Drug and Alcohol Dependence 129 (2013) 102–109 Contents lists available at SciVerse ScienceDirect Drug and Alcohol Dependence j ourna l ho me pag e: www.elsevier.com/locate/drugalcdep Decreased frontal lobe phosphocreatine levels in methamphetamine users Young-Hoon Sung a,b,∗ , Deborah A. Yurgelun-Todd a,b,c , Xian-Feng Shi a , Douglas G. Kondo a,b,c , Kelly J. Lundberg a , Erin C. McGlade a,b,c , Tracy L. Hellem a , Rebekah S. Huber a , Kristen K. Fiedler a , Renee E. Harrell a , Bethany R. Nickerson a , Seong-Eun Kim d , Eun-Kee Jeong d , Perry F. Renshaw a,b,c a Department of Psychiatry, University of Utah, Salt Lake City, UT, USA b The Brain Institute, University of Utah, 383 Colorow Drive, Salt Lake City, UT, USA c VISN 19 MIRECC, Salt Lake City, UT, USA d Department of Radiology, University of Utah, Salt Lake City, UT, USA a r t i c l e i n f o Article history: Received 10 April 2012 Received in revised form 14 August 2012 Accepted 21 September 2012 Available online 18 October 2012 Keywords: Methamphetamine Phosphorus magnetic resonance spectroscopy Phosphocreatine a b s t r a c t Background: Mitochondria-related mechanisms have been suggested to mediate methamphetamine (METH) toxicity. However, changes in brain energetics associated with high-energy phosphate metabolism have not been investigated in METH users. Phosphorus-31 ( 31 P) magnetic resonance spec- troscopy (MRS) was used to evaluate changes in mitochondrial high energy phosphates, including phosphocreatine (PCr) and -nucleoside triphosphate (-NTP, primarily ATP in brain) levels. We hypoth- esized that METH users would have decreased high-energy PCr levels in the frontal gray matter. Methods: Study participants consisted of 51 METH (age = 32.8 ± 6.7) and 23 healthy comparison (age = 31.1 ± 7.5) subjects. High-energy phosphate metabolite levels were compared between the groups and potential gender differences were explored. Results: METH users had lower ratios of PCr to total pool of exchangeable phosphate (PCr/TPP) in the frontal lobe as compared to the healthy subjects (p = .001). The lower PCr levels in METH subjects were significantly associated with lifetime amount of METH use (p = .003). A sub-analysis for gender differences revealed that female METH users, who had lower daily amounts (1.1 ± 1.0 g) of METH use than males (1.4 ± 1.7 g), had significantly lower PCr/TPP ratios than male METH users, controlling for the amount of METH use (p = .02). Conclusions: The present findings suggest that METH compromises frontal lobe high-energy phosphate metabolism in a dose-responsive manner. Our findings also suggest that the abnormality in frontal lobe high-energy phosphate metabolism might be more prominent in female than in male METH users. This is significant as decreased PCr levels have been associated with depressive symptoms, and poor responses to antidepressant treatment have been reported in those with decreased PCr levels. © 2012 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Methamphetamine (METH) abuse is a disorder characterized by compulsive METH-craving and consumption despite an appar- ent awareness of serious negative consequences. METH use has been linked to the emergence of psychotic symptoms (Iyo et al., 2004; London et al., 2004; Winslow et al., 2007) as well as morpho- logical, functional, and neurochemical abnormalities in multiple brain areas (Bae et al., 2006; Chung et al., 2007; Ernst et al., 2000; Presented in part at the 73rd annual meeting of the College on Problems of Drug Dependence, Hollywood, FL, June 18–23, 2011. ∗ Corresponding author at: 383 Colorow Drive, Room 313, The Brain Institute, University of Utah, Salt Lake City, UT 84108, USA. Tel.: +1 801 587 1555; fax: +1 801 585 5375. E-mail address: [email protected] (Y.-H. Sung). Hwang et al., 2006; Oh et al., 2005). Alterations in monoaminer- gic neurotransmission (Kokoshka et al., 1998; Ricaurte et al., 1980; Robinson and Berridge, 1993) in the frontal lobe have been related to cognitive impairments in METH users because prefrontal cortical neural networks play a central role in impaired decision-making and inhibitory control (Lubman et al., 2004; Yucel and Lubman, 2007). In METH toxicity, multiple lines of evidence suggest that dys- functional energy metabolism plays an important role. For instance, (1) impairments of mitochondrial function have been reported after administration of METH to animals including impairments in mito- chondrial electron transport chain enzyme complexes (Brown et al., 2005; Burrows et al., 2000a; Klongpanichapak et al., 2006); (2) METH toxicity involves a depletion of energy stores, evidenced by synergistic metabolic inhibition by METH resulting in depletion of striatal dopamine content (Burrows et al., 2000b); and (3) ex vivo METH exposure has been associated with oxidative cell injury 0376-8716/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved. http://dx.doi.org/10.1016/j.drugalcdep.2012.09.015

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

D
YKRa
b
c
d
a
ARRAA
KMPsP
1
beb2lb
D
Uf
0h
Drug and Alcohol Dependence 129 (2013) 102– 109
Contents lists available at SciVerse ScienceDirect
Drug and Alcohol Dependence
j ourna l ho me pag e: www.elsev ier .com/ locate /drugalcdep
ecreased frontal lobe phosphocreatine levels in methamphetamine users�
oung-Hoon Sunga,b,∗, Deborah A. Yurgelun-Todda,b,c, Xian-Feng Shia, Douglas G. Kondoa,b,c,elly J. Lundberga, Erin C. McGladea,b,c, Tracy L. Hellema, Rebekah S. Hubera, Kristen K. Fiedlera,enee E. Harrell a, Bethany R. Nickersona, Seong-Eun Kimd, Eun-Kee Jeongd, Perry F. Renshawa,b,c
Department of Psychiatry, University of Utah, Salt Lake City, UT, USAThe Brain Institute, University of Utah, 383 Colorow Drive, Salt Lake City, UT, USAVISN 19 MIRECC, Salt Lake City, UT, USADepartment of Radiology, University of Utah, Salt Lake City, UT, USA
r t i c l e i n f o
rticle history:eceived 10 April 2012eceived in revised form 14 August 2012ccepted 21 September 2012vailable online 18 October 2012
eywords:ethamphetamine
hosphorus magnetic resonancepectroscopyhosphocreatine
a b s t r a c t
Background: Mitochondria-related mechanisms have been suggested to mediate methamphetamine(METH) toxicity. However, changes in brain energetics associated with high-energy phosphatemetabolism have not been investigated in METH users. Phosphorus-31 (31P) magnetic resonance spec-troscopy (MRS) was used to evaluate changes in mitochondrial high energy phosphates, includingphosphocreatine (PCr) and �-nucleoside triphosphate (�-NTP, primarily ATP in brain) levels. We hypoth-esized that METH users would have decreased high-energy PCr levels in the frontal gray matter.Methods: Study participants consisted of 51 METH (age = 32.8 ± 6.7) and 23 healthy comparison(age = 31.1 ± 7.5) subjects. High-energy phosphate metabolite levels were compared between the groupsand potential gender differences were explored.Results: METH users had lower ratios of PCr to total pool of exchangeable phosphate (PCr/TPP) in thefrontal lobe as compared to the healthy subjects (p = .001). The lower PCr levels in METH subjects weresignificantly associated with lifetime amount of METH use (p = .003). A sub-analysis for gender differencesrevealed that female METH users, who had lower daily amounts (1.1 ± 1.0 g) of METH use than males(1.4 ± 1.7 g), had significantly lower PCr/TPP ratios than male METH users, controlling for the amount of
METH use (p = .02).Conclusions: The present findings suggest that METH compromises frontal lobe high-energy phosphatemetabolism in a dose-responsive manner. Our findings also suggest that the abnormality in frontal lobehigh-energy phosphate metabolism might be more prominent in female than in male METH users. This issignificant as decreased PCr levels have been associated with depressive symptoms, and poor responsesto antidepressant treatment have been reported in those with decreased PCr levels.. Introduction
Methamphetamine (METH) abuse is a disorder characterizedy compulsive METH-craving and consumption despite an appar-nt awareness of serious negative consequences. METH use haseen linked to the emergence of psychotic symptoms (Iyo et al.,
004; London et al., 2004; Winslow et al., 2007) as well as morpho-ogical, functional, and neurochemical abnormalities in multiplerain areas (Bae et al., 2006; Chung et al., 2007; Ernst et al., 2000;
� Presented in part at the 73rd annual meeting of the College on Problems of Drugependence, Hollywood, FL, June 18–23, 2011.∗ Corresponding author at: 383 Colorow Drive, Room 313, The Brain Institute,niversity of Utah, Salt Lake City, UT 84108, USA. Tel.: +1 801 587 1555;
ax: +1 801 585 5375.E-mail address: [email protected] (Y.-H. Sung).
376-8716/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.drugalcdep.2012.09.015
© 2012 Elsevier Ireland Ltd. All rights reserved.
