Dendrimer–protein interactions studied by tryptophan room temperature phosphorescence Edi Gabellieri a , Giovanni B. Strambini a , Dzmitry Shcharbin b , Barbara Klajnert b , Maria Bryszewska b a Istituto di Biofisica-CNR, Area della Ricerca, via G. Moruzzi 1, 56124 Pisa, Italy b Department of General Biophysics, University of Lodz, 12/16 Banacha St., 90-237 Lodz, Poland Abstract Dendrimers are a relatively new class of materials with unique molecular architectures, which provide promising opportunities for biological applications as DNA carriers and drug delivery systems. Progress in these fields, however, requires knowledge of their potential interactions with biological components at cellular and molecular level. This study utilizes Trp phosphorescence spectroscopy to examine possible perturbations of the protein native fold in solution by neutral, positively and negatively charged fifth generation polyamidoamine (PAMAM) dendrimers. Phosphorescence lifetime measurements, conducted on model proteins varying in the degree of burial of the triplet probe and in quaternary structure, show that dendrimers interact with proteins in solutions forming stable complexes in which the protein structure may be significantly altered, particularly in superficial, flexible regions of the polypeptide. Both electrostatic and non electrostatic interactions can give rise to stable complexes, whose affinity and limited number of binding sites distinguish them from mere aspecific molecular associations. Of direct relevance for the application of these polymers in the medical field, structural alterations have also been detected in human plasma proteins such as serum albumin and immunoglobulins (Ig). The above results suggest that Trp phosphorescence may provide a useful monitor for working out experimental conditions and protocols that help preserve the structural integrity of proteins in the presence of these polymers.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dendrimer–protein interactions studied by tryptophan room
temperature phosphorescence
Edi Gabellieri a, Giovanni B. Strambini
a, Dzmitry Shcharbin
b,
Barbara Klajnert b, Maria Bryszewska
b
a Istituto di Biofisica-CNR, Area della Ricerca, via G. Moruzzi 1, 56124 Pisa,
Italy
b Department of General Biophysics, University of Lodz, 12/16 Banacha St.,
90-237 Lodz, Poland
Abstract
Dendrimers are a relatively new class of materials with unique molecular
architectures, which provide promising opportunities for biological applications as DNA
carriers and drug delivery systems. Progress in these fields, however, requires knowledge of
their potential interactions with biological components at cellular and molecular level. This
study utilizes Trp phosphorescence spectroscopy to examine possible perturbations of the
protein native fold in solution by neutral, positively and negatively charged fifth generation
polyamidoamine (PAMAM) dendrimers. Phosphorescence lifetime measurements, conducted
on model proteins varying in the degree of burial of the triplet probe and in quaternary
structure, show that dendrimers interact with proteins in solutions forming stable complexes
in which the protein structure may be significantly altered, particularly in superficial, flexible
regions of the polypeptide. Both electrostatic and non electrostatic interactions can give rise to
stable complexes, whose affinity and limited number of binding sites distinguish them from
mere aspecific molecular associations. Of direct relevance for the application of these
polymers in the medical field, structural alterations have also been detected in human plasma
proteins such as serum albumin and immunoglobulins (Ig). The above results suggest that Trp
phosphorescence may provide a useful monitor for working out experimental conditions and
protocols that help preserve the structural integrity of proteins in the presence of these
polymers.

2
Introduction
Dendrimers are polymers synthesized in a step-wise manner from branched monomer
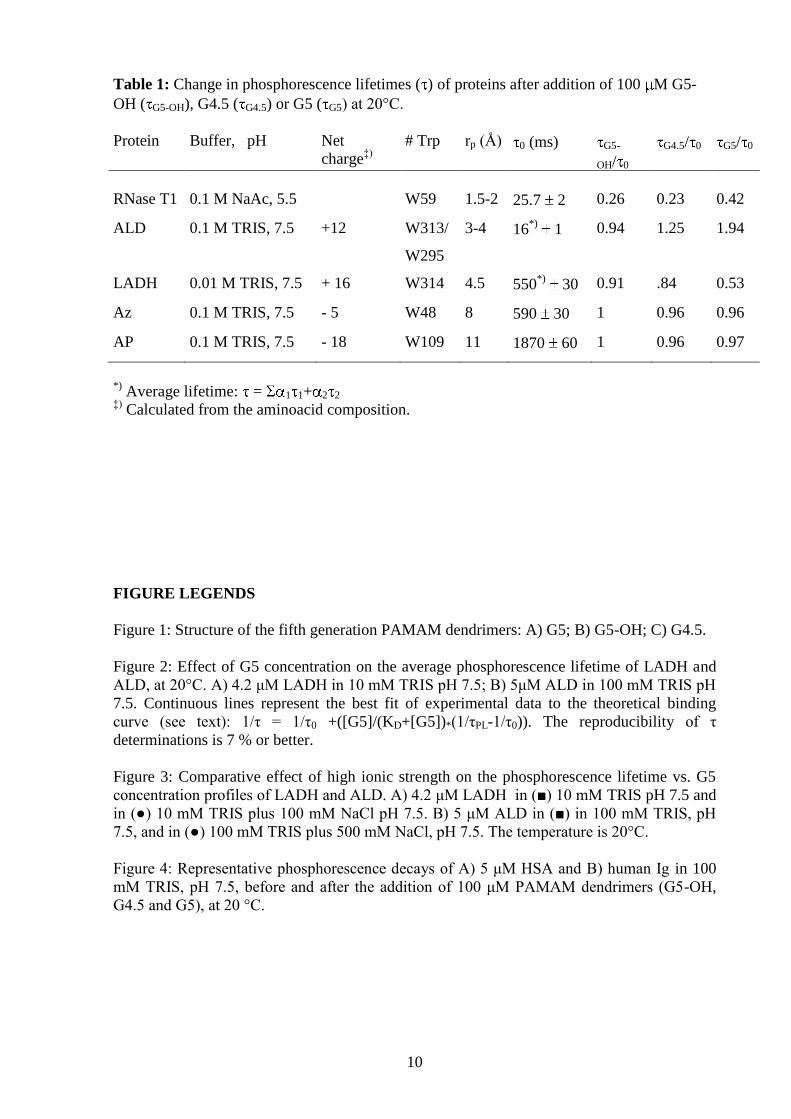
units and topologically based on the structure of a tree (1-4) (Figure 1). A typical dendrimer
consists of a multifunctional central core, branched units and surface groups. The branched
units are organized in layers called 'generations', and represent the repeating monomer unit of
these macromolecules [9]. Due to specific synthesis dendrimers possess empty internal
cavities and many superficial functional end groups, which are responsible for high solubility
and reactivity (1-3, 5, 6). Their peculiar structure makes dendrimers potentially suitable for a
