LETTER Deconstructing environmental predictability: seasonality, environmental colour and the biogeography of marine life histories Dustin J. Marshall 1 * and Scott C. Burgess 2 1 School of Biological Sciences Monash University Melbourne, 3800, Australia 2 Department of Biological Science Florida State University Tallahassee, FL, 32306, USA *Correspondence: E-mail: dustin.marshall@monash. edu Abstract Environmental predictability is predicted to shape the evolution of life histories. Two key types of environmental predictability, seasonality and environmental colour, may influence life-history evo- lution independently but formal considerations of both and how they relate to life history are exceedingly rare. Here, in a global biogeographical analysis of over 800 marine invertebrates, we explore the relationships between both forms of environmental predictability and three fundamen- tal life-history traits: location of larval development (aplanktonic vs. planktonic), larval develop- mental mode (feeding vs. non-feeding) and offspring size. We found that both dispersal potential and offspring size related to environmental predictability, but the relationships depended on both the environmental factor as well as the type of predictability. Environments that were more sea- sonal in food availability had a higher prevalence of species with a planktonic larval stage. Future studies should consider both types of environmental predictability as each can strongly affect life- history evolution. Keywords Environmental predictability, life-history theory, offspring size. Ecology Letters (2015) 18: 174–181 INTRODUCTION Environmental predictability has long been suggested to be a driver of life-history evolution. Foundational work by Co- hen, and MacArthur and Wilson predicted that different life-history strategies are favoured depending on the predict- ability of environment conditions (Cohen 1966, 1967; MacArthur & Wilson 1967). Accordingly, environmental predictability covaries with patterns of biodiversity at a range of scales (Slobodkin & Sanders 1969; Brown 1973; Botero et al. 2014; Sieferling et al. 2014). In terrestrial ver- tebrates in particular, a range of life-history traits appear to be influenced by elements of environmental predictability (Fisher et al. 2013). For example, more predictable rainfall appears to favour shorter reproductive spans and increased litter size in Madagascar (Dewar & Richard 2007). While it is clear that environmental predictability matters for life-his- tory evolution, several important knowledge gaps remain. In some instances, there is an excellent body of theory (e.g. Travis 2001) but a dearth of data on how major life-history traits evolve in response to environmental predictability, chief among these being the evolution of dispersal strategies and offspring size. Theory predicts that environmental predictability will influ- ence the evolution of dispersal strategies (Cohen & Levin 1991). Whenever there is variation in expected fitness across space, theory predicts less selection for dispersal when the environment is predictable in time (this strongly depends on the degree of spatial autocorrelation; McPeek & Holt 1992; Travis 2001). While these predictions are fairly consistent from model to model, there have been surprisingly few studies that have searched for empirical support for these predictions (Duputie & Massol 2013). More generally, manipulative studies of the evolution of dispersal are hampered by the onerous logistics of such experiments (Duputie & Massol 2013), though excellent examples exist (Fronhofer et al. 2014). One solution to these challenges is to first search for macro- ecological patterns in dispersal strategies that covary with environmental predictability, provided a reasonable proxy of dispersal can be identified (Duputie & Massol 2013). The pre- dictions from theoretical models on dispersal evolution in autocorrelated environments (e.g. Cohen & Levin 1991; McPeek & Holt 1992; Travis 2001) can then be consulted to generate a priori hypotheses about associations between dis- persal and environmental predictability and tested using the observed macroecological patterns. Environmental predictability has long been implicated in the evolution of the key life-history trait of offspring size. Classic theory predicted that environments that are highly unpredictable with stochastic mortality favour rapid matura- tion and reproduction with minimal investment in individual offspring (MacArthur 1972). More recent theory predicts the opposite pattern – unpredictable environments select for greater per offspring investment so as to buffer them from sporadic disasters (McGinley et al. 1987; Einum & Fleming 2004). Still, more theory predicts that mean offspring size should be unaffected by environmental predictability, instead mothers should increase the level of within-clutch variation in offspring size when environments are less predictable (Geritz 1995; Marshall et al. 2008). Despite the wealth of theoretical predictions, there have been few direct tests of the role of environmental predictability in driving offspring size varia- tion. Those studies that have attempted to address the issue empirically have used fairly indirect proxies of environmental © 2014 John Wiley & Sons Ltd/CNRS Ecology Letters, (2015) 18: 174–181 doi: 10.1111/ele.12402

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

LETTER Deconstructing environmental predictability: seasonality,environmental colour and the biogeography of marine lifehistories
Dustin J. Marshall1* and Scott C.
Burgess2
1School of Biological Sciences
Monash University Melbourne,
3800, Australia2Department of Biological Science
Florida State University Tallahassee,
FL, 32306, USA
*Correspondence:
E-mail: dustin.marshall@monash.
edu
AbstractEnvironmental predictability is predicted to shape the evolution of life histories. Two key types ofenvironmental predictability, seasonality and environmental colour, may influence life-history evo-lution independently but formal considerations of both and how they relate to life history areexceedingly rare. Here, in a global biogeographical analysis of over 800 marine invertebrates, weexplore the relationships between both forms of environmental predictability and three fundamen-tal life-history traits: location of larval development (aplanktonic vs. planktonic), larval develop-mental mode (feeding vs. non-feeding) and offspring size. We found that both dispersal potentialand offspring size related to environmental predictability, but the relationships depended on boththe environmental factor as well as the type of predictability. Environments that were more sea-sonal in food availability had a higher prevalence of species with a planktonic larval stage. Futurestudies should consider both types of environmental predictability as each can strongly affect life-history evolution.
KeywordsEnvironmental predictability, life-history theory, offspring size.
