Dense, small and male-biased cages exacerbate maleemale competition and reduce female choosiness in Bicyclus anynana Marie-Jeanne Holveck * , Anne-Laure Gauthier, Caroline M. Nieberding * Group of Evolutionary Ecology and Genetics, Biodiversity Research Centre, Earth and Life Institute, Universit e Catholique de Louvain (UCL), Louvain-la-Neuve, Belgium article info Article history: Received 15 October 2014 Initial acceptance 19 November 2014 Final acceptance 18 March 2015 Published online MS. number: 14-00824R Keywords: Bicyclus anynana choosiness competition density mating preference mating success sex ratio Sexual selection is increasingly recognized to depend upon, and to fluctuate with, major ecological factors in natural environments. The operational sex ratio (OSR) and population density can affect the opportunity for, and strength of, sexual selection but their effects are rarely taken into account in lab- oratory behavioural studies. In Bicyclus anynana, a model butterfly for mate choice studies, the experi- mental set-up widely used in laboratories often involves very high densities compared to the field, male- biased sex ratios and small cage volumes. We hypothesized that these conditions impede the proper expression of female mating preference by promoting male competition and sexual harassment of fe- males. Using various cage volumes, we separately manipulated OSR and density to cover the range of values used in B. anynana laboratory mating experiments and to approach field values. Male competition, quantified by the number and duration of courtships aborted by males, became stronger with increasing densities, specifically under more male-biased sex ratios, and decreasing cage volumes. Thus, male eagerness to mate was essentially due to the use of small cage volumes and very high experimental densities. Concomitantly, female choosiness, quantified by the proportion of rejections of male mating attempts, decreased with increasing densities under a male-biased sex ratio. Females also accepted more matings with decreasing cage volume, and mated more rapidly with increasing density. We conclude that the laboratory social environments frequently used to test mate choice in B. anynana, with unnat- urally high densities and male-biased sex ratios, exacerbate maleemale competition and strongly hinder female choice, biasing the estimates of the strength and direction of sexual selection that were shaped under very different natural environments in the wild. We propose a set-up for B. anynana mate choice studies that allows the assessment of female choosiness in statistically robust mating experiments. © 2015 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. Understanding the processes of pair formation is a necessary, yet neglected, step to fully grasp how secondary sexual traits and mating preferences evolve under sexual selection. It requires taking into account the fact that sexual selection, like natural selection, takes place in ecologically complex environments and examining the two major selective agents of sexual selection, mate choice and within-sex competition, in an integrative way (Miller & Svensson, 2014). It is increasingly acknowledged that mating patterns depend on ecological and social constraints, which determine the availability of potential mating partners and, as a consequence, affect the expression of within-sex competition for mate access and of between-sex mating preferences (Miller & Svensson, 2014; Widemo & Saether, 1999). Social constraints include population density (defined here as the number of individuals ready to mate in a given space) and the operational sex ratio (OSR, i.e. the ratio of sexually active males to fertilizable females; Emlen & Oring, 1977; Kvarnemo & Ahnesj€ o, 1996). Sexual selection has been shown to be affected in various ways by population density (Crowley et al., 1991; Eshel, 1979; Hubbell & Johnson, 1987; Kokko & Rankin, 2006) and OSR (Clutton-Brock & Parker, 1992; Emlen & Oring, 1977; Kokko, Klug, & Jennions, 2012; Kvarnemo & Ahnesj€ o, 1996; but see Head, Lindholm, & Brooks, 2007). On the one hand, the effects of OSR and density on competition for mate access are relatively straightforward: mate competition usually increases with density (e.g. crickets, Gryllus integer: Cade & Cade, 1992; guppies, * Correspondence: M.-J. Holveck and C. M. Nieberding, Universit e Catholique de Louvain (UCL), Earth and Life Institute (SST/ELI/ELIB), Biodiversity Research Centre, Carnoy building, Letter box L7.07.04, Croix du sud 4-5, 1348, Louvain-la-Neuve, Belgium. E-mail addresses: [email protected] (M. -J. Holveck), caroline. [email protected] (C. M. Nieberding). Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav http://dx.doi.org/10.1016/j.anbehav.2015.03.025 0003-3472/© 2015 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. Animal Behaviour 104 (2015) 229e245

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Animal Behaviour 104 (2015) 229e245
Contents lists avai
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
Dense, small and male-biased cages exacerbate maleemalecompetition and reduce female choosiness in Bicyclus anynana
Marie-Jeanne Holveck*, Anne-Laure Gauthier, Caroline M. Nieberding*
Group of Evolutionary Ecology and Genetics, Biodiversity Research Centre, Earth and Life Institute, Universit�e Catholique de Louvain (UCL),Louvain-la-Neuve, Belgium
a r t i c l e i n f o
Article history:Received 15 October 2014Initial acceptance 19 November 2014Final acceptance 18 March 2015Published onlineMS. number: 14-00824R
Keywords:Bicyclus anynanachoosinesscompetitiondensitymating preferencemating successsex ratio
* Correspondence: M.-J. Holveck and C. M. NieberdLouvain (UCL), Earth and Life Institute (SST/ELI/ELIB),Carnoy building, Letter box L7.07.04, Croix du sudBelgium.
E-mail addresses: marie-jeanne.holveck@[email protected] (C. M. Nieberding).
http://dx.doi.org/10.1016/j.anbehav.2015.03.0250003-3472/© 2015 The Association for the Study of A
Sexual selection is increasingly recognized to depend upon, and to fluctuate with, major ecologicalfactors in natural environments. The operational sex ratio (OSR) and population density can affect theopportunity for, and strength of, sexual selection but their effects are rarely taken into account in lab-oratory behavioural studies. In Bicyclus anynana, a model butterfly for mate choice studies, the experi-mental set-up widely used in laboratories often involves very high densities compared to the field, male-biased sex ratios and small cage volumes. We hypothesized that these conditions impede the properexpression of female mating preference by promoting male competition and sexual harassment of fe-males. Using various cage volumes, we separately manipulated OSR and density to cover the range ofvalues used in B. anynana laboratory mating experiments and to approach field values. Male competition,quantified by the number and duration of courtships aborted by males, became stronger with increasingdensities, specifically under more male-biased sex ratios, and decreasing cage volumes. Thus, maleeagerness to mate was essentially due to the use of small cage volumes and very high experimentaldensities. Concomitantly, female choosiness, quantified by the proportion of rejections of male matingattempts, decreased with increasing densities under a male-biased sex ratio. Females also accepted morematings with decreasing cage volume, and mated more rapidly with increasing density. We concludethat the laboratory social environments frequently used to test mate choice in B. anynana, with unnat-urally high densities and male-biased sex ratios, exacerbate maleemale competition and strongly hinderfemale choice, biasing the estimates of the strength and direction of sexual selection that were shapedunder very different natural environments in the wild. We propose a set-up for B. anynana mate choicestudies that allows the assessment of female choosiness in statistically robust mating experiments.© 2015 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Understanding the processes of pair formation is a necessary,yet neglected, step to fully grasp how secondary sexual traits andmating preferences evolve under sexual selection. It requires takinginto account the fact that sexual selection, like natural selection,takes place in ecologically complex environments and examiningthe two major selective agents of sexual selection, mate choice andwithin-sex competition, in an integrative way (Miller & Svensson,2014). It is increasingly acknowledged that mating patternsdepend on ecological and social constraints, which determine the
ing, Universit�e Catholique deBiodiversity Research Centre,4-5, 1348, Louvain-la-Neuve,
.be (M. -J. Holveck), caroline.
nimal Behaviour. Published by Els
availability of potential mating partners and, as a consequence,affect the expression of within-sex competition for mate access andof between-sex mating preferences (Miller & Svensson, 2014;Widemo & Saether, 1999). Social constraints include populationdensity (defined here as the number of individuals ready to mate ina given space) and the operational sex ratio (OSR, i.e. the ratio ofsexually active males to fertilizable females; Emlen & Oring, 1977;Kvarnemo & Ahnesj€o, 1996). Sexual selection has been shown tobe affected in various ways by population density (Crowley et al.,1991; Eshel, 1979; Hubbell & Johnson, 1987; Kokko & Rankin,2006) and OSR (Clutton-Brock & Parker, 1992; Emlen & Oring,1977; Kokko, Klug, & Jennions, 2012; Kvarnemo & Ahnesj€o, 1996;but see Head, Lindholm, & Brooks, 2007). On the one hand, theeffects of OSR and density on competition for mate access arerelatively straightforward: mate competition usually increases withdensity (e.g. crickets, Gryllus integer: Cade & Cade, 1992; guppies,
evier Ltd. All rights reserved.

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245230
Poecilia reticulata: Jirotkul, 1999b; soldier beetles, Chauliognathuspennsylvanicus: McLain, 1982) and with increasing proportions ofthe mate-limited sex (usually males; e.g. lobsters, Homerus gam-marus: Debuse, Addison, & Reynolds, 1999; Japanese medaka,Oryzias latipes: Grant, Bryant, & Soos, 1995; sand gobies, Pomato-schistus minutus: Kvarnemo, Forsgren, & Magnhagen, 1995; agilefrogs, Rana dalmatina: Lod�e, Holveck, Lesbarr�eres, & Pagano, 2004),which should lead to stronger sexual selection in the mate-limitedsex, namely to higher variance in mating success or few individualsmonopolizing more matings (OSR, Croshaw, 2010; Jones, Arguello,& Arnold, 2004; Klemme, Yl€onen,& Eccard, 2007; density, Kokko&Rankin, 2006). On the other hand, the effects of OSR and density onmating preference, and in particular on choosiness (i.e. the effortand energy devoted to mate assessment), are more complex(Jennions & Petrie, 1997; Widemo & Saether, 1999; Wong &Candolin, 2005). An OSR biased towards the mate-limited sex (i.e.usually increasing proportions of males) and high density maydecrease choosiness of the choosing sex (density, Arnqvist, 1992;Rowe, 1992; OSR, Lauer, Sih, & Krupa, 1996), via mating costssuch as increased harassment (Thornhill & Alcock, 1983) or sexualinterference (Schwagmeyer & Brown, 1983), and often correspondto frequent multiple mating (Lod�e, Holveck, & Lesbarr�eres, 2005;Lod�e et al., 2004; Rowe & Arnqvist, 2002; Rowe, Arnqvist, Sih, &Krupa, 1994; Uller & Olsson, 2008; but see these empiricalstudies: Laloi, Richard, Lecomte, Massot, & Clobert, 2004; Fitze, LeGalliard, Federici, Richard, & Clobert, 2005; and these modellingstudies: Bleu, Bessa-Gomes, & Laloi, 2012; H€ardling & Kaitala,2005). As such, a male-biased OSR and high density can decreasethe opportunity for, and strength of, sexual selection as thechoosing sex then mates more indiscriminately (e.g. OSR, Arnqvist,1992; Kokko et al., 2012; Krupa& Sih, 1993; density, Jirotkul,1999b;Mills & Reynolds, 2003; Pomfret & Knell, 2008; Rowe et al., 1994).The reverse may also be true: choosiness may increase withincreased OSR biases (Berglund, 1994; Gwynne & Simmons, 1990;Jirotkul, 1999a; Lawrence, 1986; Souroukis & Murray, 1995) anddensity (Gwynne, 1994; Palokangas, Alatalo, & Korpim€aki, 1992;Shelly & Bailey, 1992), for instance via increased variance in qual-ity of mates to choose from (Owens & Thompson, 1994) ordecreased cost of mate searching facilitating mate quality assess-ment (Crowley et al., 1991; reviewed in Kokko & Rankin, 2006),which should strengthen sexual selection. In addition, sex roles canbe reversed when the OSR is biased towards the choosing, and nottowards the competing, sex (Clark & Grant, 2010; Forsgren,Amundsen, Borg, & Bjelvenmark, 2004; Gwynne & Simmons,1990).
As shown above, OSR and density affect the opportunity for, andstrength of, sexual selection. Yet, there are two main caveats inmost studies published so far. First, the effects of both social con-straints are likely to be entangled as variation in OSR depends onthe relative densities of the sexes. One thus needs to manipulateOSR and density separately to be able to predict the consequencesof their potentially complex interactions on mate competition andmating preference. So far, few empirical studies have attempted todo so both in invertebrates (Alonso-Pimentel & Papaj, 1996;Arnqvist, 1992; Janowitz & Fischer, 2012; Saeki, Kruse, & Switzer,2005; Smith, 2007; Wang, He, Yang, Hedderley, & Davis, 2009;Wang, Yang, & Hedderley, 2008) and in vertebrates (Aronsen,Berglund, Mobley, Ratikainen, & Rosenqvist, 2013; Aronsen,Mobley, et al., 2013; Dreiss, Cote, Richard, Federici, & Clobert,2010; Elmberg, 1991; Head et al., 2007; de Jong, Wacker,Amundsen, & Forsgren, 2009; Lu, Ma, Fan, & Yu, 2010; Wackeret al., 2013). Second, it is crucial to test mate competition andmating preference in the social environment in which the pop-ulations evolved and adapted, which requires using the OSR anddensity encountered by natural populations in the wild (Fromhage,
Elgar, & Schneider, 2005; Fromhage, McNamara, & Houston, 2008;Kokko & Jennions, 2008).