variety of biomedical applications. For example, drug molecules can be attached to
dendrimers through covalent bonds. Because of the large number of terminal groups to which
drug molecules can be conjugated, dendrimers are capable to carry drugs at a high density (3,
5-8), a feature used to radiolabel antibodies (5, 7, 8). Another strategy to drug delivery is to
design dendrimers as containers to encapsulate drug molecules(5-8). Drugs entrapped inside
the dendrimer can be released slowly, which is important, for example, in reducing the
toxicity anti-tumor(5). Considering the widespread application in the biomedical field (1-10)
knowledge of their potential interactions with biological structures at the molecular,
membrane and cellular levels, is paramount. While the interaction between dendrimers and
cells has been widely studied (5, 7, 8), many aspects of their interaction with proteins are still
unknown.
Recently, we have studied the interaction between polyamidoamine (PAMAM)
dendrimers of 4th
and 5th
generations and the proteins (HSA, BSA, pure and membrane-bound
acetyl cholinesterase) by fluorescence spectroscopy, differential scanning calorimetry and
fluorescent probes (11-15). It was found that dendrimers did affect enzyme activity (11, 13),
binding properties (15) and protein conformation, their interaction depending on pH and
ionic strength (12, 14).
But there are some questions which still remain unclear: What role can the protein net
charge play in such interactions? In what manner these effects depend on protein quaternary
structure and size? How binding can differ for various proteins?
In the present investigation we extend the analysis of potential structuring effects of the
5th
generation PAMAM dendrimers by means of a sensitive spectroscopic technique based on
room temperature Trp phosphorescence. Trp phosphorescence spectroscopy has proven to be
a sensitive structural probe capable of detecting even subtle changes in protein conformation
such as those induced by ligand binding (16, 17), subunit association (18), protein-protein
interactions (19), besides variations of physical and chemical properties of the solution (20-
22). The phosphorescence spectrum reports on the polarity and homogeneity of the protein
structure around the chromophore, whereas the phosphorescence lifetime is essentially a
monitor of the local conformational flexibility, increasing 3-4 orders of magnitude from fluid
to rigid matrices (23, 24). Depending on the local flexibility of the polypeptide Trp residues
buried inside globular proteins may exhibit long-lived room temperature phosphorescence
lifetime (τ0), in the millisecond to second time range (23), which in general is promptly
affected by even minor changes in protein conformation. For this study model systems were
selected among proteins with well characterized phosphorescence properties in a wide range
of experimental conditions, known crystallographic structure and various degrees of
quaternary structure, namely, apoazurin (Az) and ribonuclease T1 (RNase T1) (monomers),
alkaline phosphatase (AP) and liver alcohol dehydrogenase (LADH) (dimers), and aldolase
(ALD) (tetramer). In general, the emission is due entirely to a single Trp residue per subunit,
which has been identified in all the above proteins except ALD. In the latter, the emission is
from either W295 or W313, or both, as the third residue, W147, is totally quenched by C149
in direct contact with it (23). From the known crystallographic structure it is possible to
determine the distance of the phosphorescence probe from the solvent as well as the charge
distribution on the protein surface. Besides these model proteins, the study of protein-

3
dendrimer interactions was extended to the serum proteins human serum albumin (HSA) and
human γ-globulins (Ig), for their direct relevance in the biomedical applications of
dendrimers. Three series of PAMAM macromolecules (G4.5, G5-OH and G5), differing in
the nature of the end groups, carboxyl, hydroxyl and amine, respectively, have permitted to
examine the influence of the chemical properties of the dendrimer surface on the protein-
dendrimer interaction.
The results point out that dendrimers bind to the proteins, also at ionic strength higher
that the physiological value, and that the electrostatic attraction is not the only mechanism
driving the interaction.
Materials and Methods
Chemicals and proteins: All chemicals were of the highest purity grade available from
commercial sources and were used without further purification.