Ecology Letters (2015) 18: 174–181
INTRODUCTION
Environmental predictability has long been suggested to bea driver of life-history evolution. Foundational work by Co-hen, and MacArthur and Wilson predicted that differentlife-history strategies are favoured depending on the predict-ability of environment conditions (Cohen 1966, 1967;MacArthur & Wilson 1967). Accordingly, environmentalpredictability covaries with patterns of biodiversity at arange of scales (Slobodkin & Sanders 1969; Brown 1973;Botero et al. 2014; Sieferling et al. 2014). In terrestrial ver-tebrates in particular, a range of life-history traits appear tobe influenced by elements of environmental predictability(Fisher et al. 2013). For example, more predictable rainfallappears to favour shorter reproductive spans and increasedlitter size in Madagascar (Dewar & Richard 2007). While itis clear that environmental predictability matters for life-his-tory evolution, several important knowledge gaps remain. Insome instances, there is an excellent body of theory (e.g.Travis 2001) but a dearth of data on how major life-historytraits evolve in response to environmental predictability,chief among these being the evolution of dispersal strategiesand offspring size.Theory predicts that environmental predictability will influ-
ence the evolution of dispersal strategies (Cohen & Levin1991). Whenever there is variation in expected fitness acrossspace, theory predicts less selection for dispersal when theenvironment is predictable in time (this strongly depends onthe degree of spatial autocorrelation; McPeek & Holt 1992;Travis 2001). While these predictions are fairly consistentfrom model to model, there have been surprisingly few studiesthat have searched for empirical support for these predictions
(Duputie & Massol 2013). More generally, manipulativestudies of the evolution of dispersal are hampered by theonerous logistics of such experiments (Duputie & Massol2013), though excellent examples exist (Fronhofer et al. 2014).One solution to these challenges is to first search for macro-ecological patterns in dispersal strategies that covary withenvironmental predictability, provided a reasonable proxy ofdispersal can be identified (Duputie & Massol 2013). The pre-dictions from theoretical models on dispersal evolution inautocorrelated environments (e.g. Cohen & Levin 1991;McPeek & Holt 1992; Travis 2001) can then be consulted togenerate a priori hypotheses about associations between dis-persal and environmental predictability and tested using theobserved macroecological patterns.Environmental predictability has long been implicated in
the evolution of the key life-history trait of offspring size.Classic theory predicted that environments that are highlyunpredictable with stochastic mortality favour rapid matura-tion and reproduction with minimal investment in individualoffspring (MacArthur 1972). More recent theory predicts theopposite pattern – unpredictable environments select forgreater per offspring investment so as to buffer them fromsporadic disasters (McGinley et al. 1987; Einum & Fleming2004). Still, more theory predicts that mean offspring sizeshould be unaffected by environmental predictability, insteadmothers should increase the level of within-clutch variation inoffspring size when environments are less predictable (Geritz1995; Marshall et al. 2008). Despite the wealth of theoreticalpredictions, there have been few direct tests of the role ofenvironmental predictability in driving offspring size varia-tion. Those studies that have attempted to address the issueempirically have used fairly indirect proxies of environmental
© 2014 John Wiley & Sons Ltd/CNRS
Ecology Letters, (2015) 18: 174–181 doi: 10.1111/ele.12402

predictability and have focused on within-clutch variation inoffspring size rather than mean offspring size per se (Einum &Fleming 2002; Marshall et al. 2008). For example, Marshallet al. (2008) used developmental mode as a proxy for environ-mental predictability in marine invertebrates – species lackinga planktonic larval phase were supposed to be better able topredict their offspring’s environment relative to species with ahighly dispersive larval phase. There is limited support for thissupposition (Burgess & Marshall 2014), and ideally, a directexamination of local environmental predictability and off-spring size would constitute a stronger test. One option is toexplore macroecological associations between offspring sizeand environmental predictability.One largely overlooked issue in considering the role of
environmental predictability in driving life-history patternsis identifying the type of predictability that matters. Predict-ability can be thought of as having two components (Col-well 1974). The first type of predictability is the regularityin the timing and magnitude of fluctuations in the averageenvironmental state, such as that caused by regular tidal orseasonal cycles (e.g., it is predictably, say, an average of30 °C in June and an average of 10 °C in January in thenorthern hemisphere). Such predictability in terms of thetiming and magnitude of the mean environmental state isexpected to influence the evolution of life-history traits suchas phenology, dispersal and offspring size (Winemiller &Rose 1992; Sheldon & Tweksbury 2014). This type of pre-dictability occurs under some combination of constancy(Colwell’s C; complete constancy is when the environmentdoes not change) and contingency (Colwell’s M; i.e. a singleperiodicity where the environmental state varies betweenseasons but is the same for all seasons for all years). Forthe purposes of our explorations, we will define these peri-odic fluctuations as ‘seasonality’ because the timescales weconsider empirically match seasonal fluctuations in environ-mental trends. The second type of predictability is thedegree to which the environment is similar between succes-sive time points, or how far into the future the environmen-tal state is likely to stay the same, independent of the meanenvironmental state. This type of predictability is deter-mined by the structure of the variation around a givenmean trend (i.e. the residual variation) and is often calledthe colour of environmental noise (Steele 1985; Vasseur &Yodzis 2004), and we will refer to as environmental colour.White noise occurs when there is no correlation betweenone measurement and the next, whereas for red noise, thereis some correlation between measurements separated by afinite timescale; this correlation decreases with increasingtimescale. The ‘redder’ the noise, the greater the correlationfor a given separation timescale. Rather than knowing if,say, the temperature will be hot in June, this type of pre-dictability is the degree to which the temperature today issimilar to the temperature x days into the future. This typeof predictability can still occur in variable environments, sois distinct from Colwell’s constancy. Autocorrelated environ-mental noise is known to be important in the evolution ofdispersal, phenotypic plasticity and offspring size (Travis2001; Sheiner & Holt 2012; Burgess & Marshall 2014),as well as the risk of demographic extinction (Ripa &
Lundberg 1996). The key distinction between seasonalityand environmental colour relevant to life histories is thatthe former is about predictability of the mean environmen-tal state, while the latter is about how long a particularenvironment state stays above or below the seasonal cycle.Environments can be equally variable, but with differenttypes and degrees of predictability. Both of these types ofpredictability (seasonality and environmental colour) haverelevance to life-history evolution in variable environmentsfor potentially different reasons (see Figure 6, Winemiller &Rose 1992), but are rarely analysed together formally. Wewould argue that given both types of predictability matterto life-history evolution, an integrative understanding ofpredictability requires analysing predictability in both sea-sonality and environmental colour simultaneously.One major group of organisms that lends itself well to mac-
roecological explorations of patterns in dispersal mode andoffspring size is marine invertebrates. Marine invertebratesshow tremendous variation in dispersal modes and offspringsize at the lowest taxonomic levels (Marshall et al. 2012). Forexample, some species produce tiny larvae that spend weeksin the plankton, whereas closely related species produce off-spring two to three orders of magnitude larger that develop inthe complete absence of a pelagic larval phase (Shanks 2009;Weersing & Toonen 2009). These differences in developmentalmode translate into differences in dispersal potential – meta-analyses confirm that the largest scales of dispersal occur inspecies with a long-lived (weeks to months) feeding pelagiclarval phase, while species with aplanktonic development tendto have much smaller scales of dispersal (cm’s to m’s) (Shanks2009). Despite the tremendous variation in the life histories ofmarine invertebrates and a long history of speculation as tothe drivers of this variation (Palmer & Strathmann 1981;Strathmann 1985), no study has determined whether dispersalpotential or offspring size in this group covaries with environ-mental predictability.Here, we present results from the first global exploration of
