Dental topography and diets of Australopithecus afarensis and early Homo Peter Ungar* Department of Anthropology, University of Arkansas, Old Main 330, Fayetteville, AR 72701, USA Received 14 November 2003; accepted 22 March 2004 Abstract Diet is key to understanding the paleoecology of early hominins. We know little about the diets of these fossil taxa, however, in part because of a limited fossil record, and in part because of limitations in methods available to infer their feeding adaptations. This paper applies a new method, dental topographic analysis, to the inference of diet from fossil hominin teeth. This approach uses laser scanning to generate digital 3D models of teeth and geographic information systems software to measure surface attributes, such as slope and occlusal relief. Because it does not rely on specific landmarks that change with wear, dental topographic analysis allows measurement and comparison of variably worn teeth, greatly increasing sample sizes compared with techniques that require unworn teeth. This study involved comparison of occlusal slope and relief of the lower second molars of Australopithecus afarensis (n=15) and early Homo (n=8) with those of Gorilla gorilla gorilla (n=47) and Pan troglodytes troglodytes (n=54). Results indicate that while all groups show reduced slope and relief in progressively more worn specimens, there are consistent differences at given wear stages among the taxa. Early Homo shows steeper slopes and more relief than chimpanzees, whereas A. afarensis shows less slope and relief than any of the other groups. The differences between the two hominin taxa are on the same order as those between the extant apes, suggesting similar degrees of difference in diet. Because these chimpanzees and gorillas differ mostly in fallback foods where they are sympatric, results suggest that the early hominins may likewise have differed mostly in fallback foods, with A. afarensis emphasizing harder, more brittle foods, and early Homo relying on tougher, more elastic foods. 2004 Published by Elsevier Ltd. Keywords: Diet; Molars; Hominins; Dental functional morphology Introduction Diet is an important key to understanding the ecology of living primates (Fleagle, 1999). It should be expected, then, that changes in feeding adaptations played an important role in the origins and early evolution of our own tribe and genus. It is difficult to determine the extent of this role, however, because of our limited knowledge of early hominin diets. This is due in part to a limited fossil record, and in part to limitations in methods available to infer dietary adaptations from the * Corresponding author. Tel.: +1-479-575-6361; fax: +1-479-575-6595 E-mail address: [email protected] (P. Ungar). Journal of Human Evolution 46 (2004) 605–622 0047-2484/04/$ - see front matter 2004 Published by Elsevier Ltd. doi:10.1016/j.jhevol.2004.03.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dental topography and diets of Australopithecus afarensisand early Homo
Peter Ungar*Department of Anthropology, University of Arkansas, Old Main 330, Fayetteville, AR 72701, USA
Received 14 November 2003; accepted 22 March 2004
Abstract
Diet is key to understanding the paleoecology of early hominins. We know little about the diets of these fossil taxa,however, in part because of a limited fossil record, and in part because of limitations in methods available to infer theirfeeding adaptations. This paper applies a new method, dental topographic analysis, to the inference of diet from fossilhominin teeth. This approach uses laser scanning to generate digital 3D models of teeth and geographic informationsystems software to measure surface attributes, such as slope and occlusal relief. Because it does not rely on specificlandmarks that change with wear, dental topographic analysis allows measurement and comparison of variably wornteeth, greatly increasing sample sizes compared with techniques that require unworn teeth. This study involvedcomparison of occlusal slope and relief of the lower second molars of Australopithecus afarensis (n=15) and early Homo(n=8) with those of Gorilla gorilla gorilla (n=47) and Pan troglodytes troglodytes (n=54). Results indicate that while allgroups show reduced slope and relief in progressively more worn specimens, there are consistent differences at givenwear stages among the taxa. Early Homo shows steeper slopes and more relief than chimpanzees, whereas A. afarensisshows less slope and relief than any of the other groups. The differences between the two hominin taxa are on the sameorder as those between the extant apes, suggesting similar degrees of difference in diet. Because these chimpanzees andgorillas differ mostly in fallback foods where they are sympatric, results suggest that the early hominins may likewisehave differed mostly in fallback foods, with A. afarensis emphasizing harder, more brittle foods, and early Homo relyingon tougher, more elastic foods.� 2004 Published by Elsevier Ltd.
Keywords: Diet; Molars; Hominins; Dental functional morphology
Introduction
Diet is an important key to understanding theecology of living primates (Fleagle, 1999). It
should be expected, then, that changes in feedingadaptations played an important role in the originsand early evolution of our own tribe and genus.It is difficult to determine the extent of this role,however, because of our limited knowledge ofearly hominin diets. This is due in part to a limitedfossil record, and in part to limitations in methodsavailable to infer dietary adaptations from the
* Corresponding author. Tel.: +1-479-575-6361; fax:+1-479-575-6595
E-mail address: [email protected] (P. Ungar).
Journal of Human Evolution 46 (2004) 605–622
0047-2484/04/$ - see front matter � 2004 Published by Elsevier Ltd.doi:10.1016/j.jhevol.2004.03.004

fossils we do have. The study described here exam-ined early hominin dental functional morphologyusing a new technique, dental topographic analy-sis. Results suggest that Australopithecus afarensismolars have less occlusal slope and relief thanchimpanzees or gorillas, whereas early Homomolars have values intermediate between those ofthe two extant apes. This implies differences in dietbetween the hominins, with early Homo having thepotential to consume tough, deformable foods(i.e., those that resist crack propagation) moreefficiently than could A. afarensis.
Background
Reconstructions of fossil primate diets are oftenbased on studies of tooth shape. Researchers haverecognized form–function relationships betweendental morphology and diet for a long time, andhave focused much attention on unraveling theserelationships (e.g., Owen, 1840–1845; Gregory,1922; Crompton and Sita-Lumsden, 1970; Kayand Hiiemae, 1974; Kay, 1975, 1977, 1978;Rosenberger and Kinzey, 1976; Maier, 1977, 1984;Kay and Hylander, 1978; Kinzey, 1978; Seligsohnand Szalay, 1978; Lucas, 1979). It is clear, forexample, that primate folivores and insectivoreshave reciprocally concave, highly crested teethadapted for shearing, whereas frugivores and hard-object specialists have rounder, flatter teethadapted for crushing and grinding.
Kay and colleagues (Kay, 1978; Kay andHylander, 1978; Kay and Covert, 1984) haverecognized that comparisons of occlusal formbetween species require a quantitative approach.Their shearing quotient (SQ), for example, iscalculated as a residual from a regression of thesummed length of mesiodistal crests over thelength of the occlusal table for a series of closelyrelated species with a given type of diet. The higherthe SQ, the longer the shearing crests relative toexpectation of a species with that diet. The lowerthe SQ, the shorter the shearing crests relative toexpectation. Comparative studies have confirmedthat folivores and insectivores have higher SQsthan closely related frugivores, and that amongfrugivores, hard-object specialists have the lowestSQ values (Anthony and Kay, 1993; Strait, 1993;
Meldrum and Kay, 1997). Studies of SQ have beenapplied to a wide range of fossil primates, leadingto important dietary inferences for many extinctspecies (e.g., Kay, 1977; Kay and Simons, 1980;Anthony and Kay, 1993; Strait, 1993; Williamsand Covert, 1994; Ungar and Kay, 1995; Fleagleet al., 1996; Meldrum and Kay, 1997; Benefit,2000).
While this approach has brought new insightsregarding the diets of some fossil primate taxa,methodological limitations have prevented themfrom being broadly applied to infer early hominindiets. The problem is that shearing crests aremeasured as distances between cusp tips and thenotches between cusps. Crest lengths change withattrition, and quickly become impossible tomeasure as cusp tips are obliterated by wear. Thus,SQ studies are limited to unworn teeth. Few, ifany, early hominin taxa are well enough repre-sented by unworn teeth to allow statistical com-parisons with extant baseline series. There are, forexample, fewer than 10 unworn M2s in the entirecollection of published australopith teeth fromSouth Africa (Ungar et al., 1999). The picturefor early Homo is even bleaker, with no avail-able unworn M2s of H. habilis, H. rudolfensis, orAfrican H. erectus.
Despite these limitations, there are intimationsin the literature that early hominins differed fromone another in functional aspects of tooth shape(Kay, 1985). Grine (1981, 1984), for example,suggested that Australopithecus africanus cheekteeth have more occlusal relief than do those ofParanthropus robustus. This is consistent with thelimited SQ results available (Ungar et al., 1999).In the latter study, the species average SQ for“gracile” australopith M2s was found to be higherthan the “robust” australopith average SQ value,though mean values for both taxa were lower thanthose of extant apes in the comparative baselineseries. This suggests that neither species was well-suited to processing tough, deformable foods, andthat P. robustus teeth would have been particularlyadept at crushing brittle, inelastic items that areless resistant to crack propagation.
No SQ analyses have been published for anyother early hominin species (again, in large partbecause of a lack of sufficient unworn teeth for
P. Ungar / Journal of Human Evolution 46 (2004) 605–622606

