Ž . Brain Research 730 1996 107–117 Research report Cyclosporin A, an inhibitor of calcineurin, impairs memory formation in day-old chicks Pauleen C. Bennett a , Weiqin Zhao a , Alfons Lawen b , Kim T. Ng a, ) a Department of Psychology, Monash UniÕersity, Victoria, Australia b Department of Biochemistry and Molecular Biology, Monash UniÕersity, Victoria, Australia Accepted 2 April 1996 Abstract Considerable evidence exists that changes in the phosphorylation state of neuronal proteins are correlated with learning and that inhibition of various protein kinases disrupts memory formation. Given the reversible nature of protein phosphorylation, a role for protein phosphatases in memory processing also seems likely. It has been shown recently that administration of the phosphatase inhibitor, okadaic w x acid, disrupts memory formation in day-old chicks, with retention deficits first appearing at approximately 40 min post-training 93 . In the present study the intracranial administration of the immunosuppressant cyclosporin A was also found to produce retention deficits in day-old chicks trained on a single-trial, passive-avoidance task, but the deficits were not significant until 85 min post-training. The difference could not be attributed to differences in the pharmacokinetics of the drugs. Since okadaic acid preferentially inhibits protein phosphatases 1 and 2A, while cyclosporin A is reported to inhibit only the Ca 2q rcalmodulin-dependent protein phosphatase, calcineurin, it is possible that different phosphatases may be involved in distinct stages of memory formation, as has been reported previously for protein kinases. The possibility that cyclosporin A may, in addition, act through inhibition of cyclophilin’s peptidyl-prolyl-cis r trans- isomerase activity is also canvassed. Keywords: Calcineurin; Cyclosporin A; Dephosphorylation; Immunosuppressant; Memory; Phosphatase 1. Introduction Protein phosphorylation has been indirectly implicated in biological information storage processes via its roles in many neuronal events which contribute to signal transduc- w x tion 90 , and through its roles in long-term potentiation Ž . w x Ž . w x LTP 88 , long-term depression LTD 61 , and neuronal w x kindling 34,91 . It has also been directly implicated in memory formation in many species, with kinase disruption via pharmacological or genetic means being consistently w x reported to impair retention 1,17,37,59,85,94,96,97 , and kinase activation being reported to facilitate retention w x 68,92,97 . Changes in phosphorylative enzyme activity and location and the phosphorylation state of neuronal proteins have also been shown to accompany memory w x formation 4,11,66,84,95 . Most studies investigating a role for phosphorylation in memory have focused exclusively on the potential involve- ment of protein kinases, known to be highly regulated ) Ž. Corresponding author. Fax: q61 3 905-3948. enzymes activated directly in response to second-mes- w x senger stimulation 78 . Phosphorylation is a dynamic and inherently reversible process, however, and phosphatases are also highly regulated enzymes which contribute ac- w x tively to signal transduction processes 8,83 . This may be of particular importance since reversibility in synaptic plasticity is widely acknowledged to be essential if current network models of learning and memory formation are to possess sufficient flexibility to serve as information stor- w x age devices 52,58,80 . Because phosphorylation offers a mechanism particularly well suited for conferring such reversibility and flexibility, it is of considerable interest that the wealth of information implicating kinase activities w x in the induction and maintenance of LTP 88 has now been joined by studies demonstrating that the inhibition of phosphatase activities enhances some forms of activity-de- w x pendent synaptic transmission 19,26,43 and prevents the w x induction of LTD in both visual cortex 47,89 and CA1 w x hippocampal pyramidal cells 61,62 . Further, habituation and associative learning have been reported to be disrupted in a Drosophila mutant deficient in one form of protein wx phosphatase 3 . Evidence such as this indicates that phos- 0006-8993r96r$15.00 Copyright q 1996 Elsevier Science B.V. All rights reserved. Ž . PII S0006-8993 96 00441-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ž .Brain Research 730 1996 107–117
Research report
Cyclosporin A, an inhibitor of calcineurin, impairs memory formation inday-old chicks
Pauleen C. Bennett a, Weiqin Zhao a, Alfons Lawen b, Kim T. Ng a,)
a Department of Psychology, Monash UniÕersity, Victoria, Australiab Department of Biochemistry and Molecular Biology, Monash UniÕersity, Victoria, Australia
Accepted 2 April 1996
Abstract
Considerable evidence exists that changes in the phosphorylation state of neuronal proteins are correlated with learning and thatinhibition of various protein kinases disrupts memory formation. Given the reversible nature of protein phosphorylation, a role for proteinphosphatases in memory processing also seems likely. It has been shown recently that administration of the phosphatase inhibitor, okadaic
w xacid, disrupts memory formation in day-old chicks, with retention deficits first appearing at approximately 40 min post-training 93 . Inthe present study the intracranial administration of the immunosuppressant cyclosporin A was also found to produce retention deficits inday-old chicks trained on a single-trial, passive-avoidance task, but the deficits were not significant until 85 min post-training. Thedifference could not be attributed to differences in the pharmacokinetics of the drugs. Since okadaic acid preferentially inhibits proteinphosphatases 1 and 2A, while cyclosporin A is reported to inhibit only the Ca2qrcalmodulin-dependent protein phosphatase, calcineurin,it is possible that different phosphatases may be involved in distinct stages of memory formation, as has been reported previously forprotein kinases. The possibility that cyclosporin A may, in addition, act through inhibition of cyclophilin’s peptidyl-prolyl-cisr trans-isomerase activity is also canvassed.
Keywords: Calcineurin; Cyclosporin A; Dephosphorylation; Immunosuppressant; Memory; Phosphatase
1. Introduction
Protein phosphorylation has been indirectly implicatedin biological information storage processes via its roles inmany neuronal events which contribute to signal transduc-
w xtion 90 , and through its roles in long-term potentiationŽ . w x Ž . w xLTP 88 , long-term depression LTD 61 , and neuronal
w xkindling 34,91 . It has also been directly implicated inmemory formation in many species, with kinase disruptionvia pharmacological or genetic means being consistently
w xreported to impair retention 1,17,37,59,85,94,96,97 , andkinase activation being reported to facilitate retentionw x68,92,97 . Changes in phosphorylative enzyme activityand location and the phosphorylation state of neuronalproteins have also been shown to accompany memory
w xformation 4,11,66,84,95 .Most studies investigating a role for phosphorylation in
memory have focused exclusively on the potential involve-ment of protein kinases, known to be highly regulated
) Ž .Corresponding author. Fax: q61 3 905-3948.
enzymes activated directly in response to second-mes-w xsenger stimulation 78 . Phosphorylation is a dynamic and
inherently reversible process, however, and phosphatasesare also highly regulated enzymes which contribute ac-
w xtively to signal transduction processes 8,83 . This may beof particular importance since reversibility in synapticplasticity is widely acknowledged to be essential if currentnetwork models of learning and memory formation are topossess sufficient flexibility to serve as information stor-
w xage devices 52,58,80 . Because phosphorylation offers amechanism particularly well suited for conferring suchreversibility and flexibility, it is of considerable interestthat the wealth of information implicating kinase activities
w xin the induction and maintenance of LTP 88 has nowbeen joined by studies demonstrating that the inhibition ofphosphatase activities enhances some forms of activity-de-
w xpendent synaptic transmission 19,26,43 and prevents thew xinduction of LTD in both visual cortex 47,89 and CA1
w xhippocampal pyramidal cells 61,62 . Further, habituationand associative learning have been reported to be disruptedin a Drosophila mutant deficient in one form of protein
w xphosphatase 3 . Evidence such as this indicates that phos-
0006-8993r96r$15.00 Copyright q 1996 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 96 00441-6

( )P.C. Bennett et al.rBrain Research 730 1996 107–117108
phatases may contribute to memory formation in vertebratespecies.
