The Pennsylvania State University The Graduate School Eberly College of Science CYANOBACTERIAL QUINOMICS STUDIES OF QUINONES IN CYANOBACTERIA A Thesis in Biochemistry, Microbiology, and Molecular Biology by Yumiko Sakuragi Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy August 2004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Pennsylvania State University
The Graduate School
Eberly College of Science
CYANOBACTERIAL QUINOMICS
STUDIES OF QUINONES IN CYANOBACTERIA
A Thesis in
Biochemistry, Microbiology, and Molecular Biology
by
Yumiko Sakuragi
Submitted in Partial Fulfillment of the Requirements
for the Degree of
Doctor of Philosophy
August 2004

The thesis of Yumiko Sakuragi has been reviewed and approved* by the following:
Date of Signature
Donald A. Bryant Ernest C. Pollard Professor in Biotechnology Professor of Biochemistry and Molecular Biology Thesis Advisor Co-Chair of Committee
John H. Golbeck Professor of Biochemistry and Biophysics Professor of Chemistry Co-Chair of Committee
J. Martin Bollinger Jr. Associate Professor of Biochemistry and Molecular Biology
A. Daniel Jones Senior Scientist
B. Tracy Nixon Associate Professor of Biochemistry and Molecular Biology Special Signatory
Robert A. Schlegel Professor of Biochemistry and Molecular Biology Head of the Department of Biochemistry and Molecular Biology
*Signatures are on file in the Graduate School.

iii
ABSTRACT
Roles and functions of isoprenoid quinones (phylloquinone, plastoquinone-9) and
α-tocopherol were investigated in cyanobacteria. Comparative genome analyses of 14
cyanobacteria suggested that phylloquinone (PhyQ) biosynthesis in most but not all
cyanobacteria occurs similarly to menaquinone biosynthesis in Escherichia coli. This was
further supported by the discovery that two cyanobacteria, Synechococcus sp. PCC 7002
and Gloeobacter violaceus PCC 7421, synthesize menaquinone-4 (MQ-4). Targeted
inactivation of the menB, menF, and menG genes resulted in the incorporation of
plastoquinone-9 (PQ-9) and demethyl-MQ or demethyl-PhyQ into Photosystem I (PS I)
complexes. In the PS I complexes containing demethyl-PhyQ, the rate of electron transfer
from A1 to the iron-sulfur clusters slowed by a factor of two, while the kinetics of the
P700+ [FA/FB]- backreaction increased by a factor of 3 to 4. These results were explained
by a lowering of the equilibrium constant between Q-/Q and FX-/FX in the demethyl-PhyQ
containing PS I complexes by a factor of ~10.
Populations of α-tocopherol mutants of the cyanobacterium Synechocystis sp.
PCC 6803, previously isolated in the presence of glucose, were found to be
phenotypically and genotypically heterogeneous. Newly isolated, “authentic” tocopherol
mutants were unable to grow in the presence of glucose at pH 7.0; this was suggested to
be due to a significant reduction of the amounts of sigA and rbcL transcripts in cells
under these conditions. The slr2031 product, which has been previously shown to be
involved in sulfur, nitrogen, and carbon metabolism, and genes encoding inorganic
carbon uptake mechanisms, were found to be constitutively down-regulated in the

iv
“authentic” tocopherol mutants. The results indicate that α-tocopherol is involved in the
transcriptional regulation of these metabolic genes and plays an important role in the
coordination of nitrogen, sulfur, and carbon metabolism in Synechocystis sp. PCC 6803.
The PQ-9 biosynthesis pathway was predicted to be similar to that for ubiquinone
biosynthesis based on comparative genome analyses of 14 cyanobacteria. However,
targeted inactivation mutagenesis of eight genes encoding putative methyltransferase
genes similar to UbiE/MenG in E. coli did not affect PQ-9 biosynthesis in Synechocystis
sp. PCC 6803. Based on the results obtained, a possible PQ-9 biosynthesis pathway is
proposed.

v
TABLE OF CONTENTS
Page List of Figures vii List of Tables xiii Acknowledgements xv Chapter 1 General Introduction 1
Cyanobacterial oxygenic photosynthesis 3 Phylloquinone and Photosystem I 6 Plastoquinone and Photosystem II 9 Role of α-tocopherol 12 Cyanobacterial quinomics 13 References 16
Chapter 2 Comparative genome analysis and identification of menaquinone-4 biosynthetic pathways in cyanobacteria
32
Abstract 33 Introduction 35 Results 38 Discussion 45 Summary 49 Materials and Methods 50 References 54
Chapter 3 Recruitment of a foreign quinone into the A1 site of Photosystem I. Interruption of menG in the phylloquinone biosynthetic pathway of Synechocystis sp. PCC 6803 results in the incorporation of 2-phytyl-1,4-naphthoquinone and alteration of the equilibrium constant for electron transfer between A1 and FX.
72
Abstract 73 Introduction 76 Results 80 Discussion 92 Summary 100 Materials and Methods 101 References 108

vi
Page Chapter 4 Physiological characterization of tocopherol biosynthesis mutants in the cyanobacterium Synechocystis sp. PCC 6803: demonstration of a conditionally lethal phenotype in the presence of glucose at pH 7.0
129
Abstract 130 Introduction 131 Results 137 Discussion 153 Summary 159 Materials and Methods 160 References 167
Chapter 5 Transcriptional regulation of the metabolic and sigma factor genes in the vitamin E-deficient mutant of Synechocystis sp. PCC 6803
194
Abstract 195 Introduction 196 Results 201 Discussion 216 Summary 220 Materials and Methods 221 References 225
Chapter 6 Comparative genome analysis and genetic study of plastoquinone biosynthetic pathway in cyanobacteria
242
Abstract 243 Introduction 245 Results 248 Discussion 258 Summary 262 Materials and Methods 263 References 266
Appendix 291

vii
LIST OF FIGURES
Page
Figure 1.1 Structure of phylloquinone, plastoquinone-9, and α-tocopherol 27
Figure 1.2 Scheme of oxygenic photosynthesis in cyanobacteria 28
Figure 1.3 The spatial arrangement and the redox potentials of the electron
transfer cofactors in Photosystem I
29
Figure 1.4 The electron transfer cofactors in Photosystem II 30
Figure 1.5 Biosynthetic pathway of PQ-9 and α-tocopherol in higher plants 31
Figure 2.1 Menaquinone biosynthetic pathway in E. coli 62
Figure 2.2 The men gene arrangements 63
Figure 2.3 Phylogenetic trees based on the amino acid sequences of MenA,
MenB, and MenD
64
Figure 2.4 HPLC profiles of solvent extracts from PS I complexes and whole
cells
65
Figure 2.5 Absorption spectra of solvent-extracted components analyzed by
HPLC
66
Figure 2.6 LC-mass spectrometric analysis of the solvent extracts from
Synechococcus sp. PCC 7002
67
Figure 2.7 Restriction maps of the menB, menF, and menG coding regions in
Synechococcus sp. PCC 7002
68
Figure 2.8 PCR analyses on genomic DNAs isolated from the menB rubA, menF,
and menG mutants of Synechococcus sp. PCC 7002
69

viii
Figure 2.9 Reverse-phase HPLC analyses of the solvent extracts from the PS I
complexes
70
Figure 2.10 Absorption spectra of eluting components analyzed for PS I
complexes isolated from Synechococcus sp. PCC 7002
71
Figure 3.1 Relationship between the molecular structure of 2-phytyl-1,4-NQ and
the axes of the electronic g-tensor
117
Figure 3.2 Construction and verification of the menG mutant strain of
Synechocystis sp. PCC 6803
118
Figure 3.3 Mass spectra recorded for pigments and quinone extracts from the PS I
complexes isolated from the wild-type and menG mutant strains of Synechocystis
sp. PCC 6803
119
Figure 3.4 77 K fluorescence emission spectra from whole cells of Synechocystis
sp. PCC 6803
120
Figure 3.5 SDS-PAGE analysis of PS I complexes isolated from the wild-type
and menG mutant strains of Synechocystis sp. PCC 6803
121
Figure 3.6 Photoaccumulated Q-band EPR spectra and simulations of PS I
complexes from Synechocystis sp. PCC 6803
122
Figure 3.7 Transient spin-polarized EPR spectra of the charge separated P700+ Q-
state in PS I complexes.
123
Figure 3.8 Simulations of the Q-band spectrum of the menG mutant carried out
for various angles between the largest principal A11 and the gxx axis.
124
Figure 3.9 Spin-polarized EPR transient of wild type and menG mutant 125

ix
Figure 3.10 Decay-associated transient EPR spectra at ambient temperature 126
Figure 3.11 Flash-induced absorbance changes at 812 nm in PS I complexes
isolated from Synechocystis sp. PCC 6803
127
Figure 3.12 Steady-state rates of flavodoxin reduction in PS I complexes isolated
from the wild-type and menG mutant strains.
128
Figure 4.1 Biosynthetic pathway of α-tocopherol in Synechocystis sp. PCC 6803 176
Figure 4.2 Colony morphologies of three tocopherol mutants previously isolated
in the presence of glucose.
177
Figure 4.3 Growth curves of wild type and cell lines derived from small and large
colonies of tocopherol mutants selected in the presence of glucose
178
Figure 4.4 PCR analysis of the genomic DNA extracted from the newly isolated
tocopherol mutants selected in the presence of glucose
179
Figure 4.5 Growth curves of wild type and the “authentic” tocopherol mutants in
the presence and in the absence of glucose
180
Figure 4.6 Growth curves of wild type and the “authentic” slr1736 mutant in the
presence of glucose
181
Figure 4.7 Growth analyses of wild type and the “authentic” slr1736 mutant of
Synechocystis sp. PCC 6803 in the presence of various carbon sources
182
Figure 4.8 Chlorophyll a and quinone contents in wild type and the “authentic”
tocopherol mutants grown in the absence and in the presence of glucose
183
Figure 4.9 Oxygen-evolution activities of the whole-chain and the Photosystem
II-dependent electron transport chains in wild type and the “authentic” tocopherol
184