Here we aimed to test whether the set-ups widely used inmating success experiments in the model butterfly Bicyclus any-nana promote sexual harassment of females by males and impedefemale mate choice, given that these set-ups probably differ fromthe natural conditions occurring in the field. This species is anexcellent model species to address the effects of varying OSR anddensity on the expression of maleemale competition and femalemating preferences. Indeed, sex roles in B. anynana switch fromconventional (with males competing for female access) toreversed sex roles across seasons (Prudic, Jeon, Cao, & Monteiro,2011). This suggests considerable flexibility in the matingbehaviour of both sexes, possibly caused by changes in OSR and/ordensity. In the wild, natural courtship in B. anynana involves aperch-and-chase strategy for mate location (Brakefield & Reitsma,1991; Breuker & Brakefield, 2002). We lack a detailed descriptionof male mate location strategies in the field, but both perch-and-chase and patrolling strategies are commonly observed in thelaboratory at all experimental densities (Joron & Brakefield, 2003;Nieberding et al., 2008; Prudic et al., 2011; Robertson &Monteiro,2005; this study). Perching males can be found locally at highdensities, where they are frequently involved in maleemalecompetition evidenced by circuit and chasing flights, such chasesbeing interrupted by repeated alightings (Brakefield & Reitsma,1991; Janowitz & Fischer, 2010; Joron & Brakefield, 2003). In thewild, the OSR is biased towards males and males can mate manytimes whereas females rarely mate more than once (Brakefield &Reitsma, 1991). There is limited available information regardingfield sex ratios and densities for B. anynana. The only publishedreport of field sex ratios of which we are aware comes from a 3-year survey in Malawi (Windig, Brakefield, Reitsma, & Wilson,1994) and mentions a sex ratio of 62% of males based on dailycapture in three bait traps. Our own field data from Uganda(Holveck, Gauthier, & Nieberding, 2013) provided an average sexratio of 84% of males (range 57e100%, N ¼ 1063 butterfliescollected once a day in 7e29 banana bait traps in six sites of0.25e6.02 ha for a total of 26 collection days). Regarding fielddensity, on average 47 Bicyclus butterflies were caught per ha persampling round (range 15e115) in Central Uganda, among whichan average of 19 butterflies/ha may belong to B. anynana species(range 8e35) considering an even species diversity (pooled datafrom three sampling methods: transect walk-and-counts, handnetting for 20e30 min per transect and banana bait trapping over2 consecutive days, Munyuli, 2012). Our own field data fromUganda (Holveck et al., 2013) produced a density of 42 ± 50B. anynana butterflies/ha per sampling round (mean ± 1SD; range1e158, N ¼ 1063 butterflies). The field density values reportedhere should be treated with caution since they strongly depend onthe sampling methods used to assess species richness (e.g. baittrapping is less than half as effective as capture by hand net;Munyuli, 2012), and were not estimated by captureerecaptureexperiments. These values nevertheless overlap estimates ofB. anynana population size by captureerecapture experiments inMalawi (with banana bait trapping or capture on natural fruit fallsfor 2 months) where field density was 3e13 butterflies/ha, asretrieved from the study of Brakefield and Reitsma (1991) aftercorrection by the relative density of B. anynana and Bicyclussafitza.
In the laboratory, numerous experimental studies of matingsuccess have recently been performed for B. anynana (30 publishedstudies; see Fig. 1). While the laboratory sex ratios (25e80% ofmales) fall within the range of field ones (62e84% of males; seeabove), most laboratory studies used a range of densities(0.0002e4 butterflies/dm3, which can be roughly converted to two

100(a)
(b)
17 610
15 2
1217 6
2 18
14 1431
3 26 2822
12 16 55307
13
14 14 31 4
14 2712 22 1
2419
19
1814
14 14
22
2731 29
272923
31
31
15
66
3125
2525 20
21
1515
151520
21
21
9
8
31
31
6
31
31
31
31
11
514
90
80
1515
15
1520
2121 21
20 25 25 25
31 31
31 31
31 31
6 6
31
23
11 3127
15 6
2826 3
276
510
6 17
2
3117
218 30 22
1215
55
13
14
14
14
16 12
14
19
1931
22
223118
27
2931
4
14
14
ReferencesLiterature surveyThis study
14
14 1 24
1412
729
8
9
−9 −7 −6 −5 −4 −3 −2 −1 0 1 2
60
50
40
30
20
10
0
100
90
80
70
60
50
40
30
20
10
40 6 8 10 12 14
70
Sex
rati
o (%
of
mal
es)
−8
2
Log-transformed density (individuals/dm3)
Log-transformed cage volume (dm3)
Figure 1. Sex ratio as a function of (a) densities and (b) cage volumes (as natural logarithms) in published B. anynana mating experiments and in our experiment. Source: Scopus,keywords ‘mating AND bicyclus anynana’ in ‘article title, abstract, keywords’ field, on 14 October 2014 and additional literature search. (1) Brakefield et al. (2001); (2) Breuker andBrakefield (2002); (3) Costanzo and Monteiro (2007); (4) Ferkau and Fischer (2006); (5) Fischer (2006); (6) Fischer et al. (2008); (7) Fischer, Zimmer, and Wedell (2009); (8)Frankino et al. (2005); (9) Frankino et al. (2007); (10) Geister and Fischer (2007); (11) Heuskin et al. (2014); (12) Janowitz and Fischer (2010); (13) Janowitz and Fischer (2011); (14)Janowitz and Fischer (2012); (15) Joron and Brakefield (2003); (16) Karl and Fischer (2013); (17) Karl et al. (2013); (18) Kehl et al. (2014); (19) Lewis and Wedell (2007); (20)Nieberding et al. (2012); (21) Nieberding et al. (2008); (22) Prudic et al. (2011); (23) Robertson and Monteiro (2005); (24) Saccheri and Brakefield (2002); (25) van Bergen et al.(2013); (26) Westerman and Monteiro (2013); (27) Westerman, Drucker, et al. (2014); (28) Westerman, Hodgins-Davis, Dinwiddie, & Monteiro (2012); (29) Westerman,Chirathivat, et al. (2014); (30) Zwaan, Zijlstra, Keller, Pijpe, and Brakefield (2008); (31) this study.
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 231
hundreds to four millions of butterflies per ha; Fig. 1a) much higherthan the natural variation occurring in the field (3e42 butterflies/ha; see above). Most laboratory studies also use small cage volumes(as low as 7 dm3; e.g. Janowitz & Fischer, 2010; Karl, Heuskin, &Fischer, 2013; Fig. 1b). In such experimental laboratory
conditions, the opportunity for ‘normal’ mate detection and chaseby males is effectively absent, and females might also be unable toreject copulation attempts by flying away from courting males(Joron & Brakefield, 2003). We hypothesize that the huge range ofenvironmental conditions in density, sex ratio and cage volume

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245232
used for B. anynana mating experiments may have led researchersto assess the effect of male eagerness to mate rather than femalemate choice on mating outcome and to discrepancies betweenpublications regarding the relative importance of male eagerness tomate (e.g. Fischer, Perlick, & Galetz, 2008; Karl et al., 2013) andfemale choosiness (Joron & Brakefield, 2003; Nieberding et al.,2008, 2012; van Bergen, Brakefield, Heuskin, Zwaan, &Nieberding, 2013) on mating outcome.
Here we thus tested the effects of the variation in densities andOSR across the range of values used in laboratory experiments,while trying to approach the (low) densities and (male-biased) sexratios encountered in the field (Fig. 1a), on maleemale competitionand female choosiness. We also used a range of cage volumes tomanipulate the ability of females to escape unwanted matings byflying away. We predicted that increasing densities, biasing the sexratio towards males or reducing cage volumes should increasemalecompetition for female access because of a reduced number of fe-males to pick per male or reduced opportunities for females toescape unwanted mating attempts. Stronger competition betweenmales should translate into more courtship attempts, more court-ships aborted by the male or decreased duration of courtshipsaborted by the male, the last two due to increased sexual inter-ference between competing males, all of which may affect matingfrequency. In response to increasing male competition, we ex-pected that female choosiness would either increase, as there aremore males to choose from, or decrease if male courtship activityleads to sexual harassment, given that the costs associated withresisting male harassment are likely to exceed mating benefits. Areduction in female choosiness should translate into a reduction inthe number of courtships aborted by the female, an increasedduration of courtships when aborted by the female (i.e. femalesshould take longer before escaping male courtship), an increasednumber of matings or a decreased duration of courtships endingwith a mating.
METHODS
Ethical Note
All procedures followed the ASAB/ABS Guidelines for the Useof Animals in Research and Belgium laws and involved an insectspecies for which a review by an institutional or governmentalregulatory body was not required. All B. anynana individualswere maintained in laboratory conditions as described in thedetailed protocol for rearing the species (Brakefield, Beldade, &Zwaan, 2009). We handled adults only to number them, whichtook less than 5 s, and to transfer them to the experimental cage.Butterflies behaved normally as soon as they were released in thecage. After the experiments, all butterflies were euthanized byfreezing (�20 �C). The parental generation was maintained in theclimate room (see below) in wet season conditions with amplefood and water until natural death or until euthanized byfreezing after egg collection for the subsequent generation wasover.
Insect Rearing
Experiments were performed with B. anynana from an outbredlaboratory stock population at Universit�e Catholique de Louvain(UCL) established from hundreds of eggs collected from a labora-tory colony in Yale (U.S.A.) itself originally established from 80gravid females collected in Malawi in 1988 (Brakefield & Reitsma,1991). Between 400 and 600 adults per generation have beenreared in each generation in order to preserve high levels of het-erozygosity (Van't Hof et al., 2005). Larvae are fed with maize, Zea
mays, and adults with moist banana, Musa acuminata (Brakefieldet al., 2009). Animals were reared, kept and tested in a largeclimate-controlled room (5.8 � 5.7 m and 2.27 m high) under astandard temperature regime (26 ± 2 �C), 12:12 h light:dark andhigh relative humidity (65 ± 5%) representing the wet season in thefield (Brakefield, 1997). For the experiment, we reared larvae ingroups of about 300 individuals in netted square cages of 200 dm3.On the day of their emergence, virgin adults were sexed, individ-ually marked on the ventral wingswith an indelible felt-tip pen andput in cylindrical hanging netted cages (diameter 30 cm, height38 cm, i.e. 26.9 dm3) with a density of 30 individuals of the samesex per cage.
Experimental Design
We assessed the effect of two social constraints, density and OSR,on the expression and relative importance of female choosiness andmale competition onmating outcome.We recordedmale and femalebehaviours in densities and sex ratios covering the range of experi-mental conditions previously used in all laboratories working withthis species (Fig. 1a). Thus, three densities (high: 0.44e0.45; inter-mediate: 0.065e0.067; low: 0.003e0.016 individuals/dm3) and threesex ratios (75, 50 and 33% of males) were tested in a crossed design.We accounted for the potential confounding effects of cage size onbutterfly behaviour by creating treatments with identical densitiesand sex ratios produced using variable numbers of individuals inthree cage volumes (small cylinder: p� 1.52 � 3.8 dm¼ 27 dm3;medium parallelepiped: 12� 6� 7 dm¼ 504 dm3; large parallele-piped: 420� 190� 215 dm¼ 17157 dm3; Table 1, Fig. 1b). Thedesign resulted in 14 different treatments and the number of repli-cates per treatment varied from one to four (Table 1). Potted maizeplants were present in the small (one plant), medium (four plants)and large experimental cages (10 plants) to allow perching andhiding. The 20 experimental trials were carried out with two cohortsfrom 25 April 2012 to 8 May 2012 and from 17 to 29 June 2012.
Experimental Procedure
We ran all experiments with the wet seasonal form as itshows rapid reproduction (Brakefield, Pijpe, & Zwaan, 2007) andhas been used by the vast majority of studies of mating success inB. anynana (i.e. in 28 of the 30 references in Fig. 1). We used8e10-day-old virgin males as all three components of the malesex pheromone, which is a major secondary sexual trait undersexual selection, are present at significant levels in the maleblend at this age (Heuskin et al., 2014; Nieberding et al., 2008,2012). We used 3e5-day-old virgin females as they readilymate in the laboratory (Brakefield et al., 2009; Nieberding et al.,2008, 2012). These ages also match those usually used inbehavioural experiments with this species (see references inFig. 1). All experimental trials occurred under full-spectrumlights and between 1600 and 1800 hours Central EuropeanTime, i.e. from 8.5 to 10.5 h after the light went on (at 0730hours), within the period of maximum courtship activity for thisspecies (Arun, San Martin, Brakefield, & Nieberding, 2014). Eachtrial lasted for 1.5 h during which the adults were placed in oneof the experimental cages. We first introduced the males into theexperimental area and left them to adapt and interact with oneanother for 1 h; we then added the females. Observations began10 min later. One of us (A.L.G.) simultaneously ran and sequen-tially observed a maximum of three experimental trials (eachfrom a different treatment; Table 1) per session, so that each trial(N ¼ 20) was observed for 10 min every 30 min, with a totalobservation duration of 30 min per trial. Direct behavioural

Table 1Sample size and occurrence of each behaviour per treatment, replicate, sex ratio, cage volume and density
TreatmentID
Replicate Sex ratio(% of males)
Cagevolume(dm3)
Density*
(individuals/dm3)Number ofindividuals
Number ofmale courtshipattempts
Number ofcourtships abortedby the male
Number ofcourtships abortedby the female
Number ofcourtships endingwith a mating
A 1 33 27 0.447 12 1 0 1 0A 2 33 27 0.447 12 6 0 5 1A 3 33 27 0.447 12 21 0 18 3A 4 33 27 0.447 12 21 0 19 2B 1 50 27 0.447 12 12 1 9 2C 1 75 27 0.447 12 13 6 6 1D 1 33 504 0.440 222 47 14 31 2E 1 76 504 0.456 230 88 14 71 3F 1 33 504 0.065 33 8 3 4 1G 1 50 504 0.067 34 32 8 23 1H 1 73 504 0.065 33 29 8 18 3I 1 33 504 0.012 6 1 1 0 0I 2 33 504 0.012 6 0 0 0 0J 1 50 504 0.008 4 2 0 2 0J 2 50 504 0.008 4 0 0 0 0K 1 75 504 0.016 8 6 1 5 0K 2 75 504 0.016 8 0 0 0 0L 1 33 17157 0.003 60 11 5 4 2M 1 50 17157 0.003 60 26 3 20 3N 1 75 17157 0.006 100 61 13 48 0d d d d Total 880 385 77 284 24
* Given constraints regarding sex ratio adjustment and availability of butterflies in the selected age range, it was not possible to get the exact same density for each densitycategory (high: 0.44e0.45; intermediate: 0.065e0.067; low: 0.003e0.016 individuals/dm3).
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 233
observations were encoded using the software The Observer 5.0(Noldus Information Technology, Wageningen, The Netherlands).
Behavioural Variables
We selected a series of courtship variables that best allowed usto assess competition for mate access and choosiness (Table 2). InB. anynana, only males initiate courtship in the wet seasonal form(Prudic et al., 2011) and the number of male courtship attempts wasused as a first indicator of male motivation to mate. We also ana-lysed the data using the courtship rate per male (see the Appendix),which yielded the same results. A courtship could be aborted by thecourting male (‘number of courtships aborted by the male’ here-after): this mostly occurred when the courting pair was disturbedby sexual interference from other males (personal observation),thus reflecting strong male competition, but it may also indicate ahigh level of male choosiness when the male stops courting lesspreferred females. A courtship could also be aborted by the femaleescaping (‘number of courtships aborted by the female’ hereafter),which probably reflects her high level of choosiness (e.g. Joron &Brakefield, 2003). Finally, a courtship could also end with a mat-ing (‘number of courtships ending with a mating’ hereafter) givingthe mating success. We used as response variables the proportions
Table 2Recorded behavioural variables together with their predicted significance in relation tomale competition and female choosiness under increasing density, sex ratio or decreasin
Variables Behavioural proxy
Number of male courtship attempts Male competition for matesNumber of courtships aborted by the male Male competition for mates (
OR male choosinessNumber of courtships aborted by the female Female choosinessNumber of courtships ending with a mating Male competition for mates ADuration of courtships aborted by the male Male competition for mates (
OR male choosinessDuration of courtships aborted by the female Female choosinessDuration of courtships ending with a mating Male competition for mates A
of courtships either aborted by the male, aborted by the female orending with a mating.