Tris(hydroxymethyl)aminomethane (TRIS), NaCl Suprapur® and sodium acetate (NaAc)
Suprapur® were from Merck (Darmstadt, Germany). PAMAM dendrimers were purchased in
methanol solution from Sigma-Aldrich Co. (St. Louis, MO). Lyophilised ribonuclease T1
(RNase T1) from Aspergillus oryzae was from Calbiochem Corp. (San Diego, CA). Horse
liver alcohol dehydrogenase (LADH) was supplied as crystalline suspension from Fluka
Chemie GmbH (Buchs SG, Switzerland). Rabbit muscle aldolase (ALD), E. coli alkaline
phosphatase (AP), human -globulins (Ig) and human serum albumin (HSA) were from
Sigma-Aldrich Co. (St. Louis, MO). Fatty acids were removed from HSA by charcoal
treatment, following the procedure described by Chen (1967) (25). Apo-azurin (Az) was
prepared by removing copper ion from Pseudomonas aeruginosa azurin, isolated and purified
following published protocols (26). The plasmid carrying the azurin sequence was a generous
gift from Prof. Alessandro Desideri (University of Tor Vergata, Rome, Italy). Water, doubly
distilled over quartz, was purified by a Milli-Q Plus system (Millipore Corporation, Bedford,
MA). All glassware used for sample preparation was conditioned in advance by standing for
24 h in 10% HCl Suprapur® (Merck, Darmstadt, Germany).
Sample preparation: Prior to sample preparation all protein stocks were dialyzed
overnight against 100 mM TRIS buffer, pH 7.5. Different buffers were used for LADH and
RNase T1: LADH was dialyzed against 10 mM TRIS, pH 7.5, whereas 100 mM NaAc/acetic
acid, pH 5.5, was used for RNase T1. The protein concentration in phosphorescence
measurements was about 5 M throughout. Small volumes (100 l) of PAMAM dendrimers
were dialyzed, using disposable dialyzer (Micro DispoDialyzer®, Spectrum Laboratories Inc.,
Rancho Dominguez, CA,) against the same buffer used for the protein. Dendrimers
concentration after dialysis was determined from absorbance to 280 nm, by comparison to
that of stock dendrimer solution.
Phosphorescence spectroscopy: For phosphorescence measurements in fluid solutions,
O2 removal was achieved by the alternative application of moderate vacuum and inlet of ultra
pure N2 (27). The samples were placed in specially designed T-shaped spectrosil quartz
cuvettes (4 mm ID round tubing in the optical section, Hellma, Mullheim/Baden, Germany)
and rocked very gently, for about 10 min, to achieve complete exchange. The cuvette was
connected to the N2/vacuum line by peek tubing (1/16”) and the sample was fully isolated
from the atmosphere by a septum (Hamilton 76003, Alltech, Lancashire, UK) plus O-ring seal
assembly (27). Based on the phosphorescence lifetime of the protein alcohol dehydrogenase
from horse liver, which exhibits one of the highest sensitivities to O2 quenching, this
procedure lowered the O2-level below 2 nM.
Phosphorescence spectra and decays were both measured with pulsed excitation ( ex =
288 nm) on a home made apparatus (27), modified to implement spectral measurements by

4
means of CCD camera. Pulsed excitation was provided by a frequency-doubled Nd/Yag-
pumped dye laser (Quanta Systems, Milan, Italy) with pulse duration of 5 ns and a typical
energy per pulse of 0.5 - 1 mJ. For spectra measurements the emission was collected at 90
from the excitation and dispersed by a 0.3 m focal length triple grating imaging spectrograph
(SpectraPro-2300i, Acton Research Corporation, Acton, MA) with a band pass ranging from
1.0 to 0.2 nm. The emission was monitored by a back-illuminated 1340x400 pixels CCD
camera (Princeton Instruments Spec-10:400B(XTE), Roper Scientific Inc., Trenton, NJ)
cooled to -60°C.
Phosphorescence decays were monitored by collecting the emission at 90° from
vertical excitation through a filter combination with a transmission window of 405-445 nm
(WG405, Lot-Oriel, Milan Italy; plus interference filter DT-Blau, Balzer, Milan, Italy). The
photomultiplier (EMI 9235QA, Middlesex, UK) was protected against fatigue from the strong
excitation/fluorescence pulse by a gating circuit that inverts the polarity of dynodes 1 and 3,
for up to 1.5 ms, after the laser pulse. The photocurrent was amplified by a current-to-voltage
converter (SR570, Stanford Research Systems, Stanford, CA) and digitized by a
computerscope system (ISC-16, RC Electronics, Santa Barbara, CA) capable of averaging
multiple sweeps. All phosphorescence decays were analyzed by a non-linear least squares
fitting algorithm (DAS6, Fluorescence decay analysis software, Horiba Jobin Yvon, Milan,
Italy).
Each spectral and lifetime determination was repeated at least three times.
Results and Discussion
The effect of G5-OH, G5 and G4.5 (100 M) on the phosphorescence spectrum and
lifetime of every protein (typically 5 M) was measured at 20 °C. The buffer is 100 mM
TRIS, pH 7.5 except for LADH (10 mM TRIS, pH 7.5) and RNase T1 (100 mM NaAc, pH
5.5), as τ0 of these proteins depend on the salt concentration (LADH) and on pH (RNase T1).
In no case the phosphorescence spectrum was affected by the dendrimer whereas the
phosphorescence lifetime of some proteins was significantly altered by it.