Hwang et al., 2006; Oh et al., 2005). Alterations in monoaminer-gic neurotransmission (Kokoshka et al., 1998; Ricaurte et al., 1980;Robinson and Berridge, 1993) in the frontal lobe have been relatedto cognitive impairments in METH users because prefrontal corticalneural networks play a central role in impaired decision-makingand inhibitory control (Lubman et al., 2004; Yucel and Lubman,2007).
In METH toxicity, multiple lines of evidence suggest that dys-functional energy metabolism plays an important role. For instance,(1) impairments of mitochondrial function have been reported afteradministration of METH to animals including impairments in mito-chondrial electron transport chain enzyme complexes (Brown et al.,2005; Burrows et al., 2000a; Klongpanichapak et al., 2006); (2)
METH toxicity involves a depletion of energy stores, evidenced bysynergistic metabolic inhibition by METH resulting in depletionof striatal dopamine content (Burrows et al., 2000b); and (3) exvivo METH exposure has been associated with oxidative cell injury
hol De
am2tthcp(22Smlb
uhcrttIdmeduept
euhe(luMd2udwp
abArawmatcel
2
2
tp
Y.-H. Sung et al. / Drug and Alco
nd apoptosis in rat cortical neuron and undifferentiated pheochro-ocytoma (PC12) cells (Cunha-Oliveira et al., 2006; Oliveira et al.,
002). Considering the evidence for mitochondrial involvement inhe potential pathophysiology of METH toxicity, it is not surprisinghat proton (1H) magnetic resonance spectroscopy (MRS) studiesave consistently reported that METH users, relative to healthyomparison subjects, have decreased levels of total creatine (phos-hocreatine plus creatine) as well as decreased N-acetylaspartateNAA, a marker of neuronal viability or integrity (Moffett et al.,007)) levels (Chang et al., 2005; Ernst et al., 2000; Nordahl et al.,002; Sailasuta et al., 2010b; Sekine et al., 2002; Smith et al., 2001;ung et al., 2007; Taylor et al., 2007). As NAA synthesis occurs pri-arily in the mitochondria (Patel and Clark, 1979), decreased NAA
evels in METH users are potentially consistent with compromisedrain energetics.
Phosphocreatine (PCr) and adenosine triphosphate (ATP) makep the PCr–ATP energy buffering system in neuronal cells that haveigh and fluctuating energy demands. The enzyme creatine kinaseontrols the transfer of a phosphate group from PCr to ADP, therebyeplenishing brain ATP. It is reported that PCr serves as a buffero maintain constant ATP levels so that ATP levels remain rela-ively stable at the cost of PCr expenditure (Schlattner et al., 2006).n the fluctuating energy requirements of neurons, mitochondrialysfunction may lead to decreased formation of phosphocreatineediated by the mitochondrial creatine kinase isoenzyme (Dolder
t al., 2001; Wallimann et al., 1998). Regarding region specificeficits, frontal hypometabolism has been reported in METH userssing positron emission tomography (PET; Kim et al., 2005; Londont al., 2004, 2005). These findings would be consistent with theotential mitochondrial abnormality and decreased PCr levels inhe frontal lobe of METH users.
Published neuroimaging studies have reported gender differ-nces in METH toxicity with favorable outcomes in female METHsers in terms of frontal glucose metabolism and white matteryperintensities/integrity (Bae et al., 2006; Chung et al., 2007; Kimt al., 2005), probably reflecting a protective effect of estrogenDhandapani and Brann, 2002). However, female psychostimu-ant users are more sensitive to the reinforcing effects than malesers (Anker and Carroll, 2011; Carroll et al., 2004). Also, femaleETH users have a higher incidence of depression and more severe
epressive symptoms than male METH users (Dluzen and Liu,008; Hser et al., 2005; Kalechstein et al., 2000). In non-METHsers, depression severity has been significantly associated withecreased brain PCr levels (Kato et al., 1992), but it is not knownhat the effects of METH use will be on brain high energy phos-hate metabolism in female METH users.
Phosphorus-31 (31P)-MRS provides a unique method to evalu-te changes in high-energy phosphate metabolites such as PCr andeta-nucleoside triphosphate (�-NTP), which arises primarily fromTP in brain (Renshaw et al., 2001). To date, there have been noeports measuring 31P-MRS metabolite levels in METH users rel-tive to healthy subjects. In this study, we aimed to investigatehether METH use significantly altered high energy phosphateetabolism. It was hypothesized that first, METH use would be
ssociated with decreased high-energy PCr levels by 31P-MRS inhe frontal cortex, and second, that altered PCr levels will be signifi-antly correlated with lifetime amount of METH use. In addition, wexplored possible gender differences in the phosphorus metaboliteevels in METH users.
. Methods
.1. Subjects
This was a cross-sectional study in which phosphorus MRS data was acquiredo examine brain metabolite alterations related to METH use. The study partici-ants consisted of 51 METH dependent subjects (age = 32.8 ± 6.7, female = 23) and 23
pendence 129 (2013) 102– 109 103
healthy comparison subjects (age = 31.1 ± 7.5, female = 11). Each individual under-went two dimensional phosphorus magnetic resonance spectroscopic imaging (2D31P-MRSI) as well as assessment of clinical and drug abuse history. METH-dependentsubjects were evaluated for the severity of their lifetime METH use (METH amount,frequency, duration, and abstinence). Inclusion criteria for METH subjects were asfollows: (1) age 18–55 years, (2) subjects who met diagnostic criteria for currentmethamphetamine abuse or dependence as their preferred drug of abuse as deter-mined by the Structured Clinical Interview for DSM-IV (SCID-IV). A board certifiedpsychiatrist took complete medical histories and physical examinations of the sub-jects. The SCID-IV was administered by a trained psychologist (ECM), and (3) METHuse within the past six months. Exclusion criteria for METH subjects included: (1)major medical or neurological disorders, including HIV seropositivity; (2) comorbidpsychiatric disorders including schizophrenia, bipolar disorder, and use of otherillicit drugs as preferred drug of abuse; (3) major sensorimotor handicaps (e.g.,deafness, blindness, and paralysis), full scale IQ < 70 or learning disabilities; and(4) contraindications to magnetic resonance imaging.
Healthy comparison (HC) subjects were recruited with the inclusion criteria(1) age 18–55 years, (2) no dependence or abuse of alcohol, METH, amphetamine,cocaine, heroin, alcohol, and cannabis, and (3) no psychiatric, neurologic, and med-ical disease identified by physical examination. The healthy subjects were matchedon age and gender with METH users. The healthy subjects had a slightly higher levelof education than METH users and this difference was controlled for statistically.Exclusion criteria for the HC subjects were the same as for the METH users. Thestudy protocol was approved by the Institutional Review Boards of the University ofUtah and the Department of Human Services of the State of Utah. Written informedconsent was obtained from all study subjects before participation.
2.2. Magnetic resonance imaging and spectroscopy data acquisition andprocessing
2.2.1. Structural MR images. Brain MR imaging was performed using a 3 TeslaSiemens scanner (Trio, Siemens AG, Erlangen, Germany) and a 31P/1H double-tunedvolume head coil (Clinical MR Solutions LLC, Brookfield, WI) for transmission andreception. To obtain high resolution T1-weighted anatomical images for tissuesegmentation and positioning MRS grids, a three dimensional chemical Mag-netization Prepared Rapid Acquisition Gradient Echo (MPRAGE) pulse sequencewas used. The parameters for the structural MRI were as follows: T1 weightedimage – TR = 2000 ms, TE = 3.37 ms, TI = 1100 ms, average number = 1, flip angle = 8◦ ,FOV = 256 mm, matrix 256 × 192 × 144, bandwidth = 300 Hz/pixel, slice thickness1.0 mm and no gap.
2.2.2. 31P magnetic resonance spectroscopy. Phosphorus spectra were acquired onthe same 3 Tesla Siemens system using the 31P/1H double-tuned volume headcoil. The spectra were obtained using a two dimensional-chemical shift imaging(2D-CSI) free induction decay (FID) pulse sequence with TR = 3000, TE = 2.3 ms, aver-age number = 36, flip angle = 90◦ , vector size = 1024, FOV = 200 mm × 200 mm, slicethickness = 2.5 cm, matrix 8 × 8, sampling bandwidth = 2.5 kHz, and voxel dimen-sion = 2.5 cm × 2.5 cm. The 2D-CSI grid was positioned covering an axial brain slicejust above an imaginary line connecting the anterior commissure and posterior com-missure. The acquisition matrix of the 2D-CSI grid was 8 × 8 with the slice thickness25 mm (Fig. 1). Shimming was performed over the excited brain volume. Since MRSdata is significantly affected by magnetic field inhomogeneity, high order advancedshimming routine on the Siemens system was used to achieve linewidths of less thanor equal to 15 Hz for the unsuppressed water signal. The proton channel was used forshimming, localization, and anatomic imaging. As an a priori region of interest (ROI),frontal lobe spectroscopic data were quantified. Also, as a control region for the com-parison, temporoparietal lobe and occipital lobe were included in the spectroscopicand statistical analyses. The frontal lobe was selected as an active ROI because ofprior reports suggesting frontal neurochemical abnormalities and hypometabolismin METH users (Kim et al., 2005; London et al., 2004, 2005; Sailasuta et al., 2010a).