x
mutants of Synechocystis sp. PCC 6803
Figure 4.10 Immunoblotting analysis for the D1 and PsbO proteins in whole cells
of wild type and three “authentic” tocopherol mutants
185
Figure 4.11 Time-dependent RT-PCR analysis of the sodB and nblA transcripts in
wild type and the “authentic” slr1736 mutant of Synechocystis sp. PCC 6803
186
Figure 4.12 Relative phycobiliprotein contents in wild type and the “authentic”
tocopherol mutants of Synechocystis sp. PCC 6803
187
Figure 4.13 Growth curves of wild type and the “authentic” slr1736 mutant the
presence of glucose at 3% (v/v) CO2 and air
188
Figure 4.14 Cultures of wild type and the “authentic” tocopherol mutants of
Synechocystis sp. PCC 6803 grown at various pH values
189
Figure 4.15 Growth curves of wild type and the “authentic” slr1736 mutant of
Synechocystis sp. PCC 6803 at various pH values in the presence of glucose in air
and 1% (v/v) CO2
190
Figure 4.16 Cultures of wild type and the “authentic” slr1736 mutant of
Synechocystis sp. PCC 6803 grown under various conditions for 48 h at pH 7.0
191
Figure 4.17 PS II-dependent O2-evolution activities of wild type and the
“authentic” slr1736 mutant in the absence and in the presence of glucose
192
Figure 4.18 Phycobiliprotein contents in wild type and the tocopherol mutants of
Synechocystis sp. PCC 6803 grown at 1% (v/v) CO2.
193
Figure 5.1 Time-dependent RT-PCR analysis on the cpcA, nblA, and sbtA
transcripts in wild type and slr1736 mutant grown at pH 7.0 and 8.0
233

xi
Figure 5.2 Time-dependent RT-PCR analysis of the alternative sigma factor
transcripts in wild-type and slr1736 mutant grown at pH 7.0 and 8.0
234
Figure 5.3 Time-dependent RT-PCR analysis of the sigA, sigG, and Ci genes in
wild-type and slr1736 mutant grown at pH 7.0 and 8.0
235
Figure 5.4 Thin-section micrographs of the wild-type cells grown in the absence
and in the presence of glucose for 24 h
236
Figure 5.5 Thin-section micrographs of the slr1736 mutant grown in the absence
and in the presence of glucose for 24 h
237
Figure 5.6 Time-dependent RT-PCR analysis of the slr2031 and CCM gene
transcripts in wild type and slr1736 mutant grown at pH 7.0 and 8.0
238
Figure 5.7 PCR analysis of the genomic DNAs extracted from the slr2031::aadA
and pmgA::aacC1 mutants
239
Figure 5.8 Cultures of wild type, the slr1736, slr2031, and pmgA mutants of
Synechocystis sp. PCC 6803 in the presence of glucose at pH 7.0 and 8.0
240
Figure 5.9 Summary and a model of the non-antioxidant role of α-tocopherol in
Synechocystis sp. PCC 6803
241
Figure 6.1 UQ biosynthesis pathway in Escherichia coli 280
Figure 6.2 Hypothetical PQ-9 biosynthesis pathway in cyanobacteria 281
Figure 6.3 An alignment of the Ycf21/UbiC homologs in cyanobacteria and
eukaryotic phototrophs
282
Figure 6.4 Phylogenetic analyses based on nucleotide sequences of 16S rRNA
and amino acid sequences of Ubi proteins in cyanobacteria and proteobacteria
283

xii
Figure 6.5 Phylogenetic trees based on amino acid sequences of UbiE homologs
in cyanobacteria
285
Figure 6.6 HPLC analysis of the sll1653 mutant of Synechocystis sp. PCC 6803
and the 2-phytyl-1,4-benzoquinone (MPBQ) standard.
286
Figure 6.7 PQ-content in whole cells of the wild-type and methyltransferase
mutant strains of Synechocystis sp. PCC 6803
287
Figure 6.8 Restriction maps of coding regions and their flanking sequences for the
UbiE homologs in Synechocystis sp. PCC 6803
288
Figure 6.9 PCR analyses of genomic DNA isolated from the single, double, and
triple methyltransferase (UbiE homolog) mutants of Synechocystis sp. PCC 6803
289
Figure 6.10 Aromatic amino acid biosynthesis and an alternative proposal for 4-
hydroxybenzoate synthesis in cyanobacteria
290

xiii
LIST OF TABLES
Page
Table 2.1 Whole genome analyses for the MQ/PhyQ biosynthesis enzymes in 14
cyanobacteria, Escherichia coli, Bacillus subtilis, Chlorobium tepidum and
Halobacterium sp. NRC
60
Table 3.1 Doubling times of Synechocystis sp. PCC 6803 wild type and the
menG mutant strains 114
Table 3.2 Chlorophyll content in cells grown photoautotrophically at 32 °C
under various illumination conditions. 115
Table 3.3 Magnetic parameters of PhyQ (A1-) and 2-phytyl-1,4-NQ in the A1
binding site 116
Table 6.1 Amino acid sequence similarities between UbiA homologs in 14
cyanobacteria and E. coli 272
Table 6.2 Amino acid sequence similarities between UbiH homologs in 14
cyanobacteria and E. coli 273
Table 6.3 Amino acid sequence similarities between UbiD homologs in 14
cyanobacteria and E. coli 274
Table 6.4 Amino acid sequence similarities between UbiX homologs in 14
cyanobacteria and E. coli 275
Table 6.5 List of the UbiE homologs in 14 cyanobacteria 276
Table 6.6 Ycf21/UbiC homologs in cyanobacteria and eukaryotic phototrophs 277
Table 6.7 Sequences of primers used for PCR analyses 278

xiv
Table 6.8 Construction of methyltransferase mutants in Synechocystis sp. PCC
6803
279

xv
ACKNOWLEDGEMENTS
I would like to thank my advisor Dr. Donald A. Bryant and my co-advisor Dr.
John H. Golbeck for their guidance and support. I would also like to acknowledge my
collaborators who have contributed to the work on phylloquinone: Dr. Gaozhong Shen,
Dr. Boris Zybailov, Dr. Art van der Est, Dr. Robert Bittl, Dr. Stephan Zech, and Dr.
Dietmar Stehlik; and my collaborators who have contributed to the work on
plastoquinone and α-tocopherol: Dr. Dean DellaPenna, Hiroshi Maeda, and Dr. Zigang
Cheng.
My special thanks are also due to the current and former members of both the
Bryant and Golbeck laboratories. Lastly, I would like to thank my husband, Dr. Niels-
Ulrik Frigaard, for his support and advice throughout my studies.

1
Chapter 1
General Introduction
Cyanobacteria are a phylogenetically coherent group of organisms, yet
morphologically, physiologically, and genetically they are very diverse (Bryant, 1994).
They are found in a variety of terrestrial and aqueous environments such as open oceans,
coastal waters, fresh water lakes, rivers, municipal reservoirs, ponds, and even kitchen
sinks in domestic households. Some perform N2 fixation, and some are capable of
chemotaxis or phototaxis or both. While the majority are obligate photoautotrophs, a few
are able to grow chemoheterotrophically. Genome sizes vary from 2 Mbp to ~15 Mbp.
Despite these differences, there is one thing that is common among all cyanobacteria: the
unique ability to perform oxygenic photosynthesis. All cyanobacteria perform oxygenic
photosynthesis, which uses CO2 as the source of carbon, water as the source electrons,
and light as the source of energy, in order to produce biomass. As a result, molecular
oxygen is produced as a byproduct (Barber and Anderson, 2002). Through the activity of
oxygenic photosynthesis, cyanobacteria have affected, and are continuing to affect, our
environment since their first appearance more than 3 billion years ago. This activity of
cyanobacteria has transformed the atmosphere of our planet from an extremely reducing
to a rather oxidizing environment, and this change has supported the evolution of diverse
forms of life on Earth. In an era of global warming, these organisms attract more
attention given that about half of the global CO2 fixation known to occur on Earth is
carried out by cyanobacteria (Bryant, 2003).