In a recent paper we reported that the cell-permeableŽ .marine toxin okadaic acid OKAD , a well-established
w xinhibitor of phosphatase activities 6,87 , impairs retentionin day-old chicks trained on a single-trial passive-avoi-
w xdance task 93 . Significant retention deficits were firstobserved some 40–50 min post-training, during the secondof a three stage sequence of memory formation previouslyestablished for this behavioural task with this speciesw x29,64,67 . OKAD is known to inhibit three of four majorphosphatase classes in vitro, completely inhibiting protein
Ž .phosphatase 2A PP2A at very low nanomolar concentra-Ž . Ž .tions 1 nM in dilute assays , protein phosphatase 1 PP1
at 100 times higher nanomolar concentrations, and cal-Ž . Ž 2q .cineurin CaN , the calciumrcalmodulin Ca rCaM -de-
pendent protein phosphatase 2B, at micromolar concentra-w xtions 41 . Since our results were obtained using nanomolar
concentrations, it was concluded that the inhibition ofPP2A or PP1 was more likely to be responsible for theobserved amnestic effect than was the inhibition of CaN.However, because it is difficult to predict in vivo drug
w xpotencies 41,57 , the possibility that the amnestic effectsof OKAD may be due to inhibition of CaN cannot bedismissed. Further, it remains possible that more than onephosphatase class is involved in information storage pro-cesses.
Ž .The immunosuppressant cyclosporin A CsA has beenshown to exert its immunosuppressive effect via the inhibi-
w xtion of CaN 13,21,23,54,55 . CsA binds to a distinctfamily of endogenous receptor proteins, collectively calledcyclophilins, forming immunosuppressant-cyclophilincomplexes which potently and selectively inhibit CaN. Thecyclophilins are ubiquitously distributed in brain tissue,being particularly prominent in granule cells of the cere-bellum and in all areas of the hippocampus and dentate
w xgyrus 15 . Since this distribution means that they tend tobe colocalized with CaN, the cell permeable CsA can beused as a probe to examine neuronal roles for this phos-phatase, various isoforms of which are also concentrated in
w xparticular brain tissues 12,35 . To date, most reportedactions of CsA appear to be mediated by the inhibition of
w xCaN’s phosphatase activity 18,60,65,76,77 .CaN is the only phosphatase known to be activated in
direct response to second messengers thought to mediatew xsignal transduction processes 48 . Further, although CaN
has a far more restricted range of substrates than othermajor phosphatases, it is known to dephosphorylate arange of important neuronal substrates, including cyto-
w x w xskeletal proteins 36 , other phosphorylative enzymes 7 ,Ž . w xN-methyl-D-aspartate NMDA channel constituents 51
and the protein kinase C substrates neurogranin, MARCKSŽ .myristoylated alanine-rich C kinase substrate and GAP-43Žgrowth-associated protein 43; also called neuromodulin,
. w xprotein F1, B-50 56,74,81 , the latter of which has beenstrongly implicated in memorial processes in the chick
w x10,95 . CaN also dephosphorylates the endogenous PP1Žinhibitors inhibitor-1 and DARPP-32 dopamine- and cyclic
. w xAMP-regulated phosphoprotein, M s32 000 42 . Sincev
the inhibition of PP1 by these enzymes depends on theirown phosphorylation by a cAMP-dependent protein ki-nase, the Ca2qrCaM-dependent activation of CaN tends toinitiate a cascade of dephosphorylation events antagonistic
w xto those mediated by cAMP 42,49 .CaN is highly concentrated in membrane fractions from
kindled rat brain, and administration of CsA reversiblyw xblocks kindling stage progression in these animals 60 . In
addition, CaN is believed to play a particularly importantrole in LTP and LTD. It has been suggested that, althoughboth CaN and the Ca2qrCaM-dependent kinase, CaMK-II,are activated in response to the same second-messenger,CaN may be activated more rapidly and in response to
2q w x w xlower levels of Ca than is CaMK-II 49 . Lisman 53has argued that activity-dependent alterations in intra-cellular Ca2q concentrations may differentially affect theactivity of CaMK-II and CaN, leading to changes in thephosphorylation state of key synaptic molecules and, hence,to subtle and bi-directional changes in synaptic efficacy.
w xConsistent with this are reports that both CaN 38,39 andw x 2qCaMK-II 24 appear to be activated in response to Ca
influx following NMDA receptor stimulation and that, inslices of rat visual cortex, the same tetanic stimulation canresult in either LTD or LTP, with LTP being induced whenthe level of depolarization in the postsynaptic cell exceedsa threshold related to NMDA receptor-gated conductances,and LTD being induced when this threshold is not ex-
w xceeded 2 . Further, while the induction of some forms ofLTP is disrupted by the administration of the CaMK-II
w xinhibitor KN-62 46 and promoted by the administrationw xof the CaN inhibitor FK 506 26 , the induction of LTD is
disrupted following the inhibition of CaN by either FK 506w xor CsA 62,89 but promoted following the inhibition of
w xCaMK-II by KN-62 25 . Thus, CaN potentially plays anessential role in activity-dependent modulation of synapticefficacy.
There is as yet no direct evidence of a role for CaN inmemory processing per se. In this paper we report theeffects of inhibiting CaN activity with CsA on memoryprocessing in neonate chicks trained on a single-trial pas-sive-avoidance task. The results are compared with those
Ž .of cyclosporin H CsH , an analogue of CsA that does notw xbind to cyclophilin 40 .
2. Materials and methods
2.1. Animals and housing conditions
Day-old black-Australorp white-Leghorn cockerels, ob-tained from a local hatchery on the morning of eachexperiment, were randomly divided into groups of twenty,each group being assigned to a row of 10 open-topped
( )P.C. Bennett et al.rBrain Research 730 1996 107–117 109
Ž .wooden boxes 20=25=20 cm which were maintainedat a temperature of 26–298C by 25 W light globes sus-pended above the boxes. Chick pellets were available adlib. Chicks were housed in pairs in order to avoid isolation
w xstress 16 , one of each pair being marked with indelibleink for the purposes of identification. In those experimentswhere chicks were required to be housed overnight, eachpair was given fresh water and food before the boxes werecovered at night to keep out excessive light. The water andcovers were removed several hours prior to testing thechicks on the following day.
2.2. BehaÕioral procedures
The single-trial, passive-avoidance learning task is de-w xscribed in detail elsewhere 63 . Briefly, after being left to
become accustomed to the laboratory for approximately 30min, each pair of chicks was presented with a small
Ž .chrome bead 2 mm diam. , mounted on a thin metal rodand coated with water. Chicks tend to peck spontaneouslyat small shiny objects and most did so within a 15–20-speriod, continuing to peck because the water coating thebead is reinforcing. Approximately 30 mins after the firsttrial, the water-coated chrome bead was presented again toeach pair of chicks for approximately 10 s. This wasfollowed some 30 min later by two further pretrainingtrials using, first, a small red glass bead and, second, a
Ž .small blue glass bead each 4 mm diam. . Each of thesewater-coated beads was presented for 10 s, with the num-ber of pecks for each chick being recorded on an on-linecomputer. Approximately 30 min after the final pretrainingtrial, each pair of chicks was presented with a red beadidentical to that used in the pretraining trial, but coated
Ž .with an aversant chemical, methylanthranilate MeA . Al-Ž .most all chicks )90% spontaneously pecked the aver-
sive training bead, typically responding with disgust reac-tions such as shaking their heads vigorously and wipingtheir beaks on the floor of the box. Again the number ofpecks was recorded for each chick. Retention tests werecarried out at various times after the training trial, involv-ing successive presentation of a dry red and a dry bluebead for 10 s each. The number of pecks for each chickwas again recorded. Whenever possible the test trials werecarried out by an experimenter blind to both the drugtreatment and to such variables as the time of training ordrug administration. Twenty different chicks were used foreach data point on each day of experimentation.
2.3. Drugs and injections
Ž .CsA and CsH kindly donated by Sandoz Ltd. wereŽ .dissolved in either dimethyl sulfoxide DMSO or ethanol
before being diluted in 154 mM NaCl to theŽ .concentration s required for each experiment. On all occa-
sions 10 ml per hemisphere of the appropriate concentra-tion or, for control purposes, an equivalent concentration
of either the DMSO or EtOH vehicle, again diluted in 154nM NaCl, was administered freehand into theneostriatalrhyperstriatal region of the forebrain at the re-quired time relative to training. This area of the chick brainis known to be metabolically active following passive-
w xavoidance training 73,79 . All injections were made usinga Hamilton repeating dispenser syringe with a 27-gaugeneedle fitted with a plastic stop to regulate injection depthto 3.5 mm. The accuracy of needle placements was ran-domly checked by histological examination.