analysis). Nevertheless, significant variationamong taxa would be expected given apparentdifferences in gross morphology. As Teaford andcoauthors (Teaford et al., 2002: 153) recentlywrote, “while it appears that H. habilis and H.rudolfensis had somewhat more occlusal relief thandid the australopiths, this difference is difficultto assess given current methods for quantifyingocclusal relief” (Fig. 1).
Dental topographic analysis
Dental topographic analysis offers a newapproach to the quantification of occlusal relief
that allows consideration of both worn andunworn teeth (Ungar and Williamson, 2000;M’Kirera and Ungar, 2003; Ungar and M’Kirera,2003; Dennis et al., in press). A laser scanner isused to generate 3D points along the surface of atooth, and these points are used in a geographicinformation system (GIS) to model and character-ize that surface as a whole. Dental topographicanalysis has the advantage over traditional SQstudies in that measurements do not rely onspecific landmarks that erode with the onset ofwear.
Recent studies of extant primate dental top-ography have shown this approach to be an
Fig. 1. Australopithecus afarensis, AL 400-1a (top), and early African Homo erectus, KNM-WT 15000 (bottom), mandibular cheekteeth. Image copyrights belong to the National Museums of Ethiopia (AL 400-1a) and National Museums of Kenya (KNM-WT15000). Specimens are scaled to approximately the same size.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 607

effective way to model and compare the shapes ofvariably worn molar teeth (Ungar and Williamson,2000). Scoring teeth by wear stage allows directcomparisons of shape data among similarly wornspecimens of different species. Such attributes asaverage surface slope, occlusal relief, and surfaceangularity or jaggedness have all been shown todiffer between chimpanzees and gorillas when con-trolling for degree of gross tooth wear (M’Kireraand Ungar, 2003; Ungar and M’Kirera, 2003; seebelow). Further, a longitudinal study of changes intooth shape in Alouatta palliata has shown thatindividuals of that species wear their teeth down insimilar ways (Dennis et al., in press). Assuming thepattern holds for other taxa, this suggests thatvariably worn teeth of different individuals can beused to represent a species, a necessary prerequisitefor studies of tooth shape changes with wear forfossil taxa.
Paleontological applications
While most early hominin species are notrepresented by enough unworn specimens of agiven tooth type for SQ analysis, there are suf-ficient numbers of worn specimens of at least sometaxa for dental topographic analysis. Australo-pithecus afarensis presents such a case. This taxonis one of the best represented Pliocene hominins,with well-dated specimens ranging from about3.7 Ma at Laetoli to 3.0 Ma at Hadar (Drake andCurtis, 1987; Kimbel et al., 1994; Lockwood et al.,2000; see also Ward et al., 1999).
Australopithecus afarensis has been thought tohave played a central role in early human evolu-tion since it was first recognized (Johanson andWhite, 1979), with numerous researchers plac-ing this species on the line leading ultimately tomodern humans (see Strait et al., 1997, for review).While recent finds have led some to favor a morecomplex hominin phylogenetic tree featuringuncertain ancestor-descendant relationships (e.g.,Leakey et al., 2001; Lieberman, 2001), A. afarensisremains important to studies of the evolution ofhuman diet. Its occlusal morphology is conserva-tive for early hominins, and its cusps lack theswollen, inflated appearance often seen in later
australopiths (White et al., 1981). This speciestherefore offers a useful model for a generalizedaustralopith from which the occlusal morphologyof later hominins, including early Homo, mighthave been derived.
Studies of occlusal form in early African Homospecies (H. rudolfensis, H. habilis, and H. erectus)might likewise provide new insights into the rolethat diet played in the origin and early evolution ofour genus. As with earlier Pliocene hominins,temporal overlap between early Homo speciesmakes it difficult to determine which may havegiven rise to modern humans. Fossils attributableto H. habilis and H. rudolfensis evidently date tobetween about 2.4 and 1.65 Ma (Wood, 1991,1992, 1999; Hill et al., 1992; Bromage et al., 1995;Kimbel et al., 1996; Sherwood et al., 2002). Thesetwo species overlap with African H. erectus nearthe end of their temporal ranges, with the latterdating from at least 1.89 Ma to 660 ka (Wood andVan Noten, 1986; Feibel et al., 1989).
It is not possible at this time to evaluate form–function relationships for individual species ofearly Homo because of very limited sample size(see Wood, 1991). We might, however, combineH. habilis, H. rudolfensis, and early African H.erectus molars into a single early Homo samplefor functional study. The justification for doingso is their evident closer evolutionary relation-ships with one another than with any knownaustralopith species (Strait and Grine, 2001).While there are no unworn M2s available forearly Homo, there are enough worn molars toevaluate differences between this taxon and earlieraustralopiths.
The study described here is the first quantitativeanalysis of worn early hominin occlusal mor-phology. A new technique, dental topographicanalysis, is applied to available Australopithecusafarensis and early Homo M2s to examine func-tional aspects of tooth form. Results for thesefossil taxa are compared both to one another andto data from an extant baseline series of chimpan-zee and gorilla M2s. Documented differences inmorphology, both between the early hominins andbetween the fossil taxa and the extant apes, offernew insights into the dietary adaptations of bothA. afarensis and early Homo.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622608

Materials and methods
Specimens used in this study
High resolution replicas of lower second molars(M2s) were prepared from original teeth of bothextant and fossil specimens following conventionalprocedures. These teeth were chosen becauseSQ studies have shown that M2 occlusal mor-phology is very effective for distinguishinghominoids on the basis of their diets (Kay, 1977).Molds of undamaged occlusal surfaces were madeusing President’s Jet (Coltene-Whaledent Corp.,Mawah, NJ) regular-body polyvinylsiloxanedental impression material. Casts were pouredusing Epotek 301 (Epoxy Technologies, Inc.,Billerica, MA) epoxy and centrifuged prior tohardening to remove bubbles that would otherwiseobscure occlusal morphology. Because laserscanners tend to have difficulty identifying surfacesof translucent epoxy, a pale pink pigment wasadded to the mix, and replicas were coated witha thin layer of Magniflux SKD-S2 Developer(Illinois Tool Works, Inc, Glenview, IL).
All available M2s of Australopithecus afarensis,Homo rudolfensis, H. habilis, and H. erectushoused at the National Museum of Ethiopia,National Museum of Kenya, National Museum ofTanzania, Transvaal Museum, and the Universityof the Witwatersrand Department of Anatomywere considered. Most specimens had to beexcluded from analysis because taphonomicdamage to the crown would affect measurementsof surface topography. The list of fossil specimensincluded in this study is presented in Table 1 (n=15for A. afarensis and n=8 for early Homo).
The comparative baseline series consisted ofvariably worn M2s of wild-shot Gorilla gorillagorilla (n=47) and Pan troglodytes troglodytes(n=54) housed at the Cleveland Museum ofNatural History. These taxa were chosen for com-parison with the fossil species because these apesare closely related to the hominins, and havesimilar molar cusp patterns. Kay and Ungar (1997)note the importance of phylogenetic control whenapplying the comparative method to infer dietfrom molar morphology. As Kay and Covert(1984) have shown, for example, fundamental
differences in tooth form between cercopithecoidsand hominoids prevent direct comparison of thesetaxa. Likewise, autapomorphic traits can evenmake it difficult to compare closely related taxa.Such is the case with orangutans, which often havecrenulations likely to affect average occlusal slopeand relief, as well as morphological changes withmolar wear.
On the other hand, chimpanzees and gorillasare appropriate for comparisons with early hom-inins. While early hominins probably did not eatthe same exact foods consumed by living Africanapes, degrees of difference in occlusal morphologycan be compared between taxa, potentially offeringnew insights into differences in mechanical proper-ties of foods eaten by the fossil taxa (see Spearsand Crompton, 1996; Kay and Ungar, 1997).
Gorilla gorilla gorilla and Pan troglodytestroglodytes are particularly useful baseline taxa forcomparison because of the modest degree to whichthey differ in the material properties of the foodsthey consume. At sites where the two taxa aresympatric, such as Lope, Gabon, chimpanzees andgorillas overlap considerably in their diets, but
Table 1Specimens used in this analysis (all M2s)
AL 128-23 A. afarensisAL 145-35 A. afarensisAL 188-1 A. afarensisAL 198-1 A. afarensisAL 207-13 A. afarensisAL 241-14 A. afarensisAL 266-1 A. afarensisAL 330-5 A. afarensisAL 333w-1a A. afarensisAL 333w-27 A. afarensisAL 333w-57 A. afarensisAL 333w-60 A. afarensisAL 400-1a A. afarensisAL 417-1a A. afarensisLH 23 A. afarensisKNM-ER 806 H. erectusKNM-ER 992 H. erectusKNM-ER 1506 H. rudolfensisKNM-ER 1802 H. rudolfensisKNM-ER 3734 Homo indet.KNM-WT 15000 H. erectusOH 16 H. habilisOH 22 H. erectus
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 609