2
Much has been learned about the molecular mechanisms of oxygenic
photosynthesis owing to decades of study, which have dissected and largely solved the
mystery of water oxidation and the detailed functions of the photosynthetic apparatus
primarily from the viewpoint of protein functions. Such a perspective, however, has
somewhat overlooked the biological importance of non-proteineous molecules that are
also involved in the process. A group of such molecules are the quinones. In
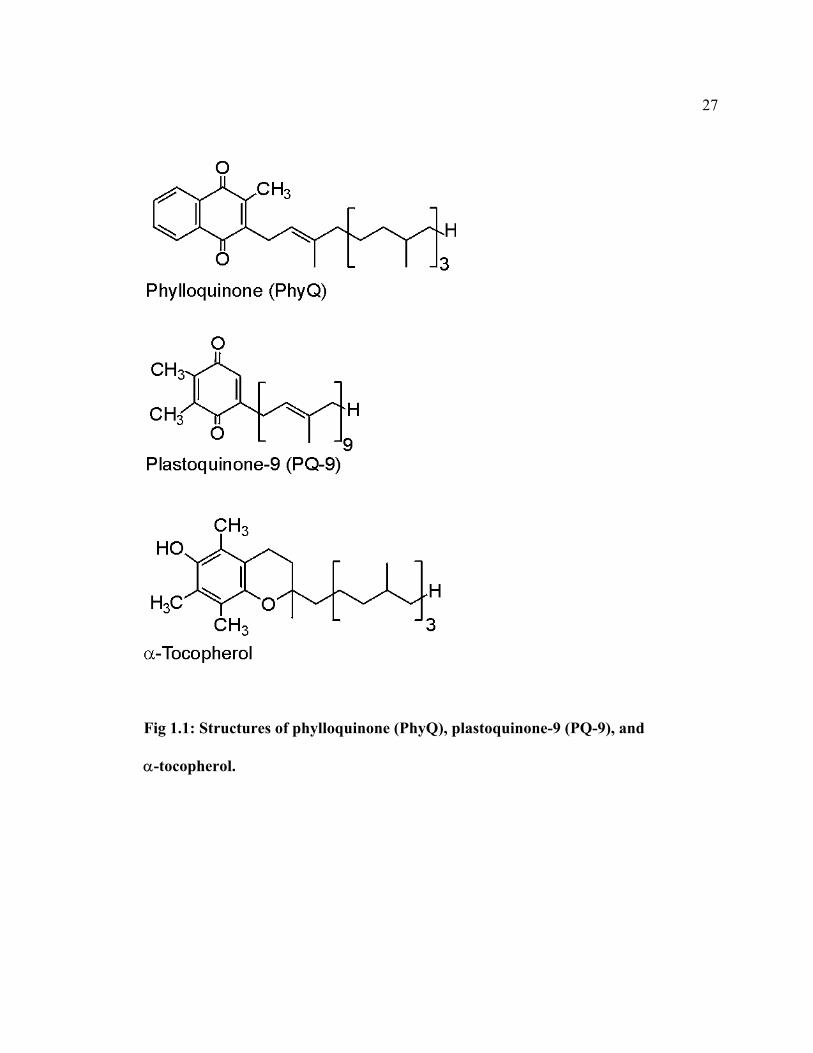
cyanobacteria these are phylloquinone (PhyQ, 2-methyl-3-phytyl-1,4-naphthoquinone,
also known as vitamin K1), plastoquinone (PQ-9, 2,3-dimethyl-5-solanyl-1,4-
benzoquinone). These two compounds are isoprenoid quinones, which are composed of a
quinonoid nucleus with a polyprenyl substituent (Fig 1.1). There are two types of quinone
substructures: 1,4-naphthoquinone (1,4-NQ), and 1,4-benzoquinone (1,4-BQ). PhyQ is a
methyl- and phytyl-substituted 1,4-NQ, while PQ-9 is a methyl- and solanyl-substituted
1,4-BQ. The phytyl substituent is a partially saturated tetra-isoprene unit (equivalent to
20 carbon atoms), while the solanyl substituent is a nona-isoprene unit (equivalent to 45
carbon atoms). α-Tocopherol (2,5,7,8-tetramethyl-6-chromanol, also known vitamin E)
is a tetramethyl-substituted chromanol, another type of quinone nucleus that is derived
from a phytyl-substituted 1,4-BQ after ring closure (Fig 1.1).
These quinones are synthesized only in oxygenic phototrophs including
cyanobacteria, algae and higher plants (Collins and Jones, 1981; Threlfall and Whistance,
1971), and their functions are tightly connected to oxygenic photosynthesis. PhyQ and
PQ-9 are bound cofactors of the Photosystem I (PS I) and Photosystem II (PS II)
complexes, respectively, and in this context they mediate electron transfer as one-electron
carriers (see below). PQ-9 also serves as a membrane-associated two-electron and two-

3
proton carrier, both in the photosynthetic electron transport and the respiratory electron
transport chains. α-Tocopherol, on the other hand, is thought to provide protection
against oxidative stress in animals and plants, although its role(s) in cyanobacteria has not
yet been demonstrated (see below).
Cyanobacterial oxygenic photosynthesis
Fig 1.2 illustrates the process of oxygenic photosynthesis in cyanobacteria. In
general, photosynthesis is described in the context of two processes known as the light
reactions and dark reactions. The light reactions are the processes in which the energy of
photons is transformed into an electrochemical membrane potential that drives ATP
synthesis and into a strong reducing power that ultimately generates NADPH. The dark
reactions are the processes in which CO2 is reduced or “fixed” into biomass at the
expense of NADPH and ATP as the reducing power and energy source, respectively.
The light reactions are catalyzed by the photosynthetic electron transport chain
that resides in the thylakoid membranes (Bryant, 1994; Hervás et al., 2003). This electron
transport chain involves 3 membrane integral protein complexes (PS I, PS II, cytochrome
b6f complex) and 2 water-soluble electron carriers (cytochrome c6 or plastocyanin, and
ferredoxin or flavodoxin), as well as the lipid-soluble electron carrier PQ-9 (Fig 1.2). X-
ray crystallographic structures of PS I, PS II, and cytochrome b6f complexes are now
available at resolutions of 2.5 Å (Jordan et al., 2001), 3.5 Å (Ferreira et al., 2004), and 3.0
Å (Kurisu et al., 2001), respectively. Upon illumination, photons absorbed by the
peripheral antenna complexes known as phycobilisomes are transferred predominantly to
the PS II complexes, in which they trigger photochemistry that drives the light-dependent

4
oxidation of water and the reduction of plastoquinone (Barry et al., 1994; Bricker and
Ghanotakis, 1996; Britt, 1996; Diner and Babcock, 1996). Water oxidation occurs in the
thylakoid lumen and results in the release of molecular oxygen and protons (2H2O O2
+ 4H+ + 4e-), while the reduction and protonation of PQ-9 occurs on the stromal side of
the membrane and releases doubly reduced plastoquinone, or plastoquinol (PQH2), into
the membrane matrix (2 PQ-9 + 4 H+ + 2 e- 2 PQH2). PQH2 shuttles electrons to the
quinol oxidation site (o) of the cytochrome b6 f complex in a diffusion-dependent fashion,
where it is reoxidized and becomes available for a new round of photochemical reduction
by the PS II complexes. At this step, the electron transfer bifurcates into two separate
pathways within the cytochrome b6f complex; one electron is transferred to the Reiske
iron-sulfur protein and subsequently to a soluble electron carrier such as cytochrome c6 or
plastocyanin in the lumen with concomitant release of two protons into the lumen, while
the other electron is transferred to heme bL and heme bH and reduces PQ-9 at the quinone
reduction site (r) near the stromal surface of the thylakoid membranes (Kallas, 1994;
Hauska et al., 1996). After two rounds of PQH2 oxidation, one PQ-9 at the quinone
reduction site becomes doubly reduced and is protonated by concomitant uptake of two
protons from the stroma (cytoplasm). The resulting PQH2 is released into the membrane
matrix where it joins the pool of PQH2 and participates in electron donation to the
cytochrome b6f complex (2PQH2 (O) + PQ (r) + 2H+Lumen 2PQ(O) + PQH2(r) + 4H+
Stroma
+ 2e-). This process is known as the Q cycle, and the result is an amplification of the
proton gradient across the membranes, which is coupled to ATP synthesis by ATP
synthase (Michell, 1976). The final steps of the electron transport chain involve the PS I
complex. Essentially, this light-dependent cytochrome c6/plastocyanin-

5
ferredoxin/flavodoxin oxidoreductase shuttles an electron from cytochrome c6 or
plastocyanin in the lumen to ferredoxin or flavodoxin in the stroma (Golbeck, 1994;
Brettel and Leibl, 2001). The resulting strong reductant donates an electron to ferredoxin-
NADPH+ oxidoreductase, which catalyzes the reduction of NADP+ to NADPH (Morand
et al., 1994).
It is noteworthy that the cyanobacterial photosynthetic electron transport chain
shares the membrane-associated PQ-9 pool and cytochrome b6f complexes with the
respiratory electron transport chain (Fig 1.2). In this case, electrons enter the electron
transport chain through NADH dehydrogenase in the form of NAD(P)H that is generated
through glycolysis and the TCA cycle, and the oxidative pentose phosphate pathway. The
terminal electron acceptor is molecular oxygen, which is reduced and protonated by
cytochrome c oxidase resulting in generation of water (Schmetterer, 1994).
NADPH and ATP generated through the photosynthetic electron transport chain
are used to fuel the dark reactions. A CO2 molecule is first incorporated into the C5
backbone of ribulose-1,5-bisphosphate, which is then split into two molecules of 3-
phosphoglycerate (Tabita, 1987, 1994). This reaction is catalyzed by ribulose-1,5-
bisphosphate carboxylase/oxygenase (RuBisCO), which is probably the most abundant
protein on Earth (Garrett and Grisham, 1999). In cyanobacteria, RuBisCO is housed
within protein-encapsulated structures known as carboxysomes (Badger and Price, 2003),
which play a role in locally increasing the CO2 concentration. 3-Phosphoglycerate is
reduced to 3-phosphoglyceraldehyde, which is processed through the reductive pentose
phosphate pathway to regenerate ribulose-1,5-bisphosphate. The processes are known as
the Calvin cycle and cause the net production of organic molecules that constitute the

