Journal of Water and Environment Technology, Vol.12, No.4, 2014 Address correspondence to Sorawit Powtongsook, Center of Excellence for Marine Biotechnology, National Center of Genetic Engineering and Biotechnology, Email: [email protected] Received May 11, 2013, Accepted January 17, 2014. - 347 - Denitrification and Dissimilatory Nitrate Reduction to Ammonium (DNRA) Activities in Freshwater Sludge and Biofloc from Nile Tilapia Aquaculture Systems Pokchat CHUTIVISUT*, Wiboonluk PUNGRASMI*, Sorawit POWTONGSOOK** , *** *Department of Environmental Engineering, Faculty of Engineering, Chulalongkorn University, 254 Phayathai Road, Pathumwan, Bangkok 10330, Thailand **Center of Excellence for Marine Biotechnology, Department of Marine Science, Faculty of Science, Chulalongkorn University, 254 Phayathai Road, Pathumwan, Bangkok 10330, Thailand ***National Center of Genetic Engineering and Biotechnology, 113 Thailand Science Park, Phahonyothin Road, Khlong Luang, Pathum Thani 12120, Thailand ABSTRACT Suspended organic sludge from freshwater and biofloc Nile tilapia systems were examined for the presence of denitrifying and dissimilatory nitrate reduction to ammonium (DNRA) activities under nitrate and sulfide stimulation. Initial nitrate concentrations at 25 and 100 mg NO 3 – -N/L were added to the freshwater sludge and biofloc samples to simulate low and high nitrate levels that are normally found in aquaculture systems. The results showed that freshwater sludge and biofloc both had denitrifying activity immediately after nitrate addition. However, ammonium accumulated in the biofloc reactors but not in the freshwater reactors, indicating the activity of DNRA in the high C/N biofloc particles. The influence of sulfide on nitrate reduction was also studied by adding different concentrations of sulfide along with 100 mg NO 3 – -N/L. The results showed that elevated sulfide concentrations partially inhibited denitrification in the freshwater sludge and caused nitrite and ammonium accumulation, in which ammonium formation was probably responsible by DNRA activity. In sulfide-added biofloc reactors, ammonium accumulated at the same level as found in the biofloc reactors without sulfide. Therefore, DNRA bacteria residing in the biofloc aquaculture system were more likely to be heterotrophs that did not use sulfide as their electron donor. Keywords: biofloc, denitrification, dissimilatory nitrate reduction to ammonium (DNRA), freshwater aquaculture sludge, Nile tilapia, sulfide INTRODUCTION Aquaculture systems generally produce inorganic nitrogen and organic suspended solid wastes that are capable of causing adverse effects if directly discharged to the environment. Recirculating aquaculture system (RAS) has been developed to overcome this problem, in which, with proper wastewater treatment process, water can be reused in aquaculture systems and the water discharge can be minimized to almost zero. Most of the treatment systems in RAS target the removal of ammonium, nitrite, and nitrate, as well as the separation of particulate organic matters (POMs) derived from fish feed, fish feces, and suspended solids (Cripps and Bergheim, 2000). However, instead of discarding the solids as organic wastes, POMs can further be used as the source of carbon for anoxic denitrification process (van Rijn et al., 1995). Normally, external carbon has to be added to a denitrifying reactor due to the low dissolved organic carbon in the aquaculture system. However, with the use of POMs, the growth of denitrifying bacteria can be promoted in a sustainable way and the organic sludge can also be stabilized along the process. These POMs are removed in a separate compartment and

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Water and Environment Technology, Vol.12, No.4, 2014
Address correspondence to Sorawit Powtongsook, Center of Excellence for Marine Biotechnology, National Center of Genetic Engineering and Biotechnology, Email: [email protected] Received May 11, 2013, Accepted January 17, 2014.
