter und physische Konstitution der Tiere angepasst werden
(KNOX et al., 2006).
2.7.2.1 Proteine
Der Proteingehalt der Ration spielt dabei eine große Rolle, da dieser in gemäßigten
Zonen naturgemäß den limitierenden Faktor der Diät darstellt und der Befall mit
gastrointestinalen Nematoden den Protein-Stoffwechsel beeinträchtigt (BOWN et al.,
1991, zitiert in HOSTE et al., 2005). So beeinflusste eine ganzjährige Ration, die 125-
130 % des Proteinbedarfs deckt (z.B. durch Supplementierung von Mais oder Soja),
sowohl die Resistenz (gemessen am Grad der Eiausscheidung), als auch Wider-
standsfähigkeit (geschätzt an Milchmenge und Milchfettgehalt) positiv (CHARTffiR et
al, 2000, zitiert in HOSTE et al., 2005). Insbesondere durch die Verbesserung der Pro-
teinversorgung vor und nach der Geburt konnte der in der Folge des PPRI auftreten-

41
den „periparturient rise" , also der vorübergehende Anstieg der Eiausscheidung vor
und nach dem Kitzen, abgeschwächt (HOSTE et al., 2005) bzw. sogar unterdrückt
werden (ETTER et al., 1999, zitiert in HOSTE et al., 2008). Eine Proteinsubstitution
kann demzufolge wesentlich zur EndoparasitenkontroUe und -prävention in Form
von reduzierter Eiausscheidung, besonders in Bezug auf die Trichostrongylose, bei-
tragen. Des Weiteren ermöglichte die Supplementierung von qualitativ hochwertigen
Proteinen die Aufrechterhaltung der Milchleistung bei Hochleistungstieren (ETTER et
al., 2000, zitiert in HOSTE et al., 2002a).
Aus ökologischen Gründen und unter dem ökonomischen Aspekt sind als Futterer-
gänzung bevorzugt regional typische, so z.B. bei der Lebensmittelherstellung anfal-
lende Nebenprodukte oder Abfälle etc., von praktischer Relevanz (KNOX et al.,
2006). Auch bereits die Supplementierung von Mineralstoffe und Non-Protein-
Nitrogen bewirkt bedeutsame Veränderungen der Pansenphysiologie, insofern sie zu
vermehrter Futteraufnahme und vermehrter mikrobieller Proteinproduktion führt und
somit mehr Proteine zur Digestion und Absorption im Dünndarm zur Verfügung ste-
hen. Dies zeigt sich nicht nur in verbesserter Produktion, sondern befähigt die Tiere
zusätzlich, die negativen Auswirkungen der Parasitosen zu relativieren (KNOX, 1996,
zitiert in WALLER, 1999). Allerdings unterliegt die Proteinsubstitution Beschränkun-
gen hinsichtlich möglicher gesundheitlicher Gefahren (Stoffwechselkrankheiten) bei
Überdosierung, ökonomischen Aspekten und Verfügbarkeit möglicher Proteine.
2.7.2.2 Energie
Zusätzlich zur Proteinsubstitution muss jedoch auch das Verhältnis Prote-
in/Energiegehalt der Ration ausgewogen sein, da der Energiegehalt der Ration für die
Ernährung der Pansenmikroorganismen verantwortlich ist, die für die Proteinsynthe-
se zuständig sind. Durch eine ausreichende Energiezufuhr stehen durch die Aktivität
der Mikroorganismen mehr mikrobielle Proteine für die intestinale Verdauung zur
Verfügung und die Pansenfunktion wird verbessert (LENG, 1991, zitiert in KNOX et
al., 2006). Bei hohem Proteingehalt und im Verhältnis zu niedriger Energiezufuhr
. werden die zusätzlichen Proteine nicht verwertet und mit dem Harn ausgeschieden
(TORRES-ACOSTA, 1999, zitiert in HOSTE et al., 2008).
Eine balancierte Ration aus z.B. 26 % Sojabohnen-Mehl und 74 % Hirse-Mehl konn-
te in Versuchen sowohl Resistenz als auch Resilience (Widerstandsfähigkeit) gegen-
über gastrointestinalen Parasiten stärken (ToRRES-AcoSTA, 2004, zitiert in ICNOX et

42
al., 2006). Untersuchungen zeigten ebenfalls, dass die Tiere von der Zufütterung mit
im Pansen verdaulicher Energie profitieren, die kostengünstiger als eine Proteinsub-
stitution ist (KNOX et al., 2006). Nachdem Soja und Mais jedoch zum humanen Le-
bensmittelspektrum gehört, wäre es aus ökologischer Sicht sinnvoll, Futtermittel zu
verwenden, die weniger in Konkurrenz zur Lebensmittelproduktion stehen, wie z.B.
Zuckerrüben-Melasse (sugar-cane-molasses), die vergleichbar gute Ergebnisse bzgl.
der verbesserten Abwehr gegenüber gastrointestinalen Nematoden zeigte und zusätz-
lich durch den Gehalt an Makro- und Spurenelementen eine wertvolle Emährungs-
funktion erfüllt (LANDA-CASIGNO et al., 2005; LENG, 1991, zitiert in KNOX et al.,
2006). Durch Energiesubstitution konnte in Versuchen eine Reduktion der Wurm-
bürde um 65 % erreicht werden (GöMEZ-RINCÖN et al., 2007, zitiert in HOSTE et al.,
2008).
2.7.2.3 Kondensierte Tannine
Als dritte bedeutsame Komponente der Rationsgestaltung mit anthelminthischen
Qualitäten sind pflanzliche Sekundärstoffe, insbesondere kondensierte Tannine (KT),
zu nennen (ATHANASIADOU et al., 2001, zitiert in HOSTE et al., 2005a). Wie bereits
erwähnt, nehmen Ziegen diese Sekundärstoffe unter artgerechten Weidebedingungen
automatisch durch Knabbern an Büschen, Bäumen etc. zu sich, so dass die Parasiten-
belastung in extensiver Haltung häufig geringer ist. In intensiveren Haltungssyste-
men, in denen Ziegen jedoch zum Grasen gezwungen sind, wirken sich nicht nur das
bodennahe Grasen, sondern auch das Fehlen der anthelminthisch wirkenden Sekun-
därstoffe nicht selten nachteilig im Sinne größerer Wurmbürden aus (HOSTE et al.,
2008).
Ein KT-Gehalt von 5-6 % (PAOLINI et al., 2003, 2003a) der Trockenmasse (TM) in
der Ration bzw. 20-40g/kg TM (NGUYEN et al., 2005) resultierte in besserer Toleranz
des Wirtstieres, reduzierter Fertilität der weiblichen Nematoden, reduziertem Schlupf
und schlechterer Entwicklung der Larven (MIN et al., 2004) einer verkleinerten
Wurmpopulation, damit verbundener, bis zu 40 % geringerer Eiausscheidung (MIN et
al., 2004) und außerdem verbesserter Proteinverdaulichkeit und Milchproduktion. Es
muss jedoch berücksichtigt werden, dass ein höherer KT-Gehalt störend auf die Pan-
senfunktionen und somit negativ auf die Proteinverdaulichkeit und die Milchproduk-
tion auswirken kann (BROOKER, 1999, zitiert in WALLER, 2006). Zu den KT-haltigen
Pflanzen zählen unter anderem Leguminosen wie die Ackerbohne (Vicia faba) oder

43
auch weißer Klee {Trifolium repens), Sericea lespedeza (Lespedeza cuneata), Heu
von Süßklee (Esparsette, Onobrychis viviifolia), Lotus-Arien (comiculatus und pe-
dunculatus), Sulla (Hedysarum coronarium), Akazienspezies und Gehölze (Stielei-
che - Quercus robur. Echte Brombeere - Rubus fructicosis. Gemeine Hasel - Cory-
lus avellana), die die Entwicklungsbiologie der Drittlarven wie auch der Adulten
negativ beeinflussen können (Übersichten bei HOSTE et al., 2005a, WAGHORN,
2007). Als besonders wirksam gegen T. circumcinta und T. colubriformis haben sich
der Hornklee - Lotus major, Sulla und auch Quebracho erwiesen, während Quebra-
cho und Süßklee (sowohl Heu als auch Silage) effektiv zur Reduktion von H. contor-
tus beitragen und auch die Fertilität von C. curticei reduzieren können (Übersichten
bei HOSTE et al., 2005a; WAGHORN, 2007).
Die exakte anthelminthische Wirkungsweise von KT ist bis dato noch nicht gänzlich
geklärt. Es bestehen drei Hypothesen, wonach KT zum einen die mit der Nahrung
aufgenommenen Proteine vor ruminalem Abbau schützen, indem sie stabile, unlösli-
che Proteinkomplexe bilden (BARRY U. MANLEY, 1986). In dieser Form gelangen sie
vermehrt über Abomasum in den Dünndarm und erhöhen dadurch die Proteinverfüg-
barkeit, was sich indirekt in einer verbesserten Emährungssituation und daraus resul-
tierend in einer besseren Resilience bzw. verbesserter Immunantwort des Wirtstieres
(BARRY U. MCNABB, 1999, zitiert in MIN et al., 2004) bezüglich negativer Auswir-
kungen von Endoparasiten auswirkt. Zum zweiten wird eine direkt anthelminthische
Wirkung vermutet, wonach sie die Anzahl der gastrointestinal lebenden Parasiten
reduzieren (HOSTE et al., 2005; WALLER, 2006a), und zum dritten üben sie eventuell
auch direkten Einfluss auf die Lebensfähigkeit der frei lebenden Stadien aus (in
Form der Beeinflussung der Entwicklung infektiöser Stadien aus den Eiern) (WAL-
LER, 2006a). Im Zusammenhang mit der Fütterung von KT kann eine bereits nach
spätestens 15 Tagen (MIN et al., 2004) signifikant reduzierte Ei- und auch Coccidien-
Oozystenausscheidung (NGUYEN et al., 2005), eine reduzierte Fertilität der Nemato-
den sowie eine reduzierte Wurmbürde beobachtet werden bzw. tragen sie zu einer
Reduktion der negativen Auswirkungen von Endoparasitosen (WAGHORN, 2007) bei.
Gleichzeitig führen sie zu einer deutlich verringerten Weidekontamination und kön-
nen durch den geringeren Infektionsdruck die Notwendigkeit von Antheminthika-
Behandlungen minimieren (KABASA et al., 2000, zitiert in MIN et al., 2004).
In jedem Fall könnte ihre Wirkung durch Beweidung entsprechend bewachsener
Weiden oder mittels wiederholter Fütterung KT-haltigen Heus genützt werden. Au-

44
ßerdem könnte dieses Vorgehen in Kombination mit anderen Strategien zur Parasi-
tenkontrolle im Sinne eines nachhaltigen Parasitenmanagements gute Ergebnisse
erzielen und somit das Weidemanagement revolutionieren (WALLER, 1999). Des
Weiteren wirkt sich eine KT-Fütterung ökologisch positiv in Form einer geringeren
Stickstoffausscheidung und geringerer Methan-Emission aus (WAGHORN U. WOOD-
WARD, 2006, zitiert in WAGHORN et al., 2007), was in Zeiten der Diskussionen um
Treibhausgase und Klimawandel wohl immer mehr an Bedeutung gewinnen wird.
2.7.2.4 Weitere Sekundärstoffe
Abgesehen von den kondensierten Tanninen wurde Chicoree, aufgrund seiner In-
haltsstoffe wie z.B. den Sesquiterpen-Laktonen in vitro nachweislich antiparasitische
Qualitäten zugesprochen (MOLAN et al., 2000, zitiert in WALLER U. THAMSBORG,
2004). Insgesamt ist unklar, ob die positiven Auswirkungen einer Supplementierung
im Sinne einer reduzierten Eiausscheidung auf eine verbesserte Immunabwehr oder
eine veränderte Zusammensetzung der Wurmpopulation zurückzuführen ist. Fest
steht jedoch, dass in Studien bei substituierter Fütterung ein Anstieg der eosinophilen
Granulozyten in Kombination mit verminderter Eiausscheidung zu beobachten war
(TORRES-ACOSTA et al., 2004, zitiert in HOSTE et al., 2008).
2.7.3 Prävention durch Vakzinierung
Trotz jüngster Studien ist in absehbarer Zeit nicht mit einer in der Praxis bereits ein-
setzbaren neuen anthelminthisch wirksamen Stoffgruppe zu rechnen, deshalb gilt der
Suche nach Alternativen zur medikamentellen Parasitenbekämpfung besonderes Au-
genmerk (HERTZBERG U. BAUER, 2002; WALLER, 2006). Neuere Versuche, Vakzinen
auf molekularer Ebene mittels parasitären Antigenmaterials herzustellen, scheiterten
nicht zuletzt an der Schwierigkeit, Antigenfraktionen des Parasitenmaterials in
kommerziellen Mengen zu produzieren. Zusätzlich verkompliziert die komplexe
Immunantwort, bestehend aus einer Kombination von lokaler Hypersensitivität mit
zell- und antikörpervermittelter sowie entzündlicher Komponente, die Entwicklung
derartiger Vakzinen (SMITH, 1999, zitiert in WALLER U. THAMSBORG, 2004). In Er-
mangelung neuer Impfstoffe werden nun althergebrachte Methoden in Form von
attenuierten, fortpflanzungsunfähigen Larven, die die Immunantwort des Wirtstieres
provozieren sollen, tendenziell wieder mehr als Prophylaxe in Betracht gezogen
(WALLER, 1999). In diesem Zusammenhang wurden bereits vor Jahren Vakzinen v.a.

45
gegen H. contortus entwickelt, die viel versprechende Ergebnisse lieferten, jedoch
bis dato aufgrund von Problemen hinsichtlich der Produktion noch nicht kommerziell
zum Einsatz gekommen sind (WALLER, 1999).
Ein zusätzliches Problem bezüglich der Wirksamkeit der Impfungen stellt zudem die
natürliche immunologische Reaktionsschwäche von Jungtieren (< 6-9 Monaten) und
trächtigen Ziegen um den Geburtstermin dar (SMITH, 1999, zitiert in WALLER U.
THAMSBORG, 2004). Langfristig bieten vor allem züchterische Maßnahmen die
Chance auf einen reduzierten Anthelminthika-Einsatz durch Selektion nematodenre-
sistenter Tiere, wobei als Selektionskriterium eine niedrige Eiausscheidungsrate hin-
zugezogen werden kann, die ein guter Indikator einer geringen Wurmbürde ist.
2.7.4 Prävention durch Zucht und Rasseauswahl
2.7.4.1 Rasse
Eine erfolgreiche Prävention muss bereits bei der Rasseauswahl beginnen. So sind
rassebedingte Unterschiede in der Empfänglichkeit gegenüber Endoparasiten bekannt
(AGUILAR-CABALLERO, 2003), die besonders in Phasen hoher Beanspruchung und
Leistung (Trächtigkeit, Laktation) in Erscheinung treten (HOSTE et al., 2005, 2008).
HOSTE et al. (2001) wiesen nach, dass das bereits diskutierte Graseverhalten auch
rassetypisch differieren kann. Das ist insofern bedeutsam, als das Fressverhalten -
wie im Kapitel 5.8 eingehender behandelt wird - Teil der Strategie der Ziegen zur
Parasitenkontrolle ist, indem sie eine Infektion durch ihre selektive Futteraufnahme
und eine höhere Widerstandskraft zu vermeiden versuchen, da die Immunantwort
bekanntlich bei Ziegen im Vergleich zu Schafen weit weniger ausgeprägt und unzu-
reichend ist (HuNTLEY, 1995, zitiert in HOSTE et al., 2005; HOSTE U. CHARTffiR,
1998a).
Grundsätzlich ist die Rasseauswahl an den Standort anzupassen. So waren Versuche,
besonders widerstandsfähige Rassen aus tropischen Regionen in gemäßigten Zonen
anzusiedeln bzw. einzukreuzen, nicht erfolgreich, da die Tiere große Schwierigkeiten
hatten, sich an Klimafaktoren, Fütterung und vor allem an das unbekannte Parasiten-
spektrum zu adaptieren (WALLER, 1999). Zusätzlich liefern die tropischen Rassen bei
weitem nicht jene Produktionsleistungen wie Rassen der gemäßigten Klimazonen.
Jüngste Bestrebungen zielen daher auf eine Decodierung der für die Unempfänglich-
keit gegenüber Parasiten verantwortlichen Gene aus widerstandsfähigen, tropischen

46
Rassen, die Bestimmung ihres Genlocus und deren Integration in das Genom öko-
nomisch-produktiver Rassen, wodurch transgene Tiere entstehen. Diese Entwicklung
vollzieht sich jedoch nur sehr langsam und muss gleichzeitig auch sehr kritisch hin-
terfragt werden, da Langzeitfolgen nur schwer bzw. nicht absehbar sind (WALLER U.
THAMSBORG, 2004).
2.7.4.2 Zucht
Eine langfristige Lösung des Parasitenproblems kann besonders auf züchterischer
Ebene erfolgen. Endoparasitenprävention bedeutet dabei, jene Tiere für die Zucht zu
selektieren, die gegenüber negativen Auswirkungen von Parasitosen unempfindlicher
(toleranter) oder sogar im Stande sind, Parasitosen mit Hilfe immunologischer Ab-
wehrmechanismen zu bekämpfen (BARGER U. DASH, 1987, zitiert in KOOPMANN et
al., 2006). Die Eiausscheidungsrate im Kot der Tiere kann als verlässlicher Parame-
ter zur Ermittlung angeborener Widerstandsfähigkeit gegen Parasiteninfektionen
angesehen werden und ist aufgrund der Heritabilität als Selektionsmerkmal geeignet
(EADY et al., 1996, zitiert in WALLER, 1999; HOSTE et al., 2008).
Wie im Kapitel 5.8.1 eingehender beschrieben, konnten HOSTE et al. (2001a) anhand
der Eiausscheidungsrate nachweisen, dass innerhalb einer Herde immer dieselben
Tiere besonders hohe Befallsintensitäten zeigen und demnach immer nur ein kleiner
Anteil der Tiere für den Großteil der ausgeschiedenen Parasitenstadien verantwort-
lich ist. Diese Erkenntnis ist für die genetische Selektion parasitenresistenter Tiere
im Hinblick auf züchterische Parasitenkontrolle von großer Bedeutung und sollte
künftig stärker genutzt werden.

47
3. MATERIAL UND METHODE
3.1 Klimadaten der untersuchten Region
Die im Folgenden verwendeten Klimadaten sind mit Genehmigung der Zentralanstalt
für Meteorologie und Geodynamik (ZAMG) in Wien der Website www.zamg.ac.at
entnommen.
3.1.1 Niederschlagsmenge
In der untersuchten Region liegt die Niederschlagsmenge im Durchschnitt (Mittel-
wert der Jahre 1971-2000) bei 1333,3 Liter pro m^ pro Jahr, demzufolge im Ver-
gleich zu z.B. Wien mit 547,9 Liter pro m^ pro Jahr mehr als doppelt so hoch. Nie-
derschlagshöhepunkte waren dabei in den Monaten Juni/Juli/August zu verzeichnen.
3.1.2 Temperatur
Die Jahresdurchschnittstemperatur von 1971-2000 liegt in der untersuchten Region
mit 8,5°C deutlich unter der von Wien mit 11,4°C. Höchsttemperaturen zeigt der Juli
mit durchschnittlich 18,1°C gegenüber 21,7°C in Wien. Temperaturtiefpunkte sind
im Januar mit durchschnittlich -1,1°C und 23,3 Frosttagen gegenüber Wien mit
-f-l,2°C und 17,1 Frosttagen zu beobachten.
3.2 Betriebe
Zur Untersuchung wurden drei, im Grenzgebiet Salzburg/Oberösterreich im Umkreis
von 21 km und auf durchschnittlich 489 m (+ 23 m) Seehöhe liegende, biologisch
wirtschaftende Milchziegenbetriebe ähnlicher Rassenstruktur mit unterschiedlichen
Haltungs- bzw. Fütterungsformen ausgesucht und jeweils im Sommer (8./9. August)
und Herbst (31. Oktober) 2007 besucht.
3.3 Befragung der Landwirte zu den Betriebsabläufen
Beim ersten Betriebsbesuch erfolgte eine Befragung der Landwirte zu allgemeinen
betrieblichen Abläufen und bereits bekannter Parasitenproblematik anhand eines
Fragebogens (siehe Kapitel 10).

48
3.4 Tierauswahl
Die Auswahl der Tiere zur rektalen Kotprobenentnahme erfolgte zufällig und um-
fasste mindestens fünf Tiere pro Haltungsgruppe. Zusätzlich wurden klinisch auffäl-
lige Tiere (Husten, Niesen, Veränderungen im Atemmuster, Abmagerung, Kotver-
schmutzungen bzw. beobachteter Durchfall, vermindertes Allgemeinverhalten,
stumpfes, struppiges Haarkleid) in die Untersuchungen mit aufgenommen.
3.5 Klinische Untersuchung
Die Tiere wurden klinisch untersucht und Auffälligkeiten wie Zahnprobleme, Nasen-
ausfluss, Husten, stumpfes, struppiges Haarkleid, Durchfall bzw. verschmutzte Anal-
region vermerkt und der Ernährungszustand anhand allgemeiner Adspektion und
zusätzlicher Palpation von Sternum und Lumbaibereich ermittelt und als Body Con-
dition Score angegeben.
3.6 Erhebung des Body Condition Score (BCS)
Die Erhebung des BCS dient zur Ermittlung der Körperkondition bzw. der Energie-
reserven eines Tieres. Diese Technik wurde zuerst für Schafe entwickelt und später
zur Anwendung bei Ziegen adaptiert (Lenden- und Brustbeinscore für Milchziegen
nach MORAND-FEHR et al., 1989), nachdem die Ziege, im Unterschied zum Schaf,
das Körperfett in lokalen Fettdepots - dazu gehören die Brustbein- sowie die Len-
denwirbelregion - speichert. Berücksichtigt werden muss außerdem, dass ein Teil
des Körperfetts bei der Ziege intraabdominal und somit nicht palpierbar liegt (VATTA
et al., 2002), so dass der palpierbare Fettanteil das Tier magerer erscheinen lässt, als
es tatsächlich ist.
Der allgemeine Ernährungszustand wurde zunächst durch Adspektion des Tieres in
Distanzbetrachtung ermittelt und in den Score von 1-5 eingeteilt (1: kachektisch, 1,5:
schlecht, 2: mindergut, 2,5-3,5: gut, 4: sehr gut, >4: adipös). Aufgrund der bereits
erwähnten lokalen Fett- und auch Proteinspeicherung der Ziegen wurde zusätzlich
die Sternal- und Lumbairegion palpiert, um die vorhandenen Fettdepots wiederum
nach folgendem Schema (LEEB, 2002; HARWOOD U. MATTHEWS, 2008) zu beurteilen:

49
Score 1 Scaxe. Z
\^'0
Ä«t 2
S:oie 5
Abb. 1 Sternaler Score 1-5, Brustkorbquer- schnitt (l=kachektisch; 1,5=schlecht; 2=tnindergut; 2,5-3,5=gut; 4=sehr gut; >4=adipös)
Abb. 2 Lumbaler Score 1-5, Rückenquerschnitt (l=kachektisch; l,5=schlecht; 2=mindergut; 2,5- 3,5=gut; 4=sehr gut; >4=adipös)
3.7 Probenentnahme
Die Kotproben wurden in der Regel rektal bzw. von während der klinischen Untersu-
chung frisch abgesetztem und einem Tier eindeutig zuzuordnendem Kot entnommen.
Etwaige, von Durchfall herrührende Verschmutzungen der Analregion wurden ver-
merkt. Der Kot jedes Einzeltieres wurde grobsinnlich nach Farbe, Konsistenz, Bei-
mengungen und Geruch beurteilt und Abweichungen von der Norm vermerkt.
Zusätzlich zu den Einzelproben wurden Sammelkotproben der einzelnen Haltungs-
gruppen aus den Laufboxen von jeweils mindestens vier verschiedenen Stellen ent-
nommen (die jeweils einzeln verpackt wurden), da sich die Gewinnung von genü-
gend Material an den Einzeltieren häufig schwierig darstellte und die gewonnene
Kotmenge der Einzelproben größtenteils nicht ausreichte, um zusätzlich zur Anrei-
cherung Eizählungen durchzuführen bzw. Kotkulturen anzusetzen.
3.8 Probenaufarbeitung
Jede Kotprobe wurde möglichst luftarm einzeln in Plastiksäckchen verpackt. Lage-
rung und Transport erfolgten unter ständiger Kühlung in einer transportablen Kühl-
box. Die Aufarbeitung und parasitologische Untersuchung der Kotproben erfolgte
am Institut für Parasitologie und Zoologie der Veterinärmedizinischen Universität
Wien. Die Einzelproben wurden mittels Sedimentations-Flotations-Verfahren ange-
reichert und einzeln ausgewertet. Anhand der Sammelkotproben wurden Eizählungen
durchgeführt und Kotkulturen angesetzt.

50
3.8.1 Kombiniertes Sedimentations-Flotations-Verfahren
Um einen Überblick über das vorhandene Parasitenspektrum zu erhalten, wurden von
allen Einzeltierkotproben Anreicherungen durchgeführt. Die Anreicherung erfolgte
mittels kombiniertem Sedimentations-Flotations-Verfahren mit Sedimentation unter
Zugabe von Leitungswasser und darauf folgender Flotation anhand der Durchmi-
schung des Sediments mit gesättigter Zuckerlösung. Von jeder Einzelprobe wurden 3
g Kot entnommen. Sofern weniger Material vorhanden war, wurde dies notiert. Die
Proben wurden in Reibschalen mit Leitungswasser versetzt und der Kot zur gleich-
mäßigen Durchmischung mittels Mörser zerrieben. Der in Wasser gelöste Kot wurde
daraufhin durch grobmaschige Metallsiebe in Reagenzgläser abgefüllt. Die Reagenz-
gläser wurden bei 2.500 Umdrehungen pro Minute (U/min) 4 Minuten (min) lang
zentrifugiert. Mittels einer Wasserstrahlpumpe wurde daraufhin der Überstand abge-
saugt und das Sediment mit gesättigter Zuckerlösung gut durchmischt. Darauf folgte
ein erneutes Zentrifugieren bei 2.500 U/min für 4 min. Von der Oberfläche des Über-
standes wurden mit davor abgeflammten und gut ausgekühlten Drahtösen mehrere
Tropfen entnommen, die sodann auf Objektträger aufgebracht wurden. Die Präparate
wurden mit Deckgläsern abgedeckt, im Durchlichtmikroskop bei zuerst schwacher
(lOOx), dann stärkerer Vergrößerung (200 bis 400x) ausgewertet und die differen-
zierten Eitypen anhand ihrer relativen Häufigkeit (vereinzelt, geringgradig (ggr),
mittelgradig (mgr), hochgradig (hgr) und Übergangsformen z.B.: ggr-mgr) für jedes
Einzeltier aufgezeichnet.
3.8.2 Modiflziertes McMaster-Verfahren
Dieses quantitative Verfahren dient dazu, anhand der Verwendung definierter Kot-
mengen und Flotationsflüssigkeit Nematodeneier und Coccidienoozysten in einer
Zählkammer („McMaster-Kammer") durch Flotation anzureichern. Die Parasitensta-
dien werden ausgezählt und als Ei- bzw. Oozystenanzahl pro Gramm Kot angegeben
(Ei/Oozysten pro Gramm, kurz epg/opg). Die Eizählungen wurden an den Sammel-
kotproben der einzelnen Haltungsgruppen vorgenommen. Von jeder der mindestens
vier Proben pro Haltungsgruppe wurde je 1 g Kot entnommen und in ein Plastik-
schälchen verbracht. Die Proben wurden mit 45 ml Zinksulfat pro 2 g Kot versetzt
und gut durchmischt. Mit Einwegpipetten wurden pro Probe alle drei Zählfelder
(wovon jedes ein Volumen von 150 |il umfasst) einer McMaster-Zählkammer be-
schickt und bei 100-facher Vergrößerung im Durchlichtmikroskop ausgezählt. Die

51
Anzahl der pro Parasitenstadium gefundenen Objekte wurde mit 50 multipliziert und
in epg angegeben.
3.8.3 Kotkultur (nach Roberts und O'Sullivan) und Auswanderverfahren (nach Baermann-Wetzel)
Nachdem die Gattungszugehörigkeit von MDS-Eiem mittels der gängigen Verfahren
der Koproskopie mit Ausnahme Nematodirus nicht identifiziert werden kann, wur-
den Kotkulturen angesetzt, um anhand der sich im Kot entwickelnden Drittlarven
eine Gattungszuordnung vornehmen zu können. Zur Identifikation wurden nur ein-
deutig differenzierbare Drittlarven hinzugezogen. Für die Kotkultur wurden von den
in den Laufboxen der Haltungsgruppen aufgesammelten Sammelkotproben je 10g
Kot entnommen, in ein Einmachglas gegeben und mit sterilen Sägespänen und Lei-
tungswasser vermengt, so dass die Kultur zwar feucht, jedoch durch die Sägespäne
aufgelockert vorlag, um den Zutritt von Sauerstoff zu gewährleisten. Die Einmach-
gläser wurden im Wärmeschrank bei 25°C etwa vier Wochen bebrütet. Eine regel-
mäßige Belüftung durch Schütteln, Kontrolle der Feuchtigkeit und - falls nötig -
erneute Zugabe von Wasser wurde durchgeführt. Eine zur Erhaltung konstanter Luft-
feuchtigkeit im Brutschrank positionierte Wasserschale verhinderte zusätzlich die
Austrocknung der Kotkulturen.
Nach der Bebrütung wurden die in Gaze eingeschlagenen Kotkulturen nach dem
Trichterauswanderverfahren nach Baermann-Wetzel in einen Trichter verbracht, mit
Wasser versetzt und über Nacht ruhen gelassen, um das Auswandern der hydrophilen
Larven aus den Kotkulturen zu bewirken. Der Abfluss des sich sammelnden Wassers
wird hierbei durch eine Klemme unterbunden. Am Folgetag wurden die Klemmen
gelöst und der flüssige Anteil in Reagenzgläser abgelassen. Die Probenröhrchen
wurden bis zur Auswertung im Kühlschrank gelagert. Zur Auswertung wurden mit
Einwegpipetten einige Tropfen vom Sediment jeden Probenröhrchens entnommen,
auf einen Objektträger aufgebracht und die noch lebenden und sich bewegenden Lar-
ven mit einem Tropfen Lugol'scher Lösung abgetötet. Zur Auswertung im Durch-
lichtmikroskop bei 40-facher Vergrößerung wurden alle bzw. höchstens 100 Larven
nach dem Schema von ECKERT et al. (2005b) differenziert und ausgezählt, um daraus
den prozentuellen Anteil der jeweiligen Gattung zu errechnen.

52
3.9 Statistische Auswertung
Die statistische Auswertung erfolgte mittels SPSS Version 14.0 für Windows. Nach-
dem die ermittelten Daten nicht normalverteilt waren, wurden Korrelationen mit dem
Korrelationskoeffizienten nach Spearman, Vergleiche zwischen mehreren Gruppen
mittels Kruskal-Wallis-Test und Vergleiche zwischen zwei Gruppen anhand des
Mann-Whitney-U-Tests berechnet und auf ihre Signifikanz geprüft.
4. ERGEBNISSE
4.1 Betriebscharakteristik und Befragungsergebnisse
Exakte Daten zur Fütterung und Rationsgestaltung sowie zur Futtermittelanalyse sind
der Diplomarbeit von Robert Wolf (in Veröffentlichung) zu entnehmen.
4.1.1 Betrieb 1 mit Auslauf und Heufütterung
Der Betrieb arbeitet seit 30 Jahren biologisch und seit 20 Jahren mit Milchziegen.
Zum Zeitpunkt der Befragung am 08. August 2007 umfasste der Betrieb 160 adulte
Milchziegen (v.a. Deutsche weiße Edelziege, teilweise Saanenziege bzw. Anglo-
Nubier eingekreuzt), 100 Jungtiere und 14 Mastböcke.
• Aufstauung
Die adulten Tiere werden entsprechend ihres Laktationsstadiums gruppiert (Hoch-,
Mittel-, Niedriglaktation, Trockenstehende) und sind in einem Offenfrontstall mit
Tiefstroh-Laufboxen und jeweils angeschlossenem und jederzeit von den Tieren
nutzbarem befestigtem Auslauf aufgestallt. Die Einstreu bleibt in den Boxen, bis sie
ca. 40 cm hoch ist. Dreimal jährlich wird gesamt entmistet und neu eingestreut, eine
Desinfektion erfolgt nicht. Der Mist wird zur Düngung der eigenen Futterwiesen
verwendet. Bis vor zwei Jahren wurde noch mit Weidegang gearbeitet, dann wurde
jedoch aufgrund wiederholter Durchfallproblematik auf befestigte Ausläufe umge-
stellt. Nach Angaben des Betriebsleiters ist das Auftreten von Durchfällen erheblich
gesunken, seitdem die Tiere keinen Zugang mehr zu Weide und zu Grünfutter haben.

