DEVELOPMENTAL BIOLOGY 185, 92–103 (1997) ARTICLE NO. DB978546 Delta-1 Is a Regulator of Neurogenesis in the Vertebrate Retina Iqbal Ahmad, Constance M. Dooley, and Dorisa L. Polk Department of Cell Biology and Anatomy, and Department of Ophthalmology, University of Nebraska Medical Center, Omaha, Nebraska 68198-6395 In the retina, cell fate determination is thought to be regulated by a series of local cell – cell interactions. Evidence suggests that retinal precursors utilize Notch-mediated intercellular signaling to regulate their fates. However, the identity of the endogenous ligand and its role in the Notch-signaling pathway is not well understood. We have identified C-Delta-1 as the putative endogenous ligand for Notch, in the developing chick retina. C-Delta-1 is coexpressed spatially and temporally with C-Notch-1 and their expression is associated with the temporal aspects of cell birth in the developing retina. This suggests that Delta – Notch signaling is utilized to maintain progenitors in an uncommitted state and that a subtle fluctua- tion in this signaling helps to sort out competent cells during successive cell-fate determination. We have tested the latter possibility in the specification of the ganglion cells. In early stages of retinal development when ganglion cells are the predominant cells born, decreasing C-Delta-1 expression with antisense oligonucleotides increases the proportion of RA4 antigen-expressing ganglion cells which are recruited predominantly in the periphery. Conversely, use of exogenous Dro- sophila Delta leads to a decrease in the RA4 antigen-expressing ganglion cells. Our results suggest that C-Delta-1 activation of the Notch pathway regulates the specification of retinal neurons in general and of ganglion cells in particular. q 1997 Academic Press precursor to follow a particular path depends on local cell INTRODUCTION interactions. The molecular and cellular mechanisms underlying cell- In the vertebrate retina neurons develop from neuroepi- fate regulation by cell interactions are not well understood. thelial cells lining the inner wall of the developing optic However, recent evidence suggests that most precursors, vesicle (reviewed in Robinson, 1991). Thymidine birthdat- both neural and nonneural, utilize a very general and evolu- ing analyses in several species have shown that these pro- tionarily conserved intercellular signaling mechanism, the genitors give rise to retinal neurons in a temporal order Notch pathway, to regulate their fates (reviewed in that is evolutionarily conserved (Sidman, 1961; Kahn, 1974; Greenwald and Rubin, 1992; Muskavitch, 1994; Artavanis- Young, 1985; Prada et al., 1991; LaVail et al., 1991). Retinal Tsakonas et al., 1991, 1995). In this pathway, Notch acts ganglion cells (RGC) are the first neurons which are born as a cellular receptor that participates in cell-fate specifica- and the bipolar cells are usually one of the last retinal cell tion by regulating the competence of a differentiating cell types to become postmitotic. In addition to this temporal to respond to epigenetic cues. The ligands for Notch belong pattern, cell fate specification in retina follows a stereotypi- to a family of membrane-anchored proteins that contain cal spatial pattern; retinal neurons are born first in the cen- EGF repeats and a cysteine-rich DSL (Delta – Serrate – Lag2) tral retina and last in the periphery with the gradient of cell domain in the extracellular region (reviewed in Kopan and differentiation spreading from the center to the periphery Nye, 1996). These putative ligands include Delta and Ser- (Harman and Beazley, 1989; Robinson, 1991; Prada et al., rate in Drosophila (Vaessin et al., 1987; Kopczynski et al., 1991; Lavail et al., 1991). Evidence from a variety of experi- 1988; Fleming et al., 1990; Thomas et al., 1991), Apx-1 and mental approaches including cell ablation experiments (Reh Lag-2 in Caenorhabditis elegans (Tax et al., 1994; Hender- and Tully, 1986)) and lineage analyses (Turner and Cepko, son et al., 1994; Gao and Kimble, 1995; Fitzgerald and 1987; Price et al., 1987; Holt et al., 1988; Wetts and Fraser, Greenwald, 1995) and Delta-1, Serrate-1, and Jagged in ver- 1988; Turner et al., 1990) suggests that the retinal precur- tebrates (Chitnis et al., 1995; Henrique et al., 1995; Lindsell et al., 1995; Myat et al., 1996). Evidence in Drosophila sug- sors are multipotential and that the decision taken by a 92 0012-1606/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved. AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DEVELOPMENTAL BIOLOGY 185, 92–103 (1997)ARTICLE NO. DB978546
Delta-1 Is a Regulator of Neurogenesisin the Vertebrate Retina
Iqbal Ahmad, Constance M. Dooley, and Dorisa L. PolkDepartment of Cell Biology and Anatomy, and Department of Ophthalmology,University of Nebraska Medical Center, Omaha, Nebraska 68198-6395
In the retina, cell fate determination is thought to be regulated by a series of local cell–cell interactions. Evidence suggeststhat retinal precursors utilize Notch-mediated intercellular signaling to regulate their fates. However, the identity of theendogenous ligand and its role in the Notch-signaling pathway is not well understood. We have identified C-Delta-1 asthe putative endogenous ligand for Notch, in the developing chick retina. C-Delta-1 is coexpressed spatially and temporallywith C-Notch-1 and their expression is associated with the temporal aspects of cell birth in the developing retina. Thissuggests that Delta–Notch signaling is utilized to maintain progenitors in an uncommitted state and that a subtle fluctua-tion in this signaling helps to sort out competent cells during successive cell-fate determination. We have tested the latterpossibility in the specification of the ganglion cells. In early stages of retinal development when ganglion cells are thepredominant cells born, decreasing C-Delta-1 expression with antisense oligonucleotides increases the proportion of RA4antigen-expressing ganglion cells which are recruited predominantly in the periphery. Conversely, use of exogenous Dro-sophila Delta leads to a decrease in the RA4 antigen-expressing ganglion cells. Our results suggest that C-Delta-1 activationof the Notch pathway regulates the specification of retinal neurons in general and of ganglion cells in particular.q 1997 Academic Press
precursor to follow a particular path depends on local cellINTRODUCTIONinteractions.
The molecular and cellular mechanisms underlying cell-In the vertebrate retina neurons develop from neuroepi- fate regulation by cell interactions are not well understood.
thelial cells lining the inner wall of the developing optic However, recent evidence suggests that most precursors,vesicle (reviewed in Robinson, 1991). Thymidine birthdat- both neural and nonneural, utilize a very general and evolu-ing analyses in several species have shown that these pro- tionarily conserved intercellular signaling mechanism, thegenitors give rise to retinal neurons in a temporal order Notch pathway, to regulate their fates (reviewed inthat is evolutionarily conserved (Sidman, 1961; Kahn, 1974; Greenwald and Rubin, 1992; Muskavitch, 1994; Artavanis-Young, 1985; Prada et al., 1991; LaVail et al., 1991). Retinal Tsakonas et al., 1991, 1995). In this pathway, Notch actsganglion cells (RGC) are the first neurons which are born as a cellular receptor that participates in cell-fate specifica-and the bipolar cells are usually one of the last retinal cell tion by regulating the competence of a differentiating celltypes to become postmitotic. In addition to this temporal to respond to epigenetic cues. The ligands for Notch belongpattern, cell fate specification in retina follows a stereotypi- to a family of membrane-anchored proteins that containcal spatial pattern; retinal neurons are born first in the cen- EGF repeats and a cysteine-rich DSL (Delta–Serrate–Lag2)tral retina and last in the periphery with the gradient of cell domain in the extracellular region (reviewed in Kopan anddifferentiation spreading from the center to the periphery Nye, 1996). These putative ligands include Delta and Ser-(Harman and Beazley, 1989; Robinson, 1991; Prada et al., rate in Drosophila (Vaessin et al., 1987; Kopczynski et al.,1991; Lavail et al., 1991). Evidence from a variety of experi- 1988; Fleming et al., 1990; Thomas et al., 1991), Apx-1 andmental approaches including cell ablation experiments (Reh Lag-2 in Caenorhabditis elegans (Tax et al., 1994; Hender-and Tully, 1986)) and lineage analyses (Turner and Cepko, son et al., 1994; Gao and Kimble, 1995; Fitzgerald and1987; Price et al., 1987; Holt et al., 1988; Wetts and Fraser, Greenwald, 1995) and Delta-1, Serrate-1, and Jagged in ver-1988; Turner et al., 1990) suggests that the retinal precur- tebrates (Chitnis et al., 1995; Henrique et al., 1995; Lindsell
et al., 1995; Myat et al., 1996). Evidence in Drosophila sug-sors are multipotential and that the decision taken by a
92
0012-1606/97 $25.00Copyright q 1997 by Academic Press
All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

93Delta–Notch Signaling in the Retina
gests that the binding of ligands activates the Notch recep- (2) In developing chick hindbrain and spinal cord C-Serrate-1 may perform the same function as C-Delta-1 in comple-tor. For example, null mutation of Delta results in the neu-
rogenic loss of function phenotype that is also observed in mentary domains during neurogenesis (Myat et al., 1996).In this study we have identified and analyzed the role ofa loss of function mutation of Notch (Heitzler and Simpson,
1991; Greenwald and Rubin, 1992). Serrate, which is closely the putative endogenous ligand that activates Notch signal-ing during retinal neurogenesis. We show that C-Delta-1related to Delta structurally and in its interaction with
Notch (Rebay et al., 1991; Muskavitch, 1994), is able to appears to be the dominant endogenous ligand for the Notchpathway in chick retina. Both C-Delta-1 and C-Notch-1rescue the neurogenic phenotype in Delta mutant embryos
(Gu et al., 1995). Similar observations that Notch signaling genes are coexpressed spatially and temporally and the lev-els of their transcripts correspond to stages of cell-fate speci-is facilitated by the activation of Notch receptor by ligands
have been reported in vertebrates. For example, the suppres- fication, suggesting the involvement of Delta–Notch sig-naling in successive cell-fate determination as well as thesion of neurogenesis by constitutively activated Notch is
mimicked by the overexpression of Delta-1 in Xenopus spatial specification of retinal neurons. We analyzed theeffect of a decrease in Delta-1 expression on the distribution(Chitnis et al., 1995) and Jagged, the rat homolog of Serrate,
suppresses the differentiation of cultured myoblasts ex- of RA4 antigen, an early marker for RGC differentiation(McLoon and Barnes, 1989; Waid and McLoon, 1995). In thepressing Notch-1 (Lindsell et al., 1995).