We also recorded the duration of courtships either aborted bythe male, aborted by the female or ending with a mating. A shortcourtship aborted by the male should reflect high male choosinessor strong male competition if the male is interrupted by othermales, in which case we do not know the level of female compe-tition and choosiness. A short courtship aborted by the femaleshould reflect high female choosiness, in which case we do notknow male competition and choosiness. While we cannot infermuch from short courtships ending with a mating, an increase inthe length of male courtships ending with a mating should reflectan increase in female choosiness (i.e. themale needs to complete alltypical steps of the courtship before the female acceptsmatingwithhim) and high mate competition for males (i.e. males keep tryingdespite unsuccessful previous courtship trials; Nieberding et al.,2008).
Statistical Analyses
Data were analysed in R v 3.1.1 (R Core Team, 2014) with a two-tailed (a ¼ 0.05) generalized linear model (GLM) or a generalizedlinear mixed model (GLMM; lme4 package) with a quasi-Poissondistribution accounting for overdispersion for the number of male
male competition and female choosiness, and with their predicted correlation withg cage volume
Predicted correlation
Positiveby means of sexual interference between males) Positive
PositiveND female choosiness Positive and negativeby means of sexual interference between males) Negative
NegativeND female choosiness Positive

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245234
courtship attempts, and with a quasi- or binomial distribution forthe proportions of courtships ending with matings, female escapesor male stops. The latter allowed us to integrate the number of malecourtship attempts into the analyses with the ‘cbind’ function(number of successes: the number of times one of the three types ofbehaviour, mating, escaping or aborting, ended a courtship; num-ber of failures: the number of times the other two types ofbehaviour ended a courtship). The three courtship duration vari-ables were analysed with linear mixed models (LMM, nlme pack-age) fitted by restricted maximum likelihood and followingBoxeCox transformations for normality reasons. All P > 0.05 for theShapiroeWilk tests on linear model residuals.
Density is a composite measure of number of individuals andcage volume, but our crossed design (Table 1) together with the useof cage volume residuals (see details below) allowed us to analysethe effects of these three factors simultaneously. We used the re-siduals of the linear regression of cage volume on density andnumber of individuals (following log transformation of the threevariables for normality reasons and to avoid potential outlier ef-fects) given that cage volume was correlated with both density andnumber of individuals (Spearman correlation: rS ¼ �0.82, P < 0.001and rS ¼ 0.32, P ¼ 0.17, respectively, for N ¼ 20 trials; rS ¼ �0.83,P < 0.001 and rS ¼ 0.51, P ¼ 0.04, respectively, for N ¼ 17 trials withobserved behaviours; see Table 1), while the latter were notcorrelated with each other (rS ¼ 0.16, P ¼ 0.5, N ¼ 20 trials;rS ¼ �0.05, P ¼ 0.9, N ¼ 17 trials).
All models were of the following form:Y ~ X1 � X2 þ X3 � X2 þ X4 þ random factor(s), with Y ¼ each of theseven response variables, X1 ¼ density, X2 ¼ sex ratio (expressed asa percentage of males; Table 1), X3 ¼ cage volume residuals,X4 ¼ number of individuals (included as a covariate to account forits variability between treatments) and random factor¼ treatmentID (N ¼ 14, Table 1) to account for replicates. In GLMMs, we addedan observation level random effect to capture the overdispersionwhen needed (e.g. Elston, Moss, Boulinier, Arrowsmith, & Lambin,2001; Holveck & Riebel, 2014). In LMMs, we also added a secondrandom factor, i.e. experimental trial (N ¼ 20, Table 1), to accountfor pseudoreplication. Full models went through a stepwise back-ward selection procedure, first on the random factor treatment ID,and then on fixed factors to reach the minimal adequate model. Wekept the model with the best fit based on the log-likelihood ratiotest. We removed treatment ID from analyses when P > 0.1. Wecalculated the variance inflation coefficients VIF to diagnose po-tential multicollinearity problems among predictor variables (Zuur,Ieno, & Elphick, 2010) in all models: all VIF were lower than 1.5showing no notable multicollinearities. Nevertheless, variableswere not fully independent (all 1.04 > VIF > 1.42), which is why wetested eachmain effect after adjusting for all othermain effects, andfitted the interaction termwhile all main effects were in the model(e.g. Alonso-Pimentel& Papaj, 1996; Wang et al., 2008). In addition,we also ensured the absence of type I errors (in comparing thesignificance level of a variable tested in isolation or together withall other main effects), which can occur when model simplificationis applied to models that are overfitted before simplification(Forstmeier & Schielzeth, 2011; Mundry & Nunn, 2009). We alsoreanalysed all datawith an information-basedmethod, which givessimilar output (see Appendix). We report estimates ± 1 SE of eachfactor together with the statistics (see Tables 3, 4) of the compari-son between the models with and without the tested factor. Allother reported means are ± 1 SD.
RESULTS
In total, we observed 385 courtship attempts among which 284were aborted by the female, 77 were aborted by the male and 24
ended with a mating, namely 70 ± 25%, 23 ± 25% and 8 ± 7%,respectively (N ¼ 17 trials; Table 1). Therefore the courtships mostoften ended with the female escaping, and more often by malestops than by matings (chi-square test for given probabilities:c2
2 ¼ 294, P < 0.001; all Holm corrected P < 0.001 for paired com-parisons). The courtships aborted by the female were significantlyshorter (4.53 ± 11.65 s, N ¼ 284) than courtships either aborted bythe male (8.56 ± 13.73 s, N ¼ 77) or ending with a mating(9.59 ± 9.89 s, N ¼ 24; log-likelihood ratio test: c2
2 ¼ 57, P < 0.001,on LMMs with or without the tested factor; Tukey contrasts formultiple comparisons: both P < 0.001).
Male Competition
The number of male courtship attempts increased significantlywith the number of individuals and tended to increase underincreasing male-biased sex ratios (Table 3, Fig. 2a, b). The propor-tion of courtships aborted by the male depended on density and onits interactionwith sex ratio (Table 3): at low densitymales stoppedcourting more often under a female-biased than a male-biased sexratio, while at high density males stopped courting more oftenunder a male-biased than a female-biased sex ratio (Fig. 3a). Theproportion of courtships aborted by the male also decreased whencage volume increased, mostly under female-biased and unbiasedsex ratios (Fig. 3b, Table 3). In addition, males aborted theircourtship more rapidly with increasing density under all sex ratios(Table 4, Fig. 4a). Altogether, these results indicate that malecompetition became stronger with increasing densities, in partic-ular under more male-biased sex ratios and decreasing cage vol-ume. Note that the effect involving cage volume can be attributed tothe variation in cage volume only as we used its residuals againstdensity and individual number.
Female Choosiness
The proportion of courtships aborted by the female dependedon density and on its interaction with sex ratio (Table 3): femalesescaped more often under a male-biased or an unbiased sex ratiothan a female-biased sex ratio at the lowest densities, and escapedmore oftenwith increasing density under a female-biased sex ratiobut less often with increasing density under a male-biased or anunbiased sex ratio (Fig. 3c). In addition, females took longer toescape courtships when density decreased (Table 4), and this wastrue in interaction with sex ratio, under a female-biased sex ratio(Table 4, Fig. 4b). Altogether, these results indicate that femaleswere choosier at lower densities under a male-biased sex ratio, andfemale choosiness was minimal at low density under a female-biased sex ratio.
Mating Success
Concomitantly with the variation in male competition (court-ships aborted by the male) and female choosiness (courtshipsaborted by the female), the proportion of courtships ending with amating depended on the interaction between cage volume and sexratio (Table 3): mating proportion decreased with increasing cagevolume under a male-biased sex ratio, while it increased withincreasing cage volume under a female-biased and an unbiased sexratio (Fig. 3d). The duration of courtships ending with a mating waslonger under a male-biased than a female-biased sex ratio at thelowest densities, and increased with density under a female-biasedsex ratio but decreased with density under a male-biased sex ratio(Table 4, Fig. 4c).
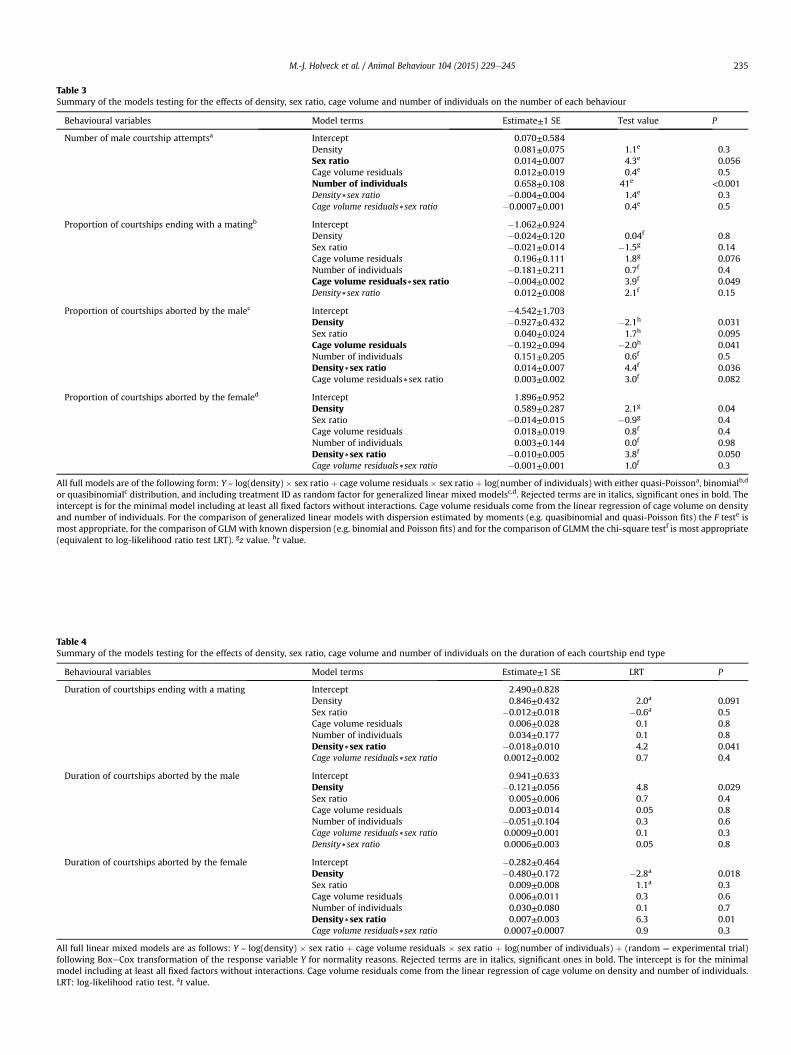
Table 3Summary of the models testing for the effects of density, sex ratio, cage volume and number of individuals on the number of each behaviour
Behavioural variables Model terms Estimate±1 SE Test value P
Number of male courtship attemptsa Intercept 0.070±0.584Density 0.081±0.075 1.1e 0.3Sex ratio 0.014±0.007 4.3e 0.056Cage volume residuals 0.012±0.019 0.4e 0.5Number of individuals 0.658±0.108 41e <0.001Density)sex ratio �0.004±0.004 1.4e 0.3Cage volume residuals)sex ratio �0.0007±0.001 0.4e 0.5
Proportion of courtships ending with a matingb Intercept �1.062±0.924Density �0.024±0.120 0.04f 0.8Sex ratio �0.021±0.014 �1.5g 0.14Cage volume residuals 0.196±0.111 1.8g 0.076Number of individuals �0.181±0.211 0.7f 0.4Cage volume residuals)sex ratio �0.004±0.002 3.9f 0.049Density)sex ratio 0.012±0.008 2.1f 0.15
Proportion of courtships aborted by the malec Intercept �4.542±1.703Density �0.927±0.432 �2.1h 0.031Sex ratio 0.040±0.024 1.7h 0.095Cage volume residuals �0.192±0.094 �2.0h 0.041Number of individuals 0.151±0.205 0.6f 0.5Density)sex ratio 0.014±0.007 4.4f 0.036Cage volume residuals)sex ratio 0.003±0.002 3.0f 0.082
Proportion of courtships aborted by the femaled Intercept 1.896±0.952Density 0.589±0.287 2.1g 0.04Sex ratio �0.014±0.015 �0.9g 0.4Cage volume residuals 0.018±0.019 0.8f 0.4Number of individuals 0.003±0.144 0.0f 0.98Density)sex ratio �0.010±0.005 3.8f 0.050Cage volume residuals)sex ratio �0.001±0.001 1.0f 0.3
All full models are of the following form: Y ~ log(density) � sex ratio þ cage volume residuals � sex ratio þ log(number of individuals) with either quasi-Poissona, binomialb,d
or quasibinomialc distribution, and including treatment ID as random factor for generalized linear mixed modelsc,d. Rejected terms are in italics, significant ones in bold. Theintercept is for the minimal model including at least all fixed factors without interactions. Cage volume residuals come from the linear regression of cage volume on densityand number of individuals. For the comparison of generalized linear models with dispersion estimated by moments (e.g. quasibinomial and quasi-Poisson fits) the F teste ismost appropriate, for the comparison of GLM with known dispersion (e.g. binomial and Poisson fits) and for the comparison of GLMM the chi-square testf is most appropriate(equivalent to log-likelihood ratio test LRT). gz value. ht value.
Table 4Summary of the models testing for the effects of density, sex ratio, cage volume and number of individuals on the duration of each courtship end type
Behavioural variables Model terms Estimate±1 SE LRT P
Duration of courtships ending with a mating Intercept 2.490±0.828Density 0.846±0.432 2.0a 0.091Sex ratio �0.012±0.018 �0.6a 0.5Cage volume residuals 0.006±0.028 0.1 0.8Number of individuals 0.034±0.177 0.1 0.8Density)sex ratio �0.018±0.010 4.2 0.041Cage volume residuals)sex ratio 0.0012±0.002 0.7 0.4
Duration of courtships aborted by the male Intercept 0.941±0.633Density �0.121±0.056 4.8 0.029Sex ratio 0.005±0.006 0.7 0.4Cage volume residuals 0.003±0.014 0.05 0.8Number of individuals �0.051±0.104 0.3 0.6Cage volume residuals)sex ratio 0.0009±0.001 0.1 0.3Density)sex ratio 0.0006±0.003 0.05 0.8
Duration of courtships aborted by the female Intercept �0.282±0.464Density �0.480±0.172 �2.8a 0.018Sex ratio 0.009±0.008 1.1a 0.3Cage volume residuals 0.006±0.011 0.3 0.6Number of individuals 0.030±0.080 0.1 0.7Density)sex ratio 0.007±0.003 6.3 0.01Cage volume residuals)sex ratio 0.0007±0.0007 0.9 0.3
All full linear mixed models are as follows: Y ~ log(density) � sex ratio þ cage volume residuals � sex ratio þ log(number of individuals) þ (random ¼ experimental trial)following BoxeCox transformation of the response variable Y for normality reasons. Rejected terms are in italics, significant ones in bold. The intercept is for the minimalmodel including at least all fixed factors without interactions. Cage volume residuals come from the linear regression of cage volume on density and number of individuals.LRT: log-likelihood ratio test. at value.