Dendrimer effects on the phosphorescence lifetime of model proteins: For each protein
table 1 reports the Trp residue responsible for the emission (24), its location relative to the
aqueous phase (rp is minimum distance from the aqueous interface), its intrinsic
phosphorescence lifetime, τ0, and the lifetime ratio, τG/τ0, representing the change in τ induced
by each dendrimer (100 M). In the case of ALD and LADH the phosphorescence decay both
prior and after the addition of the polymer is not exponential (evidence of conformational
heterogeneity) and τ represents the average lifetime τav = α1τ1+α2τ2 obtained by a two
component fit of the data. For convenience the proteins are listed in the order of increasing
burial of the probe within the globular structure (rp). For the most deeply buried Trp residues,
W48 of Az and W109 of AP, the results show that the lifetime ratio τG/τ0 is 1 with every
dendrimer, implying that the structure within the internal rigid cores (large τ0) of these
macromolecules is not affected by the polymers. Thus, the globular fold remains intact and
any perturbation of protein flexibility in the outer layer by transient or stable association
between protein and polymer does not propagate to the interior. The results obtained with the
other proteins do, however, indicate that more superficial protein sites are generally affected
by the polymers. In the case of LADH and RNase T1 every polymer causes a reduction of τ as
if, locally, the native fold became more flexible through its association with the dendrimer.
This effect is markedly larger about the most superficial W59 of RNase T1. The opposite
trend, an increase of τ by dendrimers, is observed with ALD and only by the charged
polymers G4.5 and G5. Here, the probe reports a tightening of the structure about
W295/W313, sites that, judging from the small τ0, are both rather flexible.

5
It should be pointed out that a decrease of τ may also be caused by quenching
reactions with the functional groups of the dendrimer itself or with impurities associated to
the dendrimer stock solution. However, the functional groups on the dendrimer surface, OH
for G5-OH, NH3+
for G5 and COO- for G4.5, are equivalent to the side chains of Ser, Lys and
Glu, which in proteins have been shown to be inert with respect to the phosphorescence
lifetime of Trp in direct contact with them (23, 24). As to quenching by trace impurities a non
linear dependence of the decay rate on the dendrimer concentration, with early saturation
plateaus (see below), is a strong indication that impurity quenching is not important under
these conditions.
The above perturbations of the intrinsic lifetime of buried Trp residues by dendrimers
provide direct evidence that these polymers may interact sufficiently strongly with protein
molecules to alter their conformation in peripheral regions and at the aqueous interface.
Although the number of proteins examined is too limited to establish any firm correlation
with either their chemical composition or 3D structure it appears that, beyond the above noted
correlation with rp, other factors such quaternary structure and net surface charge are not
crucial for their association with the polymers. Indeed, a significant perturbation is observed
with monomeric RNase T1 as well as with dimeric LADH and tetrameric ALD. Likewise, the
results with RNase T1 indicate that the net protein charge is not the dominant steering force of
the interaction as neutral G5-OH and oppositely charged polymers are all quite effective. For
LADH and ALD the largest effects are observed with positively charged G5 in spite the net
protein charge, at pH 7.5, is also positive (table1).
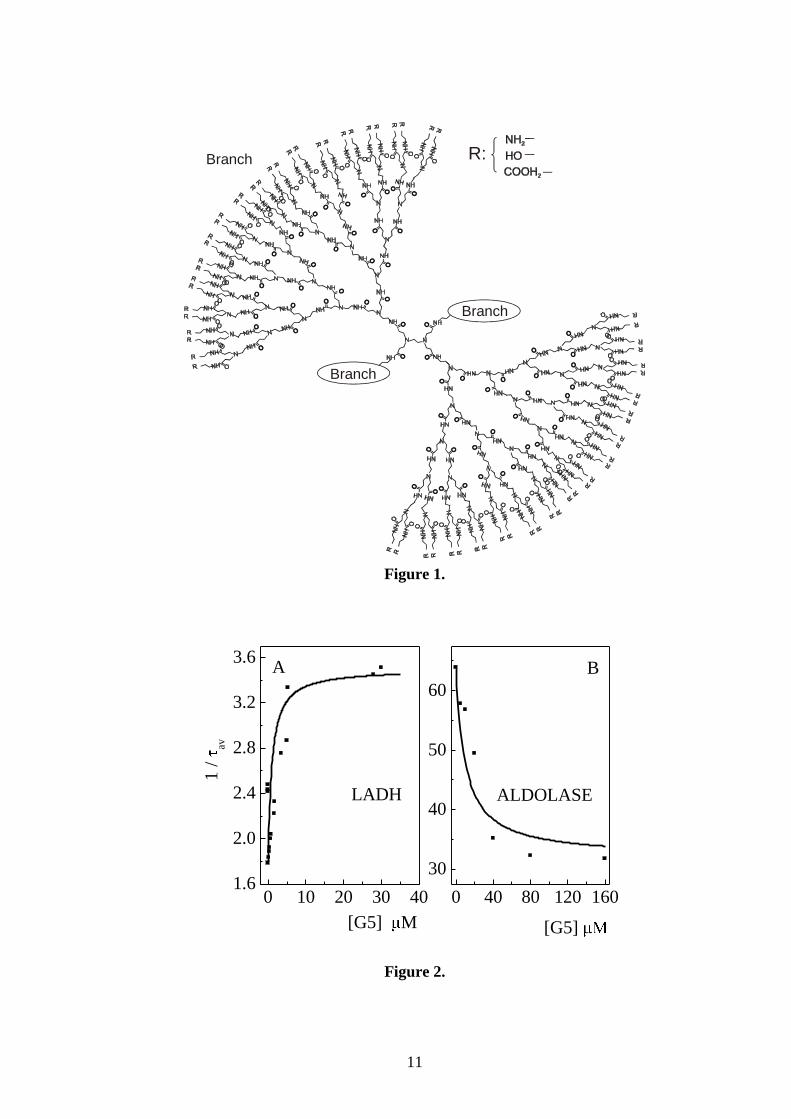
Effects of G5 concentration on of LADH and ALD: To shed light on the nature of
the interaction between proteins and dendrimers we studied the effect of increasing
concentration of G5 on τ of LADH and ALD. The systems LADH/G5 and ALD/G5 were
selected for the abundant protein supply and because G5 is among the three dendrimers that
causes the largest variation of τ, decreasing the lifetime of LADH and increasing that of ALD.