Recent observations indicate that, like in Drosophila (Ca- developing retina, RA4 immunoreactivity showed a distinctcentral to peripheral gradient and was largely absent fromgan and Ready, 1989; Fortini et al., 1993), the Notch path-
way plays a significant role in the specification of retinal the periphery at Embryonic Day (E) 4. A decrease in thelevels of C-Delta-1 transcripts in E4 retina in response toneurons in vertebrates. Analyses of the pattern of Notch-1
expression in rat retina have suggested that Notch signaling antisense treatment increased the number of RA4-positiveRGCs which appear to be recruited more in the peripheryregulates temporal as well as spatial aspects of retinal neu-
rogenesis (Ahmad et al., 1995). It is thought that the Notch than in the center of the retina. A similar increase in thenumber of RA4-positive RGCs was also observed when thepathway maintains a population of immature cells in an
uncommitted state until proper cues for differentiation be- expression of C-Notch-1 was decreased. Additionally,exposing developing retinal cells to extraneous Delta in ex-come available. For example, expression of constitutively
active Notch-1 in Xenopus retina prevents immature cells plant cultures prevented the recruitment of the RA4-posi-tive cells to the peripheral retina. These results suggest thatfrom differentiating (Dorsky et al., 1995). Conversely, a de-
crease in Notch-1 signaling promotes differentiation in the C-Delta-1 activation of the Notch pathway is required forthe temporal and spatial specification of the retinal neurons.retina (Austin et al., 1995). While these observations
strongly suggest that Notch signaling plays a significantrole in the specification of retinal neurons, the endogenousligand(s) involved in regulating the signaling have not been MATERIALS AND METHODSidentified. In Drosophila, Delta and Serrate participate inNotch signaling to facilitate different developmental deci- Embryos. Fertilized hens’ eggs were incubated in a humidifiedsions. While Delta–Notch signaling mediates lateral inhibi- chamber at 387C. Embryos were staged according to Hamburger
and Hamilton (1951).tion that leads to the segregation of neuroblasts (Musko-RT–PCR. DNase-digested total RNA (5 mg) from each develop-vitch, 1994; Artavanis-Tsakonas et al., 1995), Serrate–
mental stage was used to synthesize cDNA, using a random hex-Notch signaling participates in the organization of the wingamer as a primer (Grillo and Margolis, 1991; Ahmad, 1995). One-imaginal disc (Speicher et al., 1994; Diaz-Benjumea and Co-tenth (5 ml) of the cDNA reaction/developmental stage washen, 1995; Couso et al., 1995). However, in later develop-amplified using either Delta-1 specific (5*-CACCAGCCCGAG-mental processes both Delta and Serrate show overlappingGCCTGC-3*; 5*-GATGCACTCATCTTTCTC-3*) (Henrique et al.,
but distinct expression patterns, suggesting that develop- 1995) or Notch-1 specific (Ahmad et al., 1995; Myat et al., 1996)mental decisions in the same tissue may utilize either Ser- primers for 25-step cycles (947C, 1 min; 487C, 1 min; 727C, 1.5 min).rate or Delta, and in some cases both ligands (Gu et al., The amplification after 25-step cycles was linear as ascertained by1995). In retina, where Notch-1 activity is correlated with amplification of serially diluted cDNA. PCR products were re-
solved on 1.5% agarose gel and Southern analyses were carried outsuccessive cell-fate decisions (Ahmad et al., 1995) the possi-using C-Delta-1 or C-Notch-1 cDNA as probes (Henrique et al.,bility exists that the Notch pathway utilizes either one or1995; Myat et al., 1996). A 548-bp b-actin cDNA was amplifieddifferent ligands for the specification of different cell types.using 5 ml of cDNA/developmental stage as a measure of the consti-The observation that in a coculture study Drosophila Deltatutive expression and RNA integrity (Ahmad et al., 1995). To ascer-was able to suppress RGC specification suggests that Dro-tain the effect of antisense oligonucleotides corresponding to C-sophila Delta mimics a vertebrate ligand that activatesDelta-1 and C-Notch-1 transcripts cDNA was synthesized from
Notch during retinal development (Austin et al., 1995). equal amounts of total RNA isolated from explant culture. BecauseThat vertebrate ligand (s) could be homologs of either Delta of the small amount of total RNA extracted (approximately 5 mg)or Serrate based on the following observations: (1) In Dro- the possible variation between the samples was further controlledsophila Serrate can substitute for Delta in the activation of by monitoring the amount of cDNA synthesized using 32P-labeled
dCTP in an aliquot of cDNA reaction. RT–PCR was carried outthe Notch pathway during neurogenesis (Gu et al., 1995).
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

94 Ahmad, Dooley, and Polk
as described and stage-specific normalization of the Delta-1- and Drosophila S2 cell culture. Drosophila Delta cDNA-trans-fected and untransfected S2 cells were cultured as previously de-Notch-1-specific transcript was carried out using b-actin-amplifiedscribed (Fehon et al., 1990). To determine the effects of exogenousproduct.Delta on retinal development the lens was removed from E3.5 eyeIn situ hybridization. Probes labeled with 35S were transcribedcups giving S2 cells access to the vitreal surface of the retina. Theusing C-Delta-1 or C-Notch-1 cDNA as template. In situ hybridiza-eye cup was filled with excess S2 cells and incubated in culturetion was carried out on fresh frozen retinal sections (12 mM) asmedium containing additional S2 cells for 24 hr. After culturing,previously described (Ahmad et al., 1994, 1995).the eye cups were fixed in STF fixative for 10 min, cryoprotectedNorthern analyses. Northern analyses were performed on 20overnight in 30% sucrose, sectioned, and analyzed for RA4 immu-mg of total RNA isolated from E3.5 retina as previously describednocytochemistry.(Ahmad, 1995) using 32P-labeled C-Delta-1 or C-Notch-1 cDNA
Immunocytochemistry. Immunocytochemistry was carriedprobes. The final wash was carried out at 657C for 1 hr in 11 SSCout as previously described (McLoon and Barnes, 1989; Ahmadand 0.1% SDS.et al., 1995). Briefly, whole sections or dissociated cells wereExplant culture and antisense treatment. Retinae were har-incubated in PBS containing 5% normal goat serum and 0.2%vested from stage-specific embryos in Hanks’ balanced salt solutionTriton X-100 followed by an overnight incubation in RA4 anti-(HBSS). The explants were cultured in 96-well plates in DMEM:F12body (1:1000) or Notch-1 antibody (20F) (1:100) at 47C. The sec-medium containing 11 N2 supplement (Gibco), 1% fetal bovinetions were examined for epifluorescence using Leica DMR micro-serum (FBS), 2 mM L-glutamine, 100 U/ml penicillin, and 100 mg/scope following incubation in anti-mouse (RA4) or anti-ratml streptomycin at 377C in 95% humidity and 5% CO2. Following(Notch-1) IgG conjugated to CY3.24 hr of incubation with and without antisense, sense or missense
oligonucleotides (50 mM), the explants were fixed in Streck TissueFixative (Streck Laboratories) and cryoprotected in 30% sucrose forimmunocytochemistry. For dissociation into single cell suspension RESULTSexplants were incubated in HBSS (Ca2/ and Mg2/ free) containing0.25% trypsin, 1 mM EDTA, and 20 mg/ml DNase1 at 377C for C-Delta-1 and C-Notch-1 Transcripts Are20 min. Trypsin was neutralized by washing the tissue in HBSS Coexpressed Temporallycontaining 20% FBS. Cells were dissociated by trituration (10–15times) in the culture medium and plated at a density of 103–104 The identification of C-Delta-1 and subsequent analysescells/glass coverslip treated with 50–100 mg/ml of poly-D-lysine. of the temporal pattern of expression of C-Delta-1 and C-Cells were allowed to adhere to the coverslips for 2–3 hr at 377C Notch-1 in the developing retina were carried out by semi-before fixation in 4% paraformaldehyde. Each explant culture was quantitative RT-PCR analysis. cDNA was synthesized fromanalyzed for cell viability using the trypan blue dye-exclusion equal amounts of total RNA isolated from retina from vari-method. Experiments were performed at least three times and re-