the relationship between life history and the two types of envi-ronmental predictability: seasonality and environmental col-our. We explore biogeographical covariance among marineinvertebrate offspring size, larval developmental location(planktonic vs. aplanktonic), larval development mode (feed-ing vs. non-feeding) and environmental predictability. Ourwork follows a long history of biogeographic research in thisgroup. Thorson (1950) suggested that marine life histories var-ied predictably across latitudes. Specifically, Thorson sug-gested that species with feeding larvae were less common atthe poles because limited productivity and very cold tempera-tures precluded successful planktonic development. Eventuallyknown as ‘Thorson’s rule’, this idea was debated for over50 years, and while it has been modified significantly, many ofthe original tenets of this idea are supported today (Marshallet al. 2012). Modern assessments confirm that marine life his-tories show strong latitudinal gradients (Fernandez et al.2009) and differences in mean temperatures and mean levelsof primary productivity (measured as chlorophyll a concentra-tions) across latitudes are good predictors of some of thesepatterns (Fernandez et al. 2009). There are other importantpatterns in marine life histories that remain unresolved, how-
© 2014 John Wiley & Sons Ltd/CNRS
Letter Environmental predictability and biogeography 175

ever. Most notably, the prevalence of species with feeding lar-vae (perhaps the most prominent element Thorson’s originalhypothesis) and biogeographic patterns in offspring size formost species are not predicted by mean levels of primary pro-ductivity (Marshall et al. 2012). The failure of mean produc-tivity to explain these life-history patterns is surprising giventhat planktonic food is essential for species with obligate feed-ing larvae in order for them to complete their larval period.Empirical work confirms that mean food availability alonedoes not drive the selective advantages of larval feeding,instead, variability in larval food also matters (Miner &Vonesh 2004). Our hypothesis here is that environmental pre-dictability (both in terms of temperature and food) are associ-ated with these life-history patterns. Specifically, we predictthat: (1) taxa with a pelagic larval phase are least commonwhen food availability is predictable, because temporally auto-correlated environments do not favour dispersal (Travis 2001)and (2) for species with pelagic larvae, larvae that spendlonger in the plankton (i.e. feeding larvae) are more commonwhen food availability is unpredictable so as to increase theprobability of avoiding stochastic declines in local food avail-ability. We make no a priori predictions about the effects ofenvironmental predictability on offspring size as the underly-ing theory is ambivalent; some models predict decreases inoffspring size with decreasing predictability (Sargent et al.1987), while others (Einum & Fleming 2004) predict increasesin offspring size under unpredictable conditions. Throughoutour explorations, we consider only temporal scales of variabil-ity, we do not consider spatial scales because we lack data atsufficient spatial resolutions at global scales. Life histories areundoubtedly shaped by combinations of both spatial and tem-poral autocorrelation (Travis 2001) and we look forward toincorporating spatial autocorrelation when such data becomeavailable.
MATERIALS AND METHODS
Life-history and biophysical variables
Our analyses were based on the database of global marineinvertebrate life-history patterns published in Marshall et al.(2012) and Pringle et al. (2014). Here, we focused on larvaldevelopment and offspring size retaining the simplified classifi-cation of ‘planktonic’ and ‘aplankonic’ development fromMarshall et al. (2012). Species are classified as having plank-tonic development if at least part of the larval phase is spentin water column independent of the mother – under such aclassification, species with some development in a benthic eggcapsule but that hatches free swimming larva would thereforebe classed as planktonic. Within species with a planktonic lar-val phase, we further classified species as feeding (plankto-trophic) and non-feeding. Species were classified as having afeeding larva if food was acquired by the larva independentlyof maternal provisioning. For details of the database, how thedata were compiled, and for further discussion of the relativemerits of the simplified classification system of developmentalmode, see Marshall et al. (2012). Importantly, almost all spe-cies are shallow water inhabitants, so sea surface temperatures(SST) are likely to reflect their usual conditions (Fig. S1). The
life-history section of the database is now available fromDryad (http://dx.doi.org/10.5061/dryad.m7j72). A previousstudy also showed that phylogeny does not drive biogeo-graphical patterns in life-history traits in the species consid-ered here (Marshall et al. 2012), and so phylogeneticallycontrolled analyses were not repeated here. All classificationswere based on published studies that documented their modeof development, none were inferred indirectly.For each 1° latitude by 1° longitude grid cell where there
were life-history data, time-series data on chlorophyll a con-centration (mg/m3; MODIS 9 km) and SST (°C, 11 micronnight; MODIS-Terra 9 km) were extracted from SeaWiFS(http://disc.sci.gsfc.nasa.gov). The time-series data on chloro-phyll a and SST covered a 10-year period from July 2002 toJuly 2012. Chlorophyll a data were recorded every eight days(Dt = 8 days) and SST data were recorded every month(Dt = 1 month). We used grid cells that had < 13% of thetime series missing (75% of the grid cells for chlorophyll aand 99% of the grid cells for SST). The number of observa-tions within each grid cell was n = 461 for the chlorophyll adata and n = 121 for the SST data. Note that because datawere sparse in latitudes above 50°, we could not include thecolder, less variable polar regions. As such, the number ofspecies we could include in our analyses here was less thanthe number used in Marshall et al. (2012; ~ 800 vs. ~ 1000species respectively). Prior to any analyses, linear trends(though minor) were removed by extracting the residuals froma linear regression model fitted to the raw time series.We decomposed the time-series data on chlorophyll a and
SST into the two types of predictability: seasonality and envi-ronmental colour. To estimate seasonality, we calculated thevariance of the seasonal trend for each spatial grid point. Toestimate the seasonal trend, we binned the time-series data ofchl and SST into monthly intervals, took the average for eachmonth (i.e. average over all 10 years), then re-created a sea-sonal time-series data set on the same timescale of the originaldata using a linear interpolation between the monthly mid-points. From this seasonal trend, we calculated the varianceof the seasonal trend (denoted ‘a’, below). We also calculatedthe variance of the residual time series (i.e. the time seriesafter the seasonal trend was removed; denoted ‘b’). The frac-tion of the total variance that is due to predictable seasonalperiodicities is then a/(a+b) and is what we call ‘seasonality’.Using absolute variance as the predictor in our life-historyanalyses did not change our results qualitatively.To estimate the colour of environmental variation, we per-
formed a spectral analysis on the residual time series with the sea-sonal trend removed. The residual time series was calculated bysubtracting the corresponding seasonal value from each datapoint in the time series. Following standard techniques (Steele1985; Vasseur & Yodzis 2004), we assumed that variance scaleswith frequency (f) according to an inverse power law, 1/fb. Whitenoise (b = 0) occurs when there is no correlation between onemeasurement and the next, while for reddened noise (b > 0),there is some correlation between measurements separated by afinite timescale. The ‘redder’ the noise, the greater the correlationfor a given separation timescale. The family of 1/fb noise modelsseems to describe well the fluctuations of a wide range of physi-cal environmental variables (Steele 1985; Vasseur & Yodzis
© 2014 John Wiley & Sons Ltd/CNRS
176 D. J. Marshall and S. C. Burgess Letter
2004). Spectral analyses were performed in R (R Core Develop-ment Team, 2012) using Lomb-Scargle Periodograms to ensurethat missing values did not bias estimates of the spectral densi-ties (Press & Rybicki 1989; Glynn et al. 2006). The spectralexponent b was estimated as in Steele (1985) and Vasseur &Yodzis (2004) as the negative slope of the linear regression oflog10 spectral density against log10 frequency. Spectral densitiesand subsequent b’s were calculated between the frequencies of2/nDt and 1/2Dt, which translates to periods of 16 days to5.05 years for chlorophyll a and 30 days to 5.05 years for SST.