2.2.3. 31P-MRS data analysis. Before performing 2D fast Fourier transform (FFT) onraw data, a Hamming filter was applied to reduce the effect of the point-spread-function, and each free induction decay (FID) was line-broadened with 10 Hz ofapodization. After Fourier transformation and frequency shift correction, zero-/first-order phase correction and baseline correction with polynomial interpolationwere applied. Metabolite location error resulting from different chemical shift dis-placement was corrected along in-plane readout and phase-encoding directions.Following that, spatial filtering with a Hamming window function was implementedto reduce the signal contamination from neighboring voxels. The preprocessed 31P-MRSI data was fitted using jMRUI software (Naressi et al., 2001) with the AdvancedMethod for Accurate, Robust and Efficient Spectral (AMARES) fitting algorithm(Vanhamme et al., 1997). Metabolites of interest were PCr, �-NTP, and their ratio(Fig. 1C). Each metabolite concentration was expressed relative to the total pool ofexchangeable phosphate (TPP; Blumberg et al., 1999). Referencing 31P metabolites
to TPP facilitated effective evaluation of high energy phosphate metabolism (Amesset al., 1997; Cady et al., 2008; Iwata et al., 2008).From the registered anatomical images, tissue segmentation was performedusing FSL (FMRIB’s Software Library) software so that cerebrospinal fluid (CSF)-corrected metabolite concentrations as well as gray matter percentage in each voxel
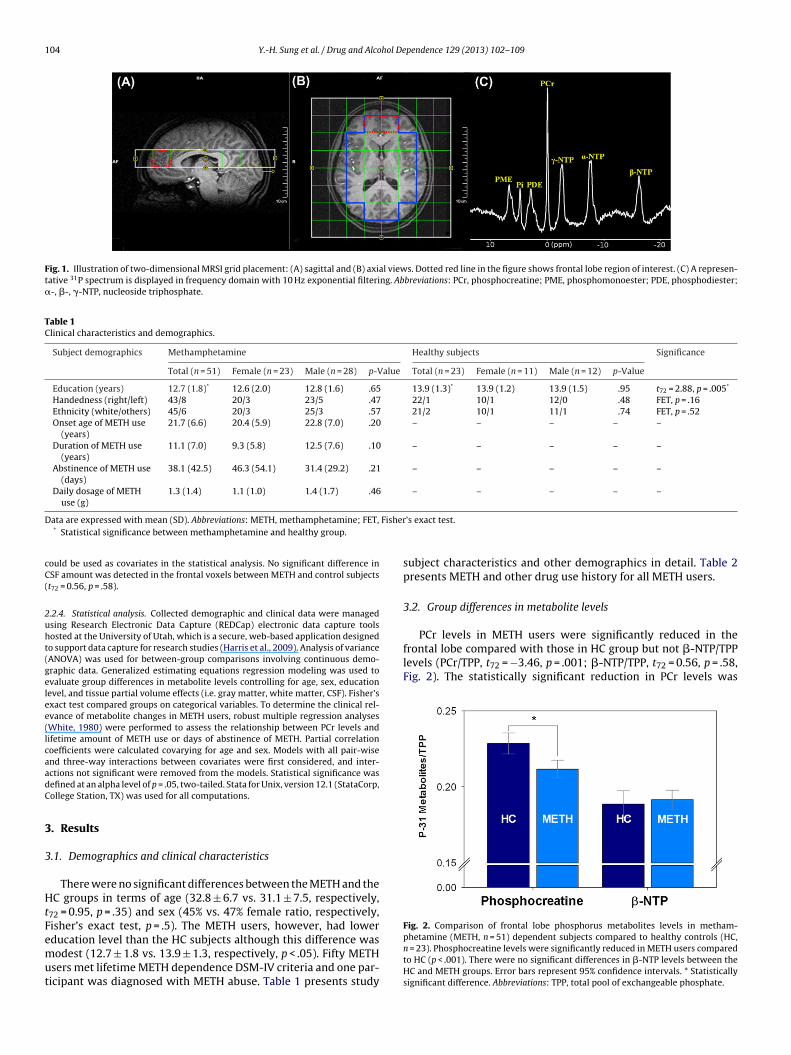
104 Y.-H. Sung et al. / Drug and Alcohol Dependence 129 (2013) 102– 109
Fig. 1. Illustration of two-dimensional MRSI grid placement: (A) sagittal and (B) axial views. Dotted red line in the figure shows frontal lobe region of interest. (C) A represen-tative 31P spectrum is displayed in frequency domain with 10 Hz exponential filtering. Abbreviations: PCr, phosphocreatine; PME, phosphomonoester; PDE, phosphodiester;�-, �-, �-NTP, nucleoside triphosphate.
Table 1Clinical characteristics and demographics.
Subject demographics Methamphetamine Healthy subjects Significance
Total (n = 51) Female (n = 23) Male (n = 28) p-Value Total (n = 23) Female (n = 11) Male (n = 12) p-Value
Education (years) 12.7 (1.8)* 12.6 (2.0) 12.8 (1.6) .65 13.9 (1.3)* 13.9 (1.2) 13.9 (1.5) .95 t72 = 2.88, p = .005*
Handedness (right/left) 43/8 20/3 23/5 .47 22/1 10/1 12/0 .48 FET, p = .16Ethnicity (white/others) 45/6 20/3 25/3 .57 21/2 10/1 11/1 .74 FET, p = .52Onset age of METH use
(years)21.7 (6.6) 20.4 (5.9) 22.8 (7.0) .20 – – – – –
Duration of METH use(years)
11.1 (7.0) 9.3 (5.8) 12.5 (7.6) .10 – – – – –
Abstinence of METH use(days)
38.1 (42.5) 46.3 (54.1) 31.4 (29.2) .21 – – – – –
Daily dosage of METHuse (g)
1.3 (1.4) 1.1 (1.0) 1.4 (1.7) .46 – – – – –
D Fisher
cC(
2uht(gelee(lcaadC
3
3
HtFemut
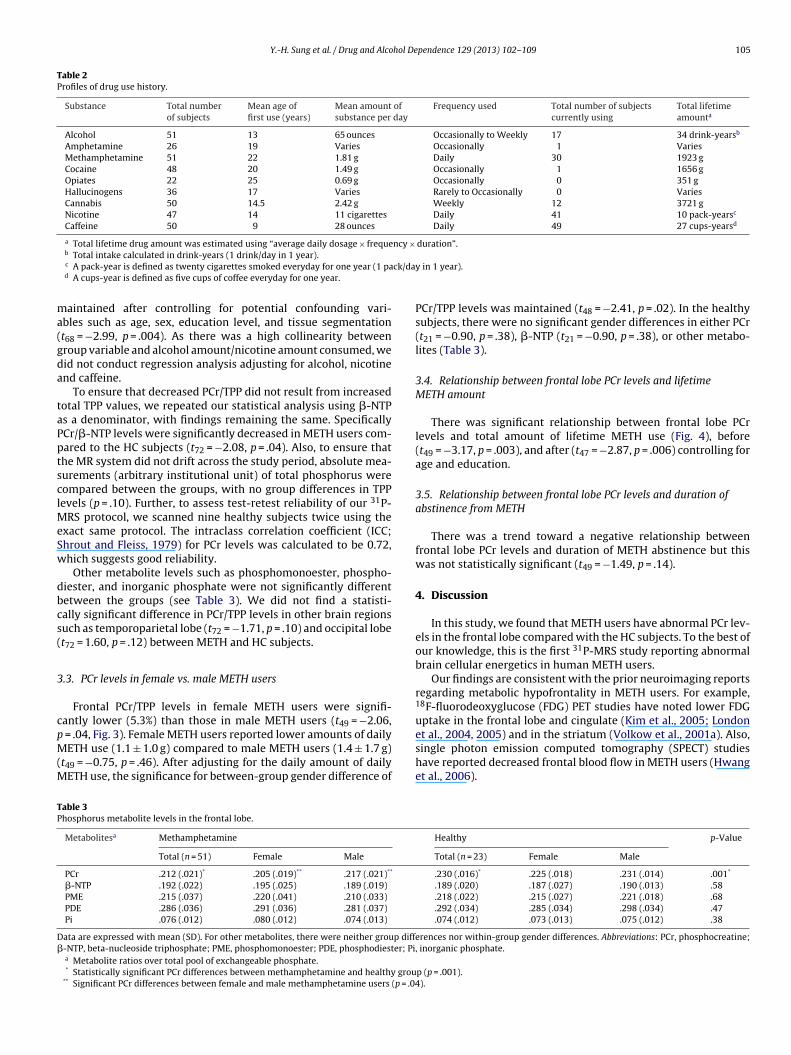
PCr levels in METH users were significantly reduced in thefrontal lobe compared with those in HC group but not �-NTP/TPPlevels (PCr/TPP, t72 = −3.46, p = .001; �-NTP/TPP, t72 = 0.56, p = .58,Fig. 2). The statistically significant reduction in PCr levels was
Fig. 2. Comparison of frontal lobe phosphorus metabolites levels in metham-
ata are expressed with mean (SD). Abbreviations: METH, methamphetamine; FET,* Statistical significance between methamphetamine and healthy group.
ould be used as covariates in the statistical analysis. No significant difference inSF amount was detected in the frontal voxels between METH and control subjectst72 = 0.56, p = .58).