ous developmental stages and used in PCR to amplify a 418-peated in triplicate. On average, 35 cells were counted in four differ-and 298-bp region in C-Delta-1 and C-Notch-1 transcripts,ent fields per treatment. The antisense, sense, and missense oligo-respectively. The specificity of amplification was furthernucleotides were synthesized at Yale University (Department ofconfirmed by Southern analyses using C-Delta-1 and C-Pathology) and University of Nebraska Medical Center (EppleyNotch-1 cDNA as probes. The earliest stage that was ana-Cancer Research Center) and purified by HPLC. Sequence were
as follows: for b-globin antisense, 5*-AGTCCAGTGCACCAT-3* lyzed for C-Delta-1 expression was E2.5, at which approxi-(Represa et al., 1991); Notch-1 antisense, 5*-CCTCCGCTGCAG- mately 15% of ganglion cells have been generated (Prada etGAGGCAATCAT-3* (Weinmaster et al., 1991; Austin et al., al., 1991). C-Delta-1 expression was detected at this stage1995); Notch-1 sense, 5*-ATGATTGCCTCCTGCAGCGCAGG-3*; and its levels increased until reaching a peak of expressionNotch-1 missense, 5*-CGTAGTGACTACAGAGCGCTCCC-3*; at E4 and E7.5 (Fig. 1A). This is the period when all theDelta-1 antisense, 5*-GACCTTCTCGCCACGCTC-3* (Henrique retinal neurons and Muller glia are in various stages of dif-et al., 1995); Delta-1 sense, 5*-GAGCGTGGCGAGAAGGTC-3*;
ferentiation (Fig. 1C). A decrease in the levels of C-Delta-1Delta-1 missense, 5*-TCGTCCACCGATCCGTCC-3*. For the eye-transcripts was observed beginning at E10. Since one of thecup culture, eyes were removed from E4 embryos, placed in HBSS,requisites for C-Delta-1 action is the presence of C-Notch-and hemisected removing the lens to ensure that the peripheral1 receptor we wanted to know if C-Delta-1 is coexpressedtips of the retina which remain devoid of RA4 immunoreactivitytemporally with C-Notch-1. Therefore, we analyzed the lev-(McLoon and Barnes, 1989) were not included in the culture. The
eye-cups were incubated for 24 hr in culture medium containing els of C-Notch-1 transcript in embryonic stages in which50 mM C-Delta-1 antisense oligonucleotide in a 96-well plate. The the expression of Delta-1 was measured. The levels of C-medium was changed once halfway through the culture. The eye Notch-1 transcripts showed a pattern of temporal expres-cups were fixed in STF fixative for 10 minutes, cryoprotected over- sion similar to that of C-Delta-1 transcripts with their high-night in 30% sucrose, sectioned, and analyzed for RA4 immunore- est levels detected at E4 and E7.5. (Fig. 1B). Therefore, theactivity. To distinguish the central from the peripheral retina we levels of C-Delta-1 expression appeared to be tightly corre-scored for retinal sections in which a labeled region was flanked
lated with that of C-Notch-1 expression and, together, theirby an unlabeled region for RA4 immunoreactivity. Because theexpression was associated with cell birth in the developingunlabeled tips were removed prior to culturing, any region devoidretina.of RA4 immunoreactivity represented true periphery. In two of
To corroborate that transcripts amplified by RT–PCRfour repeat experiments sections were obtained in both treated andwere the products of C-Delta-1 genes expressed in the devel-untreated groups in which the central and peripheral retina can be
distinguished by the above-mentioned criteria. oping retina, we carried out Northern analyses on E3.5 total
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
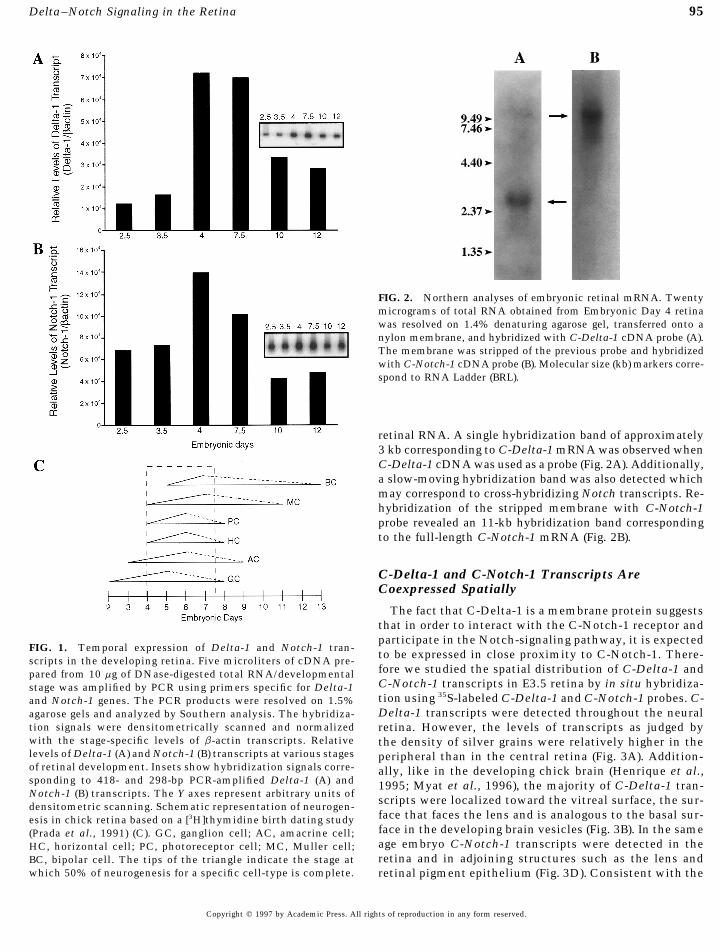
95Delta–Notch Signaling in the Retina
FIG. 2. Northern analyses of embryonic retinal mRNA. Twentymicrograms of total RNA obtained from Embryonic Day 4 retinawas resolved on 1.4% denaturing agarose gel, transferred onto anylon membrane, and hybridized with C-Delta-1 cDNA probe (A).The membrane was stripped of the previous probe and hybridizedwith C-Notch-1 cDNA probe (B). Molecular size (kb) markers corre-spond to RNA Ladder (BRL).
retinal RNA. A single hybridization band of approximately3 kb corresponding to C-Delta-1 mRNA was observed whenC-Delta-1 cDNA was used as a probe (Fig. 2A). Additionally,a slow-moving hybridization band was also detected whichmay correspond to cross-hybridizing Notch transcripts. Re-hybridization of the stripped membrane with C-Notch-1probe revealed an 11-kb hybridization band correspondingto the full-length C-Notch-1 mRNA (Fig. 2B).
C-Delta-1 and C-Notch-1 Transcripts AreCoexpressed Spatially
The fact that C-Delta-1 is a membrane protein suggeststhat in order to interact with the C-Notch-1 receptor andparticipate in the Notch-signaling pathway, it is expected
FIG. 1. Temporal expression of Delta-1 and Notch-1 tran-to be expressed in close proximity to C-Notch-1. There-scripts in the developing retina. Five microliters of cDNA pre-fore we studied the spatial distribution of C-Delta-1 andpared from 10 mg of DNase-digested total RNA/developmentalC-Notch-1 transcripts in E3.5 retina by in situ hybridiza-stage was amplified by PCR using primers specific for Delta-1tion using 35S-labeled C-Delta-1 and C-Notch-1 probes. C-and Notch-1 genes. The PCR products were resolved on 1.5%Delta-1 transcripts were detected throughout the neuralagarose gels and analyzed by Southern analysis. The hybridiza-
tion signals were densitometrically scanned and normalized retina. However, the levels of transcripts as judged bywith the stage-specific levels of b-actin transcripts. Relative the density of silver grains were relatively higher in thelevels of Delta-1 (A) and Notch-1 (B) transcripts at various stages peripheral than in the central retina (Fig. 3A). Addition-of retinal development. Insets show hybridization signals corre- ally, like in the developing chick brain (Henrique et al.,sponding to 418- and 298-bp PCR-amplified Delta-1 (A) and 1995; Myat et al., 1996), the majority of C-Delta-1 tran-Notch-1 (B) transcripts. The Y axes represent arbitrary units of
scripts were localized toward the vitreal surface, the sur-densitometric scanning. Schematic representation of neurogen-face that faces the lens and is analogous to the basal sur-esis in chick retina based on a [3H]thymidine birth dating studyface in the developing brain vesicles (Fig. 3B). In the same(Prada et al., 1991) (C). GC, ganglion cell; AC, amacrine cell;age embryo C-Notch-1 transcripts were detected in theHC, horizontal cell; PC, photoreceptor cell; MC, Muller cell;retina and in adjoining structures such as the lens andBC, bipolar cell. The tips of the triangle indicate the stage at
which 50% of neurogenesis for a specific cell-type is complete. retinal pigment epithelium (Fig. 3D). Consistent with the
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
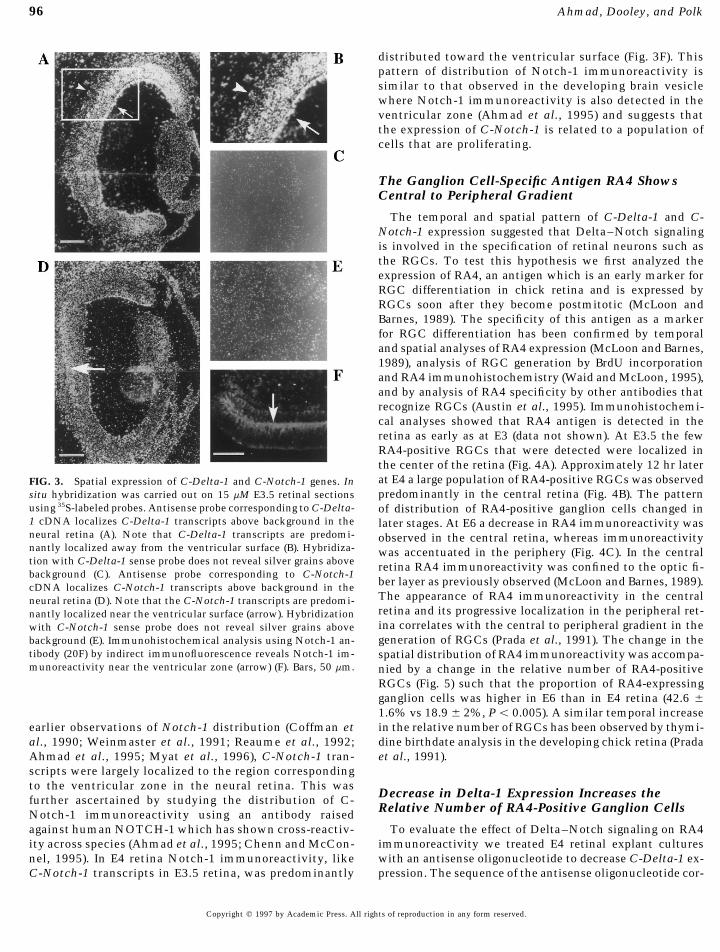
96 Ahmad, Dooley, and Polk
distributed toward the ventricular surface (Fig. 3F). Thispattern of distribution of Notch-1 immunoreactivity issimilar to that observed in the developing brain vesiclewhere Notch-1 immunoreactivity is also detected in theventricular zone (Ahmad et al., 1995) and suggests thatthe expression of C-Notch-1 is related to a population ofcells that are proliferating.