2.4. Data analysis
Chicks which failed to peck the aversive red beadduring the training trial were excluded from subsequentstatistical analyses, as were chicks which failed to peck theblue bead during the 10-s retention test. This removedchicks which failed to train, as well as those that may havefailed to peck the red bead during the test trial because ofnonspecific performance deficits. Generally, no more than
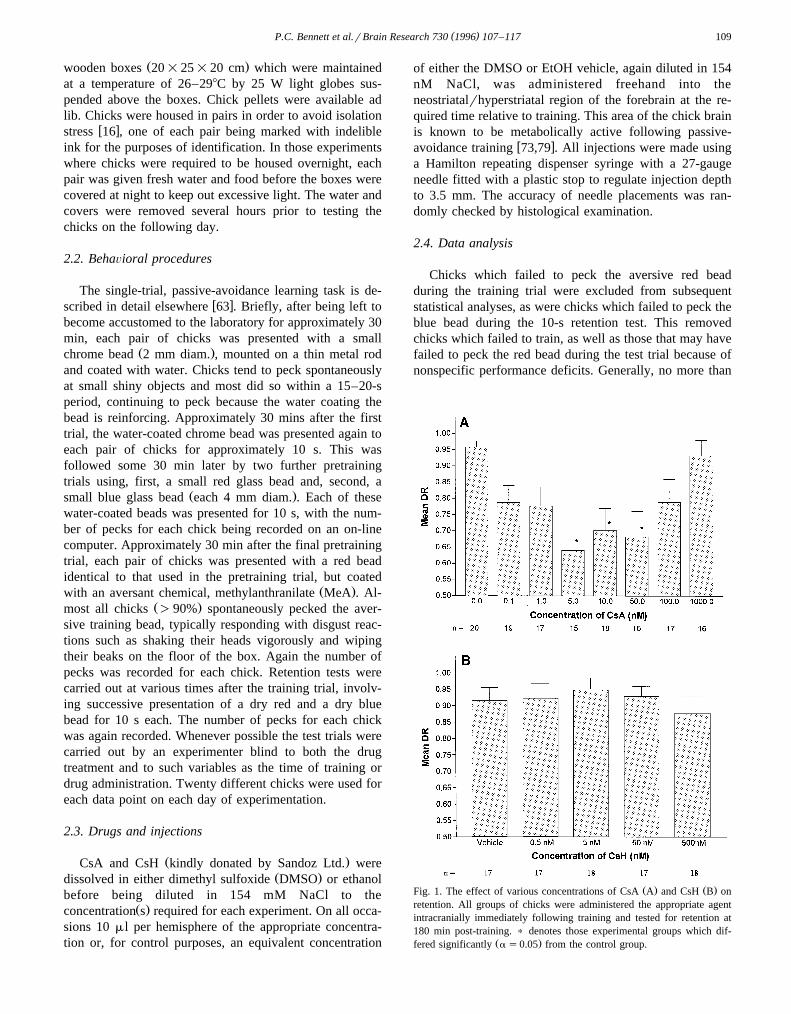
Ž . Ž .Fig. 1. The effect of various concentrations of CsA A and CsH B onretention. All groups of chicks were administered the appropriate agentintracranially immediately following training and tested for retention at180 min post-training. ) denotes those experimental groups which dif-
Ž .fered significantly a s0.05 from the control group.
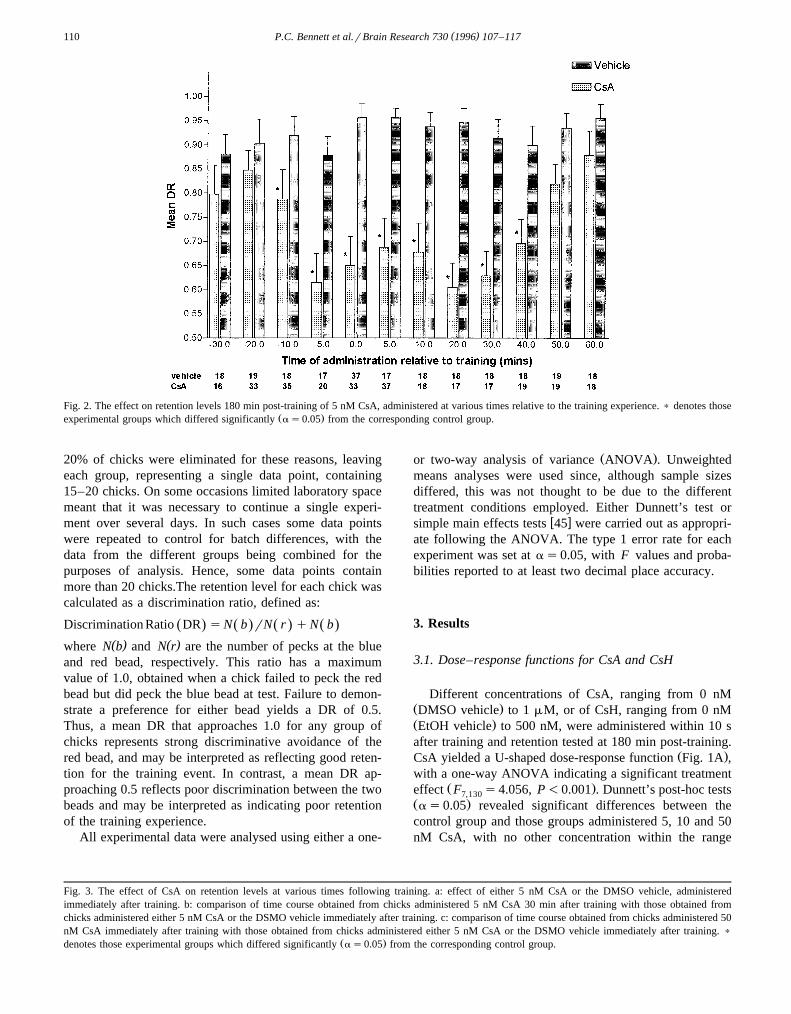
( )P.C. Bennett et al.rBrain Research 730 1996 107–117110
Fig. 2. The effect on retention levels 180 min post-training of 5 nM CsA, administered at various times relative to the training experience. ) denotes thoseŽ .experimental groups which differed significantly as0.05 from the corresponding control group.
20% of chicks were eliminated for these reasons, leavingeach group, representing a single data point, containing15–20 chicks. On some occasions limited laboratory spacemeant that it was necessary to continue a single experi-ment over several days. In such cases some data pointswere repeated to control for batch differences, with thedata from the different groups being combined for thepurposes of analysis. Hence, some data points containmore than 20 chicks.The retention level for each chick wascalculated as a discrimination ratio, defined as:
Discrimination Ratio DR sN b rN r qN bŽ . Ž . Ž . Ž .( ) ( )where N b and N r are the number of pecks at the blue
and red bead, respectively. This ratio has a maximumvalue of 1.0, obtained when a chick failed to peck the redbead but did peck the blue bead at test. Failure to demon-strate a preference for either bead yields a DR of 0.5.Thus, a mean DR that approaches 1.0 for any group ofchicks represents strong discriminative avoidance of thered bead, and may be interpreted as reflecting good reten-tion for the training event. In contrast, a mean DR ap-proaching 0.5 reflects poor discrimination between the twobeads and may be interpreted as indicating poor retentionof the training experience.
All experimental data were analysed using either a one-
Ž .or two-way analysis of variance ANOVA . Unweightedmeans analyses were used since, although sample sizesdiffered, this was not thought to be due to the differenttreatment conditions employed. Either Dunnett’s test or
w xsimple main effects tests 45 were carried out as appropri-ate following the ANOVA. The type 1 error rate for eachexperiment was set at as0.05, with F values and proba-bilities reported to at least two decimal place accuracy.
3. Results
3.1. Dose–response functions for CsA and CsH
Different concentrations of CsA, ranging from 0 nMŽ .DMSO vehicle to 1 mM, or of CsH, ranging from 0 nMŽ .EtOH vehicle to 500 nM, were administered within 10 safter training and retention tested at 180 min post-training.