.2.4. Statistical analysis. Collected demographic and clinical data were managedsing Research Electronic Data Capture (REDCap) electronic data capture toolsosted at the University of Utah, which is a secure, web-based application designedo support data capture for research studies (Harris et al., 2009). Analysis of varianceANOVA) was used for between-group comparisons involving continuous demo-raphic data. Generalized estimating equations regression modeling was used tovaluate group differences in metabolite levels controlling for age, sex, educationevel, and tissue partial volume effects (i.e. gray matter, white matter, CSF). Fisher’sxact test compared groups on categorical variables. To determine the clinical rel-vance of metabolite changes in METH users, robust multiple regression analysesWhite, 1980) were performed to assess the relationship between PCr levels andifetime amount of METH use or days of abstinence of METH. Partial correlationoefficients were calculated covarying for age and sex. Models with all pair-wisend three-way interactions between covariates were first considered, and inter-ctions not significant were removed from the models. Statistical significance wasefined at an alpha level of p = .05, two-tailed. Stata for Unix, version 12.1 (StataCorp,ollege Station, TX) was used for all computations.
. Results
.1. Demographics and clinical characteristics
There were no significant differences between the METH and theC groups in terms of age (32.8 ± 6.7 vs. 31.1 ± 7.5, respectively,
72 = 0.95, p = .35) and sex (45% vs. 47% female ratio, respectively,isher’s exact test, p = .5). The METH users, however, had lower
ducation level than the HC subjects although this difference wasodest (12.7 ± 1.8 vs. 13.9 ± 1.3, respectively, p < .05). Fifty METHsers met lifetime METH dependence DSM-IV criteria and one par-icipant was diagnosed with METH abuse. Table 1 presents study
’s exact test.
subject characteristics and other demographics in detail. Table 2presents METH and other drug use history for all METH users.
3.2. Group differences in metabolite levels
phetamine (METH, n = 51) dependent subjects compared to healthy controls (HC,n = 23). Phosphocreatine levels were significantly reduced in METH users comparedto HC (p < .001). There were no significant differences in �-NTP levels between theHC and METH groups. Error bars represent 95% confidence intervals. * Statisticallysignificant difference. Abbreviations: TPP, total pool of exchangeable phosphate.
Y.-H. Sung et al. / Drug and Alcohol Dependence 129 (2013) 102– 109 105
Table 2Profiles of drug use history.
Substance Total numberof subjects
Mean age offirst use (years)
Mean amount ofsubstance per day
Frequency used Total number of subjectscurrently using
Total lifetimeamounta
Alcohol 51 13 65 ounces Occasionally to Weekly 17 34 drink-yearsb
Amphetamine 26 19 Varies Occasionally 1 VariesMethamphetamine 51 22 1.81 g Daily 30 1923 gCocaine 48 20 1.49 g Occasionally 1 1656 gOpiates 22 25 0.69 g Occasionally 0 351 gHallucinogens 36 17 Varies Rarely to Occasionally 0 VariesCannabis 50 14.5 2.42 g Weekly 12 3721 gNicotine 47 14 11 cigarettes Daily 41 10 pack-yearsc
Caffeine 50 9 28 ounces Daily 49 27 cups-yearsd
a Total lifetime drug amount was estimated using “average daily dosage × frequency × duration”.
ck/da
ma(gda
taPptsclMeSw
dbcs(
3
cpM(M
TP
D�
b Total intake calculated in drink-years (1 drink/day in 1 year).c A pack-year is defined as twenty cigarettes smoked everyday for one year (1 pad A cups-year is defined as five cups of coffee everyday for one year.
aintained after controlling for potential confounding vari-bles such as age, sex, education level, and tissue segmentationt68 = −2.99, p = .004). As there was a high collinearity betweenroup variable and alcohol amount/nicotine amount consumed, weid not conduct regression analysis adjusting for alcohol, nicotinend caffeine.
To ensure that decreased PCr/TPP did not result from increasedotal TPP values, we repeated our statistical analysis using �-NTPs a denominator, with findings remaining the same. SpecificallyCr/�-NTP levels were significantly decreased in METH users com-ared to the HC subjects (t72 = −2.08, p = .04). Also, to ensure thathe MR system did not drift across the study period, absolute mea-urements (arbitrary institutional unit) of total phosphorus wereompared between the groups, with no group differences in TPPevels (p = .10). Further, to assess test-retest reliability of our 31P-
RS protocol, we scanned nine healthy subjects twice using thexact same protocol. The intraclass correlation coefficient (ICC;hrout and Fleiss, 1979) for PCr levels was calculated to be 0.72,hich suggests good reliability.
Other metabolite levels such as phosphomonoester, phospho-iester, and inorganic phosphate were not significantly differentetween the groups (see Table 3). We did not find a statisti-ally significant difference in PCr/TPP levels in other brain regionsuch as temporoparietal lobe (t72 = −1.71, p = .10) and occipital lobet72 = 1.60, p = .12) between METH and HC subjects.
.3. PCr levels in female vs. male METH users
Frontal PCr/TPP levels in female METH users were signifi-antly lower (5.3%) than those in male METH users (t49 = −2.06,
= .04, Fig. 3). Female METH users reported lower amounts of dailyETH use (1.1 ± 1.0 g) compared to male METH users (1.4 ± 1.7 g)
t49 = −0.75, p = .46). After adjusting for the daily amount of dailyETH use, the significance for between-group gender difference of
able 3hosphorus metabolite levels in the frontal lobe.
Metabolitesa Methamphetamine
Total (n = 51) Female Male
PCr .212 (.021)* .205 (.019)** .217 (.021)**
�-NTP .192 (.022) .195 (.025) .189 (.019)
PME .215 (.037) .220 (.041) .210 (.033)
PDE .286 (.036) .291 (.036) .281 (.037)
Pi .076 (.012) .080 (.012) .074 (.013)
ata are expressed with mean (SD). For other metabolites, there were neither group diff-NTP, beta-nucleoside triphosphate; PME, phosphomonoester; PDE, phosphodiester; Pia Metabolite ratios over total pool of exchangeable phosphate.* Statistically significant PCr differences between methamphetamine and healthy grou
** Significant PCr differences between female and male methamphetamine users (p = .04
y in 1 year).
PCr/TPP levels was maintained (t48 = −2.41, p = .02). In the healthysubjects, there were no significant gender differences in either PCr(t21 = −0.90, p = .38), �-NTP (t21 = −0.90, p = .38), or other metabo-lites (Table 3).
3.4. Relationship between frontal lobe PCr levels and lifetimeMETH amount
There was significant relationship between frontal lobe PCrlevels and total amount of lifetime METH use (Fig. 4), before(t49 = −3.17, p = .003), and after (t47 = −2.87, p = .006) controlling forage and education.
3.5. Relationship between frontal lobe PCr levels and duration ofabstinence from METH
There was a trend toward a negative relationship betweenfrontal lobe PCr levels and duration of METH abstinence but thiswas not statistically significant (t49 = −1.49, p = .14).
4. Discussion
In this study, we found that METH users have abnormal PCr lev-els in the frontal lobe compared with the HC subjects. To the best ofour knowledge, this is the first 31P-MRS study reporting abnormalbrain cellular energetics in human METH users.
Our findings are consistent with the prior neuroimaging reportsregarding metabolic hypofrontality in METH users. For example,18F-fluorodeoxyglucose (FDG) PET studies have noted lower FDGuptake in the frontal lobe and cingulate (Kim et al., 2005; London
et al., 2004, 2005) and in the striatum (Volkow et al., 2001a). Also,single photon emission computed tomography (SPECT) studieshave reported decreased frontal blood flow in METH users (Hwanget al., 2006).Healthy p-Value
Total (n = 23) Female Male
.230 (.016)* .225 (.018) .231 (.014) .001*
.189 (.020) .187 (.027) .190 (.013) .58
.218 (.022) .215 (.027) .221 (.018) .68
.292 (.034) .285 (.034) .298 (.034) .47
.074 (.012) .073 (.013) .075 (.012) .38
erences nor within-group gender differences. Abbreviations: PCr, phosphocreatine;, inorganic phosphate.
p (p = .001).).