The Ganglion Cell-Specific Antigen RA4 ShowsCentral to Peripheral Gradient
The temporal and spatial pattern of C-Delta-1 and C-Notch-1 expression suggested that Delta–Notch signalingis involved in the specification of retinal neurons such asthe RGCs. To test this hypothesis we first analyzed theexpression of RA4, an antigen which is an early marker forRGC differentiation in chick retina and is expressed byRGCs soon after they become postmitotic (McLoon andBarnes, 1989). The specificity of this antigen as a markerfor RGC differentiation has been confirmed by temporaland spatial analyses of RA4 expression (McLoon and Barnes,1989), analysis of RGC generation by BrdU incorporationand RA4 immunohistochemistry (Waid and McLoon, 1995),and by analysis of RA4 specificity by other antibodies thatrecognize RGCs (Austin et al., 1995). Immunohistochemi-cal analyses showed that RA4 antigen is detected in theretina as early as at E3 (data not shown). At E3.5 the fewRA4-positive RGCs that were detected were localized inthe center of the retina (Fig. 4A). Approximately 12 hr laterat E4 a large population of RA4-positive RGCs was observedFIG. 3. Spatial expression of C-Delta-1 and C-Notch-1 genes. Inpredominantly in the central retina (Fig. 4B). The patternsitu hybridization was carried out on 15 mM E3.5 retinal sections
using 35S-labeled probes. Antisense probe corresponding to C-Delta- of distribution of RA4-positive ganglion cells changed in1 cDNA localizes C-Delta-1 transcripts above background in the later stages. At E6 a decrease in RA4 immunoreactivity wasneural retina (A). Note that C-Delta-1 transcripts are predomi- observed in the central retina, whereas immunoreactivitynantly localized away from the ventricular surface (B). Hybridiza- was accentuated in the periphery (Fig. 4C). In the centraltion with C-Delta-1 sense probe does not reveal silver grains above retina RA4 immunoreactivity was confined to the optic fi-background (C). Antisense probe corresponding to C-Notch-1
ber layer as previously observed (McLoon and Barnes, 1989).cDNA localizes C-Notch-1 transcripts above background in theThe appearance of RA4 immunoreactivity in the centralneural retina (D). Note that the C-Notch-1 transcripts are predomi-retina and its progressive localization in the peripheral ret-nantly localized near the ventricular surface (arrow). Hybridizationina correlates with the central to peripheral gradient in thewith C-Notch-1 sense probe does not reveal silver grains abovegeneration of RGCs (Prada et al., 1991). The change in thebackground (E). Immunohistochemical analysis using Notch-1 an-
tibody (20F) by indirect immunofluorescence reveals Notch-1 im- spatial distribution of RA4 immunoreactivity was accompa-munoreactivity near the ventricular zone (arrow) (F). Bars, 50 mm. nied by a change in the relative number of RA4-positive
RGCs (Fig. 5) such that the proportion of RA4-expressingganglion cells was higher in E6 than in E4 retina (42.6 {1.6% vs 18.9 { 2%, Põ 0.005). A similar temporal increasein the relative number of RGCs has been observed by thymi-earlier observations of Notch-1 distribution (Coffman et
al., 1990; Weinmaster et al., 1991; Reaume et al., 1992; dine birthdate analysis in the developing chick retina (Pradaet al., 1991).Ahmad et al., 1995; Myat et al., 1996), C-Notch-1 tran-
scripts were largely localized to the region correspondingto the ventricular zone in the neural retina. This was
Decrease in Delta-1 Expression Increases thefurther ascertained by studying the distribution of C-Relative Number of RA4-Positive Ganglion CellsNotch-1 immunoreactivity using an antibody raised
against human NOTCH-1 which has shown cross-reactiv- To evaluate the effect of Delta–Notch signaling on RA4immunoreactivity we treated E4 retinal explant culturesity across species (Ahmad et al., 1995; Chenn and McCon-
nel, 1995). In E4 retina Notch-1 immunoreactivity, like with an antisense oligonucleotide to decrease C-Delta-1 ex-pression. The sequence of the antisense oligonucleotide cor-C-Notch-1 transcripts in E3.5 retina, was predominantly
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
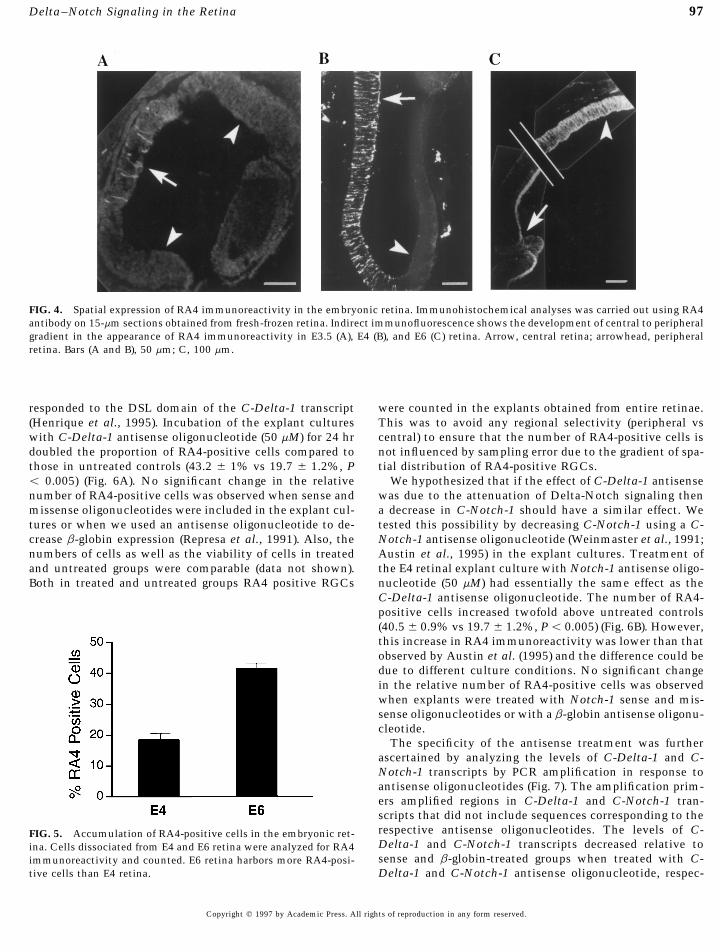
97Delta–Notch Signaling in the Retina
FIG. 4. Spatial expression of RA4 immunoreactivity in the embryonic retina. Immunohistochemical analyses was carried out using RA4antibody on 15-mm sections obtained from fresh-frozen retina. Indirect immunofluorescence shows the development of central to peripheralgradient in the appearance of RA4 immunoreactivity in E3.5 (A), E4 (B), and E6 (C) retina. Arrow, central retina; arrowhead, peripheralretina. Bars (A and B), 50 mm; C, 100 mm.
responded to the DSL domain of the C-Delta-1 transcript were counted in the explants obtained from entire retinae.This was to avoid any regional selectivity (peripheral vs(Henrique et al., 1995). Incubation of the explant cultures
with C-Delta-1 antisense oligonucleotide (50 mM) for 24 hr central) to ensure that the number of RA4-positive cells isnot influenced by sampling error due to the gradient of spa-doubled the proportion of RA4-positive cells compared to
those in untreated controls (43.2 { 1% vs 19.7 { 1.2%, P tial distribution of RA4-positive RGCs.We hypothesized that if the effect of C-Delta-1 antisenseõ 0.005) (Fig. 6A). No significant change in the relative
number of RA4-positive cells was observed when sense and was due to the attenuation of Delta-Notch signaling thena decrease in C-Notch-1 should have a similar effect. Wemissense oligonucleotides were included in the explant cul-
tures or when we used an antisense oligonucleotide to de- tested this possibility by decreasing C-Notch-1 using a C-Notch-1 antisense oligonucleotide (Weinmaster et al., 1991;crease b-globin expression (Represa et al., 1991). Also, the
numbers of cells as well as the viability of cells in treated Austin et al., 1995) in the explant cultures. Treatment ofthe E4 retinal explant culture with Notch-1 antisense oligo-and untreated groups were comparable (data not shown).