they also differ, particularly at times of fruit scar-city. At such “crunch” times, gorillas fallback ontougher, more fibrous plant parts, such as leavesand stems, whereas chimpanzees continue toexploit available ripe, succulent fruits (Tutin et al.,1991; Remis, 1997). Differences in their molartopography reflect these differences in their diets(M’Kirera and Ungar, 2003; Ungar and M’Kirera,2003).
Data acquisition and analysis
Data acquisition and analysis followed thedental topographic approach described in detailelsewhere (M’Kirera and Ungar, 2003; Ungar andM’Kirera, 2003; Dennis et al., in press). First,molar replicas were mounted on a horizontal plat-form approximating the occlusal plane. Specimenswere then rotated about the mesiodistal andbuccolingual axes using setscrews to maximize theocclusal surface in top view. Repeatability studieshave shown most measurements of specimensoriented in this manner to be precise to within onepercent (Ungar et al., 2002).
Specimens were then scanned using a modifiedSurveyor 500 laser scanner (Laser Design, Inc.,Minneapolis, MN) with an RPS 150 rapid profilesensor. This instrument collects elevation data fora surface at fixed intervals in an x,y plane. Thelateral and vertical resolutions of the instrumentwere both set to 25.4 µm for this study, producinga matrix of 1550 evenly spaced elevations, orz-values, for each 1 mm2 in the horizontal plane.Elevations were collected using two offset detec-tors to avoid missing data resulting from shadow-ing effects of steep cusps on adjacent low points.Resulting data were output as ASCII files contain-ing lists of x,y,z coordinates representing theocclusal surface of each specimen.
These ASCII files were opened as data tables inArcView 3.2 (ESRI Corp., Redlands, CA) withSpatial Analyst and 3D Analyst extensions. Toothsurfaces were interpolated using inverse distanceweighting and cropped to include only the occlusaltable (defined as the surface above a horizontalplane intersecting the lowest point on the talonidbasin when the tooth is oriented as describedabove [Ungar and Williamson, 2000]). Average
slope (in degrees) between adjacent elevations wasthen calculated for each surface. Further, ArcViewwas used to create a triangulated irregular network(TIN), which is a series of triangles connectingadjacent data points on each surface. The surfaceareas of these triangles were tallied to generate a3D surface area value for the specimen. This valuewas then divided by the projected horizontal or2D planimetric area calculated from the x,y pointdata and multiplied by 100 to give a dimensionlessindex of occlusal relief. Relief data includedhere for the chimpanzees and gorillas were firstreported by M’Kirera and Ungar (2003).
Finally, molar replicas were scored for grosswear using Scott’s (1979) method. This method,commonly used with bioarchaeological samples(see Rose and Ungar, 1998), has also proven to bea repeatable, effective approach to characterizingmolar wear of extant hominoids (M’Kirera andUngar, 2003; Ungar and M’Kirera, 2003). Scoresbased on degree of faceting or dentine exposure ofindividual cusps were summed as described byScott (1979). Specimens were arranged into sixwear stages based on their Scott score values: 1)5–9; 2) 10–14; 3) 15–19; 4) 20–24; 5) 25–29; 6)30–34.
Both average slope and relief data were ana-lyzed using SYSTAT 7.0 (Systat Software Inc.,Richmond, CA). Data were rank-transformed tomitigate violation of assumptions required ofparametric tests (Conover and Iman, 1981), andtwo-way ANOVAs with taxon and wear score asfactors were conducted for slope and occlusalrelief. This allowed assessment of the effects oftaxon and degree of wear on each model, as well asthe effects of interactions between the two factors.Bonferroni pairwise multiple-comparisons testswere used to determine sources of significant vari-ation for taxon and wear stage differences. Asstated above, small sample sizes for Homo habilis,H. rudolfensis, and African H. erectus made com-parisons of these individual species impractical, sothey were combined into a single early Homosample for analysis.
It should be noted that combining the earlyHomo sample probably conflates dietary variationamong individual species. Wood and Collard(1999) suggested, for example, that Homo habilis
P. Ungar / Journal of Human Evolution 46 (2004) 605–622610

and H. rudolfensis retained large australopith-sizemolar teeth, and thus had a “mechanically moredemanding” diet compared with H. erectus. Whilethese assertions concerning tooth size have beenchallenged (McHenry and Coffing, 2000), and thefunctional significance of molar size differencesamong higher primates remains unclear (seeUngar, 1998), it would be best to consider speciesindividually if sufficient samples of each becomeavailable in the future.
The approach presented here compares earlyhominin and extant ape specimens with similardegrees of faceting and dentine exposure. Statisti-cal analyses were limited to specimens at wearstages represented by all species (2–4). The fossilhominin specimens all fell within wear stages 1–4,whereas the extant hominoids ranged from stages2–5. This lack of complete overlap evidentlyreflects differences between taxa in enamel thick-ness. Not withstanding methodological issues con-cerning the characterization of molar enamelthickness, researchers generally agree that early
hominin molars have thicker enamel than do thoseof chimpanzees or gorillas (e.g., Grine and Martin,1988). The approach presented here allows com-parison of dental morphology between earlyhominin and extant ape specimens with similardegrees of faceting and dentine exposure. Still,given that enamel thickness can influence rate ofwear (Dean et al., 1992), comparisons of specimensat a given wear stage does not necessarily implycomparisons of individuals at similar ages.
Results
Results are presented in Figs. 2 and 3 andTables 2–4. While sample sizes for the fossilhominin taxa are small, two distinct patterns areevident. First, as with the chimpanzees andgorillas, Homo and Australopithecus afarensis eachshow decreases in both average occlusal surfaceslope and relief through successive wear stages. Inother words, molar teeth of all four taxa became
Fig. 2. Comparisons of average surface slope values for given wear stages. Wear stages correspond to Scott scores as follows: 2) 10–14,3) 15–19, 4) 20–24 (see text).
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 611

progressively flatter as they wore. Second, differ-ences between early Homo and A. afarensis andbetween each hominin and the extant ape taxa arefairly consistent for both variables at given stagesof wear. Gorilla gorilla specimens tend to havethe steepest slopes and most relief, followed byearly Homo, then Pan troglodytes, and finally A.afarensis, which has the flattest slopes with theleast relief (Fig. 4). Early Homo slope and reliefmean values are greater than those of A. afarensisin seven of eight cases (see Table 2). The slope andrelief mean values for A. afarensis are smaller thanthose of G. gorilla in seven of eight cases, andsmaller than those of P. troglodytes in all six cases.Finally, early Homo slope and relief mean valuesare smaller than those of G. gorilla in all six casesand larger than those of P. troglodytes in five of sixcases.
Despite the small sample sizes, statisticalanalyses yielded results consistent with these
observations. First, factorial ANOVAs on theranked slope and ranked relief data both evincedsignificant variation in the models (Table 3). Thereis significant variation among the taxa and amongthe wear stages for both slope and relief models.Further, neither model showed a significant inter-action between the two factors. In other words,degrees of difference between the taxa do notchange significantly at different wear stages—differences between gorillas, chimpanzees, earlyHomo, and Australopithecus afarensis evidentlyremain consistent through the wear sequence.
Bonferroni multiple comparisons tests revealthe sources of significant variation. Comparingvalues among wear stages, the second wear stagehas significantly steeper average slopes andrelief than the fourth wear stage. In addition, thethird wear stage shows significantly steeper slopesand marginally more relief than the fourth wearstage. Comparing values among taxa, as already
Fig. 3. Comparisons of mean relief values for given wear stages. Wear stages correspond to Scott scores as follows: 2) 10–14, 3) 15–19,4) 20–24 (see text).
P. Ungar / Journal of Human Evolution 46 (2004) 605–622612