- 347 -
Denitrification and Dissimilatory Nitrate Reduction to Ammonium (DNRA) Activities in Freshwater Sludge and Biofloc from Nile Tilapia Aquaculture Systems Pokchat CHUTIVISUT*, Wiboonluk PUNGRASMI*, Sorawit POWTONGSOOK**, ***
*Department of Environmental Engineering, Faculty of Engineering, Chulalongkorn University, 254 Phayathai Road, Pathumwan, Bangkok 10330, Thailand
**Center of Excellence for Marine Biotechnology, Department of Marine Science, Faculty of Science, Chulalongkorn University, 254 Phayathai Road, Pathumwan, Bangkok 10330, Thailand
***National Center of Genetic Engineering and Biotechnology, 113 Thailand Science Park, Phahonyothin Road, Khlong Luang, Pathum Thani 12120, Thailand
ABSTRACT Suspended organic sludge from freshwater and biofloc Nile tilapia systems were examined for the presence of denitrifying and dissimilatory nitrate reduction to ammonium (DNRA) activities under nitrate and sulfide stimulation. Initial nitrate concentrations at 25 and 100 mg NO3
–-N/L were added to the freshwater sludge and biofloc samples to simulate low and high nitrate levels that are normally found in aquaculture systems. The results showed that freshwater sludge and biofloc both had denitrifying activity immediately after nitrate addition. However, ammonium accumulated in the biofloc reactors but not in the freshwater reactors, indicating the activity of DNRA in the high C/N biofloc particles. The influence of sulfide on nitrate reduction was also studied by adding different concentrations of sulfide along with 100 mg NO3
–-N/L. The results showed that elevated sulfide concentrations partially inhibited denitrification in the freshwater sludge and caused nitrite and ammonium accumulation, in which ammonium formation was probably responsible by DNRA activity. In sulfide-added biofloc reactors, ammonium accumulated at the same level as found in the biofloc reactors without sulfide. Therefore, DNRA bacteria residing in the biofloc aquaculture system were more likely to be heterotrophs that did not use sulfide as their electron donor. Keywords: biofloc, denitrification, dissimilatory nitrate reduction to ammonium (DNRA),
freshwater aquaculture sludge, Nile tilapia, sulfide
INTRODUCTION Aquaculture systems generally produce inorganic nitrogen and organic suspended solid wastes that are capable of causing adverse effects if directly discharged to the environment. Recirculating aquaculture system (RAS) has been developed to overcome this problem, in which, with proper wastewater treatment process, water can be reused in aquaculture systems and the water discharge can be minimized to almost zero. Most of the treatment systems in RAS target the removal of ammonium, nitrite, and nitrate, as well as the separation of particulate organic matters (POMs) derived from fish feed, fish feces, and suspended solids (Cripps and Bergheim, 2000). However, instead of discarding the solids as organic wastes, POMs can further be used as the source of carbon for anoxic denitrification process (van Rijn et al., 1995). Normally, external carbon has to be added to a denitrifying reactor due to the low dissolved organic carbon in the aquaculture system. However, with the use of POMs, the growth of denitrifying bacteria can be promoted in a sustainable way and the organic sludge can also be stabilized along the process. These POMs are removed in a separate compartment and

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 348 -
degraded through anaerobic decomposition. Denitrifying reactors supplied with digested sludge have been found to operate successfully without the need for external carbon addition (Aboutboul et al., 1995; Klas et al., 2006; Tal et al., 2009). Hence, this is the method of choice when it comes to nitrate removal in aquaculture systems. Denitrification, however, is not the only process for nitrate reduction. Other nitrogen respiration pathways such as anaerobic ammonium oxidation (anammox), which is the transformation of nitrite and ammonium into dinitrogen gas, and dissimilatory nitrate reduction to ammonium (DNRA), which is the reduction of nitrate to nitrite followed by the reduction of nitrite to ammonium, have also been proven to contribute a significant proportion to nitrate removal in various environments (Kartal et al., 2007; Smith et al., 2007; Dong et al., 2011). Favorable conditions for these pathways are hypothesized to depend mainly on the concentration and type of electron donor, the amount of inorganic nitrogen compounds present, along with the redox potential found in those systems (van Rijn et al., 2006; Kraft et al., 2011). To date, there had been several reports on the presence of anammox bacteria in aquaculture treatment systems that use degraded sludge as the substrates for denitrification (Tal et al., 2006; Lahav et al., 2009). However, the involvement of the DNRA process in aquaculture systems is still uncertain. So far, DNRA has been found in the sediment below cages of an open water fish farm with activity up to 7-fold greater than denitrification (Christensen et al., 2000). The activity of DNRA was also detected along with autotrophic denitrification in mariculture sludge under high sulfide concentration (Sher et al., 2008). The understanding of the factors influencing the DNRA process in RAS, however, is still limited. The occurrence of DNRA is undesirable in wastewater treatment systems because it competes with denitrification and reduces the process efficiency by producing ammonium, hence nitrogen waste is still retained in the water. To control the occurrence of DNRA, conditions affecting its presence and activity need to be studied in more detail. The nitrogen pathways that can occur in aquaculture systems under oxic and anoxic conditions are shown in Fig. 1.
Fig. 1 - Nitrogen pathways that can occur in aquaculture systems under
oxic and anoxic conditions.