53
• Kitzaufzucht
Die Kitze werden nach 3-5 Tagen von den Muttertieren getrennt und danach mit Bio-
Kuhmilch-Pulver im Tränkautomat aufgezogen.
• Fütterung
Es erfolgt eine ganzjährige Heufütterung mit an die Milchleistung angepasster Zufüt-
terung von Kraftfutter. Die Hochlaktierenden (Gruppe 2) werden mit 30% Erst-
schnitt-Heu und 70% Drittschnitt-Heu ad libitum mit einer Zufütterung von 1 kg
Fertigkraftfutter und 0,25 kg Körnermais pro Tier und Tag gefüttert. Die Mittellak-
tierenden (Gruppe 3 und 4) erhalten gleiche Menge und Zusammensetzung an Heu
unter Zugabe von 0,8 kg Fertigkraftfutter und 0,15 kg Körnermais pro Tier und Tag.
An die Niedriglaktierenden werden Grundfutterreste der Gruppen 2-4 ad libitum und
jeweils 0,4 kg Fertigkraftfutter pro Tier und Tag verfüttert. Die Trockenstehenden
werden mit Heu ad libitum wie bereits oben erwähnt und einer Beifütterung von 0,5
kg Fertigkraftfutter pro Tier und Tag gefüttert.
Der Ernährungszustand wird vom Besitzer regelmäßig anhand Palpation des Brust-
beins überprüft. Tiere, die einen sehr guten Ernährungszustand aufweisen, werden in
eine Haltungsgruppe mit niedrigenergetischer Fütterung umgruppiert. Dementspre-
chend werden Tiere minderguten Ernährungszustandes in eine Haltungsgruppe mit
höherem Kraftfutteranteil umgestallt. Das unter Belüftung getrocknete Heu stammt
ausschließlich von betriebseigenen, mit eigenem Stallmist gedüngten Futterwiesen.
Der Mist wird 2-3x umgesetzt und unter Luftzufuhr verrottet. Sobald er verrottet ist,
wird er im Herbst und Frühling auf die Wiesen ausgebracht. Die Wasserversorgung
wird mittels Frischwassertränken gewährleistet.
• Zukaufe
Es werden ausschließlich Böcke zugekauft. Neu in den Bestand einzubringende Tiere
werden tierärztlich untersucht (auf Brucellose und Pseudotuberkulose getestet) und
bei Gesundheitsbestätigung durch den Grenz- oder Amtstierarzt ohne Quarantäne in
die Herde integriert. Es werden keine Ausstellungen besucht und auch sonst besteht
keine Kontaktmöglichkeit mit Tieren anderer Bestände. Kitze werden zum Teil le-
bend verkauft, zum Teil am Hof geschlachtet.

54
• Anamnese betriebsbekannter Parasitosen bzw. klinischer Auffälligkeiten
Solange die Tiere noch Weidegang hatten, bestand laut Besitzer eine regelmäßige
Durchfalls- und Verwurmungsproblematik. Seitdem die Tiere jedoch ausschließlich
in befestigten Ausläufen gehalten werden und kein Grünfutter mehr verfüttert wird -
also seit zwei Jahren - ist für den Besitzer keine generelle Endoparasitenproblematik
augenscheinlich. Im Allgemeinen werden keine Entwurmungen durchgeführt, einzig
bei Auftreten von vermindertem Allgemeinbefinden, Atemwegsproblemen, Husten
oder Durchfall und bei zugekauften Zuchtböcken aus Weidehaltung werden die be-
troffenen Tiere entwurmt. Vor allem im Sommer treten Atemwegsprobleme (Husten)
regelmäßig und gehäuft bei allen Altersgruppen auf. 2005 wurden Coccidien und ein
Befall mit kleinen Lungenwürmem unterschiedlicher Befallsintensität diagnostiziert.
Unter den Kitzen und Jungziegen treten zeitweise Probleme mit Coccidien auf, die
sich nach Angaben des Landwirtes in wechselhafter Kotkonsistenz äußern. Ein Ekto-
parasitenbefall ist nicht bekannt. Bei einmalig auftretendem vermehrtem Juckreiz
mehrerer Tiere fiel eine parasitologische Untersuchung durch den Tierarzt negativ
aus. Die Sterberate bei der Kitzaufzucht schätzt der Besitzer auf 4-5 %.
• Parasitenmanagement
Im Betrieb werden keine prophylaktischen Maßnahmen zur Parasitenbekämpfung
durchgeführt. Eine Behandlung erfolgt nur bei zugekauften Zuchtböcken aus Weide-
haltung und eventuell auffälligen Einzeltieren mit Cydectin® oral 0,1 % (Wirkstoff
Moxidectin, Fort Dodge Veterinär GmbH Deutschland, Würselen). Dabei wird die
1,5-fache für Schafe empfohlene Dosis verwendet (0,3 mg Moxidectin pro kg Kör-
pergewicht (KGW) = 1,5 ml Cydectin® 0,1 % oral/pro 5 kg KGW oral). Regelmäßige
Bestandsbehandlungen werden nicht durchgeführt. Zeitweises Auftreten von wech-
selhafter Kotkonsistenz bei Kitzen und Jungziegen führt der Landwirt auf Coccidien
zurück, die er mit homöopathischen Tropfen {Abrotanum D6, 20 Tropfen in 10 L
Trinkwasser) behandelt.
4.1.2 Betrieb 2 mit Auslauf und Grünfütterung
1998/1999 wurde der Betrieb auf biologische Bewirtschaftung umgestellt. Zum Zeit-
punkt des Betriebsbesuchs umfasste der Betrieb 180 laktierende Adulte und 70-80
Jungziegen der Rassen bunte und weiße deutsche Edelziege und braune Gebirgszie-
ge.

55
• Aufstauung
Die Ziegen sind zum Zeitpunkt des ersten Betriebsbesuchs nach Alter der Tiere in
drei adulte Gruppen (jung, mittleres Alter, alt) und eine Gruppe Jungtiere gruppiert.
Beim Herbstbesuch war jedoch eine Umgruppierung nach Laktationsstadium, ver-
gleichbar mit Betrieb 1, erfolgt. Die Tiere sind in einem umgebauten Kuhstall mit
Tiefstrohlaufboxen und nur zum Teil jederzeit nutzbaren Ausläufen aufgestallt. Im
Sommer wird die Einstreu monatlich zur Gänze gewechselt. Im Jahr 2007 wurde
erstmals auch desinfiziert (Desinfektionsmittel ist jedoch unbekannt), da bei den
Jungtieren wiederholt Probleme mit Eimerien auftraten. Entmistet und nachgestreut
wird im Sommer mindestens jeden zweiten Tag, im Winter etwas seltener, da die
Tiere das gefütterte Heu in die Boxen einbringen.
• Kitzaufzucht
Nach aktuellsten telefonischen Informationen vom Juni 2008 werden die Kitze neu-
erdings ab dem zweiten oder dritten Lebenstag in eine neu errichtete Halle umgestallt
und dort gesondert mittels Tränkautomat aufgezogen. Die Halle konnte vor Abgabe
dieser Arbeit leider nicht mehr besichtigt werden.
• Fütterung
Solange die Gruppen noch nach Alter gruppiert waren, wurde an alle Haltungsgrup-
pen dieselbe Ration aus Erstschnitt-Heu und Viertschnitt-Grünfutter und 0,5 kg Fer-
tigkraftfutter verfüttert. Mit Umgruppierung der Ziegen erfolgt die Rationsgestaltung
auch hier produktionsorientiert, so dass Tiere in Hochlaktation besseres Heu und
mehr Kraftfutter (0,6 kg/Tier) bekommen als die Niederiglaktierenden, die weniger
als 0,5 kg Kraftfutter erhalten. Das Grünfutter stammt ausschließlich von betriebsei-
genen Futterwiesen, die mit eigenem Stallmist und Gülle gedüngt werden. Der
Stallmist wird für mindestens drei Monate im Freien gelagert und wird nach erfolgter
Verrottung auf die betriebseigenen Wiesen ausgebracht. Die Wasserversorgung er-
folgt mittels Frischwassertränken im Stallbereich.
• Zukaufe
Es werden ausschließlich Zuchtböcke zugekauft und keinerlei Ausstellungen be-
sucht, so dass keine Kontaktmöglichkeiten zu Tieren aus anderen Beständen beste-
hen. Die neu zugekauften Böcke stammen aus der Schweiz und werden bereits im

56
Zuge des Imports einer Quarantäne unterzogen. Im Betrieb werden die Tiere deshalb
nicht in Quarantäne gehalten.
• Anamnese betriebsbekannter Parasitosen bzw. klinischer Auffälligkeiten
Bekannt ist die bei Jungtieren regelmäßig auftretende Kokzidiose. Der Besitzer hat
den Eindruck, dass das Auftreten der Kokzidiose fütterungsabhängig ist und gehäuft
bei Grünfutterfütterung auftritt. Im Winter bei Heufütterung ist die Problematik we-
niger ausgeprägt. Bei den Adulten konnten bereits des Öfteren Würmer beim Melken
gefunden werden, die jedoch trotz Einsenden an eine Untersuchungsanstalt nicht
näher bestimmt werden konnten. Ansonsten sind dem Besitzer bei den adulten Tieren
keine parasitär-bedingten Probleme bekannt. Die Sterberate der Kitze gibt der Besit-
zer mit 0 % an, solange die Kitze noch bei den Muttertieren sind. Nach dem Abset-
zen wird die Sterblichkeit auf 1-2 % geschätzt.
• Parasitenmanagement
Die Jungtiere werden aufgrund der regelmäßig auftretenden Kokzidiose systematisch
im Alter von 4-5 Wochen mit Baycox® 2,5 % orale Lösung für Hühner (Wirkstoff
Toltrazuril, Bayer, Leverkusen) oral behandelt (über die genaue Dosierung konnte
der Landwirt jedoch keine Auskunft geben). Der Besitzer ist 2007 erstmalig dazu
übergegangen, nach Entmistung zu desinfizieren. Etwaige Verbesserung der Situati-
on konnte zum Zeitpunkt des Besuches noch nicht bemerkt werden, allerdings wurde
erst einmal desinfiziert. In der neuen Halle zur Kitzaufzucht wird derzeit nicht desin-
fiziert.
Entwurmungen werden bei Jungziegen vor der Geburt und bei den Altziegen regel-
mäßig in der Trockenstehperiode, im Abstand von ca. 1,5-2 Jahren, durchgeführt.
Der Besitzer sieht diesbezüglich als Problem, dass die Abkitztermine über ein halbes
Jahr streuen und somit immer nur einzelne Tiere, nicht jedoch der gesamte Bestand,
entwurmt werden. Zur Entwurmung wird meistens Panacur® 2,5 % orale Suspension
(Wirkstoff Fenbendazol, Intervet, Wien; 7,5 mg Fenbendazol pro kg KGW = 1,5 ml
Panacur® 2,5% orale Suspension pro 5 kg KGW oral), kürzlich Cydectin®0,l % oral
(Wirkstoff Moxidectin, Fort Dodge Veterinär GmbH Deutschland, Würselen; Dosie-
rung siehe oben) - beides in 1,5-facher Schafdosierung - eingesetzt, wobei nach ge-
schätztem Körpergewicht dosiert wird.

57
4.1.3 Betrieb 3 mit ganzjähriger Weidehaltung
Der Betrieb arbeitet seit 1993/1994 biologisch. Zum Zeitpunkt des Betriebsbesuches
im August 2007 umfasste der Betrieb 170-180 adulte Milchziegen, 30 Jungziegen
und 50-60 Kitze der Rassen Deutsche Edelziege (weiß und bunt), Anglo-Nubier,
Burenziege, Bündner Strahlenziege bzw. Kreuzungen zwischen den Rassen. Die Kit-
ze werden zum Teil zum Eigenbedarf geschlachtet oder zur Weiterzucht genützt.
• Aufstauung
Alle laktierenden Ziegen werden zusammen in einer Großgruppe gehalten, lediglich
die Trockenstehenden werden von der Gruppe separiert und mit den Jährlingen auf-
gestallt. Jährlinge und Kitze sind in jeweils eigenen Gebäuden untergebracht, die
getrennt voneinander begehbar sind. Alle Tiere werden in Tiefstrohsystemen gehal-
ten. Jährlinge und Laktierende haben ganzjährigen Zugang zu Weideflächen, Kitze
permanente Nutzungsmöglichkeit von Ausläufen. Dreimal jährlich werden die Stal-
lungen zur Gänze entmistet und mit Urgesteinsmehl zur Desinfektion bestreut. Die
Boxen werden nach Bedarf nachgestreut. Der Stallmist wird l-2x pro Jahr umge-
schichtet und für gewöhnlich ein Jahr gelagert, bevor er auf die Weiden ausgebracht
wird. Die Tiere haben ganzjährigen Weidegang (auch bei Schnee). Auf einer der
Weiden befindet sich eine permanent-feuchte Stelle, an der eine besondere Vegetati-
on wächst, die die Tiere sehr gerne fressen. Einmal im Jahr dürfen sie an dieser Stelle
grasen. Trotzdem wurden beim Schlachten bisher erst sehr selten und nur sehr ver-
einzelt Leberegel gefunden.
• Kitzaufzucht
Die Kitze bleiben ca. vier Tage bei der Mutter. Wenn mehrere Ziegen in kurzem Ab-
stand nacheinander gebären, wird mitunter bis zu zehn Tage mit der Trennung ge-
wartet, um für mehrere Kitze gleichzeitig mit der - anfangs gewässerten - Bio-
Kuhmilch-Fütterung aus einem am Boden stehenden Becken zu beginnen. Heu und
Kraftfutter stehen zur freien Verfügung, weibliche und männliche Jungtiere werden
getrennt gehalten.
• Fütterung
Im Sommer bildet die Weide, d.h. dementsprechend Grünfutter zusammen mit 1,1 kg
Kraftfutter die tägliche Futterration pro Tier. Im Winter wird statt Grünfutter Heu

58
zusammen mit 1,1 kg Kraftfutter verfüttert. Die Frischwasserversorgung wird ganz-
jährig über eine über den Melkstand geführte Rundleitung gewährleistet.
• Zukaufe
Es werden ausschließlich Böcke aus kontrollierten Zuchten zugekauft. Die neu zuge-
kauften Böcke werden sofort in die bestehende Herde und das für sie vorgesehene
Entwurmungsschema integriert.
• Anamnese betriebsbekannter Parasitosen bzw. klinischer Auffälligkeiten
Bei den Kitzen tritt regelmäßig Durchfall auf, jedoch scheint dieser laut Beobachtun-
gen des Besitzers häufig an zu kalt verfütterte Kuhmilch, die zur Aufzucht verwendet
wird, gekoppelt zu sein. Zur Zeit des Besuchs im August befinden sich laut Besitzer
zwei innerhalb kurzer Zeit (wenige Tage) hochgradig abgemagerte adulte Tiere im
Bestand, die Durchfall zeigen, seit einigen Tagen keine Milch mehr geben und vor-
rangig in die parasitologischen Untersuchungen inkludiert werden sollen. Bei allen
Altersgruppen wird regelmäßig und vermehrt in den Sommermonaten Husten beo-
bachtet. Eine Lungenwurmproblematik ist den Besitzern bekannt. Leberegel konnten
bei den Schlachtungen nur vereinzelt nachgewiesen werden. Eine Coccidien-
Problematik ist nicht bekannt. Die Sterblichkeit der Kitze wird mit 40 % (60 von 150
Kitzen pro Jahr) angegeben und tritt vornehmlich in den ersten vier Tagen auf
• Parasitenmanagement
Der Weidegang erfolgt nach dem Prinzip des Weidewechsels mit Portionsweide, die
nach Abgrasen gewechselt wird. Wenn möglich, sind die Tiere maximal 10 Tage auf
demselben Wiesenstück. Der darauf folgende Aufwuchs wird als Heu geemtet und
das entsprechende Weidestück erst dann wieder durch Tiere beweidet (genaues Zeit-
intervall ist nicht bekannt). Weidewechsel ist bei Jährlingen und Kitzen aufgrund der
örtlichen Begebenheiten nicht möglich.
Entwurmungen werden nicht prophylaktisch und ausschließlich bei klinisch auffälli-
gen Tieren (mit Durchfall, stumpfem Haarkleid...) zu Beginn der Trockenstehperio-
de durchgeführt und erfolgen mit Panacur® 2,5% orale Suspension (Wirkstoff Fen-
bendazol, Intervet, Wien) in der doppelten Dosierung für Schafe (10 mg Fenbendazol
pro kg KGW = 2 ml Panacur® 2,5% orale Suspension pro 5 kg KGW oral). Das Ge-
wicht der Tiere wird dabei geschätzt. Jungtiere, die zur Schlachtung bestimmt sind.

59
werden nicht entwurmt, die übrigen zweimalig vor dem ersten Weidegang im Alter
von drei Monaten. Coccidien-Behandlungen werden nicht durchgeführt.
4.2 Ergebnisse der klinischen Untersuchung
Bei der klinischen Untersuchung der Tiere fielen im Betrieb 1 zwei Tiere wegen ih-
rer kotverschmierten Afterregion, ein Tier mit weicherem Kot und ein Tier mit
Durchfall auf. Zwei Tiere zeigten Husten, eines davon auch struppiges Haarkleid, ein
Tier Nasenausfluss, ein Tier sowohl Husten als auch Nasenausfluss und ein Tier ne-
ben Husten und Nasenausfluss auch erhöhte innere Körpertemperatur. Diese Tiere
waren jedoch nicht dieselben wie diejenigen mit auffälligem Kotbefund. In Betrieb 2
wurde ebenfalls bei zwei Tieren eine kotverschmierte Analregion und bei vier Tieren
weicherer Kot bemerkt. Eines der beiden Tiere mit kotverschmierter Analregion war
außerdem hochgradig abgemagert. Respiratorische Symptome lagen bei keinem der
Tiere vor. In Betrieb 3 waren zwei Tiere mit Durchfall, struppigem Haarkleid und
hochgradiger Abmagerung zu verzeichnen, die bereits seit einigen Tagen zusätzlich
wegen verminderten Allgemeinbefindens, Inappetenz und nahezu ausbleibender
Milchleistung aufgefallen waren. Alle anderen untersuchten Tiere in Betrieb 3 zeig-
ten keinerlei klinische Auffälligkeiten.
4.3 Ergebnisse des Body Condition Scorings
Die prozentuale Verteilung des BCS zeigte sich für jeden Betrieb unterschiedlich. So
lagen - beide Betriebsbesuche zusammengenommen - im Betrieb 3 80 % der Tiere
im Idealbereich mit einem BCS von 2,5 bis 3,5, während 18 % der Tiere einen unter-
durchschnittlichen und 2 % einen überdurchschnittlichen BCS aufwiesen. Im Ver-
gleich dazu lag die prozentuale Verteilung im Betrieb 1 bei 11,5 % mit unterdurch-
schnittlichem, bei nur 69,2 % mit idealem und 19,2 % mit überdurchschnittlichem
BCS, während der Betrieb 2 den geringsten Anteil an unterdurchschnittlichem BCS
mit 7,7 % zeigte, mit 73,1 % mit idealem BCS im Mittelfeld und mit 19,2 % mit ü-
berdurchschnittlichem Ernährungszustand gleichauf mit Betrieb 1 lag (siehe Tab. 1).

60
Tabelle 1: Prozentuale Verteilung des BCS im betrieblichen Vergleich (Angabe in %; n=Anzahl der untersuchten Tiere)
Betrieb BCS < 2,5 2,5 < BCS < 3,5 BCS > 3,5
1 (n-78) 11,5 69,2 19,2
2 (n=52) 7,7 73,1 19,2
3(n=50) 18,0 80,0 2,0
Im Sommer lagen die prozentualen Anteile der Tiere mit BCS < 2,5 für alle Betriebe
in einem ähnlichen Bereich. Einen BCS im Bereich von 2,5 bis incl. 3,5 zeigten am
häufigsten Tiere in Betrieb 3, während die Anteile in Betrieb 1 und 2 niedriger wa-
ren. Tiere mit BCS > 3,5 konnten in Betrieb 3 kaum und in Betrieb 1 und 2 bei ca.
einem Fünftel der Tiere beobachtet werden (siehe Tab. 2).
Tabelle 2: Prozentuale Verteilung des BCS innerhalb der Betriebe im Sommer (Angabe in %; n=Anzahl der untersuchten Tiere)
Betrieb BCS < 2,5 2,5 < BCS < 3,5 BCS > 3,5
l(n=43) 16,3 60,5 23,3
2(n=27) 14,8 63,0 22,2
3 (n=24) 16,7 79,2 4,2
Im Herbst zeigten ca. ein Fünftel der Tiere in Betrieb 3 einen BCS < 2,5, während
dieser Anteil in Betrieb 1 und 2 sehr gering war bzw. 0% betrug. Einen BCS im Be-
reich von 2,5 bis incl. 3,5 zeigten alle Betriebe zu annähernd gleichen Anteilen, was
bedeutet, dass dieser Anteil in Betrieb 1 und 2 um ca. 20% gestiegen war, während er
in Betrieb 3 ungefähr gleich geblieben war. Der Anteil der Tiere mit BCS > 3,5 zeig-
te in allen Betrieben eine fallende Tendenz im Vergleich zum Sommer und sank in
Betrieb 3 auf 0 % ab (siehe Tab. 3).
Tabelle 3: prozentuale Verteilung des BCS innerhalb der Betriebe im Herbst (Angabe in %; n^Anzahl der untersuchten Tiere)
Betrieb BCS < 2,5 2,5 < BCS < 3,5 BCS > 3,5
1(n=35) 5,7 80,0 14,3
2(n=25) 0,0 84,0 16,0
3 (n=26) 19,2 80,8 0,0

61
• Statistische Auswertung
Betriebsübergreifend zeigte sich eine schwach signifikante Korrelation bezüglich des
Zusammenhangs der Ausscheidungsrate von Trichuris und dem BCS (r=-0,156 mit
p<0,05). Dieser Zusammenhang konnte im jahreszeitlichen Vergleich jedoch aus-
schließlich im Sommer festgestellt werden (r=-0,229 mit p<0,05). Im Herbst konnte
keine Korrelation zwischen BCS und dem Auftreten vermehrter Ei- bzw. Oozyste-
nausscheidung nachgewiesen werden.
Bei der betriebsspezifischen Betrachtung zeigte sich innerhalb Betrieb 1 keine Korre-
lation zwischen Eiausscheidungsrate und BCS, während in Betrieb 2 eine signifikan-
te Korrelation zwischen BCS und ausgeschiedenen Strongyloides-Eiem (r=0,297 mit
p<0,05) nachweisbar war. Auch in Betrieb 3 konnte keine Korrelation zwischen BCS
und dem Ausmaß der Eiausscheidung verzeichnet werden.
4.4 Ergebnisse der parasitologischen Untersuchungen
4.4.1 Kombiniertes Sedimentations-Flotations-Verfahren
Bei allen Betrieben war ein - jedoch unterschiedlich starker - Befall mit Eimerien,
MDS, Strongyhides, Trichuris und Skrjabinema, jedoch mit unterschiedlicher Präva-
lenz ausscheidender Tiere, nachweisbar (siehe Tab. 4). Eier von Moniezia konnten
jeweils, allerdings nur sehr vereinzelt, in Betrieb 1 im Sommer und Herbst, in Be-
trieb 3 im Herbst gefunden werden. Eier von Nematodirus wurden ausschließlich im
Betrieb 3 und auch hier nur im Sommer nachgewiesen.
4.4.1.1 Betriebliche Unterschiede
• Befallsraten im Betriebsvergleich
In Betrieb 2 und 3 waren alle untersuchten Tiere Eimerien-positiv, in Betrieb 1 ge-
ring weniger. Die Ausscheider-Prävalenz für MDS-Eier hingegen variierte stark im
Betriebsvergleich: Betrieb 1 zeigte deutlich weniger positive Proben als Betrieb 2
und 3. Deutliche Unterschiede waren auch für Skrjabinema zu verzeichnen, dessen
Eier in Betrieb 3 deutlich seltener zu finden waren bzw. auch für Trichuris, dessen
Eier wiederum in Betrieb 3 wesentlich häufiger beobachtet werden konnten. Für die
Prävalenz von Strongyloides-^oüiiven Proben lagen in allen Betrieben annähernd
ähnliche Ergebnisse vor, wobei in Betrieb 2 am meisten positive Proben und in Be-

62
trieb 1 am wenigsten positive Proben nachgewiesen werden konnten. Moniezia konn-
te in geringem Maße nur in Betrieb 1 und 3, Nematodirus ausschließlich in Betrieb 3
gefunden werden (siehe Tab. 4).
Tabelle 4: Prävalenz ausscheidender Tiere pro Betrieb (Angabe in %; n=Anzahl der untersuchten Tiere; E.=Eimerien; St.-Strongyloides; Sk.=Skrjabinema; T.-Trichuris; M.=Moniezia; N.=Nematodints)
Betrieb E. MDS St. Sk. T. M. N.
1(n=75) 92,0 64,0 64,0 58,7 9,3 2,7 0,0
2(n=51) 100,0 100,0 80,4 60,8 15,7 0,0 0,0
3(n=48) 100,0 95,8 75,0 20,8 50,0 4,2 8,3
• Befallsintensitäten im betrieblichen Vergleich
Der Median der Ausscheidungsrate für Eimerien-Oozysten war für Betrieb 1 und 2
gleich hoch, für Betrieb 3 deutlich höher. Gleiche Verhältnisse lagen für die Aus-
scheidung von MDS-Eiem vor, wovon in Betrieb 3 am meisten Eier zu finden waren.
Auch bezüglich Trichuris lag Betrieb 3 über Betrieb 1 und 2, allerdings war die
durchschnittliche Ausscheidungsrate generell niedrig. Bezüglich der Strongyloides-
Eier lieferten Betrieb 2 und 3 ähnliche Werte, während in Betrieb 1 weniger Eier
gefunden werden konnten. Skrjabinema-Eier waren hingegen mit einem Median von
0,0 deutlich weniger in Betrieb 3 zu verzeichnen, während Betrieb 2 den höchsten
Median aufwies. Moniezia konnte jeweils nur vereinzelt in Betrieb 1 und 3, Nemato-
dirus nur in Betrieb 3 nachgewiesen werden, weswegen der Median für diese Parasi-
ten in allen Betrieben 0,0 betrug (siehe Tab. 5).
Tabelle 5: Median der Ausscheidungsraten für die Einzelkotproben im Betriebsvergleich (n=Anzahl der untersuchten Tiere; 0=negativ, l=vereinzelt, 2=geringgradig, 3=gering- bis mittelgra- dig, 4=mittelgradig, 5=mittel- bis hochgradig, 6=hochgradig; E.=:Eimerien; St.-Strongyloides; Sk.-Skrjabinema; T.-Trichuris; M.=Moniezia; N.=Nematodirus)
Betrieb E. MDS St. Sk. T. M. N.
1(n=75) 2,0 2,0 1,0 1,0 0,0 0,0 0,0
2(n=51) 2,0 2,0 2,0 2,0 0,0 0,0 0,0
3(n=48) 4,0 4,0 2,0 0,0 0,5 0,0 0,0

63
• Statistische Auswertung
Betriebsübergreifend ließ sich für fast alle nachgewiesenen Parasiten, ausgenommen
Moniezia, eine signifikante positive Korrelation zwischen der Höhe der Ausschei-
dungsrate und der Haltungsform nachweisen, wobei die Ausscheidungsraten in Be-
trieb 1 tendenziell am niedrigsten und in Betrieb 3 am höchsten waren. Eine gegen-
sätzliche Tendenz in Form einer negativen Korrelation konnte einzig für Skrjabine-
ma verzeichnet werden (siehe Tab. 6).
Tabelle 6: Korrelation zwischen Ausscheidungsmenge und Haltung (Angabe als Korrelationskoeffizienten r; *=p<0,05; **=p<0,01; St.=Strongyloides\ Sk.-Skrjabinema; T.-Trichuris; N. -Nematodirus)
Eimerien MDS St. Sk. T. N.
Haltung 0,319** 0,585** 0,242** -0,253** 0,394** 0,206**
Im Vergleich der durchschnittlichen Ausscheidungsraten unterschieden sich die Be-
triebe im Kruskal-Wallis-Test bezüglich der nachgewiesenen Eier bzw. Oozysten
signifikant in allen differenzierten Parasiten (ausgenommen Moniezia und Nematodi-
rus, durch deren vereinzeltes Auftreten keine repräsentativen statistischen Ergebnisse
gewonnen werden konnten und die demnach hier nicht berücksichtigt werden). Bei
der durchschnittlichen Häufigkeit des Auftretens von Eimerien-Oozysten lag der
Betrieb 3 deutlich über Betrieb 1 und 2 (p=0,000). Der Unterschied zwischen diesen
war jedoch wiederum nicht signifikant. Bezüglich der durchschnittlichen Ausschei-
dung von MDS-Eiem unterschieden sich die einzelnen Betriebe signifikant
(p=0,000), wobei Betrieb 1 die geringste und Betrieb 3 die höchste Ausscheidungsra-
te zu verzeichnen hatte. Auch im Hinblick auf Strongyloides zeigte Betrieb 3 die
höchsten durchschnittlichen Ausscheidungsmengen, während Betrieb 1 die signifi-
kant geringsten aufwies (p=0,001). Betrieb 3 lieferte hingegen signifikant geringere
Werte bei der Ausscheidung von Skrjabinema-E'iem als Betrieb 1 und 2 (p=0,000).
Die durchschnittliche Ausscheidungsmenge für Trichuris-Eier lag im Betrieb 3 signi-
fikant höher (p=0,000), während sich Betrieb 1 und 2 nicht signifikant unterschieden.
In der Zusammenschau der betrieblichen Untersuchungsergebnisse konnten außer-
dem Zusammenhänge hinsichtlich des parallelen Auftretens mancher Parasiten-Eier
bzw. -oozysten beobachtet werden. So zeigten sich betriebsübergreifend signifikante
Korrelationen betreffend des gemeinsamen Auftretens von Magen-Darm-
Strongyliden (MDS) mit Eimerien, Strongyloides und Trichuris sowie von Eimerien
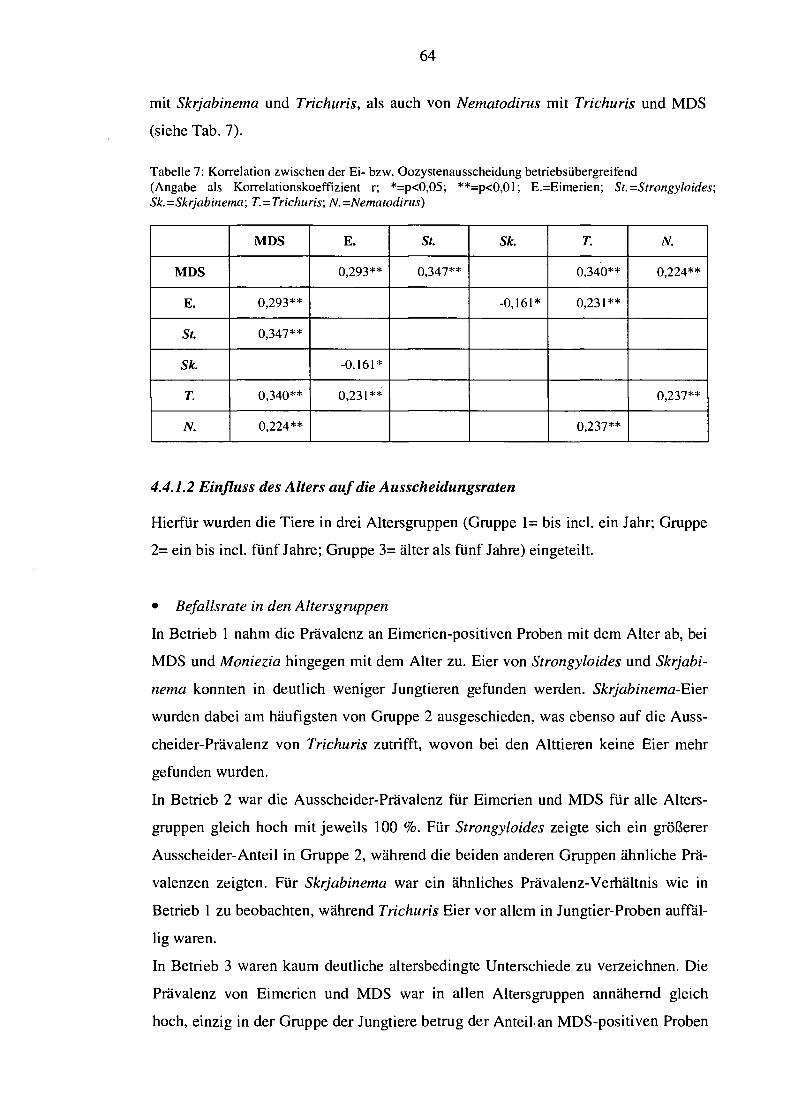
64
mit Skrjabinema und Trichuris, als auch von Nematodirus mit Trichuris und MDS
(siehe Tab. 7).
Tabelle 7: Korrelation zwischen der Ei- bzw. Oozystenausscheidung betriebsübergreifend (Angabe als Korrelationskoeffizient r; *=p<0,05; **=p<0,01; E.=Eimerien; St.=Strongyloides; Sk.-Skrjabinema; T.=Tricfiuris; N.=Nematodirus)
MDS E. St. Sk. T. N.
MDS 0,293** 0,347** 0,340** 0,224**
E. 0,293** -0,161* 0,231**
St. 0,347**
Sk. -0,161*
T. 0,340** 0,231** 0,237**
N. 0,224** 0,237**
4.4.1.2 Einßuss des Alters auf die Ausscheidungsraten
Hierfür wurden die Tiere in drei Altersgruppen (Gruppe 1= bis incl. ein Jahr; Gruppe
2= ein bis incl. fünf Jahre; Gruppe 3= älter als fünf Jahre) eingeteilt.
• Befallsrate in den Altersgruppen
In Betrieb 1 nahm die Prävalenz an Eimerien-positiven Proben mit dem Alter ab, bei
MDS und Moniezia hingegen mit dem Alter zu. Eier von Strongyloides und Skrjabi-
nema konnten in deutlich weniger Jungtieren gefunden werden. Skrjabinema-Eier
wurden dabei am häufigsten von Gruppe 2 ausgeschieden, was ebenso auf die Auss-
cheider-Prävalenz von Trichuris zutrifft, wovon bei den Alttieren keine Eier mehr
gefunden wurden.
In Betrieb 2 war die Ausscheider-Prävalenz für Eimerien und MDS für alle Alters-
gruppen gleich hoch mit jeweils 100 %. Für Strongyloides zeigte sich ein größerer
Ausscheider-Anteil in Gruppe 2, während die beiden anderen Gruppen ähnliche Prä-
valenzen zeigten. Für Skrjabinema war ein ähnliches Prävalenz-Verhältnis wie in
Betrieb 1 zu beobachten, während Trichuris Eier vor allem in Jungtier-Proben auffäl-
lig waren.
In Betrieb 3 waren kaum deutliche altersbedingte Unterschiede zu verzeichnen. Die
Prävalenz von Eimerien und MDS war in allen Altersgruppen annähernd gleich
hoch, einzig in der Gruppe der Jungtiere betrug der Anteil an MDS-positiven Proben