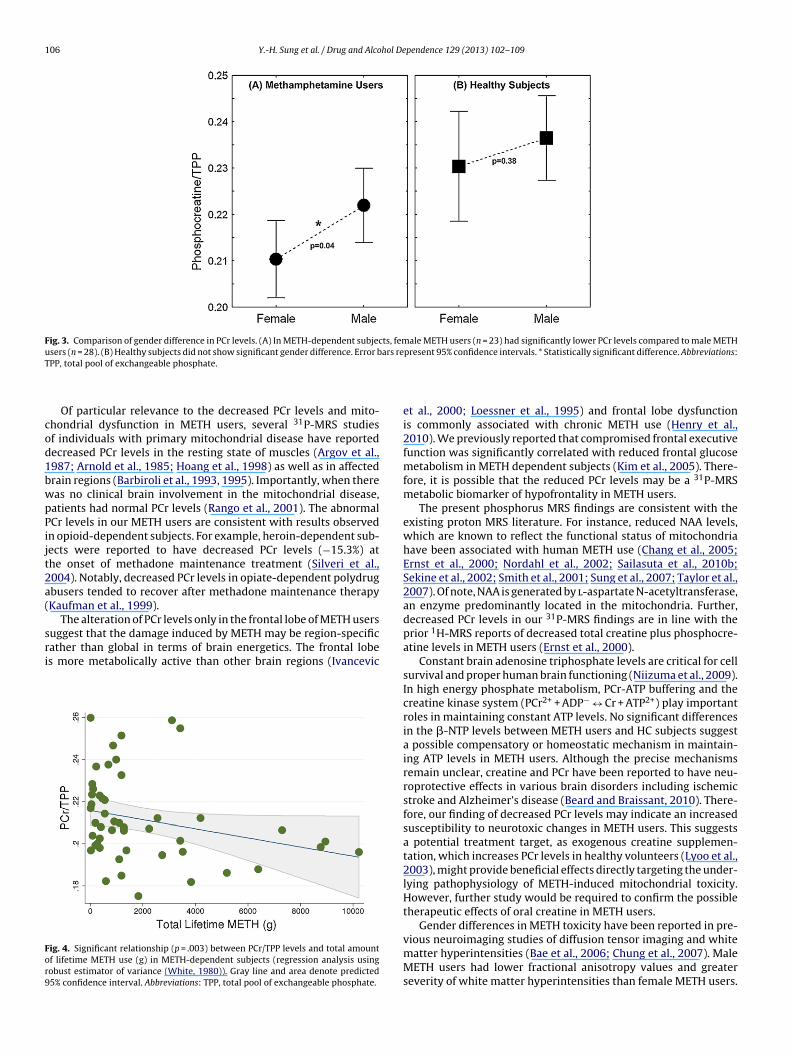
106 Y.-H. Sung et al. / Drug and Alcohol Dependence 129 (2013) 102– 109
Fig. 3. Comparison of gender difference in PCr levels. (A) In METH-dependent subjects, female METH users (n = 23) had significantly lower PCr levels compared to male METHu ars reT
cod1bwpPijt2a(
sri
For9
sers (n = 28). (B) Healthy subjects did not show significant gender difference. Error bPP, total pool of exchangeable phosphate.
Of particular relevance to the decreased PCr levels and mito-hondrial dysfunction in METH users, several 31P-MRS studiesf individuals with primary mitochondrial disease have reportedecreased PCr levels in the resting state of muscles (Argov et al.,987; Arnold et al., 1985; Hoang et al., 1998) as well as in affectedrain regions (Barbiroli et al., 1993, 1995). Importantly, when thereas no clinical brain involvement in the mitochondrial disease,atients had normal PCr levels (Rango et al., 2001). The abnormalCr levels in our METH users are consistent with results observedn opioid-dependent subjects. For example, heroin-dependent sub-ects were reported to have decreased PCr levels (−15.3%) athe onset of methadone maintenance treatment (Silveri et al.,004). Notably, decreased PCr levels in opiate-dependent polydrugbusers tended to recover after methadone maintenance therapyKaufman et al., 1999).
The alteration of PCr levels only in the frontal lobe of METH usersuggest that the damage induced by METH may be region-specificather than global in terms of brain energetics. The frontal lobes more metabolically active than other brain regions (Ivancevic
ig. 4. Significant relationship (p = .003) between PCr/TPP levels and total amountf lifetime METH use (g) in METH-dependent subjects (regression analysis usingobust estimator of variance (White, 1980)). Gray line and area denote predicted5% confidence interval. Abbreviations: TPP, total pool of exchangeable phosphate.
present 95% confidence intervals. * Statistically significant difference. Abbreviations:
et al., 2000; Loessner et al., 1995) and frontal lobe dysfunctionis commonly associated with chronic METH use (Henry et al.,2010). We previously reported that compromised frontal executivefunction was significantly correlated with reduced frontal glucosemetabolism in METH dependent subjects (Kim et al., 2005). There-fore, it is possible that the reduced PCr levels may be a 31P-MRSmetabolic biomarker of hypofrontality in METH users.
The present phosphorus MRS findings are consistent with theexisting proton MRS literature. For instance, reduced NAA levels,which are known to reflect the functional status of mitochondriahave been associated with human METH use (Chang et al., 2005;Ernst et al., 2000; Nordahl et al., 2002; Sailasuta et al., 2010b;Sekine et al., 2002; Smith et al., 2001; Sung et al., 2007; Taylor et al.,2007). Of note, NAA is generated by l-aspartate N-acetyltransferase,an enzyme predominantly located in the mitochondria. Further,decreased PCr levels in our 31P-MRS findings are in line with theprior 1H-MRS reports of decreased total creatine plus phosphocre-atine levels in METH users (Ernst et al., 2000).
Constant brain adenosine triphosphate levels are critical for cellsurvival and proper human brain functioning (Niizuma et al., 2009).In high energy phosphate metabolism, PCr-ATP buffering and thecreatine kinase system (PCr2+ + ADP− ↔ Cr + ATP2+) play importantroles in maintaining constant ATP levels. No significant differencesin the �-NTP levels between METH users and HC subjects suggesta possible compensatory or homeostatic mechanism in maintain-ing ATP levels in METH users. Although the precise mechanismsremain unclear, creatine and PCr have been reported to have neu-roprotective effects in various brain disorders including ischemicstroke and Alzheimer’s disease (Beard and Braissant, 2010). There-fore, our finding of decreased PCr levels may indicate an increasedsusceptibility to neurotoxic changes in METH users. This suggestsa potential treatment target, as exogenous creatine supplemen-tation, which increases PCr levels in healthy volunteers (Lyoo et al.,2003), might provide beneficial effects directly targeting the under-lying pathophysiology of METH-induced mitochondrial toxicity.However, further study would be required to confirm the possibletherapeutic effects of oral creatine in METH users.
Gender differences in METH toxicity have been reported in pre-
vious neuroimaging studies of diffusion tensor imaging and whitematter hyperintensities (Bae et al., 2006; Chung et al., 2007). MaleMETH users had lower fractional anisotropy values and greaterseverity of white matter hyperintensities than female METH users.
hol De
ImlbbweoMstf(TMiAMblTtiAPsbviHsc
aammPlrotPaFmrp2
4
iTim2fAwP
mi
Y.-H. Sung et al. / Drug and Alco
n the present 31P-MRS study, however, decreased PCr levels wereore prominent in females. The exact mechanism responsible for
ower PCr levels in female METH users is unclear and has yet toe elucidated. However, since lower brain levels of PCr have alsoeen associated with severe depression (Kato et al., 1992) andorse outcomes in response to antidepressant treatment (Iosifescu
t al., 2008), our findings may be related to the higher incidencef depression in female METH users (Semple et al., 2007). Indeed,ETH abuse is associated with increased risk of depression and
uicide attempts compared to rates observed in the general popula-ion (Glasner-Edwards et al., 2009). Also, METH abusers have higheremale/male ratios (46%) than heroin (11%) or cocaine (29%) abusersCohen et al., 2007; Holdcraft and Iacono, 2004; Hser et al., 2008).herefore, the relatively large reduction of PCr levels in femaleETH users highlights the potential consequences of bioenergetic
mpairment that merit special clinical attention in female subjects.nother possible underlying mechanism for lower PCr in femaleETH users may be related to the effect of gonadal hormones on
rain PCr levels. For example, estrogen has been reported to stimu-ate brain-specific cytosolic creatine kinase activity (Kaye, 1983).herefore, the higher estrogen levels in females may contributeo the decreased PCr levels through the forward (i.e. regenerat-ng ATP) creatine kinase activity in the PCr-ATP buffer system.lthough it was not statistically significant, we found slightly lowerCr levels in female healthy subjects. Interestingly, a recent PETtudy demonstrated decreased prefrontal metabolism in femalesut not in male cocaine users (Volkow et al., 2011). This obser-ation suggests that gonadal hormones may be associated withncreased risk of abnormal high energy phosphate metabolism.owever, further study will be required to clarify the relation-
hip between hormone levels and brain high energy phosphatehanges.
The significant relationship between total amount of METH usend the PCr levels illustrates that heavy METH users might manifests an abnormality in high energy phosphates in a dose-responsiveanner. These findings raise the possibility that heavy METH usersay benefit from agents which facilitate the recovery of frontal lobe
Cr levels. A non-significant negative association between frontalobe PCr levels and duration of METH abstinence suggests thatecovery of PCr levels may not occur during the moderate levelsf simple abstinence (95% confidence limits: 26–50 days). Indeed,he negative trend suggests that longer abstinence may result inCr levels that remain low or even decline further. These findingsre consistent with persistently lower striatal, but not thalamic,DG metabolism in METH users compared to HC even after 12–17onths of abstinence (Wang et al., 2004), as well as significantly
educed striatal dopamine transporter levels in METH users com-ared to HC even after 11 months of abstinence (Volkow et al.,001b).