6
metabolic precursors for the biosynthesis of proteins, nucleic acids, sugars, pigments, and
other metabolites. The net production of biomass in the Calvin cycle can be expressed as
the following equation:
6 CO2 + 12 NADPH + 18 ATP C6H12O6 + 12 NADP+ + 18 ADP + 18 Pi
PhyQ and Photosystem I
The PS I complex consists of twelve protein subunits, 96 Chl a molecules, 22 β-
carotene molecules, two PhyQ molecules, three iron-sulfur clusters, and some structural
lipids and water molecules (Jordan et al., 2001). Tightly bound electron transfer cofactors
are found within the core of the complex formed by PsaA and PsaB proteins. These
included a chlorophyll a (Chl a) dimer (P700), two Chl a monomers (A0), two PhyQ
molecules (A1), and three [4Fe-4S] clusters (denoted as FX, FA, FB). These cofactors are
arranged in two branches related by C2 symmetry. The spatial arrangement of these
cofactors and their estimated in situ redox potentials are shown in Fig. 1.3. Upon
photoexcitation, the primary donor P700 reduces the primary acceptor, A0, resulting in
the initial charge separated state [P700+ A0-]. Subsequent electron transfer to the
secondary acceptor A1 stabilizes the charge-separated state [P700+ A0 A1-] and minimizes
the rapid charge recombination that occurs between P700+ and A0-. The electron is
subsequently transferred to FX [P700+ A0 A1 FX-], and then to the terminal acceptors FA
[P700+ A0 A1 FX FA-] and FB [P700+ A0 A1 FX FA FB
-], which are located in the peripheral
PsaC subunit. The reduction of ferredoxin or flavodoxin takes place upon direct protein-
protein interaction between these acceptors and the stromal proteins: PsaC, PsaD, and
PsaE (Zhao et al., 1991, 1993; Li et al., 1991; Golbeck, 1994). On the lumenal side, the

7
photooxidized P700+ accepts an electron from reduced cytochrome c6 or plastocyanin
(Durán et al., 2004).
Since the first discovery of PhyQ in oxygenic phototrophs by Dam in 1941 (cited
in Hauska, 1988), the role of PhyQ in PS I as the secondary acceptor A1 was obscure for
more than two decades. After its initial discovery by Thornber and coworkers in the
1970’s, in the 1980s it was found to be exclusively associated with PS I (Thornber et al.,
1976; Takahashi et al., 1985; Shoeder and Locau, 1986) and to participate in electron
transfer as shown by spin-polarized EPR analysis (Petersen et al., 1987) and optical
kinetic analysis (Brettel et al., 1987; see also reviews by Golbeck, 1994, 2003 and
references therein). Further supporting evidence was derived from solvent extraction
studies, in which extraction of PhyQ from PS I with hexane or ether resulted in a rapid,
nano-second-scale charge recombination between P700+ and A0-. Stable charge
separation was restored by the addition of exogenous PhyQ (Biggins and Mathis, 1988;
Itoh and Iwaki, 1989).
This in vitro reconstitution method was subsequently used to study the structural
and thermodynamic requirements for A1 by introducing diverse quinones of abiotic origin
into the A1 site, including benzoquinones, naphthoquinones, and anthraquinones with
various ring substitutions (Iwaki and Itoh, 1989, 1991; Itoh and Iwaki, 1991; see also
review by Itoh 2001). These authors concluded that the successful restoration of stable
charge separation occurs only when quinones with redox potentials between those of A0
and FX are used. They additionally found that neither the structure of the head group nor
the presence of an alkyl substituent is important. This conclusion contradicted earlier
studies, in which the presence of an alkyl substituent was shown to be essential when

8
hexane/hexane-methanol-extracted PS I complexes were studied (Biggins and Mathis,
1998). These contradictory interpretations imply that different solvent-extraction methods
affect the architecture of PS I differently; hence, the inconsistency in conclusions may
reflect artifacts of the methods employed. Such artifacts can be avoided if “biologically”
or “physiologically” modified materials are used. Materials that have not gone through
harsh extraction processes, which result in co-extraction of other important PS I
constituents such as water, Chl a, carotenoid, and lipid molecules (Biggins and Mathis,
1988; Itoh and Iwaki, 1989), should produce more consistent and interpretable results.
Some of these molecules are located in the vicinity of the quinone-binding site; in
particular, as revealed by 2.5Å-resolution crystallographic structure (Jordan et al., 2002),
β-carotenes and chlorophylls form hydrophobic contacts with the phytyl side chain of
PhyQ and seem to provide structural stability for this important electron carrier.
Genetic manipulation of the PhyQ biosynthetic pathway was first introduced by
Chitnis and colleagues (Johnson et al., 2000) and provided an alternative and very
powerful experimental system, by which the selective elimination of PhyQ from PS I
could be achieved in vivo. Based on the menaquinone biosynthetic pathway in
Escherichia coli (Meganathan, 2001), a part of the PhyQ biosynthetic pathway in
Synechocystis sp. PCC 6803 was first predicted. Targeted insertional mutagenesis of
genes encoding dihydroxynaphthoate phytyltransferase (MenA) and
dihydroxynaphthoate-CoA synthase (MenB) successfully interrupted the PhyQ
biosynthesis in the mutant strains (Johnson et al., 2000). The PS I complexes isolated
from these mutants completely lacked PhyQ and accumulated PQ-9 (Johnson et al., 2000;
Zybailov et al., 2000; Semenov et al., 2000). As confirmed by continuous-wave EPR,

9
electron spin-polarized transient EPR, and electron spin-echo modulation experiments
(Zybailov et al., 2000), the orientation of the carbonyl bonds relative to the membrane
normal and the distance between PQ-9 and P700+ were the same as with PhyQ in the
wild-type PS I complexes. In the PS I complexes isolated from the mutants, PQ-9 was
shown to participate in electron transfer from A0 to FX (Zybailov et al., 2000; Semenov et
al., 2000), although the rate of forward electron transfer from A1 to FX slowed at least by
a factor of 100. The estimated in situ redox potential of PQ-9 differs from that of PhyQ
by ca. 130 mV (Shinkarev et al., 2002), which is consistent with the faster rate of charge
recombination between P700+ and FeS- in PS I (Semenov et al., 2000). Despite this
significant defect in the efficiency of forward electron transfer, the PS I complexes
containing PQ-9 supported nearly 85% of the wild-type activity of PS I, as measured by
the rate of cytochrome c6:flavodoxin oxidoreduction (Johnson et al., 2000). More recent
studies have shown that PS I complexes are functional when the A1 site is occupied by
PhyQ, PQ-9 and even anthraquinones with various substituents (Golbeck et al., 2001;
Zybailov, 2003). The combined results indicate that PS I has an innate capacity to
accommodate and use a wide range of redox-active quinones with various structures and
substituents.
PQ-9 and Photosystem II
Cyanobacterial Photosystem II is a light-driven, water:plastoquinone
oxidoreductase and is composed of at least 19 protein subunits, seven carotenoid
molecules, two pheophytin molecules, two PQ-9 molecules, two bicarbonate molecules,
one none-heme Fe, one heme b, and one heme c (Zouni et al., 2001; Ferreira et al.,

10
2004). Four manganese atoms and one calcium atom are bound to the oxygen-evolving
complex that is attached to the lumenal surface of the PS II complex and is responsible
for water oxidation. As shown in Fig 1.4, the electron transfer cofactors are arranged in
two branches that are related by a pseudo-C2 symmetry axis within the core of the PS II
complex that is formed by the D1 and D2 proteins. Aside from the differences in the
specific types of the cofactors and their arrangement, the principle of the photochemistry
is largely the same in PS II as in PS I. Upon photoexcitation of a Chl a monomer (PD1,
the primary donor P680), an electron is transferred to a pheophytin molecule (PheoD1)
(Danielius et al., 1987). This initial charge separation [P680+ Pheo-] is further stabilized
by electron transfer to a tightly bound PQ-9 molecule (QA) [P680+ Pheo QA-], which is
then followed by transfer to a second PQ-9 molecule (QB) [P680+ Pheo QA QB-] (Renger,
1992; Barry et al., 1994). The second electron transfer doubly reduces QB, which, after
protonation, dissociates from the complex into the thylakoid membrane matrix (Bouges-
Bocquet, 1973; Velthuys and Amesz, 1974; Govindjee and van Rensen, 1993). The
oxidized P680+ is reduced by the redox-active tyrosine Z (TyrZ, Y161 in the D1 protein)
(Gerken et al., 1988; Diner and Babcock, 1996), which is then reduced by a tetra-
manganese cluster that catalyzes the abstraction of electrons from water molecules
(Bricker and Ghanotakis, 1996). With the exception of QA, all redox-active cofactors that
are involved in electron transfer are in principle bound by subunit D1 (PsbA) (Ferreira et
al., 2004).
Prolonged exposure to high light intensity is known to cause irreversible damage
to PS II, and this phenomenon is generally known as photoinhibition (see review by Aro
et al., 1993, and references therein). Under such conditions, QA becomes doubly reduced