demonstrated elsewhere (M’Kirera and Ungar,2003; Ungar and M’Kirera, 2003), Gorilla gorillaM2s have significantly steeper slopes and moreocclusal relief than those of Pan troglodytes.Furthermore, early Homo molars have signifi-cantly less occlusal relief than those of G. gorillaand marginally significantly steeper slopes thanthose of Australopithecus afarensis. Finally, A.
afarensis has significantly lower slope values andless occlusal relief than G. gorilla.
In sum, despite small sample sizes, there aretrends evident in the data. First, there is a tendencyfor the molar teeth of all four taxa to becomeflatter with wear. Second, differences among thetaxa remain about the same at all observed stagesof wear. Gorilla M2s have the steepest slopes with
Table 2Summary Statistics
Wear Australopithecus Gorilla Homo Pan
Mean SD N Mean SD N Mean SD N Mean SD N
A. Slope1 37.89 – 1 – – – 37.92 – 1 – – –2 32.01 6.154 4 37.75 5.036 7 36.5o 4.677 2 32.88 5.859 53 25.46 3.810 7 36.29 2.665 10 35.83 9.519 3 30.15 5.771 284 25.21 2.450 2 32.13 5.069 14 26.52 3.296 2 26.48 4.680 185 – – – 27.53 4.290 13 – – – 25.69 8.607 26 32.05 – 1 32.14 6.445 3 – – – 29.31 – 1
B. Relief1 150.61 – 1 – – – 152.14– 1 – – –2 139.34 6.206 4 163.86 9.411 7 154.06 18.627 2 143.59 7.774 53 129.29 13.641 7 164.88 9.279 10 143.08 8.770 3 139.4 10.701 284 135.30 9.155 2 152.51 14.309 14 131.86 10.074 2 136.00 10.023 185 – – – 142.35 6.734 13 – – – 125.71 7.027 26 169.90 – 1 139.39 3.619 3 – – – 148.05 – 1
Table 3Analysis of slope data (data rank transformed)
A. Factorial ANOVAEffect SS df MS F pTaxon 23349 3 7783 8.607 0.000Wear class 15207 2 7603 8.409 0.000Interaction 3708 6 618 0.683 0.663Error 81385 90 904
B. Bonferroni multiple comparison testsTaxon Australopithecus Gorilla HomoGorilla 52.295b
Homo 34.493a �17.802Pan 19.172 �33.123b �15.321
Wear stage 2 33 �17.6454 �41.358b �23.713b
ap<0.10.bp<0.05.
Table 4Analysis of relief data (data rank transformed)
A. Factorial ANOVAEffect SS df MS F pTaxon 47217 3 15739 20.040 0.000Wear class 6585 2 3292 4.192 0.018Interaction 3912 6 652 0.830 0.550Error 70685 90 785
B. Bonferroni multiple comparison testsTaxon Australopithecus Gorilla HomoGorilla 52.342b
Homo 18.837 �33.505b
Pan 9.555 �42.787b �9.282
Wear stage 2 33 �13.3974 �28.144b �14.747a
ap<0.10.bp<0.05.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 613
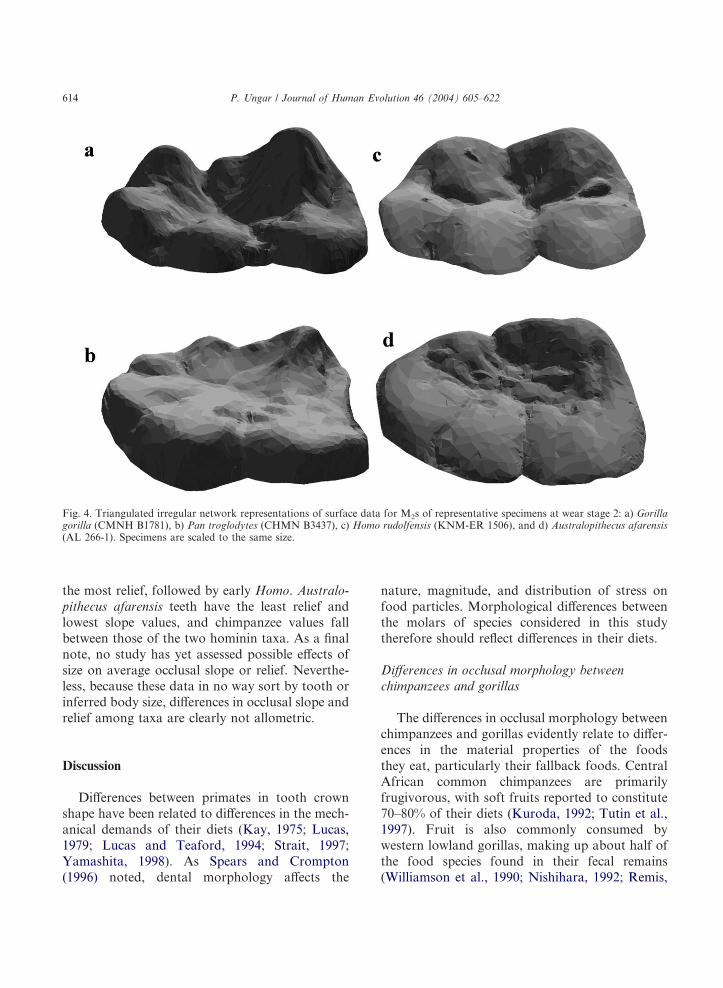
the most relief, followed by early Homo. Australo-pithecus afarensis teeth have the least relief andlowest slope values, and chimpanzee values fallbetween those of the two hominin taxa. As a finalnote, no study has yet assessed possible effects ofsize on average occlusal slope or relief. Neverthe-less, because these data in no way sort by tooth orinferred body size, differences in occlusal slope andrelief among taxa are clearly not allometric.
Discussion
Differences between primates in tooth crownshape have been related to differences in the mech-anical demands of their diets (Kay, 1975; Lucas,1979; Lucas and Teaford, 1994; Strait, 1997;Yamashita, 1998). As Spears and Crompton(1996) noted, dental morphology affects the
nature, magnitude, and distribution of stress onfood particles. Morphological differences betweenthe molars of species considered in this studytherefore should reflect differences in their diets.
Differences in occlusal morphology betweenchimpanzees and gorillas
The differences in occlusal morphology betweenchimpanzees and gorillas evidently relate to differ-ences in the material properties of the foodsthey eat, particularly their fallback foods. CentralAfrican common chimpanzees are primarilyfrugivorous, with soft fruits reported to constitute70–80% of their diets (Kuroda, 1992; Tutin et al.,1997). Fruit is also commonly consumed bywestern lowland gorillas, making up about half ofthe food species found in their fecal remains(Williamson et al., 1990; Nishihara, 1992; Remis,
Fig. 4. Triangulated irregular network representations of surface data for M2s of representative specimens at wear stage 2: a) Gorillagorilla (CMNH B1781), b) Pan troglodytes (CHMN B3437), c) Homo rudolfensis (KNM-ER 1506), and d) Australopithecus afarensis(AL 266-1). Specimens are scaled to the same size.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622614

1997; Doran et al., 2002). Differences and simi-larities in food preferences are most obvious wherethese taxa are sympatric and have access to thesame resources. At Lope, Gabon, for example,dietary overlap is substantial, with gorillasreported to consume 73% of the food species eatenby chimpanzees (Tutin and Fernandez, 1985).Differences between the two taxa are notable attimes of fruit scarcity though, when gorillas fallback on tougher, more fibrous foods (such asleaves and stems) than those eaten by chimpanzees(Tutin et al., 1991; Remis, 1997).
Several studies have shown that gorillas havelonger shearing crests and steeper cusps than dochimpanzees (Kay, 1978; Hartman, 1986; Uchida,1998). These occlusal differences clearly reflectdifferences in the mechanical demands of theirdiets. Tougher fallback items taken by gorillasrequire long tooth–tooth contact times, and steepplanes of contact for shearing and slicing (Kay andHiiemae, 1974; Lucas, 1979; Strait, 1997; Uchida,1998). Flatter surfaces of chimpanzee molars arebetter suited to crushing and grinding, and extract-ing juices from fruit pulp. Given analogies betweenshearing crest length and occlusal relief andbetween cusp steepness and crown slope, datapresented here likewise reflect reported differencesin diet between these two taxa (M’Kirera andUngar, 2003; Ungar and M’Kirera, 2003). In sum,more occlusal relief and steeper cusps suggestefficiency in fracturing tougher, more deformablefoods, whereas less occlusal relief and less steepcusps suggest efficiency in fracturing brittle, lessdeformable foods.
It should be reiterated that differences in occlu-sal morphology between P. t. troglodytes and G. g.gorilla evidently reflect differences in fallbackresources rather than preferred foods. While bothtaxa evidently prefer soft fruits when available,differences in occlusal morphology apparentlyallow the gorillas to take advantage of fallbackfoods that are less accessible to the chimpanzees.The idea that occlusal differences can relate toimportant secondary foods rather than more com-monly consumed preferred items is not new.Kinzey (1978) noted, for example, that whileCallicebus moloch and C. torquatus are both pri-marily frugivorous, the former have longer shear-
ing crests for slicing leaves and the latter havelarger talonid basins for crushing insect chitin. Hereasoned that dental morphology therefore reflectsadaptations not only to primary foods, but also toless frequently eaten but still critical ones.
As an aside, it can be noted that mountaingorillas (Gorilla gorilla beringei) are regularlyreported to consume more leaves, stems and otherfibrous foods year-round than western lowlandgorillas (Watts, 1984, Doran et al., 2002). Manymountain gorillas live in higher altitude forestswith fewer available fruit species. Differential fruitavailability, rather than food preference, mightthen explain diet differences between the G. g.beringei and G. g. gorilla subspecies. This is con-sistent with observations that mountain gorillas atsites with greater fruit availability consume morefruit (McNeilage, 2001; Robbins and McNeilage,2003). While food preferences can be difficult toassess, it might therefore be that mountain gorillasat “marginal” sites perpetually fallback on morefibrous, less desirable foods. If so, gorilla occlusalmorphology would facilitate the dietary flexibilitynecessary to allow this.
Dental topography and early hominin diets
Relationships between tooth morphology anddiet in chimpanzees and gorillas provide a goodcomparative baseline for inferring aspects of dietin early hominins. The dental topography data forAustralopithecus afarensis and early Homo can beinterpreted in this context, and results comparedwith those from previous studies on the diets ofthese fossil taxa.
Australopithecus afarensisSeveral workers have noted that Australo-
pithecus afarensis had smaller incisors than extantchimpanzees, but larger molar teeth (McHenry,1988; Picq, 1990; Teaford and Ungar, 2000; Woodand Richmond, 2000). According to conventionalwisdom, dental allometry, taken with evidence ofthick tooth enamel and thick mandibular corpora,suggest intensive mastication without extensiveincisal preparation. Researchers have suggested,following these lines of evidence, a diet thatincluded nuts, seeds, and underground storage
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 615