65
lediglich 86,7 %. Die Prävalenz von Skrjabinema war in Gruppe 3 etwas geringer,
während Trichuris-Eitx bei Jungtieren gering häufiger zu beobachten waren. Monie-
zia konnte einzig in Gruppe 3, Nematodirus in jeweils vergleichbaren Anteilen nach-
gewiesen werden (siehe Tab. 8).
Tabelle 8: Prävalenz der Ausscheider in den Altersgruppen (Angabe in %; n=Anzahi der untersuchten Tiere; E.=Eimerien; St.=Strongyloides; Sk.=Skrjabinema\ T.=Tnchuris\ M.=Moniezia\ N.-Nematodirus)
Betrieb Alter E. MDS St. Sk. T. M. N.
1
1 (n=16) 100,0 0,0 6,2 18,7 6,2 0,0 0,0
2(n=47) 91,5 78,7 80,9 74,5 12,8 2,1 0,0
3(n=I2) 83,3 91,7 75,0 50,0 0,0 8.3 0,0
2
1(n=16) 100,0 100,0 68,7 31,2 43,7 0,0 0,0
2(n-20) 100,0 100,0 95,0 80,0 5,0 0,0 0,0
3(n=13) 100,0 100,0 69,2 69,2 0,0 0,0 0,0
3
1(n-15) 100,0 86,7 80,0 20,0 60,0 0,0 6,7
2(n=17) 100,0 100,0 70,6 29,4 47,1 0,0 11,8
3(n=15) 100,0 100,0 73,3 13,3 40,0 13,3 6,7
• Befallsintensität in den Altersgruppen
In Betrieb 1 waren vor allem Unterschiede zwischen Jungtieren und Adulten evident.
So war die Ausscheidungsrate von Eimerien-Oozysten in der Gruppe der Jungtiere
wesentlich höher, während alle anderen Parasiteneier deutlich seltener beobachtet
werden konnten (Median 0,0). Der Median für MDS- und Strongyloides-Eier war bei
den adulten Tieren demnach deutlich höher. Die Altersgruppen 2 und 3 unterschie-
den sich hingegen kaum: einzig bezüglich Skrjabinema konnte eine höhere Aus-
scheidungsrate in Altersgruppe 2 beobachtet werden. Die Ausscheidungsrate für die
übrigen Parasiten war in allen Altersgruppen sehr niedrig.
In Betrieb 2 zeichnete sich indes der Unterschied zwischen den Altergruppen weni-
ger deutlich ab. Die Ausscheidungsrate für Eimerien, MDS und Strongyloides zeigte
demnach ähnliche Werte. Unterschiede konnten bei Skrjabinema beobachtet werden,
wo die Ausscheidungsrate in Altersgruppe 2 am höchsten und bei den Jungtieren am
niedrigsten lag. Die Ausscheidungsrate für die übrigen Parasiten war generell sehr
niedrig.
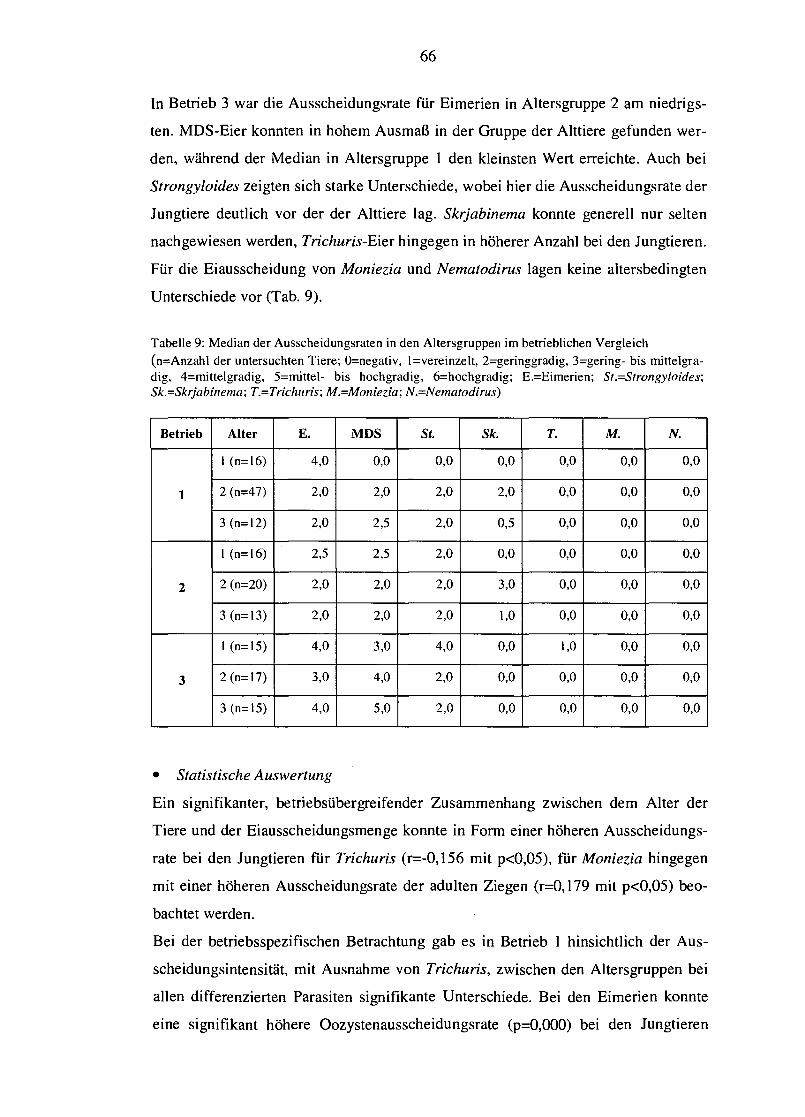
66
In Betrieb 3 war die Ausscheidungsrate für Eimerien in Altersgruppe 2 am niedrigs-
ten. MDS-Eier konnten in hohem Ausmaß in der Gruppe der Alttiere gefunden wer-
den, während der Median in Altersgruppe 1 den kleinsten Wert erreichte. Auch bei
Strongyloides zeigten sich starke Unterschiede, wobei hier die Ausscheidungsrate der
Jungtiere deutlich vor der der Alttiere lag. Skrjabinema konnte generell nur selten
nachgewiesen werden, Trichuris-Eier hingegen in höherer Anzahl bei den Jungtieren.
Für die Eiausscheidung von Moniezia und Nematodirus lagen keine altersbedingten
Unterschiede vor (Tab. 9).
Tabelle 9: Median der Ausscheidungsraten in den Altersgruppen im betrieblichen Vergleich (n=Anzahl der untersuchten Tiere; 0=negativ, l=vereinzelt, 2=geringgradig, 3=gering- bis mittelgra- dig, 4=mittelgradig, 5=mittel- bis hochgradig, 6=hochgradig; E.=Eimerien; St.-Strongyloides; Sk.=Skrjabinema; T=Trichuris; M.=Moniezia; N.=Nematodirus)
Betrieb Alter E. MDS St. Sk. T. M. A^.
1
1 (n=16) 4,0 0,0 0,0 0,0 0,0 0,0 0,0
2(n=47) 2,0 2,0 2,0 2,0 0,0 0,0 0,0
3(n=12) 2,0 2,5 2,0 0,5 0,0 0,0 0,0
2
1 (n=i6) 2,5 2,5 2,0 0,0 0,0 0,0 0,0
2(n=20) 2,0 2,0 2,0 3,0 0,0 0,0 0,0
3(n=i3) 2,0 2,0 2,0 1,0 0,0 0,0 0,0
3
1(n=15) 4,0 3,0 4,0 0,0 1,0 0,0 0,0
2(n=17) 3,0 4,0 2,0 0,0 0,0 0,0 0,0
3(n=15) 4,0 5,0 2,0 0,0 0,0 0,0 0,0
• Statistische Auswertung
Ein signifikanter, betriebsübergreifender Zusammenhang zwischen dem Alter der
Tiere und der Eiausscheidungsmenge konnte in Form einer höheren Ausscheidungs-
rate bei den Jungtieren für Trichuris (r=-0,156 mit p<0,05), für Moniezia hingegen
mit einer höheren Ausscheidungsrate der adulten Ziegen (r=0,179 mit p<0,05) beo-
bachtet werden.
Bei der betriebsspezifischen Betrachtung gab es in Betrieb 1 hinsichtlich der Aus-
scheidungsintensität, mit Ausnahme von Trichuris, zwischen den Altersgruppen bei
allen differenzierten Parasiten signifikante Unterschiede. Bei den Eimerien konnte
eine signifikant höhere Oozystenausscheidungsrate (p=0,000) bei den Jungtieren

67
gegenüber den Adulten verzeichnet werden, wobei sich Gruppe 2 und 3 nicht signifi-
kant voneinander unterschieden, jedoch insgesamt ein fallende Tendenz der Befalls-
intensität auffällig war. Im Gegensatz zu den Eimerien lag die Eiausscheidungsrate
der MDS bei den Jungtieren signifikant unter der der adulten Tiere (p=0,000), die
Unterschiede zwischen Gruppe 2 und 3 waren jedoch auch hier nicht signifikant.
Ebenso wie bei den MDS war die Ausscheidungsrate von Strongyloides-Eiem in der
Gruppe 1 signifikant niedriger (p=0,000) als bei den adulten Tieren, zwischen denen
wiederum keine Unterschiede vorlagen. Bei Skrjabinema ergab sich nur zwischen
Gruppe l und Gruppe 2 ein signifikanter Unterschied (p=0,001), wobei die Aus-
scheidungsrate der Gruppe 2 deutlich höher war. Hier lag also in Gruppe 2 die höchs-
te Ausscheidungsrate vor, jedoch unterschieden sich Gruppe 1 und 3, aber auch 2
und 3 nicht signifikant voneinander.
In Betrieb 2 konnten altersabhängige Unterschiede bei Skrjabinema und Trichuris
nachgewiesen werden, wobei die Unterschiede ausschließlich zwischen Jungtieren
und Adulten, jedoch nicht innerhalb der Adulten vorlagen. Die Jungtiere wiesen eine
signifikant niedrigere Eiausscheidung von Skrjabinema gegenüber der Gruppe 2
(p=0,001) auf, unterschieden sich hinsichtlich der Gruppe 3 jedoch nicht signifikant.
Der Infektionsverlauf ist also vergleichbar mit Betrieb 1. Die Befallsintensität der
Jungtiere mit Trichuris lag hier signifikant über der der adulten Tiere (p=0,001).
In Betrieb 3 zeigte sich eine signifikante Altersabhängigkeit in der Befallsintensität
bei MDS, wobei sich die Unterschiede wiederum nur auf den Vergleich zwischen
Jungtieren und Adulten beschränkten und zwischen den adulten Gruppen keine signi-
fikanten Abweichungen zu verzeichnen waren. Die MDS-Ausscheidungsrate der
Jungtiere lag deutlich unter der beider adulter Gruppen (p=0,005), was sich mit den
Beobachtungen in Betrieb 1 deckt. Die Jungtiere schieden außerdem deutlich mehr
(p=0,051) Strongyloides-Eier aus als die Gruppe 2, das Ergebnis war jedoch knapp
außerhalb der statistischen Signifikanz. Dies steht in Kontrast zu Beobachtungen im
ersten Betrieb, wo die Ausscheidungsrate der Jungtiere signifikant unter der der adul-
ten Tiere lag. Die Unterschiede zu höheren Ausscheidungsraten in Gruppe 3 waren
deutlich, jedoch ebenfalls nicht signifikant.
4.4.1.3 Saisonale Variation der Eiausscheidung
Innerhalb der Betriebe konnten jahreszeitliche Variationen in der Ei- bzw. Oo-
zystenausscheidungsrate beobachtet werden.

68
• Befallsrate im jahreszeitlichen Vergleich
Im Herbst war die Prävalenz der Eimerien-Ausscheider für Betrieb 1 niedriger als im
Sommer, während in Betrieb 2 und 3 weiterhin alle getesteten Tiere positiv waren.
Bezüglich der MDS-Eier zeigte sich in Betrieb 1 ebenfalls ein gegenüber Sommer
sinkender Anteil an Ausscheidern, während in Betrieb 2 und 3 annähernd alle Kot-
proben positiv waren. Das Auftreten von Strongyloides-Exem war in Betrieb 1 und 3
im Herbst seltener als im Sommer, während der Anteil an Ausscheidern in Betrieb 2
auf annähernd 100 % anstieg. Der Anteil an SA:r/a£'mema-Ausscheidern in Betrieb 1
und 2 war zwar geringer als im Sommer, jedoch immer noch doppelt so hoch wie in
Betrieb 3, wo der Anteil allerdings leicht angestiegen war. Die Prävalenz an Trichu-
m-Eiem erreichte in Betrieb 3 im Sommer wie auch im Herbst ähnliche Werte und
lag jeweils um ein Vielfaches höher als in Betrieb 1 und 2. Moniezia-E'ier konnten
vereinzelt in Betrieb 1 und 3 nachgewiesen werden, während Nematodir us-Eier nur
im Sommer in Betrieb 3 beobachtet werden konnten (siehe Tab. 10).
Tabelle 10: Prävalenz ausscheidender Tiere im jahreszeitlichen Vergleich (Angabe in %;n=Anzahl der untersuchten Tiere; Monat 0=Sommer, Monat l=Herbst; E.^Eimerien; St.=Strongyloides\ Sk.=Skrjabinema; T.=Trichuris; M.-Moniezia; N.-Nematodirus)
Betrieb Monat E. MDS St. Sk. T. M. N.
1 0 (n=43) 100,0 79,1 76,7 62,8 7,0 2,3 0,0
1(n-32) 81,2 43,7 46,9 53,1 12,5 3,1 0,0
2 0(n=27) 100,0 100,0 66,7 70,4 14,8 0,0 0,0
1 (n=22) 100,0 100,0 95,5 50,0 18,2 0,0 0,0
3
0(n=24) 100,0 91,7 79,2 16,7 45,8 0,0 16,7
1 (n=24) 100,0 100,0 70,8 25,0 54,2 8,3 0,0
• Befallsintensität im jahreszeitlichen Vergleich
Der Median der Ausscheidungsrate für Eimerien-Oozysten zeigte in Betrieb 1 und 3
im Sommer geringere Werte als im Herbst, während in Betrieb 2 das Gegenteil zu
beobachten war. Eine ähnliche Tendenz konnte ebenso bezüglich den MDS-Eier ver-
zeichnet werden. Strongyloides-Eier wurden in Betrieb 1 vermehrt im Sommer aus-
geschieden, während der Medianwert in Betrieb 2 und 3 keine jahreszeitlichen Un-
terschiede zeigte. Hinsichtlich SA:r/a^mema-Ausscheidungsraten zeigten sich wieder-
um in Betrieb 1 und 3 keine Unterschiede im Medianwert, während in Betrieb 2 die
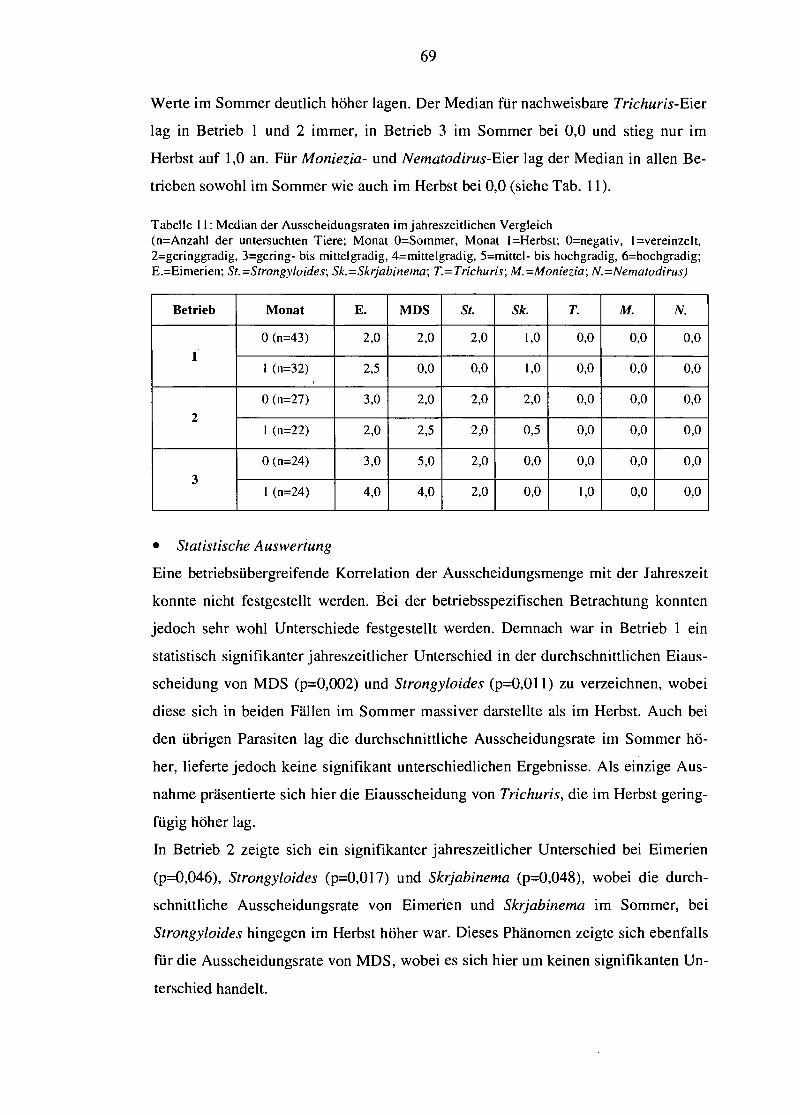
69
Werte im Sommer deutlich höher lagen. Der Median für nachweisbare Trichuris-Eier
lag in Betrieb 1 und 2 immer, in Betrieb 3 im Sommer bei 0,0 und stieg nur im
Herbst auf 1,0 an. Für Moniezia- und Nematodirus-E\er lag der Median in allen Be-
trieben sowohl im Sommer wie auch im Herbst bei 0,0 (siehe Tab. 11).
Tabelle 11: Median der Ausscheidungsraten im jahreszeitlichen Vergleich (n=Anzahl der untersuchten Tiere; Monat O^Sommer, Monat l=Herbst; 0=negativ, l=vereinzelt, 2=geringgradig, 3=gering- bis mittelgradig, 4=niittelgradig, 5=mittel- bis hochgradig, 6=hochgradig; E.=Eimerien; St.=Strongyloides; Sk.=Skrjabinema\ T.=Trichuris; M.-Moniezia; N.=Nematodirus)
Betrieb Monat E. MDS St. Sk. T. M. N.
1 0 (n=43) 2,0 2,0 2,0 1,0 0,0 0,0 0,0
1 (n=32) 2,5 0,0 0,0 1,0 0,0 0,0 0,0
2 0(n=27) 3,0 2,0 2,0 2,0 0,0 0,0 0,0
1 (n=22) 2,0 2,5 2,0 0,5 0,0 0,0 0,0
3 0(n=24) 3,0 5,0 2,0 0,0 0,0 0,0 0,0
1 (n=24) 4,0 4,0 2,0 0,0 1,0 0,0 0,0
• Statistische Auswertung
Eine betriebsübergreifende Korrelation der Ausscheidungsmenge mit der Jahreszeit
konnte nicht festgestellt werden. Bei der betriebsspezifischen Betrachtung konnten
jedoch sehr wohl Unterschiede festgestellt werden. Demnach war in Betrieb 1 ein
statistisch signifikanter jahreszeitlicher Unterschied in der durchschnittlichen Eiaus-
scheidung von MDS (p=0,002) und Strongyloides (p=0,011) zu verzeichnen, wobei
diese sich in beiden Fällen im Sommer massiver darstellte als im Herbst. Auch bei
den übrigen Parasiten lag die durchschnittliche Ausscheidungsrate im Sommer hö-
her, lieferte jedoch keine signifikant unterschiedlichen Ergebnisse. Als einzige Aus-
nahme präsentierte sich hier die Eiausscheidung von Trichuris, die im Herbst gering-
fügig höher lag.
In Betrieb 2 zeigte sich ein signifikanter jahreszeitlicher Unterschied bei Eimerien
(p=0,046), Strongyloides (p=0,017) und Skrjabinema (p=0,048), wobei die durch-
schnittliche Ausscheidungsrate von Eimerien und Skrjabinema im Sommer, bei
Strongyloides hingegen im Herbst höher war. Dieses Phänomen zeigte sich ebenfalls
für die Ausscheidungsrate von MDS, wobei es sich hier um keinen signifikanten Un-
terschied handelt.
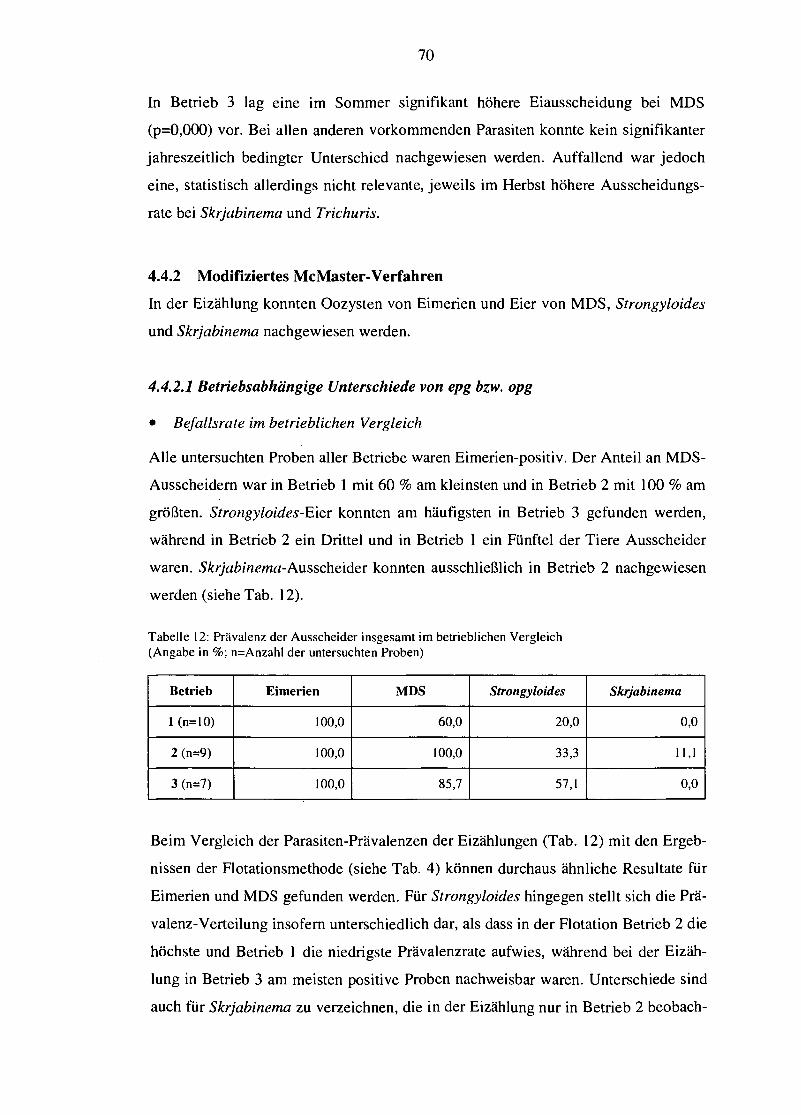
70
In Betrieb 3 lag eine im Sommer signifikant höhere Eiausscheidung bei MDS
(p=0,000) vor. Bei allen anderen vorkommenden Parasiten konnte kein signifikanter
jahreszeitlich bedingter Unterschied nachgewiesen werden. Auffallend war jedoch
eine, statistisch allerdings nicht relevante, jeweils im Herbst höhere Ausscheidungs-
rate bei Skrjabinema und Trichuris.
iXA.l Modiflziertes McMaster-Verfahren
In der Eizählung konnten Oozysten von Eimerien und Eier von MDS, Strongyloides
und Skrjabinema nachgewiesen werden.
4.4.2.1 Betriebsabhängige Unterschiede von epg bzw. opg
• Befallsrate im betrieblichen Vergleich
Alle untersuchten Proben aller Betriebe waren Eimerien-positiv. Der Anteil an MDS-
Ausscheidem war in Betrieb 1 mit 60 % am kleinsten und in Betrieb 2 mit 100 % am
größten. Strongyloides-Extx konnten am häufigsten in Betrieb 3 gefunden werden,
während in Betrieb 2 ein Drittel und in Betrieb 1 ein Fünftel der Tiere Ausscheider
waren. Skrjabinema-A.us%ch&\d&v konnten ausschließlich in Betrieb 2 nachgewiesen
werden (siehe Tab. 12).
Tabelle 12: Prävalenz der Ausscheider insgesamt im betrieblichen Vergleich (Angabe in %; n=Anzahl der untersuchten Proben)
Betrieb Eimerien MDS Strongyloides Skrjabinema
l(n=10) 100,0 60,0 20,0 0,0
2 (n=9) 100,0 100,0 33,3 11,1
3 (n=7) 100,0 85,7 57,1 0,0
Beim Vergleich der Parasiten-Prävalenzen der Eizählungen (Tab. 12) mit den Ergeb-
nissen der Flotationsmethode (siehe Tab. 4) können durchaus ähnliche Resultate für
Eimerien und MDS gefunden werden. Für Strongyloides hingegen stellt sich die Prä-
valenz-Verteilung insofern unterschiedlich dar, als dass in der Flotation Betrieb 2 die
höchste und Betrieb 1 die niedrigste Prävalenzrate aufwies, während bei der Eizäh-
lung in Betrieb 3 am meisten positive Proben nachweisbar waren. Unterschiede sind
auch für Skrjabinema zu verzeichnen, die in der Eizählung nur in Betrieb 2 beobach-