11
and leaves the binding pocket (Vass et al., 1992). This leads to rapid charge
recombination between P680+ and Pheo- and to formation of the triplet P680T. P680T can
readily react with molecular oxygen in its vicinity and generate singlet oxygen (1O2).
Singlet oxygen, and other reactive oxygen species generated from it, are responsible for
the photodamage. Photoinhibition can also occur under normal light intensity, when the
donation of electrons to P680 occurs slower than their removal by the acceptor side
(Chen et al., 1995; Aro et al, 1993; Anderson and Chow, 2002). In such cases, highly
oxidizing Tyr Z+ and P680+ cation radicals are formed, which can extract electrons from
the surrounding environment and also cause irreversible damage to PS II. In either case,
the D1 protein is rapidly degraded (Gong and Ohad, 1991; Philbrick et al., 1991; Aro et
al., 1993).
Genetic manipulation of the quinone species present in the PS II quinone-binding
sites has not been demonstrated. The PQ-9 biosynthesis pathway has been intensively
studied, and the complete pathway is known in higher plants (Threlfall and Whistance,
1971). The synthesis occurs from the precursor homogentisate after two enzymatic steps
involving homogentisate solanyltransferase and 2-methyl-6-solanyl-1,4-benzoquinone
(MSBQ) methyltransferase (Soll et al., 1980, 1985; Collakova and DellaPenna, 2001;
Cheng et al., 2003; also see review by Threlfall and Whistance, 1971) (Fig. 1.5). This
pathway in plants overlaps with the α-tocopherol biosynthesis pathway, since both utilize
homogentisate as a precursor, and both pathways share the step that introduces the second
methyl group, a reaction which is catalyzed by MPBQ/MSBQ methyltransferase (Cheng
et al. 2003) (Fig 1.5). Recent studies have shown that the cyanobacterial PQ-9
biosynthetic pathway involves neither homogentisate (Dänhardt et al., 2002) nor MPBQ

12
activity (Cheng et al., 2003). Therefore, PQ-9 seems to be synthesized by a completely
different pathway in cyanobacteria.
Role of α-Tocopherol
α-Tocopherol was first discovered by Evans and Bishop (1922) as a factor
essential for reproduction in rats. Forty years later, its antioxidant activity was recognized
by Epstein and colleagues (Epstein et al., 1966). Because α-tocopherol is an essential
component of our diet, much work since then has been done on its significance in animal
systems. Studies in animals, animal cell cultures, and artificial membranes have shown
that tocopherols scavenge or quench various reactive oxygen species and lipid oxidation
by-products that would otherwise propagate lipid peroxidation chain reactions in
membranes (Kamal-Eldin and Appelqvist, 1996). Upon interaction with a prooxidant, α-
tocopherol can undergo one-electron transfer and form a relatively stable tocopheroxyl
cation radical, which is then recycled back to α-tocopherol by an antioxidant network
consisting of ascorbate, glutathione, and NADPH/NADH (see review by Packer et al.,
2001). α-Tocopherol can also undergo two-electron transfer and form α-
tocopherylquinone. In higher plants, this antioxidant function of α-tocopherol has been
discussed in connection with protection against oxidative stress caused by various
environmental factors (Munné-Bosch and Leonor, 2000). In the eukaryotic green algae
Chlamydomonas reinhardtii, the herbicide-mediated interruption of the α-tocopherol
biosynthetic pathway rendered PS II more susceptible to oxidative stress induced by an

13
extreme high light illumination (Trebst et al., 2002). Under these conditions, the D1
protein was rapidly degraded.
In addition to these antioxidant functions, other “non-antioxidant” functions
related to modulation of signaling and transcriptional regulation in mammals have also
been reported (Chan et al., 2001; Azzi et al., 2002; Ricciarelli et al., 2002). For example,
α-tocopherol has been shown to bind to phospholipase A2 specifically at the substrate-
binding pocket and to act as a competitive inhibitor, which thereby decreased the release
of arachidonic acid for eicosanoid synthesis (Chandra et al., 2002). α-Tocopherol has
also been suggested to modulate the phosphorylation state of protein kinase Cα in rat
smooth-muscle cells, possibly via phosphorylation of protein phosphatase 2A (Ricciarelli
et al., 1998). α-Tocopherol is also directly involved in transcriptional regulation in
animals, including the expression of the genes encoding liver collagen αI, α-tocopherol
transfer protein, and α-tropomyosin collagenase (Yamaguchi et al., 2001; Azzi et al.,
2002). Whether α-tocopherol in cyanobacteria performs one or both of these roles has not
yet been demonstrated.
Cyanobacterial Quinomics
The goal of this study was to develop experimental systems in which the type and
amounts of the quinone species found in cyanobacteria could be manipulated in vivo.
Such systems would allow the functions of the complexes in which they occur to be
studied. This has been accomplished by using comparative genome analyses, combined
with a reverse genetic approach, to predict and verify the biosynthetic pathways of PhyQ,

14
α-tocopherol, and PQ-9 (Chapter 2, 3, 6). Consequences of altering the nature of these
quinone species are studied in the context of PS I function (Chapter 3), stress responses
(Chapter 4), and gene regulation (Chapter 5). To express the relatively wide scope of and
inclusiveness of these studies, I introduce the term “quinomics” to describe my
investigations of the cyanobacterial “quinome”.

15
ABBREVIATIONS
BQ benzoquinone
Chl a chlorophyll a
DHNA 1,4-dihydroxy-2-naphthoate
MPBQ 2-methyl-6-phytyl-1,4-benzoquinone
MSBQ 2-methyl-6-solanyl-1,4-benzoquinone
NQ naphthoquinone
PhyQ phylloquinone
PQ-9 plastoquinone-9
PQH2 plastoquinol
PS I Photosystem I
PS II Photosystem II
RuBisCO ribulose-1,5-bisphosphate carboxylase/oxygenase

16
REFERENCES
Andersson JM, Chow WS (2002) Structural and functional dynamics of plant
Photosystem II. Phil Trans R Soc Lond B 357: 1421-1430
Aro E-M, Girgin T, Andersson B (1993) Photoinhibition of Photosystem II. Inactivation,
protein damage and turn over. Biochim Biophys Acta 1143: 113-134
Azzi A, Ricciarelli R, Zingg JM (2002) Non-antioxidant molecular functions of α-
tocopherol (vitamin E). FEBS Lett 519: 8-10
Badger MR, Price GD (2003) CO2 concentrating mechanisms in cyanobacteria:
molecular components, their diversity and evolution. J Exp Bot 54: 609-622
Barber J, Anderson JM (2002) Introduction. Phil Trans R Soc Lond B 357: 1325-1328
Barry BA, Boerner RJ, de Paula JC (1994) The use of cyanobacteria in the study of the
structure and function of Photosystem II. In: Bryant DA (ed) Molecular Biology of
Cyanobacteria. pp 217-257. Kluwer Academic Publishers, Dordrecht
Biggins J and Mathis P (1988) Functional role of vitamin K1 in Photosystem I of the
cyanobacterium Synechocystis 6803. Biochemistry 27: 1494-1500
Bouges-Bocquet B (1973) Electron transfer between the two photosystems in spinach
chloroplasts. Biochim Biophys Acta 314: 250-256

17
Brettel K, Sétif P, Mathis P (1987) Flash-induced absorption changes in Photosystem I at
low temperature: evidence that the electron acceptor A1 is vitamin K1. FEBES Lett 203:
22-224
Brettel K, Leibl W (2001) Electron transfer in Photosystem I. Biochim Biophys Acta
1507: 100-114
Bricker TM, Ghanotakis DF (1996) Introduction to oxygen evolution and the oxygen-
evolving complex. In: Ort DR, Yocum CF (eds), Oxygenic Photosynthesis: The Light
Reactions. pp 113-136. Kluwer Academic Publishers, Dordrecht
Britt RD (1996) Oxygen evolution. In: Ort DR, Yocum CF (eds), Oxygenic
Photosynthesis: The Light Reactions. pp 137-164. Kluwer Academic Publishers,
Dordrecht
Bryant DA (1994) The Molecular Biology of Cyanobacteria, Kluwer Academic
Publisher, Dordrecht
Bryant DA (2003) The beauty in small things revealed. Proc Natl Acad Sci USA 17:
9647-9640
Chan SS, Monteiro HP, Schindler F, Stern A, Junqueira VB. (2001) α-Tocopherol
modulates tyrosine phosphorylation in human neutrophils by inhibition of protein kinase
C activity and activation of tyrosine phosphatases. Free Radic Res 35: 843-856
Chandra V, Jasti J, Kaur P, Betzel C, Srinivasan A, Singh TP (2002) First structural
evidence of a specific inhibition of phospholipase A2 by α-tocopherol (vitamin E) and its

18
implications in inflammation: crystal structure of the complex formed between
phospholipase A2 and α-tocopherol at 1.8 Å resolution. J Mol Biol 320: 215-222
Chen G-X, Blubaugh DJ, Homann PH, Golbeck JH, Cheniae GM (1995) Superoxide
contributes to the rapid inactivation of specific secondary donors of the Photosystem II
reaction center during photodamage of manganese-depleted Photosystem II membranes.
Biochemistry 34: 2317-2332
Cheng Z, Sattler S, Maeda H, Sakuragi Y, Bryant DA, DellaPenna D (2003) Highly
divergent methyltransferase catalyzes a conserved reaction in tocopherol and
plastoquinone synthesis in cyanobacteria and photosynthetic prokaryotes. Plant Cell 15:
2343-2356
Collakova E and DellaPenna D (2001) Isolation and functional analysis of homogentisate
phytyltransferase from Synechocystis sp. PCC 6803 and Arabidopsis. Plant Physiol 127:
1-12
Collins MD and Jones D (1981) Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiological Reviews 45: 316-354
Cramer WA, Knaff DB (1991) Energy Transduction in Biological Membranes. pp 239-
298. Springer-Verlag, New York
Dähnhardt D, Falk J, Appel J, van der Kooij TAW, Schulz-Friedrich R, Krupinska K
(2002) The hydroxyphenylpyruvate dioxygenase from Synechocystis sp. PCC 6803 is not
required for plastoquinone biosynthesis. FEBS Lett 523: 177-181