As indicated in figure 2, A and B, the addition of micro-molar quantities of G5 is sufficient to
produce the large changes of τ observed in these proteins. A plateau is reached just above 5
and 40 µM G5 for LADH and ALD, respectively. In both cases the 1/ vs. G5 concentration
profile is roughly hyperbolic, reminiscent of a binding curve. This finding suggests that
collisions between LADH/ALD and G5 change the protein conformation, presumably by
producing a bimolecular complex.
A simple scheme to account for the change of protein P upon transient complex
formation with a non-phosphorescent ligand L is as follows:
kon
P* + L ↔ P
*L
↓ koff ↓
kP kPL
where kP and kPL are the phosphorescence decay rate constant of free protein (P*) and of the
protein complex (P*L), respectively. Under rapid equilibrium conditions (kP, kPL < kon, koff)
the overall decay rate, 1/τ, is governed by
1/τ = fP kP + (1-fP) kPL
where fP is the fraction of free protein, which is related to the dissociation constant, KD =
koff/kon = fP[L]/(1- fP). Alternatively, when the equilibrium is slow the phosphorescence decay
is non exponential with lifetimes τ0 = 1/kP and τPL = 1/kPL and amplitudes equal to fP and (1-
fP), respectively. Because the decay of both proteins is throughout heterogeneous it is not
possible to distinguish rapid from slow equilibrium regimes and apply a rigorous analysis. In
either case, however, the average decay rate is governed by the dissociation constant and

6
therefore the profiles of figure 2 provide a rough estimate of KD. The best fit of lifetime data,
assuming rapid equilibrium conditions, yields dissociation constants of 1.5 μM for the
complex LADH/G5 and 10 μM for the complex ALD/G5, respectively. Theoretical curves
corresponding to these values of KD are drawn in figure 2. Compared to affinities of 10-6
-10-9
M observed for specific protein-protein complexes (28), these estimates suggest that the
affinity of G5 for LADH and ALD is comparable to or just weaker than the subunit affinity in
some multimeric proteins. These complexes are characterized by a dissociation free energy
ΔGD (ΔGD = -RTlnKD) in the range of 7 to 8 kcal mol-1
.
Another feature of these complexes is the relative small number of potential binding
sites. From the profiles of figure 2 we note that saturation is reached at a protein:G5 ratio of
about 1:1 for LADH and 1:4 for ALD. These correspond to one binding site per dimer for the
former and one binding site per subunit for the latter. Considering a Mw of 80,000 and
160,000 for the dimer and tetramer, respectively, it is clear that in either case the protein
surface could accommodate many more G5 molecules (Mw of 28,000). Hence, binding of G5
to these proteins, far from being limited by steric hindrance, appears to be directed to specific
regions of the protein surface.
Effect of ionic strength on G5 binding to LADH and ALD:. Electrostatic forces are
expected to play a preeminent role in the interaction between charged macromolecules, a
contribution that becomes strongly attenuated at high ionic strength (µ). Both ALD (pI = 8.4-
9.15) and LADH (pI = 8.7) are positively charged at pH 7.5, and the net charge calculated
from the amino acid sequence is +12 for ALD and +16 for LADH. Because the G5 surface is
also positively charged, having the primary amino groups of PAMAM dendrimers a pKa = 9
(29), we may expect that by increasing µ the dendrimer would form even stronger complexes
with the above proteins. To assess the importance of electrostatic “binding” in the protein/G5
interaction, the phosphorescence lifetime vs. G5 concentration profiles were monitored also at
higher ionic strength, through the addition of NaCl.
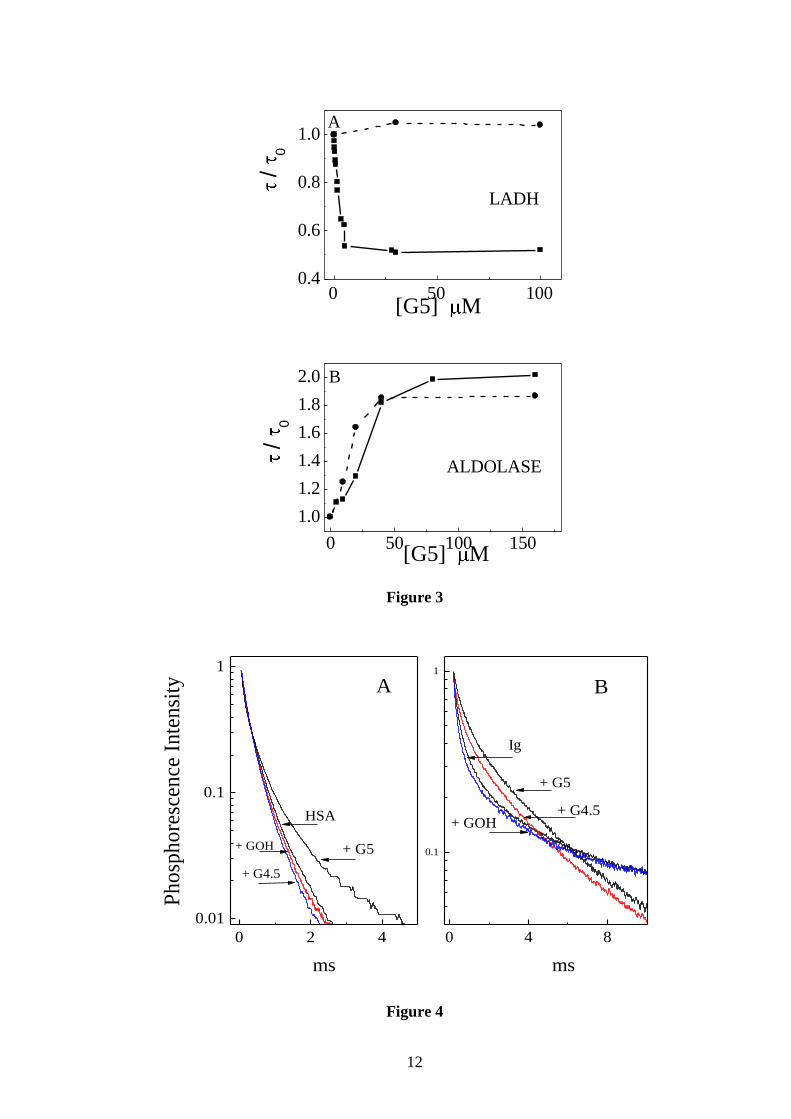
We first point out that the intrinsic lifetime of both LADH and ALD vary with ionic
strength, increasing slightly in the case of ALD, decreasing for LADH. For this reason τ0
refers to the intrinsic lifetime of the protein at the value of µ under examination. Figure 3A
compares the τG5/τ0 profiles of LADH at values of equal to 0.004 and 0.1 M. Rather
surprising, in the presence of 100 mM salt the dendrimer effect on the lifetime is totally
abolished. The response to increased µ is opposite to that expected from the net protein
charge. As pointed out above both protein and dendrimer are positively charged, so that
increasing µ would reduce the charge repulsion between macromolecules and favor their