71
tet werden konnte, während in der Flotation Betrieb 1 und 2 zu annähernd gleichen
Teilen positive Proben lieferte und Betrieb 3 deutlich geringere Prävalenzen aufwies.
• Befallsintensität im betrieblichen Vergleich
Eklatante Unterschiede zeigten sich in Bezug auf die Anzahl an MDS-Eiem, aber
auch hinsichtlich der Eimerien, bei denen Betrieb 3 deutlich höhere Medianwerte der
Oozysten-Anzahl aufwies. Die Resultate für Strongyloides waren für Betrieb 2 und 3
ähnlich, während Betrieb 1 geringere Werte lieferte. Eier von Skrjabinema waren nur
in Betrieb 2 nachweisbar (siehe Tab. 13).
Tabelle 13: Median der Eizählungsergebnisse im Betriebsvergleich (n=Anzahl der untersuchten Proben; Angabe des epg/opg-Median mit Minimum- (=Min) und Maxi- mumwert (=Max))
Betrieb Wert Eimerien MDS Strongyloides Skrjabinema
l(n=IO) Median
Min
Max
500,0
350,0
6400,0
50,0
0,0
250,0
0,0
0,0
100,0
0,0
0,0
0,0
2 (n=9)
Median
Min
Max
650,0
100,0
3650,0
450,0
100,0
1650,0
0,0
0,0
350,0
0,0
0,0
50,0
3 (n=7) Median
Min
Max
1000,0
450,0
9850,0
850,0
0,0
3150,0
50,0
0,0
300,0
0,0
0,0
0,0
Der Vergleich der Befallsintensitäten der Eizählung (siehe Tab. 13) mit der Flotation
(siehe Tab. 5) erbrachte eine tendenziell gute Übereinstimmung der Untersuchungs-
ergebnisse, d.h. Betrieb 3 zeigte sowohl in der Flotation wie auch in der Eizählung
deutlich höhere Ergebnisse bei MDS und Eimerien als die diesbezüglich ähnliche
Werte liefernden Betriebe 1 und 2. Strongyloides-Eier wurden in beiden Untersu-
chungsmethoden in Betrieb 1 in geringstem Ausmaß gefunden, während Betrieb 2
und 3 ähnliche Resultate zeigten. Skrjabinema-Eitr konnten in der Eizählung nur in
Betrieb 2, in der Flotation in geringerem Maße auch in Betrieb 1 nachgewiesen wer-
den.
• Statistische Auswertung
Eine statistisch signifikante Korrelation zwischen Eiausscheidung und Haltung konn-
te für die Eier von MDS ermittelt werden (r=0,661 mit p<0,01). Dieser Unterschied
zwischen den Betrieben zeigte sich in signifikant niedrigeren (p=0,002) epg-Werten
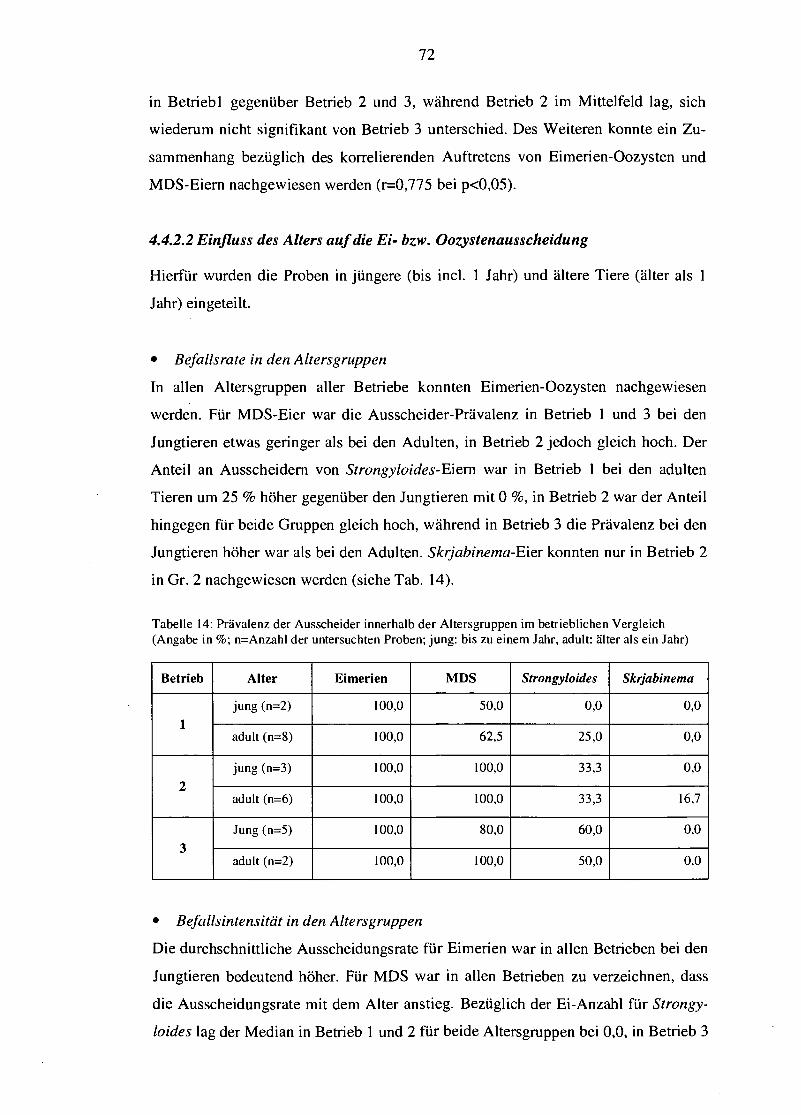
72
in Betriebt gegenüber Betrieb 2 und 3, während Betrieb 2 im Mittelfeld lag, sich
wiederum nicht signifikant von Betrieb 3 unterschied. Des Weiteren konnte ein Zu-
sammenhang bezüglich des korrelierenden Auftretens von Eimerien-Oozysten und
MDS-Eiem nachgewiesen werden (r=0,775 bei p<0,05).
4.4.2.2 Einfluss des Alters auf die Ei- bzw. Oozystenausscheidung
Hierfür wurden die Proben in jüngere (bis incl. t Jahr) und ältere Tiere (älter als 1
Jahr) eingeteilt.
• Befallsrate in den Altersgruppen
In allen Altersgruppen aller Betriebe konnten Eimerien-Oozysten nachgewiesen
werden. Für MDS-Eier war die Ausscheider-Prävalenz in Betrieb 1 und 3 bei den
Jungtieren etwas geringer als bei den Adulten, in Betrieb 2 jedoch gleich hoch. Der
Anteil an Ausscheidern von Strongyloides-Eiem war in Betrieb 1 bei den adulten
Tieren um 25 % höher gegenüber den Jungtieren mit 0 %, in Betrieb 2 war der Anteil
hingegen für beide Gruppen gleich hoch, während in Betrieb 3 die Prävalenz bei den
Jungtieren höher war als bei den Adulten. Skrjabinema-Eier konnten nur in Betrieb 2
in Gr. 2 nachgewiesen werden (siehe Tab. 14).
Tabelle 14: Prävalenz der Ausscheider innerhalb der Altersgruppen im betrieblichen Vergleich (Angabe in %; n=Anzahl der untersuchten Proben; jung: bis zu einem Jahr, adult: älter als ein Jahr)
Betrieb Alter Eimerien MDS Strongyloides Skrjabinema
1 jung (n=2) 100,0 50,0 0,0 0,0
adult (n=8) 100,0 62,5 25,0 0,0
2 jung (n==3) 100,0 100,0 33,3 0,0
adult (n=6) 100,0 100,0 33,3 16,7
3 Jung (n=5) 100,0 80,0 60,0 0,0
adult (n=2) 100,0 100,0 50,0 0,0
• Befallsintensität in den Altersgruppen
Die durchschnittliche Ausscheidungsrate für Eimerien war in allen Betrieben bei den
Jungtieren bedeutend höher. Für MDS war in allen Betrieben zu verzeichnen, dass
die Ausscheidungsrate mit dem Alter anstieg. Bezüglich der Ei-Anzahl für Strongy-
loides lag der Median in Betrieb 1 und 2 für beide Altersgruppen bei 0,0, in Betrieb 3
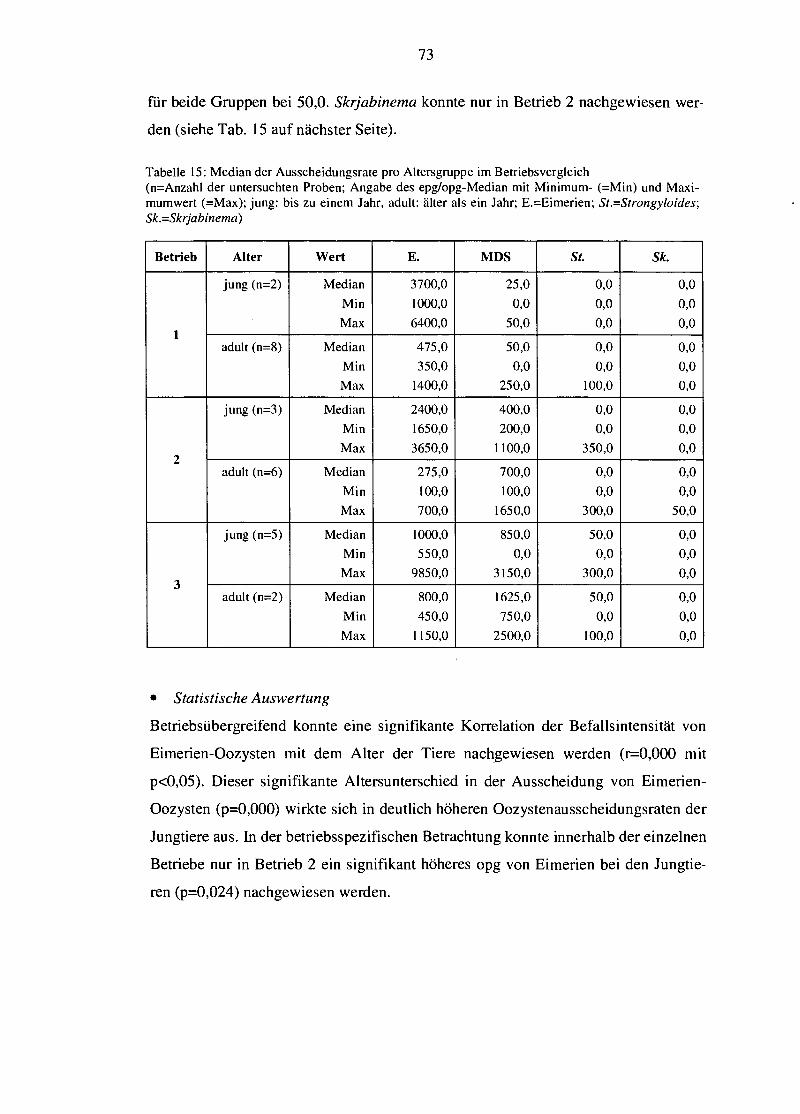
73
für beide Gruppen bei 50,0. Skrjabinema konnte nur in Betrieb 2 nachgewiesen wer-
den (siehe Tab. 15 auf nächster Seite).
Tabelle 15: Median der Ausscheidungsrate pro Altersgruppe im Betriebsvergleich (n=Anzahl der untersuchten Proben; Angabe des epg/opg-Median mit Minimum- (=Min) und Maxi- mumwert (=Max); jung: bis zu einem Jahr, adult: älter als ein Jahr; E.=Eimerien; St.=Strongyloides; Sk.=Skrjabinemä)
Betrieb Alter Wert E, MDS St. Sk.
1
jung (n=2) Median
Min
Max
3700,0
1000,0
6400,0
25,0
0,0
50,0
0,0
0,0
0,0
0,0
0,0
0,0
adult (n=8) Median
Min
Max
475,0
350,0
1400,0
50,0
0,0
250,0
0,0
0,0
100,0
0,0
0,0
0,0
2
jung (n=3) Median
Min
Max
2400,0
1650,0
3650,0
400,0
200,0
1100,0
0,0
0,0
350,0
0,0
0,0
0,0
adult (n=6) Median
Min
Max
275,0
100,0
700,0
700,0
100,0
1650,0
0,0
0,0
300,0
0,0
0,0
50,0
3
jung (n=5) Median
Min
Max
1000,0
550,0
9850,0
850,0
0,0
3150,0
50,0
0,0
300,0
0,0
0,0
0,0
adult (n=2) Median
Min
Max
800,0
450,0
1150,0
1625,0
750,0
2500,0
50,0
0,0
100,0
0,0
0,0
0,0
• Statistische Auswertung
Betriebsübergreifend konnte eine signifikante Korrelation der Befallsintensität von
Eimerien-Oozysten mit dem Alter der Tiere nachgewiesen werden (r=0,000 mit
p<0,05). Dieser signifikante Altersunterschied in der Ausscheidung von Eimerien-
Oozysten (p=0,000) wirkte sich in deutlich höheren Oozystenausscheidungsraten der
Jungtiere aus. In der betriebsspezifischen Betrachtung konnte innerhalb der einzelnen
Betriebe nur in Betrieb 2 ein signifikant höheres opg von Eimerien bei den Jungtie-
ren (p=0,024) nachgewiesen werden.

74
4.4.2.3 Saisonale Variation der Ausscheidungsraten (epg und opg)
• Befallsrate im jahreszeitlichen Vergleich
In beiden Jahreszeiten konnten in allen Kotproben aller Betriebe Eimerien-Oozysten
nachgewiesen werden. Die Prävalenz der MDS-Ausscheider war in Betrieb 1 im
Sommer höher, in Betrieb 2 jeweils 100 % und in Betrieb 3 im Herbst höher. Beim
Anteil der Strongyloides-Ausschsider war in allen Betrieben eine steigende Tendenz
von Sommer auf Herbst zu verzeichnen. Skrjabinema war nur in Betrieb 2 und hier
nur im Herbst nachweisbar (siehe Tab. 16).
Tabelle 16: Prävalenz der Ausscheider im jahreszeitlichen Vergleich (Angabe in %; n=Anzahl der untersuchten Proben)
Betrieb Monat Eimerien MDS Strongyloides Skrjabinema
1 Sommer (n=6) 100,0 83,3 16,7 0,0
Herbst (n=4) 100,0 25,0 25,0 0,0
2 Sommer(n=5) 100,0 100,0 0,0 0,0
Herbst (n=4) 100,0 100,0 75,0 25,0
3 Sommer (n=4) 100,0 75,0 25,0 0,0
Herbs t(n=3) 100,0 100,0 100,0 0,0
• Befallsintensität im jahreszeitlichen Vergleich
In allen Betrieben konnte eine deutliche Reduktion der Eiausscheidung hinsichtlich
MDS mit fortschreitendem Jahresverlauf beobachtet werden. Ebenso verhielt es sich
für die Eimerien-Oozystenausscheidung in Betrieb 1 und 2, während in Betrieb 3 im
Herbst gering höhere Oozystenausscheidungsraten nachweisbar waren. Bezüglich
der S^rongy/o/i/e^-Ausscheidungsrate war in den Betrieben 2 und 3 eine Zunahme im
Herbst gegenüber Sommer zu verzeichnen, während der Medianwert in Betrieb 1 für
beide Jahreszeiten bei 0,0 lag. Skrjabinema konnte einzig in Betrieb 2 und auch hier
nur im Herbst nachgewiesen werden (siehe Tab. 17).
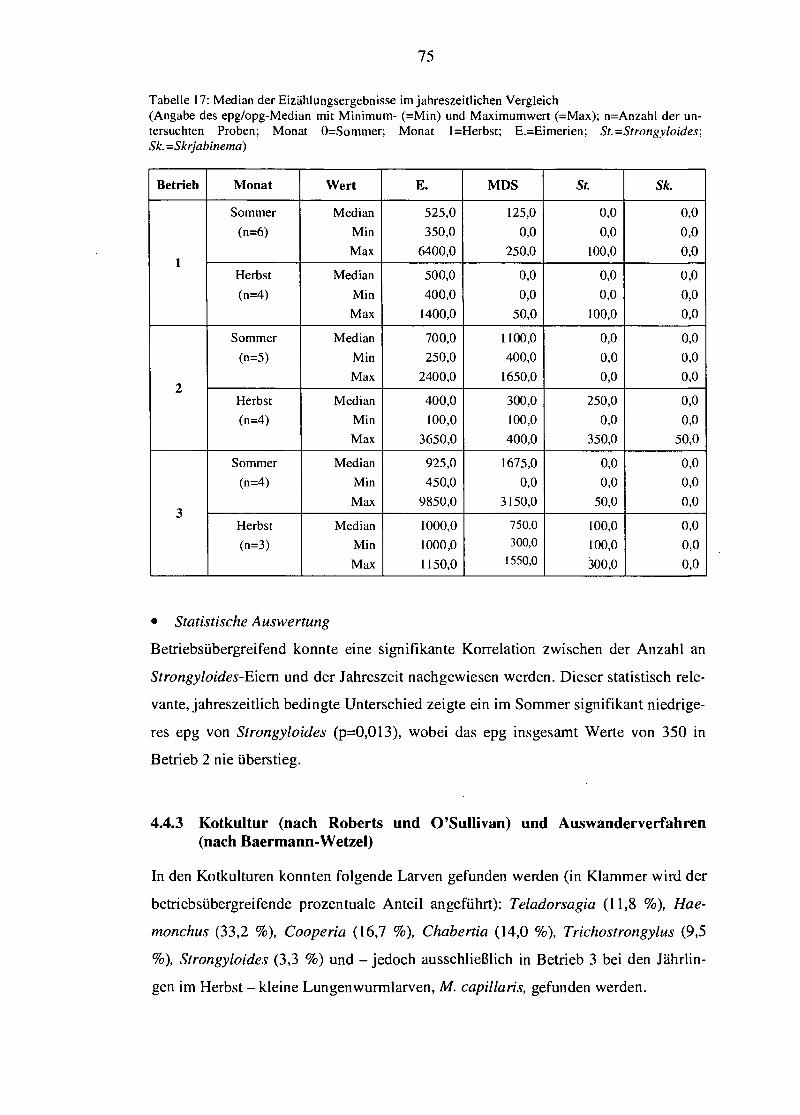
75
Tabelle 17: Median der Eizählungsergebnisse im jahreszeitlichen Vergleich (Angabe des epg/opg-Median mit Minimum- (=Min) und Maximumwert (=Max); n=Anzahl der un- tersuchten Proben; Monat 0=Sommer; Monat l=Herbst; E.=Eimerien; St.=Strongyloides\ Sk. -Skrjabinemä)
Betrieb Monat Wert E. MDS St. Sk.
1
Sommer
(n=6)
Median
Min
Max
525,0
350,0
6400,0
125,0
0,0
250,0
0,0
0,0
100,0
0,0
0,0
0,0
Herbst
(n=4)
Median
Min
Max
500,0
400,0
1400,0
0,0
0,0
50,0
0,0
0,0
100,0
0,0
0,0
0,0
2
Sommer
(n=5)
Median
Min
Max
700,0
250,0
2400,0
1100,0
400,0
1650,0
0,0
0,0
0,0
0,0
0,0
0,0
Herbst
(n-4)
Median
Min
Max
400,0
100,0
3650,0
300,0
100,0
400,0
250,0
0,0
350,0
0,0
0,0
50,0
3
Sommer
(n=4)
Median
Min
Max
925,0
450,0
9850,0
1675,0
0,0
3150,0
0,0
0,0
50,0
0,0
0,0
0,0
Herbst
(n=3)
Median
Min
Max
1000,0
1000,0
1150,0
750,0 300,0
1550,0
100,0
100,0
300,0
0,0
0,0
0,0
• Statistische Auswertung
Betriebsübergreifend konnte eine signifikante Korrelation zwischen der Anzahl an
Strongyloides-Eiem und der Jahreszeit nachgewiesen werden. Dieser statistisch rele-
vante, jahreszeitlich bedingte Unterschied zeigte ein im Sommer signifikant niedrige-
res epg von Strongyloides (p=0,013), wobei das epg insgesamt Werte von 350 in
Betrieb 2 nie überstieg.
4.4.3 Kotkultur (nach Roberts und O'SuIlivan) und Auswanderverfahren (nach Baermann-Wetzel)
In den Kotkulturen konnten folgende Larven gefunden werden (in Klammer wird der
betriebsübergreifende prozentuale Anteil angeführt): Teladorsagia (11,8 %), Hae-
monchus (33,2 %), Cooperia (16,7 %), Chabertia (14,0 %), Trichostrongylus (9,5
%), Strongyloides (3,3 %) und - jedoch ausschließlich in Betrieb 3 bei den Jährlin-
gen im Herbst - kleine Lungenwurmlarven, M. capillaris, gefunden werden.

76
4.4.3.1 Betriebsabhängige Unterschiede der Larvenprävalenz
• Befallsrate im Betriebsvergleich
Der Anteil von Teladorsagia-positiven Kotproben war in Betrieb 1 geringer als in
den beiden anderen Betrieben. Haemonchus-Larven konnten hingegen häufiger in
Betrieb 2 gefunden werden. Betrieb 3 zeigte im Hinblick auf Larven von Cooperia
und Strongyloides eine deutlich geringere Prävalenz als die beiden anderen Betriebe,
während Trichostrongylus-Larvcn hier häufiger als in Betrieb 1 und 2 nachgewiesen
werden konnten. Chabertia-Larvcn waren mit 100 % an positiven Proben am häu-
figsten in Betrieb 2, mit 40 % an positiven Proben in Betrieb 1 am seltensten nach-
weisbar (Tab. 18).
Tabelle 18: Anteil positiver Proben im Betriebsvergleich (Angabe in %; n=Anzahl der untersuchten Proben; Tela.—Teladorsagia; Haemo.—Haemonchus; Coop.=Coopena; Chab.-Chabertia; Tricho.=Trichostrongylus; Strongy.-Strongyloides)
Betrieb Tela. Haemo. Coop. Chab. Tricho. Strongy.
l(n=10) 60,0 40,0 40,0 40,0 50,0 50,0
2 (n=9) 88,9 77,8 44,4 100,0 55,6 55,6
3 (n=7) 85,7 42,9 14,3 85,7 71,4 28,6
• Befallsintensität im betrieblichen Vergleich
Die prozentuale Häufigkeit der Larven zeigte zum Teil große betriebliche Unter-
schiede. So erreichte der Anteil an Teladorsagia-Larven in Betrieb 1 einen deutlich
geringeren Maximalwert, während der Maximalwert in Betrieb 2 und 3 ähnlich hoch
war. Für Haemonchus lagen die Maximalwerte aller Betriebe in einem ähnlich hohen
Bereich. Cooperia-Larven erreichten in Betrieb 1 Anteile von bis zu 95 %, während
die Maximalwerte in Betrieb 2 und 3 deutlich geringer waren. Chabertia-Larven
konnten in besonders hohen Anteilen in Betrieb 2 beobachtet werden, während der
Maximalwert in Betrieb 3 im Mittelfeld lag und in Betrieb 1 die geringsten Anteile
zu finden waren. Die Larven von Trichostrongylus erreichten die höchsten Anteile in
Betrieb 3, die geringsten in Betrieb 1. Die Anteile für Strongyloides verhielten sich
für alle Betriebe niedrig. M. capillaris konnten ausschließlich in Betrieb 3 in der
Gruppe der Jährlinge im Herbst, hier jedoch in immenser Anzahl, nachgewiesen
werden (siehe Tab. 19).

77
Tabelle 19: Mittelwert der Larvenanteile mit Minimal- und Maximalwert im Betriebsvergleich (Angabe des Mittelwerts (=Mittel) Minimal- (=Min) und Maximalwerts (=Max) in %; n=Anzahl der untersuchten Proben; Tela.=Teladorsagia; Haemo.- Haemonchus; Coop.-Cooperia; Chab.=Chabertia; Tricho.-Trichostrongylus; Strongy. -Strongy-loides)
Betrieb Wert Tela. Haemo. Coop. Chat. Tricho. Strongy.
l(n=IO) Mittel 2,6 32,7 35,5 2,4 2,2 4,7
Min 0,0 0,0 0,0 0,0 0,0 0,0
Max 11,0 97,0 95,0 20,0 8,0 23,0
2 (n-9) Mittel 17,0 46,0 7,1 22,1 4,4 3,3
Min 0,0 0,0 0,0 3,0 0,0 0,0
Max 50,0 96,0 27,3 70,5 25,0 24,0
3 (n=7) Mittel 18,3 17,6 2,1 20,0 26,6 1,1 Min 0,0 0,0 0,0 0,0 0,0 0,0
Max 41,0 90,0 15,0 48,0 74,0 7,0
• Statistische Auswertung
Eine statistisch signifikante Korrelation zwischen Larvenprävalenz und Haltungs-
form ergab sich für Teladorsagia (r=0,502 mit p<0,01) und Chabertia (r=0,530 mit
p<0,01). Diese betriebsabhängigen Unterschiede im prozentuellen Anteil der Lar-
venprävalenz zeigten sich bei Teladorsagia mit p=0,033 und Chabertia mit p=0,004.
Der Anteil von Teladorsagia-Larwen war in Betrieb 1 signifikant geringer als in den
beiden anderen Betrieben, wobei er in Betrieb 3 am höchsten war. Ähnlich verhielten
sich die Chabertia-Larvenanteile, die ebenfalls für Betrieb 1 am geringsten ausfielen,
während sie in Betrieb 2 den höchsten Wert erreichten. Es ergaben sich weiters Zu-
sammenhänge hinsichtlich des korrelierenden Auftretens von Parasitenlarven. So
korrelierte die Larvenprävalenz von Teladorsagia, Chabertia und Trichostrongylus,
außerdem von Haemonchus, Cooperia und Strongyloides (siehe Tab. 20).
Tabelle 20: Korrelation der Larvenprävalenz betriebsübergreifend (Angabe als Korrelationskoeffizient r; *=p<0,05; **=p<0,01; Tela.=Teladorsagia; Haemo.= Hae- monchus; Coop.=Cooperia; Chab.=Chabertia; Tricho.=Trichostrongylus; Strongy.= Strongyloides)
Tela. Haemo. Coop. Chab. Tricho. Strongy.
Tela. 0,642** 0,461*
Haemo. -0,547** 0,637**
Coop. -0,547** -0,455*
Chat. 0,642**
Tricho. 0,461*
Strongy. 0,637** -0,455*
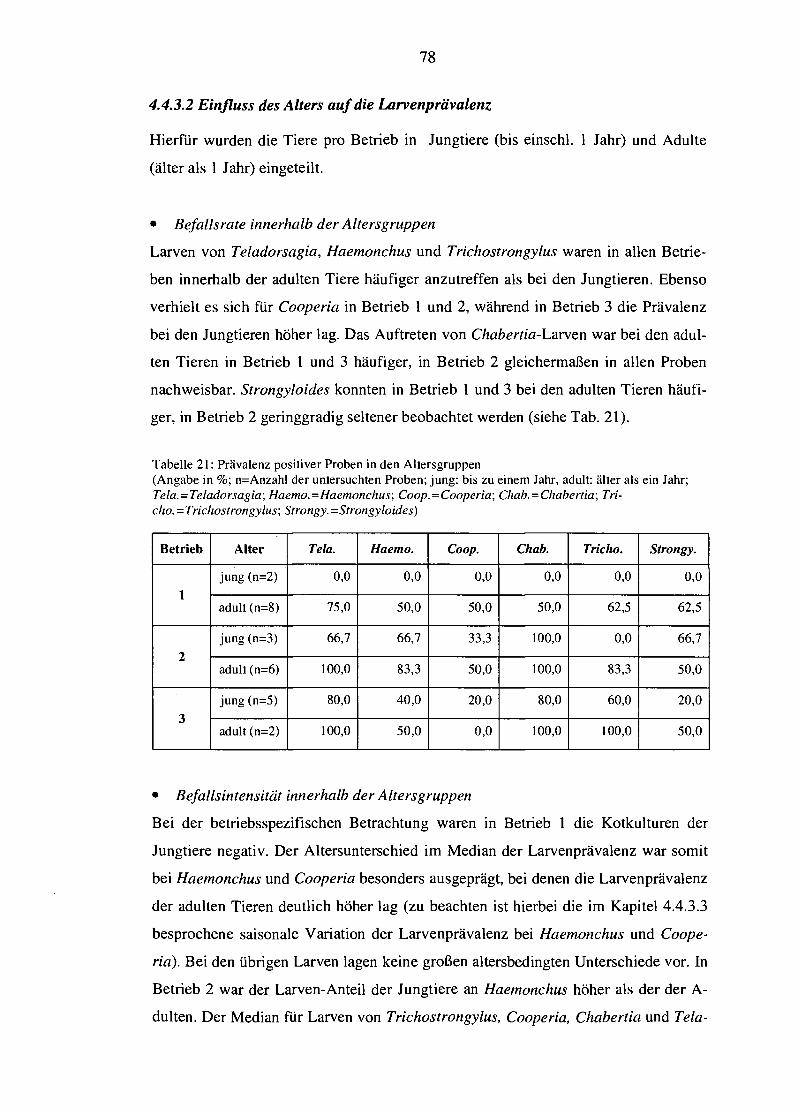
78
4.4.3.2 Einfluss des Alters auf die Larvenprävalenz
Hierfür wurden die Tiere pro Betrieb in Jungtiere (bis einschl. 1 Jahr) und Adulte
(älter als 1 Jahr) eingeteilt.
• Befallsrate innerhalb der Altersgruppen
Larven von Teladorsagia, Haemonchus und Trichostrongylus waren in allen Betrie-
ben innerhalb der adulten Tiere häufiger anzutreffen als bei den Jungtieren. Ebenso
verhielt es sich für Cooperia in Betrieb 1 und 2, während in Betrieb 3 die Prävalenz
bei den Jungtieren höher lag. Das Auftreten von Chabertia-har\en war bei den adul-
ten Tieren in Betrieb l und 3 häufiger, in Betrieb 2 gleichermaßen in allen Proben
nachweisbar. Strongyloides konnten in Betrieb 1 und 3 bei den adulten Tieren häufi-
ger, in Betrieb 2 geringgradig seltener beobachtet werden (siehe Tab. 21).
Tabelle 21: Prävalenz positiver Proben in den Altersgruppen (Angabe in %; n=Anzahl der untersuchten Proben; jung: bis zu einem Jahr, adult: älter als ein Jahr; Tela.—Teladorsagia; Haemo.= Haemonchus; Coop.=Coopena; Chab.=Chabertia; Tri- cho.-Trichostrongylus; Strongy. =Strongyloides)
Betrieb Alter Tela. Haemo. Coop. Chab. Tricho. Strongy.
1 jung (n=2) 0,0 0,0 0,0 0,0 0,0 0,0
adult (n=8) 75,0 50,0 50,0 50,0 62,5 62,5
2 jung (n=3) 66,7 66,7 33,3 100,0 0,0 66,7
adult (n=6) 100,0 83,3 50,0 100,0 83,3 50,0
3 jung (n=5) 80,0 40,0 20,0 80,0 60,0 20,0
adult (n=2) 100,0 50,0 0,0 100,0 100,0 50,0
• Befallsintensität innerhalb der Altersgruppen
Bei der betriebsspezifischen Betrachtung waren in Betrieb 1 die Kotkulturen der
Jungtiere negativ. Der Altersunterschied im Median der Larvenprävalenz war somit
bei Haemonchus und Cooperia besonders ausgeprägt, bei denen die Larvenprävalenz
der adulten Tieren deutlich höher lag (zu beachten ist hierbei die im Kapitel 4.4.3.3
besprochene saisonale Variation der Larvenprävalenz bei Haemonchus und Coope-
ria). Bei den übrigen Larven lagen keine großen altersbedingten Unterschiede vor. In
Betrieb 2 war der Larven-Anteil der Jungtiere an Haemonchus höher als der der A-
dulten. Der Median für Larven von Trichostrongylus, Cooperia, Chabertia und Tela-
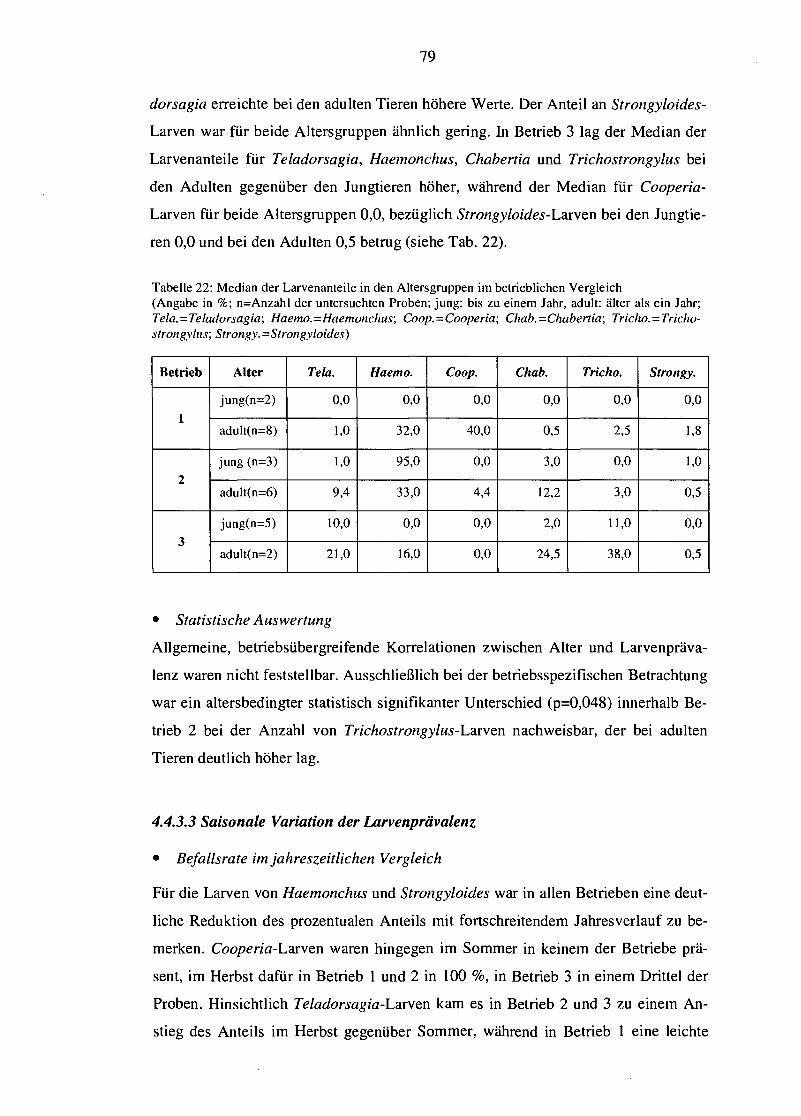
79
dorsagia erreichte bei den adulten Tieren höhere Werte. Der Anteil an Strongyloides-
Larven war für beide Altersgruppen ähnlich gering. In Betrieb 3 lag der Median der
Larvenanteile für Teladorsagia, Haemonchus, Chabertia und Trichostrongylus bei
den Adulten gegenüber den Jungtieren höher, während der Median für Cooperia-
Larven für beide Altersgruppen 0,0, bezüglich Strongyloides-LdxwQn bei den Jungtie-
ren 0,0 und bei den Adulten 0,5 betrug (siehe Tab. 22).
Tabelle 22: Median der Larvenanteile in den Altersgruppen im betrieblichen Vergleich (Angabe in %; n=Anzahl der untersuchten Proben; jung: bis zu einem Jahr, adult: älter als ein Jahr; Tela.=Teladorsagia; Haemo.=Haemonchus; Coop.=Cooperia; Chab.=Chabertia\ Tricho.=Triclio- strongylus; Strongy. =Strongyloides)
Betrieb Alter Tela. Haemo. Coop. Chab. Tricho. Strongy.
1 jung(n=2) 0,0 0,0 0,0 0,0 0,0 0,0
adult(n=8) 1,0 32,0 40,0 0,5 2,5 1,8
2 jung(n=3) 1,0 95,0 0,0 3,0 0,0 1,0
adult(n=6) 9,4 33,0 4,4 12,2 3,0 0,5
3 jung(n=5) 10,0 0,0 0,0 2,0 11,0 0,0
adult(n=2) 21,0 16,0 0,0 24,5 38,0 0,5
• Statistische Auswertung
Allgemeine, betriebsübergreifende Korrelationen zwischen Alter und Larvenpräva-
lenz waren nicht feststellbar. Ausschließlich bei der betriebsspezifischen Betrachtung
war ein altersbedingter statistisch signifikanter Unterschied (p=0,048) innerhalb Be-
trieb 2 bei der Anzahl von Trichostrongylus-havven nachweisbar, der bei adulten
Tieren deutlich höher lag.
4.4.3.3 Saisonale Variation der Larvenprävalenz
• Befallsrate im jahreszeitlichen Vergleich
Für die Larven von Haemonchus und Strongyloides war in allen Betrieben eine deut-
liche Reduktion des prozentualen Anteils mit fortschreitendem Jahresverlauf zu be-
merken. Cooperia-harven waren hingegen im Sommer in keinem der Betriebe prä-
sent, im Herbst dafür in Betrieb 1 und 2 in 100 %, in Betrieb 3 in einem Drittel der
Proben. Hinsichtlich Teladorsagia-Larwen kam es in Betrieb 2 und 3 zu einem An-
stieg des Anteils im Herbst gegenüber Sommer, während in Betrieb 1 eine leichte