19
Danielius RV, Satoh K, van Kan PJM, Plijter JJ, Nuijs AN, van Gorkom HI (1987) The
primary reaction of Photosystem II in D1-D2-cytochrome b-559 complex. FEBS Lett
213: 241-244
Diner BA, Babcock GT (1996) Structure, dynamics, and energy conversion efficiency in
Photosystem II. In: Ort DR, Yocum CF (eds), Oxygenic Photosynthesis: The Light
Reactions. pp 213-247. Kluwer Academic Publishers, Dordrecht
Durán RV, Hervás M, de la Rosa MA, Navarro JA (2004) The efficient functioning of
photosynthesis and respiration in Synechocystis sp. PCC 6803 strictly requires the
presence of either cytochrome c6 or plastocyanin. J Biol Chem 279: 7229-7233
Epstein SS, Forsyth J, Saporoschetz IB, Mantel N (1966) An exploratory investigation on
the inhibition of selected photosensitizers by agents of varying antioxidant activity.
Radiat Res 28: 322-335
Evans HM, Bishop BKS (1922) Fetal resorption. Science 55: 650
Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S (2004) Architecture of the
photosynthetic oxygen-evolving center. Science 303: 1831-1838
Garrett RH, Grisham CM (1999) Biochemistry, second edition. Harcourt Brace College
Publishers, Fort Worth
Gerken S, Brettel K, Schlodder E, Witt HT (1988) Optical characterization of the
immediate electron donor to chlorophyll αII+ in O2-evolving Photosystem II complexes.

20
Tyrosine as possible electron carrier between chlorophyll αII and the water-oxidizing
manganese complex. FEBS Lett 237: 69-75
Golbeck JH (2003) The binding of cofactors to Photosystem I analyzed by spectroscopic
and mutagenic methods. Annu Rev Biophys Biomol Struct 32: 237-256
Golbeck JH (1994) Photosystem I in cyanobacteria. In: Bryant DA (ed), The Molecular
Biology of Cyanobacteria, pp 319-360. Kleuwer Academic Publishers, Dordrecht
Golbeck JH, Zybailov B, Shalome E, Shen G (2001) Biological incorporation of
anthraquinone into the A1 site of Photosystem I. Presented in 29th Annual meeting of the
American society of photobiology. Chicago, IL. July 7th-12th
Gong HS, Ohad I (1991) The PQ/PQH2 ratio and occupancy of Photosystem II-QB site by
plastoquinone control the degradation of D1 protein during photoinhibition in vivo. J Biol
Chem 266: 21293-21299
Govindjee, van Rensen JJS (1993) Photosystem II reaction center and bicarbonate. In:
Deisenhofer J, Norris JR (eds), Photosynthetic Reaction Center, Vol I, pp 357-389.
Academic Press, San Diego
Hauska G (1988) Phylloquinone in Photosystem I. Are quinones the secondary-electron
acceptors in all types of photosynthetic reaction centers. Trends Biochem Sci 13: 415-416
Hauska G, Schütz M, Büttner M (1996) The cytochrome b6f complex-composition,
structure and function. In: Ort DR, Yocum CF (eds), Oxygenic Photosynthesis: The Light
Reactions. pp 377-398. Kluwer Academic Publishers, Dordrecht

21
Hervás M, Navarro JA, De La Rosa MA. (2003) Electron transfer between
membrane complexes and soluble proteins in photosynthesis. Acc Chem Res 36:
798-805.
Itoh S and Iwaki M (1989) Vitamin K1 (phylloquinone) restores the turnover of FeS
centers in the ether-extracted spinach PS I particles. FEBS Lett 243: 47-52
Itoh S and Iwaki S (1991) Full replacement of the function of the secondary electron
acceptor phylloquinone (=vitamin K1) by non-quinone carbonyl compounds in green
plant Photosystem I photosynthetic reaction center. Biochemistry 30: 5340-5346
Itoh S, Iwaki M, Ikegami I (2001) Modification of Photosystem I reaction center by the
extraction and exchange of chlorophylls and quinones. Biochim Biophys Acta 1507: 115-
138
Iwaki M and Itoh S (1989) Electron transfer in spinach Photosystem I reaction center
containing benzo-, naphtho- and anthraquinones in place of phylloquinone. FEBS Lett
256: 11-16
Iwaki M and Itoh S (1991) Structure of the phylloquinone-binding (Qф) site in green
plant Photosystem I reaction centers: the affinity of quinones and quinoid compounds for
Qф site. Biochemistry 30: 5347-5352
Johnson TW, Shen G, Zybailov B, Folling D, Reategui R, Beauparlant S, Vassiliev IR,
Bryant DA, Jones AD, Golbeck JH, Chitnis PR (2000) Recruitment of a foreign quinone
into the A1 site of Photosystem I: I. Genetic and physiological characterization of

22
phylloquinone biosynthetic pathway mutants in Synechocystis sp. PCC 6803. J Biol
Chem 275: 8523-8530
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauß N (2001) Three-dimensional
structure of cyanobacterial Photosystem I at 2.5 Å resolution. Nature 411: 896-899
Kallas T (1994) The Cytochrome b6f complex. In: Bryant DA (ed), Molecular Biology of
Cyanobacteria. pp 259-317. Kluwer Academic Publishers, Dordrecht
Kamal-Eldin A, Appelqvist L-A (1996) The chemistry and antioxidant properties of
tocopherols and tocotrienols. Lipids 31: 671-701
Kurisu G, Zhang H, Smith JL, Cramer WA (2003) Structure of the cytochrome b6f
complex of oxygenic photosynthesis: tuning the cavity. Science 302: 1009-1014
Li N, Zhao JD, Warren PV, Warden JT, Bryant DA, Golbeck JH (1991) PsaD is required
for the stable binding of PsaC to the Photosystem I core protein of Synechococcus sp.
PCC 6301. Biochemistry 30: 7863-7872
Meganathan R (2001) Biosynthesis of menaquinone (vitamin K2) and ubiquinone
(coenzyme Q): a perspective on enzymatic mechanisms. Vitam Horm 61: 173-218
Michell P (1976) Possible molecular mechanisms of the protonmotive function of
cytochrome systems. J Theo Biol 62: 327-367
Morand LZ, Cheng RH, Krogmann DW, Ho KK (1994) Soluble electron transfer
catalysts of cyanobacteria. In: Bryant DA (ed), Molecular Biology of Cyanobacteria. pp
381-407. Kluwer Academic Publishers, Dordrecht

23
Munné-Bosch S, Leonor A (2000) The function of tocopherols and tocotrienols in plants.
Critic Rev Plant Sci 21: 31-57
Packer L, Webber SU, Rimbach G (2001) Molecular aspects of α-tocotrienol antioxidant
action and cell signaling. J Nutr 131: 369S-373S
Petersen, J, Stehlik D, Gast P, Thurnauer M (1987) Comparison of the electron spin
polarized spectrum founding plant Photosystem I and in iron-depleted bacterial reaction
centers with time-resolved K-band EPR; evidence that the Photosystem I acceptor A1 is a
quinone. Photosynth Res 14: 15-29
Philbrick JB, Diner BA, Zilinskas BA (1991) Construction and characterization of
cyanobacterial mutants lacking the manganese-stabilizing polypeptide of Photosystem II.
J Biol Chem 266: 13370-13376
Renger G (1992) Energy transfer and trapping in Photosystem II. In: Barber J (ed), The
Photosystems: Structure, Function and Molecular Biology, Topics in Photosynthesis,
Vol. 11, pp 45-99. Elsevier Science Publishers, Amsterdam
Ricciarelli R, Tasinato A, Clément S, Özer NK, Boscoboinik D, Azzi A (1998) α-
Tocopherol specifically inactivates cellular protein kinase Cα by changing its
phosphorylation state. Biochem J 334: 243-249
Ricciarelli R, Zingg J-M, Azzi A (2002) The 80th anniversary of vitamin E: beyond its
antioxidant properties. Biol Chem 383: 457-465

24
Schmetterer G (1994) Cyanobacterial Respiration. In: Bryant DA (ed), Molecular
Biology of Cyanobacteria. pp 409-435. Kluwer Academic Publishers, Dordrecht.
Semenov AY, Vassiliev IR, van der Est A, Mamedov MD, Zybailov B, Shen G, Stehlik
D, Diner BA, Chitnis PR, Golbeck JH (2000) Recruitment of a foreign quinone into the
A1 site of Photosystem I: Altered kinetics of electron transfer in phylloquinone
biosynthetic pathway mutants studied by time-resolved optical, EPR, and electromagnetic
techniques. J Biol Chem 275: 23429-23438
Shinkarev VP, Zybailov B, Vassiliev IR, Golbeck JH (2002) Modeling of the P700+
charge recombination kinetics with phylloquinone and plastoquinone in the A1 site of
Photosystem I. Biophys J 83: 2885-2897
Shoeder HU and Lockau W (1986) Phylloquinone copurified with the large subunits of
Photosystem I. FEBS Lett 199: 23-27
Soll J, Kemmerling M, Schultz G. (1980) Tocopherol and plastoquinone synthesis in
spinach chloroplasts subfractions. Arch Biochem Biophys 204: 544-550
Soll J, Schultz G, Joyard J, Douce R, Block MA (1985) Localization and synthesis of
prenylquinones in isolated outer and inner envelope membranes from spinach
chloroplast. Arch Biochem Biophys 238: 290-299
Tabita FR (1987) Carbon dioxide fixation and its regulation in cyanobacteria. In: Fay P,
Baalen CV (eds), The Cyanobacteria. pp 95-117 Elesvier Science Publishers, Amsterdam