NO2-NH4
+
NO3-
NO2-
NO
N2O
N2
NH4++N2
NO2-
NH4+
NO3- Nitrification
Denitrification
Anammox
Oxic
Anoxic
Organic Nitrogen
Organic Nitrogen
DNRA

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 349 -
During the organic sludge digestion, sulfide was also found to be released and stimulates the community of autotrophic denitrifying bacteria under anoxic conditions (Cytryn et al., 2003; Schwermer et al., 2010; Shao et al., 2010). Since a sulfide-rich environment has been hypothesized to favor DNRA (Brunet and Garcia-Gil, 1996; Sher et al., 2008), the presence of the DNRA process is to be expected in this system and should also be an important pathway for nitrate reduction. Apart from sulfide, the high C/N ratio is another factor that promotes the growth of DNRA bacteria (van Rijn et al., 2006; Schreier et al., 2010). The high C/N condition can be found in the biofloc system, which is a type of aquaculture that uses external carbon substrates to enhance the growth of heterotrophic bacteria in the fish tank (Crab et al., 2012). Ammonium uptake by microbial biomass in the biofloc, thus, eliminates nitrogen wastes without the need for slow-growing autotrophic nitrifiers. This high C/N ratio should also support the growth of DNRA bacteria, hence, under oxygen depleted conditions, ammonium regeneration is also likely to occur in the biofloc system. The objective of this study was to investigate the occurrence of denitrification and DNRA processes in sludge taken from freshwater and biofloc aquaculture systems. Activities of denitrifying and DNRA bacteria in both sludge types were compared by monitoring the change in nitrate, nitrite, and ammonium concentrations in batch reactors. The effects of sulfide on nitrate reduction in both freshwater sludge and biofloc were also studied by adding different concentrations of sulfide to the sludge samples. Results of the experiment were discussed in terms of the presence of different nitrogen transformation pathways in the systems as well as the effects of environmental conditions on the occurrence of these microbial processes. MATERIALS AND METHODS Sources of aquaculture sludge Freshwater aquaculture sludge was taken from 4-m3 indoor Nile tilapia culture tank with internal nitrification media for ammonium removal. Fish density in the tank was equal to 9.5 kg/m3 with suspended solid concentration of 126.0 ± 17.4 mg/L. A biofloc sample was collected from 800-L Nile tilapia tank with fish density of 9 kg/m3. Starch was added daily to this biofloc aquaculture system as the external carbon source to provide a C:N ratio of 20:1. Suspended solid concentration in the biofloc aquaculture system was equal to 498.3 ± 30.1 mg/L. Evaluation of sludge digestion and nitrate reduction Sludge digestion and nitrate reduction experiments were conducted using 4 sets of 1-L DURAN® bottles (Schott, Germany), 2 reactors for freshwater sludge and 2 reactors for biofloc sample, at room temperature (30°C), with approximately 10 g (dry weight) of sludge per liter. The reactors were operated in batch mode by providing nitrate at the initial concentrations of 25 and 100 mg NO3
–-N/L to each sludge sample using NaNO3 as the source of NO3
–-N. Each reactor was supplemented with nutrients containing (per liter): 3.8 g of Na2HPO4, 1.5 g of K2HPO4, 0.1 g of MgSO4·7H2O, and 2 mL of trace element solution (van Rijn et al., 1996). Trace element solution added consisted of (per liter): 50.0 g of EDTA, 22.0 g of ZnSO4·7H2O, 7.34 g of CaCl2·2H2O, 5.06 g of MnCl2·4H2O, 4.99 g of FeSO4·7H2O, 0.195 g of NaMoO4·2H2O, 1.57 g of CuSO4·5H2O, and 1.61 g of CoCl2·6H2O (Vishniac and Santer, 1957). The medium was stirred with a

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 350 -