Both in treated and untreated groups RA4 positive RGCs nucleotide (50 mM) had essentially the same effect as theC-Delta-1 antisense oligonucleotide. The number of RA4-positive cells increased twofold above untreated controls(40.5 { 0.9% vs 19.7 { 1.2%, P õ 0.005) (Fig. 6B). However,this increase in RA4 immunoreactivity was lower than thatobserved by Austin et al. (1995) and the difference could bedue to different culture conditions. No significant changein the relative number of RA4-positive cells was observedwhen explants were treated with Notch-1 sense and mis-sense oligonucleotides or with a b-globin antisense oligonu-cleotide.
The specificity of the antisense treatment was furtherascertained by analyzing the levels of C-Delta-1 and C-Notch-1 transcripts by PCR amplification in response toantisense oligonucleotides (Fig. 7). The amplification prim-ers amplified regions in C-Delta-1 and C-Notch-1 tran-scripts that did not include sequences corresponding to therespective antisense oligonucleotides. The levels of C-FIG. 5. Accumulation of RA4-positive cells in the embryonic ret-Delta-1 and C-Notch-1 transcripts decreased relative toina. Cells dissociated from E4 and E6 retina were analyzed for RA4sense and b-globin-treated groups when treated with C-immunoreactivity and counted. E6 retina harbors more RA4-posi-
tive cells than E4 retina. Delta-1 and C-Notch-1 antisense oligonucleotide, respec-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
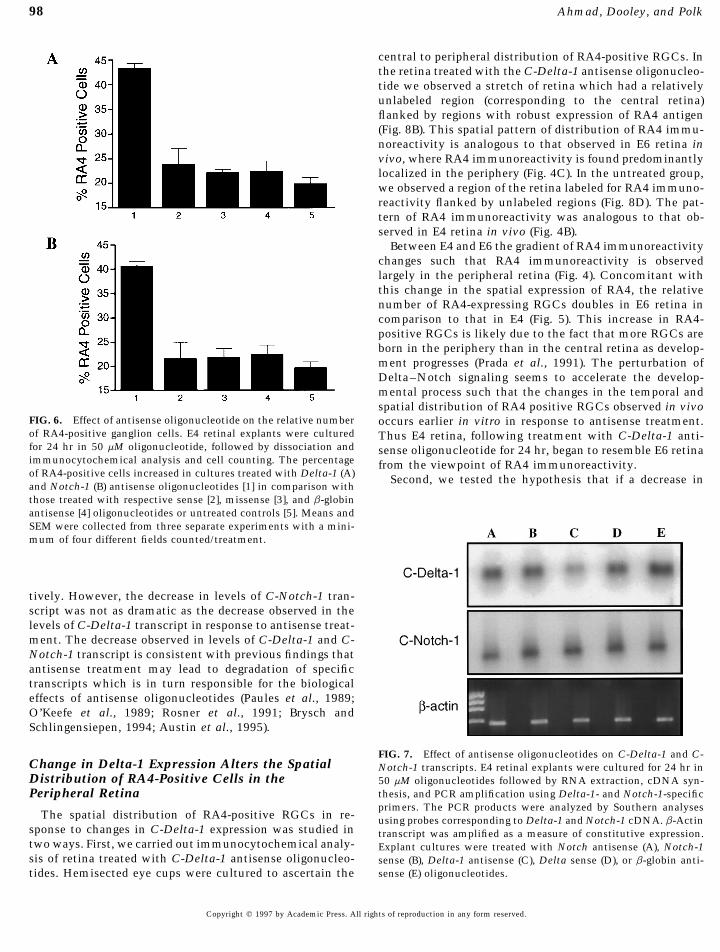
98 Ahmad, Dooley, and Polk
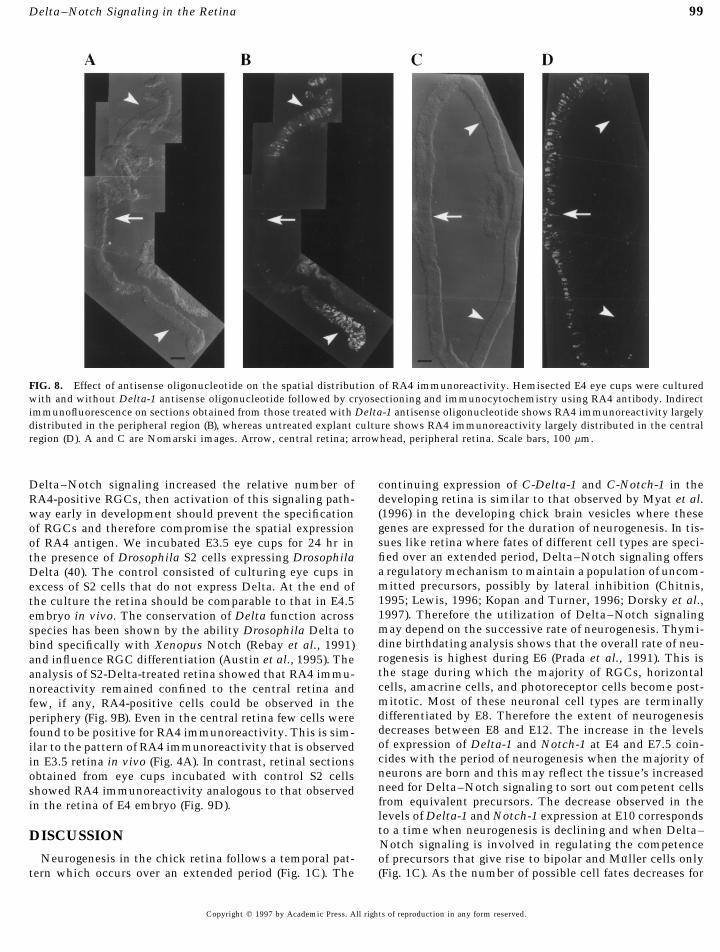
central to peripheral distribution of RA4-positive RGCs. Inthe retina treated with the C-Delta-1 antisense oligonucleo-tide we observed a stretch of retina which had a relativelyunlabeled region (corresponding to the central retina)flanked by regions with robust expression of RA4 antigen(Fig. 8B). This spatial pattern of distribution of RA4 immu-noreactivity is analogous to that observed in E6 retina invivo, where RA4 immunoreactivity is found predominantlylocalized in the periphery (Fig. 4C). In the untreated group,we observed a region of the retina labeled for RA4 immuno-reactivity flanked by unlabeled regions (Fig. 8D). The pat-tern of RA4 immunoreactivity was analogous to that ob-served in E4 retina in vivo (Fig. 4B).
Between E4 and E6 the gradient of RA4 immunoreactivitychanges such that RA4 immunoreactivity is observedlargely in the peripheral retina (Fig. 4). Concomitant withthis change in the spatial expression of RA4, the relativenumber of RA4-expressing RGCs doubles in E6 retina incomparison to that in E4 (Fig. 5). This increase in RA4-positive RGCs is likely due to the fact that more RGCs areborn in the periphery than in the central retina as develop-ment progresses (Prada et al., 1991). The perturbation ofDelta–Notch signaling seems to accelerate the develop-mental process such that the changes in the temporal andspatial distribution of RA4 positive RGCs observed in vivo
FIG. 6. Effect of antisense oligonucleotide on the relative number occurs earlier in vitro in response to antisense treatment.of RA4-positive ganglion cells. E4 retinal explants were cultured Thus E4 retina, following treatment with C-Delta-1 anti-for 24 hr in 50 mM oligonucleotide, followed by dissociation and sense oligonucleotide for 24 hr, began to resemble E6 retinaimmunocytochemical analysis and cell counting. The percentage from the viewpoint of RA4 immunoreactivity.of RA4-positive cells increased in cultures treated with Delta-1 (A) Second, we tested the hypothesis that if a decrease inand Notch-1 (B) antisense oligonucleotides [1] in comparison withthose treated with respective sense [2], missense [3], and b-globinantisense [4] oligonucleotides or untreated controls [5]. Means andSEM were collected from three separate experiments with a mini-mum of four different fields counted/treatment.
tively. However, the decrease in levels of C-Notch-1 tran-script was not as dramatic as the decrease observed in thelevels of C-Delta-1 transcript in response to antisense treat-ment. The decrease observed in levels of C-Delta-1 and C-Notch-1 transcript is consistent with previous findings thatantisense treatment may lead to degradation of specifictranscripts which is in turn responsible for the biologicaleffects of antisense oligonucleotides (Paules et al., 1989;O’Keefe et al., 1989; Rosner et al., 1991; Brysch andSchlingensiepen, 1994; Austin et al., 1995).