80
Reduktion der Prävalenz zu beobachten war. Der Anteil an Chabertia-Larwtn verrin-
gerte sich von Sommer auf Herbst in Betrieb 1, blieb in Betrieb 2 gleich und stieg in
Betrieb 3 an. Trichostrongylus-Larven waren in Betrieb 2 zu jeweils annähernd glei-
chen, in Betrieb 1 zu gleichen Teilen nachweisbar, während in Betrieb 3 eine Ver-
dopplung der Prävalenz von Sommer auf Herbst zu verzeichnen war (Tab. 23).
Tabelle 23: Larven-Prävalenzen im jahreszeitlichen Vergleich (Angabe in %; n=Anzahrder untersuchten Proben; Tela.-Tela-dorsagia; Haemo.-Haemonchus; Coop.=Cooperia\ Chab.=Chabertia\ Tricho.-Trichostrongylus; Strongy.=Strongyloides)
Betrieb Monat Tela. Haemo. Coop. Chat. Tricho. Strongy.
1 Sommer (n=6) 66,7 66,7 0,0 50,0 50,0 66,7
Herbst (n=4) 50,0 0,0 100,0 25,0 50,0 25,0
2 Sommer (n=5) 80,0 100,0 0,0 100,0 60,0 100,0
Herbst (n=4) 100,0 50,0 100,0 100,0 50,0 0,0
3 Sommer (n=4) 75,0 75,0 0,0 75,0 50,0 50,0
Herbst (n=3) 100,0 0,0 33,3 100,0 100,0 0,0
• Befallsintensität im jahreszeitlichen Vergleich
Der Median-Wert von Teladorsagia-Larven stieg in allen Betrieben von Sommer auf
Herbst an. Ähnlich verhielten sich die Anteile für Cooperia-Larven in Betrieb 1 und
2, während der Median in Betrieb 3 jeweils 0,0 betrug. Bei Haemonchus-Larven war
hingegen in allen Betrieben eine mehr oder weniger deutliche Reduktion des Anteils
im Herbst gegenüber Sommer bemerkbar. In Betrieb 2 und 3 war ein Zuwachs des
Anteils an Chabertia-Larvcn zu beobachten, während in Betrieb 1 ein leichtes Ab-
sinken nachweisbar war. Die Medianwerte für Trichostrongylus waren allgemein
sehr gering und erreichten nur in Betrieb 3 im Herbst höhere Anteile. Ähnlich nied-
rig, zum Großteil mit einem Medianwert von 0,0, stellten sich auch die Werte für
Strongyloides-harven dar (siehe Tab. 24).
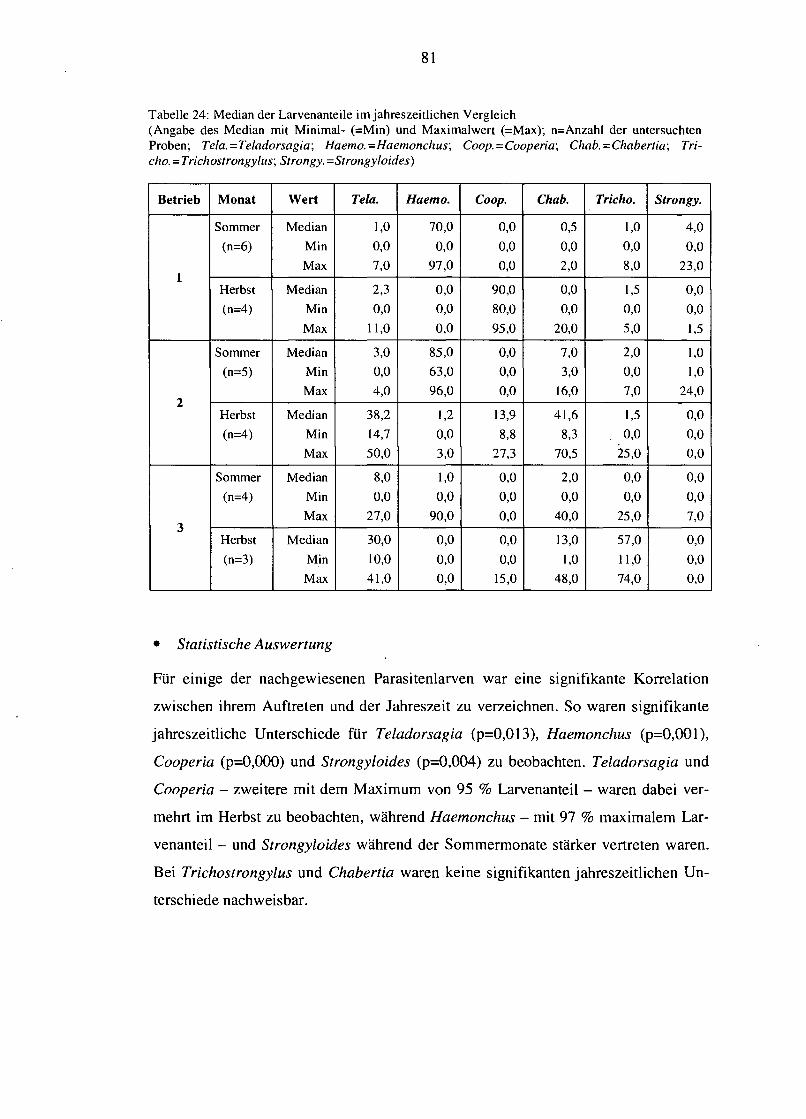
81
Tabelle 24: Median der Larvenanteile im jahreszeitlichen Vergleich (Angabe des Median mit Minimal- (=Min) und Maximalwert (=Max); n=Anzahl der untersuchten Proben; Tela.=Teladorsagia; Haemo.-Haemonchus; Coop.=Cooperia; Chab. = Chabertia; Tri- cho.-Trichostrongylus; Strongy.-Strongyloides)
Betrieb Monat Wert Tela. Haento. Coop. Chab. Tricho. Strongy.
Sommer Median 1,0 70,0 0,0 0,5 1,0 4,0
(n=6) Min 0,0 0,0 0,0 0,0 0,0 0,0
1 Max 7,0 97,0 0,0 2,0 8,0 23,0
Herbst Median 2,3 0,0 90,0 0,0 1,5 0,0
(n=4) Min 0,0 0,0 80,0 0,0 0,0 0,0
Max 11,0 0,0 95,0 20,0 5,0 1,5
Sommer Median 3,0 85,0 0,0 7,0 2,0 1,0
(n=5) Min 0,0 63,0 0,0 3,0 0,0 1,0
2 Max 4,0 96,0 0,0 16,0 7,0 24,0
Herbst Median 38,2 1,2 13,9 41,6 1,5 0,0
(n=4) Min 14,7 0,0 8,8 8,3 . 0,0 0,0
Max 50,0 3,0 27,3 70,5 25,0 0,0
Sommer Median 8,0 1,0 0,0 2,0 0,0 0,0
(n-4) Min 0,0 0,0 0,0 0,0 0,0 0,0
3 Max 27,0 90,0 0,0 40,0 25,0 7,0
Herbst Median 30,0 0,0 0,0 13,0 57,0 0,0
(n=3) Min 10,0 0,0 0,0 1,0 11,0 0,0
Max 41,0 0,0 15,0 48,0 74,0 0,0
• Statistische Auswertung
Für einige der nachgewiesenen Parasitenlarven war eine signifikante Korrelation
zwischen ihrem Auftreten und der Jahreszeit zu verzeichnen. So waren signifikante
jahreszeitliche Unterschiede für Teladorsagia (p=0,013), Haemonchus (p=0,001),
Cooperia (p=0,000) und Strongyloides (p=0,004) zu beobachten. Teladorsagia und
Cooperia - zweitere mit dem Maximum von 95 % Larvenanteil - waren dabei ver-
mehrt im Herbst zu beobachten, während Haemonchus - mit 97 % maximalem Lar-
venanteil - und Strongyloides während der Sommermonate stärker vertreten waren.
Bei Trichostrongylus und Chabertia waren keine signifikanten jahreszeitlichen Un-
terschiede nachweisbar.

82
5. DISKUSSION
Ziel dieser Arbeit war es, das Ausmaß der Endoparasitenbelastung anhand der Ei-
bzw. Oozystenausscheidungsrate in drei biologisch bewirtschafteten Milchziegenbe-
trieben mit jeweils unterschiedlicher Haltungsform im Sommer sowie im Spätherbst
zu erheben. Die jeweiligen Ergebnisse wurden zu Haltungssystem, BCS, Tieralter
und Jahreszeit in Beziehung gesetzt, um etwaig vorhandene Korrelationen aufzuzei-
gen und anhand dieser Resultate Erkenntnisse zur möglichen Optimierung des be-
triebsüblichen Parasitenmanagements zu gewinnen.
Bei der klinischen Untersuchung waren in allen Betrieben in gleichem Maße verein-
zelt Tiere mit pathologischem Kotbefund und/oder respiratorischen Symptomen auf-
fällig. Die parasitologischen Untersuchungen zeigten überwiegend ein ähnliches Pa-
rasitenspektrum für alle drei Betriebe, einzig Eier von Moniezia und Nematodirus
konnten nicht überall nachgewiesen werden. Große Unterschiede lagen jedoch hin-
sichtlich der Befallsintensität vor. Hier zeigte Betrieb 3 (ganzjährige Weidehaltung)
bezüglich weideassoziierten bzw. haltungsunabhängigen Parasiten deutlich höhere
Ausscheidungsraten als Betriebe 1 (Auslauf und Heufütterung) und 2 Auslauf und
Grünfütterung), die sich weniger deutlich voneinander unterschieden, wobei jedoch
tendenziell Betrieb 1 geringere Befallsintensitäten aufwies. Es lag demnach eine sta-
tistisch signifikante Korrelation der Befallsintensität aller Parasiten - mit Ausnahme
von Moniezia - mit der Haltungsform vor. Auffallend war jedoch, dass sich eine
deutlich höhere Befallsintensität, wie sie generell in Betrieb 3 und bei Einzeltieren in
Betrieb 1 und 2 vorzufinden war, nicht in eindeutig geringerem BCS widerspiegelte.
So war der Anteil der Tiere mit BCS im Idealbereich in Betrieb 3 sogar höher als in
Betrieb 1 und 2. Demzufolge konnte keine signifikante Korrelation zwischen BCS
und der Ausscheidungsrate nachgewiesen werden. Signifikante Zusammenhänge
zeigten sich hingegen zwischen dem Tieralter und der Ausscheidungsrate einiger
Parasiten. Dies traf besonders auf die Ausscheidungsraten von Eimerien zu, die bei
Jungtieren maßgeblich höher waren. Saisonale Variationen der Ausscheidungsraten
konnten innerhalb der einzelnen Betriebe durchaus beobachtet werden, wobei die
weideassoziierten Parasiten im Herbst tendenziell seltener nachweisbar waren, wäh-
rend die stallassoziierten Parasiten zum Teil höhere Ausscheidungsraten zeigten. Bei
der betriebsübergreifenden Betrachtung konnte allerdings kein allgemeingültiger
Zusammenhang zwischen Ausscheidungsraten und Jahreszeit gefunden werden.

83
5.1 Methodenkritik
Um die Ergebnisse dieser Arbeit objektivieren zu können, wäre es notwendig, die
empirische Basis zu verbreitem, d.h. noch mehr Betriebe in die Untersuchungen auf-
zunehmen, da sich betriebsabhängige und individuelle tierbedingte Unterschiede bei
geringer Betriebsanzahl stark auf die Befunde auswirken. Außerdem birgt die zum
Teil kleine Stichprobenanzahl die Gefahr, verzerrte Ergebnisse zu liefern, was insbe-
sondere bei der statistischen Auswertung zu berücksichtigen ist. Die hier gewonne-
nen Ergebnisse erheben demzufolge keinen Anspruch auf Allgemeingültigkeit.
Der Vergleich der Untersuchungsergebnisse aus Flotation und Eizählung hinsichtlich
der Parasiten-Prävalenz und Befallsintensität zeigte durchaus vergleichbare Resulta-
te, insbesondere für MDS und Eimerien. Allfällige Unterschiede können nicht zuletzt
auf eine sehr kleine und dementsprechend nicht repräsentative Stichprobenanzahl bei
den Einzahlungen zurückgeführt werden.
Eizählungen jeden Einzeltieres wären sehr aufschlussreich, insbesondere bei starker
Befallsintensität wie z.B. in Betrieb 3, um die Ergebnisse differenzierter wiedergeben
zu können, da die hier erfolgte Graduierung in ggr. bis hgr. Unterschiede nur be-
grenzt wiedergeben kann. Dies könnte auch die Ursache für zum Teil statistisch nicht
nachweisbare Korrelationen und signifikante Unterschiede sein. In den meisten Fäl-
len war es nicht möglich, ausreichend Kotmaterial von den Einzeltieren zu gewinnen,
um zusätzlich zum Anreicherungsverfahren für jedes Tier Eizählungen durchführen
zu können, so dass die Eizählungen an Sammelkotproben aus den Haltungsgruppen
unternommen wurden.
Zusätzlich wäre die Durchführung von Sektionen aufschlussreich, um einen umfas-
senden Einblick in das Ausmaß der Parasitenbürde und das vorhandene Endoparasi-
tenspektrum der Tiere und dessen exaktes Ausmaß gewinnen zu können. Außerdem
erlauben Sektionen die weitere Differenzierung der Parasiten anhand ihrer adulten
Stadien und ermöglichen zusätzlich die Besichtigung aller Organe, was etwa im Hin-
blick auf einen Leberegel- oder Lungenwurmbefall in den hier untersuchten Betrie-
ben von großem Interesse wäre. So konnten nur diejenigen Parasiten nachgewiesen
werden, für deren Detektion die Methodik der Koproskopie geeignet ist, wodurch ein
Befall mit Parasiten, die nicht über ihre Ei- bzw. Oozystenausscheidung diagnosti-
zierbar sind, nicht ausgeschlossen werden kann. Das könnte eine mögliche Ursache
dafür sein, dass bei keinem der Betriebe Trematoden, nur vereinzelt Cestoden und als
Parasiten der Atemwege ausschließlich M. capillaris (und auch diese nur in Betrieb

84
3) nachgewiesen werden konnten. Interessant bezüglich des Parasitenspektrums wäre
zusätzlich die genaue Differenzierung der vorliegenden Eimerien-Oozysten anhand
sporulierter Stadien .
In weiterer Folge wäre die Untersuchung der Milchleistungsdaten der Einzeltiere im
Hinblick auf etwaige Korrelationen zu Endoparasitosen äußerst aufschlussreich, da
das Laktationsstadium nachweislich einen bedeutsamen Faktor hinsichtlich der Emp-
fänglichkeit gegenüber Parasiteninfektionen und der Eiausscheidungsrate darstellt
(siehe Kapitel 5.6). Sehr interessant wäre zudem der Vergleich von biologischen mit
konventionellen Betrieben mit unterschiedlichen Haltungsformen.
5.2 Haltungs- und Fütterungspraxis der Betriebe
• Betrieb 1
In Betrieb 1 beruht das Endoparasitenmanagement in erster Linie auf der Haltungs-
form, d.h. dem - nach wiederholter Endoparasitenproblematik - bewussten Umstieg
von Weide- auf Auslaufhaltung. Die ganzjährige Heufütterung stellt eine zusätzliche
Reduktion des Infektionsrisikos mit Parasiten dar. Aus rein parasitologischer Sicht ist
diese Haltungsform demnach als effiziente Methode zur Parasitenkontrolle zu sehen,
zeigte doch der Großteil der Ziegen geringe Eiausscheidungsraten. Sie muss jedoch
mit den im Jahr 2010 in Kraft tretenden Bestimmungen zur Regelung des Auslaufs
bzw. Weidegangs in biologischer Viehwirtschaft in Einklang gebracht werden und
im Hinblick darauf kritisch hinterfragt werden.
Die leistungsorientierte Fütterung und die Anpassung der Ration an die Körperkondi-
tion sind im Hinblick auf die Parasitenprophylaxe als durchaus förderlich zu beurtei-
len. Von großer Bedeutung sind dabei die exakten Futtermittelanalysen des Nähr-
stoffgehaltes, die der Diplomarbeit von Robert Wolf (in Veröffentlichung) entnom-
men werden können. Interessant wäre weiters die Fragestellung, ob die angewandte
Technik der Mistaufbereitung ausreichend ist, um den Großteil der im Stallmist ent-
haltenen infektiöse Parasitenstadien unschädlich zu machen und um somit eine Kon-
tamination der Futterwiesen zu vermeiden. Zusätzlich stellt die Heuaufbereitung in
Form der Trocknung in jedem Fall eine potentielle Reduktion der Infektionsgefahr
über das Futter dar.

85
• Betrieb 2
Auch hier basiert ein Teil des Parasitenmanagements auf der Auslaufhaltung, wobei
die Ausläufe hier zum Teil bewachsen sind. Im Sommer wird Grünfutter zugefüttert,
was ein höheres Risiko für eine mögliche Parasiteninfektion mit sich bringt. Die im
Vergleich zu Betrieb 1 höhere Parasitenbelastung könnte nicht zuletzt Folge dieses
Umstands sein.
Die erfolgte Umgruppierung der Tiere nach Laktationsstadium ist hinsichtlich einer
somit eher gewährleisteten Deckung des Nährstoffbedarfs durch leistungsorientierte
Fütterung als positiv zu bewerten. Durchaus aufschlussreich wäre die Untersuchung
auf sich daraus langfristig ergebende Veränderungen hinsichtlich der Eiausschei-
dungsraten. Erfolg versprechend zur Coccidien-KontroUe ist außerdem die Umstal-
lung der Kitze in eine neue Halle, die jedoch erst vor kurzer Zeit erfolgte, weshalb
noch keine Erfahrungsberichte des Landwirtes im Hinblick auf eine etwaige Verbes-
serung der Situation vorliegen. Gleichzeitig sollte versucht werden, die Oozyste-
nausscheidungsrate der Muttertiere und damit den Infektionsdruck für die Kitze zu
senken, um damit die Kontamination der neuen Stallungen durch die umgestallten
Kitze so gering wie möglich zu halten. Begleitende regelmäßige Reinigungsmaß-
nahmen wären zusätzlich vorteilhaft.
• Betrieb 3
In der Weidehaltung ist das Risiko einer Konfrontation mit Endoparasiten naturge-
mäß gegeben. Auch die im Zuge dieser Arbeit untersuchten Tiere von Betrieb 3 zeig-
ten Endoparasiteninfektionen unterschiedlichen Ausmaßes. Umso bedeutsamer ist es,
in ein nachhaltiges Endoparasitenmanagement bei Weidehaltung den Aspekt Wei-
demanagement einzubinden. Demzufolge ist das im Betrieb 3 praktizierte Modell des
Weidewechsels als durchaus wichtig und positiv zu betrachten. Über die Effizienz
entscheidet jedoch das Beweidungsintervall der Portionsweiden, das in jedem Fall an
örtliche Gegebenheiten angepasst werden muss und vom Landwirt nicht exakt ange-
geben werden konnte. Zu bedenken ist hierbei die bereits erwähnte Problematik der
Wechselbeweidung in gemäßigtem Klima aufgrund der erforderlichen langen Ruhe-
pausen zwischen der Beweidung. Der Grad der Inaktivierung von Parasitenstadien
durch die Lagerung des Stallmistes müsste eingehend untersucht werden. In Anbet-
racht einer optimierten Deckung des Nährstoffbedarfes der Ziegen könnte die, wie in
Betrieb 1 und 2 praktizierte, Gruppierung nach Laktationsstadium in Betracht gezo-

86
gen werden, nachdem eine individuelle Anpassung der Ration an die Produktions-
leistung derzeit nur schwer, eventuell mit Transponderfütterung, durchführbar ist und
im Moment alle Ziegen die selbe Ration erhalten. Eine diesbezügliche Optimierung
könnte sich durchaus positiv auf die Widerstandsfähigkeit der Ziegen gegenüber En-
doparasitosen auswirken.
5.3 Betriebliche Endoparasitenkontrolle und mögliche Optimierung
• Betrieb 1
Bis auf einzelne Ausnahmen, vor allem bei Coccidien, ist der Endoparasitenbefall als
geringgradig einzustufen, wobei jedoch synthetische Anthelminthika eine Nebenrolle
spielen und nur restriktiv eingesetzt werden. Grundsätzlich entspricht die Minimie-
rung der Anthelminthika-Behandlungen den Rahmenbedingungen der biologischen
Viehwirtschaft und ist durchaus zu befürworten, solange keine massiven, aus Grün-
den des Tierschutzes bedenklichen Parasitosen vorliegen.
Der im Betrieb übliche Behandlungsmodus kann jedoch trotz der geringen Behand-
lungshäufigkeit mit Gefahren vor allem bezüglich einer Anthelminthika-
Resistenzbildung assoziiert sein. Im Hinblick auf eine potentielle Resistenzentwick-
lung sollte zum einen der eingesetzte Wirkstoff des Anthelminthikums regelmäßig
gewechselt werden, zum anderen erfolgt die Dosierung nach geschätztem Körperge-
wicht, wodurch das Risiko einer Unterdosierung besteht. Nachdem es sich um Ein-
zeltierentwurmungen handelt, wäre das genaue Abwiegen und Dosieren nach tat-
sächlichem Körpergewicht mit überschaubarem Aufwand verbunden und aus Grün-
den der Anwendungssicherheit zu empfehlen. Zusätzlich wird anstelle der - in der
Literatur empfohlenen - doppelten die 1,5-fache Dosis für Schafe angewandt, so dass
die Gefahr der Unterdosierung wächst. Außerdem erfolgt der Anthelminthika-Einsatz
bei Zuchtböcken ohne exakte Diagnostik des tatsächlichen Parasitenbefalls mittels
Breitspektrum-Anthelminthika, was ebenso zur Resistenzentwicklung beitragen
kann. Eine weitere Parasitengefahr birgt der Zukauf dieser Zuchtböcke, zumal die
Tiere nicht in Quarantäne gehalten werden und weder der Parasiten- noch der
Anthelminthika-Resistenzstatus der neuen Tiere bekannt sind. So kann es leicht zur
Einschleppung resistenter Parasitenstämme in die bereits bestehende Herde kommen.
Insofern wäre eine separate Haltung der neuen Tiere mit begleitender parasitologi-

87
scher Untersuchung und - wenn nötig - gezielter antiparasitärer Therapie mittels
Antiparasitika mit engem Wirkspektrum die optimale Lösung.
Der gänzliche Verzicht auf synthetische Antiparasitika in Bezug auf die Coccidien-
Bekämpfung schlägt sich insbesondere im Sommer in zum Teil erhöhten Oozyste-
nausscheidungsmengen (opg von 6400) bei den Kitzen nieder. Diesbezüglich sollten
eventuell zusätzliche unterstützende Maßnahmen wie z.B.: Desinfektion, häufigere
Entmistung oder auch der gezielte Einsatz synthetischer Kokzidiostatika bei klinisch
apparenten Infektionen in Betracht gezogen werden, da die Kokzidiose aufgrund
geringerer Gewichtszunahmen zu massiven Produktionsverlusten und auch zu To-
desfällen führen kann und abgesehen vom ökonomischen auch einen Tierschutzas-
pekt darstellt.
• Betrieb 2
Die Eiausscheidungsraten liegen - allgemein betrachtet - über denen von Betrieb 1,
sie sind jedoch - obwohl hier regelmäßig aufgrund der sonst anfallenden Wartezeiten
in der Trockenstehperiode entwurmt wird - als mittelgradig einzustufen. Als Alterna-
tive zur regelmäßigen prophylaktischen Entwurmung aller Tiere könnte als Optimie-
rungsmaßnahme über eine eventuelle Umstellung der - nicht ganz korrekt „biologi-
schen" - prophylaktischen Entwurmungspraxis auf das bereits eingehend behandelte
selektive Behandlungsregime nachgedacht werden, da innerhalb einer Herde nicht
alle Tiere im gleichen Ausmaß infiziert sind. Zusätzlich sind die Entwurmungen
nach derzeitigem Schema zeitlich über mehrere Monate gestreut, da sie immer nur in
der Trockenstehperiode erfolgen und könnten dadurch an Effizienz einbüßen. Der
ineffiziente Einsatz von Anthelminthika ist hingegen sehr teuer, da er meistens wie-
derholt werden muss oder erhoffte Erfolge ausbleiben und es trotzdem zu Produkti-
onsverlusten kommt. Eine selektive Behandlung auffälliger Tiere und das zusätzliche
Fasten vor der geplanten Durchführung könnten somit gleichzeitig die Effizienz der
Entwurmungsmaßnahmen erhöhen und Kosten sparen, obwohl bei Entwurmungen
während der Laktation Wartezeiten berücksichtigt werden müssen. Dabei wäre hier
die Behandlung von Einzeltieren eher geeignet als die Entwurmung ganzer Risiko-
gruppen, da die Tiere inzwischen nach Laktationsstadium (was sich mitunter mit den
Risikogruppen deckt) gruppiert werden. Die Entwurmung einer ganzen Haltungs-
gruppe könnte demnach die Resistenzentwicklung fördern, nachdem alle Parasiten
dieser Gruppe dem Selektionsdruck des Anthelminthikums ausgesetzt sind.

88
Positiv anzumerken ist hingegen der Wirkstoffwechsel (von Fenbendazol auf Moxi-
dectin), der weiterhin regelmäßig durchgeführt werden sollte. Der Einsatz von Breit-
bandanthelminthika ist allerdings auch hier kritisch zu hinterfragen und sollte im
Idealfall durch eine - an die diagnostizierte Parasitose angepasste - Therapie mit
spezifisch wirksamen Stoffgruppen ersetzt werden, zumal besonders die Gruppe der
Benzimidazole, wozu Fenbendazol (Panacur®) zählt, als höchst gefährdet bezüglich
Resistenzentwicklung gilt. Wie auch bereits für Betrieb 1 empfohlen, sollte die ein-
gesetzte Dosierung das 2-fache der empfohlenen Schafdosis betragen und außerdem
nach genauem Körpergewicht oder nach dem schwersten Tier der Herde erfolgen,
um eine Unterdosierung zu vermeiden. Nachdem in diesem Betrieb ebenfalls Zucht-
böcke zugekauft werden, wären auch hier die bereits für Betrieb 1 angeführten Qua-
rantänemaßnahmen vor Integration des zugekauften Tieres in die bestehende Herde
aus denselben Gründen ratsam.
Zu hoffen ist, dass mit Umstallung der Kitze in eine neu errichtete Halle die Not-
wendigkeit der prophylaktischen Kokzidiostatika-Behandlung abnimmt. Vorsorgli-
che Maßnahmen wie z.B. die häufige Entmistung mit regelmäßiger Desinfektion
bzw. die rechtzeitige Diagnostik und (homöopathische) Behandlung bei wechselhaf-
ter Kotkonsistenz könnten ein Wiederaufflammen der allgemeinen Problematik e-
ventuell verhindern.
• Betrieb 3
Erwartungsgemäß waren in Betrieb 3 die Eiausscheidungsraten der primär weideas-
soziierten Parasiten am höchsten und erreichten zum Teil massive Ausmaße, so dass
parasitär-bedingte Produktionsverluste nicht ausgeschlossen werden können, obwohl
die Tiere zum Großteil einen guten Allgemeinzustand zeigten. Die genaue Beobach-
tung der Milchleistung wäre angesichts des zum Teil hohen Befalls hier sehr auf-
schlussreich.
Bisher wird nur bei vorliegendem Parasitosenverdacht und jeweils nur das entspre-
chende Tier entwurmt. Die Effizienz der Entwurmungen könnte hier durch den
Wechsel zu einem strategischen, selektiven Behandlungsmodus erhöht werden. Al-
lerdings bietet sich in diesem Betrieb die Behandlung unter Berücksichtigung von
Risikogruppen z.B. der Hochlaktierenden an, da alle melkenden Tiere gemeinsam
gehalten werden und so trotz Behandlung einer gesamten Risikogruppe ein ausrei-
chend großes Refugium der unbehandelten Tiere zurückbleibt. Ein regelmäßiger

89
Wirkstoffwechsel ist auch hier äußerst ratsam. Positiv hervorzuheben ist die vom
Landwirt praktizierte Verabreichung von Anthelminthika in der doppelten empfohle-
nen Schafdosierung. Eine mögliche Effizienzsteigerung durch eine dem Körperge-
wicht angepasste Dosierung sowie durch Fasten der Tiere vor geplanter Behandlung
wurden bereits erwähnt und werden auch hier empfohlen. Zusätzlich bieten eventuell
einige der in Kapitel 2.6 besprochenen Behandlungsmöglichkeiten in naher Zukunft
eine Alternative bzw. Ergänzung zum Einsatz synthetischer Anthelminthika.
Dringender Handlungsbedarf besteht bezüglich der hohen Sterblichkeitsrate von 40
% der Kitze innerhalb der ersten Lebenstage, da damit nicht zuletzt hohe Produkti-
onsverluste verbunden sind. Eine umfassende Aufklärung der Ursache wäre unbe-
dingt notwendig. Nicht auszuschließen ist ein Zusammenhang mit Toxoplasmen, die
durch junge Katzen übertragen werden und durch Schädigung der Föten für lebens-
schwache Kitze verantwortlich sein können. Da am Hof einige Katzen zur Mausbe-
kämpfung gehalten werden, wäre eine Toxoplasmen-Prävalenz nicht überraschend.
Nachdem ausschließlich junge Katzen als Toxoplasmen-Ausscheider fungieren und
damit die Kontaminationsquelle darstellen, wäre ein denkbarer Lösungsansatz, die
vorhandenen Katzen kastrieren zu lassen, um die Anzahl der Jungtiere zu reduzieren.
Gegen Toxoplasmen als Ursache für die hohe Kitzsterblichkeit spricht jedoch die als
vereinzelt angegebene Abortrate, die vom Besitzer auf zum Teil beobachtete Stoßun-
fälle zurückgeführt wird. Die zum Teil massive Coccidien-Prävalenz ist als Ursache
für die hohe Sterberate der Kitze zwar nicht wahrscheinlich, da die Kitze vor allem
innerhalb der ersten vier Lebenstagen sterben und dies für Coccidien-bedingte Aus-
fälle zu früh erscheint. Trotzdem sollten Maßnahmen zur Eindämmung des Cocci-
dien-Befalls in Betracht gezogen werden, um damit einhergehende etwaige Produk-
tionsverluste zu minimieren.
5.4 BCS und dessen Korrelation mit der Endoparasitenbelastung
Die Erhebung des BCS dient zur Ermittlung der Körperkondition bzw. der Energie-
reserven eines Tieres. Diese Technik wurde zuerst für Schafe entwickelt und später
für die Anwendung bei Ziegen adaptiert (Lenden- und Brustbeinscore für Milchzie-
gen nach MORAND-FEHR et al., 1989), da die Ziege im Unterschied zum Schaf das
Körperfett in lokalen Fettdepots - dazu gehören die Brustbein- sowie die Lendenwir-
belregion - speichert. Berücksichtigt werden muss unbedingt, dass ein Teil des Kör-