association. Evidently, the sharp effect of µ implies that G5 binds to LADH predominantly
through an electrostatic attraction and that must involve a specific, negatively charged patch
of the protein surface.
The effect of µ is totally different with ALD where the change in lifetime induced by
G5 is maintained up to 0.5 M NaCl. In this case the main effect of the salt is towards
increasing the affinity of the complex (figure 3B), presumably a result of diminished charge
repulsion between like charged macromolecules. The invariance to ionic strength of the
ALD/G5 complex points out that forces other than electrostatic attraction (H-bonding, van der
Waals and hydrophobic) are responsible for the association of G5 to the ALD. This is a
significant, if surprising, result for it demonstrates that charged dendrimers can make relative
strong associations to proteins also through non ionic interactions, namely without the
formation of salt bridges at the polymer-protein interface. Because burial of charged groups at
the protein-dendrimer interface is costly in free energy terms, it probably requires that the
ionization state of the functional groups in the contact region changes to neutrality. After all
the pKa of the amino group in G5 is considerably lower than that of an isolated primary amine
(~ 9 vs. 12-13) and, further, only a few terminal groups of the dendrimer need to be involved
in the complex. Energetic considerations (30) conducted on protein-protein associations

7
indicate that to make a stable complex, each molecule involved needs to bury a surface area of
about 600 Å2. Considering that on G5 the surface area available per terminal group is
estimated to be 276 Å2 (31) 2-3 functional groups comprise an area comparable in size to
those found in aspecific protein-protein associations (32).
Interaction of dendrimers with plasma proteins HSA and -globulins: HSA has a sole
Trp residue (W214) buried within a deep cleft of the globular structure with the edge of the
benzene ring exposed to the solvent (33). Its phosphorescence emission is characterized by a
short-lived non exponential decay, the lifetime heterogeneity reflecting the presence of
multiple Trp environments/protein conformations in solution. Fig 4A shows the effect of each
dendrimer (100 µM) on the decay of HSA in 100 mM Tris, pH 7.5. Significant changes in the
decay rate are found in the presence of G5, whose effect is to widen the distribution to
shorter and longer-lived components, relative to HSA alone or in the presence of G5-OH and
G4.5. Lifetimes of sub milliseconds or shorter are due to quenching reactions with
neighboring Cys, His or Tyr side-chains (23). The crystal structure of HSA shows that H242
can come in direct contact with the indole ring whereas C200 is about 11 Ǻ from it, just
outside of its interaction range (33). Evidently, binding of G5 to HSA induces subtle
alterations in the protein conformation bringing the quenching side-chains either closer or
farther to the indole ring.
The human Ig has 11 Trp residues distributed among light and heavy chains and, further, no
assignment has been made yet of the room temperature phosphorescence emission to
individual residues. As shown in figure 4B the phosphorescence decay of Ig in 100 mM Tris,
pH 7.5, is highly heterogeneous with lifetime components ranging from sub milliseconds to
40 ms. Whereas neutral G5-OH does not affect the phosphorescence decay, positively and
negatively charged dendrimers alter considerably the decay profile and in a surprisingly
similar manner. As a result the lifetime of short-lived components increased 3-4 folds while
the long components have been shortened. In either case the change of τ is a clear
manifestation that the structure about some Trp residues has been altered, presumably
increasing the separation from quenching side-chains as well by enhancing the flexibility of
structured regions harboring the residues exhibiting the longer phosphorescence lifetimes.
Conclusions
The sensitivity of Trp phosphorescence to perturbations of the native fold has pointed out that
dendrimers tend to interact with proteins in solution forming complexes in which the protein
structure may be significantly altered. The results obtained with model proteins show that
structural changes are probably confined to superficial, flexible regions of the polypeptide.