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 235

(a) (b)5
3
2
1
0
5
3
2
1
0
1 2
R2 = 0.66
R2 = 0.05
Sex ratio
4 5 6 30 40 50 60 70 80
4N
um
ber
of m
ale
cou
rtsh
ip a
ttem
pts
3
Number of individuals
4
Figure 2. Number of male courtship attempts (as natural logarithm) as a function of (a) the number of individuals (as natural logarithm) and (b) sex ratio. Linear regression lines areshown with their corresponding R2 values. Jitter range is 0.1 on the x-axis and 0.07 on the y-axis to see overlapping points.
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245236
DISCUSSION
The aim of this study was to investigate whether variation indensity, sex ratios and cage volumes used in B. anynana laboratorybehavioural experiments affect female choosiness and malecompetition, two parameters that determine the evolution of malesecondary sexual traits and of female mating preference in nature.Our results show, consistently for most behavioural variables, aninteraction of sex ratio with density or cage volume on behaviouralvariables associated with male competition and female choosiness.Male competition became stronger with increasing densities, inparticular under more male-biased sex ratios, and decreasing cagevolumes. Female choosiness, quantified by female escapingbehaviour, became weaker with increasing densities under a male-biased sex ratio and stronger with increasing densities under afemale-biased sex ratio.
Classical sexual selection theory (Emlen & Oring, 1977) pre-dicts that sexual selection should be stronger under sex ratiosbiased towards the mate-limited sex (traditionally considered asbeing the males), as revealed by a higher variance in matingsuccess of males, or few males monopolizing a lot of females. Yet,this pattern is not consistent across studies which suggests thatthe opportunity for, and strength of, sexual selection may stronglydepend upon density, once density and sex ratio effects areproperly disentangled (e.g. Arnqvist, 1992; Aronsen, Berglund,et al., 2013; Dreiss et al., 2010; Elmberg, 1991; de Jong et al.,2009; Lu et al., 2010; Wang et al., 2009). Our results revealedthat density, either alone or in interaction with sex ratio, had asignificant effect on male and female behavioural traits moreoften than sex ratio (Tables 3, 4). Modern sexual selection theoryalso indicates that reproductive (mating or breeding) costs may bea more direct predictor of the intensity of sexual selection thanOSR (empirically, Dreiss et al., 2010; Fitze & Le Galliard, 2008;theoretically, Kokko & Johnstone, 2002; Kokko & Monaghan,2001): if female mating costs counteract or exceed the matingbenefits, female choosiness may remain stable or decrease,respectively, under high densities or a male-biased sex ratio. Inthis respect, we found that B. anynana males tended to increasetheir courtship effort (i.e. number of attempts) with increasingbias towards males, as expected by a wealth of empirical andtheoretical studies (empirical, e.g. Carrillo, Danielson-François,
Siemann, & Meffert, 2012; Forsgren et al., 2004; Head et al.,2007; de Jong, Forsgren, Sandvik, & Amundsen, 2012; Wackeret al., 2013; theoretical, Emlen & Oring, 1977; Kokko & Rankin,2006; Kvarnemo & Ahnesj€o, 1996). As male courtship efforts didnot depend on density, we inferred that the increase in malecourtship was due to a higher relative number of males and not toa higher mate encounter rate, which shows that the level ofcourtship activity was directly due to the level of maleemalecompetition. Importantly, exacerbation of male competition withincreasing density under a male-biased sex ratio (as quantified bythe increasing number, and decreasing duration, of courtshipsaborted by males) had a negative effect on the opportunity forsexual selection: female ability to escape from male mating at-tempts decreased with increasing density, females accepted morematings with decreasing cage volume, and mated more rapidlywith increasing density. Therefore, our results agree with thetheoretical prediction that if females suffer from increasedharassment for copulation (i.e. more courtship attempts andsexual interference) in male-biased or high-density populations(Kokko & Johnstone, 2002; Kokko & Monaghan, 2001; Kokko &Rankin, 2006), the cost of choosiness for females (i.e. repeatedlyescaping male courtship attempts) may increase and females maybecome more likely to accept copulations indiscriminately toavoid harassment. Evidence has accumulated in insects that fe-males become less choosy with increased density and male-biased sex ratio (density, Arnqvist, 1992; Rowe, 1992; OSR, Laueret al., 1996) and that females find it more challenging todiscriminate between secondary sexual traits when they are facedwith a greater number of signalling individuals (Hutchinson,2005). In addition, sexual interference in stopping or delayingmating may reduce the mating success of rivals while improvingthat of the harassers through increased access to reproductivepartners (e.g. Deno€el, 2003), and, under such conditions, alter-native mating tactics (e.g. coercive or manipulative) such assneaking copulations can circumvent mate choice (Byrne &Roberts, 2004; Crespi, 1988; Lod�e et al., 2004; Shine, Langkilde,& Mason, 2003), thus potentially counteracting sexual selectionresulting from conventional mating tactics (Kokko & Rankin,2006). Consequently, under extreme demographic/social con-straints such as those often used for B. anynana in the laboratory(Fig. 1), there is a risk that the variation in mating success between

1 (a) (b)
(c) (d)
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
−6 −16 −9 −2 5 12−5 −3 −2 −1 0
−6 −5 −3
Cage volume
−2 −1 0
R2 = 0.07
R2 = 0.23
R2 = 0.28
R2 = 0.73
R2 = 0.62
R2 = 0.07
R2 = 0.3
R2 = 0.2
R2 = 0.61R2 = 0.22
R2 = 0.04
R2 = 0.21
1
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
1
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0.2
0.1
0
0
−16 −9 −2 5 12
Prop
orti
on o
f co
urt
ship
s ab
orte
d b
y th
e fe
mal
e
Prop
orti
on o
f co
urt
ship
s en
din
g w
ith
a m
atin
g
0.9Pr
opor
tion
of
cou
rtsh
ips
abor
ted
by
the
mal
e0.9
Prop
orti
on o
f co
urt
ship
s ab
orte
d b
y th
e m
ale
−4
−4
Density
Figure 3. Proportion of courtships (a) aborted by the male in relation to density (as natural logarithm), (b) aborted by the male in relation to cage volume (as residuals againstdensity and number of individuals), (c) aborted by the female in relation to density (as natural logarithm) and (d) ending with a mating in relation to cage volume (as residuals) persex ratio (male-biased: pale grey circles and dotted line; equal: dark grey circles and dashed line; female-biased: black). Linear regression lines with their corresponding R2 valuesare shown per sex ratio. Jitter range is 0.1 on the x-axis and 0.0007 on the y-axis to see overlapping points. Note the smaller y-axis scale of (d) as compared to the others.
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 237
males becomes uncoupled from the variation in mate quality, thusreducing the opportunity and strength of sexual selection (e.g.OSR, Arnqvist, 1992; Kokko et al., 2012; Krupa& Sih,1993; density,Jirotkul, 1999b; Mills & Reynolds, 2003; Pomfret & Knell, 2008;Rowe et al., 1994).
In B. anynana, there has long been a general agreement thatfemale choice should prevail in the wet seasonal form (van Bergenet al., 2013; Brakefield et al., 2001; Costanzo & Monteiro, 2007;Frankino, Zwaan, Stern, & Brakefield, 2007; Nieberding et al.,2008, 2012; Prudic et al., 2011; Robertson & Monteiro, 2005;Westerman, Chirathivat, Schyling, & Monteiro, 2014;Westerman, Drucker, & Monteiro, 2014; Westerman, Hodgins-Davis, Dinwiddie, & Monteiro, 2012; Westerman & Monteiro,2013). This is notably based on the facts that females do rejectcourting males, rarely mate more than once in the field and in thelaboratory (van Bergen et al., 2013; Brakefield et al., 2001;Brakefield & Reitsma, 1991; Joron & Brakefield, 2003;Nieberding et al., 2008, 2012), and gain neither direct benefits(no evidence for nuptial gifts, Ferkau & Fischer, 2006; M€olleman,Zwaan,& Brakefield, 2004; but see Prudic et al., 2011; Westerman,
Drucker, et al., 2014, who suggest that females may receive abeneficial nuptial gift from dry season males) nor fitness benefitsfrom multiple matings while suffering survival costs (Fischer,2007). Yet, other research in B. anynana supports the hypothesisthat male behaviour (male sexual harassment, male competition)may be a stronger determinant of mating outcome than femalechoice, as evidenced by male courtship persistence and aggres-siveness in ‘latching onto the female's abdomen to copulate’ (e.g.Fischer et al., 2008; Geister & Fischer, 2007; Janowitz & Fischer,2010, 2012; Karl et al., 2013; Kehl et al., 2014). We suggest thatour results provide an explanation for this seemingly oppositepattern regarding the relative importance of female choice versusmale courtship activity on mating outcome across B. anynanamating experiments. Our results showed that the percentage ofcourtships leading to mating varied from 0 to 20% depending oncage volume and sex ratio in our experiments. This is not anec-dotal since our experiments occurred at lower densities and inlarger cages than the lowest densities and largest cages, respec-tively, used by some other teams (roughly half of the studies;Fig. 1). Moreover, we clearly showed that the importance of male
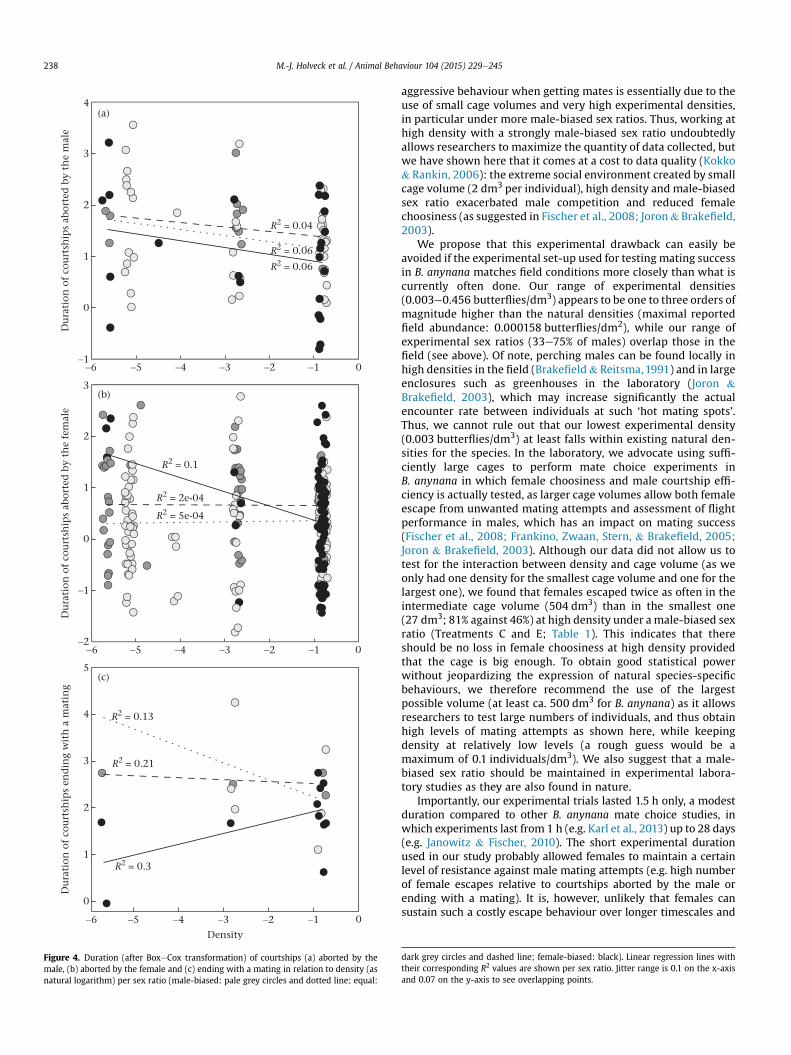
4(a)
(b)
(c)
2
1
0
3
2
1
0
−1
−2
5
4
3
2
1
0
−1−6 −4 −3 −2 −1 0
R2 = 0.04
R2 = 0.06
R2 = 0.06
R2 = 0.1
R2 = 2e-04
R2 = 5e-04
R2 = 0.13
R2 = 0.21
R2 = 0.3
−6 −4 −3 −2 −1 0
−6 −4 −3 −2 −1 0
Du
rati
on o
f co
urt
ship
s ab
orte
d b
y th
e fe
mal
eD
ura
tion
of
cou
rtsh
ips
end
ing
wit
h a
mat
ing
3
Du
rati
on o
f co
urt
ship
s ab
orte
d b
y th
e m
ale
−5
−5
−5
Density
Figure 4. Duration (after BoxeCox transformation) of courtships (a) aborted by themale, (b) aborted by the female and (c) ending with a mating in relation to density (asnatural logarithm) per sex ratio (male-biased: pale grey circles and dotted line; equal:
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245238
aggressive behaviour when getting mates is essentially due to theuse of small cage volumes and very high experimental densities,in particular under more male-biased sex ratios. Thus, working athigh density with a strongly male-biased sex ratio undoubtedlyallows researchers to maximize the quantity of data collected, butwe have shown here that it comes at a cost to data quality (Kokko& Rankin, 2006): the extreme social environment created by smallcage volume (2 dm3 per individual), high density and male-biasedsex ratio exacerbated male competition and reduced femalechoosiness (as suggested in Fischer et al., 2008; Joron& Brakefield,2003).