90
perfetts bei der Ziege intraabdominal und somit nicht palpierbar liegt (VATTA et al.,
2002), so dass die Palpation das Tier eventuell magerer erscheinen lässt, als es tat-
sächlich ist.
In den im Zuge dieser Arbeit untersuchten Betrieben war zu bemerken, dass die Tie-
re von Betrieb 3, die eine statistisch signifikant höhere Befallsintensität an verschie-
denen Endoparasiten aufweisen, bezüglich ihres BCS im subjektiven Eindruck wie
auch in errechneten Anteilen - mit Ausnahme zweier Tiere beim ersten Betriebsbe-
such, die in sehr schlechtem Ernährungszustand waren - ein ausgesprochen homoge-
nes Bild darboten. Allerdings lag der Anteil der Tiere mit BCS<2,5 im Vergleich mit
den beiden anderen Betrieben höher, während Tiere mit BCS>3,5 nur vereinzelt und
bei Betrieb 2 und 3 häufiger anzutreffen waren. Hingegen wies Betrieb 1 mit der
weitgehend geringsten Parasitenbelastung den kleinsten Prozentsatz an Tieren mit
idealem BCS von 2,5 bis 3,5 auf.
Die Ergebnisse dieser Arbeit zeigen somit keine eindeutige Korrelation zwischen
BCS und Parasitenbelastung und decken sich insofern mit Ergebnissen vorangegan-
gener Untersuchungen (VATTA et al., 2002). Auch nach CABARET et al. (2002) hat
sich die Ermittlung des BCS am Einzeltier als nicht nützlich zur Parasitenkontrolle
im Rahmen eines Managementprogramms erwiesen. In Studien von KOOPMANN et
al. (2006) konnte zusätzlich keine maßgebliche Korrelation zwischen erhobenen
FAMACHA®-Werten (siehe Kapitel 5.9) und Entwicklung der Körperkondition ge-
funden werden (VAN WYK U. BATH, 2002; VAN WYK et al., 2006). Grund dafür, dass
der BCS nicht verlässlich mit der Endoparasitenbelastung korreliert, könnte die mul-
tiple Faktorenabhängigkeit des BCS sein, so dass er selten eindeutig auf einen allei-
nigen Aspekt wie z.B. Endoparasiten zurückgeführt werden kann. So kann der BCS
als Spiegel der Emährungssituation z.B. deutlich geringere Werte in Zeiten geringe-
rer Nährstoffversorgung bzw. reduzierter Nahrungsaufnahme und/oder im Zuge er-
höhten Nährstoffbedarfs wie z.B. während des Wachstums, in der Trächtigkeit, wäh-
rend der Laktation, aber auch bei Endoparasiteninfektionen zeigen (VATTA et al.,
2002). Dazu kommen jedoch noch weitere umweltbedingte Faktoren wie z.B. klima-
tische Bedingungen, Haltungsform (Bewegungsmöglichkeiten), Futterangebot, Rati-
onsgestaltung und Nährstoffgehalt, die einen maßgeblichen Einfluss auf den Ernäh-
rungszustand eines Tieres ausüben. Zusätzlich spielen tierspezifische Aspekte wie
Gesundheits- und Immunstatus (z.B. Krankheiten wie Para- oder Pseudotuberkulo-
se), Laktations- bzw. Reproduktionsstadium (Hochlaktation, Trächtigkeit...), Alter,

91
Fressverhalten (Rangordnung), Vermögen der Futteraufnahme (z.B. Zahnprobleme,
Kaustörungen, Erkrankungen des Bewegungsapparates etc.), Stoffwechsellage,
Stress etc. eine große Rolle.
Bei der Beurteilung des BCS ist jedoch unbedingt der Laktationsstand zu berücksich-
tigen, d.h. ein BCS von 4 in der Trockenstehperiode muss anders beurteilt werden als
derselbe Wert während der Hochlaktation; niedrige BCS-Werte während der Hoch-
laktation sind eher zu erwarten als z.B. in der Trockenstehperiode. Zu erkennen ist,
dass in Betrieb 1 und 2 der Anteil an Tieren mit BCS > 3,5 deutlich höher ist bzw.
Tiere mit BCS < 2,5 in Betrieb 3 häufiger zu beobachten sind. Diese Ergebnisse bes-
tätigen den Eindruck, dass sich ein starker Parasitenbefall nicht immer offensichtlich
in verminderter Körperkondition und schlechterem Allgemeinzustand manifestieren
muss und kann als Bestätigung für die Erkenntnisse von VAN WYK et al. (2006) hin-
zugezogen werden, wonach keine zwingende Beziehung zwischen dem Ausmaß der
Eiausscheidungsrate und einer klinischen Helminthose vorliegt. Daraus ist zu schlie-
ßen, dass hohe Eiausscheidungen keinen Rückschluss auf das Vermögen des
Wirtstieres zulassen, die Helminthose kontrollieren zu können, so dass durchaus be-
obachtet werden kann, dass die Eiausscheidungsraten klinisch apparenter Tiere nied-
riger sein können, als jene klinisch inapparenter Tiere mit guter immunologischer
Widerstandskraft bzw. „Resilience", d.h. der Fähigkeit, die Parasitenbelastung bzw.
deren negative Nebeneffekte kontrollieren bzw. minimieren und trotzdem die Pro-
duktionsleistung aufrechterhalten zu können (HOSTE et al., 2002a; VAN WYK et al.,
2006). Des Weiteren konnten im Vergleich mit den beiden anderen Betrieben in Be-
trieb 3 keinerlei vermehrte klinische Auffälligkeiten verzeichnet werden, die auf ein
massives Endoparasitenproblem schließen ließen, was ebenfalls als Anzeichen einer
guten „Resilience" gesehen werden kann (VAN WYK et al., 2006). Die in Betrieb 3
beobachteten beiden abgemagerten Tiere waren anamnestisch bereits seit einigen
Tagen auffällig (plötzliche Abmagerung, reduzierte Futteraufnahme, fehlende
Milchproduktion, struppiges Haarkleid, vermindertes Allgemeinverhalten), wobei
sich deren Zustand nach Angaben des Landwirtes rapide verschlechterte. In der Kot-
untersuchung zeigte jedoch nur eines der beiden Tiere einen außerordentlich starken
Befall mit MDS. Fraglich ist, ob der überdurchschnittlich starke Befall die Folge
einer Allgemeinerkrankung und damit assoziierter Immunschwäche, oder die primäre
Ursache des plötzlichen Verfalls war. Nach der Anamnese der Landwirte zu urteilen,
ist hingegen die Strongylidose als alleinige Ursache für den rapiden Verfall eher un-

92
wahrscheinlich, zumal das zweite Tier parasitologisch dem Betriebsbild entsprach,
jedoch nicht besonders auffällig war. Beide Tiere wurden nach dem Besuch ge-
schlachtet, jedoch im Zuge dieser Arbeit nicht pathologisch untersucht.
In Anlehnung an die Resultate unserer Untersuchung ist ein über- oder unterdurch-
schnittlicher BCS also nicht auf eine einzelne Ursache zurückführbar, sondern muss
in größerem Zusammenhang gesehen werden. Der BCS ist demnach - isoliert be-
trachtet - als Indikator zur Früherkennung von Endoparasitosen aufgrund seiner mul-
tifaktoriellen Abhängigkeit nicht geeignet, kann jedoch ergänzend im Zusammen-
spiel und unter Einbeziehung bzw. Berücksichtigung anderer Parameter durchaus
hilfreiche Zusatzinformationen über den Zustand des Bestandes und des Einzeltieres
liefern. So besteht etwa ein direkter Zusammenhang zwischen BCS und Hämatokrit,
so dass im Zuge einer Infektion mit H. contortus und T. colubriformis ein Absinken
des BCS proportional zum Hämatokrit beobachtet werden konnte (HOSTE U. CHARI-
TER, 1993).
Als aussagekräftigere Indikatoren für die Effizienz der Wirts-Resilience haben sich
hingegen die regelmäßige Gewichtskontrolle und die Detektion plötzlicher Ge-
wichtsschwankungen erwiesen (BISSET et al., 1991, zitiert in VAN WYK et al., 2006).
Bei schlechterer Körperkondition wird aber, ungeachtet ihrer Ursache, in jedem Fall
die Futtermittelergänzung durch Substitution von Proteinen und Energie empfohlen,
um einer Unterversorgung frühzeitig entgegenzuwirken und das Tier vorsorglich
gegen negative Auswirkungen einer Endoparasitose zu schützen (COOP u. KYRIAZA-
KIS, 1999, zitiert in VATTA et al., 2002) (siehe Kapitel 2.7.2).
Detaillierte Untersuchungen bezüglich der Objektivierbarkeit der palpatorischen
BCS-Ermittlung bzw. der Reproduzierbarkeit der Palpationsbefunde sowie des Ver-
gleichs der Aussagekraft der Messung des Brustumfanges mit der ultraschallgestütz-
ten Messung der Rückenfettdicke sind der Diplomarbeit von Robert WOLF (in Veröf-
fentlichung) zu entnehmen.
5.5 Einfluss des Haltungssystems
Da jeder der drei untersuchten Betriebe für ein unterschiedliches Haltungsmanage-
ment steht, soll im Folgen unter diesem Gesichtspunkt auf Unterschiede und Ge-
meinsamkeiten eingegangen werden. Zu bedenken ist jedoch, dass die hier gewonne-
nen Erkenntnisse aufgrund der geringen Anzahl an Betrieben nicht als allgemeingül-
tig gesehen werden dürfen.

93
5.5.1 Endoparasitenspektrum
In den untersuchten Betrieben konnte ein ähnliches Endoparasitenspektrum gefunden
werden. Unterschiede zeigten sich nur bezüglich Moniezia und Nematodirus, die
nicht in allen Betrieben und generell nur vereinzelt auftraten. Das in dieser Arbeit
erhobene Parasitenspektrum deckt sich auch weitgehend mit Ergebnissen bereits vo-
rangegangener Studien (REHBEIN et al., 1998; SILVESTRE et al., 2000, PAPADOPOU-
LOS et al., 2003). So konnten auch hier bei 89 % der adulten Tiere Eier von Magen-
Darm-Strongyliden (im Sommer vorwiegend Haemonchus und Strongyhides, im
Herbst Teladorsagia, Cooperia, Chabertia und Trichostrongylus) in unterschiedli-
cher Menge im Kot gefunden werden, während von den Jungtieren (bis ein Jahr alt)
69 % positiv waren. Das gefundene Larvenspektrum deckt sich weitgehend mit Un-
tersuchungen von SILVESTRE et al. (2000) und von EYSKER et al. (2005), die in ihrer
Studie in erster Linie Larven von Haemonchus (> 90%), Teladorsagia, Trichostron-
gylus, Cooperia und Chabertia nachweisen konnten.
Coccidienoozysten konnten bei 95 % der Adulten und 100 % der Jungtiere nachge-
wiesen werden, was mit Untersuchungsergebnissen von BORGSTEEDE U. DERCKSEN
(1996), KOUDELA U. BOKOVä (1998) und BALICKA-RAMISZ (1999) weitgehend über-
einstimmt. Kritisch zu hinterfragen ist allerdings, in welchem Ausmaß die Anzahl
der ausgeschiedenen Eier/Oozysten proportional zur Anzahl der vorhandenen, fort-
pflanzungsfähigen adulten Endoparasiten anzusehen ist (KOOPMANN et al., 2006).
Die Umstellung von konventioneller auf biologische Viehwirtschaft geht generell mit
einem im Vergleich zu konventionellen Betrieben breiteren Endoparasitenspektrum
in Bio-Betrieben einher, wobei die Umstellung in der Milchwirtschaft sich weniger
massiv auswirkt, da sich die Haltungsbedingungen im Vergleich zu konventionellen
Betrieben hier meist nur minimal verändern (THAMSBORG et al., 1999, zitiert in CA-
BARET et al., 2002).
In diesem Zusammenhang als bedeutsam erweist sich die Studie von SILVESTRE et al.
(2000), demzufolge eine höhere Speziesdiversität der Endoparasiten, wie sie häufig
in Weidebetrieben, nicht jedoch beim hier untersuchten Weidebetrieb, zu beobachten
ist, negativ mit der jeweiligen Infektionsintensität korreliert. Mit anderen Worten
resultiert ein breiteres Parasitenspektrum in einer balancierten und geringeren Be-
fallsintensität der einzelnen Spezies, wobei die dahinter stehenden Mechanismen
noch unbekannt sind (SILVESTRE et al., 2000; CABARET et al., 2002). Ein möglicher
Grund hierfür könnte sein, dass das Wirtstier eine Infektion mit einem breiteren Pa-

94
rasitenspektrum besser kontrollieren kann als eine hohe Infektionsintensität mit nur
wenigen Spezies. Eine andere Möglichkeit könnte die Konkurrenz innerhalb der Pa-
rasiten im Hinblick auf Nahrungsressourcen sein. Nach SILVESTRE et al. (2000) sinkt
die Infektionsintensität mit der Speziesdiversität, mit kleinerer Besatzdichte bzw.
mehr zur Verfügung stehender Weidefläche pro Tier, sie steigt jedoch mit zuneh-
mendem Alter der Herde. Die Speziesdiversität wird dabei wesentlich von der Her-
kunft der gruppierten Tiere zum Zeitpunkt der Herdengründung beeinflusst, da im
weiteren Verlauf weitgehend versucht wird, bei Zukaufen ausschließlich uninfizierte
Tiere in bereits bestehende Herden einzubringen (CABARET u. GASNffiR, 1994, zitiert
in Silvestre et al., 2000). Ein Zusammenhang zwischen Weidefläche und Speziesdi-
versität konnte bisher nicht nachgewiesen werden (SILVESTRE et al., 2000).
5.5.2 Befallsintensität
• Allgemeines zur Beurteilung der Befallsintensität anhand von Eizählungen
Nach LLOYD (1987) ist ein epg von 500-2000 Nemtatodeneiem als Indikator für eine
subklinische Infektion anzusehen, während bei einem epg von mehr als 2000 klinisch
apparente Infektionen erwartet werden. Bei Jungtieren wies PEARSON (1988) bereits
ab einem epg von 1000 Symptome einer parasitär-bedingten Gastroenteritis nach.
Zu beachten ist jedoch, dass diese Zusammenhänge nicht linear und stark von der
Spezieszusammensetzung der Parasitenpopulation abhängig sind. Die Eizählung re-
flektiert nach MCKENNA (1985) mit hoher Aussagekraft die tatsächliche Menge an
den im Verdauungstrakt des Wirtstieres lebenden Parasiten (REHBEIN et al., 1998;
HOSTE et al., 2001).
• Befallsintensität in den Betrieben
Die Ergebnisse dieser Arbeit zeigen, dass der Zusammenhang von Haltungsform und
Parasiten einer differenzierten Betrachtung bedarf. Zwar unterschieden sich die im
Zuge dieser Diplomarbeit untersuchten biologischen Betriebe zum Teil signifikant
bezüglich der Eiausscheidungsmengen bzw. auch im prozentuellen Anteil auftreten-
der Larven, jedoch weniger hinsichtlich des Parasitenspektrums, das sich - mit Aus-
nahme von Moniezia und Nematodirus, die allerdings insgesamt nur sehr vereinzelt
auftraten und deshalb hier nicht ausführlich behandelt werden - für alle Betriebe
gleich präsentierte.

95
Grundsätzlich kann zwischen vor allem weide- und hauptsächlich stallassoziierten
bzw. haltungsunabhängigen Endoparasiten unterschieden werden. Diese Differenzie-
rung könnte auch an den im Zuge dieser Arbeit untersuchten Betrieben vorgenom-
men werden. Vor allem innerhalb der Jungtiere ist zu beobachten, dass die Unter-
schiede zwischen den Betrieben nicht eindeutig der Haltung zugeschrieben werden
können. Vor dem Hintergrund, dass es sich bei den Jungtieren in erster Linie um
Coccidien-Infektionen handelt und Coccidien unabhängig vom Haltungssystem re-
gelmäßig auftreten, ist dies jedoch nicht überraschend. Die Werte des opg sind je-
doch durchwegs hoch angesiedelt, wobei Betrieb 3 die höchsten Werte zeigt. Über
gezielte Therapien und unterstützende Maßnahmen wie z.B. Desinfektion sollte
nachgedacht werden, um wirtschaftliche Verluste zu minimieren.
Die Coccidieninfektionen, die bei Jungtieren häufig eine Verbreitung von bis zu
100 % und auch bei adulten Tieren 65-80 % erreichen können (BORGSTEEDE U.
DERCKSEN, 1996; BALICKA-RAMISZ, 1999), werden von den Faktoren der Haltung
und dem Carrier-Status der Muttertiere als Infektionsquelle für die Jungtiere insofern
nachhaltig beeinflusst, als dass - einhergehend mit einer zunehmenden Intensivie-
rung der Ziegenhaltung - Zunahmen der Befallsintensität verzeichnet werden können
(BALICKA-RAMISZ, 1999). Deutlich ist auch der Unterschied in der Prävalenz der
Kokzidiose nach Umstellung konventioneller Betriebe auf biologische Bewirtschaf-
tung, die im Zuge der Einführung des Tiefstreusystems an Häufigkeit gewonnen hat
(CABARET et al., 2002).
Dass hingegen keine herausragenden betrieblichen Unterschiede bezüglich MDS bei
den Jungtieren zu verzeichnen waren, lässt vermuten, dass sich die Haltungsform erst
mit fortschreitendem Alter maßgeblicher auf die Befallsintensität auswirkt. Auch
hinsichtlich Strongyloides- und Trichuris-Eiem ist Ähnliches zu beobachten. Zu be-
denken ist hier allerdings, dass die Haltungsbedingungen der Jungtiere in allen Be-
trieben ähnlich (Tiefstreuboxen mit Auslauf) waren. Mit fortschreitendem Alter stel-
len sich hingegen die für den Weidebetrieb erwarteten höheren Befallsraten bezüg-
lich MDS und Trichuris ein.
Aus diesem Grund ist bei der Weidehaltung eine genaue Überwachung und Erhe-
bung der Endoparasitenbelastung besonders bedeutsam, da die Weide - aufgrund der
besseren Überlebensmöglichkeiten für die Parasiten - deutlich mehr Infektionsmög-
lichkeiten mit infektiösen Endoparasitenstadien bietet als Haltungssysteme mit befes-
tigten Ausläufen. Umso wichtiger wird gutes Herden- und Gesundheitsmanagement

96
als primäre Strategie im Öko-Landbau, wobei zu beachten ist, dass trotz verschie-
denster Managementstrategien zur Endoparasitenbekämpfung bei fast allen Tiere in
ökologisch wirtschaftenden Betrieben Endoparasiten nachgewiesen werden können
(KoOPMANN, 2005) und demnach eine Endoparasitenfreiheit wohl kaum erreicht
werden kann. Ziel in der biologischen Landwirtschaft muss es viel mehr sein, ein
balanciertes Verhältnis zwischen Pathogenen und Tieren herzustellen (THAMSBORG
et al., 1999).
Die Tragweite der Parasiteninfektion steht in direktem Zusammenhang mit der Men-
ge der aufgenommenen infektiösen Stadien, was wiederum eng an die Art der Futter-
aufnahme gekoppelt ist. Demnach ist das Infektionsrisiko für Ziegen, die die Mög-
lichkeit der selektiven Futteraufnahme von Büschen und Stauden haben, geringer, als
für Ziegen, die zum ausschließlichen Grasen gezwungen sind, da parasitäre Entwick-
lungsstadien eher in Bodennähe anzutreffen sind. Bezüglich des Fressverhaltens
spielen jedoch auch Rassenunterschiede eine Rolle: so konnte in Versuchen festge-
stellt werden, dass Angora-Ziegen fast ausschließlich grasen und kaum an Büschen
und Stauden knabbern, während Saanen-Ziegen dies bevorzugen. Damit einher gin-
gen deutlich höhere Eiausscheidungsraten der Angora-Ziegen gegenüber den Saa-
nen-Ziegen (HOSTE et al., 2001). Die Infektionsintensität, die Speziesdiversität sowie
die Lebenszyklen der Nematoden werden auch stark von medikamenteilen Antipara-
sitika-Behandlungen beeinflusst, wobei die Speziesdiversität zusätzlich von Anthel-
minthika-Resistenzen moduliert werden kann (CABARET, 2000, zitiert in SILVESTRE
et al., 2000).
Das Ausmaß der Weidekontamination ergibt sich wiederum aus der in den Tieren
präsenten Parasitenmenge, nachdem jedes Parasitenstadium den Verdauungstrakt des
Wirtes durchlaufen muss, bevor es auf die Weide gelangt. Somit ist sie abhängig von
physiologischen und metabolischen Prozessen des Wirtstieres und den Bestrebungen
des Betriebs hinsichtlich der Reduktion des Infektionsrisikos (WALLER, 2006a).
Während in dieser Studie bezüglich Strongyloides keine signifikanten haltungsbe-
dingten Unterschiede nachgewiesen werden können, ist für Skrjabinema eine deutli-
che Stallassoziation feststellbar. Bereits BORGSTEEDE U. DERCKSEN (1996) wiesen
darauf hin, dass Infektionen mit Eimerien, S. papillosus und S. ovis auch bei reiner
Stallhaltung auftreten können. Intensivere Haltungsformen scheinen außerdem prä-
destinierend für das vermehrte Auftreten von Anthelminthika-Resistenzen zu sein,
was nicht zuletzt auf den mit erhöhter Parasitenprävalenz in Intensivhaltung verbun-

97
denen steigenden Antiparasitika-Einsatz zurückgeführt werden kann, so dass Resis-
tenzen in extensiver Haltung seltener nachgewiesen werden (HERTZBERG U. BAUER,
2000).
Wie später im Kapitel 5.8 eingehender behandelt, bietet der Weidegang zudem die
für die Immunitätsbildung nötige Kontaktmöglichkeit mit Parasiten und bildet auf
diesem Weg die Basis zur Krankheitsprophylaxe, also dem Grundanliegen der öko-
logischen Landwirtschaft. Die Pflicht der Landwirte, nach biologischen Produktions-
richtlinien Weidegang zu gewährleisten, stellt zudem einen der wesentlichen Unter-
schiede zur konventionellen Tierhaltung dar und ist als „Qualitätskennzeichen des
Öko-Landbaus" (KOOPMANN, 2005) nicht selten eines der Hauptargumente des öko-
logisch sensibilisierten Konsumenten, biologische statt konventionell erzeugter Pro-
dukte zu kaufen.
5.6 Einfluss des Alters
In unseren Untersuchungen konnten ebenfalls signifikante Unterschiede bezüglich
der Eiausscheidung zwischen Jungtieren und Adulten festgestellt werden, unter den
adulten Tieren waren jedoch keine signifikanten altersbedingten Unterschiede nach-
weisbar. So war das Infektionsausmaß insbesondere bei Coccidien bei Jungtieren
deutlich höher als bei den adulten Tieren. Dies stimmt sowohl mit den Erkenntnissen
von BORGSTEEDE u. DERCKSEN (1996), REHBEIN et al. (1998), als auch mit KOUDELA
u. BOKOVÄ (1998) sowie BALICKA-RAMISZ (1999) überein, die nachweisen konnten,
dass das Infektionsausmaß insbesondere bei Coccidien altersabhängig und bei Jung-
tieren deutlich höher ist, was jedoch nicht zwangsläufig mit klinischen Erscheinun-
gen einhergehen muss, deren Ausmaß vor allem speziesabhängig ist. Besonders in-
nerhalb^der Coccidienarten gibt es große Pathogenitätsunterschiede und nicht jede
Altersstufe wird in gleichem Maße von den selben Spezies besiedelt, so konnten et-
wa in einer tschechischen Studien bei Kitzen primär E. arloingi, bei älteren Tieren E.
ninakohlyakimovae am häufigsten nachgewiesen werden (KoUDELA u. BOKOVÄ,
1998). Vermutlich treten bei Jungtieren häufiger hochpathogene Arten auf, die auch
bei geringerer Befallsintensität zu Krankheitssymptomen führen. Bei adulten Tieren
sind demzufolge zwar häufig vergleichbare Mengen, allerdings eventuell weniger
pathogene Spezies vertreten, so dass Infektionen klinisch inapparent bzw. subkli-
nisch verlaufen. Grund für den milderen oder subklinischen Krankheitsverlauf könn-
te jedoch auch die nach Coccidien-Erstinfektionen beobachtete gute Immunitätsin-

98
duktion der pathogenen Arten sein, die ältere Tiere vor erneuten Erkrankungen
schützt (BORGSTEEDE u. DERCKSEN, 1996). Nachdem im Zuge dieser Arbeit keine
nähere Differenzierung der Coccidien durchgeführt wurde, können keine Aussagen
über die eventuell altersbedingte Artenverteilung getroffen werden.
Bei den übrigen nachgewiesenen Parasiten konnte - wie auch bei LLOYD u. SoULSBY
(1978, zitiert in REHBEIN et al., 1998) und RICHARD et al. (1990, zitiert in HOSTE et
al., 2008) beschrieben - eine Zunahme der Befallsintensität mit zunehmendem Alter
beobachtet werden. Besonders deutlich war dieses Phänomen bezüglich Eiausschei-
dungsanzahl von MDS zu beobachten, wonach diese bei alten Tieren häufig signifi-
kant höher war als bei Jungtieren. Innerhalb der adulten Tiere lagen keine altersbe-
dingten Unterschiede mehr vor. Die Eiausscheidung nimmt hier also zwischen dem
ersten Lebensjahr und der Adoleszenz zu und ist im adulten Lebensabschnitt viel-
leicht weniger altersabhängig als vielmehr von anderen Faktoren wie Laktationssta-
dium, Immunität, Ernährung etc. bestimmt. Auch bei Strongyloides ist eine Altersab-
hängigkeit nur im Vergleich zwischen Jungtier und adultem Tier nachweisbar, wäh-
rend in späteren Jahren offensichtlich ebenso der Einfluss oben genannter Faktoren
überwiegt. Den auch noch bei alten Tieren häufig anzutreffenden hohen Ausschei-
dungsraten von Nematodeneiem könnte die bereits häufig nachgewiesene unzurei-
chende Fähigkeit der Ziege zugrunde liegen, auf eine Endoparasiteninfektion immu-
nologisch adäquat im Sinne einer effizienten Immunantwort reagieren zu können
(siehe Kapitel 5.8).
Bei Schafen liegen sehr klare altersbedingte Unterschiede in der Endoparasitenbe-
lastung vor. Demnach weisen jüngere Tiere üblicherweise höhere Befallsraten auf als
ältere. Dieses Phänomen kann bei Ziegen generell zwar auch beobachtet werden, es
ist jedoch weniger deutlich ausgeprägt (POMROY et al., 1986, zitiert in HOSTE et al.,
2008). Etliche Studien (unter anderem HOSTE et al., 1993, 1999, 2001, 2002b) bele-
gen, dass Erstlaktierende sehr häufig eine höhere Eiausscheidung aufweisen, von der
auf das Vorhandensein einer größeren Wurmbürde geschlossen werden kann und die
sehr wahrscheinlich auf eine noch nicht oder nicht ausreichend entwickelte Immuni-
tät zurückgeführt werden kann. In anderen Studien konnte, wie bereits erwähnt, hin-
gegen Gegenteiliges beobachtet werden, wonach häufig auch alte Tiere eine hohe
Eiausscheidungsrate aufweisen.
Insgesamt kann der Faktor Alter jedoch trotz widersprüchlicher Ergebnisse als
Parameter für eine selektive Antiparasitika-Behandlung hinzugezogen werden. Dem-

99
nach handelt es sich vor allem bei Jungtieren, Erstlaktierenden und Hochlaktierenden
um Risikogruppen, die im selektiven Behandlungsregime unbedingt berücksichtigt
werden müssen. Individuelle Schwankungen in der Eiausscheidung, wie sie von
HOSTE et al. (2002b) nachgewiesen wurden, müssen dabei jedoch ebenfalls bedacht
werden, indem man etwa versucht, die Hauptausscheider zu eruieren, die eine
Hauptkontaminations- und -Infektionsquelle für die gesamte Umgebung und die rest-
liche Herde darstellen. So zeichnen sich hier bereits die beiden grundlegenden Mög-
lichkeiten der selektiven Behandlung ab, nämlich zum einen die Einteilung der Tiere
in Untergruppen nach Alter und Laktationsstadium und zum anderen die individuelle
Risikobeurteilung und Identifikation einzelner Hauptausscheider.
5.7 Einfluss von Jahreszeit und Klima
Bei den im Rahmen dieser Studie untersuchten Betrieben waren deutliche jahreszeit-
liche Unterschiede in Bezug auf die Eiausscheidungsmenge und die Larvenprävalenz
bemerkbar. Die Eiausscheidungsrate zeigte bezüglich der meisten Parasiten höhere
Tendenzen beim Betriebsbesuch im Sommer bzw. lag sogar signifikant höher als im
Herbst. Bei aus den Kotkulturen determinierten Larven war eine deutliche Präferenz
von Haemonchus und auch Strongyloides in den warmen Monaten erkennbar, wäh-
rend die übrigen Arten im Jahresverlauf zu kumulieren schienen und im Herbst ver-
mehrt nachweisbar waren.
Wie auch schon in den im Zuge dieser Arbeit untersuchten Betrieben, konnten saiso-
nale Schwankungen der Ei- bzw. Oozystenausscheidung bei Ziegen auch schon in
vorangegangenen Studien nachgewiesen werden (BALICKA-RAMISZ, 1999; PAPADO-
POULOS et al., 2003), wobei der jahreszeitliche Einfluss sich primär nicht signifikant
auf die vorhandene Anzahl adulter Würmer sondern nur auf die Eiausscheidungsrate
auswirkte. Nachdem im Rahmen dieser Diplomarbeit keine Sektionen durchgeführt
werden konnten, kann diesbezüglich keine Aussage getroffen werden.
Jegliche Art von Umweltfaktoren beeinflussen nachhaltig die Prävalenz von Parasi-
tenstadien in der Wirtsumgebung und somit auch die tägliche Aufnahme infektiöser
Parasitenstadien durch das grasende Wirtstier, die somit starken saisonalen Schwan-
kungen unterliegen kann. Faktoren wie Niederschlagsmenge und Temperatur sind
dabei von besonderer Bedeutung, indem sie für saisonalen Peaks Höhepunkte an auf
der Weide verfügbaren Parasitenstadien verantwortlich sind. Diese Höhepunkte sind
jedoch nicht nur klimatischen (z.B. Larven sind bei warmem, feuchten Bedingungen

100
aktiver da sie unter diesen Verhältnissen die Fähigkeit zur aktiven Beweglichkeit
haben (somit ist das Übertragungsrisiko höher), überleben jedoch bei kühlen, feuch-
ten Bedingungen länger; SOUTHCOTT et al., 1976, zitiert in BARGER, 1999) sondern
auch tierspezifischen (z.B. herabgesetzte immunologische Widerstandsfähigkeit der
Geißen um den Zeitpunkt der Geburt, dem „periparturient rise", oder während der
Laktation; ROBERTS et al., 1991, zitiert in BARGER, 1999) Schwankungen unterwor-
fen und führen meist unmittelbar zum Auftreten klinisch apparenter Endoparasitosen.
Ziel eines nachhaltigen und erfolgreichen Parasitenmanagements muss es daher sein,
genau jene Höhepunkte zu erkennen, sie frühzeitig zu bekämpfen oder besser sie zu
verhindern (BARGER, 1999). Sinnvoll wäre es, diese saisonalen Schwankungen be-
züglich antiparasitärer Therapien zu nützen. Aus parasitologischer Sicht durchaus
effizient erscheint z.B. eine Antiparasitika-Behandlung in den Monaten, in denen die
Parasitenbelastung der Weideflächen klimatisch bedingt gering ist, so dass die Ge-
fahr der Reinfektion nach erfolgter Behandlung minimiert werden kann (BARGER,
1999), andererseits muss dies in Anbetracht der Resistenzproblematik wiederum kri-
tisch hinterfragt werden (Näheres dazu siehe Kapitel 2.5.1).
5.7.1 Allgemeine klimatische Faktoren
Klimatische Faktoren üben nicht nur in Form von saisonalen Variationen hinsichtlich
der auf der Weide verfügbaren Parasitenstadien, sondern auch auf die allgemeine
Speziesprävalenz einen bedeutenden Einfluss aus. Demzufolge spielen Haemonchus
und Cooperia vor allem in subtropischen bis tropischen Regionen die größte Rolle,
während Teladorsagia und Nematodirus vermehrt in gemäßigten Zonen auftritt und
Trichostrongylus ubiquitär präsent ist (WALLER, 2006a). Allerdings konnte in unse-
rer Studie eine hohe Prävalenz an Haemonchus und Cooperia nachgewiesen werden,
während Nematodirus nur vereinzelt gefunden werden konnte.
5.7.2 Niederschlagsmenge
Die Niederschlagsmenge beeinflusst laut vorangegangenen Studien nachhaltig die
Lebensdauer der infektiösen Parasitenstadien. Das feuchte Milieu in der untersuchten
Region allgemein und insbesondere in den Sommermonaten bietet somit einen idea-
len Lebensraum für viele Endoparasiten, was zu beträchtlichem Infektionsdruck auf
bewachsenen Ausläufen und Weiden führen kann.
Untersuchungen von PAPADOPOULOS et al. (2003) ergaben einen Höhepunkt der Ei-