25
Tabita FR (1994) The biochemistry and molecular regulation of carbon dioxide
metabolism in cyanobacteria. In: Bryant DA (ed), Molecular Biology of Cyanobacteria.
pp 437-467. Kluwer Academic Publishers, Dordrecht
Takahashi Y, Hirota K, Katoh S (1985) Multiple forms of P700-chlorophyll a-protein
complex from Synechococcus sp.: the iron, quinone and carotenoid contents. Photosynth
Res 6: 183-192
Threlfall DR and Whistance GR (1971) Biosynthesis of isoprenoid quinones and
chromanols. In: Goodwin TW (ed), Aspects of Terpenoid Chemistry and Biochemistry,
12: 357-404. Academic Press, New York
Trebst A, Depka Bm Hollander-Czytko H (2002) A specific role for tocopherol and of
chemical singlet oxygen quenchers in the maintenance of Photosystem II structure and
function in Chlamydomonas reinhardtii. FEBS Lett 516: 156-160
Vass I, Styring S, Hundal T, Koivuniemi A, Aro E-M, and Andersson B (1992)
Reversible and irreversible intermediates during photoinhibition of Photosystem II: stable
reduced QA species promote chlorophyll triplet formation. Proc Natl Acad Sci 89: 1408-
1412
Velthuys BR, Amesz J (1974) Charge accumulation at the reducing side of system 2 of
photosynthesis. Biochim Biophys Acta 333: 85-94