organs. It has been emphasized, however, that A.afarensis probably had a mixed forest-savannaresource adaptation rather than a hard-objectspecialization. These hominins would probablystill have preferred soft, sugar-rich fruits, butwould have been able to make better use of hard,brittle resources as fallback foods given seasonalavailability of favored items (Picq, 1990; Teafordand Ungar, 2000; Ungar and Teaford, 2001; seeConklin-Brittain et al., 1998). Ryan and Johanson(1989) also proposed a mixed adaptation for A.afarensis based on dental microwear features ontheir anterior teeth. This suggested to them theconsumption of both closed forest plants and opensavanna foods, such as seeds, roots, and rhizomes.
Results presented here for Australopithecusafarensis indicate less crown relief and less slopingocclusal surfaces at given stages of wear than ineither gorillas or chimpanzees. This implies moreefficient fracture of brittle, less deformable foodsbut less efficient fracture of tough, more deform-able foods. The average degrees of differencebetween chimpanzees and A. afarensis in occlusalslope and relief are certainly no more, and likelyeven less than that between the chimpanzees andgorillas. Given that the extant ape diets differmostly in fallback foods, it is reasonable to suggestthat the A. afarensis diet may also have differedfrom that of chimpanzees largely in fallbackresources, which could have included more hard,brittle items. This is consistent with dietary recon-structions of Ryan and Johanson (1989) and Picq(1990).
Early HomoThere is less consensus concerning the dietary
adaptations of early Homo in the literature thanthere is for Australopithecus afarensis. Severalmodels have stressed changing resources associ-ated with spreading savannas around the time ofthe Plio-Pleistocene boundary (see Cerling, 1992;Behrensmeyer et al., 1997; Potts, 1998). Many havesuggested an increase in vertebrate tissue con-sumption based largely on the longstandingassumed relationships between Oldowan tools, evi-dence of butchery, and fossils of early Homo (e.g.,Leakey et al., 1964). Physiological models andethnographic/ethological analogy have been cited
in support of this scenario (Milton, 1987; Aielloand Wheeler, 1995; Stanford and Bunn, 2001, andreferences therein). Others have suggested thatsavanna plants (especially their undergroundstorage organs, or USOs) rather than animalsprovided keystone resources for early Homo,particularly H. erectus (O’Connell et al., 1999;Wrangham et al., 1999).
The fossil remains of early Homo have, how-ever, provided little concrete evidence one way orthe other. Some have suggested that early Homo,especially H. habilis and H. rudolfensis, had rela-tively large incisors, implying increased consump-tion of foods requiring incisal preparationcompared with many australopiths (Jolly, 1970;Teaford et al., 2002). Others have suggested thatreduced molar size in early Homo, compared withaustralopiths, indicates the consumption of amechanically less demanding diet—though there isno consensus concerning whether this size decreasebegan with H. habilis and H. rudolfensis or withearly African H. erectus (Wood and Collard, 1999;McHenry and Coffing, 2000). Scenarios suggestingconsumption of a mechanically less demandingdiet may receive some support from observationsthat early Homo, especially H. erectus, had thinnermolar enamel than most australopiths (see Beynonand Wood, 1986).
If the small samples considered here arerepresentative of their taxa, dental topographicanalysis indicates that early Homo is intermediatebetween chimpanzees and gorillas in slope andocclusal relief, and therefore, was probably inter-mediate in the fracture properties of the foods towhich it was adapted. Early Homo could havemore efficiently fractured tough, pliant foodsthan could chimpanzees, or for that matter, A.afarensis. Thinner enamel in early Homo wouldalso be consistent with the consumption of tougherfoods, as sharp edges at the sites of dentinexposure with wear would improve shredding andslicing efficiency (Kay, 1981; Ungar and M’Kirera,2003).
What sorts of tough foods might have beenavailable to early Homo? Research on mechanicalproperties of foods that would have been availableto early hominins is very limited. Nevertheless,tubers, especially larger ones (Baritelle and Hyde,
P. Ungar / Journal of Human Evolution 46 (2004) 605–622616

1999), are often fairly brittle, whereas mammaliansoft tissues tend to be tough and elastic (Lucas andPeters, 2000). Thus, meat seems more likely tohave been a key tough-food resource for earlyHomo than would have USOs. It has also beennoted that USOs are of limited nutritional value(Schoeninger et al., 2001), and so would not havemade very good keystone resources.
Increased occlusal relief and steeper slopedcusps yield sharper cutting surfaces that wouldgive animal tissues less of an opportunity to stretchand absorb energy, thereby thwarting the majortoughening mechanism. Thus, increased consump-tion of animal products may have played a rolein the dental adaptations of the earliest membersof our genus. Indeed, researchers have con-sidered hunting and meat eating to be key elementsin human evolution since the time of Darwin(1871).
If so, it may be argued that environmentalchange during the Plio-Pleistocene provided earlyHomo with the motive, and technological innova-tion offered them the opportunity for a new dietaryadaptation that included more meat than eaten bytheir hominin predecessors. Cerling (1992) notedthat following 2.5 Ma, C4 grasslands spread acrossEast Africa, concomitant with periodic fluctua-tions in climate (Behrensmeyer et al., 1997; Potts,1998). Conventional wisdom associates adaptiveradiations of both Paranthropus and Homo withthese environmental changes (Stanley, 1992; DeMenocal, 1995; Vrba et al., 1995). This suggestsan increasingly mosaic environment that wouldhave included fewer C3 foods and more savannaresources, including larger-bodied ungulates.
It is also compelling that the earliest evidence ofstone tool manufacture and animal processing(Semaw et al., 1997, 2003; De Heinzelin et al.,1999) are found in deposits dated to about 2.5 Ma,roughly coincident with the first appearance ofHomo (Hill et al., 1992; Bromage et al., 1995;Kimbel et al., 1996; Suwa et al., 1996; Wood, 1999;Sherwood et al., 2002). Further, most sites yieldingstone tools and bones modified by early homininspostdate 2.0 Ma (Blumenschine and Masao, 1991),with the earliest large concentrations roughlycoincident with the appearance of African Homoerectus (Feibel et al., 1989; Wood, 1991).
Differences between the homininsThe teeth of Australopithecus afarensis have
been said to present a “primitive morphologicalcomposite that parallels extant and MiddleMiocene apes” (White et al., 1981: 466; see alsoPicq, 1990). White and coauthors (White et al.,1981) note, for example, that the lower molarcusps of A. afarensis lack the swollen, inflatedappearance often seen in later australopiths, suchas A. africanus and Paranthropus spp. The post-canine dentition of A. afarensis may thereforepresent a generalized australopith morphotypefrom which early Homo occlusal morphologycould have been derived.
Dental topographic differences between Austra-lopithecus afarensis and early Homo are of aboutthe same magnitude as differences between Pantroglodytes troglodytes and Gorilla gorilla gorilla.As noted above, these chimpanzees and gorillasoverlap considerably in diet, but do differ some-what, particularly in fallback resources consumedduring seasonal crunch times. Differences in dietbetween A. afarensis and early Homo may likewisehave been mostly in fallback resources.
Raymond Dart’s ideas concerning predatorynature of hominins were predicated on observa-tions that savanna baboons fallback on huntingduring harsh dry seasons when preferred resourcesare unavailable. He wrote “baboons anywherebecome hunting mammals under the pressure ofseasonal or environmental need” (Dart, 1953: 215).Perhaps, then, the transition to Homo may haveinvolved a shift toward the use of meat as afallback resource given more open or variableenvironments. Hopefully, other approaches to thereconstruction of diet, such as bone and toothchemistry analyses, will provide further evidencewith which to evaluate this hypothesis.
Caveats
Such hypotheses must, however, be temperedby the acknowledgement that, while a great dealof research has focused on the paleoenviron-mental context of the Plio-Pleistocene hominins(Cerling, 1992; contributions in Vrba et al., 1995;Behrensmeyer et al., 1997; Potts, 1998; Sept, 2000),we cannot know the full complement of foods
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 617