Both electrostatic and non electrostatic interactions can give rise to stable complexes, whose
affinity and limited number of binding sites distinguish them from mere, aspecific molecular
associations. Of direct relevance for the application of these polymers in the medical field,
structural alterations have also been detected in two plasma proteins. The above results
suggest that Trp phosphorescence may also provide a useful monitor for working out
experimental conditions and protocols that help preserve the structural integrity of proteins in
the presence of these polymers.
Acknowledgement
Dr. Dzmitry Shcharbin is a beneficiary of a Marie Curie International Incoming Fellowship
within the 6th EU Framework Programme (grant 510018).

8
REFERENCES
(1) Fischer, M., and Vogtle, F. (1999) Dendrimers: From design to application - A
progress report. Angewandte Chemie-International Edition 38, 885-905.
(2) Frechet, J. M. J. (1994) Functional Polymers and Dendrimers - Reactivity, Molecular
Architecture, and Interfacial Energy. Science 263, 1710-1715.
(3) Hawker, C. J., and Frechet, J. M. J. (1992) Unusual Macromolecular Architectures -
the Convergent Growth Approach to Dendritic Polyesters and Novel Block
Copolymers. Journal of the American Chemical Society 114, 8405-8413.
(4) Tomalia, D. A., Naylor, A. M., and Goddard, W. A. (1990) Starburst Dendrimers -
Molecular-Level Control of Size, Shape, Surface-Chemistry, Topology, and
Flexibility from Atoms to Macroscopic Matter. Angewandte Chemie-International
Edition in English 29, 138-175.
(5) Cloninger, M. J. (2002) Biological applications of dendrimers. Current Opinion in
Chemical Biology 6, 742-748.
(6) Matthews, O. A., Shipway, A. N., and Stoddart, J. F. (1998) Dendrimers - Branching
out from curiosities into new technologies. Progress in Polymer Science 23, 1-56.
(7) Aulenta, F., Hayes, W., and Rannard, S. (2003) Dendrimers: a new class of
nanoscopic containers and delivery devices. European Polymer Journal 39, 1741-
1771.
(8) Esfand, R., and Tomalia, D. A. (2001) Poly(amidoamine) (PAMAM) dendrimers:
from biomimicry to drug delivery and biomedical applications. Drug Discovery Today
6, 427-436.
(9) Diederich, F., and Felber, B. (2002) Supramolecular chemistry of dendrimers with
functional cores. Proceedings of the National Academy of Sciences of the United
States of America 99, 4778-4781.
(10) Ozaki, S., DeWald, D. B., Shope, J. C., Chen, J., and Prestwich, G. D. (2000)
Intracellular delivery of phosphoinositides and inositol phosphates using polyamine
carriers. Proceedings of the National Academy of Sciences of the United States of
America 97, 11286-11291.
(11) Klajnert, B., Sadowska, M., and Bryszewska, M. (2004) The effect of
polyamidoamine dendrimers on human erythrocyte membrane acetylcholine sterase
activity. Bioelectrochemistry 65, 23-26.
(12) Klajnert, B., Stanislawska, L., Bryszewska, M., and Palecz, B. (2003) Interactions
between PAMAM dendrimers and bovine serum albumin. Biochimica Et Biophysica
Acta-Proteins and Proteomics 1648, 115-126.
(13) Shcharbin, D., Jokiel, M., Klajnert, B., and Bryszewska, M. (2006) Effect of
dendrimers on pure acetylcholinesterase activity and structure. Bioelectrochemistry
68, 56-59.
(14) Shcharbin, D., Klajnert, B., and Bryszewska, M. (2005) The effect of PAMAM
dendrimers on human and bovine serum albumin at different pH and NaCl
concentrations. Journal of Biomaterials Science-Polymer Edition 16, 1081-1093.
(15) Shcharbin, D., Klajnert, B., Mazhul, V., and Bryszewska, M. (2005) Dendrimer
interactions with hydrophobic fluorescent probes and human serum albumin. Journal
of Fluorescence 15, 21-28.
(16) Cioni, P., Piras, L., and Strambini, G. B. (1989) Tryptophan phosphorescence as a
monitor of the structural role of metal ions in alkaline phosphatase. Eur J Biochem
185, 573-9.
(17) Strambini, G. B., and Gonnelli, M. (1990) Tryptophan luminescence from liver
alcohol dehydrogenase in its complexes with coenzyme. A comparative study of
protein conformation in solution. Biochemistry 29, 196-203.

9
(18) Strambini, G. B., Cioni, P., and Puntoni, A. (1989) Relationship between the
conformation of glutamate dehydrogenase, the state of association of its subunit, and
catalytic function. Biochemistry 28, 3808-14.
(19) Gabellieri, E., and Strambini, G. B. (1994) Conformational changes in proteins
induced by dynamic associations. A tryptophan phosphorescence study. Eur J
Biochem 221, 77-85.
(20) Cioni, P., and Strambini, G. B. (2002) Tryptophan phosphorescence and pressure
effects on protein structure. Biochimica Et Biophysica Acta-Protein Structure and
Molecular Enzymology 1595, 116-130.
(21) Gabellieri, E., and Strambini, G. B. (2001) Structural perturbations of azurin deposited
on solid matrices as revealed by trp phosphorescence. Biophys J 80, 2431-8.
(22) Strambini, G. B., and Gabellieri, E. (1996) Proteins in frozen solutions: Evidence of
ice-induced partial unfolding. Biophysical Journal 70, 971-976.
(23) Gonnelli, M., and Strambini, G. B. (2005) Intramolecular quenching of tryptophan
phosphorescence in short peptides and proteins. Photochemistry and Photobiology 81,
614-622.