magnetic stirrer to maintain the homogenous state of the medium. Water samples were taken every 30 or 60 minutes depending on the initial concentration of nitrate. Effects of sulfide on nitrate reduction Effects of sulfide on nitrate reduction was studied by adding sulfide (Na2S·H2O) at the initial concentrations of 50 and 100 mg S2–/L in freshwater sludge, and 25 and 50 mg S2–/L in biofloc samples. The experiments were conducted in batch mode using 4 sets of 1-L Duran® bottles at room temperature (30°C). Nitrate was added as NaNO3 at 100 mg NO3
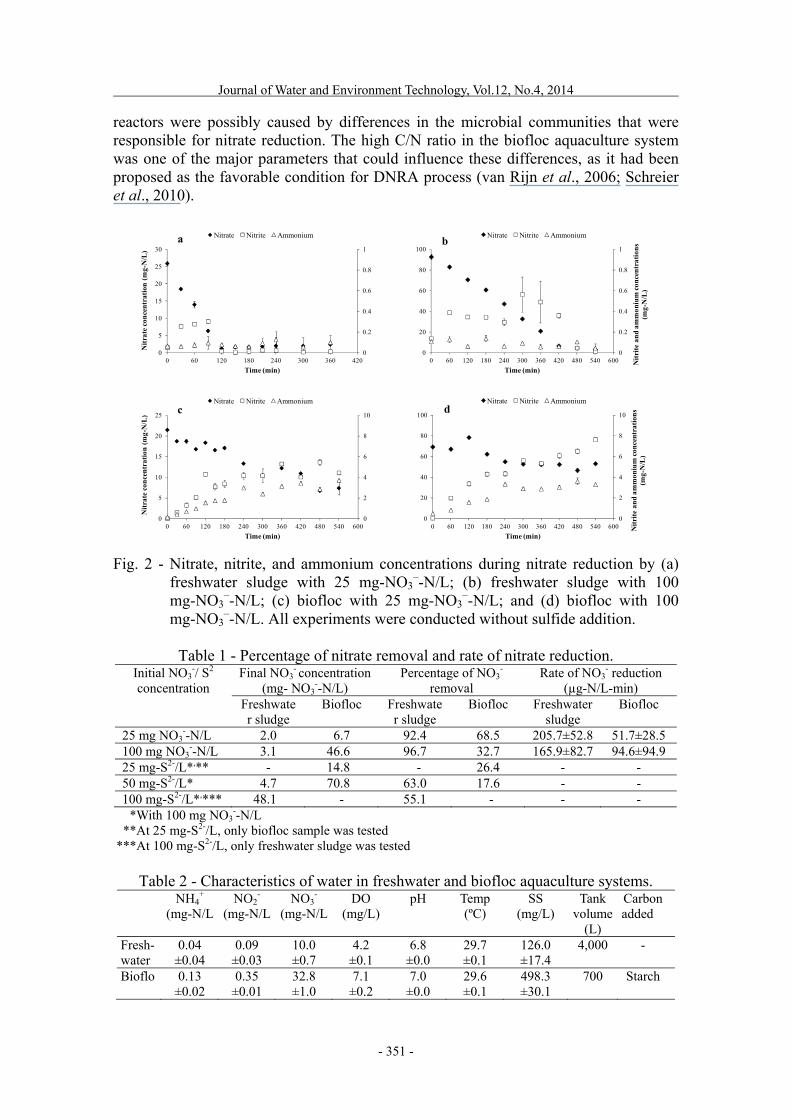
–-N/L for all sulfide concentrations. The culture medium supplemented and water samplings were similar to those of the previous experiment. Analytical methods Nitrate was analyzed using an ultraviolet spectrophotometric method (PowerWave XS2 Microplate Spectrophotometer, BioTek Instruments, USA) according to Standard Methods for Water and Wastewater Analysis (APHA, 1992). Nitrite and ammonium were both analyzed colorimetrically using the sulfanilamide method (APHA, 1992) and the salicylate-hypochlorite method (Bower and Holm-Hansen, 1980), respectively. Sulfide fixation was performed by adding 2.5-mL of sample to 1-mL of 5% zinc acetate solution (Cytryn et al., 2003), then the concentration of sulfide was measured colorimetrically using the methylene blue method (Cline, 1969). RESULTS AND DISCUSSION Nitrate reduction in freshwater sludge and biofloc reactors Nitrate reduction in freshwater sludge and biofloc reactors was monitored by measuring the changes in nitrate, nitrite, and ammonium concentrations in the reactors. The results are shown in Fig. 2. For freshwater sludge reactors, nitrate decreased rapidly at the rates of 205.7 ± 52.8 and 165.9 ± 82.7 µg-N/L-min for the initial concentrations of 25 and 100 mg NO3
–-N/L (as indicated in Table 1), respectively. A slight increase in nitrite was found during nitrate reduction for both of the freshwater sludge reactors, indicating the transformation of nitrate to nitrite. Ammonium in both of the freshwater sludge reactors was low and comparable to the concentration found in the fish tank from where the sludge was originally taken (0.04 ± 0.04 mg-N/L, as shown in Table 2). Changes in nitrate, nitrite, and ammonium concentrations indicated that denitrifying bacteria were already present in the freshwater aquaculture system and could function readily under anoxic conditions, which can be seen from the concomitant reduction of nitrate and nitrite. The activity of DNRA could not be detected under either nitrate concentration, which otherwise should result in the accumulation of ammonium in the system. For the biofloc reactors, nitrate reduction rates were equal to 51.7 ± 28.5 and 94.6 ± 94.9 µg-N/L-min for the nitrate concentrations of 25 and 100 mg NO3
–-N/L, respectively. These reduction rates were slower than those found in the freshwater sludge reactors, while nitrite and ammonium were found to accumulate in the biofloc reactors. Peaks of nitrite and ammonium both exceeded the toxicity threshold for aquatic animals, which is not desirable for water recycling. The absence of ammonium accumulation in freshwater sludge showed that ammonium release from sludge decomposition was unlikely to occur at this stage, and that ammonium increase in the biofloc reactors was likely caused by DNRA bacteria that originally resided in the biofloc sample. Hence, the differences in nitrogen transformation patterns in freshwater sludge and biofloc
Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 351 -
reactors were possibly caused by differences in the microbial communities that were responsible for nitrate reduction. The high C/N ratio in the biofloc aquaculture system was one of the major parameters that could influence these differences, as it had been proposed as the favorable condition for DNRA process (van Rijn et al., 2006; Schreier et al., 2010).
Fig. 2 - Nitrate, nitrite, and ammonium concentrations during nitrate reduction by (a)
freshwater sludge with 25 mg-NO3–-N/L; (b) freshwater sludge with 100
mg-NO3–-N/L; (c) biofloc with 25 mg-NO3
–-N/L; and (d) biofloc with 100 mg-NO3
–-N/L. All experiments were conducted without sulfide addition.