FIG. 7. Effect of antisense oligonucleotides on C-Delta-1 and C-Change in Delta-1 Expression Alters the Spatial Notch-1 transcripts. E4 retinal explants were cultured for 24 hr inDistribution of RA4-Positive Cells in the 50 mM oligonucleotides followed by RNA extraction, cDNA syn-
thesis, and PCR amplification using Delta-1- and Notch-1-specificPeripheral Retinaprimers. The PCR products were analyzed by Southern analyses
The spatial distribution of RA4-positive RGCs in re- using probes corresponding to Delta-1 and Notch-1 cDNA. b-Actinsponse to changes in C-Delta-1 expression was studied in transcript was amplified as a measure of constitutive expression.two ways. First, we carried out immunocytochemical analy- Explant cultures were treated with Notch antisense (A), Notch-1sis of retina treated with C-Delta-1 antisense oligonucleo- sense (B), Delta-1 antisense (C), Delta sense (D), or b-globin anti-
sense (E) oligonucleotides.tides. Hemisected eye cups were cultured to ascertain the
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
99Delta–Notch Signaling in the Retina
FIG. 8. Effect of antisense oligonucleotide on the spatial distribution of RA4 immunoreactivity. Hemisected E4 eye cups were culturedwith and without Delta-1 antisense oligonucleotide followed by cryosectioning and immunocytochemistry using RA4 antibody. Indirectimmunofluorescence on sections obtained from those treated with Delta-1 antisense oligonucleotide shows RA4 immunoreactivity largelydistributed in the peripheral region (B), whereas untreated explant culture shows RA4 immunoreactivity largely distributed in the centralregion (D). A and C are Nomarski images. Arrow, central retina; arrowhead, peripheral retina. Scale bars, 100 mm.
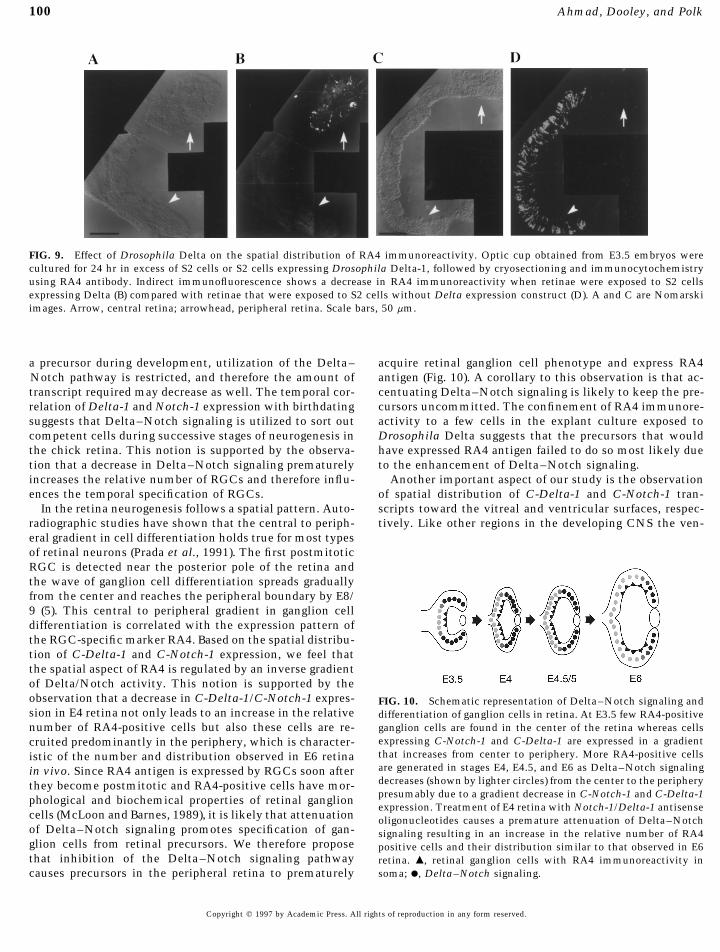
Delta–Notch signaling increased the relative number of continuing expression of C-Delta-1 and C-Notch-1 in thedeveloping retina is similar to that observed by Myat et al.RA4-positive RGCs, then activation of this signaling path-(1996) in the developing chick brain vesicles where theseway early in development should prevent the specificationgenes are expressed for the duration of neurogenesis. In tis-of RGCs and therefore compromise the spatial expressionsues like retina where fates of different cell types are speci-of RA4 antigen. We incubated E3.5 eye cups for 24 hr infied over an extended period, Delta–Notch signaling offersthe presence of Drosophila S2 cells expressing Drosophilaa regulatory mechanism to maintain a population of uncom-Delta (40). The control consisted of culturing eye cups inmitted precursors, possibly by lateral inhibition (Chitnis,excess of S2 cells that do not express Delta. At the end of1995; Lewis, 1996; Kopan and Turner, 1996; Dorsky et al.,the culture the retina should be comparable to that in E4.51997). Therefore the utilization of Delta–Notch signalingembryo in vivo. The conservation of Delta function acrossmay depend on the successive rate of neurogenesis. Thymi-species has been shown by the ability Drosophila Delta todine birthdating analysis shows that the overall rate of neu-bind specifically with Xenopus Notch (Rebay et al., 1991)rogenesis is highest during E6 (Prada et al., 1991). This isand influence RGC differentiation (Austin et al., 1995). Thethe stage during which the majority of RGCs, horizontalanalysis of S2-Delta-treated retina showed that RA4 immu-cells, amacrine cells, and photoreceptor cells become post-noreactivity remained confined to the central retina andmitotic. Most of these neuronal cell types are terminallyfew, if any, RA4-positive cells could be observed in thedifferentiated by E8. Therefore the extent of neurogenesisperiphery (Fig. 9B). Even in the central retina few cells weredecreases between E8 and E12. The increase in the levelsfound to be positive for RA4 immunoreactivity. This is sim-of expression of Delta-1 and Notch-1 at E4 and E7.5 coin-ilar to the pattern of RA4 immunoreactivity that is observedcides with the period of neurogenesis when the majority ofin E3.5 retina in vivo (Fig. 4A). In contrast, retinal sectionsneurons are born and this may reflect the tissue’s increasedobtained from eye cups incubated with control S2 cellsneed for Delta–Notch signaling to sort out competent cellsshowed RA4 immunoreactivity analogous to that observedfrom equivalent precursors. The decrease observed in thein the retina of E4 embryo (Fig. 9D).levels of Delta-1 and Notch-1 expression at E10 correspondsto a time when neurogenesis is declining and when Delta–DISCUSSIONNotch signaling is involved in regulating the competence
Neurogenesis in the chick retina follows a temporal pat- of precursors that give rise to bipolar and Muller cells only(Fig. 1C). As the number of possible cell fates decreases fortern which occurs over an extended period (Fig. 1C). The
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
100 Ahmad, Dooley, and Polk
FIG. 9. Effect of Drosophila Delta on the spatial distribution of RA4 immunoreactivity. Optic cup obtained from E3.5 embryos werecultured for 24 hr in excess of S2 cells or S2 cells expressing Drosophila Delta-1, followed by cryosectioning and immunocytochemistryusing RA4 antibody. Indirect immunofluorescence shows a decrease in RA4 immunoreactivity when retinae were exposed to S2 cellsexpressing Delta (B) compared with retinae that were exposed to S2 cells without Delta expression construct (D). A and C are Nomarskiimages. Arrow, central retina; arrowhead, peripheral retina. Scale bars, 50 mm.
a precursor during development, utilization of the Delta– acquire retinal ganglion cell phenotype and express RA4antigen (Fig. 10). A corollary to this observation is that ac-Notch pathway is restricted, and therefore the amount of
transcript required may decrease as well. The temporal cor- centuating Delta–Notch signaling is likely to keep the pre-cursors uncommitted. The confinement of RA4 immunore-relation of Delta-1 and Notch-1 expression with birthdating
suggests that Delta–Notch signaling is utilized to sort out activity to a few cells in the explant culture exposed toDrosophila Delta suggests that the precursors that wouldcompetent cells during successive stages of neurogenesis in
the chick retina. This notion is supported by the observa- have expressed RA4 antigen failed to do so most likely dueto the enhancement of Delta–Notch signaling.tion that a decrease in Delta–Notch signaling prematurely
increases the relative number of RGCs and therefore influ- Another important aspect of our study is the observationof spatial distribution of C-Delta-1 and C-Notch-1 tran-ences the temporal specification of RGCs.