(24) Gonnelli, M., and Strambini, G. B. (1995) Phosphorescence Lifetime of Tryptophan in
Proteins. Biochemistry 34, 13847-13857.
(25) Chen, R. F. (1967) Removal of Fatty Acids from Serum Albumin by Charcoal
Treatment. J. Biol. Chem. 242, 173-181.
(26) van de Kamp, M., Silvestrini, M. C., Brunori, M., van Beeumen, J., Hali, F. C., and
Canters, G. W. (1990) Involvement of the Hydrophobic Patch of Azurin in the
Electron-Transfer Reactions with Cytochrome-C551 and Nitrite Reductase. European
Journal of Biochemistry 194, 109-118.
(27) Strambini, G. B., Kerwin, B. A., Mason, B. D., and Gonnelli, M. (2004) The triplet-
state lifetime of indole derivatives in aqueous solution. Photochemistry and
Photobiology 80, 462-470.
(28) Nooren, I. M. A., and Thornton, J. M. (2003) Structural characterisation and
functional significance of transient protein-protein interactions. Journal of Molecular
Biology 325, 991-1018.
(29) Cakara, D., Kleimann, J., and Borkovec, M. (2003) Microscopic protonation equilibria
of poly(amidoamine) dendrimers from macroscopic titrations. Macromolecules 36,
4201-4207.
(30) Cyrus Chothia, J. J. (1975) Principles of protein-protein recognition. Nature 256, 705-
708.
(31) Meltzer, A. D., Tirrell, D. A., Jones, A. A., Inglefield, P. T., Hedstrand, D. M., and
Tomalia, D. A. (1992) Chain Dynamics in Poly(Amido Amine) Dendrimers - a Study
of C-13 Nmr Relaxation Parameters. Macromolecules 25, 4541-4548.
(32) Bahadur, R. P., Chakrabarti, P., Rodier, F., and Janin, J. (2004) A dissection of
specific and non-specific protein - Protein interfaces. Journal of Molecular Biology
336, 943-955.
(33) Bhattacharya, A. A., Curry, S., and Franks, N. P. (2000) Binding of the general
anesthetics propofol and halothane to human serum albumin - High resolution crystal
structures. Journal of Biological Chemistry 275, 38731-38738.
10
Table 1: Change in phosphorescence lifetimes ( ) of proteins after addition of 100 M G5-
OH ( G5-OH), G4.5 ( G4.5) or G5 ( G5) at 20°C.
Protein Buffer, pH Net
charge‡)
# Trp rp (Å) 0 (ms) G5-
OH/ 0 G4.5/ 0 G5/ 0
RNase T1 0.1 M NaAc, 5.5 W59 1.5-2 25.7 2 0.26 0.23 0.42
ALD 0.1 M TRIS, 7.5 +12 W313/
W295
3-4 16*)
1 0.94 1.25 1.94
LADH 0.01 M TRIS, 7.5 + 16 W314 4.5 550*)
30 0.91 .84 0.53
Az 0.1 M TRIS, 7.5 - 5 W48 8 590 30 1 0.96 0.96
AP 0.1 M TRIS, 7.5 - 18 W109 11 1870 60 1 0.96 0.97
*)
Average lifetime: = 1 1+ 2 2 ‡)
Calculated from the aminoacid composition.
FIGURE LEGENDS
Figure 1: Structure of the fifth generation PAMAM dendrimers: A) G5; B) G5-OH; C) G4.5.
Figure 2: Effect of G5 concentration on the average phosphorescence lifetime of LADH and
ALD, at 20°C. A) 4.2 μM LADH in 10 mM TRIS pH 7.5; B) 5μM ALD in 100 mM TRIS pH
7.5. Continuous lines represent the best fit of experimental data to the theoretical binding
curve (see text): 1/τ = 1/τ0 +([G5]/(KD+[G5])*(1/τPL-1/τ0)). The reproducibility of τ
determinations is 7 % or better.
Figure 3: Comparative effect of high ionic strength on the phosphorescence lifetime vs. G5
concentration profiles of LADH and ALD. A) 4.2 μM LADH in (■) 10 mM TRIS pH 7.5 and
in (●) 10 mM TRIS plus 100 mM NaCl pH 7.5. B) 5 μM ALD in (■) in 100 mM TRIS, pH
7.5, and in (●) 100 mM TRIS plus 500 mM NaCl, pH 7.5. The temperature is 20°C.
Figure 4: Representative phosphorescence decays of A) 5 μM HSA and B) human Ig in 100
mM TRIS, pH 7.5, before and after the addition of 100 μM PAMAM dendrimers (G5-OH,
G4.5 and G5), at 20 °C.
11
Branch
Branch
Branch
R:
Figure 1.
0 10 20 30 401.6
2.0
2.4
2.8
3.2
3.6
0 40 80 120 160
30
40
50
60
1 /
av
[G5] M
A
LADH
[G5]
B
ALDOLASE
Figure 2.
12
0 50 1000.4
0.6
0.8
1.0
0 50 100 150
1.0
1.2
1.4
1.6
1.8
2.0
[G5] M
[G5] M
B
A
LADH
/ 0
/ 0
ALDOLASE
Figure 3
0 4 8
0.1
1
0 2 4
0.01
0.1
1
ms
B
Ig
+ G5
+ G4.5+ GOH
Ph
osp
ho
resc
ence
In
ten
sity
ms
A
+ G5+ GOH
+ G4.5
HSA
Figure 4
Related Documents