We propose that this experimental drawback can easily beavoided if the experimental set-up used for testing mating successin B. anynana matches field conditions more closely than what iscurrently often done. Our range of experimental densities(0.003e0.456 butterflies/dm3) appears to be one to three orders ofmagnitude higher than the natural densities (maximal reportedfield abundance: 0.000158 butterflies/dm2), while our range ofexperimental sex ratios (33e75% of males) overlap those in thefield (see above). Of note, perching males can be found locally inhigh densities in the field (Brakefield& Reitsma,1991) and in largeenclosures such as greenhouses in the laboratory (Joron &Brakefield, 2003), which may increase significantly the actualencounter rate between individuals at such ‘hot mating spots’.Thus, we cannot rule out that our lowest experimental density(0.003 butterflies/dm3) at least falls within existing natural den-sities for the species. In the laboratory, we advocate using suffi-ciently large cages to perform mate choice experiments inB. anynana in which female choosiness and male courtship effi-ciency is actually tested, as larger cage volumes allow both femaleescape from unwanted mating attempts and assessment of flightperformance in males, which has an impact on mating success(Fischer et al., 2008; Frankino, Zwaan, Stern, & Brakefield, 2005;Joron & Brakefield, 2003). Although our data did not allow us totest for the interaction between density and cage volume (as weonly had one density for the smallest cage volume and one for thelargest one), we found that females escaped twice as often in theintermediate cage volume (504 dm3) than in the smallest one(27 dm3; 81% against 46%) at high density under a male-biased sexratio (Treatments C and E; Table 1). This indicates that thereshould be no loss in female choosiness at high density providedthat the cage is big enough. To obtain good statistical powerwithout jeopardizing the expression of natural species-specificbehaviours, we therefore recommend the use of the largestpossible volume (at least ca. 500 dm3 for B. anynana) as it allowsresearchers to test large numbers of individuals, and thus obtainhigh levels of mating attempts as shown here, while keepingdensity at relatively low levels (a rough guess would be amaximum of 0.1 individuals/dm3). We also suggest that a male-biased sex ratio should be maintained in experimental labora-tory studies as they are also found in nature.
Importantly, our experimental trials lasted 1.5 h only, a modestduration compared to other B. anynana mate choice studies, inwhich experiments last from 1 h (e.g. Karl et al., 2013) up to 28 days(e.g. Janowitz & Fischer, 2010). The short experimental durationused in our study probably allowed females to maintain a certainlevel of resistance against male mating attempts (e.g. high numberof female escapes relative to courtships aborted by the male orending with a mating). It is, however, unlikely that females cansustain such a costly escape behaviour over longer timescales and
dark grey circles and dashed line; female-biased: black). Linear regression lines withtheir corresponding R2 values are shown per sex ratio. Jitter range is 0.1 on the x-axisand 0.07 on the y-axis to see overlapping points.

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 239
particularly in small cages at high densities and under male-biasedsex ratios (e.g. Janowitz & Fischer, 2012) because escape requiresexplosive take-off acceleration performance from resting, which isone of themost energetically demanding types of aerial locomotion(Dudley, 2000). We thus also recommend working on short time-scales (a few hours to a couple of days), in particular at high den-sities and strongly male-biased sex ratios, as we showed here thatin such conditions, at least for 1.5 h, female mate choice could stillbe expressed to some extent. Alternatively, we recommenddecreasing density and increasing cage volume if experimentalduration is increased to days (as in Joron & Brakefield, 2003;Nieberding et al., 2008, 2012).
Of note, we quantified male competition using two behaviouralproxies, namely the number of male courtship attempts and thenumber of courtships aborted by males, i.e. the number of malesexual interferences. Here, we want to acknowledge aweakness inour behavioural scoring, namely that we did not record whetherthe abortion of the courtships by the courting males did or did notoccur following sexual interference by other males; we thusadvocate distinguishing clearly between these two behaviours asthey probably reflect different selective agents, namely malecompetition in the case of sexual interference versus male matechoice. We found that the number of interferences increasedwhen density increased under a male-biased sex ratio. In natureand in the laboratory, male competition may be affected by thestrategy used by males to locate females, namely a perch-and-chase strategy, i.e. males taking off to follow a passing female, ora patrolling strategy, i.e. take-off and flying in search of perchedfemales (Brakefield & Reitsma, 1991; Joron & Brakefield, 2003;Nieberding et al., 2008; Prudic et al., 2011; Robertson &Monteiro, 2005; this study). There is no clear association be-tween these two strategies and the propensity of males to courtfemales or to interrupt their courtship as both behaviours may beperformed by either perching-and-chasing or patrolling males.Very high density used in the laboratory may break the perching/site defence strategies and affect male competition as observed inthe wild at high density where individual males can switch flex-ibly between two or more mating strategies (e.g. in butterflies,Alcock, 1994; Alcock & O'Neill, 1986; in ungulates, Isvaran, 2005).There are no such data to our knowledge for B. anynana, but thedensity threshold at which such breakdown occurs is likely to bevery high in this species as in other butterflies in which territorydefence seems to be adopted whenever possible and indepen-dently of density (Bergman et al., 2007; Bergman & Wiklund,2009a, 2009b; Kemp, 2001; Peixoto & Benson, 2009). Absence offield data quantifying the different types of male behavioursprevents us from comparing our laboratory results with the dy-namics of mate competition in the wild.
Our results also suggest that males chose which females tocourt under the female-biased sex ratio. First, males interruptedtheir courtship a lot at low densities, as compared to a similardensity with higher relative number of males, which is unlikelyto be due to sexual interference between males at such lowdensity, and may instead indicate that males increased theireffort in assessing multiple females (i.e. increased male choosi-ness). Second, males strongly decreased their persistency tocourt with increasing densities, suggesting that males did notcourt indiscriminately at higher density despite a higher mateencounter rate. Such male mate choice, expressed throughcourtship preferences, has already been reported in some species(Hebets & Sullivan-Beckers, 2010), was previously suggested inB. anynana under a female-biased sex ratio (Westerman, Drucker,et al., 2014) and can even sometimes occurs when OSR is skewedtowards males (Servedio, 2007). Nevertheless, in our study whenthe OSR was biased towards females, sex role did not seem to be
totally reversed as female choosiness appeared to increase withincreasing densities: females escaped more often and morerapidly from male mating attempts with increasing density.Therefore, mutual mate choice appeared to vary with densityunder the female-biased sex ratio in our study, suggesting thatmutual mate choice in B. anynana occurs in the wet seasonalform and not only at the level of the seasonal sex role reversal ofthe species (Prudic et al., 2011; Westerman, Chirathivat, et al.,2014).
We conclude that unnaturally high densities and sex ratios infavour of males may strongly bias estimates of the strength anddirection of sexual selection in B. anynana, provided that the re-ported effects of social factors on mating behaviour and success doaffect the opportunity for sexual selection and the selection onmale secondary sexual traits (e.g. Wacker et al., 2013; but see Headet al., 2007). Although studies using demographic factors outsidethe natural species-specific range provide valuable information ontheir possible effects on sexual selection, they provide little infor-mation on how important these effects are in shaping secondarysexual traits and sexual preferences for them in natural pop-ulations, and, in particular for B. anynana, on the evolution of thetwo sexually selected traits, dorsal forewing eyespot pupils and sexpheromone composition (Costanzo & Monteiro, 2007; Nieberdinget al., 2008, 2012; Robertson & Monteiro, 2005; van Bergen et al.,2013).
Acknowledgments
We thank two anonymous referees for their comments on themanuscript and Christophe Pels for help with the rearing. Theproject was supported by the Universit�e Catholique de Louvain UCL(Grant ARC 10/15-031 Acad�emie Louvain and FSR grant 372 605031to C.M.N., and construction of the environmentally friendly insec-tarium for B. anynana) and by the FRS-FNRS Belgian National Sci-ence Foundation (FRFC grant 2.4560.11 and CDR grant FNRS CDRJ.0069.14 to C.M.N.). M.J.H. is a FRS-FNRS postdoctoral fellow andA.L.G. a FRIA Ph.D. student. This is publication BRC 335 of theBiodiversity Research Centre (UCL).
References
Aho, K., Derryberry, D., & Peterson, T. (2014). Model selection for ecologists: theworldviews of AIC and BIC. Ecology, 95(3), 631e636. http://dx.doi.org/10.1890/13-1452.1.
Alcock, J. (1994). Alternative mate-locating tactics in Chlosyne californica (Lepi-doptera, Nymphalidae). Ethology, 97(2), 103e118.
Alcock, J., & O'Neill, K. M. (1986). Density-dependent mating tactics in the greyhairstreak, Strymon melinus (Lepidoptera: Lycaenidae). Journal of Zoology, 209,105e113.
Alonso-Pimentel, H., & Papaj, D. R. (1996). Operational sex ratio versus genderdensity as determinants of copulation duration in the walnut fly, Rhagoletisjuglandis (Diptera: Tephritidae). Behavioral Ecology and Sociobiology, 39(3),171e180. http://dx.doi.org/10.1007/s002650050278.
Arnqvist, G. (1992). Pre-copulatory fighting in a water strider: inter-sexual conflictor mate assessment? Animal Behaviour, 43(4), 559e567. http://dx.doi.org/10.1016/0003-3472(92)90079-O.
Aronsen, T., Berglund, A., Mobley, K. B., Ratikainen, I. I., & Rosenqvist, G. (2013). Sexratio and density affect sexual selection in a sex-role reversed fish. Evolution,67(11), 3243e3257. http://dx.doi.org/10.1111/evo.12201.
Aronsen, T., Mobley, K. B., Berglund, A., Sundin, J., Billing, A. M., & Rosenqvist, G.(2013). The operational sex ratio and density influence spatial relationshipsbetween breeding pipefish. Behavioral Ecology, 24(4), 888e897. http://dx.doi.org/10.1093/beheco/art019.
Arun, A., San Martin, G., Brakefield, P. M., & Nieberding, C. M. (2014). PBAN: Aconserved molecular mechanism for regulating sex pheromone biosynthesis acrossLepidoptera. Manuscript in preparation.
Barber, J. J., & Ogle, K. (2014). To P or not to P? Ecology, 95(3), 621e626. http://dx.doi.org/10.1890/13-1402.1.
van Bergen, E., Brakefield, P. M., Heuskin, S., Zwaan, B. J., & Nieberding, C. M. (2013).The scent of inbreeding: a male sex pheromone betrays inbred males. Pro-ceedings of the Royal Society B: Biological Sciences, 280(1758), 20130102. http://dx.doi.org/10.1098/rspb.2013.0102.

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245240
Berglund, A. (1994). The operational sex ratio influences choosiness in a pipefish.Behavioral Ecology, 5(3), 254e258.
Bergman, M., Gotthard, K., Berger, D., Olofsson, M., Kemp, D. J., & Wiklund, C. (2007).Mating success of resident versus non-resident males in a territorial butterfly.Proceedings of the Royal Society B: Biological Sciences, 274(1618), 1659e1665.http://dx.doi.org/10.1098/rspb.2007.0311.
Bergman, M., & Wiklund, C. (2009a). Differences in mate location behaviours be-tween residents and nonresidents in a territorial butterfly. Animal Behaviour,78(5), 1161e1167. http://dx.doi.org/10.1016/j.anbehav.2009.08.003.
Bergman, M., & Wiklund, C. (2009b). Visual mate detection and mate flight pursuitin relation to sunspot size in a woodland territorial butterfly. Animal Behaviour,78(1), 17e23. http://dx.doi.org/10.1016/j.anbehav.2009.02.005.
Bleu, J., Bessa-Gomes, C., & Laloi, D. (2012). Evolution of female choosiness andmating frequency: effects of mating cost, density and sex ratio. Animal Behav-iour, 83(1), 131e136. http://dx.doi.org/10.1016/j.anbehav.2011.10.017.
Brakefield, P. M. (1997). Phenotypic plasticity and fluctuating asymmetry as re-sponses to environmental stress in the butterfly Bicyclus anynana. InR. R. Bijlsma, & V. Loeschcke (Eds.), Environmental stress, adaptation and evo-lution (pp. 65e78). Basel, Switzerland: Birkh€auser.
Brakefield, P. M., Beldade, P., & Zwaan, B. J. (2009). The African butterfly Bicyclusanynana: a model for evolutionary genetics and evolutionary developmentalbiology. In R. R. Behringer, A. D. Johnson, & R. E. Krumlauf (Eds.), Emerging modelorganisms: A laboratory manual (Vol. 1, pp. 291e329). New York, NY: Cold SpringHarbor Laboratory.
Brakefield, P. M., El Filali, E., Van der Laan, R., Breuker, C. J., Saccheri, I. J., & Zwaan, B.(2001). Effective population size, reproductive success and sperm precedence inthe butterfly, Bicyclus anynana, in captivity. Journal of Evolutionary Biology, 14(1),148e156. http://dx.doi.org/10.1046/j.1420-9101.2001.00248.x.
Brakefield, P. M., Pijpe, J., & Zwaan, B. J. (2007). Developmental plasticity andacclimation both contribute to adaptive responses to alternating seasons ofplenty and of stress in Bicyclus butterflies. Journal of Biosciences, 32(3),465e475. http://dx.doi.org/10.1007/s12038-007-0046-8.
Brakefield, P. M., & Reitsma, N. (1991). Phenotypic plasticity, seasonal climate andthe population biology of Bicyclus butterflies (Satyridae) in Malawi. EcologicalEntomology, 16(3), 291e303.
Breuker, C. J., & Brakefield, P. M. (2002). Female choice depends on size but notsymmetry of dorsal eyespots in the butterfly Bicyclus anynana. Proceedings ofthe Royal Society B: Biological Sciences, 269(1497), 1233e1239. http://dx.doi.org/10.1098/rspb.2002.2005.
Burnham, K. P., & Anderson, D. R. (2002) (2nd ed.). Model selection and multimodelinference: A practical information-theoretic approach (2nd ed.), (Vol. 488). NewYork, NY: Springer.
Burnham, K. P., & Anderson, D. R. (2014). P values are only an index to evidence:20th- vs. 21st-century statistical science. Ecology, 95(3), 627e630. http://dx.doi.org/10.1890/13-1066.1.
Byrne, P. G., & Roberts, J. D. (2004). Intrasexual selection and group spawning inquacking frogs (Crinia georgiana). Behavioral Ecology, 15(5), 872e882. http://dx.doi.org/10.1093/beheco/arh100.
Cade, W. H., & Cade, E. S. (1992). Male mating success, calling and searchingbehaviour at high and low densities in the field cricket, Gryllus integer. AnimalBehaviour, 43(1), 49e56.
Carrillo, J., Danielson-François, A., Siemann, E., & Meffert, L. (2012). Male-biased sexratio increases female egg laying and fitness in the housefly, Musca domestica.Journal of Ethology, 30(2), 247e254. http://dx.doi.org/10.1007/s10164-011-0317-6.
Clark, L., & Grant, J. W. A. (2010). Intrasexual competition and courtship in femaleand male Japanese medaka, Oryzias latipes: effects of operational sex ratio anddensity. Animal Behaviour, 80(4), 707e712. http://dx.doi.org/10.1016/j.anbehav.2010.07.007.
Clutton-Brock, T. H., & Parker, G. A. (1992). Potential reproductive rates and theoperation of sexual selection. Quarterly Review of Biology, 67(4), 437e456.
Costanzo, K., & Monteiro, A. (2007). The use of chemical and visual cues in femalechoice in the butterfly Bicyclus anynana. Proceedings of the Royal Society B:Biological Sciences, 274(1611), 845e851. http://dx.doi.org/10.1098/rspb.2006.3729.