In the retina neurogenesis follows a spatial pattern. Auto- scripts toward the vitreal and ventricular surfaces, respec-tively. Like other regions in the developing CNS the ven-radiographic studies have shown that the central to periph-
eral gradient in cell differentiation holds true for most typesof retinal neurons (Prada et al., 1991). The first postmitoticRGC is detected near the posterior pole of the retina andthe wave of ganglion cell differentiation spreads graduallyfrom the center and reaches the peripheral boundary by E8/9 (5). This central to peripheral gradient in ganglion celldifferentiation is correlated with the expression pattern ofthe RGC-specific marker RA4. Based on the spatial distribu-tion of C-Delta-1 and C-Notch-1 expression, we feel thatthe spatial aspect of RA4 is regulated by an inverse gradientof Delta/Notch activity. This notion is supported by theobservation that a decrease in C-Delta-1/C-Notch-1 expres- FIG. 10. Schematic representation of Delta–Notch signaling andsion in E4 retina not only leads to an increase in the relative differentiation of ganglion cells in retina. At E3.5 few RA4-positive
ganglion cells are found in the center of the retina whereas cellsnumber of RA4-positive cells but also these cells are re-expressing C-Notch-1 and C-Delta-1 are expressed in a gradientcruited predominantly in the periphery, which is character-that increases from center to periphery. More RA4-positive cellsistic of the number and distribution observed in E6 retinaare generated in stages E4, E4.5, and E6 as Delta–Notch signalingin vivo. Since RA4 antigen is expressed by RGCs soon afterdecreases (shown by lighter circles) from the center to the peripherythey become postmitotic and RA4-positive cells have mor-presumably due to a gradient decrease in C-Notch-1 and C-Delta-1phological and biochemical properties of retinal ganglionexpression. Treatment of E4 retina with Notch-1/Delta-1 antisense
cells (McLoon and Barnes, 1989), it is likely that attenuation oligonucleotides causes a premature attenuation of Delta–Notchof Delta–Notch signaling promotes specification of gan- signaling resulting in an increase in the relative number of RA4glion cells from retinal precursors. We therefore propose positive cells and their distribution similar to that observed in E6that inhibition of the Delta–Notch signaling pathway retina. m, retinal ganglion cells with RA4 immunoreactivity in
soma; l, Delta–Notch signaling.causes precursors in the peripheral retina to prematurely
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

101Delta–Notch Signaling in the Retina
tricular surface in the embryonic retina represents the prin- successive C-Delta-1 activation of the Notch receptor isrequired in order for retinal precursors to follow a specificciple site of cell division (Robinson, 1991). Committed cells
withdraw from the cell-cycle and begin to migrate away fate both temporally and spatially. This is especially criticalwhen numerous cell types are being generated over an ex-from the ventricular surface toward the vitreal surface
where they settle and undergo terminal differentiation. tended period. The fact that C-Delta-1 and C-Notch-1 areexpressed in spatiotemporal concert not only suggests theConsistent with the fact that the activation of the Notch
pathway suppresses differentiation (Coffman et al., 1993; involvement of Notch signaling in sorting out fates of reti-nal neurons in general but also implies that C-Delta-1 mayDorsky, 1995; Austin et al., 1995), C-Notch-1 is expressed
in ventricular zone which mostly harbors dividing progeni- be the dominant ligand in this signaling pathway. The lattersuggestion is supported by the finding that Serrate, anothertors. In contrast, C-Delta-1 transcripts are localized in cells
situated largely toward the vitreal surface. This differential ligand of Notch identified in chick, has not been detectedin the developing retina and therefore may not contributedistribution of C-Notch-1 and C-Delta-1 transcripts is simi-
lar to that observed in the developing chick hind brain and to retinal neurogenesis in this animal model (Myat et al.,1996; Ahmad et al., 1996). While we limited this report tospinal cord (Myat et al., 1996) and suggests a very early
distinction of two population of cells in the developing ret- RGC differentiation, it will be necessary to examine theeffects of perturbing Delta–Notch signaling on the re-ina; those that are expressing C-Notch-1 are proliferating
progenitors and those expressing C-Delta-1 represent a pop- maining retinal cell types.ulation of cells which are either proliferating but poisedto exit mitosis (precursors) or those which have recentlywithdrawn from the cell cycle (nascent neurons). Our study ACKNOWLEDGMENTScannot distinguish between the two possibilities for the C-Delta-1-expressing cells but evidence for both does exist. We thank Steve McLoon for RA4 antibody, David Ish-Horowicz
for cDelta-1 and cNotch-1 cDNA, Spyros Artavanis-Tsakonas forFirst, Henrique et al. (1995), using the double-staining tech-S2 cells and Notch-1 antibody, Wally Thoreson and Harsha Acharyanique to identify BrdU incorporation and C-Delta-1 tran-for critical reading of the manuscript, Huai Yun Han for technicalscripts, have shown that both C-Delta-1-expressing precur-assistance, and Walter Williams and Bill Wassom for photographysors (C-Delta-1/BrdU/) and nascent neurons (C-Delta-and graphics. This work was supported by NEI (R2910313).1/BrdU0) coexist in the developing neural tube, albeit in
different proportions. Second, Dorsky et al. (1997) haveshown that Delta-1-expressing cells in Xenopus retina are REFERENCESpredisposed toward adopting differentiated phenotypes asthey are released from lateral inhibition due to the lack of
Ahmad, I. (1995). Mash-1 is expressed during rod photoreceptorNotch-activating signal (see below). The spatial distributiondifferentiation and binds an E-box element, Eopsin-1 in the rat
of C-Notch-1- and C-Delta-1-expressing cells which are in opsin gene. Dev. Brain Res. 90, 184–189.different stages of differentiation may facilitate lateral inhi- Ahmad, I., Leinders-Zufall, T., Kocsis, J. D., Shepherd, G. M., Zu-bition to maintain a population of uncommitted retinal pro- fall, F., and Barnstable, C. J. (1994). Retinal ganglion cells expressgenitors whose competence can be regulated by subtle fluc- a cGMP-gated cation conductance activatable by nitric oxide do-
nors. Neuron 12, 155–165.tuations in Delta–Notch signaling. This is similar to mech-Ahmad, I., Polk, D., and Dooley, C. (1996). Expression of the puta-anism proposed by Myat et al. (1996) in the developing
tive Notch-1 ligands, Delta-1 and Jagged in the vertebrate retina.brain. Cells that express C-Delta-1 are precursors poised toInvest. Ophthalmol. Visual Sci. Res. 375S, 200.exit mitosis and/or nascent neurons. As they migrate out
Ahmad, I., Zagouras, P., and Artavanis-Tsakonas, S. (1995). Involve-of the ventricular zone they interact with proliferating cellsment of Notch-1 in mammalian retinal neurogenesis: Associa-which express C-Notch-1. This interaction delivers the lat-tion of Notch-1 activity with both immature and terminally dif-
eral inhibition and keeps the proliferating cell from differ- ferentiated cells. Mech. Dev. 53, 73–85.entiating prematurely. However, according to the feedback Artavanis-Tsakonas, S., Delidakis, C., and Fehon, R. G. (1991). Theloop mechanism proposed in Drosophila (Heitzler et al., Notch locus and cell biology of neuroblast segregation. Annu.1996) and Xenopus (Dorsky et al., 1997) the activation of Rev. Cell Biol. 7, 427–452.
Artavanis-Tsakonas, S., Matsuno, K., and Fortini, M. (1995). NotchNotch signaling is likely to reduce the expression of C-signalling. Science 268, 225–232.Delta-1 in proliferating cells, thereby reducing the Notch-
Austin, C. P., Feldman, D. E., Ida, J. A., and Cepko, C. L. (1995).activating signal on the neighboring cells which are express-Vertebrate retinal ganglion cells are selected from competent pro-ing C-Delta-1. The consequence would be that C-Delta-1-genitors by the action of Notch. Development 121, 3637–3650.expressing cells will be released from lateral inhibition and
Brysch, W., and Schlingensiepen, K. (1994). Design and applicationbecome competent to differentiate. Therefore a perturba-of antisense oligonucleotides in cell culture, in vivo and as thera-
tion in the Delta–Notch signaling, either by misexpression peutic agents. Cell. Mol. Biol. 14, 557–568.of Delta-1 (Dorsky et al., 1997) or by decreasing C-Delta-1/ Cagan, R. L., and Ready, D. F. (1989). Notch is required for succes-C-Notch-1 expression as carried out in our study will result sive cell decisions in the developing Drosophila retina. Genesin premature differentiation of precursors. Dev. 3, 1099–1112.
Chen, A., and McConnel, S. K. (1995). Cleavage orientation and theBased on the results presented it can be suggested that
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

102 Ahmad, Dooley, and Polk
asymmetric inheritance of Notch-1 immunoreactivity in mam- (1996). Genes of the Enhancer of Split and Achaete–Scute com-plexes are required for a regulatory loop between Notch and Deltamalian neurogenesis. Cell 82, 631–641.during lateral signalling in Drosophila. Development 122, 161–Chitnis, A., Henrique, D., Lewis, J., Ish-Horowicz, D., and Kitner,171.C. (1995). primary neurogenesis in Xenopus embryos regulated
by a homologue of the Drosophila neurogenic gene Delta. Nature Heitzler, P., and Simpson, P. (1991). The choice of cell fate in theepidermis of Drosophila. Cell 64, 1083–1092.375, 761–766.
Chitnis, A. J. (1995). The role of Notch in lateral inhibition and Henderson, S. T., Gao, D., Lambie, E. J., and Kimble, J. (1994). Lag-2 may encode a signaling ligand for GLP-1 and LIN-12 receptorscell fate specification. Mol. Cell. Biol. 6, 311–321.of C. elegans. Development 120, 2913–2924.Coffman, C., Harris, W., and Kitner, C. R. (1990). Xotch, the xeno-
pus homolog of Drosophila Notch. Science 249, 1438–1441. Henrique, D., Adam, J., Myat, A., Chitnis, A., Lewis, J., and Ish-Horowicz, D. (1995). Expression of Delta homologue in prospec-Coffman, C., Skogland, P., Harris, W. A., and Kitner, C. R. (1993).tive neurons in chick. Nature 375, 787–790.Expression of an extracellular deletion of Xotch diverts cell fate
in the Xenopus embryos. Cell 73, 659–671. Holt, C. E., Bretsch, T. W., Ellis, H. M., and Harris, W. (1988). Cellu-lar determination in the Xenopus retina is independent of lineageCouso, J. P., Knust, E., and Martinez Arias, S. (1995). Serrate andand birthdate. Neuron 1, 15–26.Wingless cooperate to induce vestigial gene expression and wing
formation in Drosophila. Curr. Biol. 5, 1437–1448. Kahn, A. J. (1974). An audioradiographic analysis of the time ofappearance of neurons in the developing chick neural retina. Dev.Diaz Benjumea, F. J., and Cohen, S. M. (1995). Serrate signalBiol. 38, 30–40.through Notch to establish a wingless-dependent organizer at the
dorsal/ventral compartment boundary of the Drosophila wing. Kopan, R., and Nye, J. S. (1995). Vertebrate ligands for Notch. Curr.Biol. 5, 966–969.Development 121, 4215–4225.