Ž .CsA yielded a U-shaped dose-response function Fig. 1A ,with a one-way ANOVA indicating a significant treatment
Ž .effect F s4.056, P-0.001 . Dunnett’s post-hoc tests7,130Ž .as0.05 revealed significant differences between thecontrol group and those groups administered 5, 10 and 50nM CsA, with no other concentration within the range
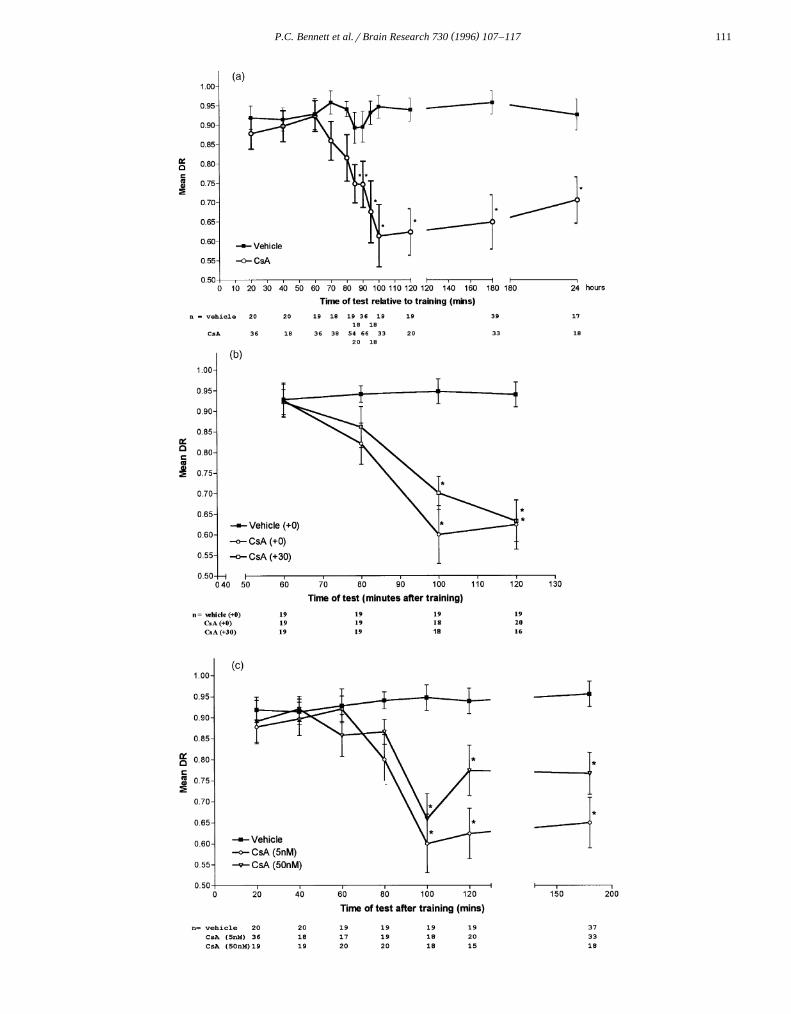
Fig. 3. The effect of CsA on retention levels at various times following training. a: effect of either 5 nM CsA or the DMSO vehicle, administeredimmediately after training. b: comparison of time course obtained from chicks administered 5 nM CsA 30 min after training with those obtained fromchicks administered either 5 nM CsA or the DSMO vehicle immediately after training. c: comparison of time course obtained from chicks administered 50nM CsA immediately after training with those obtained from chicks administered either 5 nM CsA or the DSMO vehicle immediately after training. )
Ž .denotes those experimental groups which differed significantly as0.05 from the corresponding control group.
( )P.C. Bennett et al.rBrain Research 730 1996 107–117 111

( )P.C. Bennett et al.rBrain Research 730 1996 107–117112
tested resulting in significant retention deficits. In contrastto CsA, CsH appeared to have no effect on retention levels
Ž .at any of the concentrations used Fig. 1B , with a one-wayŽANOVA showing no significant treatment effect F s4,82
.0.433, Ps0.784 .
3.2. Time of administration function for CsA
To determine when, relative to training, CsA had to beŽ .administered in order to affect retention, CsA 5 nM or
the DMSO vehicle was administered to groups of chicks atvarious times either before or after the training trial, withretention being tested approximately 180 min post-trainingŽ .Fig. 2 . A two-way ANOVA revealed a significant main
Ž .effect for drug condition F s121.97, P-0.001 and1,493
a significant interaction effect between drug condition andŽ .time of administration F s2.77, P-0.01 . Simple11,493
main effects analyses confirmed that retention levels inŽ .experimental groups were significantly as0.05 lower
than those in corresponding control groups at all times ofdrug administration between y10 min and q40 mininclusive. Differences across time of administration weresignificant for cyclosporin A-treated groups of chicks butnot for vehicle-treated groups.
3.3. Retention functions for CsA
Ž .CsA 5 nM or the DMSO vehicle was administeredimmediately after training and retention tested after vari-ous intervals. Retention deficits in CsA-treated chicks wereapparent after 60 min post-training, with marked differ-ences between control and experimental groups remaining
Ž .evident up to 24 h later Fig. 3a . A two-way ANOVAŽrevealed significant main effects for drug condition F1,628
. Žs92.03, P-0.001 and time of test F s3.23, P-11,628. Ž0.001 and a significant interaction effect F s3.91,11,628
.P-0.001 . Simple main effects tests confirmed that thereŽ .were significant differences as0.05 between control
and experimental groups at all training-test intervals from85 min to 24 h. Differences in retention levels weresignificant across training-test intervals for groups admin-istered CsA but not for those administered the DMSOvehicle.
To determine whether the time of appearance of reten-tion deficits was due simply to the time at which CsA wasadministered, 5 nM CsA was administered to chicks 30min after training and retention tested at various timesafter drug administration. Because of ethical constraints onthe unnecessary repetition of data points, the data obtainedfrom the corresponding CsA- and DMSO-treated groups inthe previous experiment were used for comparison. Thiswas thought to be acceptable since the DSMO vehicleshowed no significant effect on retention levels, regardless
Žof the time of administration relative to training see Fig..2 . The retention functions for the two different times of
CsA administration were virtually identical across the
Ž .training-retention intervals sampled Fig. 3b . A two-wayANOVA revealed significant main effects for drug condi-
Ž . Žtion F s21.23, P-0.001 , and time of test F s2,222 3,222.12.85, P-0.001 , and a significant interaction effect
Ž .F s3.91, P-0.001 . Post hoc simple main effects6,222Ž .analyses revealed a significant as0.05 time of test
effect for chicks administered CsA either immediatelyafter training or at 30 min post-training, but not for chicksgiven DMSO. Significant simple main effects for drugcondition were obtained for the 100 and 120 min training-test interval.
To determine whether the time at which retentiondeficits emerged may depend on the concentration of CsAused, chicks were administered 50 nM CsA immediately
Žafter training and tested at various times post-training Fig..3c . For reasons stated above, the retention function for
these chicks was compared with the corresponding CsAand DMSO functions in Fig. 3a. A two-way ANOVA
Žyielded significant main effects for drug condition F2,370. Žs28.83, P-0.001 and time of test F s6.88, P-6,370
. Ž0.001 and a significant interaction effect F s3.03,12,370.P-0.001 , with simple time of test main effects beingŽ .significant as0.05 for 5 nM CsA-treated and 50 nM
CsA-treated chicks, but not for DMSO-treated chicks.Significant simple drugs main effects were obtained for the100, 120 and 180 min training-test intervals. While the 50nM CsA-treated chicks tended to show higher levels of
Ž .retention than the 5 nM CsA-treated chicks see Fig. 1A ,Newman–Keuls post-hoc tests indicate that these differ-ences were not significant.
3.4. RetrieÕal processes
It is possible that the effects of CsA may have been onretrieval processes rather than on memory formation as
Fig. 4. The effect on retention levels one day following training of either5 nM CsA or the DMSO vehicle administered 2 h prior to test. )
indicates that retention levels in both groups receiving CsA immediatelyŽ .after training on the first day differed significantly a s0.05 from those
receiving DMSO at the same time. The drug condition at time of test didnot significantly affect retention levels.