Crespi, B. J. (1988). Alternative male mating tactics in a thrips: effects of sex ratiovariation and body size. American Midland Naturalist, 119(1), 83e92.
Croshaw, D. A. (2010). Quantifying sexual selection: a comparison of competingindices with mating system data from a terrestrially breeding salamander.Biological Journal of the Linnean Society, 99(1), 73e83. http://dx.doi.org/10.1111/j.1095-8312.2009.01337.x.
Crowley, P. H., Travers, S. E., Linton, M. C., Cohn, S. L., Sih, A., & Sargent, R. C. (1991).Mate density, predation risk, and the seasonal sequence of mate choices: adynamic game. American Naturalist, 137(4), 567e596. http://dx.doi.org/10.1086/285184.
De Valpine, P. (2014). The common sense of P values. Ecology, 95(3), 617e621. http://dx.doi.org/10.1890/13-1271.1.
Debuse, V. J., Addison, J. T., & Reynolds, J. D. (1999). The effects of sex ratio on sexualcompetition in the European lobster. Animal Behaviour, 58(5), 973e981. http://dx.doi.org/10.1006/anbe.1999.1213.
Deno€el, M. (2003). Effect of rival males on the courtship of paedomorphic andmetamorphic Triturus alpestris (Amphibia: Salamandridae). Copeia, (3),618e623.
Dreiss, A. N., Cote, J., Richard, M., Federici, P., & Clobert, J. (2010). Age- and sex-specific response to population density and sex ratio. Behavioral Ecology,21(2), 356e364. http://dx.doi.org/10.1093/beheco/arp198.
Dudley, R. (2000). The biomechanics of insect flight: Form, function, evolution.Princeton, NJ: Princeton University Press.
Ellison, A. M., Gotelli, N. J., Inouye, B. D., & Strong, D. R. (2014). P values, hypothesistesting, and model selection: it's d�ej�a vu all over again. Ecology, 95(3), 609e610.http://dx.doi.org/10.1890/13-1911.1.
Elmberg, J. (1991). Factors affecting male yearly mating success in the common frog,Rana temporaria. Behavioral Ecology and Sociobiology, 28(2), 125e131. http://dx.doi.org/10.1007/BF00180989.
Elston, D. A., Moss, R., Boulinier, T., Arrowsmith, C., & Lambin, X. (2001). Analysis ofaggregation, a worked example: numbers of ticks on red grouse chicks. Para-sitology, 122(5), 563e569. http://dx.doi.org/10.1017/s0031182001007740.
Emlen, S. T., & Oring, L. W. (1977). Ecology, sexual selection, and evolution of matingsystems. Science, 197(4300), 215e223.
Eshel, I. (1979). Sexual selection, population density, and availability of mates.Theoretical Population Biology, 16(3), 301e314.
Ferkau, C., & Fischer, K. (2006). Costs of reproduction in male Bicyclus anynana andPieris napi butterflies: effects of mating history and food limitation. Ethology,112(11), 1117e1127. http://dx.doi.org/10.1111/j.1439-0310.2006.01266.x.
Fischer, K. (2006). Reduced mating vigor in selection lines of the butterfly Bicyclusanynana. Journal of Insect Behavior, 19(5), 657e668. http://dx.doi.org/10.1007/s10905-006-9057-9.
Fischer, K. (2007). Control of reproduction and a survival cost to mating in femaleBicyclus anynana butterflies. Ecological Entomology, 32(6), 674e681. http://dx.doi.org/10.1111/j.1365-2311.2007.00922.x.
Fischer, K., Perlick, J., & Galetz, T. (2008). Residual reproductive value andmalematingsuccess: older males do better. Proceedings of the Royal Society B: Biological Sci-ences, 275(1642), 1517e1524. http://dx.doi.org/10.1098/rspb.2007.1455.
Fischer, K., Zimmer, K., & Wedell, N. (2009). Correlated responses to selection onfemale egg size in male reproductive traits in a butterfly. Evolutionary Ecology,23(3), 389e402. http://dx.doi.org/10.1007/s10682-007-9233-1.
Fitze, P. S., & Le Galliard, J. F. (2008). Operational sex ratio, sexual conflict and theintensity of sexual selection. Ecology Letters, 11(5), 432e439. http://dx.doi.org/10.1111/j.1461-0248.2008.01158.x.
Fitze, P. S., Le Galliard, J. F., Federici, P., Richard, M., & Clobert, J. (2005). Conflict overmultiple-partner mating between males and females of the polygynandrouscommon lizards. Evolution, 59(11), 2451e2459.
Forsgren, E., Amundsen, T., Borg, Å. A., & Bjelvenmark, J. (2004). Unusually dynamicsex roles in a fish. Nature, 429(6991), 551e554. http://dx.doi.org/10.1038/nature02562.
Forstmeier, W., & Schielzeth, H. (2011). Cryptic multiple hypotheses testing in linearmodels: overestimated effect sizes and the winner's curse. Behavioral Ecologyand Sociobiology, 65(1), 47e55. http://dx.doi.org/10.1007/s00265-010-1038-5.
Frankino, W. A., Zwaan, B. J., Stern, D. L., & Brakefield, P. M. (2005). Natural selectionand developmental constraints in the evolution of allometries. Science,307(5710), 718e720. http://dx.doi.org/10.1126/science.1105409.
Frankino,W. A., Zwaan, B. J., Stern, D. L., & Brakefield, P.M. (2007). Internal and externalconstraints in the evolution of morphological allometries in a butterfly. Evolution,61(12), 2958e2970. http://dx.doi.org/10.1111/j.1558-5646.2007.00249.x.
Fromhage, L., Elgar, M. A., & Schneider, J. M. (2005). Faithful without care: theevolution of monogyny. Evolution, 59(7), 1400e1405.
Fromhage, L., McNamara, J. M., & Houston, A. I. (2008). A model for the evolutionarymaintenance of monogyny in spiders. Journal of Theoretical Biology, 250(3),524e531. http://dx.doi.org/10.1016/j.jtbi.2007.10.008.
Geister, T. L., & Fischer, K. (2007). Testing the beneficial acclimation hypothesis:temperature effects on mating success in a butterfly. Behavioral Ecology, 18(4),658e664. http://dx.doi.org/10.1093/beheco/arm024.
Grant, J. W. A., Bryant, M. J., & Soos, C. E. (1995). Operational sex ratio, mediated bysynchrony of female arrival, alters the variance of male mating success inJapanese medaka. Animal Behaviour, 49(2), 367e375. http://dx.doi.org/10.1006/anbe.1995.9998.
Gwynne, D. T. (1994). Sexual selection and sexual differences in morman crickets(Orthoptera: Tettigoniidae, Anabrus simplex). Evolution, 38, 1011e1022.
Gwynne, D. T., & Simmons, L. W. (1990). Experimental reversal of courtship roles inan insect. Nature, 346(6280), 172e174.
H€ardling, R., & Kaitala, A. (2005). The evolution of repeated mating under sexualconflict. Journal of Evolutionary Biology, 18(1), 106e115. http://dx.doi.org/10.1111/j.1420-9101.2004.00795.x.
Head, M. L., Lindholm, A. K., & Brooks, R. (2007). Operational sex ratio and densitydo not affect directional selection on male sexual ornaments and behavior.Evolution, 62(1), 135e144. http://dx.doi.org/10.1111/j.1558-5646.2007.00277.x.
Hebets, E. A., & Sullivan-Beckers, L. (2010). Mate choice and learning. In M. D. Breed,& J. Moore (Eds.), Encyclopedia of Animal Behavior (Vol. 2, pp. 389e393).Amsterdam, The Netherlands: Elsevier.
Heuskin, S., Vanderplanck, M., Bacquet, P., Holveck, M. J., Kaltenpoth, M., Engl, T.,et al. (2014). The composition of cuticular compounds indicates body parts, sexand age in the model butterfly Bicyclus anynana. Frontiers in Ecology and Evo-lution, 2, 37. http://dx.doi.org/10.3389/fevo.2014.00037.
Holveck, M. J., Gauthier, A. L., & Nieberding, C. M. (2013). [Field collection of Bicyclusanynana butterflies in Uganda]. Unpublished raw data.
Holveck, M. J., & Riebel, K. (2014). Female zebra finches learn to prefer more thanone song and from more than one tutor. Animal Behaviour, 88, 125e135. http://dx.doi.org/10.1016/j.anbehav.2013.11.023.
Hubbell, S. P., & Johnson, L. K. (1987). Environmental variance in lifetime matingsuccess, mate choice, and sexual selection. American Naturalist, 130(1),91e112.

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 241
Hutchinson, J. M. C. (2005). Is more choice always desirable? Evidence and argu-ments from leks, food selection, and environmental enrichment. Biological Re-views of the Cambridge Philosophical Society, 80(1), 73e92. http://dx.doi.org/10.1017/S1464793104006554.
Isvaran, K. (2005). Variation in male mating behaviour within ungulate pop-ulations: patterns and processes. Current Science, 89(7), 1192e1199.
Janowitz, S. A., & Fischer, K. (2010). Costing reproduction: effects of mating op-portunity on mating success in male Bicyclus anynana butterflies. BehavioralEcology and Sociobiology, 64(12), 1999e2006. http://dx.doi.org/10.1007/s00265-010-1011-3.
Janowitz, S. A., & Fischer, K. (2011). Opposing effects of heat stress on male versusfemale reproductive success in Bicyclus anynana butterflies. Journal of ThermalBiology, 36(5), 283e287. http://dx.doi.org/10.1016/j.jtherbio.2011.04.001.
Janowitz, S. A., & Fischer, K. (2012). Polyandry in Bicyclus anynana butterflies re-sults from sexual conflict over mating. Ethology. http://dx.doi.org/10.1111/eth.12017.
Jennions, M. D., & Petrie, M. (1997). Variation in mate choice and mating preferences:a review of causes and consequences. Biological Reviews of the Cambridge Phil-osophical Society, 72(2), 283e327. http://dx.doi.org/10.1017/S0006323196005014.
Jirotkul, M. (1999a). Operational sex ratio influences female preference and male-male competition in guppies. Animal Behaviour, 58(2), 287e294. http://dx.doi.org/10.1006/anbe.1999.1149.
Jirotkul, M. (1999b). Population density influences male-male competition inguppies. Animal Behaviour, 58(6), 1169e1175. http://dx.doi.org/10.1006/anbe.1999.1248.
Jones, A. G., Arguello, J. R., & Arnold, S. J. (2004). Molecular parentage analysis inexperimental newt populations: the response of mating system measures tovariation in the operational sex ratio. American Naturalist, 164(4), 444e456.http://dx.doi.org/10.1086/423826.
de Jong, K., Forsgren, E., Sandvik, H., & Amundsen, T. (2012). Measuring matingcompetition correctly: available evidence supports operational sex ratio theory.Behavioral Ecology, 23(6), 1170e1177. http://dx.doi.org/10.1093/beheco/ars094.
de Jong, K.,Wacker, S., Amundsen, T., & Forsgren, E. (2009). Do operational sex ratio anddensity affect mating behaviour? An experiment on the two-spotted goby. AnimalBehaviour, 78(5), 1229e1238. http://dx.doi.org/10.1016/j.anbehav.2009.08.006.
Joron, M., & Brakefield, P. M. (2003). Captivity masks inbreeding effects on malemating success in butterflies. Nature, 424, 191e194. http://dx.doi.org/10.1038/nature01713.
Karl, I., & Fischer, K. (2013). Old male mating advantage results from sexual conflictin a butterfly. Animal Behaviour, 85(1), 143e149. http://dx.doi.org/10.1016/j.anbehav.2012.10.018.
Karl, I., Heuskin, S., & Fischer, K. (2013). Dissecting the mechanisms underlying oldmale mating advantage in a butterfly. Behavioral Ecology and Sociobiology, 67(5),837e849. http://dx.doi.org/10.1007/s00265-013-1507-8.
Kehl, T., Burmeister, M. F. W. T., Donke, E., K€ohn, N. A. K., Metschke, K., Pfender, D.,et al. (2014). Pheromone blend does not explain old male mating advantage in abutterfly. Ethology. http://dx.doi.org/10.1111/eth.12287.
Kemp, D. J. (2001). Investigating the consistency of mate-locating behavior in theterritorial butterfly Hypolimnas bolina (Lepidoptera: Nymphalidae). Journal ofInsect Behavior, 14(1), 129e147. http://dx.doi.org/10.1023/A:1007809915296.
Klemme, I., Yl€onen, H., & Eccard, J. A. (2007). Reproductive success of male bankvoles (Clethrionomys glareolus): the effect of operational sex ratio and body size.Behavioral Ecology and Sociobiology, 61(12), 1911e1918. http://dx.doi.org/10.1007/s00265-007-0431-1.
Kokko, H., & Jennions, M. D. (2008). Parental investment, sexual selection and sexratios. Journal of Evolutionary Biology, 21(4), 919e948. http://dx.doi.org/10.1111/j.1420-9101.2008.01540.x.
Kokko, H., & Johnstone, R. A. (2002). Why is mutual mate choice not the norm?Operational sex ratios, sex roles and the evolution of sexually dimorphic andmonomorphic signalling. Philosophical Transactions of the Royal Society B: Bio-logical Sciences, 357(1419), 319e330. http://dx.doi.org/10.1098/rstb.2001.0926.
Kokko, H., Klug, H., & Jennions, M. D. (2012). Unifying cornerstones of sexual se-lection: operational sex ratio, Bateman gradient and the scope for competitiveinvestment. Ecology Letters, 15(11), 1340e1351. http://dx.doi.org/10.1111/j.1461-0248.2012.01859.x.
Kokko, H., & Monaghan, P. (2001). Predicting the direction of sexual selection. EcologyLetters, 4(2), 159e165. http://dx.doi.org/10.1046/j.1461-0248.2001.00212.x.
Kokko, H., & Rankin, D. J. (2006). Lonely hearts or sex in the city? Density-dependent effects in mating systems. Philosophical Transactions of the RoyalSociety B: Biological Sciences, 361(1466), 319e334. http://dx.doi.org/10.1098/rstb.2005.1784.
Krupa, J. J., & Sih, A. (1993). Experimental studies onwater strider mating dynamics:spatial variation in density and sex ratio. Behavioral Ecology and Sociobiology,33(2), 107e120. http://dx.doi.org/10.1007/BF00171662.
Kvarnemo, C., & Ahnesj€o, I. (1996). The dynamics of operational sex ratios andcompetition for mates. Trends in Ecology and Evolution, 11(10), 404e408. http://dx.doi.org/10.1016/0169-5347(96)10056-2.
Kvarnemo, C., Forsgren, E., & Magnhagen, C. (1995). Effects of sex ratio on intra- andinter-sexual behaviour in sand gobies. Animal Behaviour, 50(6), 1455e1461.http://dx.doi.org/10.1016/0003-3472(95)80002-6.