Table 1 - Percentage of nitrate removal and rate of nitrate reduction. Initial NO3
-/ S2 concentration
Final NO3- concentration
(mg- NO3--N/L)
Percentage of NO3-
removal Rate of NO3
- reduction (µg-N/L-min)
Freshwater sludge
Biofloc Freshwater sludge
Biofloc Freshwater sludge
Biofloc
25 mg NO3--N/L 2.0 6.7 92.4 68.5 205.7±52.8 51.7±28.5
100 mg NO3--N/L 3.1 46.6 96.7 32.7 165.9±82.7 94.6±94.9
25 mg-S2-/L*,** - 14.8 - 26.4 - - 50 mg-S2-/L* 4.7 70.8 63.0 17.6 - - 100 mg-S2-/L*,*** 48.1 - 55.1 - - -
*With 100 mg NO3--N/L
**At 25 mg-S2-/L, only biofloc sample was tested ***At 100 mg-S2-/L, only freshwater sludge was tested
Table 2 - Characteristics of water in freshwater and biofloc aquaculture systems. NH4
+
(mg-N/LNO2
-
(mg-N/LNO3
-
(mg-N/LDO
(mg/L)pH Temp
(ºC) SS
(mg/L) Tank
volume (L)
Carbon added
Fresh- water
0.04 ±0.04
0.09 ±0.03
10.0 ±0.7
4.2 ±0.1
6.8 ±0.0
29.7 ±0.1
126.0 ±17.4
4,000 -
Bioflo 0.13 ±0.02
0.35 ±0.01
32.8 ±1.0
7.1 ±0.2
7.0 ±0.0
29.6 ±0.1
498.3 ±30.1
700 Starch
0
0.2
0.4
0.6
0.8
1
0
5
10
15
20
25
30
0 60 120 180 240 300 360 420
Nit
rate
con
cen
trat
ion
(m
g-N
/L)
Time (min)
a Nitrate Nitrite Ammonium
0
0.2
0.4
0.6
0.8
1
0
20
40
60
80
100
0 60 120 180 240 300 360 420 480 540 600
Time (min)
bNitrate Nitrite Ammonium
Nit
rite
an
d a
mm
oniu
m c
once
ntr
atio
ns
(mg-
N/L
)
0
2
4
6
8
10
0
5
10
15
20
25
0 60 120 180 240 300 360 420 480 540 600
Nit
rate
con
cen
trat
ion
(m
g-N
/L)
Time (min)
cNitrate Nitrite Ammonium
0
2
4
6
8
10
0
20
40
60
80
100
0 60 120 180 240 300 360 420 480 540 600
Time (min)
dNitrate Nitrite Ammonium
Nit
rite
an
d a
mm
oniu
m c
once
ntr
atio
ns
(mg-
N/L
)

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 352 -
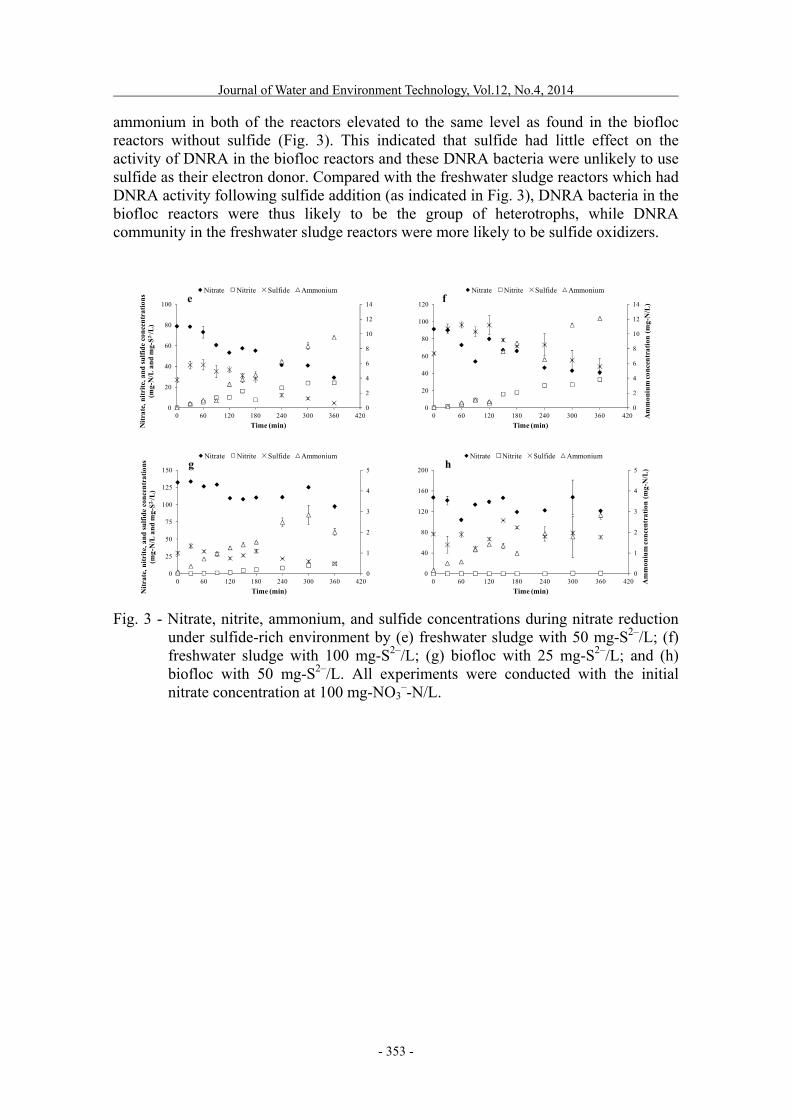
When comparing the results from these two sludge types, denitrifying bacteria in the freshwater sludge seem to be more active than those found in the biofloc reactors. This might be due, in part, to the differences in the fish tank volumes from where the two sludge samples were originally taken. With the larger freshwater aquaculture system (as indicated in Table 2), water mixing by aeration was not effective enough to suspend most of the sludge in the water column. Accumulation of the bottom sludge layer was hence usually found, which has microaerobic or anoxic conditions that favor the growth of denitrifying bacteria. The low nitrate concentration (10.0 ± 0.7 mg-N/L, as shown in Table 2) in the freshwater compared to the biofloc aquaculture system also supports this hypothesis that denitrifiers were present and functionally active in the freshwater sludge sample. On the contrary, the biofloc aquaculture system had a much smaller fish tank volume which meant that the water mixing was sustained well by the strong aeration. An accumulation of nitrate was found in the biofloc aquaculture system (32.8 ± 1.0 mg-N/L, Table 2) as a result of the aerobic nitrification process in the biofloc particles (Nootong et al., 2011). The bottom sludge layer accumulation in the fish tank thus plays a significant role in controlling the fate of nitrate in aquaculture systems. Effects of sulfide on nitrate