101
ausscheidungsrate im Frühling. Abgesehen vom häufig beobachteten „periparturient-
rise", also einem durch eine Immunschwäche vor und nach der Geburt bedingten
temporären Anstieg der Eiausscheidung (GiBBS, 1986, zitiert in ZAJAC, 2006), könn-
te dies nicht zuletzt auf die infolge vermehrter Regenfälle höhere Infektionsrate zu-
rückzuführen sein, so dass nach Ablauf der Präpatenzzeit ein Anstieg der Eiauss-
cheidung zu verzeichnen ist. (NffiZEN et al., 1996). Ein zweiter kleinerer Peak war in
dieser Studie daraufhin im Herbst zu beobachten, wo ebenfalls höhere Nieder-
schlagsmengen verzeichnet wurden und deshalb als Ursache für vermehrte Infekti-
onsraten gesehen werden können.
5.7.3 Temperatur
Auch die Temperaturen stellen nachweislich einen wichtigen Faktor dar, insofern
Larven bei warmen und trockenen Bedingungen generell eine weitaus kürzere Über-
lebensrate zeigen als bei kühlem und feuchtem Klima (BARGER, 1999). Die allge-
mein kühleren Temperaturen in der untersuchten Region bieten in Verbindung mit
hohen Niederschlagsmengen demnach gute Überlebensbedingungen für die Parasi-
ten. Die sehr kalten Wintermonate in der untersuchten Region schaffen jedoch einen
zum Teil suboptimalen Endoparasitenlebensraum. Dies spiegeln auch die Ergebnisse
dieser Arbeit wieder, wonach eine deutliche Kälte-Sensitivität von H. contortus
(BARGER, 1999) zu erkennen war und auch von Strongyloides nachgewiesen werden
konnte, so dass in den Kotkulturen der im Herbst entnommenen Proben nur noch
vereinzelt Larven dieser Parasiten zu finden waren. Hier wäre demnach eine Behand-
lung während der Wintermonate aus parasitologischer Sicht sehr effizient, um einen
nachhaltigen Effekt zu erzielen und die Gefahr der Reinfektion zu minimieren, aller-
dings ist auch hier an die Anthelminthika-Resistenz-Problematik zu erinnern.
5.8 Die Bedeutung immunologischer Faktoren
Bei den im Zuge dieser Arbeit untersuchten Tieren waren starke Variationen in der
Eiausscheidung nachweisbar. Innerhalb einer Haltungsgruppe konnten sowohl Tiere
mit hohen als auch Tiere mit geringer Eiausscheidungsrate gefunden werden, obwohl
die Haltungsbedingungen (Fütterung, Aufstauung etc.) für alle Tiere gleich vorlagen
und die Gruppierung - zumindest in Betrieb 1 und 2 - leistungsorientiert erfolgte.
Neben Faktoren wie Alter, Produktionsleistung etc. bestehen demnach sehr individu-

102
eile Varianzen in der Empfänglichkeit der Tiere gegenüber Parasiteninfektionen, die
nicht zuletzt auf immunologische Faktoren zurückgeführt werden können.
Die Konfrontation des Organismus mit einem körperfremden Antigen führt bei intak-
ter Immunlage des Individuums zu einer Immunantwort, die mitunter einen lang an-
haltenden immunologischen Schutz im Falle einer wiederholten Konfrontation mit
demselben Antigen zur Folge hat. Im Unterschied zu mikrobiellen Infektionen, die
eine prompte und aggressive Immunreaktion induzieren, rufen Nematoden eine sich
langsam entwickelnde Immunantwort hervor. Die Ursache dafür ist die im Wirt bis
zu Wochen dauernde Vollendung des Replikationszyklus der Nematoden, wodurch
nur langsam ausreichende Mengen an Antigen-Information erreicht werden, die zur
Antigen-Erkennung und Mobilisierung des Wirt-Immunsystems nötig sind (DiNEEN
et al., 1965) Außerdem induzieren Nematoden verschiedene Formen von immunolo-
gischer Toleranz, um ihr längeres Überleben zu gewährleisten (DiNEEN, 1963, zitiert
in WALLER, 2006a).
Eine Immunreaktion kann folgende Strategien hinsichtlich einer Konfrontation mit
einer Endoparasiteninfektion verfolgen: 1. verminderte Etablierung der infektiösen
Parasitenstadien (messbar anhand der tatsächlichen Wurmmenge in der Sektion); 2.
reduziertes Wachstum der adulten Tiere (Messung der Länge der Würmer); 3. redu-
zierte Fertilität der Parasitenweibchen (Messung der Eiausscheidung, Ermittlung der
Eiproduktion in utero); und 4. die Expulsion adulter Würmer. Die Etablierung der
endogenen Stadien kann bei der Immunantwort der Ziegen gegenüber Endoparasiten
in nur sehr geringem Umfang, eine Expulsion von Würmern im Allgemeinen nicht
beobachtet werden (BALIC et al., 2000, zitiert in HOSTE et al., 2008).
Das Ausmaß einer Infektion mit gastrointestinalen Nematoden ist beim Wiederkäuer
sehr stark von einer effektiven Immunantwort bzw. der Fähigkeit des Wirtstieres
abhängig, die Wurmpopulation immunologisch zu regulieren (HOSTE et al., 2001).
Einer Immunantwort muss jedoch - wie bereits erwähnt - immer eine Konfrontation
mit dem jeweiligen Antigen vorangehen. Dementsprechend muss auch bei Endopara-
siten eine direkte Exposition sichergestellt sein, um langfristigen immunologischen
Schutz etablieren zu können. Die Präsenz einer gewissen Endoparasitenpopulation
scheint also notwendig, um immunologische Abwehrmechanismen gegenüber neu
aufgenommenen Parasitenstadien zu stimulieren. Aus diesem Grund wirken sich
höchst effiziente Parasitenkontrollprogramme in Ermangelung des für eine Immun-
reaktion nötigen Antigens negativ auf die Entwicklung einer anhaltenden Immun-

103
antwort aus, so dass es zum Phänomen der „Underexposition", d.h. der zu geringen
Antigen-Exposition, kommt. Die Tiere bleiben infolgedessen bei späterer Exposition
gegenüber bzw. nach Behandlungsende weiterhin empfänglich (THAMSBORG et al.,
1999).
Des Weiteren konnte beobachtet werden, dass die Immunregulation der Eiausschei-
dungsrate bei unbehandelten Tieren effizienter funktioniert, was bedeuten würde,
dass eine negative Interaktion zwischen Anthelminthika-Behandlungen und der Effi-
zienz einer wirtseigenen Immunregulation besteht (HOSTE U. CHARTIER, 1998; HOSTE
et al., 2002d). Diese Erkenntnis kann im selektiven Behandlungsverfahren genutzt
werden (HOSTE et al., 2002c).
5.8.1 Aggregierte Parasitenverteilung
Wie bereits erwähnt, lagen bei den für diese Studie untersuchten Tieren große
Schwankungen hinsichtlich der Parasitenverteilung innerhalb der Haltungsgruppen
vor. Die Ursache für diese Varianzen scheint nicht zuletzt in der individuellen Im-
munreaktion des Einzeltieres zu liegen.
Wie bereits erwähnt, scheint die Fähigkeit zur Etablierung einer effizienten Immun-
antwort bei der Ziege von vornherein schlechter ausgebildet zu sein bzw. langsamer
zu verlaufen als. beim Schaf, wodurch sie empfänglicher für parasitäre Infektionen
ist (HOSTE U. CHARTIER, 1998a). Allerdings unterliegt die Fähigkeit zur Etablierung
einer effizienten Immunabwehr sehr stark individuellen Schwankungen, demzufolge
können einige Tiere grundsätzlich immunologisch wirksamer reagieren als andere
(HOSTE et al., 2001a, 2002b), so dass bei wiederholten Betriebsbesuchen höchst-
wahrscheinlich dieselben Tiere hohe Eiausscheidungsraten aufweisen werden, dem-
nach die Verteilung der Parasiten ungleichmäßig, also aggregiert, vorliegt. Wenige
Tiere sind epidemiologisch als Ausscheider großer Eimengen als Hauptkontaminati-
onsquelle für die Umgebung von größter Bedeutung (HOSTE et al., 2002a, 2002b).
Die gezielte Probenentnahme bei denselben Tieren über einen längeren Zeitraum
wäre auch für unsere Studie sehr aufschlussreich, um eine Aussage über eine Un-
gleichverteilung der Parasiten innerhalb derselben Herde treffen zu können. Nach-
dem die Tiere für unsere Untersuchungen jedoch randomisiert ausgewählt wurden,
ergaben sich keinerlei zufällige Überschneidungen in der Tierauswahl zwischen den
beiden Besuchen von Sommer und Herbst.
In Untersuchungen von ETTER et al. (2000) konnte bei immunologisch aktiveren Tie-

104
ren eine höhere Zelldichte an eosinophilen Granulozyten, Mastzellen und Monozyten
in der Darmschleimhaut nachgewiesen werden. Diese Tatsache scheint unmittelbar
mit einer geringeren Empfindlichkeit der Tiere zusammen zu hängen. Laut ETTER et
al. (2000) könnte zwischen der höheren Dichte an eosinophilen Granulozyten in der
Darmschleimhaut und einer geringeren Wurmbürde, sowie zwischen den Mastzellen
und Monozyten und der Regulation der Eiausscheidung ein direkter Zusammenhang
bestehen. Das Phänomen der aggregierten Parasitenverteilung könnte man in jedem
Fall im Parasitenmanagement im Sinne der selektiven Behandlung jener Tiere nützen
(HOSTE et al., 2002b).
5.8.2 „Resilience" oder die immunologische Belastbarkeit
Die immunologische Widerstandsfähigkeit des Wirtstieres spielt insofern eine sehr
bedeutende Rolle, als klinisch apparente Parasitosen im Vergleich zu inapparenten
Tieren nicht selten mit einer niedrigeren Eiausscheidungsrate der Erkrankten ver-
knüpft sind. Das kann darauf zurückgeführt werden, dass die Fähigkeit des Wirtes,
die Wurmbürde und deren negative Auswirkungen zu kontrollieren, nicht zwingend
an der Eiausscheidungsrate ablesbar ist und das Ausmaß der Infektion, ab dem eine
klinische Symptomatik auftritt, sehr individuell und von Tier zu Tier verschieden ist
(VAN WYK et al., 2006). Auch bei Coccidien konnte beobachtet werden, dass manche
Kitze mit sehr hohen Oozystenausscheidungsraten oft nur subklinisch infiziert sind
(KOUDELA U. BOKOVÄ, 1998).
5.8.3 Immunität in Laktation und Trächtigkeit
Die Doppelbelastung durch Milchproduktion und Zucht in den hier untersuchten
Betrieben stellt hohe Ansprüche an den Organismus und das Immunsystem der Tiere.
Generell können hohe nutritive und metabolische Anforderungen wie z.B. in Träch-
tigkeit und Laktation sehr leicht zu einem Ungleichgewicht zwischen Nährstoffbe-
darf und Nährstoffaufnahme führen. Bei der Ziege resultiert dies in der Umschich-
tung aller zur Verfügung stehenden Nährstoffe zu Lasten anderer Kompartimente -
wie dem Immunsystem - zur Deckung des erhöhten Bedarfs. Dies führt bei Schafen
zur „peri-parturient relaxation of immunity (PPRI)" (TEMBELY et al., 1998) genann-
ten Immunschwäche, die bereits in der Spätlaktation beginnt und bis zum Einsetzen
der neuen Laktationsphase andauert (Überblick bei HOSTE et al., 2008). Unter diesem

105
Gesichtspunkt erscheint es auch verständlich, dass die Immunitätsbildung nach ei-
nem ähnlichen Phänomen milchleistungsabhängig ist und bei Hochleistungstieren als
besonders schlecht angesehen werden kann. Das erklärt, dass eben jene Tiere im
Vergleich zu Tieren mit geringerer Milchleistung vermehrt unter den negativen
Auswirkungen eines Parasitenbefalls leiden bzw. diese stärker zeigen und auch als
Eiausscheider besonders bedeutsam sind (HOSTE U. CHARTIER, 1993; CHARTffiR u.
HOSTE, 1997; HOSTE U. CHARTffiR, 1998, HOSTE et al., 2002b, 2005). Entscheidend
für eine höhere Parasitenbelastung bei Hochleistungstieren scheint auch der erhöhte
Nahrungsbedarf und die damit verbundene steigende Grasaufnahme, die die Auf-
nahme parasitärer Entwicklungsstadien (Eier, Oozysten, Larven etc.) erleichtert
(HOSTE et al., 2001).
Während bei Milchschafen keine Unterschiede in der Eiausscheidung in Hinsicht auf
die altersbedingte Milchleistung nachgewiesen werden konnten, müssen auch junge
Milchziegen in der ersten Laktation als besonders gefährdet eingestuft werden (HOS-
TE et al., 2001a, 2002b, 2006). Diesbezüglich gibt es jedoch konträre Ansichten, und
verschiedene Untersuchungen erbrachten widersprüchliche Ergebnisse. Gründe für
eine niedrigere Belastung könnten etwa sein, dass Erstlaktierende zum einen einem
anderen Management unterworfen sind und häufig getrennt von den adulten Milch-
ziegen auf eigenen Weiden gehalten werden, so dass eine geringere Eiausscheidung
allein dadurch erklärbar wäre. Zum anderen könnte die erhöhte Empfänglichkeit für
Endoparasiten auch ein Resultat fehlender vorangegangener Kontaktmöglichkeiten
mit Nematoden sein, was wiederum nachhaltig auf die Aufzuchtbedingungen der
Jungziegen zurückzuführen ist. Eine dritte Ursache einer geringeren Parasitenbelas-
tung der Jungziegen könnte das geringere Körpergewicht sein, wodurch es seltener
zu einer Anthelminthika-Unterdosierung kommt, als bei adulten Ziegen (HOSTE et al.
2002b). Nachgewiesen werden konnte ebenso, dass Geißen um den Zeitpunkt der
Geburt bzw. in der Phase der Laktation ihre im Vorfeld erlangte Immunität gegen
gastrointestinale Nematoden vorübergehend einbüßen, und es dadurch zu einem vo-
rübergehenden Anstieg der Eiausscheidung kommt. Somit stellen Muttertiere zu die-
sen Zeitpunkten (bei Geburtstermin im Frühling demnach im HerbstAVinter) eine
Hauptkontaminationsquelle für die Umgebung dar, der das neugeborene Kitz gänz-
lich ausgesetzt ist. Demzufolge wäre eine anthelminthische Behandlung nach der
Belegung empfehlenswert. Die im Kapitel 2.7.2.1 eingehender besprochene Supple-
mentierung von qualitativ hochwertigen Proteinen soll der Explosion der Wurmbürde

106
und dem damit verbundenen Anstieg der Eiausscheidung entgegenwirken (DONALD-
SON, 1997, zitiert in BARGER, 1999).
5.8.4 Strategien der Ziege zum Schutz vor Endoparasiten
Abgesehen von Alter, Laktationsstadium und Trächtigkeit ist das Graseverhalten ein
wesentlicher Faktor für die Endoparasitenbelastung. In den hier untersuchten Betrie-
ben war es nur den Ziegen in Betrieb 3 möglich, zu grasen und die Ration zu einem
Großteil selbst zu gestalten, während die Futterbeschaffenheit der Tiere in den Aus-
laufbetrieben von der vom Landwirt festgelegten Ration determiniert wird. Liegt eine
Kontamination des verfütterten Heus oder Grünfutters mit infektiösen Parasitensta-
dien vor, so hat die Ziege keine Möglichkeit, die Infektion zu vermeiden. Wenn Zie-
gen jedoch ihr arttypisches Graseverhalten, also nicht das für Schafe typische Gra-
sen, sondern vielmehr das selektive Knabbern an Bäumen und Büschen, ausüben
können, sind sie weniger auf eine aktive Immunantwort gegenüber Endoparasiten
angewiesen, da sie augrund der bodenfemen Futteraufnahme keinem hohen Infekti-
onsrisiko unterliegen und im Vergleich zu Schafen seltener Parasiteninfektionen zei-
gen (POMROY et al., 1986, zitiert in HOSTE et al., 2001, 2008). Die Abwehrstrategie
der Ziegen scheint also vielmehr darauf abzuzielen, durch selektive Futteraufnahme
von Pflanzen und ihrer Sekundärstoffe eine Infektion von vornherein zu vermeiden.
Dies ist jedoch bei der Auslaufhaltung generell nicht und bei Weidehaltung auch nur
dann möglich, wenn die Weidebeschaffenheit in Form der vorhandenen Vegetation
die Aufnahme von Sekundärstoffen durch selektive Futteraufnahme zulässt. Dem-
nach kann nicht jede Weide diesem Anspruch Folge leisten. In intensiverer Haltung
haben Ziegen in Ermangelung der bereits erwähnten extensiven Weiden sogar nur
selten die Möglichkeit, ihrem artspezifischen Futteraufnahmeverhalten nachzugehen.
Infolge dessen werden Ziegen vermehrt zum Grasen gezwungen, was durch die Bo-
dennähe ein größeres Infektionsrisiko in sich birgt (HOSTE et al., 2001; HOSTE et al.
2006a, zitiert in HOSTE et al., 2008).
Generell investieren Ziegen, im Gegensatz zu Schafen, die ihre Ressourcen primär
zur effizienten Immunabwehr nützen, also weniger Energie in die Parasitenabwehr in
Form der Etablierung einer Immunantwort, als vielmehr in die Optimierung einer
verbesserten Resilience zur Minimierung negativer Nebeneffekte einer Infektion.
Sind sie nun zur aktiven Immunabwehr gezwungen, so geht dies zu Lasten der Pro-
duktionsleistung (HOSTE et al., 2008).

107
5.9 Frühdiagnostik
Die Früherkennung infizierter Tiere ist bezüglich eines nachhaltigen Parasitenmana-
gements besonders bedeutsam, da diese durch häufig sehr hohe Eiausscheidungsraten
das Infektionsrisiko der Herde immens erhöhen. Der Landwirt spielt dabei eine zent-
rale Rolle. Bei der im Zuge dieser Arbeit durchgeführten Befragung der Landwirte
wurden in erster Linie Symptome wie Gewichtsverlust, Durchfall und Husten als
auslösende Faktoren für die Durchführung einer Endoparasitenbehandlung angege-
ben. Einer der Landwirte gab an, regelmäßig den Ernährungszustand, insbesondere
bei auffälligen Tieren, durch Palpation zu ermitteln.
Die Bestimmung des Ernährungszustandes anhand des BCS ist sicherlich eine der
praxisrelevantesten Methoden zur Überprüfung der Körperkondition, da sie vom
Landwirt selbst und ohne zusätzliche Hilfsmittel durchgeführt werden kann. Die Un-
tersuchungen dieser Arbeit ergaben jedoch, dass der BCS keinen verlässlichen Indi-
kator zur Erkennung von Endoparasitosen darstellt. In vorangegangenen Studien
erwies sich hingegen eine Kombination aus konsequentem monatlichen Monitoring
der Produktionsresultate (Evaluierung der täglichen Gewichtszunahme) und der In-
fektionsrate anhand der Ermittlung der durchschnittlichen Eiausscheidungsrate als
effizient, um die Dringlichkeit einer Bestandsbehandlung bzw. die Notwendigkeit
einer selektiven Behandlung besonders empfänglicher Tiergruppen oder auch Einzel-
tiere jungen Fleischrindem in Argentinien zu ermitteln (Übersicht bei CABARET et
al., 2002; HOSTE et al., 2002a).
Neben der Körperkondition und der Eiausscheidung können Methoden wie die Mes-
sung der Immunantwort und Blutuntersuchungen - zur Identifikation subklinischer
Teladorsagiose anhand erhöhter Pepsinogenwerte; (HöGLUND et al., 2001, zitiert in
CABARET et al., 2002), oder die Messung anorganischer Phosphate - hinzugezogen
werden. Beide Parameter geben dabei Aufschluss über eventuell vorhandene und
parasitär bedingte Schleimhautläsionen in Abomasum und Darmtrakt (Review siehe
HOSTE et al., 2002d). Diese Methoden sind jedoch mit hohem finanziellem und zeit-
lichem Aufwand verbunden und können nicht vom Landwirt selbst durchgeführt
werden. Außerdem kann auch das klinische Vorhandensein und Ausmaß von Anämie
als Hinweis auf vorliegende Infektionen genützt werden (MALAN U. VAN WYK,
1992), was bei dem so genannten FAMACHA® System angewandt wird.

108
• FAMACHA® System
Das bereits erwähnte FAMACHA® System hingegen basiert auf der klinischen Iden-
tifikation anämischer (= blutarmer) Individuen innerhalb einer Herde und kann durch
den Landwirt selbst erfolgen. Das System basiert auf der Beurteilung der Schleim-
hautfarbe eines Tieres, woraufhin der ermittelte Farbton mit einer Farbtabelle vergli-
chen wird, anhand der man eruieren kann, ob und in welchem Ausmaß eine Anämie
vorliegt (WALLER, 1999; KOOPMANN et al., 2006). Ergebnisse bei Schafen zeigen,
dass diese Methode vergleichbar verlässliche Ergebnisse im Vergleich zum „Gold
Standard", der Erhebung des Hämatokrit, und sogar leicht bessere als die Ermittlung
der Eiausscheidungsrate liefert (VAN WYK AND BATH, 2002, zitiert in VAN WYK et
al., 2006). Allerdings beschränkt sich diese Detektionsmethode ausschließlich auf
mit H. contortus infizierte Tiere, was eine zusätzliche Überwachung anderer, in ge-
mäßigten Klimazonen weit häufigerer Endoparasitosen wie Trichostrongylose, Tela-
dorsagiose und Nematodirus-lnicküonen z.B. anhand regelmäßiger Gewichtskontrol-
le als Parameter der Wirts-„Resilience" etc. unabdingbar macht (BISSET et al., 1991,
zitiert in VAN WYK et al., 2006; WALLER, 1999). Da jedoch in den für diese Arbeit
untersuchten Betrieben die Haemonchose sehr wohl verbreitet ist, wäre eine diesbe-
zügliche Überwachung sicherlich ratsam.
Die in der Regel sehr verlässliche FAMACHA®-Methode zur Detektion und Früher-
kennung einer Haemonchus-lnfekixon ist aufgrund der Einzeltierbeurteilung aller-
dings mit nicht unerheblichem Zeitaufwand verbunden und ist aus diesem Grund
wohl eher nur Kleinbetrieben vorbehalten. Nachdem es sich bei unseren Betrieben
nicht und bei Biobetrieben allgemein selten um Großbetriebe handelt, stellt dieses
System eine zukunftsträchtige Möglichkeit zur Prävention Haemonchus-hedmgiQV
Verluste dar, indem selektiv nur anämische Tiere behandelt werden, was - wie be-
reits beschrieben - nicht zuletzt die Ausbreitung von Resistenzen zu verzögern hilft.

109
6. SCHLUSSFOLGERUNG
Die Parasitenbelastung ist nur einer von vielen Parametern der Tiergesundheit und
muss im Kontext mit dem Allgemeinzustand und der Produktionsleistung der Tiere
gesehen werden. Das Ausmaß der Parasitenbelastung wird zudem von vielen Fakto-
ren moduliert, wovon einer sicherlich die Haltungsform ist. Trotzdem darf der Para-
siten-Aspekt - meiner Meinung nach - in der Diskussion pro/contra Weidegang nicht
in dem Sinne überbewertet werden, dass man ihn als alleinigen Maßstab für die
Tauglichkeit eines Haltungssystems bzw. als einzigen Indikator der Tiergesundheit
darstellt. Statt den Weidegang als eines der Hauptmerkmale der biologischen Wie-
derkäuerhaltung als solchen in Frage zu stellen, sollte sich eine konstruktive Diskus-
sion viel mehr mit der Umsetzbarkeit des Weidegangs, sich daraus ergebenden Risi-
ken, deren Bewältigung aber auch möglichen Chancen und Vorteilen - wie z.B.: e-
ventuell geringeren Produktionskosten- auseinandersetzen.
7. ZUSAMMENFASSUNG
Ziele der Arbeit
Das Hauptanliegen dieser Arbeit war, das Parasitenspektrum und die Befallsintensi-
tät in drei biologischen Milchziegen-Betrieben mit unterschiedlichen Haltungssyste-
men zu erheben. Zusätzlich wurde die Validität des Body Condition Score (BCS) als
Parasitosen-Indikator und seine Praxisrelevanz zur Früherkennung einer Endoparasi-
tose überprüft. Die Untersuchungsergebnisse wurden auf Abhängigkeiten von Hal-
tungssystem, Tieralter sowie auf saisonale Schwankungen untersucht.
Material und Methode
Pro Betrieb (Betrieb 1: Auslauf und Heufütterung, Betrieb 2: Auslauf und Grünfütte-
rung, Betrieb 3: ganzjährige Weidehaltung) wurden im Sommer und Herbst 2007 für
jede Haltungsgruppe fünf bzw. mindestens 20 Einzeltierkotproben rektal sowie
Sammelkotproben aus den Boxen entnommen und parasitologisch mittels Anreiche-
rungsverfahren, Eizählungen und Kotkulturen untersucht. Auffällige Befunde bei der
klinischen Untersuchung der Tiere wurden vermerkt und der Ernährungszustand mit-
tels Body Condition Score (BCS) erhoben. Die Daten wurden mittels SPSS auf signi-
fikante Korrelationen zu Haltungsform, BCS, Alter und Jahreszeit geprüft.

no
Ergebnisse
In allen Betrieben konnten Eimerien, Magen-Darm-Strongyliden (MDS; Haemon-
chus, Teladorsagia, Trichostrongylus, Cooperia, Chabertia), Strongyloides und
Skrjabinema nachgewiesen werden. Nematodirus wurde ausschließlich in Betrieb 3,
Moniezia in Betrieb 1 und 3 beobachtet. Tendenziell konnten typische Weideparasi-
ten wie MDS aber auch Eimerien vermehrt in Betrieb 3 gefunden werden, während
stallassoziierte Parasiten in Betrieb 1 und 2 überwogen. Eine statistisch signifikante
und praxisrelevante Korrelation zwischen BCS und der Befallsintensität konnte nicht
nachgewiesen werden. Altersbedingte Unterschiede zeigten sich in einer höheren
Eimerienoozysten-Ausscheidungsrate bei Jungtieren, sowie höheren Ausscheidungs-
raten von MDS-Eiem bei adulten Tieren. Saisonale Variationen zeigte die Prävalenz
für weidetypische Parasiten, die im Sommer wesentlich höher war, während stallty-
pische Parasiten vermehrt im Herbst zu finden waren. Auffällig war die variierende
Befallsintensität innerhalb einer Herde/Haltungsgruppe, die Hinweis auf die indivi-
duell differierende Immunantwort der Ziegen sein könnte.
8. SUMMARY
PATTISS-KLINGEN, Barbara: Endoparasite burden and management in dairy
goats from organic farms with special reference to farm type and feeding.
Introduction
The increasing number of goat-farms and organic livestock during the last years re-
flect growing economic importance of these production fields. Factors causing pro-
duction losses become even more relevant - gastrointestinal (GI) parasitism can be
seen as one of them. Endoparasites can be present in high numbers especially in or-
ganic livestock - last but not least because of the restricted possibilities to use con-
ventional medicine. The main objective of this study was to determine the endopara-
sitic spectrum and the intensity of infection on three organic dairy goat farms in simi-
lar climatic conditions. Each of them represents a special housing system. The valid-
ity of the body condition score (BCS) as an indicator of endoparasite infections for
early diagnosis was tested additionally. Possible correlations between dependent on
housing system, age and seasonal changes were evaluated.

Ill
Materials and Methods
Three organic milk-goat-farms of different housing systems (farm 1 with outdoor
access and hay-feeding, farm 2 with outdoor access and seasonal forage-feeding,
farm 3 with all-season pasture management) were included in this study and visited
in summer and autumn 2007. The farmers were interrogated previously according to
a questionnaire on general farm dates, livestock husbandry and their endoparasite-
management. Five samples per group of animals or at least 20 samples per farm were
gathered rectally from individuals and additionally pooled faecal samples from the
floor were collected. The samples were examined parasitologically by sedimenta-
tion/flotation, faecal egg counts and coprocultures. Clinical findings of interest were
noted and the BCS was determined assessing sternal- and lumbar region. The ob-
tained data were analysed for significant correlations and differences concerning the
parasite-prevalence considering the housing system, BCS, age and season using
SPSS.
Results
Significant differences between the farms were found for infection level and egg
excretion but not for the parasite-spectrum. Eimeria, strongylid nematodes (Haemon-
chus, Teladorsagia, Trichostrongylus, Cooperia and Chabertia), Strongyloides and
Skrjabinema of varying degrees could be found on all farms; however, Nematodirus
was detected exclusively on farm 3 and Moniezia only sporadically on farm 1 and 3.
Clear differences between farms could be registered concerning typical pasture-
parasites such as strongyles, but also for Eimeria. Farm 3 showed the highest, farm 1
usually the lowest rate of egg/oocyst-excretion, whereas the excretion rates of farm 2
ranged in between. A statistically significant correlation of practical relevance be-
tween BCS and intensity of infection was not evident; hence BCS must be seen as an
inappropriate tool for early diagnosis of endoparasitic infections. However, obvious
significant differences of infection-rate could be documented between adults and
kids (until one year of age), particularly concerning Eimeria - with higher prevalence
in kids and young goats - and strongyles with higher excretion-rates in adults. Fur-
thermore seasonal changes of the parasite-spectrum and intensity of affection could
be found with higher rates of pasture parasites in summer and higher rates of indoor
parasites in autumn. Larvae of Haemonchus and Strongyloides could be predomi-
nantly observed in summer but little in autumn whereas for Teladorsagia, Trichos-

112
trongylus, Chabertia and Cooperia the opposite was noted. Intensity of infection was
highly varying within flocks. This could be explained by different efficiency of indi-
vidual immune response.

113
9. LITERATURVERZEICHNIS
AGUILAR-CABALLERO, A.J., ToRRES-AcosTA, J.F.J., OJEDA-ROBERTOS, N. (2003), Premunization of Criollo kids by use of a single viable dose of Haemonchus con- tortus before natural infection with gastrointestinal nematodes. 19* International Conference of the World Association for the Advancement of Veterinary Parasi- tology, 259.
ATHANASIADOU, S., KYRIAZAKIS, I., JACKSON, P., COOP, R.L. (2001), Direct anthelmintic effects of condensed tannins towards different gastrointestinal nematodes of sheep: in vitro and in vivo studies. Vet. Parasitol. 99, 205-219.
BALIC, A., BOWLES, V.M., MEEUSEN, E.N.T. (2000), The immunobiology of gastro- intestinal nematode infections in ruminants. Adv. Parasitol. 45, 181-241.
BALICKA-RAMISZ, A. (1999), Studies on coccidiosis in goats in Poland. Vet. Parasi- tol. 81, 347-349.
BANG, K.S., FAMILTON, A.S., SYKES, A.R. (1990), Effect of copper oxide wire parti- cle treatment on establishment of major gastrointestinal nematodes in lambs. Res. Vet. Sei. 49, 132-137.
BARGER, I.A., SOTHCOTT, W.H. (1978), Parasitism and production in weaner sheep grazing alternately with cattle. Australian J. Exp. Agr. Anim. Hus. 18, 340-346.
BARGER, I.A., DASH, K.M. (1987), Repeatability of ovine faecal egg counts and blood packed cell volumes in Haemonchus contortus infections. Int. J. Parasitol. 17, 977-980.
BARGER, I.A. (1997), Control by management. Vet. Parasitol. 72, 493-506.
BARGER, I.A. (1999), The role of epidemiological knowledge and grazing manage- ment for helminth control in small ruminants. Int. J. Parasitol. 29, 41-47.
BARRY, T.N., MANLEY, T.R. (1986), Interrelationships between the concentrations of total condensed tannin, free condensed tannin and lignin in Lotus sp. and other possible consequences in ruminant nutrition. J. Sei. Food Agr. 37, 248-254.
BARRY, T.N., MCNABB, W.C. (1999), The implications of condensed tannins on the nutritive value of temperate forages fed to ruminants. Br. J. Nutr. 81, 263-272.
BAUER, C. (2006): Untersuchungsmethoden. In: SCHNIEDER, T. (Hrsg.): Veterinär- medizinische Parasitologie. Parey Verlag, Berlin, 84-104.
BISSET, S.A., VLASSOFF, A., WEST, C.J. (1991): Breeding sheep for resistan- ce/tolerance to internal parasites. Veterinary Continuing Education - Massey University 134, 83-91.
BORGSTEEDE, F.H.M., DERCKSEN, D.P. (1996): Coccidial and helminth infections in goats kept indoors in the Netherlands. Vet. Parasitol. 61, 321-326.
BOWMAN, D.D. (1999): Helminths. In: Georgi's Parasitology for veterinarians, Saunders, St. Louis, 115-244.
BowN, M.D., PoPPi, D.P., SYKES, A.R. (1991): The effect of post ruminal infusion of protein or energy on the pathophysiology of Trichostrongylus colubriformis in- fection and body composition in lambs. Aust. J. Agr. Res. 42, 253-267.