Covariance among biophysical variables
Initial explorations showed that the coefficient of variation inSST (CV SST) showed extremely strong negative covariancewith mean SST. To avoid colinearity issues, CV SST wasexcluded from further analyses. Higher temperatures should beconsidered synonymously with less variable temperatures. Noother variables showed strong covariance with each other andimportantly, seasonality and environmental colour did not co-vary.Mean SST and CV SST both vary with latitude such that
lower latitudes are warmer and less variable than higher lati-tudes. The other variables of interest showed less straightfor-ward associations with latitude (Fig. S2). Generally, higherlatitudes had higher mean productivity and were more vari-able. Seasonality in temperature showed associations with lati-tude – higher latitudes were more seasonal in temperature.
Analysis of life-history variables with biophysical variables andmodel performance
After centring our predictors, we used multiple logisticregression to relate our biophysical variables to larval devel-opmental location (aplanktonic vs. planktonic) and larval
developmental mode (feeding vs. non-feeding) and multipleregression to relate them offspring size. We used standardmodel reduction approaches to simplify our models whereappropriate (Quinn & Keough 2002).
RESULTS
Larval dispersal and developmental mode
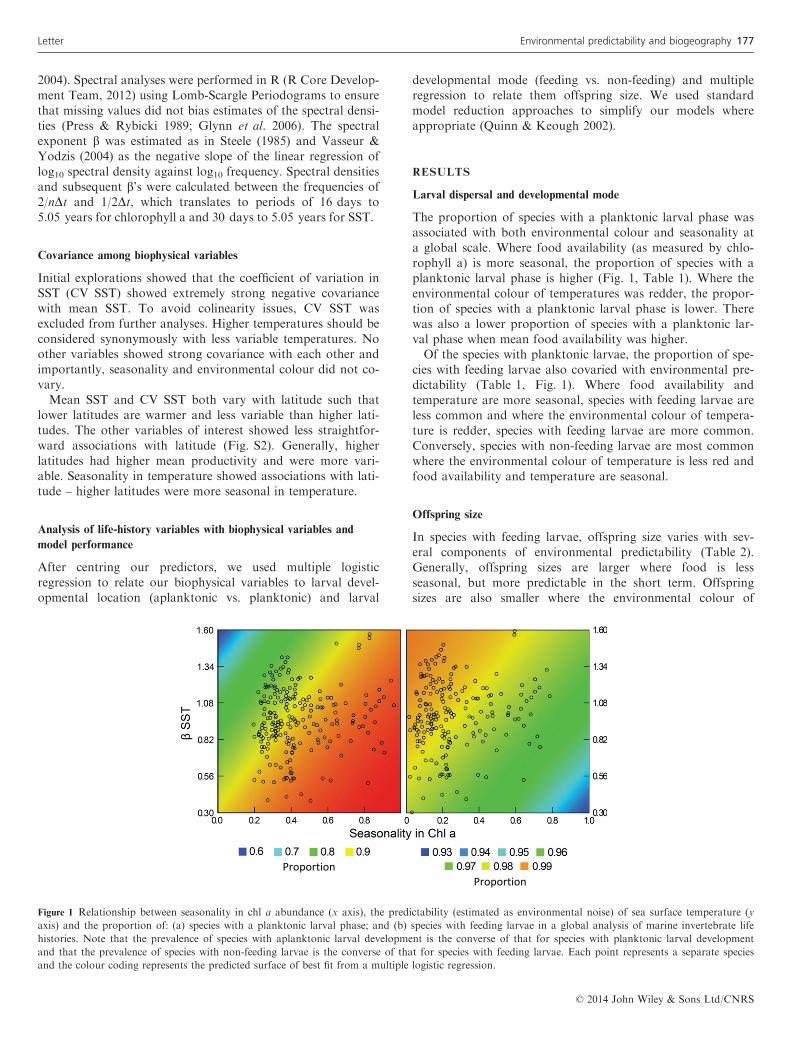
The proportion of species with a planktonic larval phase wasassociated with both environmental colour and seasonality ata global scale. Where food availability (as measured by chlo-rophyll a) is more seasonal, the proportion of species with aplanktonic larval phase is higher (Fig. 1, Table 1). Where theenvironmental colour of temperatures was redder, the propor-tion of species with a planktonic larval phase is lower. Therewas also a lower proportion of species with a planktonic lar-val phase when mean food availability was higher.Of the species with planktonic larvae, the proportion of spe-
cies with feeding larvae also covaried with environmental pre-dictability (Table 1, Fig. 1). Where food availability andtemperature are more seasonal, species with feeding larvae areless common and where the environmental colour of tempera-ture is redder, species with feeding larvae are more common.Conversely, species with non-feeding larvae are most commonwhere the environmental colour of temperature is less red andfood availability and temperature are seasonal.
Offspring size
In species with feeding larvae, offspring size varies with sev-eral components of environmental predictability (Table 2).Generally, offspring sizes are larger where food is lessseasonal, but more predictable in the short term. Offspringsizes are also smaller where the environmental colour of
ProportionProportion
Figure 1 Relationship between seasonality in chl a abundance (x axis), the predictability (estimated as environmental noise) of sea surface temperature (yaxis) and the proportion of: (a) species with a planktonic larval phase; and (b) species with feeding larvae in a global analysis of marine invertebrate lifehistories. Note that the prevalence of species with aplanktonic larval development is the converse of that for species with planktonic larval developmentand that the prevalence of species with non-feeding larvae is the converse of that for species with feeding larvae. Each point represents a separate speciesand the colour coding represents the predicted surface of best fit from a multiple logistic regression.