( )P.C. Bennett et al.rBrain Research 730 1996 107–117 113
such. To determine if this was the case, four groups of 20chicks were trained on one day, with two groups receivinga 5 nM concentration of CsA immediately after trainingand the other two groups receiving the DMSO vehicle. Onthe following day one of the drug-treated groups wasadministered another 5 nM dose of CsA, with the otherbeing administered the DMSO vehicle. The two vehicle-treated groups from the previous day were also adminis-tered either CsA or the DMSO vehicle. All groups weretested for retention approximately 2 h after drug adminis-tration. The results show that both groups administeredCsA immediately after training demonstrated poorer reten-tion than the control groups when tested the next day,
Žregardless of the drug administered 2 h prior to test Fig..4 . A two-way ANOVA yielded a significant main effect
Žfor the drug condition on the first day F s7.69,1,57.P-0.01 . Since there was no significant main effect for
the drug condition at the time of test on the second dayŽ .F s0.13, Ps0.715 and no significant interaction1,57
Ž .between the two variables F s0.56, Ps0.459 , post-1,57
hoc tests of simple main effects were not conducted.
4. Discussion
Our results clearly demonstrate that CsA impairs reten-tion in chicks trained on a single-trial passive-avoidancetask. The effect was dose-dependent, most prominent whenadministration took place between 10 min before and 40min after learning, and did not reach significance until atleast 85 min post-training whether 5 nM CsA was givenimmediately or 30 min post-training, or 50 nM CsA wasgiven immediately after training. Importantly, the memoryimpairment induced by CsA persisted for at least 24 hpost-training and was not apparent in chicks trained on oneday and administered the drug two hours prior to test onthe second day. Hence, it would appear that CsA selec-tively and permanently disrupts a relatively late stage ofmemory formation in the chick. Further, given the failureof CsH to induce similar effects, non-specific drug effectsare unlikely to account for the observed retention deficits.
If the amnestic effects of CsA are indeed mediated byits inhibition of CaN, it is of interest to contrast theseeffects with those reported previously for the less specific
Ž .phosphatase inhibitor, okadaic acid OKAD , particularlysince, while the retention deficits induced by CsA did notbecome significant until 85 min post-training, those in-duced by OKAD were significant by 40–50 min post-train-
w xing 93 . Memory for the passive avoidance task in thechick has been shown to involve three behaviorally andpharmacologically distinct, but sequentially dependent,
Ž .stages: a short-term stage STM , formed by 5 min anddecaying after 10 min post-training; an intermediate-term
Ž .stage ITM , formed by 20 min and decaying after 50 minŽ .post- training; and a long-term stage LTM , available by
w x60 min post-training 29,64,67 . Hence, within the frame-
work of this model, OKAD and CsA would appear todisrupt ITM and LTM, respectively. This supports ourearlier conclusion that the amnestic effects induced bynanomolar concentrations of OKAD were most probablydue to inhibition of PP1 andror PP2A, rather than CaN, aconclusion also consistent with the fact that OKAD wasineffective when administered after 10 min post-training,while CsA induced retention deficits when given as late as40 min post-training.
Together, the results from the use of OKAD and CsAsuggest that phosphatases may play at least two temporallydistinct roles in memory formation. Although this has notpreviously been demonstrated, a dual role for phosphatasesin memory formation would be consistent with reports thatthe pharmacological inhibition of different kinases alsoselectively disrupts quite distinct stages of complex pro-
w xcesses such as LTP 69,88 and of memory formationitself. In the chick, inhibition of CaMK II, protein kinase Cor protein kinase A has been shown to result in retentiondeficits during STM, ITM, and LTM, respectivelyw x Ž w x.94,96,97 see also ref. 82 . Preliminary results from ourlaboratory also indicate that blockade of tyrosine-kinaseactivity by genistein induces significant retention losses
Ž .during LTM Whitechurch et al., in prep. . It would seem,therefore, that memory formation may involve a complexcascade of phosphorylative events, each mediated by adifferent enzyme class, each occurring at a specific timefollowing learning, and each essential for the eventualformation of a relatively permanent memory trace.
The nature of the cellular memory mechanisms dis-rupted by the postulated inhibition of CaN in the presentexperiments remains unclear. CaN appears to be activated
w xin response to NMDA-receptor stimulation 38,39 , how-ever, and to be involved in the induction of LTD in CA1
w x w xhippocampal pyramidal cells 62 and visual cortex 89 .Administration of less specific phosphatase inhibitors thanCsA prevents the induction of LTD in CA1 hippocampalpyramidal cells, presumably by inhibiting PP1 andror
w xPP2A 61 . Since these phosphatases are not known to bedirectly regulated by recognized second-messenger sys-tems, and since changes in synaptic transmission of thetype modelled by LTP and LTD are generally dependent
w xon increased concentrations of intracellular calcium 9 , itis possible that PP1 may decrease the strength of synaptictransmission in vitro after being activated in response to aCa2qrCaM-dependent stimulation of CaN activity and thesubsequent dephosphorylation of endogenous PP1 in-hibitors such as Inhibitor-1 or DARPP-32. This is consis-tent with a recent demonstration that inhibition of CaNalso prevents the induction of LTD in CA1 hippocampal
w xpyramidal cells 62 .The relevance of processes like LTP and LTD to mem-
ory processing in the chick remains to be unequivocallydemonstrated and the present study provides no directevidence that CsA impairs memory via the inhibition ofPP1 and the disruption of an LTD-like phenomenon. It is

( )P.C. Bennett et al.rBrain Research 730 1996 107–117114
of interest, however, that several different pharmacologicalagents have now been found to disrupt memory formationin the chick after the formation, at approximately 60 minpost-training, of the long-term, protein synthesis-dependent
w xstage which was previously 29 assumed to be relativelypermanent. More importantly, perhaps, many of these sameagents have previously been implicated in LTP. Hence, inour own laboratory, it has been found that NMDA, amino-
Ž .3-hydroxy-5-methylisoxazole-4-propionic acid AMPAand metabotropic glutamate receptor antagonists impairmemory formation, with retention deficits induced by thesedrugs first becoming apparent at 80–90 min post-trainingw x71,72 . Tyrosine kinase inhibitors have recently been found
Ž .to have a similar effect Whitechurch et al., in prep. and,from an independent laboratory using a similar trainingmodel, it has also been reported that an inhibitor ofarachidonic acid synthesis induces amnesia at 75 min
w xpost-training 44 . It is therefore becoming clear that whilethe original three-stage model of memory in the chickremains a useful heuristic within which to interpret newexperimental results, the actual memory formation processcontinues beyond 60 min and may perhaps involve anLTPrLTD-like modification of synaptic plasticity at ap-proximately 80–90 min.
At this stage, any conclusion that the amnestic effectsof CsA are due to inhibition of CaN must remain some-what speculative. To date, most studies using CsA toinvestigate cellular events report that this agent acts pri-
w xmarily via the inhibition of CaN 18,60,65,76,77 . For therecently described function of CsA as a substance P recep-tor antagonist, however, concentrations of CsA about twoorders of magnitude higher than those required to inhibit
w xCaN are necessary 30 . Inhibition of phosphatase activity,therefore, appears unlikely to account for the effects de-scribed in this study. CsA also inhibits the peptidyl-prolyl
Ž . Ž .cisr trans isomerase PPIase activity of cyclophilin s ,which have been postulated to be involved in protein
w xfolding 20,22,27,28,50,75 . Because cyclophilins are ubiq-uitously distributed in brain tissue and have no known
w xassociation with CaN apart from colocalization 15 , theymay be expected to have important roles independently oftheir pharmacologically-induced inhibition of CaN activity.Indeed, a small number of CsA-induced effects have been
w xreported to be independent of CaN inhibition 5,70,86 .Such observations support claims that the catalysis ofprotein folding by cyclophilins may serve a critical but as
w xyet unidentified cellular function 20,22 and make it im-possible to rule out the inhibition of PPIase activity as afactor in the CsA disruption of memory processing re-ported here. Indeed a role for PPIase activity in informa-tion storage processes may not be surprising given asubstantial body of literature demonstrating that proteinsynthesis plays an essential role in the formation of mem-
w xory in many species 14 . Since the biphasic effect ofcyclosporin A revealed in our dose-response study is con-sistent with the drug having more than one effect, this
possibility is currently being investigated with agents whichinhibit PPIase activity without affecting CaN activity, andby experiments measuring both PPIase activity and CaNactivity in the chick brain at various times followingadministration of CsA. It should be noted, however, thatmany agents produce a U-shaped dose effect on memory
w xformation, see for example 32,72 . The reason for this isnot known, but it has been a common finding since initialstudies using rodents reported that the level of arousalassociated with a training experience, or the administrationof arousal-associated hormones, produces a dose-depen-
w xdent, U-shaped, retention function 31–33 . It may there-fore reflect homeostatic mechanisms which protect theorganism against excessive stress, whether it be caused byarousal or by the administration of high concentrations ofsome pharmacological agents.