available in the past, let alone infer their materialproperties. Furthermore, we cannot attribute theearliest stone tools and cut-marked bones to earlyHomo with complete confidence given the presenceof both Australopithecus garhi (Asfaw et al., 1999)and Paranthropus aethiopicus (Walker et al., 1986)in East Africa at about 2.5 Ma. Nor can weattribute the first large concentrations of thesearchaeological remains to a specific hominintaxon, given the contemporaneity of H. habilis, H.rudolfensis, and Paranthropus boisei. Still, it isreasonable to assume that early H. erectus inAfrica ate vertebrate tissues given Plio-Pleistocenearchaeological evidence in Eurasia, where no otherhominin species have so far been found (e.g.,Dennell et al., 1988; Gabunia and Vekua, 1995).Most would, likewise, agree that H. rudolfensis andH. habilis probably also made and used Oldowantools to process animal tissues.
Tool use introduces another possible confound-ing variable for interpreting dental morphology inearly Homo. Most tools made by non-humanprimates and other animals are used for foodacquisition and processing (e.g., Hunt, 1996; Foxet al., 1999; Whiten et al., 1999). If early Homobecame increasingly reliant on tools (Oakley,1962), selective pressures on their jaws and teethmay have changed along with properties of foodsas they were altered by preparation (Brace et al.,1991). If selective pressures changed accordingly,this could make it even more difficult to useform–function relationships to reconstruct thediets of these hominins.
Conclusions
In sum, dental topographic analysis of Austra-lopithecus afarensis and early Homo add to ourunderstanding of the dietary adaptations of thesetaxa. First, it is clear that, as with results forchimpanzees and gorillas, average cusp slope andocclusal relief decreased in both fossil taxa as theirmolar teeth wore down. It is also clear that differ-ences between taxa are about the same at givenwear stages, suggesting that variably worn fossilteeth can be included in functional analyses.Results presented here indicate that, at given wear
stages, early Homo teeth have more occlusal reliefthan chimpanzees and less than gorillas, whereasA. afarensis molars have less occlusal relief thanthe other taxa studied. This suggests that earlyHomo was capable of more efficiently consumingtough, elastic foods than are chimpanzees or thanwas A. afarensis. In contrast, A. afarensis was wellsuited to crushing hard, brittle foods. Differencesbetween the hominins are similar in degree to thosebetween the extant African apes, suggesting thatA. afarensis may have taken harder, more brittlefallback foods, whereas early Homo may haverelied more on tough fallback resources, perhapsincluding meat.
Acknowledgements
Tooth molds used in this study were preparedby Fred Grine, Alejandro Perez-Perez, MarkTeaford, and me as part of another study, andmuch of the chimpanzee and gorilla GIS data werecollected by Francis M’Kirera. I am grateful to thereviewers and Bill Kimbel for their helpful com-ments. I am also grateful to Alan Walker, RichKay, Peter Lucas, Jerry Rose, Mike Plavcan,Francis M’Kirera, John Dennis, John Wilson,Malcolm Williamson, Fred Limp, and especiallyFred Grine and Mark Teaford for discussions andideas exchanged over the past decade. Further,Diane Doran, Michelle Remis, and Amy Vedderprovided valuable insights on gorilla diets. Finally,I thank Bill Kimbel and Meave Leakey for grant-ing access to specimens, and the Governmentsand National Museums of Ethiopia, Kenya andTanzania and curators at the Transvaal Museum,the University of the Witwatersrand, and theCleveland Museum of Natural History for permis-sion to study collections in their care. This researchwas funded by the LSB Leakey Foundation andNSF grant SBR 9804882.
References
Aiello, L.C., Wheeler, P., 1995. The expensive tissue hypothesis.Curr. Anthrop. 36, 199–221.
Anthony, M.R.L., Kay, R.F., 1993. Tooth form and dietin ateline and alouattine primates: reflections on thecomparative method. Am. J. Sci. 293A, 356–382.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622618

Asfaw, B., White, T.D., Lovejoy, O., Latimer, B., Simpson, S.,Suwa, G., 1999. Australopithecus garhi: a new species ofearly hominid from Ethiopia. Science 284, 629–635.
Baritelle, A.L., Hyde, G.M., 1999. Effect of tuber size on failureproperties of potato tissue. Trans. ASAE 42, 159–161.
Behrensmeyer, A.K., Todd, N.E., Potts, R., McBrinn, G.E.,1997. Late Pliocene faunal turnover in the Turkana Basin,Kenya and Ethiopia. Science 278, 1589–1594.
Benefit, B.R., 2000. Old World monkey origins and diver-sification: an evolutionary study of diet and dentition.In: Whitehead, P.F., Jolly, C.J. (Eds.), Old WorldMonkeys. Cambridge University Press, Cambridge, pp.133–179.
Beynon, A.D., Wood, B.A., 1986. Variations in enamel thick-ness and structure in East African hominids. Am. J. Phys.Anthrop. 70, 177–193.
Blumenschine, R.J., Masao, F.T., 1991. Living sites at OlduvaiGorge, Tanzania? Preliminary landscape archaeologyresults in the basal Bed II lake margin zone. J. Hum. Evol.21, 451–462.
Brace, C.L., Smith, B.H., Hunt, K.D., 1991. What big teeth youhad grandma! Human tooth size, past and present. In:Kelley, M.A., Larsen, C.S. (Eds.), Advances in DentalAnthropology. Wiley-Liss Press, New York, pp. 33–57.
Bromage, T.G., Schrenk, F., Zonneveld, F., 1995. Paleoanthro-pology of the Malawi Rift: an early hominid mandible fromthe Chiwondo Beds, northern Malawi. J. Hum. Evol. 28,71–108.
Cerling, T.E., 1992. Development of grasslands andsavannas in East Africa during the Neogene. Paleogeogr.,Palaeoclimatol., Paleoecol. 97, 241–247.
Conklin-Brittain, N.K., Wrangham, R.W., Hunt, K.D., 1998.Dietary response of chimpanzees and cercopithecines toseasonal variation in fruit abundance II. Macronutrients.Int. J. Primatol. 19, 971–998.
Conover, W.J., Iman, R.L., 1981. Rank transformations as abridge between parametric and nonparametric statistics.Am. Statistician 35, 124–129.
Crompton, A.W., Sita-Lumsden, A.G., 1970. Functionalsignificance of therian molar pattern. Nature 227, 197–199.
Dart, R.A., 1953. The predatory transition from ape to man.Int. Anthrop. Ling. Rev. 1, 201–217.
Darwin, C., 1871. The Descent of Man. Appleton andCompany, New York.
Dean, M.C., Jones, M.E., Pilley, J.R., 1992. Natural history oftooth wear, continuous eruption and periodontal disease inwild shot great apes. J. Hum. Evol. 22, 23–39.
De Heinzelin, J., Clark, J.D., White, T., Hart, W., Renne, P.,Woldegabriel, G., Beyene, Y., Vrba, E., 1999. Environmentand behavior of 2.5-million-year-old Bouri hominids.Science 284, 625–629.
De Menocal, P.B., 1995. Plio-Pleistocene African climate.Science 270, 53–59.
Dennell, R.W., Rendell, H., Hailwood, E., 1988. Early toolmaking in Asia: two million year old artefacts in Pakistan.Antiquity 62, 98–106.
Dennis, J.C., Ungar, P.S., Teaford, M.F., Glander, K.E., inpress. Dental topography and molar wear in Alouattapalliata from Costa Rica. Am. J. Phys. Anthrop.
Doran, D.M., McNeilage, A., Greer, D., Boician, C., Mehlan,P., Shah, N., 2002. Western lowland gorilla diet andresource availability: new evidence, cross-site comparisons,and reflections on indirect sampling methods. Am. J.Primatol. 58, 91–116.
Drake, R., Curtis, G.H., 1987. Geochronology of the Laetolifossil localities. In: Leakey, M.D., Harris, J.M. (Eds.),Laetoli: A Pliocene Site in Northern Tanzania. ClarendonPress, Oxford, pp. 490–496.
Feibel, C., Brown, F.H., McDougall, I., 1989. Stratigraphiccontext of fossil hominids from the Omo Group deposits:northern Turkana Basin, Kenya and Ethiopia. Am. J. Phys.Anthrop. 78, 595–622.
Fleagle, J.G., 1999. Primate Adaptation and Evolution, seconded. Academic Press, New York.
Fleagle, J.G., Kay, R.F., Anthony, M.R.L., 1996. Fossil NewWorld monkeys. In: Kay, R.F., Madden, R.H., Cifelli,R.L., Flynn, J.J. (Eds.), Vertebrate Paleontology inthe Neotropics. Smithsonian Institution, Washington DC,pp. 473–495.
Fox, E.A., Sitompul, A.F., van Schaik, C.P., 1999. Intelligenttool use in wild Sumatran orangutans. In: Parker, S., Miles,L., Mitchell, A. (Eds.), The Mentality of Gorillas andOrangutans. Cambridge University Press, Cambridge, pp.99–117.
Gabunia, L., Vekua, A., 1995. A Plio-Pleistocene hominid fromDmanisi, East Georgia, Caucasus. Nature 373, 509–512.
Gregory, W.K., 1922. The Origin and Evolution of HumanDentition. Williams and Wilkins, Baltimore.
Grine, F.E., 1981. Trophic differences between ‘gracile’ and‘robust’ australopithecines: a scanning electron microscopeanalysis of occlusal events. S. Afr. J. Sci. 77, 203–230.
Grine, F.E., 1984. Deciduous molar microwear of SouthAfrican australopithecines. In: Chivers, D.J., Wood, B.A.,Bilsborough, A. (Eds.), Food Acquisition and Processing inPrimates. Plenum, New York, pp. 525–534.
Grine, F.E., Martin, L.B., 1988. Enamel thickness anddevelopment in Australopithecus and Paranthropus. In:Grine, F.E. (Ed.), Evolutionary History of the “Robust”Australopithecines. Aldine de Gruyter, New York, pp.3–42.
Hartman, S.E., 1986. A cladistic analysis of hominoid molars.J. Hum. Evol. 17, 489–502.
Hill, A., Ward, S., Deino, A., Curtis, G., Drake, R., 1992.Earliest Homo. Nature 355, 719–722.
Hunt, G.R., 1996. Manufacture and use of hook-tools by NewCaledonian crows. Nature 379, 249–251.
Johanson, D.C., White, T.D., 1979. A systematic assessment ofearly African hominids. Science 203, 321–330.
Jolly, C.J., 1970. The seed-eaters: a new model of hominiddifferentiation based on a baboon analogy. Man 5, 1–26.
Kay, R.F., 1975. Allometry in early hominids (letter). Science189, 63.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 619