reduction Percentages of nitrate removal after sulfide addition were found to be lower than those without sulfide for both freshwater sludge and biofloc reactors (Table 1). Despite the fact that sulfide can contribute as an electron donor for autotrophic denitrifying bacteria (Cytryn et al., 2005a; Cytryn et al., 2005b; Shao et al., 2010), certain concentrations of sulfide also inhibit the enzymes responsible for denitrification (Brunet and Garcia-Gil, 1996). Slower rates of nitrate reduction under high sulfide (as shown in Fig. 3) could be the result of the partial inhibition of the denitrifying bacterial community. Nevertheless, the fluctuation of nitrate under elevated sulfide conditions made the calculation of exact nitrate reduction rates somewhat unreliable, especially for the biofloc reactors in which nitrate only decreased slightly. However, the decrease in sulfide concentration simultaneously with nitrate was found in the freshwater sludge reactors, which was possibly caused by sulfide-oxidizing denitrifiers. Furthermore, sulfide at 50 and 100 mg-S2–/L in the freshwater sludge reactors could induce significant accumulation of nitrite up to 24.2 and 32.9 mg-N/L (Fig. 3), respectively, which were substantially higher than those found in the freshwater sludge reactors without sulfide addition (Fig. 2). The increase in nitrite suggested that nitrite reduction was the rate-limiting step in the denitrification process under this condition, where nitrate seems to be continuously transformed to nitrite. Ammonium in the sulfide-added freshwater sludge reactors also accumulated to 9.5 mg-N/L for 50 mg-S2–/L and 12.1 mg-N/L for 100 mg-S2–/L. This implies that another microbial pathway, which was likely to be DNRA, competed with denitrifying bacteria under the high sulfide conditions in the freshwater sludge reactors. This result was in accordance with the previous findings that sulfide could drive DNRA activity, as suggested by the works of Brunet and Garcia-Gil (1996) and Sher et al. (2008). As for the biofloc reactors, sulfide also had certain inhibitory effect on nitrate reduction by lowering the removal of nitrate to 26.4 and 17.6 percent for 25 and 50 mg-S2–/L, respectively (as shown in Table 1). In addition, for 25 mg-S2–/L, nitrite accumulated to 14.2 mg-N/L but hardly increased in the biofloc reactor with 50 mg-S2–/L, suggesting that nitrate reduction step was hindered by the higher sulfide concentration. Furthermore,
Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 353 -
ammonium in both of the reactors elevated to the same level as found in the biofloc reactors without sulfide (Fig. 3). This indicated that sulfide had little effect on the activity of DNRA in the biofloc reactors and these DNRA bacteria were unlikely to use sulfide as their electron donor. Compared with the freshwater sludge reactors which had DNRA activity following sulfide addition (as indicated in Fig. 3), DNRA bacteria in the biofloc reactors were thus likely to be the group of heterotrophs, while DNRA community in the freshwater sludge reactors were more likely to be sulfide oxidizers.