© 2014 John Wiley & Sons Ltd/CNRS
Letter Environmental predictability and biogeography 177
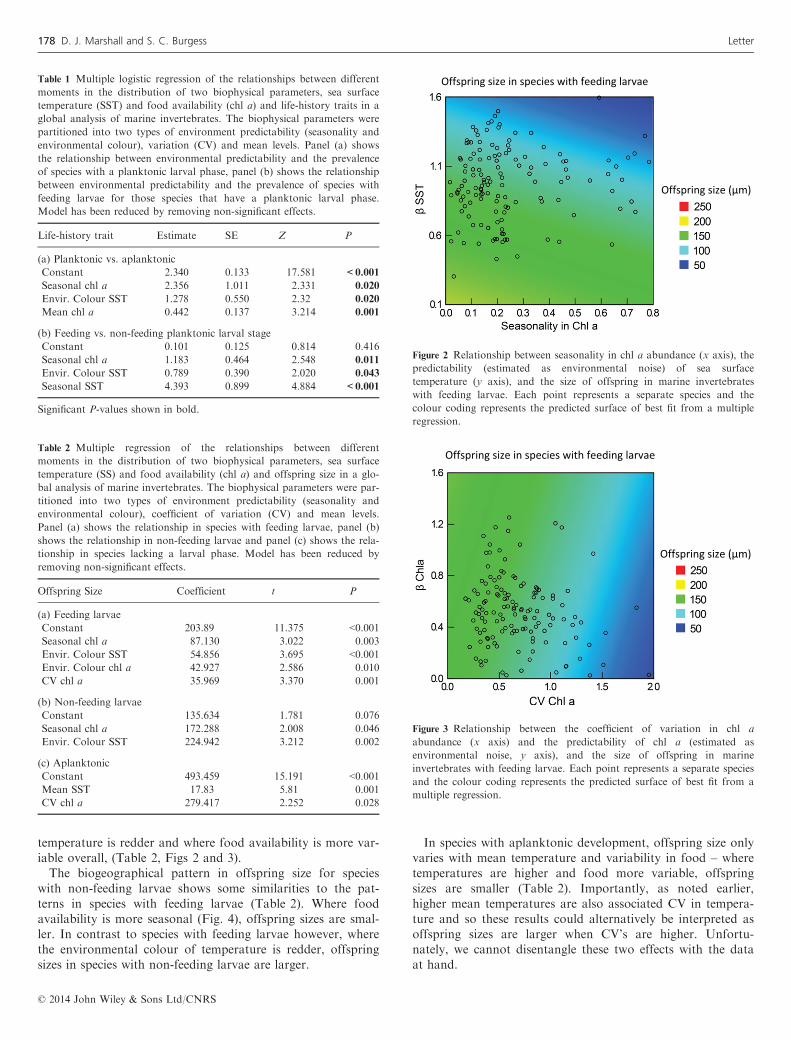
temperature is redder and where food availability is more var-iable overall, (Table 2, Figs 2 and 3).The biogeographical pattern in offspring size for species
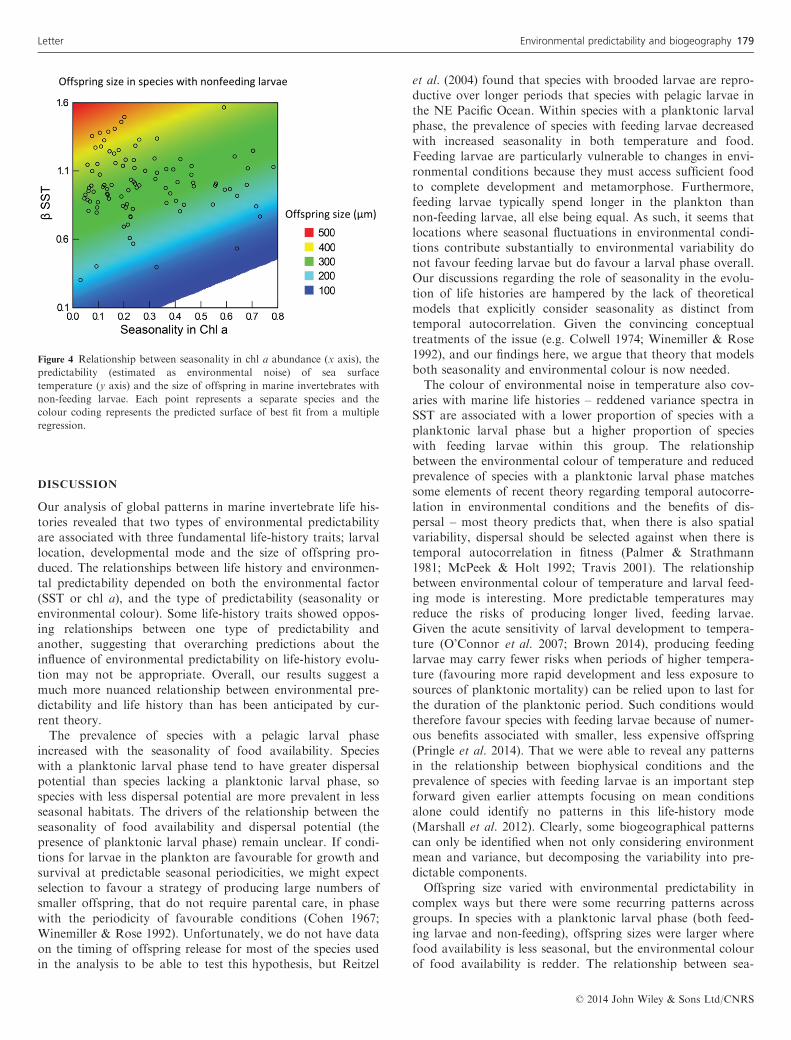
with non-feeding larvae shows some similarities to the pat-terns in species with feeding larvae (Table 2). Where foodavailability is more seasonal (Fig. 4), offspring sizes are smal-ler. In contrast to species with feeding larvae however, wherethe environmental colour of temperature is redder, offspringsizes in species with non-feeding larvae are larger.
In species with aplanktonic development, offspring size onlyvaries with mean temperature and variability in food – wheretemperatures are higher and food more variable, offspringsizes are smaller (Table 2). Importantly, as noted earlier,higher mean temperatures are also associated CV in tempera-ture and so these results could alternatively be interpreted asoffspring sizes are larger when CV’s are higher. Unfortu-nately, we cannot disentangle these two effects with the dataat hand.
Table 2 Multiple regression of the relationships between differentmoments in the distribution of two biophysical parameters, sea surfacetemperature (SS) and food availability (chl a) and offspring size in a glo-bal analysis of marine invertebrates. The biophysical parameters were par-titioned into two types of environment predictability (seasonality andenvironmental colour), coefficient of variation (CV) and mean levels.Panel (a) shows the relationship in species with feeding larvae, panel (b)shows the relationship in non-feeding larvae and panel (c) shows the rela-tionship in species lacking a larval phase. Model has been reduced byremoving non-significant effects.
Offspring Size Coefficient t P
(a) Feeding larvaeConstant 203.89 11.375 <0.001Seasonal chl a !87.130 !3.022 0.003Envir. Colour SST !54.856 !3.695 <0.001Envir. Colour chl a 42.927 2.586 0.010CV chl a !35.969 !3.370 0.001
(b) Non-feeding larvaeConstant 135.634 1.781 0.076Seasonal chl a !172.288 !2.008 0.046Envir. Colour SST 224.942 !3.212 0.002
(c) AplanktonicConstant 493.459 15.191 <0.001Mean SST !17.83 !5.81 0.001CV chl a !279.417 !2.252 0.028
Offspring size (µm)
Offspring size in species with feeding larvae
Figure 2 Relationship between seasonality in chl a abundance (x axis), thepredictability (estimated as environmental noise) of sea surfacetemperature (y axis), and the size of offspring in marine invertebrateswith feeding larvae. Each point represents a separate species and thecolour coding represents the predicted surface of best fit from a multipleregression.