Kay, R.F., 1977. Diets of early Miocene African hominoids.Nature 268, 628–630.
Kay, R.F., 1978. Molar structure and diet in extantCercopithecidae. In: Butler, P.M., Joysey, K.A. (Eds.),Development, Function, and Evolution of Teeth. AcademicPress, New York, pp. 309–339.
Kay, R.F., 1981. The nut-crackers: a new theory of the adap-tations of the Ramapithecinae. Am. J. Phys. Anthrop. 55,141–151.
Kay, R.F., 1985. Dental evidence for the diet ofAustralopithecus. A. Rev. Anthrop. 14, 315–341.
Kay, R.F., Covert, H.H., 1984. Anatomy and behavior ofextinct primates. In: Chivers, D.J., Wood, B.A.,Bilsborough, A. (Eds.), Food Acquisition and Processing inPrimates. Plenum Press, New York, pp. 467–508.
Kay, R.F., Hiiemae, K.M., 1974. Jaw movement and tooth usein recent and fossil primates. Am. J. Phys. Anthrop. 40,227–256.
Kay, R.F., Hylander, W.L., 1978. The dental structure ofmammalian folivores with special reference to primates andPhalangeroidea (Marsupialia). In: Montgomery, G.G.(Ed.), The Ecology of Arboreal Folivores. SmithsonianInstitution, Washington, DC, pp. 173–191.
Kay, R.F., Simons, E.L., 1980. The ecology of oligoceneAfrican Anthropoidea. Int. J. Primatol. 1, 21–37.
Kay, R.F., Ungar, P.S., 1997. Dental evidence for diet insome Miocene catarrhines with comments on the effects ofphylogeny on the interpretation of adaptation. In: Begun,D.R., Ward, C., Rose, M. (Eds.), Function, Phylogeny andFossils: Miocene Hominoid Evolution and Adaptations.Plenum Press, New York, pp. 131–151.
Kimbel, W.H., Johanson, D.C., Rak, Y., 1994. The first skulland other new discoveries of Australopithecus afarensis atHadar, Ethiopia. Nature 368, 449–451.
Kimbel, W.H., Walter, R.C., Johanson, D.C., Reed, K.E.,Aronson, J.L., Assefa, Z., Marean, C.W., Eck, G.G., Bobe,R., Hovers, E., Rak, Y., Vondra, C., Yemane, T., York, D.,Chen, Y., Evensen, N.M., Smith, P.E., 1996. Late PlioceneHomo and Oldowan tools from the Hadar Formation(Kada Hadar Member), Ethiopia. J. Hum. Evol. 31,549–561.
Kinzey, W.G., 1978. Feeding behavior and molar features intwo species of titi monkey. In: Chivers, D.J., Herbert, J.(Eds.), Recent Advances in Primatology, Volume 1:Behavior. Academic Press, London, pp. 373–385.
Kuroda, S., 1992. Ecological interspecies relationshipsbetween gorillas and chimpanzees in the Ndoki-NouabaleReserve, Northern Congo. In: Itoigawa, N., Sugiyama, Y.,Sackett, G.P., Thomson, R.K.R. (Eds.), Topics inPrimatology, Vol. 2: Behavior, Ecology and Conservation.University of Tokyo Press, Tokyo, pp. 385–394.
Leakey, L.S.B., Tobias, P.V., Napier, J.R., 1964. A new speciesof the genus Homo from Olduvai Gorge. Nature 202, 7–9.
Leakey, M.G., Spoor, F., Brown, F.H., Gathogo, P.N., Kiarie,C., Leakey, L.N., McDougall, I., 2001. New hominin genusfrom eastern Africa shows diverse middle Pliocene lineages.Nature 410, 433–440.
Lieberman, D.E., 2001. Another face in our family tree. Nature410, 419–420.
Lockwood, C.A., Kimbel, W.H., Johanson, D.C., 2000.Temporal trends and metric variation in the mandiblesand dentition of Australopithecus afarensis. J. Hum. Evol.39, 23–55.
Lucas, P.W., 1979. The dental-dietary adaptations ofmammals. Neus jahrbuch Geol. Palaeontol. 8, 486–512.
Lucas, P.W., Peters, C.R., 2000. Function of postcanine toothshape in mammals. In: Teaford, M.F., Smith, M.M.,Ferguson, M.W.J. (Eds.), Development, Function andEvolution of Teeth. Cambridge University Press,Cambridge, pp. 282–289.
Lucas, P.W., Teaford, M.F., 1994. Functional morphologyof colobine teeth. In: Davies, A.G., Oates, J.F. (Eds.),Colobine Monkeys: Their Ecology, Behaviour andEvolution. Cambridge University Press, Cambridge, pp.173–203.
M’Kirera, F., Ungar, P.S., 2003. Occlusal relief changes withmolar wear in Pan troglodytes troglodytes and Gorilla gorillagorilla. Am. J. Primatol. 60, 31–42.
Maier, W., 1977. Die evolution der bilophodonten molaren derCercopithecoidea. Z. Morph. Anthrop. 68, 25–56.
Maier, W., 1984. Tooth morphology and dietary specialization.In: Chivers, D.J., Wood, B.A., Bilsborough, A. (Eds.),Food Acquisition and Processing in Primates. PlenumPress, New York, pp. 467–508.
McHenry, H.M., 1988. New estimates of body weight in earlyhominids and their significance to encephalization andmegadontia in “robust” australopithecines. In: Grine, F.E.(Ed.), Evolutionary History of the “Robust”Australopithecines. Aldine Press, New York, pp. 133–148.
McHenry, H.M., Coffing, K., 2000. Australopithecus to Homo:transformations in body and mind. A. Rev. Anthrop. 29,125–146.
McNeilage, A., 2001. Diet and habitat use of two mountaingorilla groups in contrasting habitats in the Virungas. In:Robbins, M.M., Sicotte, P., Stewart, K.J. (Eds.), MountainGorillas: Three Decades of Research at Karisoke.Cambridge University Press, Cambridge, pp. 265–292.
Meldrum, D.J., Kay, R.F., 1997. Nucicruptor rubicae, a newpitheciin seed predator from the Miocene of Colombia. Am.J. Phys. Anthrop. 102, 407–428.
Milton, K., 1987. Primate diets and gut morphology: implica-tions for hominid evolution. In: Harris, M., Ross, E.B.(Eds.), Food and Evolution: Toward a Theory of HumanFood Habits. Temple University Press, Philadelphia, pp.93–115.
Nishihara, T., 1992. A preliminary report on the feeding habitsof western lowland gorillas (Gorilla gorilla gorilla) in theNdoki Forest, Northern Congo. In: Itoigawa, N., Sugiyama,Y., Sackett, G.P., Thompson, R.K.R (Eds.), Topics inPrimatology, Vol. 2: Behavior, Ecology and Conservation.University of Tokyo Press, Tokyo, pp. 225–240.
O’Connell, J.F., Hawkes, K., Blurton-Jones, N.J., 1999.Grandmothering and the evolution of Homo erectus. J.Hum. Evol. 36, 461–485.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622620