Fig. 3 - Nitrate, nitrite, ammonium, and sulfide concentrations during nitrate reduction
under sulfide-rich environment by (e) freshwater sludge with 50 mg-S2–/L; (f) freshwater sludge with 100 mg-S2–/L; (g) biofloc with 25 mg-S2–/L; and (h) biofloc with 50 mg-S2–/L. All experiments were conducted with the initial nitrate concentration at 100 mg-NO3
–-N/L.
0
2
4
6
8
10
12
14
0
20
40
60
80
100
0 60 120 180 240 300 360 420
Time (min)
eNitrate Nitrite Sulfide Ammonium
Nit
rate
, nit
rite
, an
d s
ulf
ide
con
cen
trat
ion
s (m
g-N
/L a
nd
mg-
S2-
/L)
0
2
4
6
8
10
12
14
0
20
40
60
80
100
120
0 60 120 180 240 300 360 420
Time (min)
fNitrate Nitrite Sulfide Ammonium
Am
mon
ium
con
cen
trat
ion
(m
g-N
/L)
0
1
2
3
4
5
0
25
50
75
100
125
150
0 60 120 180 240 300 360 420
Time (min)
gNitrate Nitrite Sulfide Ammonium
Nit
rate
, nit
rite
, an
d s
ulf
ide
con
cen
trat
ion
s (m
g-N
/L a
nd
mg-
S2-
/L)
0
1
2
3
4
5
0
40
80
120
160
200
0 60 120 180 240 300 360 420
Time (min)
hNitrate Nitrite Sulfide Ammonium
Am
mon
ium
con
cen
trat
ion
(m
g-N
/L)

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 354 -
CONCLUSIONS When comparing nitrate reduction rates between freshwater sludge and biofloc without sulfide addition, it was found that the freshwater sludge had higher nitrate reduction rates without any undesirable ammonium and nitrite accumulation. However, with sulfide added to the freshwater sludge reactors, significant increases in both nitrite and ammonium were observed. This indicates the partial inhibition of denitrification along with an increase in DNRA activity under elevated sulfide conditions. For the biofloc reactors, DNRA was detected at the same level for all of the reactors with or without sulfide addition. Since sulfide had little influence on DNRA activity in the biofloc reactors, these DNRA bacteria were likely to be heterotrophs that did not use sulfide as the source of energy. This was in contrast to the DNRA bacteria found in the freshwater sludge reactors which seem more likely to be sulfide oxidizers. The understanding of the DNRA process could provide useful information that can be used to solve the periodic ammonium accumulation in the denitrifying reactor. However, confirmation with more precise techniques, e.g. molecular methods with genes related to the DNRA pathway, is recommended to prove the presence and activity of DNRA bacteria in aquaculture systems. Nevertheless, the results of this study have shown that denitrifying and DNRA activities exist in both freshwater and biofloc aquaculture systems and can perform activities that drive the transformation of nitrogen when conditions are favorable. ACKNOWLEDGEMENTS This research was financially supported by the Integrated Innovation Academic Center, Chulalongkorn University Centenary Academic Development Project (CU56-FW14). It received partial support from the Higher Education Research Promotion and National Research University Project of Thailand, Office of the Higher Education Commission and the Ratchadaphiseksomphot Endowment Fund (FW1017A). Additional support was obtained from the Chulalongkorn University Graduate Scholarship to commemorate the 72nd Anniversary of His Majesty King Bhumibol Adulyadej. Equipment used was provided by the Thai Government Stimulus Package 2 (TKK2555) under the Project for the Establishment of Comprehensive Center for Innovative Food, Health Products and Agriculture. REFERENCES Aboutboul Y., Arviv R. and van Rijn J. (1995) Anaerobic treatment of intensive fish
culture effluents: volatile fatty acid mediated denitrification. Aquaculture, 133(1), 21-32.
APHA (1992) Standard methods for the examination of water and wastewater, 18th ed., American Public Health Association/American Water Works Association/Water Environment Federation, New York, USA.
Bower C. E. and Holm-Hansen T. (1980) A salicylate-hypochlorite method for determining ammonia in seawater. Can. J. Fish. Aquat. Sci., 37(5), 794-798.
Brunet R. C. and Garcia-Gil L. J. (1996) Sulfide-induced dissimilatory nitrate reduction to ammonia in anaerobic freshwater sediments. FEMS Microbiol. Ecol., 21(2), 131-138.
Christensen P. B., Rysgaard S., Sloth N. P., Dalsgaard T. and Schwaerter S. (2000)

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 355 -
Sediment mineralization, nutrient fluxes, denitrification and dissimilatory nitrate reduction to ammonium in an estuarine fjord with sea cage trout farms. Aquat. Microb. Ecol., 21(1), 73-84.
Cline J. D. (1969) Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr., 14(3), 454-458.
Crab R., Defoirdt T., Bossier P. and Verstraete W. (2012) Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture, 356-357, 351-356.
Cripps S. J. and Bergheim A. (2000) Solids management and removal for intensive land-based aquaculture production systems. Aquacult. Eng., 22(1-2), 33-56.