Table 1 Multiple logistic regression of the relationships between differentmoments in the distribution of two biophysical parameters, sea surfacetemperature (SST) and food availability (chl a) and life-history traits in aglobal analysis of marine invertebrates. The biophysical parameters werepartitioned into two types of environment predictability (seasonality andenvironmental colour), variation (CV) and mean levels. Panel (a) showsthe relationship between environmental predictability and the prevalenceof species with a planktonic larval phase, panel (b) shows the relationshipbetween environmental predictability and the prevalence of species withfeeding larvae for those species that have a planktonic larval phase.Model has been reduced by removing non-significant effects.
Life-history trait Estimate SE Z P
(a) Planktonic vs. aplanktonicConstant 2.340 0.133 !17.581 <0.001Seasonal chl a 2.356 1.011 !2.331 0.020Envir. Colour SST !1.278 0.550 2.32 0.020Mean chl a !0.442 0.137 3.214 0.001
(b) Feeding vs. non-feeding planktonic larval stageConstant 0.101 0.125 0.814 0.416Seasonal chl a !1.183 0.464 !2.548 0.011Envir. Colour SST 0.789 0.390 2.020 0.043Seasonal SST !4.393 0.899 !4.884 <0.001
Significant P-values shown in bold.
Offspring size (µm)
Offspring size in species with feeding larvae
Figure 3 Relationship between the coefficient of variation in chl aabundance (x axis) and the predictability of chl a (estimated asenvironmental noise, y axis), and the size of offspring in marineinvertebrates with feeding larvae. Each point represents a separate speciesand the colour coding represents the predicted surface of best fit from amultiple regression.
© 2014 John Wiley & Sons Ltd/CNRS
178 D. J. Marshall and S. C. Burgess Letter
DISCUSSION
Our analysis of global patterns in marine invertebrate life his-tories revealed that two types of environmental predictabilityare associated with three fundamental life-history traits; larvallocation, developmental mode and the size of offspring pro-duced. The relationships between life history and environmen-tal predictability depended on both the environmental factor(SST or chl a), and the type of predictability (seasonality orenvironmental colour). Some life-history traits showed oppos-ing relationships between one type of predictability andanother, suggesting that overarching predictions about theinfluence of environmental predictability on life-history evolu-tion may not be appropriate. Overall, our results suggest amuch more nuanced relationship between environmental pre-dictability and life history than has been anticipated by cur-rent theory.The prevalence of species with a pelagic larval phase
increased with the seasonality of food availability. Specieswith a planktonic larval phase tend to have greater dispersalpotential than species lacking a planktonic larval phase, sospecies with less dispersal potential are more prevalent in lessseasonal habitats. The drivers of the relationship between theseasonality of food availability and dispersal potential (thepresence of planktonic larval phase) remain unclear. If condi-tions for larvae in the plankton are favourable for growth andsurvival at predictable seasonal periodicities, we might expectselection to favour a strategy of producing large numbers ofsmaller offspring, that do not require parental care, in phasewith the periodicity of favourable conditions (Cohen 1967;Winemiller & Rose 1992). Unfortunately, we do not have dataon the timing of offspring release for most of the species usedin the analysis to be able to test this hypothesis, but Reitzel
et al. (2004) found that species with brooded larvae are repro-ductive over longer periods that species with pelagic larvae inthe NE Pacific Ocean. Within species with a planktonic larvalphase, the prevalence of species with feeding larvae decreasedwith increased seasonality in both temperature and food.Feeding larvae are particularly vulnerable to changes in envi-ronmental conditions because they must access sufficient foodto complete development and metamorphose. Furthermore,feeding larvae typically spend longer in the plankton thannon-feeding larvae, all else being equal. As such, it seems thatlocations where seasonal fluctuations in environmental condi-tions contribute substantially to environmental variability donot favour feeding larvae but do favour a larval phase overall.Our discussions regarding the role of seasonality in the evolu-tion of life histories are hampered by the lack of theoreticalmodels that explicitly consider seasonality as distinct fromtemporal autocorrelation. Given the convincing conceptualtreatments of the issue (e.g. Colwell 1974; Winemiller & Rose1992), and our findings here, we argue that theory that modelsboth seasonality and environmental colour is now needed.The colour of environmental noise in temperature also cov-
aries with marine life histories – reddened variance spectra inSST are associated with a lower proportion of species with aplanktonic larval phase but a higher proportion of specieswith feeding larvae within this group. The relationshipbetween the environmental colour of temperature and reducedprevalence of species with a planktonic larval phase matchessome elements of recent theory regarding temporal autocorre-lation in environmental conditions and the benefits of dis-persal – most theory predicts that, when there is also spatialvariability, dispersal should be selected against when there istemporal autocorrelation in fitness (Palmer & Strathmann1981; McPeek & Holt 1992; Travis 2001). The relationshipbetween environmental colour of temperature and larval feed-ing mode is interesting. More predictable temperatures mayreduce the risks of producing longer lived, feeding larvae.Given the acute sensitivity of larval development to tempera-ture (O’Connor et al. 2007; Brown 2014), producing feedinglarvae may carry fewer risks when periods of higher tempera-ture (favouring more rapid development and less exposure tosources of planktonic mortality) can be relied upon to last forthe duration of the planktonic period. Such conditions wouldtherefore favour species with feeding larvae because of numer-ous benefits associated with smaller, less expensive offspring(Pringle et al. 2014). That we were able to reveal any patternsin the relationship between biophysical conditions and theprevalence of species with feeding larvae is an important stepforward given earlier attempts focusing on mean conditionsalone could identify no patterns in this life-history mode(Marshall et al. 2012). Clearly, some biogeographical patternscan only be identified when not only considering environmentmean and variance, but decomposing the variability into pre-dictable components.Offspring size varied with environmental predictability in
complex ways but there were some recurring patterns acrossgroups. In species with a planktonic larval phase (both feed-ing larvae and non-feeding), offspring sizes were larger wherefood availability is less seasonal, but the environmental colourof food availability is redder. The relationship between sea-
Offspring size (µm)
Offspring size in species with nonfeeding larvae
Figure 4 Relationship between seasonality in chl a abundance (x axis), thepredictability (estimated as environmental noise) of sea surfacetemperature (y axis) and the size of offspring in marine invertebrates withnon-feeding larvae. Each point represents a separate species and thecolour coding represents the predicted surface of best fit from a multipleregression.