Oakley, K.P., 1962. The earliest tool-makers. In: Kurth, G.(Ed.), Evolution und Hominisation. Festchrift zum 06.Geburtstage von Gerhard Heberer, Stuttgart, pp. 157–169.
Owen, R., 1845, Odontography. 1 and 2. Hippolyte BaillierePress, London.
Picq, P., 1990. The diet of Australopithecus afarensis: anattempted reconstruction. C. R. Acad. Sci. Ser. II 311,725–730.
Potts, R., 1998. Environmental hypotheses of homininevolution. Yearb. phys. Anthrop. 41, 93–136.
Remis, M.J., 1997. Western lowland gorillas (Gorilla gorillagorilla) as seasonal frugivores: use of variable resources.Am. J. Primatol. 43, 87–109.
Robbins, M.M., McNeilage, A., 2003. Home range andfrugivory patterns in Bwindi Impenetrable National Park,Uganda. Int. J. Primatol. 24, 467–491.
Rose, J.C., Ungar, P.S., 1998. Gross dental wear and dentalmicrowear in historical perspective. In: Alt, K.W., Rosing,F.W., Teschler-Nicola, M. (Eds.), Dental Anthropology:Fundamentals, Limits, Perspectives. Gustav FischerPublishers, Stuttgart, pp. 349–386.
Rosenberger, A.J., Kinzey, W.G., 1976. Functional patterns ofmolar occlusion in platyrrhine primates. Am. J. Phys.Anthrop. 45, 281–297.
Ryan, A.S., Johanson, D.C., 1989. Anterior dental microwearin Australopithecus afarensis. J. Hum. Evol. 18, 235–268.
Scott, E.C., 1979. Dental wear scoring technique. Am. J. phys.Anthrop. 51, 213–218.
Schoeninger, M.J., Bunn, H.T., Murray, S.S., Marlett, J.A.,2001. Composition of tubers used by Hadza foragers ofTanzania. J. Food Compos. Analysis 14, 15–25.
Seligsohn, D., Szalay, F.S., 1978. Relationship between naturalselection and dental morphology: tooth function and diet inLepilemur and Hapalemur. In: Butler, P.M., Joysey, K.A.(Eds.), Development, Function and Evolution of Teeth.Academic Press, London, pp. 289–307.
Semaw, S., Renne, P., Harris, J.W.K., Feibel, C.S., Bernor,R.L., Fesseha, N., Mowbray, K., 1997. 2.5 million-year-oldstone tools from Gona, Ethiopia. Nature 385, 333–336.
Semaw, S., Rogers, M.J., Quade, J., Renne, P.R., Butler, R.F.,Dominguez-Rodrigo, M., Stout, D., Hard, W.S., Pickering,T., Simpson, S.W., 2003. 2.6-Million-year-old stone toolsand associated bones from OGS-6 and OGS-7 Gona, Afar,Ethiopia. J. Hum. Evol. 45, 169–177.
Sept, J., 2000. Modeling the edible landscape. In: Stanford,C.B., Bunn, H.T. (Eds.), Meat-Eating and HumanEvolution. Oxford University Press, Oxford, pp. 73–100.
Sherwood, R.J., Ward, S.C., Hill, A., 2002. The taxonomicstatus of the Chemeron temporal (KNM-BC 1). J. Hum.Evol. 42, 153–184.
Spears, I.R., Crompton, R.H., 1996. The mechanical signifi-cance of the occlusal geometry of great ape molars in foodbreakdown. J. Hum. Evol. 31, 517–535.
Stanford, C.B., Bunn, H.T., 2001. Meat-Eating and HumanEvolution. Oxford University Press, Oxford.
Stanley, S.M., 1992. An ecological theory for the origin ofHomo. Paleobiology 18, 237–257.
Strait, D.S., Grine, F.E., 2001. The systematics of Australo-pithecus garhi. Ludus Vitalis 9, 109–135.
Strait, D.S., Grine, F.E., Moniz, M.A., 1997. A reappraisal ofearly hominid phylogeny. J. Hum. Evol. 32, 17–82.
Strait, S.G., 1993. Differences in occlusal morphology and molarsize in frugivores and faunivores. J. Hum. Evol. 25, 471–482.
Strait, S.G., 1997. Tooth use and the physical properties offoods. Evol. Anthrop. 5, 199–211.
Suwa, G., White, T.D., Howell, F.C., 1996. Mandibular post-canine dentition from the Shungura Formation, Ethiopia:crown morphology, taxonomic allocations, and Plio-Pleistocene hominid evolution. Am. J. Phys. Anthrop. 101,247–282.
Teaford, M.F., Ungar, P.S., 2000. Diet and the evolution of theearliest human ancestors. Proc. Natl. Acad. Sci. 97,13506–13511.
Teaford, M.F., Ungar, P.S., Grine, F.E., 2002. Paleontologicalevidence for the diets of African Plio-Pleistocene homininswith special reference to early Homo. In: Ungar, P.S.,Teaford, M.F. (Eds.), Human Diet: Its Origins andEvolution. Bergin and Garvey Publishers, Westport, CT,pp. 143–166.
Tutin, C.E.G., Fernandez, M., 1985. Food consumed bysympatric populations of Gorilla g. gorilla and Pan. t.troglodytes in Gabon: some preliminary data. Int. J.Primatol. 6, 27–43.
Tutin, C.E., Ham, R., White, L.J.T., Harrison, M.J.S., 1997.The primate community of the Lope Reserve in Gabon:diets, responses to fruit scarcity and effects on biomass. Am.J. Primatol. 42, 1–24.
Tutin, C.E.G., Fernandez, M., Rogers, M.E., Williamson,W.A., McGrew, W.C., 1991. Foraging profiles of sympatriclowland gorillas and chimpanzees in the Lope Reserve,Gabon. Phil. Trans. R. Soc. Lond. B. 334, 179–186.
Uchida, A., 1998. Variation in tooth morphology of Gorillagorilla. J. Hum. Evol. 34, 55–70.
Ungar, P.S., 1998. Dental allometry, morphology and wear asevidence for diet in fossil primates. Evol. Anthrop. 6,205–216.
Ungar, P.S., Dennis, J.C., Wilson, J., Grine, F., 2002. Quanti-fication of tooth crown shape by dental topographicanalysis. Am. J. Phys. Anthrop. 34(Suppl.), 158–159.
Ungar, P.S., Kay, R.F., 1995. The dietary adaptations ofEuropean Miocene catarrhines. Proc. Natl. Acad. Sci. 92,5479–5481.
Ungar, P.S., M’Kirera, F., 2003. A solution to the worn toothconundrum in primate functional anatomy. Proc. Natl.Acad. Sci. 100, 3874–3877.
Ungar, P.S., Teaford, M.F., 2001. The dietary split betweenapes and the earliest human ancestors. In: Tobias, P.V.,Raath, M.A., Moggi-Chcchi, J., Doyle, G.A. (Eds.),Humanity from African Naissance to Coming Millennia.Firenze University Press, Florence, pp. 337–354.
Ungar, P.S., Teaford, M.F., Grine, F.E., 1999. A preliminarystudy of molar occlusal relief in Australopithecus africanusand Paranthropus robustus. Am. J. Phys. Anthrop.28(Suppl.), 269.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622 621

Ungar, P., Williamson, M., 2000. Exploring the effects of toothwear on functional morphology: a preliminary study usingdental topographic analysis. Palaeontologia Electronica 3,http://palaeo-electronica.org/2000_1/ gorilla/issue1_00.htm.
Vrba, E.S., Denton, G.H., Partridge, T.C., Gurkle, L.H., 1995.Paleoclimate and Evolution with Emphasis on HumanOrigins. Yale University Press, New Haven, CT.
Walker, A., Leakey, R.E., Harris, J.M., Brown, F.H., 1986.2.5-Myr Australopithecus boisei from West of LakeTurkana, Kenya. Nature 322, 517–522.
Ward, C., Leakey, C., Walker, A., 1999. The new hominidspecies Australopithecus anamensis. Evol. Anthrop. 7,197–205.
Watts, D., 1984. Composition and variability in mountaingorilla diets in the Central Virungas. Am. J. Primatol. 7,323–356.
White, T.D., Johanson, D.C., Kimbel, W.H., 1981. Australo-pithecus africanus: its phylogenetic position reconsidered. S.Afr. J. Sci. 77, 445–470.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T.,Reynolds, V., Sugiyama, Y., Tutin, C.E.G., Wrangham,R.W., Boesch, C., 1999. Cultures in chimpanzees. Nature399, 682–685.
Williams, B.A., Covert, H.H., 1994. New early Eocene anapto-morphine primate (Omomyidae) from the Washakie Basin,Wyoming, with comments on the phylogeny and paleobiol-ogy of anaptomorphines. Am. J. Phys. Anthrop. 93,323–340.
Williamson, E.A, Tutin, C.E., Rogers, M.E., Fernandez, M.,1990. Composition of the diet of the lowland gorillas atLope in Gabon. Am. J. Primatol. 21, 265–277.
Wood, B.A., 1991. Koobi Fora Research Project, Vol. 4:Hominid Cranial Remains. Oxford University Press,Oxford.
Wood, B.A., 1992. Origin and evolution of the genus Homo.Nature 355, 783–790.
Wood, B.A., 1999. Plio-Pleistocene hominins from the BaringoBasin, Kenya. In: Andrews, P., Banham, P. (Eds.), LateCenozoic Environments and Hominid Evolution: A Tributeto Bill Bishop. Geographic Society, London, pp. 113–122.
Wood, B.A., Van Noten, F.L., 1986. Preliminary observationson the BK 8518 mandible from Baringo, Kenya. Am. J.Phys. Anthrop. 69, 117–127.
Wood, B., Collard, M., 1999. The human genus. Science 284,65–71.
Wood, B., Richmond, B.G., 2000. Human evolution: taxonomyand paleobiology. J. Anat. 196, 19–60.
Wrangham, R.W., Jones, J.H., Laden, G., Pilbeam, D.,Conklin-Brittain, N.L., 1999. The raw and the stolen:cooking and the ecology of human origins. Curr. Anthrop.40, 567–594.
Yamashita, N., 1998. Functional dental correlates of foodproperties in five Malagasy lemur species. Am. J. Phys.Anthrop. 106, 169–188.
P. Ungar / Journal of Human Evolution 46 (2004) 605–622622
Related Documents