Dorsky, R. I., Chang, S. W., Rapaport, D. H., and Harris, W. A. Kopan, R., and Turner, D. L. (1996). The Notch pathway: Democ-racy and aristocracy in the selection of cell fate. Curr. Opin.(1997). Regulation of neuronal diversity in the Xenopus retina
by Delta signalling. Nature 385, 67–70. Neurobiol. 6, 594–601.Kopczynski, C. C., Alton, A. K., Fechtel, K., Kooh, P. J., andDorsky, R. I., Rapaport, D. H., and Harris, W. A. (1995). Xotch in-
hibits cell differentiation in the Xenopus retina. Neuron 14, 487– Muskavitvh, M. A. T. (1988). Delta, a Drosophila neurogenicgene, is transcriptionally complex and encodes a protein related496.to blood coagulation factors and epidermal growth factor of verte-Fehon, R. G., Kooh, P. J., Rebay, I., Regan, C. L., Xu, T., Muska-brates. Genes Dev. 2, 1723–1735.vitch, M. A. T., and Artavanis-Tsakonas, S. (1990). Molecular in-
teractions between the protein products of the neurogenic loci La Vail, M. M., Rapaport, D. H., and Rakic, P. (1991). Cytogenesisin the monkey retina. J. Comp. Neurol. 309, 86–114.Notch and Delta, two EGF-homologous genes in Drosophila. Cell
61, 523–534. Lewis, J. (1996). Neurogenic genes and vertebrate neurogenesis.Curr. Opin. Biol. 6, 3–10.Fitzgerald, K., and Greenwald, I. (1995). Interchangeability of
Caenorhabditis elegans DSL proteins and intrinsic signalling ac- Lindsell, C. E., Shawber, C. J., Boulter, J., and Weinmaster, G.(1995). Jagged: A mammalian ligand that activates Notch-1. Celltivity of their extracellular domain in vivo. Development 121,
4275–4282. 80, 909–917.McLoon, S. C., and Barnes, R. B. (1989). Early differentiation of reti-Fleming, R. J., Scottgale, T. M., Diederick, R. J., and Artavanis-Tsa-
konas, S. (1990). The gene Serrate encodes a putative EGF-like nal ganglion cells: An axonal protein expressed by premigratoryand migrating retinal ganglion cells. J. Neurosci. 9, 1424–1432.transmembrane protein essential for proper ectodermal develop-
ment. Genes Dev. 4, 2188–2201. Muskavitch, M. A. T. (1994). Delta-Notch signaling and Drosophilacell fate choice. Dev. Biol. 166, 415–430.Fortini, M. E., Rebay, I., and Artavanis-Tsakonas, S. (1993). An acti-
vated Notch receptor blocks cell-fate commitment in the devel- Myat, A., Henrique, D., Ish-Horowicz, D., and Lewis, J. (1996). Aoping Drosophila eye. Nature 365, 555–567. chick homologue of Serrate and its relationship with Notch and
Delta homologues during central neurogenesis. Dev. Biol. 174,Gao, D., and Kimble, J. (1995). APX-1 can substitute for its homologlag2 to direct cell interactions throughout Caenorhabditis eleg- 233–247.ans development. Proc. Natl. Acad. Sci. USA 92, 9839–9842. O’Keefe, S. J., Wolfe, H., Kiessling, A. A., and Cooper, G. M. (1989).
Microinjection of antisense c-mos oligonucleotide prevents mei-Greenwald, I., and Rubin, G. M. (1992). Making a difference: Therole of cell–cell interactions in establishing separate identities osis II in the maturing mouse egg. Proc. Natl. Acad Sci. USA 86,
7038–7042.for equivalent cells. Cell 68, 271–281.Grillo, M., and Margolis, F. L. (1991). Use of reverse transcriptase Paules, R. S., Buccione, R., Moschel, R. C., Vande Woude, G. F.,
and Kelly, T. J. (1989). Mouse mos protooncogene product is pres-polymerase chain reaction to monitor expression of intronlessgenes. Biotechniques 9, 262–268. ent and functions during oogenesis. Proc. Natl. Acad Sci. USA
86, 5395–5399.Gu, Y., Hukriede, N. A., and Fleming, R. J. (1995). Serrate expres-sion can functionally replace Delta activity during neurobalst Prada, C. , Puga, J., Perez-Mendez, L., Lopez, R., and Ramirez, G.
(1991). Spatial and temporal patterns of neurogenesis in the chicksegregation in the Drosophila embryo. Development 121, 535–544. retina. Eur. J. Neurosci. 3, 559–569.
Price, J., Turner, D., and Cepko, C. (1987). Lineage analysis in theHamburger, V., and Hamilton, H. L. (1951). A series of normalstages in the development of the chick embryo. J. Morphol. 88, vertebrate nervous system by retrovirus mediated gene transfer.
Proc. Natl. Acad Sci. USA 84, 156–160.49–92.Harman, A. M., and Beazley, L. D. (1989). Generation of retinal Reaume, A. G., Canlon, R. A., Zirgibl, R., Yamaguchi, T. P., and
Rossant, J. (1992). Expression analysis of Notch homologin thecells in the Wallaby Setonix brachyurus (quokka). Neuroscience28, 219–232. mouse embryo. Dev. Biol. 154, 377–387.
Rebay, I., Fleming, R. J., Fehon, R. G., Cherbas, L., Cherbas, P.,Heitzler, P., Bourouis, M., Ruel, L., Carteret, C., and Simpson, P.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba

103Delta–Notch Signaling in the Retina
and Artavanis-Tsakonas (1991). Specific EGF-repeats of Notch gene Serrate encodes an EGF-Like transmembrane protein witha complex expression pattern in embryos and wing disc. Develop-mediate interactions with Delta and Serrate: Implications of
Notch as a multifunctional receptor. Cell 67, 687–699. ment 111, 749–761.Turner, D. L., and Cepko, C. L. (1987). A common progenitor forReh, T. A., and Tully, T. (1986). Regulation of tyrosine-hydroxy-
neurons and glia persists in rat retina late in development. Naturelase-containing amacrine cell number in larval frog retina. Dev.238, 131–136.Biol. 114, 463–469.
Turner, D. L., Snyder, E. Y., and Cepko, C. L. (1990). Lineage-inde-Represa, J., Leon, Y., and Miner, C. (1991). The int-2 proto-oncogenependent determination of cell type in the embryonic mouse ret-is responsible for the induction of the inner ear. Nature 353, 561–ina. Neuron 4, 833–845.563.
Vaessin, H., Bremer, K. A., Knust, E., and Campos-Ortega, J. A.Robinson, S. R. (1991). Development of the mammalian retina. In(1987). The neurogenesis gene Delta of Drosophila melanogaster‘‘Neuroanatomy of Visual Pathways and Their Development’’ (B.is expressed in neurogenic territories and encodes a putativeDreher and S. R. Robinson, Eds.), Vol. 3, pp. 69–128. Macmillan,transmembrane protein with EGF-like repeats. EMBO J. 6, 3430–UK.3440.Rosner, M. H., Desanto, R. J., Arnheiter, H., and Staredt, L. M.
Waid, D. K., and McLoon, S. C. (1995). Immediate differentiation(1991). Oct-3 is a maternal factor required for the first mouseof ganglion cells following mitosis in the developing retina. Neu-embryonic division. Cell 64, 1103–1110.ron 14, 117–124.Sidman, R. L. (1961). Histogenesis of mouse retina studied with
Weinmaster, G., Roberts, V. J., and Lemke, G. (1991). A homologthymidine 3H. In ‘‘Structure of the Eye’’ (G. K. Smelser, Ed.), pp.of Drosophila Notch expressed during mammalian development.487–506. Academic Press, New York.Development 113, 199–205.Speicher, S. A., Thomas, U., Hinz, U., and Knust, E. (1994). The
Wetts, R., and Fraser, S. E. (1988). Multipotent precursors can giveSerrate locus of Drosophila and its role in morphogenesis of therise to all major cell types of the frog retina. Science 239, 1142–wing imaginal discs: Control of cell proliferation. Development1145.120, 535–544.
Young, R. W. (1985). Cell differentiation in the retina of the mouse.Tax, F. E., Yeargers, J. J., and Thomas, J. H. (1994). Sequence of C.Anat. Rec. 212, 499–205.elegans lag2 reveals a cell-signalling domain shared with Delta
and Serrate of Drosophila. Nature 368, 150–154. Received for publication December 19, 1996Accepted February 20, 1997Thomas, U., Speicher, S. A., and Knust, E. (1991). The Drosophila
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8546 / 6x22$$$$$1 04-09-97 20:11:01 dba
Related Documents