114
BoYAZOGLU, J., HATZIMINAOGLOU, I., MORAND-FEHR, P. (2005): The role of the goat in society: past, present and perspectives for the future. Small Rum. Res. 60, 13-23.
BROOKER, J.D. (Hrsg.) (1999): Tannins in livestock and human nutrition. Proceed- ings of an International Workshop, Adelaide, Australia, May 31-June 2 1999, ACIAR Proceedings Nr. 92, 171.
CABARET, J., GASNIER, N. (1994): Farm history and breeding management influ- ences on the intensity and specific diversity of nematode infection of dairy goats. Vet. Parasitol. 53, 219-232.
CABARET, J. (1986): 167 plantes pour soigner les animaux. Phythotherapie Veterinaire, Ed. Point Veterinaire, Maisons-Alfort, 192.
CABARET, J. (2000): Anthelmintic resistance in goats: from fictions to facts, in: Proc. 7"" Int. Conf. Goats, Tours, 15.-21 Mai 2000, 793-794.
CABARET, J., BOUILHOL, M., MAGE, C. (2002): Managing helminths of ruminants in organic farming. Vet. Res. 33, 625-640.
CHARTIER, C, HOSTE, H. (1997): Response to challenge infection with Haemonchus contortus and Trichostrongylus colubriformis in dairy goats. Differences between high and low producers. Vet. Parasitol. 73, 267-276.
CHARTIER, C, HOSTE, H. (1997a): La therapeutique anthelminthique chez les caprins. Le Point Veterinaire 28, 125-132.
CHARTIER, C, ETTER, E., HOSTE, H., PORS, I., MALLEREAU, M.P., BROQUA, C,
MALLET, S., KOCH, C, MASSE, A. (2000): Effects of the initial level of milk pro- duction and of the dietary protein intake on the course of natural nematode infec- tion in dairy goats. Vet. Parasitol. 92, 1-13.
CHARTIER, C, SOUBIRAC, F., PORS, I., SILVESTRE, A., HUBERT, J., COUQUET, C,
CABARET, J. (2001): Prevalence of anthelmintic resistance in gastrointestinal nematodes of dairy goats under extensive management conditions in southwest- em France. Journal of Helminthology 75, 325-330.
COOP, R.L., KYRIAZAKIS, I. (1999): Nutrition-parasite interaction. Vet. Parasitol. 84, 187-204.
DINEEN, J.K. (1963): Immunological aspects of parasitism. Nature 197, 268-269.
DINEEN, J.K., DONALD, A.D., WAGLAND, B.M., TURNER, J.H. (1965): The dynamics of the host-parasite relationship. Part n. The response of sheep to primary and secondary infection with Nematodirus spathiger. Parasitology 55, 163-171.
DONALDSON, J. (1997): The effect of dietary protein on the establishment and matu- ration of nematode populations in adult sheep. In: BARRELL, G.K. (Hrsg.): Sus- tainable control of internal parasites in ruminants. Christchurch: Lincoln Univer- sity, 193-201.
EADY, S.J., WOOLASTON, R.R., MORTIMER, S.I. (1996): Resistance to nematodes in Merino sheep: sources of genetic variation. Australian Journal of Agricultural Research 47, 895-915.
ECKERT, J., FRIEDHOFF, K.T., ZAHNER, H., DEPLAZES, P. (2005a): Stamm Nemato- zoa. Unterstamm Nematoda. In: Lehrbuch der Parasitologie für die Tiermedizin. Enke Vertag, Stuttgart, 209-338.

115
ECKERT, J., ERmoHOFF, K.T., ZAHNER, H., DEPLAZES, P. (2005b): Diagnostik. In: Lehrbuch der Parasitologie für die Tiermedizin. Enke Verlag, Stuttgart, 491.
ETTER, E., CHARTIER, C, HOSTE, H., PORS, L, BORGIDA, L.P. (1999): The influence of nutrition on the periparturient rise in faecal egg counts in dairy goats: results from a two-year study. Revue de Medicine Veterinaire 150, 975-980.
ETTER, E., HOSTE, H., CHARTIER, C, PORS, I., KOCH, C, BROQUA, C, COUTINEAU, H.
(2000): The effect of two levels of dietary protein on resistance and resilience of dairy goats experimentally infected with Trichostrongylus colubriformis: com- parison between high and low producers. Vet. Res. 31, 247-258.
EYSKER, M. (1997): Some aspects of inhibited development of trichostrongylids in ruminants. Vet. Parasitol. 72, S. 265-283.
EYSKER, M.; BAKKER, N.; KOOYMAN, F.N.J., PLOEGER, H.W. (2005): The possibili- ties and limitations of evasive grazing as a control measure for parasitic gastroen- teritis in small ruminants in temperate climates. Vet. Parasitol. 129, S. 95-104.
GiBBS, H.C. (1986): Hypobiosis and the periparturient rise in sheep. The Veterinary Clinics of North America. Food Animal Practice 2, 345-353.
GILLHAM, R.J., OBENDORF, D.L. (1985): Therapeutic failure of levamisole in dairy goats. Aust. Vet. J. 62, 426-427.
GITHIGIA, S.M., THAMSBORG, S.M., LARSEN, M. (2001): Effectiveness of grazing management in controlling gastrointestinal nematodes in weaner lambs on pas- ture in Denmark. Vet. Parasitol. 99, 15-27.
GöMEZ-RINCöN, C, URIARTE, J., VALDERRABANO, J. (2007): Effect of nemato- phagous Duddingtonia flag runs and energy supplementation on the epidemiology of naturally infected kids. Vet. Res. 38, 141-150.
GR0NVOLD, J., WOLSTRUP, J., NANSEN, P., HENRIKSEN, S.A., LARSEN, M.,
BRESCIANI, J. (1993): Biological control of nematode parasites in cattle with nematode-trapping fungi: a survey of Danish studies. Vet. Parasitol. 48, S 311- 325.
GR0NVOLD, J., NANSEN, P., HENRIKSEN, S.A., LARSEN, M., WOLSTRUP, J., BRESCIANI, J., RAWAT, H., FRIBERT, L. (1996): Induction of traps by Ostertagia ostertagi larvae, chlamydospore production and growth rate in the nematode- trapping fungus Duddingtonia flagrans. Journal of Helminthology 70, 291-297.
HARWOOD, D., MATTHEWS, J. (2008): Goats: medicine and surgery. In: ROBERTS, V., SCOTT-PARK, F. (Hrsg.): BSAVA Manual of Farm Pets, Wiley-VCH, Weinheim, 110-141.
HERTZBERG, H., BAUER, C. (2000): Anthelminthika-Resistenzen bei Magen-Darm- Strongyliden von Schafen und Ziegen: Aktuelles über Verbreitung, Epidemiolo- gie, Vorbeugemaßnahmen und Alternativen zum Anthelminthika-Einsatz. Berli- ner und Münchener Tierärztliche Wochenschrift 113, 122-128.
HÖGLUND, J., SVENSSON, C, HESSLE, A. (2001): A field survey of internal parasites in calves on organic farms in southwestern Sweden. Vet. Parasitol. 99, 113-128.
HOSTE, H., CHARITER, C. (1993): Comparison of the effects on milk production of concurrent infection with Haemonchus contortus and Trichostrongylus colubri- formis in high- and low-producing dairy goats. Am. J. Vet. Res. 54, 1886-1893.

116
HOSTE, H., CHARTffiR, C. (1998): Response to challenge infection with Haemonchus
contortus and Trichostrongylus colubriformis in dairy goats. Consequences on milk production. Vet. Parasitol. 74, 43-54.
HOSTE, H., CHARTER, C. (1998a): Resistance des chevres aux strongyloses gastrointestinale: differences avec les moutons. Point Vet. 29, 69-74.
HOSTE, H., CHARTER, C, ETTER, E., GOUDEAU, C, SOUBIRAC, F., LE FRILEUX, Y.
(2000): A questionnaire survey on the practices adopted to control gastrointestinal nematode parasitism in dairy goat farms in France. Vet. Res. Comm. 24, 459-469.
HOSTE, H., LEVEQUE, H., DORCHES, PH. (2001): Comparison of nematode infections of the gastrointestinal tract in Angora and dairy goats in a rangeland enviroment: relations with the feeding behaviour. Vet. Parasitol. 101, 127-135.
HOSTE, H., LE FRILEUX, Y., POMMARET, A. (2001a): Distribution and repeatability of faecal egg counts and blood parameters in dairy goats naturally infected with gas- trointestinal nematodes. Res. Vet. Sei. 70, 57-60.
HOSTE, H. (2001b): Adaptive physiological processes in the host during gastrointes- tinal parasitism. Int. J. Parasitol. 31, 231-244.
HOSTE, H., LE FRILEUX, Y., POMMARET, A. (2002): Comparison of selective and sys- tematic treatments to control nematode infection of the digestive tract in dairy goats. Vet. Parasitol. 106, 345-355.
HOSTE, H., CHARTER, C, LE FRILEUX, Y. (2002a): Control of gastrointestinal para- sitism with nematodes in dairy goats by treating the host category at risk. Vet. Res. 33, 531-545.
HOSTE, H., LE FREEUX, Y., GOUDEAU, C, CHARTER, C, BROQUA, C, BERGEAUD,
J.P. (2002b): Distribution and repeatability of nematode faecal egg counts in dairy goats: a farm survey and implications for worm control. Res. Vet. Sei. 72, 211-215.
HOSTE, H., CHARTER, C, LE FREEUX, Y., GOUDEAU, C, BROQUA, C, PORS, I., BERGEAUD, J.P., DORCHES, PH. (2002C): Targeted application of anthelmintics to control trichostrongylosis in dairy goats: result from a 2-year survey in farms. Vet. Parasitol. 110, 101-108.
HOSTE, H., LE FRILEUX, Y., POMMARET, A. (2002d): Comparison of selective and
systematic treatments to control nematode infection of the digestive tract in dairy goats. Vet. Parasitol. 106, 345-355.
HOSTE, H., TORRES-ACOSTA, J.F., PAOLINI, V., AGUILAR-CABALLERO, A., ETTER, E.,
LE FREEUX, Y., CHARTER, C, BROQUA, C. (2005): Interactions between nutrition and gastrointestinal infections with parasitic nematodes in goats. Small Rum. Res. 60, 141-151.
HOSTE, H., GAELARD, L., LE FREEUX, Y. (2005a): Consequences of the regular dis- tribution of sainfoin hay on gastrointestinal parasitism with nematodes and milk production in dairy goats. Small Rum. Res. 59, 265-271.
HOSTE, H., RULE, A.C., PREVOT, F., BERGEAUD, J.P., GRISEZ, C, DE LA FARGE, F.,
JACQUET, P., DORCHES, P. (2006): Differences in receptivity to gastrointestinal infections with nematodes in dairy ewes: Influence of age and of the level of milk production. Small Rum. Res. 63, 150-155.

117
HOSTE, H., JACKSON, F., ATHANASIADOU, S., THAMSBORG, S.M., HOSKIN, S.O. (2006a): The effect of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 6, 253-261.
HOSTE, H., TORRES-ACOSTA, J.F.J., AGUILAR-CABALLERO, A.J. (2008): Nutrition- parasite interactions in goats: is immunoregulation involved in the control of gas- trointestinal nematodes?. Parasite Immunol. 30, 79-88.
HuNTLEY, J.F., PATTERSON, M., MACKELLAR, A., JACKSON, F., STEVENSON, L.M., COOP, R.L. (1995): A comparison of the mast cell and eosinophil responses of sheep and goats to gastrointestinal nematode infections. Res. Vet. Sei. 58, 5-10.
JACKSON, F., COOP, R.L. (2000): The development of anthelmintic resistance in sheep nematodes. Parasitology 120, 95-107.
KABASA, J.D., OBUDA-ASBL, J., TER MEULEN, U. (2000): The effect of oral admini- stration of polyethylene glycol on faecal helminth egg counts in pregnant goats grazed on browse containing condensed tannins. Trop. Anim. Health Prod. 32, 73-86.
KAHN, L.P., WATSON, D.L. (2003): Sustainable Control of Internal Parasites of Sheep (SCIPS) - A Summary of recent and Current Research on Control of Internal Parasites of Sheep. University of Sydney, Faculty of Veterinary-Science.
KAMINSKY, R., DUCRAY, P., JUNG, M., CLOVER, R., RUFENER, L., BOUVIER, J.,
WEBER, S.S., WENGER, A., WmLAND-BERCHAUSEN, S., GOEBEL, T., GAUVRY, N.,
PAUTRAT, F., SKRIPSKY, T., FROELICH, O., KOMOIN-OKA, C, WESTLUND, B.,
SLUDER, A., MASER, P. (2008): A new class of anthelmintics effective against drug-resistant nematodes. Nature 452 (7184), 176-180.
KAPLAN, R.M. (2004) Drug resistance in nematodes of veterinary importance: a status report. Trends Parasitol. 20, 477-481.
KETTLE, P.R., VLASSOFF, A., REID, T.C, HORTON, C.T. (1983): A survey of control measures used by milking goat farmers and of anthelmintic resistance on their farms. New Zeal. Vet. J. 31, 139-143.
KNOX, M.R. (1996): Integrated control programs using medicated blocks. In: LE-
JAMBRE, L.F., KNOX, M.R. (Hrsg.): Sustainable control of parasites in small ru- minants. ACIAR, Canberra, 141-145.
KNOX, M.R., TORRES-ACOSTA, J.F.J., AGUILAR-CABALLERO, A.J. (2006): Exploiting the effect of dietary supplementation of small ruminants on resilience and resis- tance against gastrointestinal nematodes. Vet. Parasitol. 139, 385-393.
KOOPMANN, R. (2005): Resistente Magen-Darm-Würmer - Neue Empfehlungen für die Behandlung bei Wiederkäuern. Ökologie und Landbau 136/4, 24-26.
KOOPMANN, R., HOLST, C, EPE, C. (2006): Erfahrungen mit der FAMACHA®-Eye- Colour-Karte zur Identifizierung von Schafen und Ziegen für die gezielte anthelminthische Behandlung. Berl. Münch. Tierärztl. Wochenschr. 119, 436- 442.
KOUDELA, B., BOKOVÄ, A. (1998): Coccidiosis in goats in the Czech Republic. Vet. Parasitol. 76, 261-267.
LAMPKIN, N. (2000): Organic farming in the European Union - overview, policies and perspectives. In: ALLARD, G., DAVID, C, HENNING, J. (Hrsg): Organic Agri- culture INRA. Les coUoques de I'INRA, Paris, 95, 23-35.

118
LANDA-CANSIGNO, E., FRISENDAHL, E., TORRES-ACOSTA, J.F.J., AGUILAR-
CABALLERO, A.J., HOSTE, H., VARGAS-MAGANA, J.J. (2005): Comparing the ef- fect of two sources of energy supplements (molasses or maize) on the resilience and resistance against GI nematodes in browsing goats. In: Proc.20* Conf. WAAVP, New Zealand, October 2005,139.
LARSEN, M. (1999): Biological control of helminths. Int. J. Parasitol. 29, 139-146.
LEATHWICK, D.M., VLASSOFF, A., BARLOW, N.D. (1995): A model for nematodiasis in New Zealand lambs: the effect of drenching regime and grazing management on the development of anthelmintic resistance. Int. J. Parasitol. 25, 1479-1490.
LEEB, C. (2002): Body condition scoring in goats. Goat Veterinary Society Meeting, Swindon, UK.
LENG, R.A. (1991): Optimising herbivore nutrition. In: Ho, Y.W., WONG, H.K., AB-
DULLAH, N., TAJUDDIN, Z.A. (Hrsg.): Recent Advances on the nutrition of herbi- vores. Proc. 3'^'' Int. Symp. Nutrition Herbivores, Pulau Pinang, Malaysia, July 1991, Malaysian Society for Animal Production, UPM, Serdang, Malaysia, 269- 281.
LLOYD, S. (1987): Endoparasitic disease in goats. Goat Vet. Soc. J. 8, 32-39.
LLOYD, S., SOULSBY, E.J.L. (1978): Survey of parasites in dairy goats. Am. J. Vet. Res. 39, 1057-1059.
LUND, V., ALGERS, B. (2003): Research on animal health and welfare in organic farming - a literature review. Livestock Prod. Sei. 80, 55-68.
MAGE, C, BOUILHOL, M., ARCHIMBAUD, E., JARDIN, Y., BUSCATTO, O. (1998): Parasitisme en production d'agneau d'herbe en agriculture biologique. Diagnostic des conduits d'elevage et epidemiologie parasitaire. Institut de I'elevage, Compte Rendu n° 9983220, 33.
MAINGI, N., BJ0RN, H., THAMSBORG, S.M., B0GH, H.O., NANSEN, P. (1996): A sur- vey of anthelmintic resistance in nematode parasites of goats in Denmark. Vet. Parasitol. 66, 53-66.
MAINGI, N., BJ0RN, H., THAMSBORG, S.M., DANGOLLA, A., KYVSGAARD, N.C.
(1996a): A questionnaire survey of nematode parasite control practices on goat farms in Denmark. Vet. Parasitol. 66, 25-37.
MALAN, F.S., VAN WYK, J.A. (1992): The packed cell volume and colour of the con- junctivae as aids for monitoring Haemonchus contortus infestations in sheep. In: Proc. South African Vet. Assoc, Grahamstown, South Africa, 7-10 September, 139.
MCKENNA, P.B. (1985): Diagnosis of gastrointestinal nematode parasitism in goats. Proc. Course Goat Husbandry Med., New Zealand Vet. Assoc, November 1985, Palmerston North, New Zealand, 86-95.
MCKENNA, P.B. (1990): The detection of anthelmintic resistance by faecal egg count reduction test: An examination of some of the factors affecting performance and interpretation. New Zeal. Vet. J. 38, 142-147.
MCKENNA, P.B., ALLAN, CM., TAYLOR, M.J. (1996) The effectiveness of benzimi- dazole-levamisole combination drenches in the presence of resistance to both Benzimidazole and levamisole anthelmintics in New Zealand sheep. New Zeal. Vet. J. 44, 116-118.

119
MICHEL, J.F., CAWTHORNE, R.J.G., ANDERSON, R.M., ARMOUR, J., CLARKSON, M.J., THOMAS, R.J. (1983): Resistance to anthelmintics in Britain: husbandry practices and selective pressure. In: BORGSTEEDE, F.H.M, HENRIKSEN, S.A., OVER, H.J.
(Hrsg.): Facts and Reflections IV. Resistance of parasites to anthelmintics. Cen- tral Veterinary Institute, Lelystad, 41-59.
MICHEL, J.F. (1985): Strategies for the use of anthelmintics in livestock and their implications for the development of drug resistance. Parasitology 90, 621-628.
MIN, B.R., POMROY, W.E., HART, S.P., SAHLU, T. (2004): The effect of short-term consumption of a forage containing condensed tannins on gastro-intestinal nema- tode parasite infections in grazing wether goats. Small Rum. Res. 51, 279-283.
MOLAN, A.L. et al. (2000): Effects of condensed tannins and sesquiterpene lactones extracted from chicory on the viability of deer lungworm larvae. Proc. New Zeal. Soc. Anim. Prod. 60, 26-29.
MORAND-FEHR, P., HERVIEU, J., SANTUCCI, P. (1989): Notation de I'etat corporel: a vos stylos!. La chevre 175, 39-42.
NGUYEN, T.M., BINH, D.V., 0RSKOV, E.R. (2005): Effect of foliages containing con- densed tannins and on gastrointestinal parasites. Anim. Feed Sei. Tec. 121, 77- 87.
NIEZEN, J.H., CHARLESTON, W.A.G., HODGSON, J., MACKAY, A.D., LEATHWICK,
D.M. (1996): Controlling internal parasites in grazing ruminants without recourse to anthelmintics: approaches, experiences and prospects. Int. J. Parasitol. 26, 983- 992.
PAOLINI, V., FRAYSSINES, A.. DE LA FAROE, F., DORCHIES, P.H., HOSTE, H. (2003): Efficacy of condensed tannins on established populations and on incoming larvae of Trichostrongylus colubriformis and Teladorsagia circumcinta in goats. Vet. Res. 34,331-339.
PAOLINI, V., BERGEAUD, J.P., GRISEZ, C, PREVOT, F., DORCHIES, P.H., HOSTE, H.
(2003a): Effects of condensed tannins on goats experimentally infected with Haemonchus contortus. Vet. Parasitol. 113, 253-261.
PAPADOPOULOS, E., ARSENOS, G., SOTIRAKI, S., DELIGL\NNIS, C, LAINAS, T., ZYGOY-
lANNis, D. (2003): The epizootiology of gastrointestinal nematode parasites in Greek dairy breeds of sheep and goats. Small Rum. Res. 47, 193-202.
PEARSON, A.B. (1988): Gastro intestinal parasitism in south island goats. Proceed- ings of the 18th Seminary of the New Zealand Sheep & Beef Cattle Society, New Zealand Veterinary Association, 28-36.
PERRY, B.D., RANDOLPH, T.F. (1999): Improving the assessment of the economic impact of parasitic diseases and of their control in production animals. Vet. Para- sitol. 84, 145-168.
PIETROSEMOLI, S., OLAVEZ, R., MONTILLA, T., CAMPOS, Z. (1999): Empleo de hojas de Neem {Azadirachta indica) en control de nematodos gastrointestinales de bo- vinos a pastoreo. Rev. Fac. Agron. (LUZ) 16, Supl. 1, 220-225.
POMROY, W.E., LAMBERT, M.G., BETTERIDGE, K. (1986): Comparison of faecal strongylate egg counts of goats and sheep on the same pasture. New Zeal. Vet. J. 34, 36-37.

120
POMROY, W.E., CHARLESTON, W.A G. (1989): Failure of young goats to acquire re- sistance to Haemonchus contortus. New Zeal. Vet. J. 37, 23-26.
PRICHARD, R.K., HALL, C.A., KELLY, J.D., MARTIN, I.C, DONALD, A.D. (1980): The problem of anthelmintic resistance in nematodes. Aust. Vet. J. 56, 239-250.
REHBEIN, S., VISSER, M., WINTER, R. (1998): Helminthenbefall bei Ziegen in Deutschland. Berl. Munch. Tierärztl. Wochenschr. Ill, 427-431.
REQUEJO-FERNäNDEZ, J.A., MARTINEZ, A., MEANA, A., ROJO-VAZQUEZ, F.A., Oso- RO, K., ORTEGA-MORA, L.M. (1997): Anthelmintic resistance in nematode parasi- tes from goats in Spain. Vet. Parasitol. 73, 83-88.
RICHARD, S., CABARET, J., CABOURG, C. (1990): Genetic and environmental factors associated with nematode infections of dairy goats in North Western France. Vet. Parasitol. 36, 237-243.
RiNALDi, L., VENEZIANO, V., CRINGOLI, G. (2007): Dairy goat production and the importance of gastrointestinal strongyle parasitism. Trans. R. Soc. Trop. Med. Hyg. 101, 745-746.
ROBERTS, M.G., GRENFELL, B.T. (1991): The population dynamics of nematode in- fections of ruminants: periodic perturbations as a model for management. IMA J. Math. Appl. Med. Biol. 8, 83-93.
RODERICK, S., HOVI, M., SHORT, M. (1999): Animal health and welfare issues in or- ganic livestock farms in the UK: results of a producer survey. Occasional Publi- cation-Brit. Soc. Anim. Sei. 23, 109-112.
SCHNIEDER, T. (2006): Helminthosen der Wiederkäuer. In: SCHNIEDER, T. (Hrsg.): Veterinärmedizinische Parasitologie. Parey Verlag, Berlin, 166-234.
SCHüTTE, A. (1994): 1st Forschung in der Veterinärhomöopathie gerechtfertigt?. Beri. Munch. Tierärztl. Wochenschr. 107, 229-236.
SiLVESTRE, A., CHARTER, C, SAUVE, C, CABARET, J. (2000): Relationship between helminth species diversity, intensity of infection and breeding management in dairy goats. Vet. Parasitol. 94, 91-105.
SMITH, M.C, SHERMAN, D.M. (1994): Digestive System - Helminth Diseases. In: Goat Medicine, Lea&Febiger, Pennsylvania, 321-358.
SMITH, W.D. (1999): Prospects for vaccines of helminth parasites of grazing rumi- nants. Int. J. Parasitol. 28, 17-24.
SOUTHCOTT, W.H., MAJOR, G.W., BARGER, I.A. (1976): Seasonal pasture contamina- tion and availability of nematodes for grazing sheep. Aust. J. Agric. Res. 27, 277- 286.
STEPEK, G., BEHNKE, J.M., BUTTLE, D.J., DUCE, I.R. (2004): Natural plant cysteine proteinases as anthelmintics. Trends Parasitol. 20, 322-327.
SYKES, A.R., GREER, A.W. (2003): Effects of parasitism on the nutrient economy of sheep. Australian J. Exp. Agr. Anim. Hus. 43, 1393-1398.
TEMBELY, S., LAHLOU-KASSI, A.. REGE, J.E.O., MUKASA-MUGERWA, E., ANINDO, D.,
SOVANI, S., BAKER, R.L. (1998): Breed and season effects on the peri-parturient rise in nematode egg output in indigenous ewes in a cool tropical environment. Vet. Parasitol. 77, 123-132.

121
TENTER, A.M. (2006): Protozoeninfektionen der Wiederkäuer. In: SCHNffiDER, T. (Hrsg.): Veterinärmedizinische Parasitologie. Parey Verlag, Berlin, 119-165.
THAMSBORG, S.M., ROEPSTORFF, A., LARSEN, M. (1999): Integrated and biological control of parasites in organic and conventional production systems. Vet. Parasi- tol.84, 169-186.
TORRES-ACOSTA, J.F.J. (1999): Supplementary feeding and the control of gastroin- testinal nematodes of goats in Yucatan, Mexico. PhD of The Royal Veterinary College, University of London.
TORRES-ACOSTA, J.F.J., JACOBS, D.E., AGUILAR-CABALLERO, A., SANDOVAL-
CASTRO, C, MAY-MARTINEZ, M., COB-GALERA, L.A. (2004): The effect of sup- plementary feeding on the resilience and resistance of browsing Criollo kids against natural gastrointestinal nematode infections during the rainy season in tropical Mexico. Vet. Parasitol. 124, 217-238.
VAN WYK, J.A. (2001): Refugia - overlooked as perhaps the most potent factor con- cerning the development of anthelmintic resistance. Onderstepoort J. Vet. Res. 68, 55-67.
VAN WYK, J.A., BATH, G.F. (2002): The FAMACHA®-system for managing hae- monchosis in sheep and goats by clinically identifying individual animals for treatment. Vet. Res. 33, 509-529.
VAN WYK, J.A., HOSTE, H., KAPLAN, R.M., BESIER, R.B. (2006): Targeted selective treatment for worm management - How do we sell rational programs to farmers?. Vet. Parasitol. 139, 336-346.
VATTA, A.F., KRECEK, R.C., LETTY, B.A., VAN DER LINDE, M.J., MOTSWATSWE,
P.W., HANSEN, J.W. (2002): Effect of nematode burden as assessed by means of faecal egg counts on body condition in goats farmed under resource-poor condi- tions in South Africa. Vet. Parasitol. 108, 247-254.
WAGHORN, G., WOODWARD, S.L. (2006): Ruminant contributions to methane and global warming - a New Zealand perspective. In: BHATTI, J.S., LAL, R., APPS,
M.J., PRICE, M.A. (Hrsg.): Climate Change and Managed Ecosystems. Taylor and Francis, Boca Raton, 233-260.
WAGHORN, G. (2007): Beneficial and detrimental effects of dietary condensed tan- nins for sustainable sheep and goat production - Progress and challenges. Animal Feed Science and Technology, doi:10.1016/j.anifeedsci.2007.09.013.
WAGHORN, T.S., LEATHWICK, D.M., CHEN, L.-Y., SKIPP, R.A. (2003): Efficacy of the nematode-trapping fungus Duddingtonia flagrans against three species of gastro- intestinal nematodes in laboratory faecal cultures from sheep and goats. Vet. Parasitol. 118, 227-234.
WALLER, P.J., DASH, K.M., BARGER, I.A., LEJAMBRE, L.F., PLANT, J. (1995): Anthelmintic resistance in nematode parasites of sheep - learning from the Aus- tralian experience. Vet. Rec. 136, 411-413.
WALLER, P.J. (1999): International approaches to the concept of integrated control of nematode parasites of livestock. Int. J. Parasitol. 29, 155-164.
WALLER, P.J. (2003): Global perspectives on nematode parasite control in ruminant livestock: the need to adopt alternatives to chemotherapy, with emphasis on bio- logical control. Anim. Health Res. Rev. 4, 35-43.

122
WALLER, P.J., THAMSBORG, S. M. (2004): Nematode control in „green" ruminant production systems. Trends Parasitol. 20, 493-497.
WALLER, P.J. (2006): From discovery to development: Current industry perspectives for the development of novel methods of helminth control in livestock. Vet. Para- sitol. 139, 1-14.
WALLER, P.J. (2006a): Sustainable nematode parasite control strategies for ruminant livestock by grazing management and biological control. Anim. Feed Sei. Tec. 126, 277-289.
WOLF, R. (in Veröffentlichung): Validierbarkeit des manuellen BCS mittels Maß- band, Körperwaage und Ultraschall bei Milchziegen und Abhängigkeit des BCS von Laktationsstadium und Fütterung. Diplomarbeit an der Veterinärmedizini- schen Universität Wien.
ZAJAC, A.M. (2006): Gastrointestinal Nematodes of Small Ruminants: Life Cycle, Anthelmintics and Diagnosis. Vet. Clin. North Am. Food Anim. Pract. 22, 529- 541.
Gesetze und Verordnungen:
Verordnung (EWG) Nr. 2092/91 des Rates vom 24. Juni 1991 über den ökologischen Landbau und die entsprechende Kennzeichnung der landwirtschaftlichen Erzeugnisse und Lebensmittel (Amtsblatt der Europäischen Gemeinschaften Nr. L 198/1 vom 22. Juli 1991) - konsolidierte Fassung, 61. ergänzte Auflage vom 15. Januar 2007.

123
10. ANHANG
Fragebogen Bio-Ziegen-Betriebe
Fragen zum Betrieb allgemein;
1. Wie viele Tiere sind zurzeit auf dem Betrieb? Anteilig wie viele pro Alters- gruppe (Adulte/Jährlinge/Kitze)?
2. Welches Produktionsziel (Fleisch, Milch) wird verfolgt?
3. Besteht Kontakt zu Tieren anderer Betriebe (Ausstellungen...)/ Zukaufe? Wenn ja, woher wird zugekauft? Kommen neue Tiere in „Quarantäne", wenn ja, wie lange und wie ist die Vorgehensweise bzgl. potentieller Parasitosen?
4. Fütterung?
5. Aufstauung: wie oft wird gemistet, neu eingestreut, desinfiziert, grundgerei- nigt...?
6. In welchem Alter werden die Kitze von ihren Müttern getrennt? Wie und wo erfolgt die Kitzaufzucht?
7. Haben die Tiere die Möglichkeit der Nutzung von Auslauf/Weide? Ab und bis wann im Jahr (Weide)? Wird ein Weidewechsel durchgeführt? Wenn ja, wie oft?
8. Werden Weide/ Futterwiesen mit Stallmist gedüngt? Wie erfolgt die Mistauf- bereitung?
Fragen zu Parasitosen:
1. Sind Parasitosen im Betrieb bekannt? Wenn ja, welche? Worauf wird im Zu- sammenhang mit der Erkennung von Parasiten besonders geachtet bzw. wel- che Anzeichen erwecken den Parasitenverdacht?
2. Zeigen einzelne Tiere Auffälligkeiten wie Gewichtsverlust, Durchfall, Hus- ten, Niesen...? Wenn ja, sind bestimmte Altersgruppen davon betroffen? Tre- ten diese Auffälligkeiten zu bestimmten Jahreszeiten auf? Wie hoch ist Ster- berate bei Kitzen?
3. Wie sieht das Parasitenmanagement des Betriebes aus: Prophylaxe, Behand- lungen (Präparate und Dosierung), Haltungsmanagement (häufigeres Ausmis- ten, Desinfizieren, Fütterung, Weide Wechsel...)? Konnte die Situation durch bereits bestehende Maßnahmen verbessert werden?
Related Documents