.1. Study limitations
When considering our findings, several factors should be takennto consideration. The 31P-MRS data are reported by ratio toPP with no absolute 31P metabolite concentration. Although thiss a standard practice in the MRS literature, decreased PCr/TPP
ay have originated from increased TPP levels (Jansen et al.,006; Jayakumar et al., 2010). However, even when we use a dif-erent denominator such as �-NTP, we observe similar results.lso, metabolite ratio measures have been compared favorablyith absolute measures (Klunk et al., 1994). Therefore, decreased
Cr/TPP ratios are likely to reflect decreased PCr levels.Female subjects were not scanned at the same phase of the
enstrual cycle in the present study. Since prior published stud-es suggest hormonal levels are related to subjective reports of
pendence 129 (2013) 102– 109 107
stimulation after d-amphetamine administration (Reed et al.,2010; White et al., 2002), phosphorus metabolite levels may alsofluctuate across the menstrual cycle in the females, placing thelack of the control of menstrual phase as a limitation of this study.However, since the female subjects were randomly scanned forboth controls and METH users, we believe that the variance in PCrlevels was similar between groups, and hence the menstrual phasevariation did not significantly affect our findings.
Multiple comparisons were performed in statistical analysis forour metabolites of interest, PCr and �-NTP. As we used an a priorihypothesis in our research design, corrections for multiple com-parisons were not attempted. Therefore, readers should be awareof possible type I errors arising from our analysis.
The METH subject enrollment criteria involved the inclusion ofsubjects who identified METH as their drug of choice. Therefore, thecharacteristics of our cohort may not represent general METH usersand our findings may not be generalized to other METH-dependentpopulations. On the other hand, all METH users abused other drugsto some degree and decreased PCr levels in METH users may be, inpart, due to polysubstance abuse. Self report of drug use history inMETH subjects might have resulted in some degree of uncertaintyincluding underreport of drug use behaviors.
Because the location of our 2D-CSI grid was immediately supe-rior to the AC-PC line, we could not include subgenual cingulateor cerebellum as regions-of-interest to evaluate whether thesebrain regions have abnormalities in high energy phosphorusmetabolism. Since the subgenual cingulate is reported to beassociated with depression (Drevets et al., 2008; Mayberg et al.,2005), a study design including this brain region will be importantin future studies.
5. Conclusion
Overall, the present study provides evidence of altered high-energy phosphate metabolism in METH users. Our findings suggestthat (1) METH use is associated with dose-dependent decreasesin high-energy PCr levels, which may imply a decreased ener-getic buffer due to mitochondrial dysfunction; and (2) femaleMETH users may be more vulnerable than male METH users interms of high-energy phosphate neurochemistry. Further study iswarranted to explore the relationship between the altered phos-phorus metabolism and cognitive as well as psychiatric symptoms.Also, a longitudinal study design would shed light on within-subject changes in high energy phosphate metabolism followinglong-term abstinence and/or treatment. Efforts to characterize therelationship between the changes in high energy phosphates andpsychiatric symptomatology have the potential to generate noveltreatment strategies for METH toxicity.
Role of funding source
This study was supported by funding from NIH 1R01DA027135(PFR). The funding agency had no further role in study design; inthe collection, analysis and interpretation of data; in the writing ofthe report; or in the decision to submit the paper for publication.
Contributors
Authors Yurgelun-Todd, Sung, and Renshaw designed the studyand wrote the protocol. Author Sung wrote the first draft of
the manuscript and undertook the statistical analysis. AuthorsShi, Kondo, Lundberg, McGlade, Hellem, Huber, Fiedler, Harrell,Nickerson, Kim and Jeong participated in data collection and inter-pretation. Authors Renshaw and Yurgelun-Todd wrote the final
1 hol De
da
C
na
R
A
A
A
A
B
B
B
B
B
B
B
B
C
C
C
C
C
C
D
D
D
D
E
08 Y.-H. Sung et al. / Drug and Alco
raft of the manuscript. All authors contributed to and havepproved the final manuscript.
onflict of interest
Dr. Renshaw is a consultant for Kyowa Hakko and Ridge Diag-ostics. Dr. Yurgelun-Todd is a consultant for Eli Lilly, and Novartis,nd has research support from Kyowa Hakko.
eferences
mess, P.N., Penrice, J., Cady, E.B., Lorek, A., Wylezinska, M., Cooper, C.E., D’Souza, P.,Tyszczuk, L., Thoresen, M., Edwards, A.D., Wyatt, J.S., Reynolds, E.O., 1997. Mildhypothermia after severe transient hypoxia-ischemia reduces the delayed risein cerebral lactate in the newborn piglet. Pediatr. Res. 41, 803–808.
nker, J.J., Carroll, M.E., 2011. Females are more vulnerable to drug abuse than males:evidence from preclinical studies and the role of ovarian hormones. Curr. Top.Behav. Neurosci. 8, 73–96.
rgov, Z., Bank, W.J., Maris, J., Peterson, P., Chance, B., 1987. Bioenergetic hetero-geneity of human mitochondrial myopathies: phosphorus magnetic resonancespectroscopy study. Neurology 37, 257–262.
rnold, D.L., Taylor, D.J., Radda, G.K., 1985. Investigation of human mitochondrialmyopathies by phosphorus magnetic resonance spectroscopy. Ann. Neurol. 18,189–196.
ae, S.C., Lyoo, I.K., Sung, Y.H., Yoo, J., Chung, A., Yoon, S.J., Kim, D.J., Hwang, J.,Kim, S.J., Renshaw, P.F., 2006. Increased white matter hyperintensities in malemethamphetamine abusers. Drug Alcohol Depend. 81, 83–88.
arbiroli, B., Montagna, P., Cortelli, P., Iotti, S., Lodi, R., Barboni, P., Monari, L., Lugaresi,E., Frassineti, C., Zaniol, P., 1995. Defective brain and muscle energy metabolismshown by in vivo 31P magnetic resonance spectroscopy in nonaffected carriersof 11778 mtDNA mutation. Neurology 45, 1364–1369.
arbiroli, B., Montagna, P., Martinelli, P., Lodi, R., Iotti, S., Cortelli, P., Funicello, R.,Zaniol, P., 1993. Defective brain energy metabolism shown by in vivo 31P MRspectroscopy in 28 patients with mitochondrial cytopathies. J. Cereb. Blood FlowMetab. 13, 469–474.
eard, E., Braissant, O., 2010. Synthesis and transport of creatine in the CNS: impor-tance for cerebral functions. J. Neurochem. 115, 297–313.
lumberg, R.M., Taylor, D.L., Yue, X., Aguan, K., McKenzie, J., Cady, E.B., Weiner, C.P.,Mehmet, H., Edwards, A.D., 1999. Increased nitric oxide synthesis is not involvedin delayed cerebral energy failure following focal hypoxic-ischemic injury to thedeveloping brain. Pediatr. Res. 46, 224–231.
rown, J.M., Quinton, M.S., Yamamoto, B.K., 2005. Methamphetamine-induced inhi-bition of mitochondrial complex. II. Roles of glutamate and peroxynitrite. J.Neurochem. 95, 429–436.
urrows, K.B., Gudelsky, G., Yamamoto, B.K., 2000a. Rapid and transientinhibition of mitochondrial function following methamphetamine or 3,4-methylenedioxymethamphetamine administration. Eur. J. Pharmacol. 398,11–18.
urrows, K.B., Nixdorf, W.L., Yamamoto, B.K., 2000b. Central administration ofmethamphetamine synergizes with metabolic inhibition to deplete striatalmonoamines. J. Pharmacol. Exp. Ther. 292, 853–860.
ady, E.B., Iwata, O., Bainbridge, A., Wyatt, J.S., Robertson, N.J., 2008. Phosphorusmagnetic resonance spectroscopy 2 h after perinatal cerebral hypoxia-ischemiaprognosticates outcome in the newborn piglet. J. Neurochem. 107, 1027–1035.
arroll, M.E., Lynch, W.J., Roth, M.E., Morgan, A.D., Cosgrove, K.P., 2004. Sex andestrogen influence drug abuse. Trends Pharmacol. Sci. 25, 273–279.
hang, L., Ernst, T., Speck, O., Grob, C.S., 2005. Additive effects of HIV and chronicmethamphetamine use on brain metabolite abnormalities. Am. J. Psychiatry 162,361–369.
hung, A., Lyoo, I.K., Kim, S.J., Hwang, J., Bae, S.C., Sung, Y.H., Sim, M.E., Song,I.C., Kim, J., Chang, K.H., Renshaw, P.F., 2007. Decreased frontal white-matterintegrity in abstinent methamphetamine abusers. Int. J. Neuropsychopharma-col. 10, 765–775.
ohen, J.B., Greenberg, R., Uri, J., Halpin, M., Zweben, J.E., 2007. Women withmethamphetamine dependence: research on etiology and treatment. J. Psy-choactive Drugs Suppl. 4, 347–351.