In sum, intracranial administration of cyclosporin Asignificantly disrupts memory formation in chicks trainedon a single-trial passive-avoidance task. The results sug-gest multiple roles for protein phosphatases in memoryformation. Inhibition of PP1 andror PP2A activity by themarine toxin OKAD disrupts an early, transient, stage inthe sequence of events leading to permanent information
w xstorage 93 . In contrast, CsA appears to disrupt memoryformation during a later, relatively stable, protein synthe-sis-dependent stage, possibly at least partially throughinhibition of calcineurin activity. The precise mechanismsunderlying each of these pharmacological effects remain tobe unequivocally established, but there seems little doubtthat protein phosphatases, like protein kinases, play essen-tial roles in the molecular mechanisms underlying informa-tion storage in vertebrate species.
Acknowledgements
We gratefully acknowledge the support of the Aus-tralian Research Council in this project, and the generosityof Sandoz Ltd. in donating CsA and CsH. The authorswish to thank the Department of Psychology, La TrobeUniversity, for allowing some of the experiments to becarried out in their laboratory.
References
w x1 Abeliovich, A., Paylor, R., Chen, C., Kim, J.J., Wehner, J.M. andTonegawa, S., PKCg mutant mice exhibit mild deficits in spatial
Ž .and contextual learning, Cell, 75 1993 1263–1271.w x2 Artola, A., Brocher, S. and Singer, W., Different voltage-dependent¨
thresholds for inducing long-term depression and long-term potentia-Ž .tion in slices of rat visual cortex, Nature, 347 1990 69–72.
w x3 Asztalos, Z., von Wegerer, J., Wustmann, G., Dombradi, V., Gausz,´J., Spatz, H.-C. and Friedrich, P., Protein phosphatase 1-deficientmutant Drosophila is affected in habituation and associative learn-
Ž .ing, J. Neurosci., 13 1993 924–930.w x4 Bank, B., DeWeer, A., Kuzirian, A.M., Rasmussen, H. and Alkon,
D.L., Classical conditioning induces long-term translocation of pro-

( )P.C. Bennett et al.rBrain Research 730 1996 107–117 115
tein kinase C in rabbit hippocampal CA1 cells, Proc. Natl. Acad.Ž .Sci. USA, 85 1988 1988–1992.
w x5 Bell, A., Wernli, B. and Franklin, R.M., Roles of peptidyl-prolylcis-trans isomerase and calcineurin in the mechanisms of antimalar-ial action of cyclosporin A, FK506 and rapamycin, Biomed. Phar-
Ž .macol., 48 1994 495–503.w x6 Bialojan, C. and Takai, A., Inhibitory effect of a marine-sponge
toxin, okadaic acid, on protein phosphatases, Biochem. J., 256Ž .1988 283–290.
w x7 Blumenthal, D.K., Takio, K., Hansen, R.S. and Krebs, E.G.,Dephosphorylation of a cAMP- dependent protein kinase regulatory
Ž .subunit type II by calmodulin-dependent protein phosphatase, J.Ž .Biol. Chem., 261 1986 8140–8145.
w x8 Brautigan, D.L., Protein phosphatases, Rec. Prog. Horm. Res., 49Ž .1994 197–214.
w x9 Brocher, S., Artola, A. and Singer, W., Intracellular injection ofCa2q chelators blocks induction of long-term depression in rat
Ž .visual cortex, Proc. Natl. Acad. Sci. USA, 89 1992 123–127.w x10 Bullock, S., De Graan, P.N.E., Oestreicher, A.B., Gispen, W.H. and
Rose, S.P.R., Identification of a 52 kDa chick brain membraneprotein showing changed phosphorylation after passive avoidance
Ž . Ž .training as B-50 GAP-43 , Neurosci. Res. Commun., 6 1990181–186.
w x11 Burchuladze, R., Potter, J. and Rose, S.P.R., Memory formation inthe chick depends on membrane-bound protein kinase C, Brain Res.,
Ž .535 1990 131–138.w x12 Chung, E., Dvorozniak, M.T., Van Woert, M.H. and Li, H.C.,
Regional distribution of calciumrcalmodulin-dependent phosphataseactivity of calcineurin in rat brain, Res. Commun. Chem. Pathol.
Ž .Pharmacol., 64 1989 357–371.w x13 Clipstone, N.A. and Crabtree, G.R., Identification of calcineurin as a
key signalling enzyme in T-lymphocyte activation, Nature, 357Ž .1992 695–697.
w x14 Davis, H.P. and Squire, L.R., Protein synthesis and memory: aŽ .review, Psychol. Bull., 96 1984 518-559.
w x15 Dawson, T.M., Steiner, J.P., Lyons, W.E., Fotuhi, M., Blue, M. andSnyder, S.H., The immunophilins, FK506 binding protein and cy-clophilin, are discretely localized in the brain: relationship to cal-
Ž .cineurin, Neuroscience, 62 1994 569–580.w x16 De Vaus, J.E., Gibbs, M.E. and Ng, K.T., Effects of social isolation
Ž .on memory formation, BehaÕ. Neural Biol., 29 1980 473–480.w x17 Drain, P., Folkers, E. and Quinn, W.C., cAMP-dependent protein
kinase and the disruption of learning in transgenic flies, Neuron, 6Ž .1991 71–82.
w x18 Enslen, H. and Soderling, T.R., Roles of calmodulin-dependentprotein kinases and phosphatase in calcium-dependent transcription
Ž .of immediate early genes, J. Biol. Chem., 269 1994 20872–20877.w x19 Figurov, A., Boddeke, H. and Muller, D., Enhancement of AMPA-
mediated synaptic transmission by the protein phosphatase inhibitorŽ .calyculin A in rat hippocampal slices, Eur. J. Neurosci., 5 1993
1035–1041.w x20 Fischer, G., Peptidyl-prolyl cisr trans isomerases and their effec-
Ž .tors, Angew. Chem. Int. Ed. Engl., 33 1994 1415–1436.w x21 Friedman, J. and Weissman, I., Two cytoplasmic candidates for
immunophilin action are revealed by affinity for a new cyclophilin:Ž .one in the presence and one in the absence of CsA, Cell, 66 1991
799–806.w x22 Fruman, D.A., Burakoff, S.J. and Bierer, B.E., Immunophilins in
Ž .protein folding and immunosuppression, FASEB J., 8 1994 391–400.
w x23 Fruman, D.A., Klee, C.B., Bierer, B.E. and Burakoff, S.J., Cal-cineurin phosphatase activity in T lymphocytes is inhibited by FK
Ž .506 and cyclosporin A, Proc. Natl. Acad. Sci. USA, 89 19923686–3690.
w x24 Fukunaga, K., Stoppini, L., Miyamoto, E. and Muller, D., Long-termpotentiation is associated with an increased activity of
Ca2qrcalmodulin-dependent protein kinase II, J. Biol. Chem., 268Ž .1993 7863–7867.
w x25 Funauchi, M., Tsumoto, T., Nishigori, A., Yoshimura, Y. andHidaka, H., Long-term depression is induced in Ca2qrcalmodulin
Ž .kinase-inhibited visual cortex neurons, NeuroReport, 3 1992 173–176.
w x26 Funauchi, M., Haruta, H. and Tsumoto, T., Effects of an inhibitorfor calciumrcalmodulin-dependent protein phosphatase, calcineurin,on induction of long-term potentiation in rat visual cortex, Neurosci.
Ž .Res., 19 1994 269–278.w x27 Galat, A., Peptidylproline cis-trans-isomerases: immunophilins, Eur.