© 2014 John Wiley & Sons Ltd/CNRS
Letter Environmental predictability and biogeography 179

sonality of food and offspring size supports earlier suggestionsthat seasonal environments, with predictable fluctuationsevery year, are effectively more benign than less seasonal envi-ronments (Slobodkin & Sanders 1969; but see Winemiller &Rose 1992). Across a range of taxa, including marine inverte-brates, mothers tend to make smaller offspring when food isabundant, and larger offspring when food is scarce (Fox 1997;Allen et al. 2008; Bashey 2008). Smaller offspring may repre-sent a smaller investment from mothers such that they canproduce more offspring for a given amount of resource. Whenfood is scarce, mothers must produce larger offspring so thatoffspring have sufficient resources but when food is abundant,mothers can produce smaller offspring and enjoy a fecunditybenefit from the reduction in per offspring investment (Mar-shall et al. 2006). Surprisingly, this relationship between pre-dictability and offspring size differed when we also consideredthe other type of predictability - the colour of environmentalnoise.In species with feeding larvae, offspring size increases with
redder environmental noise in food availability but decreaseswith redder environmental noise in temperature. When thenoise in food availability was redder, offspring sizes were larger– the opposite relationship as to that for increased seasonality.Increased investment associated with higher predictability infood in the shorter term could be interpreted as support for the‘safe-harbour’ hypothesis (Sargent et al. 1987), when conditionsare likely to be good for extended periods of time (i.e. whenenvironmental noise is red and mean conditions are favour-able), increased investment might be favoured because stochas-tic mortality (which favours smaller offspring sizes) is reduced.Interestingly, species that lack a pelagic larval phase showed norelationship between offspring size and environmental predict-ability – this group represents those species with the largest off-spring sizes and one hypothesis for this group is thatpredictability in food availability is less important as very largeoffspring are buffered from such effects (Einum & Fleming2004). We are uncomfortable with these explanations, however,for several reasons. First, the relationship between offspring sizeand the environmental colour of temperature reversed in signbetween species with feeding larvae and species with non-feed-ing larvae, suggesting that any generic hypotheses about largeroffspring size being better in more predictable conditions areinadequate. These contrasting relationships between offspringsize and predictability suggest that current offspring size theoryhas not considered the role of environmental predictability ade-quately. Indeed, current offspring size theory makes no distinc-tion between the two types of environmental predictability andan important next step will be to explore their roles theoreti-cally. Of course, the patterns we observed may not represent anadaptive response to these conditions at all – unfortunately,biogeographical patterns do not reveal causality, but they dogenerate hypotheses for future work. We should also note thatmarine invertebrates exhibit significant phenotypic plasticitywith regard to offspring size (Allen et al. 2008). Environmentalnoise will interact with phenotypic plasticity in complex ways(Burgess & Marshall 2014) so the extent to which mothers cantrack local conditions and modify their offspring accordingly islikely to vary across the continua of predictabilities observedhere.
Our analyses show that using independent measures of twotypes of environmental predictability revealed nuanced bioge-ographic patterns that were not visible using more traditionalapproaches. The additional information on predictabilityoffered by combining our seasonality measure (which corre-lates with the contingency element of predictability from Col-well’s measure of predictability) and our measure ofenvironmental colour is that we investigate the timing andmagnitude of events in a fluctuating environment as well asthe degree of autocorrelated stochastic variation. Our resultsshowed that seasonality and environmental noise can haverelationships with spatial life-history patterns that are inopposite directions (e.g. offspring size varied negatively withseasonality but positively with environmental colour). The lat-ter (environmental colour) exposes the relative importancehidden periodicities that may be missed by only considering asingle period of fluctuation (e.g. seasonal), which does haveimportant biological consequences (Vasseur & Yodzis 2004).Furthermore, future work comparing multiple models of auto-correlation (e.g., 1/f vs autoregressive vs. moving average) willalso allow investigations of the biological importance of thecorrelation structure of the variable of interest (Cuddington &Yodzis 1999). Note that some of the information obtainedfrom spectral analyses can be obtained using Colwell’smethod, but spectral analysis provides much more informa-tion for much less work, and reduces the potential to misshidden periodicities (Stearns 1981). Many macroecologicalstudies use Colwell’s measure of predictability. We recom-mend a combination of methods to estimate the multiple typesof predictability to access more potential for greater biologicalinsight into macroecological patterns.
ACKNOWLEDGEMENTS
This manuscript benefited from comments on an earlier versionof this manuscript by anonymous reviewers, Jeb Byers, RichardStrathmann, Bob Warner, Grant Duffy and Stephanie Kamel,as well as very helpful discussions with Michel Loreau and Ja-mie Pringle. DJM was supported by grants from the AustralianResearch Council and SCB was supported by the Centre forPopulation Biology Fellowship from UC-Davis.
AUTHORSHIP
DJM and SCB analysed the data and wrote the manuscript.
REFERENCES
Allen, R.M., Buckley, Y.M. & Marshall, D.J. (2008). Offspring sizeplasticity in response to intraspecific competition: an adaptive maternaleffect across life-history stages. Am. Nat., 171, 225–237.
Bashey, F. (2008). Competition as a selective mechanism for largeroffspring size in guppies. Oikos, 117, 104–113.
Botero, C.A., Dor, R., McCain, C.M. & Safran, R.J. (2014).Environmental harshness is positively correlated with intraspecificdivergence in mammals and birds. Mol. Ecol., 23, 259–268.
Brown, J.H. (1973). Species diversity of seed-eating desert rodents in sanddune habitats. Ecology, 54, 775–787.
Brown, J.H. (2014). Why marine islands are farther apart in the tropics.Am. Nat., 183, 842–846.
© 2014 John Wiley & Sons Ltd/CNRS
180 D. J. Marshall and S. C. Burgess Letter

Burgess, S.C. & Marshall, D.J. (2014). Adaptive parental effects: theimportance of estimating environmental predictability and offspringfitness appropriately. Oikos, 123, 769–776.
Cohen, D. (1966). Optimizing reproduction in a randomly varyingenvironment. J. Theor. Biol., 12, 119–129.
Cohen, D. (1967). Optimizing reproduction in a randomly varyingenvironment when a correlation may exist between the conditions atthe time a choice has to be made and the subsequent outcome. J.Theor. Biol., 16, 1–14.
Cohen, D. & Levin, S.A. (1991). Dispersal in patchy environments: theeffects of temporal and spatial structure. Theor. Popul. Biol., 39, 63–99.
Colwell, R.K. (1974). Predictability, constancy, and contingency ofperiodic phenomena. Ecology, 55, 1148–1153.
Cuddington, K.M. & Yodzis, P. (1999). Black noise and populationpersistence. Proc. Roy. Soc. Lond. Ser. B., 266, 969–973.
Dewar, R.E. & Richard, A.F. (2007). Evolution in the hypervariableenvironment of Madagascar. Proc. Natl. Acad. Sci. USA, 104, 13723–13727.
Duputie, A. & Massol, F. (2013). An empiricist’s guide to theoreticalpredictions on the evolution of dispersal. Interface Focus, 3, 20130028.
Einum, S. & Fleming, I.A. (2002). Does within-population variation infish egg size reflect maternal influences on optimal values. Am. Nat.,160, 756–765.
Einum, S. & Fleming, I.A. (2004). Environmental unpredictability andoffspring size: conservative versus diversified bet-hedging. Evol. Ecol.Res., 6, 443–455.
Fernandez, M., Astorga, A., Navarrete, S.A., Valdovinos, C. &Marquet, P.A. (2009). Deconstructing latitudinal species richnesspatterns in the ocean: does larval development hold the clue? Ecol.Lett., 12, 601–611.