unha-Oliveira, T., Rego, A.C., Cardoso, S.M., Borges, F., Swerdlow, R.H., Macedo, T.,de Oliveira, C.R., 2006. Mitochondrial dysfunction and caspase activation in ratcortical neurons treated with cocaine or amphetamine. Brain Res. 1089, 44–54.
handapani, K.M., Brann, D.W., 2002. Protective effects of estrogen and selectiveestrogen receptor modulators in the brain. Biol. Reprod. 67, 1379–1385.
luzen, D.E., Liu, B., 2008. Gender differences in methamphetamine use andresponses: a review. Gend. Med. 5, 24–35.
older, M., Wendt, S., Wallimann, T., 2001. Mitochondrial creatine kinase in contactsites: interaction with porin and adenine nucleotide translocase, role in perme-ability transition and sensitivity to oxidative damage. Biol. Signals Recept. 10,93–111.
revets, W.C., Savitz, J., Trimble, M., 2008. The subgenual anterior cingulate cortexin mood disorders. CNS Spectrosc. 13, 663–681.
rnst, T., Chang, L., Leonido-Yee, M., Speck, O., 2000. Evidence for long-term neuro-toxicity associated with methamphetamine abuse: a 1H MRS study. Neurology54, 1344–1349.
pendence 129 (2013) 102– 109
Glasner-Edwards, S., Marinelli-Casey, P., Hillhouse, M., Ang, A., Mooney, L.J., Raw-son, R., Methamphetamine Treatment Project Corporate, A., 2009. Depressionamong methamphetamine users: association with outcomes from the Metham-phetamine Treatment Project at 3-year follow-up. J. Nerv. Ment. Dis. 197,225–231.
Harris, P.A., Taylor, R., Thielke, R., Payne, J., Gonzalez, N., Conde, J.G., 2009. Researchelectronic data capture (REDCap) – a metadata-driven methodology and work-flow process for providing translational research informatics support. J. Biomed.Inform. 42, 377–381.
Henry, B.L., Minassian, A., Perry, W., 2010. Effect of methamphetamine dependenceon everyday functional ability. Addict. Behav. 35, 593–598.
Hoang, T.Q., Bluml, S., Dubowitz, D.J., Moats, R., Kopyov, O., Jacques, D., Ross, B.D.,1998. Quantitative proton-decoupled 31P MRS and 1H MRS in the evaluation ofHuntington’s and Parkinson’s diseases. Neurology 50, 1033–1040.
Holdcraft, L.C., Iacono, W.G., 2004. Cross-generational effects on gender differencesin psychoactive drug abuse and dependence. Drug Alcohol Depend. 74, 147–158.
Hser, Y.I., Evans, E., Huang, Y.C., 2005. Treatment outcomes among women and menmethamphetamine abusers in California. J. Subst. Abuse Treat. 28, 77–85.
Hser, Y.I., Huang, D., Brecht, M.L., Li, L., Evans, E., 2008. Contrasting trajectories ofheroin, cocaine, and methamphetamine use. J. Addict. Dis. 27, 13–21.
Hwang, J., Lyoo, I.K., Kim, S.J., Sung, Y.H., Bae, S., Cho, S.N., Lee, H.Y., Lee, D.S., Renshaw,P.F., 2006. Decreased cerebral blood flow of the right anterior cingulate cortexin long-term and short-term abstinent methamphetamine users. Drug AlcoholDepend. 82, 177–181.
Iosifescu, D.V., Bolo, N.R., Nierenberg, A.A., Jensen, J.E., Fava, M., Renshaw, P.F., 2008.Brain bioenergetics and response to triiodothyronine augmentation in majordepressive disorder. Biol. Psychiatry 63, 1127–1134.
Ivancevic, V., Alavi, A., Souder, E., Mozley, P.D., Gur, R.E., Benard, F., Munz, D.L., 2000.Regional cerebral glucose metabolism in healthy volunteers determined by flu-ordeoxyglucose positron emission tomography: appearance and variance in thetransaxial, coronal, and sagittal planes. Clin. Nucl. Med. 25, 596–602.
Iwata, O., Iwata, S., Bainbridge, A., De Vita, E., Matsuishi, T., Cady, E.B., Robertson,N.J., 2008. Supra- and sub-baseline phosphocreatine recovery in developingbrain after transient hypoxia-ischaemia: relation to baseline energetics, insultseverity and outcome. Brain 131, 2220–2226.
Iyo, M., Sekine, Y., Mori, N., 2004. Neuromechanism of developing metham-phetamine psychosis: a neuroimaging study. Ann. N. Y. Acad. Sci. 1025, 288–295.
Jansen, J.F., Backes, W.H., Nicolay, K., Kooi, M.E., 2006. 1H MR spectroscopy of thebrain: absolute quantification of metabolites. Radiology 240, 318–332.
Jayakumar, P.N., Gangadhar, B.N., Venkatasubramanian, G., Desai, S., Velayudhan, L.,Subbakrishna, D., Keshavan, M.S., 2010. High energy phosphate abnormalitiesnormalize after antipsychotic treatment in schizophrenia: a longitudinal 31PMRS study of basal ganglia. Psychiatry Res. 181, 237–240.
Kalechstein, A.D., Newton, T.F., Longshore, D., Anglin, M.D., van Gorp, W.G., Gawin,F.H., 2000. Psychiatric comorbidity of methamphetamine dependence in a foren-sic sample. J. Neuropsychiatry Clin. Neurosci. 12, 480–484.
Kato, T., Takahashi, S., Shioiri, T., Inubushi, T., 1992. Brain phosphorous metabolismin depressive disorders detected by phosphorus-31 magnetic resonance spec-troscopy. J. Affect. Disord. 26, 223–230.
Kaufman, M.J., Pollack, M.H., Villafuerte, R.A., Kukes, T.J., Rose, S.L., Mendelson, J.H.,Cohen, B.M., Renshaw, P.F., 1999. Cerebral phosphorus metabolite abnormalitiesin opiate-dependent polydrug abusers in methadone maintenance. PsychiatryRes. 90, 143–152.
Kaye, A.M., 1983. Enzyme induction by estrogen. J. Steroid Biochem. 19, 33–40.Kim, S.J., Lyoo, I.K., Hwang, J., Sung, Y.H., Lee, H.Y., Lee, D.S., Jeong, D.U., Renshaw, P.F.,
2005. Frontal glucose hypometabolism in abstinent methamphetamine users.Neuropsychopharmacology 30, 1383–1391.
Klongpanichapak, S., Govitrapong, P., Sharma, S.K., Ebadi, M., 2006. Attenuation ofcocaine and methamphetamine neurotoxicity by coenzyme Q10. Neurochem.Res. 31, 303–311.
Klunk, W.E., Xu, C.J., Panchalingam, K., McClure, R.J., Pettegrew, J.W., 1994. Analysisof magnetic resonance spectra by mole percent: comparison to absolute units.Neurobiol. Aging 15, 133–140.
Kokoshka, J.M., Metzger, R.R., Wilkins, D.G., Gibb, J.W., Hanson, G.R., Fleckenstein,A.E., 1998. Methamphetamine treatment rapidly inhibits serotonin, but not glu-tamate, transporters in rat brain. Brain Res. 799, 78–83.
Loessner, A., Alavi, A., Lewandrowski, K.U., Mozley, D., Souder, E., Gur, R.E., 1995.Regional cerebral function determined by FDG-PET in healthy volunteers: nor-mal patterns and changes with age. J. Nucl. Med. 36, 1141–1149.
London, E.D., Berman, S.M., Voytek, B., Simon, S.L., Mandelkern, M.A., Mon-terosso, J., Thompson, P.M., Brody, A.L., Geaga, J.A., Hong, M.S., Hayashi, K.M.,Rawson, R.A., Ling, W., 2005. Cerebral metabolic dysfunction and impaired vig-ilance in recently abstinent methamphetamine abusers. Biol. Psychiatry 58,770–778.
London, E.D., Simon, S.L., Berman, S.M., Mandelkern, M.A., Lichtman, A.M., Bramen,J., Shinn, A.K., Miotto, K., Learn, J., Dong, Y., Matochik, J.A., Kurian, V., Newton, T.,Woods, R., Rawson, R., Ling, W., 2004. Mood disturbances and regional cerebralmetabolic abnormalities in recently abstinent methamphetamine abusers. Arch.Gen. Psychiatry 61, 73–84.
Lubman, D.I., Yucel, M., Pantelis, C., 2004. Addiction, a condition of compulsivebehaviour? Neuroimaging and neuropsychological evidence of inhibitory dys-
regulation. Addiction 99, 1491–1502.Lyoo, I.K., Kong, S.W., Sung, S.M., Hirashima, F., Parow, A., Hennen, J., Cohen, B.M.,Renshaw, P.F., 2003. Multinuclear magnetic resonance spectroscopy of high-energy phosphate metabolites in human brain following oral supplementationof creatine-monohydrate. Psychiatry Res. 123, 87–100.