Ž .J. Biochem., 216 1993 689–707.w x28 Galat, A. and Metcalfe, S.M., Peptidylproline cisr trans isomerases,
Ž .Prog. Biophys. Molec. Biol., 63 1995 67–118.w x29 Gibbs, M.E. and Ng, K.T., Psychobiology of memory: towards a
Ž .model of memory formation, BiobehaÕ. ReÕ., 1 1977 113–136.w x30 Gitter, B.D., Waters, D.C., Threlkeld, P.G., Lovelace, A.M., Mat-
sumoto, K. and Bruns, R.F., Cyclosporin A is a substance PŽ .tachykinin NK1 receptor antagonist, Eur. J. Pharmacol., 289Ž .1995 439–446.
w x31 Gold, P.E. and Delanoy, R.L. ACTH modulation of memory storageprocessing, In J. Martinez, R. Jensen, R. Messing, H. Rigter and J.
Ž .McGaugh Eds. , Endogenous Peptides and Learning and MemoryProcesses, Academic Press, New York, 1981, pp. 79–98.
w x32 Gold, P.E. and Van Buskirk, R., Enhancement and impairment ofmemory processes with post-trial injections of adrenocorticotrophic
Ž .hormone, BehaÕ. Biol, 16 1976 387–400.w x33 Gold, P.E., Van Buskirk, R. and Haycock, J.W., Effects of posttrain-
ing epinephrine injections on retention of avoidance training in mice,Ž .BehaÕ. Biol., 20 1977 197–204.
w x34 Goldenring, J.R., Wasterlain, C.G., Oestreicher A.B., deGraan,P.N.E., Farber, D.B., Glaser, G. and DeLorenzo, R.J., Kindlinginduces a long-lasting change in the activity of a hippocampalmembrane calmodulin-dependent protein kinase system, Brain Res.,
Ž .377 1986 47–53.w x35 Goto, S., Matsukado, Y., Mihara, Y., Inoue, N. and Miyamoto, E.,
The distribution of calcineurin in rat brain by light and electronmicroscopic immunohistochemistry and enzyme-immunoassay,
Ž .Brain Res., 397 1986 161–172.w x36 Goto, S., Yamamoto, H., Fukunaga, K., Iwasa, T., Matsukado, Y.
and Miyamoto, E., Dephosphorylation of microtubule-associatedprotein 2, t factor and tubulin by calcineurin, J. Neurochem., 45Ž .1985 276–283.
w x37 Griffith, L.C., Verselis, L.M., Aitken, K.M., Kyriacou, C.P., Danho,W. and Greenspan, R.J., Inhibition of calciumrcalmodulin-depen-dent protein kinase in Drosophila disrupts behavioral plasticity,
Ž .Neuron, 10 1993 501–509.w x38 Halpain, S., Girault, J.-A. and Greengard, P., Activation of NMDA
receptors induces dephosphorylation of DARPP-32 in rat striatalŽ .slices, Nature, 343 1990 369–372.
w x39 Halpain, S. and Greengard, P., Activation of NMDA receptorsinduces rapid dephosphorylation of the cytoskeletal protein MAP2,
Ž .Neuron, 5 1990 237–246.w x40 Handschumacher, L.E., Harding, M.W., Rice, J., Drugger, R.J. and
Speicher, D.W., Cyclophilin: a specific cytosolic binding protein forŽ .cyclosporin A, Science, 226 1984 544–547.
w x41 Hardie, D.G., Haystead, T.A.J. and Sim, A.T.R., Use of okadaicacid to inhibit protein phosphatases in intact cells, Methods Enzy-
Ž .mol., 201 1991 469–476.w x42 Hemmings, H.C. Jr. and Greengard, P., DARPP-32, a dopamine-reg-
Ž .ulated phosphoprotein, Prog. Brain Res., 69 1986 149–159.w x43 Herron, C.E. and Malenka, R.C., Activity-dependent enhancement
of synaptic transmission in hippocampal slices treated with theŽ .phosphatase inhibitor calyculin A, J. Neurosci., 14 1994 6013–
6020.w x44 Holscher, C. and Rose, S.P.R. Inhibitors of phospholipase A 2

( )P.C. Bennett et al.rBrain Research 730 1996 107–117116
produce amnesia for a passive avoidance task in the chick, BehaÕ.Ž .Neural. Biol., 61 1994 225–232.
w x45 Howell, D.C., Statistical Methods for Psychology, PWS-Kent,Boston, 1987.
w x46 Ito, I., Hidaka, H. and Sugiyama, H., Effects of KN-62, a specificinhibitor of calciumrcalmodulin-dependent protein kinase 11, onlong-term potentiation in the rat hippocampus, Neurosci. Lett., 121Ž .1991 119–121.
w x47 Kirkwood, A. and Bear, M.F., Homosynaptic long-term depressionŽ .in the visual cortex, J. Neurosci., 14 1994 3404–3412.
w x48 Klee, C.B., Draetta, G.F. and Hubbard, M.J., Calcineurin, AdÕ.Ž .Enzymol., 61 1988 149–200.
w x49 Klee, C.B., Concerted regulation of protein phosphorylation andŽ .dephosphorylation by calmodulin, Neurochem. Res., 16 1991
1059–1065.w x50 Lang, K., Schmid, F.X. and Fischer, G., Catalysis of protein folding
Ž .by prolyl isomerase, Nature, 329 1987 268–270.w x51 Lieberman, D.N. and Mody, I., Regulation of NMDA channel
function by endogenous Ca2q-dependent phosphatase, Nature, 369Ž .1994 235–239.
w x52 Linden, D.J., Long-term synaptic depression in the mammalianŽ .brain, Neuron, 12 1994 457–472.
w x53 Lisman, J., A mechanism for the Hebb and the anti-Hebb processesunderlying learning and memory, Proc. Natl. Acad. Sci. USA, 86Ž .1989 9574–9578.
w x54 Liu, J., Albers, M.W., Wandless, T.J., Luan, S., Alberg, D.G.,Belshaw, P.J., Cohen, P., MacKintosh, C., Klee, C.B. and Schreiber,S.L., Inhibition of T cell signaling by immunophilin-ligand com-plexes correlates with loss of calcineurin phosphatase activity, Bio-
Ž .chemistry, 31 1992 3896–3901.w x55 Liu, J., Farmer, J.D. Jr., Lane, W.S., Friedman, J., Weissman, I. and
Schreiber, S.L., Calcineurin is a common target of cyclophilin-Ž .cyclosporin A and FKBP-FK506 complexes, Cell, 66 1991 807–
815.w x56 Liu, Y. and Storm, D.R., Dephosphorylation of neuromodulin by
Ž .calcineurin, J. Biol. Chem., 264 1989 12800–12804.w x57 MacKintosh, C. and MacKintosh, R.W., Inhibitors of protein kinases
Ž .and phosphatases, Trends Bioc. Sci., 19 1994 444–448.w x58 Malenka, R.C., Long-term depression: not so depressing after all,
Ž .Proc. Natl. Acad. Sci. USA, 90 1993 3121–3123.w x59 Mathis, C., Lehmann, J. and Ungerer, A., The selective protein
kinase C inhibitor, NPC 15437, induces specific deficits in memoryŽ .retention in mice, Eur. J. Pharmacol., 220 1992 107–110.
w x60 Moia, L.J.M.P., Matsui, H., de Barros, G.A.M., Tomizawa, K.,Miyamoto, K., Kuwata, Y., Tokuda, M., Itano, T. and Hatase, O.,Immunosuppressants and calcineurin inhibitors, cyclosporin A andFK506, reversibly inhibit epileptogenesis in amygdaloid kindled
Ž .rats, Brain Res., 648 1994 337–341.w x61 Mulkey R.M., Herron, C.E. and Malenka, R.C., An essential role for
protein phosphatases in hippocampal long-term depression, Science,Ž .261 1993 1051–1055.
w x62 Mulkey, R.M., Endo, S., Shenolikar, S. and Malenka, R.C., Involve-ment of a calcineurinrinhibitor-1 phosphatase cascade in hippocam-
Ž .pal long-term depression, Nature, 369 1994 486–488.w x63 Ng, K.T. and Gibbs, M.E., A biological model for memory forma-
Ž .tion. In H.J. Markowitsch Ed. , Information Processing in theBrain, Hans Huber, Toronto, 1988, pp. 151–178.
w x64 Ng, K.T. and Gibbs, M.E., Stages in memory formation: a review.Ž .In R.J. Andrew Ed. , Neural and BehaÕioral Plasticity: the Use of
the Domestic Chick as a Model, Oxford University Press, Oxford,1991, pp. 351–369.
w x65 Nichols, R.A., Suplick, G.R. and Brown, J.M., Calcineurin-mediatedprotein dephosphorylation in brain nerve terminals regulates the
Ž .release of glutamate, J. Biol. Chem., 269 1994 23817–23823.w x66 Olds, J.L., Golski, S., McPhie, D.L., Olton, D., Mishkin, M. and
Alkon, D.L., Discrimination learning alters the distribution of pro-Ž .tein kinase C in the hippocampus of rats, J. Neurosci., 10 1990
3707–3713.w x67 Patterson, T.A., Alvarado, M.C., Rosenzweig, M.R. and Bennett,
E.L., Time courses of amnesia development in two areas of theŽ .chick forebrain, Neurochem. Res., 13 1986 643–647.
w x68 Paylor, R., Rudy, J.W. and Wehner, J.M., Acute phorbol estertreatment improves spatial learning performance in rats, BehaÕ.