Laloi, D., Richard, M., Lecomte, J., Massot, M., & Clobert, J. (2004). Multiple paternityin clutches of common lizard Lacerta vivipara: data from microsatellite markers.Molecular Ecology, 13(3), 719e723. http://dx.doi.org/10.1046/j.1365-294X.2004.02102.x.
Lauer, M. J., Sih, A., & Krupa, J. J. (1996). Male density, female density and inter-sexual conflict in a stream-dwelling insect. Animal Behaviour, 52(5), 929e939.http://dx.doi.org/10.1006/anbe.1996.0241.
Lavine, M. (2014). Comment on murtaugh. Ecology, 95(3), 642e645. http://dx.doi.org/10.1890/13-1112.1.
Lawrence, W. S. (1986). Male choice and competition in Tetraopes tetraophthalmus:effects of local sex ratio variation. Behavioral Ecology and Sociobiology, 18(4),289e296. http://dx.doi.org/10.1007/BF00300006.
Lewis, Z., & Wedell, N. (2007). Effect of adult feeding on male mating behaviour inthe butterfly, Bicyclus anynana (Lepidoptera: Nymphalidae). Journal of InsectBehavior, 20(2), 201e213. http://dx.doi.org/10.1007/s10905-007-9075-2.
Lod�e, T., Holveck, M. J., & Lesbarr�eres, D. (2005). Asynchronous arrival pattern,operational sex ratio and occurrence of multiple paternities in a territorialbreeding anuran, Rana dalmatina. Biological Journal of the Linnean Society, 86(2),191e200. http://dx.doi.org/10.1111/j.1095-8312.2005.00521.x.
Lod�e, T., Holveck, M. J., Lesbarr�eres, D., & Pagano, A. (2004). Sex-biased predationby polecats influences the mating system of frogs. Proceedings of the RoyalSociety B: Biological Sciences, 271, S399eS401. http://dx.doi.org/10.1098/rsbl.2004.0195.
Lu, X., Ma, X., Fan, L., & Yu, T. (2010). Opportunity for sexual selection arises atmoderate densities in the frog Rana chensinensis: an experimental approach.Journal of Ethology, 28(2), 257e262. http://dx.doi.org/10.1007/s10164-009-0179-3.
McLain, D. K. (1982). Density dependent sexual selection and positive phenotypicassortative mating in natural populations of the soldier beetle, Chauliognathuspennsylvanicus. Evolution, 36(6), 1227e1235.
Miller, C. W., & Svensson, E. I. (2014). Sexual selection in complex environments.Annual Review of Entomology, 59, 427e445.
Mills, S. C., & Reynolds, J. D. (2003). Operational sex ratio and alternative repro-ductive behaviours in the European bitterling, Rhodeus sericeus. BehavioralEcology and Sociobiology, 54(2), 98e104. http://dx.doi.org/10.1007/s00265-003-0616-1.
M€olleman, F., Zwaan, B. J., & Brakefield, P. M. (2004). The effect of male sodium dietand mating history on female reproduction in the puddling squinting bushbrown Bicyclus anynana (Lepidoptera). Behavioral Ecology and Sociobiology,56(4), 404e411. http://dx.doi.org/10.1007/s00265-004-0789-2.
Mundry, R. (2011). Issues in information theory-based statistical inference e acommentary from a frequentist's perspective. Behavioral Ecology and Sociobio-logy, 65(1), 57e68. http://dx.doi.org/10.1007/s00265-010-1040-y.
Mundry, R., & Nunn, C. L. (2009). Stepwise model fitting and statistical inference:turning noise into signal pollution. American Naturalist, 173(1), 119e123. http://dx.doi.org/10.1086/593303.
Munyuli, M. B. T. (2012). Butterfly diversity from farmlands of central Uganda.Psyche, 2012, 481509. http://dx.doi.org/10.1155/2012/481509.
Murtaugh, P. A. (2009). Performance of several variable-selection methods appliedto real ecological data. Ecology Letters, 12(10), 1061e1068. http://dx.doi.org/10.1111/j.1461-0248.2009.01361.x.
Murtaugh, P. A. (2014a). In defense of P values. Ecology, 95(3), 611e617. http://dx.doi.org/10.1890/13-0590.1.
Murtaugh, P. A. (2014b). Rejoinder. Ecology, 95(3), 651e653. http://dx.doi.org/10.1890/13-1858.1.
Nieberding, C. M., de Vos, H., Schneider, M. V., Lassance, J. M., Estramil, N.,Andersson, J., et al. (2008). The male sex pheromone of the butterfly Bicyclusanynana: towards an evolutionary analysis. PLoS One, 3(7). http://dx.doi.org/10.1371/journal.pone.0002751.
Nieberding, C. M., Fischer, K., Saastamoinen, M., Allen, C. E., Wallin, E. A.,Hedenstr€om, E., et al. (2012). Cracking the olfactory code of a butterfly: thescent of ageing. Ecology Letters, 15(5), 415e424.
Owens, I. P. F., & Thompson, D. B. A. (1994). Sex differences, sex ratios and sex roles.Proceedings of the Royal Society B: Biological Sciences, 258(1352), 93e99. http://dx.doi.org/10.1098/rspb.1994.0148.
Palokangas, P., Alatalo, R. V., & Korpim€aki, E. (1992). Female choice in the kestrelunder different availability of mating options. Animal Behaviour, 43(4),659e665. http://dx.doi.org/10.1016/0003-3472(92)90087-P.
Peixoto, P. E. C., & Benson, W. W. (2009). Seasonal effects of density on territoryoccupation by males of the satyrine butterfly Paryphthimoides phronius (Butler1867). Journal of Ethology, 27(3), 489e496. http://dx.doi.org/10.1007/s10164-008-0147-3.
Pomfret, J. C., & Knell, R. J. (2008). Crowding, sex ratio and horn evolution in a SouthAfrican beetle community. Proceedings of the Royal Society B: Biological Sciences,275(1632), 315e321. http://dx.doi.org/10.1098/rspb.2007.1498.
Prudic, K. L., Jeon, C., Cao, H., & Monteiro, A. (2011). Developmental plasticity insexual roles of butterfly species drives mutual sexual ornamentation. Science,331(6013). http://dx.doi.org/10.1126/science.1197114.
R Core Team (2014). R: A language and environment for statistical computing. Vienna,Austria: R Foundation for Statistical Computing. http://www.R-project.org.
Robertson, K. A., & Monteiro, A. (2005). Female Bicyclus anynana butterflies choosemales on the basis of their dorsal UV-reflective eyespot pupils. Proceedings ofthe Royal Society B: Biological Sciences, 272(1572), 1541e1546. http://dx.doi.org/10.1098/rspb.2005.3142.
Rowe, L. (1992). Convenience polyandry in a water strider: foraging conflicts andfemale control of copulation frequency and guarding duration. Animal Behav-iour, 44(2), 189e202. http://dx.doi.org/10.1016/0003-3472(92)90025-5.

Table A1Summary of the models testing for the effects of density, sex ratio, cage volume andnumber of individuals on the courtship rate per male
Model terms Estimate±1 SE F P
Intercept 3.386±1.210Density 0.372±0.166 5.0 0.04Sex ratio �0.011±0.018 0.4 0.6Cage volume residuals 0.041±0.040 1.1 0.3Number of individuals �0.169±0.260 0.4 0.5Density)sex ratio �0.009±0.010 0.8 0.4Cage volume residuals)sex ratio �0.002±0.002 0.5 0.5
The full generalized linear model (GLM) is of the following form: Y ~ log(density)� sex ratio þ cage volume residuals � sex ratio þ log(number of individuals) with anormal distribution. Rejected terms are in italics, significant ones in bold. Theintercept is for the minimal model including at least all fixed factors without in-teractions. Cage volume residuals come from the linear regression of cage volume ondensity and number of individuals. For the comparison of GLM with dispersion
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245242
Rowe, L., & Arnqvist, G. (2002). Sexually antagonistic coevolution in a mating sys-tem: combining experimental and comparative approaches to address evolu-tionary processes. Evolution, 56(4), 754e767.
Rowe, L., Arnqvist, G., Sih, A., & Krupa, J. J. (1994). Sexual conflict and the evolu-tionary ecology of mating patterns: water striders as a model system. Trends inEcology and Evolution, 9(8), 289e293.
Saccheri, I. J., & Brakefield, P. M. (2002). Rapid spread of immigrant genomes intoinbred populations. Proceedings of the Royal Society B: Biological Sciences,269(1495), 1073e1078. http://dx.doi.org/10.1098/rspb.2002.1963.
Saeki, Y., Kruse, K. C., & Switzer, P. V. (2005). The social environment affects mateguarding behavior in Japanese beetles, Popillia japonica. Journal of Insect Science,5. http://dx.doi.org/10.1673/031.005.1801.
Schwagmeyer, P. L., & Brown, C. H. (1983). Factors affecting male-male competitionin thirteen-lined ground squirrels. Behavioral Ecology and Sociobiology, 13(1),1e6. http://dx.doi.org/10.1007/BF00295069.
Servedio, M. R. (2007). Male versus female mate choice: sexual selection and theevolution of species recognition via reinforcement. Evolution, 61(12),2772e2789. http://dx.doi.org/10.1111/j.1558-5646.2007.00247.x.
Shelly, T. E., & Bailey, W. J. (1992). Experimental manipulation of mate choice bymale katydids: the effect of female encounter rate. Behavioral Ecology and So-ciobiology, 30(3e4), 277e282. http://dx.doi.org/10.1007/BF00166713.
Shine, R., Langkilde, T., & Mason, R. T. (2003). The opportunistic serpent: male gartersnakes adjust courtship tactics to mating opportunities. Behaviour, 140(11e12),1509e1526. http://dx.doi.org/10.1163/156853903771980693.
Smith, C. C. (2007). Independent effects of male and female density on sexualharassment, female fitness, and male competition for mates in the westernmosquitofish Gambusia affinis. Behavioral Ecology and Sociobiology, 61(9),1349e1358. http://dx.doi.org/10.1007/s00265-007-0365-7.
Souroukis, K., & Murray, A. M. (1995). Female mating behavior in the field cricket,Gryllus pennsylvanicus (Orthoptera: Gryllidae) at different operational sex ratios.Journal of Insect Behavior, 8(2), 269e279.
Spanos, A. (2014). Recurring controversies about P values and confidence intervalsrevisited. Ecology, 95(3), 645e651. http://dx.doi.org/10.1890/13-1291.1.
Stanton-Geddes, J., De Freitas, C. G., & De Sales Dambros, C. (2014). In defense of Pvalues: comment on the statistical methods actually used by ecologists. Ecology,95(3), 637e642. http://dx.doi.org/10.1890/13-1156.1.
Thornhill, R., & Alcock, J. (1983). The evolution of insect mating systems. Cambridge,MA: Harvard University Press.
Uller, T., & Olsson, M. (2008). Multiple paternity in reptiles: patterns and processes.Molecular Ecology, 17(11), 2566e2580. http://dx.doi.org/10.1111/j.1365-294X.2008.03772.x.
Van't Hof, A. E., Zwaan, B. J., Saccheri, I. J., Daly, D., Bot, A. N. M., & Brakefield, P. M.(2005). Characterization of 28 microsatellite loci for the butterfly Bicyclusanynana. Molecular Ecology Notes, 5(1), 169e172. http://dx.doi.org/10.1111/j.1471-8286.2005.00870.x.
Wacker, S., Mobley, K., Forsgren, E., Myhre, L. C., de Jong, K., & Amundsen, T. (2013).Operational sex ratio but not density affects sexual selection in a fish. Evolution,67(7), 1937e1949. http://dx.doi.org/10.1111/evo.12092.
Wang, Q., He, X. Z., Yang, L., Hedderley, D., & Davis, L. K. (2009). Biased sex ratio andlow population density increase male mating success in the bug Nysius huttoni(Heteroptera: Lygaeidae). Naturwissenschaften, 96(1), 87e91. http://dx.doi.org/10.1007/s00114-008-0455-z.
Wang, Q., Yang, L., & Hedderley, D. (2008). Function of prolonged copulation inNysius huttoni White (Heteroptera: Lygaeidae) under male-biased sex ratio andhigh population density. Journal of Insect Behavior, 21(2), 89e99. http://dx.doi.org/10.1007/s10905-007-9110-3.
Westerman, E. L., Chirathivat, N., Schyling, E., & Monteiro, A. (2014). Mate prefer-ence for a phenotypically plastic trait is learned, and may facilitate preference-phenotype matching. Evolution, 68(6), 1661e1670. http://dx.doi.org/10.1111/evo.12381.
Westerman, E. L., Drucker, C. B., & Monteiro, A. (2014). Male and female matingbehavior is dependent on social context in the butterfly Bicyclus anynana.Journal of Insect Behavior, 1e18. http://dx.doi.org/10.1007/s10905-014-9441-9.
Westerman, E. L., Hodgins-Davis, A., Dinwiddie, A., & Monteiro, A. (2012). Biasedlearning affects mate choice in a butterfly. Proceedings of the National Academyof Sciences of the United States of America, 109(27), 10948e10953.
Westerman, E. L., & Monteiro, A. (2013). Odour influences whether females learn toprefer or to avoid wing patterns of male butterflies. Animal Behaviour, 86(6),1139e1145. http://dx.doi.org/10.1016/j.anbehav.2013.09.002.
Widemo, F., & Saether, S. A. (1999). Beauty is in the eye of the beholder: causes andconsequences of variation in mating preferences. Trends in Ecology & Evolution,14(1), 26e31. http://dx.doi.org/10.1016/S0169-5347(98)01531-6.
Windig, J. J., Brakefield, P. M., Reitsma, N., & Wilson, J. G. M. (1994). Seasonal pol-yphenism in the wild: survey of wing patterns in five species of Bicyclus but-terflies in Malawi. Ecological Entomology, 19(3), 285e298.
Wong, B. B. M., & Candolin, U. (2005). How is female mate choice affected by malecompetition? Biological Reviews, 80(4), 559e571. http://dx.doi.org/10.1017/S1464793105006809.
Zuur, A. F., Ieno, E. N., & Elphick, C. S. (2010). A protocol for data exploration to avoidcommon statistical problems. Methods in ecology and evolution, 1, 3e14.
Zwaan, B. J., Zijlstra, W. G., Keller, M., Pijpe, J., & Brakefield, P. M. (2008). Potentialconstraints on evolution: sexual dimorphism and the problem of protandry inthe butterfly Bicyclus anynana. Journal of Genetics, 87(4), 395e405. http://dx.doi.org/10.1007/s12041-008-0062-y.