Cytryn E., Gelfand I., Barak Y., van Rijn J. and Minz D. (2003) Diversity of microbial communities correlated to physiochemical parameters in a digestion basin of a zero-discharge mariculture system. Environ. Microbiol., 5(1), 55-63.
Cytryn E., Minz D., Gelfand I., Neori A., Gieseke A., de Beer D. and van Rijn J. (2005a) Sulfide-oxidizing activity and bacterial community structure in a fluidized bed reactor from a zero-discharge mariculture system. Environ. Sci. Technol., 39(6), 1802-1810.
Cytryn E., van Rijn J., Schramm A., Gieseke A., de Beer D. and Minz D. (2005b) Identification of bacteria potentially responsible for oxic and anoxic sulfide oxidation in biofilters of a recirculating mariculture system. Appl. Environ. Microbiol., 71(10), 6134-6141.
Dong L. F., Sobey M. N., Smith C. J., Rusmana I., Phillips W., Stott A., Osborn A. M. and Nedwell D. B. (2011) Dissimilatory reduction of nitrate to ammonium, not denitrification or anammox, dominates benthic nitrate reduction in tropical estuaries. Limnol. Oceanogr., 56(1), 279-291.
Kartal B., Kuypers M. M. M., Lavik G., Schalk J., Op den Camp H. J. M., Jetten M. S. M. and Strous M. (2007) Anammox bacteria disguised as denitrifiers: nitrate reduction to dinitrogen gas via nitrite and ammonium. Environ. Microbiol., 9(3), 635-642.
Klas S., Mozes N. and Lahav O. (2006) A conceptual, stoichiometry-based model for single-sludge denitrification in recirculating aquaculture systems. Aquaculture, 259(1-4), 328-341.
Kraft B., Strous M. and Tegetmeyer H. E. (2011) Microbial nitrate respiration – Genes, enzymes and environmental distribution. J. Biotechnol., 155(1), 104-117.
Lahav O., Massada I. B., Yackoubov D., Zelikson R., Mozes N., Tal Y. and Tarre S. (2009) Quantification of anammox activity in a denitrification reactor for a recirculating aquaculture system. Aquaculture, 288(1-2), 76-82.
Nootong K., Pavasant P. and Powtongsook S. (2011) Effects of organic carbon addition in controlling inorganic nitrogen concentrations in a biofloc system. J. World Aquacult. Soc., 42(3), 339-346.
Schreier H. J., Mirzoyan N. and Saito K. (2010) Microbial diversity of biological filters in recirculating aquaculture systems. Curr. Opin. Biotechnol., 21(3), 318-325.
Schwermer C. U., Ferdelman T. G., Stief P., Gieseke A., Rezakhani N., van Rijn J., de Beer D. and Schramm A. (2010) Effect of nitrate on sulfur transformations in sulfidogenic sludge of a marine aquaculture biofilter. FEMS Microbiol. Ecol., 72(3), 476-484.
Shao M., Zhang T. and Fang H. H. (2010) Sulfur-driven autotrophic denitrification: diversity, biochemistry, and engineering applications. Appl. Environ. Microbiol.,

Journal of Water and Environment Technology, Vol.12, No.4, 2014
- 356 -
88(5), 1027-1042. Sher Y., Schneider K., Schwermer C. U. and van Rijn J. (2008) Sulfide-induced nitrate
reduction in the sludge of an anaerobic digester of a zero-discharge recirculating mariculture system. Water Res., 42(16), 4386-4392.
Smith C. J., Nedwell D. B., Dong L. F. and Osborn A. M. (2007) Diversity and abundance of nitrate reductase genes (narG and napA), nitrite reductase genes (nirS and nrfA), and their transcripts in estuarine sediments. Appl. Environ. Microbiol., 73(11), 3612-3622.
Tal Y., Schreier H. J., Sowers K. R., Stubblefield J. D., Place A. R. and Zohar Y. (2009) Environmentally sustainable land-based marine aquaculture. Aquaculture, 286(1-2), 28-35.
Tal Y., Watts J. E. M. and Schreier H. J. (2006) Anaerobic ammonium-oxidizing (anammox) bacteria and associated activity in fixed-film biofilters of a marine recirculating aquaculture system. Appl. Environ. Microbiol., 72(4), 2896-2904.
van Rijn J., Fonarev N. and Berkowitz B. (1995) Anaerobic treatment of intensive fish culture effluents: digestion of fish feed and release of volatile fatty acids. Aquaculture, 133(1), 9-20.
van Rijn J., Tal Y. and Barak Y. (1996) Influence of volatile fatty acids on nitrite accumulation by a Pseudomonas stutzeri strain isolated from a denitrifying fluidized bed reactor. Appl. Environ. Microbiol., 62(7), 2615-2620.
van Rijn J., Tal Y. and Schreier H. J. (2006) Denitrification in recirculating systems: Theory and applications. Aquacult. Engineer., 34(3), 364-376.
Vishniac W. and Santer M. (1957) The thiobacilli. Bacteriol. Rev., 21(3), 195-213.
Related Documents