Ž .Brain Res., 45 1991 189–193.w x69 Reymann, K.G., Brodemann, R., Kase, H. and Matthies, H., In-
hibitors of calmodulin and protein kinase C block different phases ofŽ .hippocampal long-term potentiation, Brain Res., 461 1988 388–
392.w x70 Richter, A., Davies, D.E. and Alexander, P., Growth inhibitory
effects of FK506 and cyclosporin A independent of inhibition ofŽ .calcineurin, Biochem. Pharmacol., 49 1995 367–373.
w x71 Rickard, N.S. and Ng, K.T., Blockade of metabotropic glutamatereceptors prevents long-term memory consolidation, Brain Res.
Ž .Bull., 36 1995 355–359.w x72 Rickard, N.S., Poot, A.C., Gibbs, M.E. and Ng, K.T., Both non-
NMDA and NMDA glutamate receptors are necessary for memoryŽ .consolidation in the day-old chick, BehaÕ. Neural Biol., 62 1994
33–40.w x73 Rose, S.P.R. and Csillag, A., Passive avoidance training results in
lasting changes in deoxyglucose metabolism in left hemisphereŽ .regions of chick brain, BehaÕ. Neural Biol., 44 1985 315–324.
w x74 Schrama, L.H., Heemskerk, F.M.J. and de Graan, P.N.E., Dephos-phorylation of protein kinase C phosphorylated B-50rGAP-43 bythe calmodulin-dependent phosphatase calcineurin, Neurosci. Res.
Ž .Commun., 5 1989 141–147.w x75 Schreiber, S.L., Chemistry and biology of the immunophilins and
Ž .their immunosuppressive ligands, Science, 251 1991 283–287.w x76 Schwaninger, M., Blume, R., Oetjen, E., Lux, G. and Knepel, W.,
Inhibition of cAMP-responsive element-mediated gene transcriptionby cyclosporin A and FK506 after membrane depolarization, J. Biol.
Ž .Chem., 268 1993 23111–23115.w x77 Schwaninger, M., Blume, R., Kruger, M., Lux, G., Oetjen, E. and¨
Knepel, W., Involvement of the Ca2q-dependent phosphatase cal-cineurin in gene transcription that is stimulated by cAMP through
Ž .cAMP Response Elements, J. Biol. Chem., 270 1995 8860–8866.w x78 Scott, J.D. and Soderling, T.R., Serinerthreonine protein kinases,
Ž .Curr. Opin. Neurobiol., 2 1992 289–295.w x79 Sedman, G., O’Dowd, B., Rickard, N., Gibbs, M.E. and Ng, K.T.,
Brain metabolic activity associated with long-term memory consoli-Ž .dation, Mol. Neurobiol., 5 1991 351–354.
w x80 Sejnowski, T.J., Synaptic mechanisms for long-term depression,Ž .Curr. Biol., 1 1991 38–40.
w x81 Seki, K., Chen, H.-C. and Huang, K.-P., Dephosphorylation ofprotein kinase C substrates, neurogranin, neuromodulin and MAR-CKS, by calcineurin and protein phosphatases 1 and 2A, Arch.
Ž .Biochem. Biophys., 316 1995 673–679.w x82 Serrano, P.A., Beniston, D.S., Oxonian, M.G., Rodriguez, W.A.,
Rosenzweig, M.R. and Bennett, E.L., Differential effects of proteinkinase inhibitors and activators on memory formation in the 2-day-old
Ž .chick, BehaÕ. Neural Biol., 61 1994 60–72.w x83 Shenolikar, S., Protein serinerthreonine phosphatases: new avenues
Ž .for cell regulation, Annu. ReÕ. Cell Biol., 10 1994 55–86.w x84 Sheu, F.-S., McCabe, B.J., Horn, G. and Routtenberg, A., Learning
selectively increases protein kinase C substrate phosphorylation inspecific regions of the chick brain, Proc. Natl. Acad. Sci. USA, 90Ž .1993 2705–2709.
w x85 Silva, A.J., Paylor, R., Wehner, J.M. and Tonegawa, S., Impairedspatial learning in a-calcium-calmodulin kinase II mutant mice,
Ž .Science, 257 1992 206–211.w x86 Stamnes, M.A., Shieh, B.-H., Chuman, L., Harris, G.L. and Zuker,

( )P.C. Bennett et al.rBrain Research 730 1996 107–117 117
C.S., The cyclophilin homolog ninaA is a tissue-specific integralmembrane protein required for the proper synthesis of a subset of
Ž .Drosophila rhodopsins, Cell, 65 1991 219–227.w x87 Suganuma, M., Fujiki, H., Okabe, S., Nishiwaki, S., Brautigan, D.,
Ingebritsen, T.S. and Rosner, M.R., Structurally different membersof the okadaic acid class selectively inhibit protein serinerthreonine
Ž .but not tyrosine phosphatase activity, Toxicon, 30 1992 873–878.w x88 Suzuki, T., Protein kinases involved in the expression of long-term
Ž .potentiation, Int. J. Biochem., 26 1994 735–744.w x89 Torii, N., Kamishita, T., Otsu, Y. and Tsumoto, T., An inhibitor for
calcineurin, FK506, blocks induction of long-term depression in ratŽ .visual cortex, Neurosci. Lett., 185 1995 1–4.
w x90 Walaas, S.I. and Greengard, P., Protein phosphorylation and neu-Ž .ronal function, Pharmacol. ReÕ., 43 1991 299–349.
w x91 Yamagata, Y., Obata, K., Greengard, P. and Czernik, A.J., Increasein synapsin 1 phosphorylation implicates a presynaptic component in
Ž .septal kindling, Neuroscience, 64 1995 1–4.w x92 Yang, H.C. and Lee, E.H.Y., Protein kinase C activation facilitates
Ž .memory retention in rats, Chinese J. Physiol., 36 1993 115–123.
w x93 Zhao, W.-Q., Bennett, P., Sedman, G.L. and Ng, K.T., The impair-ment of long-term memory formation by the phosphatase inhibitor
Ž .okadaic acid, Brain Res. Bull., 36 1995 557–561.w x94 Zhao, W.-Q., Bennett, P., Rickard, N., Sedman, G.L., Gibbs, M.E.
and Ng, K.T., The involvement of Ca2qrcalmodulin-dependentprotein kinase in memory formation in day-old-chicks, Neurobiol.
Ž .Learning Memory, 65 1996 in press.w x95 Zhao, W.-Q., Ng, K.T. and Sedman, G.L., Passive avoidance learn-
ing induced change in GAP43 phosphorylation in day-old chicks,Ž .Brain Res. Bull., 36 1995 11–17.
w x96 Zhao, W.-Q., Polya, G.M., Wang, B.H., Gibbs, M.E., Sedman, G.L.and Ng, K.T., Inhibitors of cAMP-dependent protein kinase impairlong-term memory formation in day-old chicks, Neurobiol. Learning
Ž .Memory, 64 1995 106–118.w x97 Zhao, W.-Q., Sedman, G.L., Gibbs, M.E. and Ng, K.T., Effect of
PKC inhibitors and activators on memory, BehaÕ. Brain Res., 60Ž .1994 151–160.
Related Documents