Fisher, D.O., Dickman, C.R., Jones, M.E. & Blomberg, S.P. (2013).Sperm competition drives the evolution of suicidal reproduction inmammals. Proc. Natl. Acad. Sci. USA, 110, 17910–17914.
Fox, C.W. (1997). The ecology of body size in a seed beetle, Statorlimbatus: persistence of environmental variation across generations?Evolution, 51, 1005–1010.
Fronhofer, E.A., Stelz, J.M., Lutz, E., Poethke, H.J. & Bonte, D. (2014).Spatially correlated extinctions select for less emigration but largerdispersal distances in the spider mite Tetranychus urticae. Evolution, 68,1838–1844.
Geritz, S.A.H. (1995). Evolutionarily stable seed polymorphism andsmall-scale spatial variation in seedling density. Am. Nat., 146, 685–707.
Glynn, E.F., Chen, J. & Mushegian, A.R. (2006). Detecting periodicpatterns in unevenly spaced gene expression time series using Lomb-Scargle periodograms. Bioinformatics, 22, 310–316.
MacArthur, R. (1972). Geographical Ecology. Harper and Rowe, NewYork.
MacArthur, R.H. & Wilson, E.O. (1967). The Theory of IslandBiogeography. Princeton University Press, Princeton, New Jersey.
Marshall, D.J., Cook, C.N. & Emlet, R.B. (2006). Offspring size effectsmediate competitive interactions in a colonial marine invertebrate.Ecology, 87, 214–225.
Marshall, D.J., Bonduriansky, R. & Bussiere, L.F. (2008). Offspring sizevariation within broods as a bet-hedging strategy in unpredictableenvironments. Ecology, 89, 2506–2517.
Marshall, D.J., Krug, P.J., Kupriyanova, E.K., Byrne, M. & Emlet, R.B.(2012). The biogeography of marine invertebrate life histories. Annu.Rev. Ecol. Evol. Syst., 43, 97–114.
McGinley, M.A., Temme, D.H. & Geber, M.A. (1987). Parentalinvestment in offspring in variable environments: theoretical andempirical considerations. Am. Nat., 130, 370–398.
McPeek, M.A. & Holt, R.D. (1992). The evolution of dispersal in spatiallyand temporally varying environments. Am. Nat., 140, 1010–1027.
Miner, B.G. & Vonesh, J.R. (2004). Effects of fine rain environmentalvariability on morphological plasticity. Ecol. Lett., 7, 794–801.
O’Connor, M.I., Bruno, J.F., Gaines, S.D., Halpern, B.S., Lester, S.E.,Kinlan, B.P. et al. (2007). Temperature control of larval dispersal and
the implications for marine ecology, evolution, and conservation. Proc.Natl. Acad. Sci. USA, 104, 1266–1271.
Palmer, A.R. & Strathmann, R.R. (1981). Scale of dispersal in varyingenvironmnets and its implications for life-histories of marineinvertebrates. Oecologia, 48, 308–318.
Press, W.H. & Rybicki, G.B. (1989). Fast algorithm for spectral analysisof unevenly sampled data. Astrophys. J., 338, 277–280.
Pringle, J.M., Byers, J.E., Pappalardo, P., Wares, J.P. & Marshall, D.J.(2014). Circulation constrains the evolution of larval developmentmodes and life histories in the coastal ocean. Ecology, 95, 1022–1032.
Quinn, G.P. & Keough, M.J. (2002). Experimental Design and DataAnalysis for Biologists. Cambridge University Press, Cambridge, U. K.
R Development Core Team (2012). R: A language and environment forstatistical computing. R Foundation for Statistical Computing, Vienna,Austria.
Reitzel, A.M., Miner, B.G. & McEdward, L.R. (2004). Relationshipsbetween spawning date and larval development time for benthic marineinvertebrates: a modeling approach. Mar. Ecol. Prog. Ser., 280, 13–23.
Ripa, J. & Lundberg, P. (1996). Noise colour and the risk of populationextinctions. Proc. Roy. Soc. Lond. Ser. B., 263, 1751–1753.
Sargent, R.C., Taylor, P.D. & Gross, M.R. (1987). Parental care and theevolution of egg size in fishes. Am. Nat., 129, 32–46.
Shanks, A.L. (2009). Pelagic larval duration and dispersal distancerevisited. Biol. Bull., 216, 373–385.
Sheiner, S.M. & Holt, R.D. (2012). The genetic of phenotypic plasticity.X. Variation versus uncertainty. Ecol. Evol., 3, 751–767.
Sheldon, K.S. & Tweksbury, J.J. (2014). The impact of seasonality intemperature on thermal tolerance and elevational range size. Ecology,95, 2134–2143.
Sieferling, I., Proulx, R. & Wirth, C. (2014). Disentangling theenvironmental-heterogeneity-species-diversity relationship along agradient of a human footprint. Ecology, 95, 2084–2095.
Slobodkin, L.B. & Sanders, H.L. (1969). On contribution ofenvironmental predictability to species diversity. Brookhaven Symp.Biol., 22, 82–95.
Stearns, S.C. (1981). On measuring fluctuating environments:predictability, constancy, and contingency. Ecology, 62, 185–199.
Steele, J.H. (1985). A comparison of terrestrial and marine ecologicalsystems. Nature, 313, 355–358.
Strathmann, R. (1985). Feeding and non-feeding larval development andlife-history evolution in marine invertebrates. Annu. Rev. Ecol. Syst.,16, 339–361.
Thorson, G. (1950). Reproductive and larval ecology of marine bottominvertebrates. Biol. Rev., 25, 1–45.
Travis, J.M.J. (2001). The color of noise and the evolution of dispersal.Ecol. Res., 16, 157–163.
Vasseur, D.A. & Yodzis, P. (2004). The color of environmental noise.Ecology, 85, 1146–1152.
Weersing, K. & Toonen, R.J. (2009). Population genetics, larval dispersal,and connectivity in marine systems. Mar. Ecol. Prog. Ser., 393, 1–12.
Winemiller, K.O. & Rose, K.A. (1992). Patterns of life-historydiversification in North American fishes: implications for populationregulation. Can. J. Fish Aquat. Sci., 49, 2196–2218.
SUPPORTING INFORMATION
Additional Supporting Information may be downloaded viathe online version of this article at Wiley Online Library(www.ecologyletters.com).
Editor, James ByersManuscript received 11 August 2014First decision made 11 September 2014Manuscript accepted 19 November 2014
© 2014 John Wiley & Sons Ltd/CNRS
Letter Environmental predictability and biogeography 181
Related Documents









![MODELING SEASONALITY IN AVIAN INFLUENZA H5N1Breban et al. [4] model seasonality and environmental transmission of low pathogenic avian influenza (LPAI) viruses in wild birds with](https://static.cupdf.com/doc/110x72/5e4ff4b6a699067e2614dfde/modeling-seasonality-in-avian-influenza-h5n1-breban-et-al-4-model-seasonality.jpg)