hol De
M
M
N
N
N
O
O
P
R
R
R
R
R
S
S
S
S
Y.-H. Sung et al. / Drug and Alco
ayberg, H.S., Lozano, A.M., Voon, V., McNeely, H.E., Seminowicz, D., Hamani,C., Schwalb, J.M., Kennedy, S.H., 2005. Deep brain stimulation for treatment-resistant depression. Neuron 45, 651–660.
offett, J.R., Ross, B., Arun, P., Madhavarao, C.N., Namboodiri, A.M., 2007. N-Acetylaspartate in the CNS: from neurodiagnostics to neurobiology. Prog.Neurobiol. 81, 89–131.
aressi, A., Couturier, C., Devos, J.M., Janssen, M., Mangeat, C., de Beer, R., Graveron-Demilly, D., 2001. Java-based graphical user interface for the MRUI quantitationpackage. MAGMA 12, 141–152.
iizuma, K., Endo, H., Chan, P.H., 2009. Oxidative stress and mitochondrial dysfunc-tion as determinants of ischemic neuronal death and survival. J. Neurochem. 109(1), 133–138.
ordahl, T.E., Salo, R., Possin, K., Gibson, D.R., Flynn, N., Leamon, M., Galloway, G.P.,Pfefferbaum, A., Spielman, D.M., Adalsteinsson, E., Sullivan, E.V., 2002. Low N-acetyl-aspartate and high choline in the anterior cingulum of recently abstinentmethamphetamine-dependent subjects: a preliminary proton MRS study mag-netic resonance spectroscopy. Psychiatry Res. 116, 43–52.
h, J.S., Lyoo, I.K., Sung, Y.H., Hwang, J., Kim, J., Chung, A., Park, K.S., Kim, S.J., Ren-shaw, P.F., Song, I.C., 2005. Shape changes of the corpus callosum in abstinentmethamphetamine users. Neurosci. Lett. 384, 76–81.
liveira, M.T., Rego, A.C., Morgadinho, M.T., Macedo, T.R., Oliveira, C.R., 2002. Toxiceffects of opioid and stimulant drugs on undifferentiated PC12 cells. Ann. N. Y.Acad. Sci. 965, 487–496.
atel, T.B., Clark, J.B., 1979. Synthesis of N-acetyl-l-aspartate by rat brain mitochon-dria and its involvement in mitochondrial/cytosolic carbon transport. Biochem.J. 184, 539–546.
ango, M., Bozzali, M., Prelle, A., Scarlato, G., Bresolin, N., 2001. Brain activationin normal subjects and in patients affected by mitochondrial disease withoutclinical central nervous system involvement: a phosphorus magnetic resonancespectroscopy study. J. Cereb. Blood Flow Metab. 21, 85–91.
eed, S.C., Levin, F.R., Evans, S.M., 2010. The effects of progesterone pretreatment onthe response to oral d-amphetamine in Women. Horm. Behav. 58, 533–543.
enshaw, P.F., Parow, A.M., Hirashima, F., Ke, Y., Moore, C.M., Frederick Bde, B.,Fava, M., Hennen, J., Cohen, B.M., 2001. Multinuclear magnetic resonance spec-troscopy studies of brain purines in major depression. Am. J. Psychiatry 158,2048–2055.
icaurte, G.A., Schuster, C.R., Seiden, L.S., 1980. Long-term effects of repeated methy-lamphetamine administration on dopamine and serotonin neurons in the ratbrain: a regional study. Brain Res. 193, 153–163.
obinson, T.E., Berridge, K.C., 1993. The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res. Brain Res. Rev. 18, 247–291.
ailasuta, N., Abulseoud, O., Harris, K.C., Ross, B.D., 2010a. Glial dysfunction in absti-nent methamphetamine abusers. J. Cereb. Blood Flow Metab. 30, 950–960.
ailasuta, N., Abulseoud, O., Hernandez, M., Haghani, P., Ross, B.D., 2010b. Metabolicabnormalities in abstinent methamphetamine dependent subjects. Subst. Abuse2010, 9–20.
chlattner, U., Tokarska-Schlattner, M., Wallimann, T., 2006. Mitochondrial cre-atine kinase in human health and disease. Biochim. Biophys. Acta 1762,
164–180.ekine, Y., Minabe, Y., Kawai, M., Suzuki, K., Iyo, M., Isoda, H., Sakahara, H., Ashby Jr.,C.R., Takei, N., Mori, N., 2002. Metabolite alterations in basal ganglia associatedwith methamphetamine-related psychiatric symptoms. A proton MRS study.Neuropsychopharmacology 27, 453–461.
pendence 129 (2013) 102– 109 109
Semple, S.J., Zians, J., Strathdee, S.A., Patterson, T.L., 2007. Psychosocial and behav-ioral correlates of depressed mood among female methamphetamine users. J.Psychoactive Drugs Suppl. 4, 353–366.
Shrout, P.E., Fleiss, J.L., 1979. Intraclass correlations: uses in assessing rater reliabil-ity. Psychol. Bull. 86, 420–428.
Silveri, M.M., Pollack, M.H., Diaz, C.I., Nassar, L.E., Mendelson, J.H., Yurgelun-Todd,D.A., Renshaw, P.F., Kaufman, M.J., 2004. Cerebral phosphorus metabolite andtransverse relaxation time abnormalities in heroin-dependent subjects at onsetof methadone maintenance treatment. Psychiatry Res. 131, 217–226.
Smith, L.M., Chang, L., Yonekura, M.L., Grob, C., Osborn, D., Ernst, T., 2001. Brain pro-ton magnetic resonance spectroscopy in children exposed to methamphetaminein utero. Neurology 57, 255–260.
Sung, Y.H., Cho, S.C., Hwang, J., Kim, S.J., Kim, H., Bae, S., Kim, N., Chang, K.H., Daniels,M., Renshaw, P.F., Lyoo, I.K., 2007. Relationship between N-acetyl-aspartate ingray and white matter of abstinent methamphetamine abusers and their historyof drug abuse: a proton magnetic resonance spectroscopy study. Drug AlcoholDepend. 88, 28–35.
Taylor, M.J., Schweinsburg, B.C., Alhassoon, O.M., Gongvatana, A., Brown, G.G.,Young-Casey, C., Letendre, S.L., Grant, I., Group, H., 2007. Effects of humanimmunodeficiency virus and methamphetamine on cerebral metabolites mea-sured with magnetic resonance spectroscopy. J. Neurovirol. 13, 150–159.
Vanhamme, L., van den Boogaart, A., Van Huffel, S., 1997. Improved method foraccurate and efficient quantification of MRS data with use of prior knowledge.J. Magn. Reson. 129, 35–43.
Volkow, N.D., Chang, L., Wang, G.J., Fowler, J.S., Franceschi, D., Sedler, M.J., Gatley,S.J., Hitzemann, R., Ding, Y.S., Wong, C., Logan, J., 2001a. Higher cortical andlower subcortical metabolism in detoxified methamphetamine abusers. Am. J.Psychiatry 158, 383–389.
Volkow, N.D., Chang, L., Wang, G.J., Fowler, J.S., Leonido-Yee, M., Franceschi, D.,Sedler, M.J., Gatley, S.J., Hitzemann, R., Ding, Y.S., Logan, J., Wong, C., Miller,E.N., 2001b. Association of dopamine transporter reduction with psychomotorimpairment in methamphetamine abusers. Am. J. Psychiatry 158, 377–382.
Volkow, N.D., Tomasi, D., Wang, G.J., Fowler, J.S., Telang, F., Goldstein, R.Z., Alia-Klein,N., Wong, C., 2011. Reduced metabolism in brain “control networks” followingcocaine-cues exposure in female cocaine abusers. PLoS One 6, e16573.
Wallimann, T., Dolder, M., Schlattner, U., Eder, M., Hornemann, T., O’Gorman, E.,Ruck, A., Brdiczka, D., 1998. Some new aspects of creatine kinase (CK): com-partmentation, structure, function and regulation for cellular and mitochondrialbioenergetics and physiology. Biofactors 8, 229–234.
Wang, G.J., Volkow, N.D., Chang, L., Miller, E., Sedler, M., Hitzemann, R., Zhu, W.,Logan, J., Ma, Y., Fowler, J.S., 2004. Partial recovery of brain metabolism inmethamphetamine abusers after protracted abstinence. Am. J. Psychiatry 161,242–248.
White, H., 1980. A heteroskedasticity-consistent covariance matrix estimator and adirect test for heteroskedasticity. Econometrica 48, 817–838.
White, T.L., Justice, A.J., de Wit, H., 2002. Differential subjective effects of d-amphetamine by gender, hormone levels and menstrual cycle phase. Pharmacol.Biochem. Behav. 73, 729–741.
Winslow, B.T., Voorhees, K.I., Pehl, K.A., 2007. Methamphetamine abuse. Am. Fam.Physician 76, 1169–1174.
Yucel, M., Lubman, D.I., 2007. Neurocognitive and neuroimaging evidence ofbehavioural dysregulation in human drug addiction: implications for diagnosis,treatment and prevention. Drug Alcohol Rev. 26, 33–39.
Related Documents