APPENDIX
Courtship rate per male
RationaleIncreasing male numbers in behavioural experiments neces-
sarily increases male competition for matings as there are fewerfemales to get per male (see references above). Accordingly, we seethat increasingmale numbers increased overall the number of malecourtships, which necessarily increased the pressure for females toescape unwanted matings. This relationship, however, does not tellus whether individual males did or did not react to their increasednumbers by courting more, but in both cases, the effect on femaleswas that they had to choose to accept themating or not, more often.Thus, whether females were experiencing more male courtshipssimply because they were surrounded by more males or becausemales were increasing their individual courtship effort both lead tomore pressure on females for mating. This said, we neverthelessanalysed the effects of individual numbers, cage volume, densityand sex ratio on the courtship rate per male in order to infer andclarify whether the individual effort of males to court increased inresponse to the increase in maleemale competition.
MethodAs we did not follow males individually, we calculated the
courtship rate permale by dividing the number of courtships by thenumber of males in each experimental trial.
ResultsThe courtship rate per male increased with density (Tables A1,
A2, Fig. A1), but this relationship was driven by two outliers(both from a female-biased sex ratio) and does not hold withoutthem (F1 ¼ 2.4, P ¼ 0.14, R2 ¼ 0.15). Accordingly, there was no effectof density on the number of male courtships at the population level(Tables 3, A2). Neither the number of individuals, sex ratio, cagevolume nor any of the tested interactions (Tables A1, A2) affectedthe courtship rate per male. Although the number of courtships inthe population increased with increasing numbers of individuals(Fig. 2a), individual males did not court more because they weremore numerous and although the number of courtships in thepopulation increased with an increasing proportion of males(Fig. 2b), sex ratio did not affect courtship behaviour of individualmales. The latter means that females were courted more in thepopulation when the proportion of males increased, and thereforethat the competition between males to access females increasedeven though there was no direct effect of sex ratio on the courtshipbehaviour of individual males.
estimated by moments (e.g. Gaussian fits) the F test is most appropriate.

Table A2Set of models, ordered by DAICc (or DQAICc for number of male courtship attempts) specified by rank (lowest on top), explaining each behaviour
Behavioural variable Model terms k DAICc wi
Intercept Cage volumeresiduals
Density Number ofindividuals
Sex ratio Cage volumeresiduals)sex ratio
Density)sexratio
Number of malecourtship attempts
0.022 NA 0.061 0.659 0.014 NA NA 4 0.0 0.4�0.228 0.003 NA 0.690 0.013 NA NA 4 0.7 0.30.542 NA 0.332 0.664 0.004 NA �0.005 5 2.8 0.10.582 0.011 0.064 0.713 NA NA NA 4 3.5 0.10.070 0.012 0.081 0.658 0.014 NA NA 5 3.8 0.1
�0.233 0.012 NA 0.692 0.013 0.000 NA 5 4.8 0.00.535 0.008 0.326 0.663 0.005 NA �0.004 6 7.5 0.00.073 0.010 0.082 0.657 0.014 0.000 NA 6 8.7 0.00.575 0.047 0.381 0.687 0.002 �0.001 �0.006 7 12.9 0.02.038 0.003 0.182 NA 0.026 NA NA 4 36.5 0.02.093 �0.049 0.193 NA 0.026 0.001 NA 5 39.9 0.02.412 0.000 0.366 NA 0.019 NA �0.003 5 40.1 0.02.541 �0.058 0.415 NA 0.018 0.001 �0.004 6 44.1 0.0
Courtship rate per male 2.976 0.044 0.388 �0.197 NA NA NA 5 0 0.32.931 0.041 0.354 NA �0.013 NA NA 5 0.07 0.33.457 NA 0.369 �0.164 �0.013 NA NA 5 0.89 0.24.114 0.035 0.798 NA �0.038 NA �0.009 6 3.08 0.12.853 0.127 0.336 NA �0.013 �0.002 NA 6 3.6 0.04.692 NA 0.856 �0.145 �0.040 NA �0.010 6 3.68 0.03.386 0.041 0.372 �0.169 �0.011 NA NA 6 3.7 0.02.331 0.040 NA �0.070 �0.017 NA NA 5 5.31 0.04.114 0.131 0.811 NA �0.040 �0.002 �0.010 7 7.08 0.04.469 0.036 0.794 �0.152 �0.035 NA �0.009 7 7.48 0.03.268 0.120 0.354 �0.151 �0.011 �0.002 NA 7 8.02 0.02.227 0.162 NA �0.051 �0.017 �0.002 NA 6 8.44 0.04.419 0.125 0.807 �0.131 �0.037 �0.002 �0.009 8 12.5 0.0
Proportion of courtshipsending with a mating
�0.433 NA 0.000 �0.309 �0.020 NA NA 4 0.0 0.2�0.433 0.000 NA �0.309 �0.020 NA NA 4 0.0 0.2�1.049 0.198 NA �0.179 �0.020 ¡0.004 NA 5 0.3 0.2�1.571 NA �0.698 �0.365 0.008 NA 0.014 5 0.4 0.2�1.591 0.233 �0.019 NA �0.024 ¡0.004 NA 5 1.0 0.1�1.023 0.005 0.048 �0.401 NA NA NA 4 2.3 0.1�1.331 0.006 0.003 NA �0.025 NA NA 4 2.7 0.0�2.398 0.013 �0.577 NA �0.003 NA 0.012 5 3.8 0.0�0.433 0.000 0.000 �0.309 �0.020 NA NA 5 4.1 0.0�2.163 0.207 �0.368 NA �0.011 �0.004 0.007 6 5.0 0.0�1.062 0.196 �0.024 �0.181 �0.021 �0.004 NA 6 5.2 0.0�1.599 0.005 �0.693 �0.360 0.008 NA 0.014 6 5.3 0.0�1.698 0.150 �0.587 �0.302 0.004 �0.003 0.012 7 9.1 0.0
Proportion of courtshipsaborted by the male
�2.490 ¡0.037 ¡0.153 0.177 NA NA NA 6 0.0 0.3�2.008 ¡0.036 ¡0.146 NA 0.004 NA NA 6 0.6 0.2�2.216 ¡0.028 NA 0.163 0.004 NA NA 6 1.2 0.2�2.435 NA �0.102 0.161 0.003 NA NA 6 1.6 0.1�3.475 ¡0.034 ¡0.737 NA 0.031 NA 0.011 7 4.2 0.0�4.030 NA �0.733 0.168 0.033 NA 0.012 7 5.1 0.0�2.032 �0.136 �0.140 NA 0.005 0.002 NA 7 5.3 0.0�2.669 �0.037 �0.153 0.175 0.003 NA NA 7 6.0 0.0�2.149 �0.124 NA 0.132 0.004 0.002 NA 7 6.1 0.0�3.923 �0.194 �0.908 NA 0.040 0.003 0.014 8 8.6 0.0�4.190 �0.036 �0.749 0.185 0.031 NA 0.011 8 11.0 0.0�2.592 �0.131 �0.148 0.147 0.004 0.002 NA 8 12.4 0.0�4.542 �0.192 �0.927 0.151 0.040 0.003 0.014 9 17.7 0.0
Proportion of courtshipsaborted by the female
0.804 0.022 0.090 NA 0.007 NA NA 6 0.0 0.31.117 0.021 0.081 0.011 NA NA NA 6 0.6 0.20.598 0.015 NA �0.001 0.006 NA NA 6 0.9 0.20.707 NA 0.058 �0.002 0.007 NA NA 6 1.1 0.21.905 0.018 0.589 NA �0.014 NA ¡0.009 7 2.3 0.11.940 NA 0.612 0.006 �0.017 NA �0.010 7 3.1 0.10.799 0.038 0.088 NA 0.007 0.000 NA 7 6.0 0.00.828 0.022 0.090 �0.007 0.007 NA NA 7 6.0 0.00.570 0.038 NA 0.008 0.006 0.000 NA 7 6.8 0.02.061 0.082 0.663 NA �0.017 �0.001 �0.011 8 8.9 0.01.896 0.018 0.589 0.003 �0.014 NA �0.009 8 9.8 0.00.799 0.038 0.088 0.000 0.007 0.000 NA 8 13.5 0.01.950 0.084 0.661 0.031 �0.017 �0.001 �0.011 9 18.6 0.0
Duration of courtshipsending with a mating
1.603 �0.023 0.089 NA 0.013 NA NA 6 0.0 0.22.535 0.006 0.820 NA �0.010 NA ¡0.018 7 0.1 0.22.476 NA 0.805 0.032 �0.011 NA ¡0.017 7 0.1 0.21.584 �0.018 NA �0.084 0.017 NA NA 6 0.5 0.1
(continued on next page)
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 243
Table A2 (continued )
Behavioural variable Model terms k DAICc wi
Intercept Cage volumeresiduals
Density Number ofindividuals
Sex ratio Cage volumeresiduals)sex ratio
Density)sexratio
1.570 NA 0.060 �0.080 0.019 NA NA 6 0.8 0.12.226 �0.035 0.111 0.020 NA NA NA 6 1.7 0.11.614 0.019 0.101 NA 0.013 �0.001 NA 7 4.1 0.01.730 �0.022 0.081 �0.059 0.015 NA NA 7 4.3 0.02.786 �0.054 1.011 NA �0.016 0.001 �0.023 8 4.5 0.01.549 0.003 NA �0.074 0.016 0.000 NA 7 4.8 0.02.490 0.006 0.846 0.034 �0.012 NA �0.018 8 5.2 0.01.693 0.016 0.095 �0.037 0.014 �0.001 NA 8 9.1 0.02.756 �0.053 1.023 0.019 �0.017 0.001 �0.023 9 10.4 0.0
Duration of courtshipsaborted by the male
0.925 NA ¡0.121 �0.050 0.005 NA NA 6 0.0 0.20.691 0.003 ¡0.128 NA 0.005 NA NA 6 0.2 0.21.247 0.005 ¡0.121 �0.058 NA NA NA 6 0.7 0.20.652 �0.060 ¡0.133 NA 0.005 0.001 NA 7 1.5 0.10.941 0.003 �0.121 �0.051 0.005 NA NA 7 2.4 0.10.887 NA �0.133 �0.048 0.005 NA 0.000 7 2.4 0.10.618 0.003 �0.158 NA 0.006 NA 0.001 7 2.6 0.10.549 �0.061 �0.176 NA 0.007 0.001 0.001 8 3.9 0.00.751 �0.056 �0.130 �0.020 0.005 0.001 NA 8 4.0 0.01.490 0.003 NA �0.106 0.005 NA NA 6 4.7 0.00.910 0.003 �0.131 �0.049 0.005 NA 0.000 8 4.9 0.00.623 �0.059 �0.168 �0.012 0.006 0.001 0.001 9 6.5 0.01.409 �0.029 NA �0.092 0.005 0.001 NA 7 6.9 0.0
Duration of courtshipsaborted by the female
�0.221 0.006 �0.491 NA 0.010 NA 0.008 7 0.0 0.3�0.216 NA �0.455 0.031 0.008 NA 0.007 7 0.2 0.3�0.063 �0.035 �0.433 NA 0.008 0.001 0.007 8 1.1 0.2�0.282 0.006 �0.480 0.030 0.009 NA 0.007 8 2.0 0.1�0.077 �0.034 �0.433 0.005 0.008 0.001 0.007 9 3.2 0.10.479 NA �0.065 0.056 �0.007 NA NA 6 4.1 0.00.628 0.002 �0.069 NA �0.006 NA NA 6 4.4 0.00.691 �0.048 �0.052 NA �0.006 0.001 NA 7 5.2 0.00.213 0.005 �0.054 0.033 NA NA NA 6 5.4 0.00.541 0.001 NA 0.061 �0.006 NA NA 6 5.5 0.00.688 �0.053 NA 0.024 �0.006 0.001 NA 7 6.1 0.00.470 0.002 �0.067 0.057 �0.007 NA NA 7 6.2 0.00.612 �0.044 �0.053 0.027 �0.007 0.001 NA 8 7.2 0.0
Models' coefficients (or NA for missing) are given for each model term, k being their total number. The significant model terms, as detected by stepwise model selection, are inbold and the ‘best’ subsets of models (DAIC � 2) in italics.
R2 = 0.23
−6 −4 −3 −2 −1 0
6
4
3
2
1
0
5
Cou
rtsh
ip r
ate
per
mal
e
−5
Density
Figure A1. Courtship rate per male as a function of density (as natural logarithm). Thelinear regression line is shownwith its corresponding R2 value. Jitter range is 0.1 on thex-axis and 0.07 on the y-axis to see overlapping points.
M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245244
Data analyses with information theory-based method
RationaleAs there is considerable debate about the performance of
stepwise model selection, used for data analyses in this paper (Aho,Derryberry, & Peterson, 2014; Barber & Ogle, 2014; Burnham &Anderson, 2014; De Valpine, 2014; Ellison, Gotelli, Inouye, &Strong, 2014; Lavine, 2014; Mundry, 2011; Mundry & Nunn,2009; Murtaugh, 2009, 2014a, 2014b; Spanos, 2014; Stanton-Geddes, De Freitas, & De Sales Dambros, 2014), we here presentthe analyses of all data with information theory-based model se-lection (Burnham & Anderson, 2002).
MethodWe used the second-order Akaike's information criterion, AICc,
which includes a correction term when sample sizes are small incomparison to the number of estimated parameters, or quasi-AICc,QAICc, a modification of AICc for overdispersed count data wherequasi-likelihood adjustments are required. Models of each set wereordered from ‘best’ (lowest AIC) to ‘worst’ (highest AIC). We reportAIC differences (DAIC ¼ AICi � AICminimum): models with DAICi � 2are considered to give a good approximation of the data, whereasmodels with 4 < DAICi < 7 have considerably less support(Burnham & Anderson, 2002). We also calculated normalizedAkaike weights (wi) for each model, which gives the relative like-lihood of the model i, given the data (Burnham & Anderson, 2002).

M.-J. Holveck et al. / Animal Behaviour 104 (2015) 229e245 245
Of note, in the information theory approach, we only included asubset of all possible models (namely 13 different models, seeTable A2) that is comparable to the set of models we used forstepwise model selection. Indeed, our design is not fully factorial,which means that the explanatory variables are not all fully inde-pendent. We were aware of this issue, and this is why we used theresiduals of the linear regression of cage volume on density andnumber of individuals and each main effect was tested afteradjusting for other main effects.
ResultsThe first model including the significant interaction effect of
density)sex ratio (as found by stepwise model selection, Table 3)has considerably less support only for the proportion of courtshipsaborted by the male (Table A2) and is almost within the ‘best’subset for the proportion of courtships aborted by the female(Table A2). Reassuringly, all other ‘best’ subsets of models(DAICc � 2) include the significant predictors as found by backwardstepwise model selection (Table A2).
Related Documents