26
Zhao JD, Warren PV, Li N, Bryant DA, Golbeck JH (1990) Reconstitution of electron
transport in Photosystem I with PsaC and PsaD proteins expressed in Escherichia coli.
FEBS Lett 276: 175-180
Zhao J, Snyder WB, Muhlenhoff U, Rhiel E, Warren PV, Golbeck JH, Bryant DA (1993)
Cloning and characterization of the psaE gene of the cyanobacterium Synechococcus sp.
PCC 7002: characterization of a psaE mutant and overproduction of the protein in
Escherichia coli. Mol Microbiol 9: 183-194.
Zouni A, Witt H-T, Kern J, Fromme P, Krauß N, Saenger W, Orth P (2001) Crystal
structure of Photosystem II from Synechococcus elongatus at 3.8 A resolution. Nature
409: 739-743
Zybailov B (2003) Modified Quinone Acceptor in Photosystem I. The Pennsylvania State
University, Ph.D. Thesis
Zybailov B, van der Est A, Zech SG, Teutloff C, Johnson TW, Shen G, Bittle R, Stehlik
D, Chitnis PR, Golbeck JH (2000) Recruitment of a foreign quinone into the A1 site of
Photosystem I: II. Structural and functional characterization of phylloquinone
biosynthetic pathway mutants by electron paramagnetic resonance and electron-nuclear
double resonance spectroscopy. J Biol Chem 275: 8531-8539
Yamaguchi J, Iwamoto T, Kida S, Masushige S, Yamada K, Esashi T (2001) Tocopherol-
associated proteins is a ligand-dependent transcriptional activator. Biochem Biophys Res
Commun 285: 295-299
27
Fig 1.1: Structures of phylloquinone (PhyQ), plastoquinone-9 (PQ-9), and
α-tocopherol.
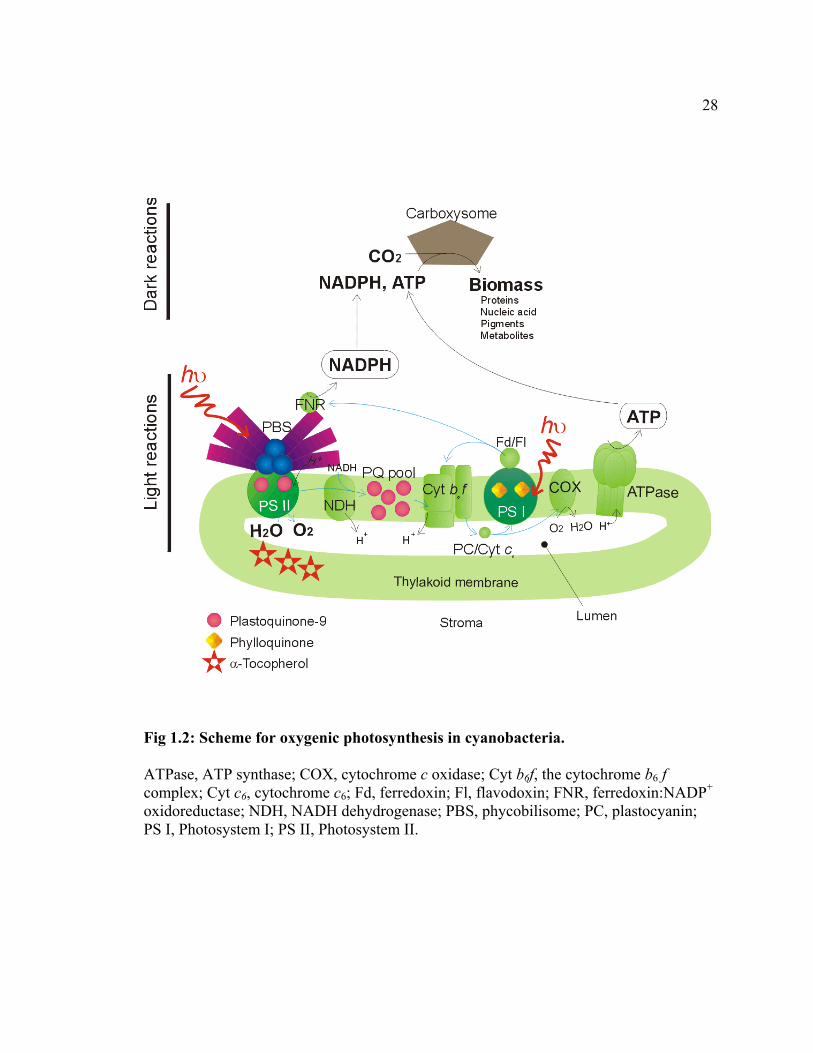
28
Fig 1.2: Scheme for oxygenic photosynthesis in cyanobacteria.
ATPase, ATP synthase; COX, cytochrome c oxidase; Cyt b6f, the cytochrome b6 f complex; Cyt c6, cytochrome c6; Fd, ferredoxin; Fl, flavodoxin; FNR, ferredoxin:NADP+ oxidoreductase; NDH, NADH dehydrogenase; PBS, phycobilisome; PC, plastocyanin; PS I, Photosystem I; PS II, Photosystem II.
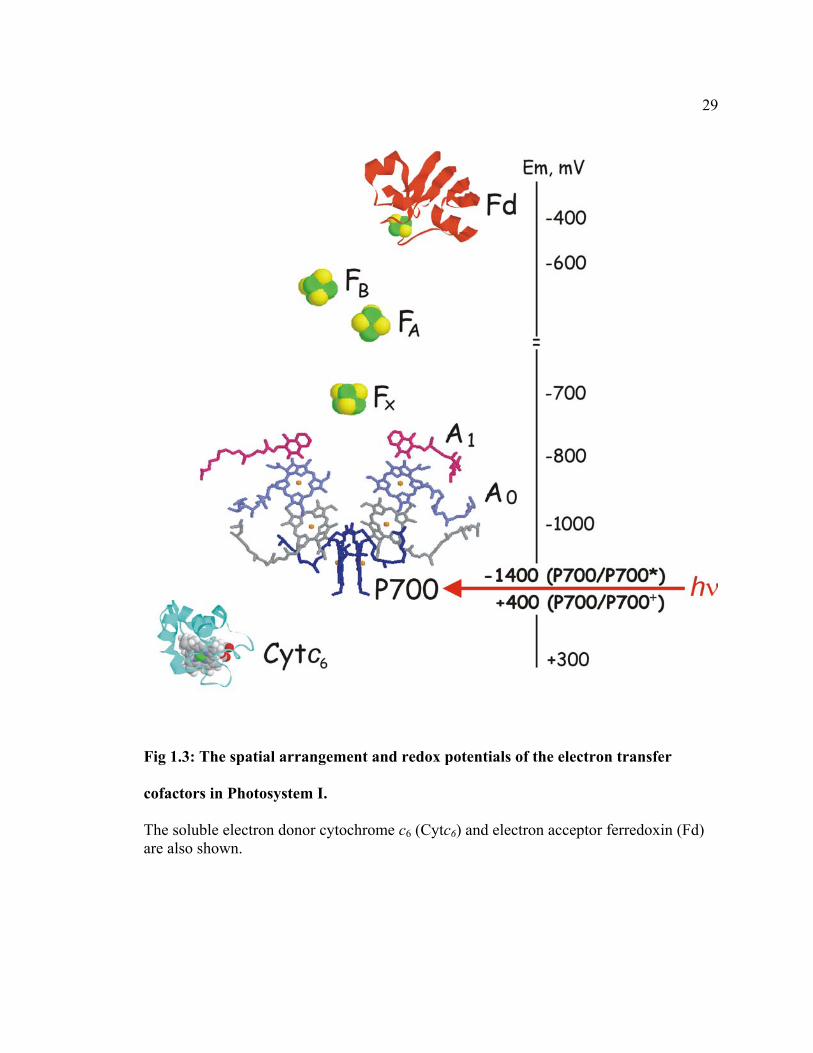
29
Fig 1.3: The spatial arrangement and redox potentials of the electron transfer
cofactors in Photosystem I.
The soluble electron donor cytochrome c6 (Cytc6) and electron acceptor ferredoxin (Fd) are also shown.
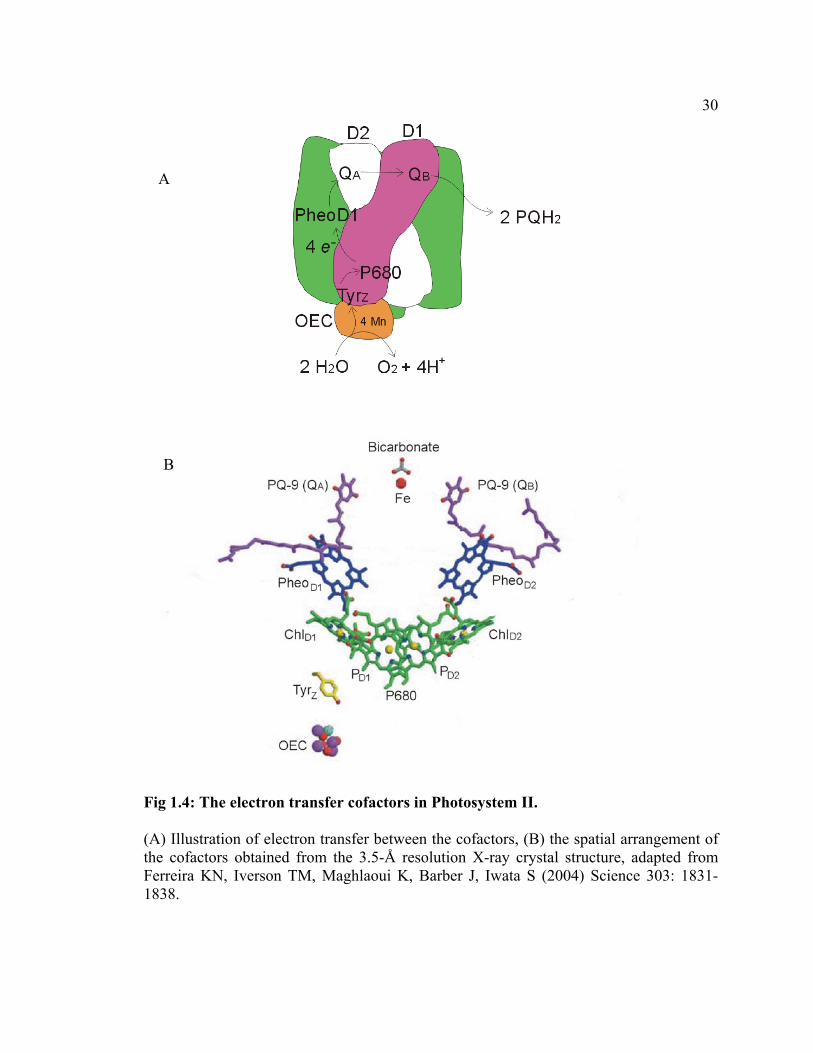
30
Fig 1.4: The electron transfer cofactors in Photosystem II.
(A) Illustration of electron transfer between the cofactors, (B) the spatial arrangement of the cofactors obtained from the 3.5-Å resolution X-ray crystal structure, adapted from Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S (2004) Science 303: 1831-1838.
A
B
31
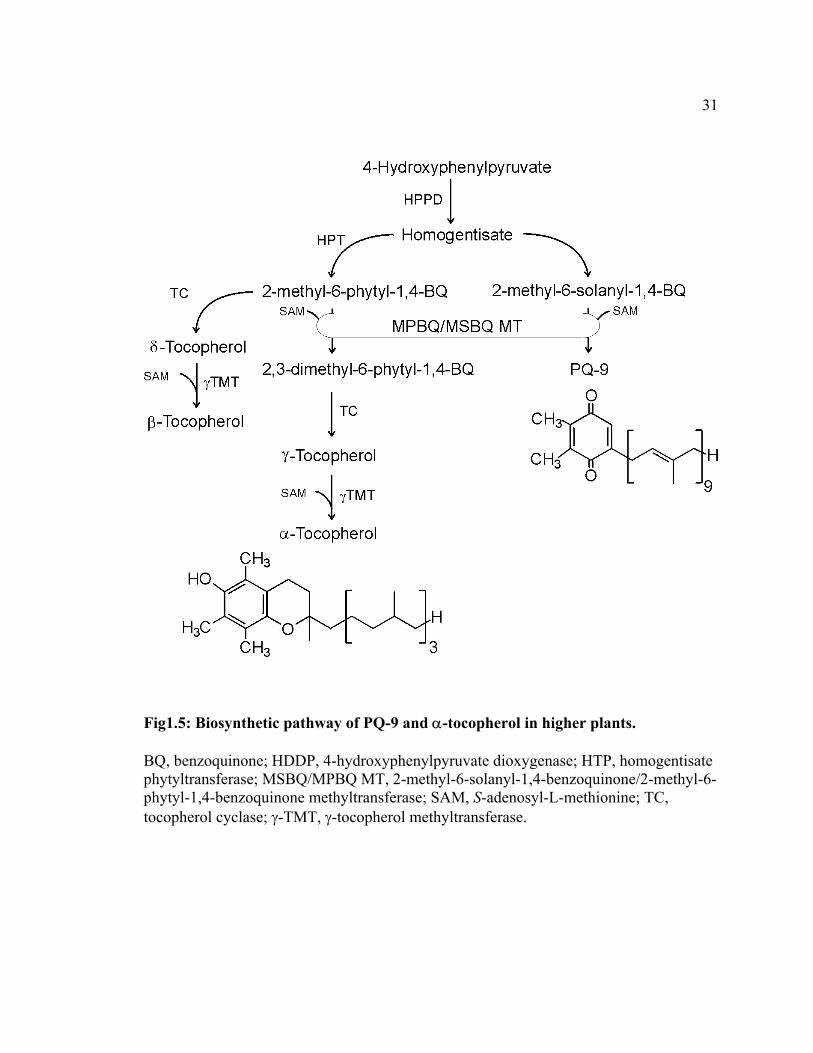
Fig1.5: Biosynthetic pathway of PQ-9 and α-tocopherol in higher plants.
BQ, benzoquinone; HDDP, 4-hydroxyphenylpyruvate dioxygenase; HTP, homogentisate phytyltransferase; MSBQ/MPBQ MT, 2-methyl-6-solanyl-1,4-benzoquinone/2-methyl-6-phytyl-1,4-benzoquinone methyltransferase; SAM, S-adenosyl-L-methionine; TC, tocopherol cyclase; γ-TMT, γ-tocopherol methyltransferase.

32
Chapter 2
Comparative Genome Analysis and Identification of Menaquinone-4 Biosynthetic
Pathways in Cyanobacteria
Publications:
Yumiko Sakuragi, Boris Zybailov, Gaozhong Shen, Ramakrishnan Balasubramanian,
Bruce A. Diner, Irina Karygina, Yulia Pushkar, Dietmar Stehlik, Donald A. Bryant and
John H. Golbeck, Recruitment of a foreign quinone into the A1 site of Photosystem I.
Spectroscopic characterization of a menB rubA double deletion mutant in Synechococcus
sp. PCC 7002 containing plastoquinone-9 but devoid of FX, FB, FA, in preparation
Yumiko Sakuragi and Donald A. Bryant, a review article ‘Genetic Manipulation of the
Quinone Pathway in Photosystem I’ in Photosystem I: the Plastocyanin:Ferredoxin
Oxidoreductase in Photosynthesis’, Golbeck JH (ed), in preparation

33
ABSTRACT
Phylloquinone (PhyQ, 2-methyl-3-phytyl-1,4-naphthoquinone) is an electron transfer
cofactor at the A1 site of Photosystem I and is synthesized only in oxygenic phototrophs
such as cyanobacteria, algae, and higher plants. The biosynthetic pathway of PhyQ was
investigated by means of comparative genomics using the menaquinone (MQ)
biosynthesis genes of Escherichia coli as queries. The homologs of menA, menB, menC,
menD, menE, menF, menG and menH were found to be conserved in most of the
cyanobacteria. In some of these organisms, these genes formed clusters similar to those
found in Escherichia coli, Bacillus subtilis, and Chlorobium tepidum, suggesting that the
PhyQ biosynthetic pathway has evolved from that for MQ. In support of this hypothesis it
was demonstrated that Gloeobacter violaceus and Synechococcus sp. PCC 7002 wild-
type strains synthesize MQ-4 (2-methyl-3-geranylgeranyl-1,4-naphthoquinone) that
functions at the A1 site in Photosystem I complexes. Targeted insertional mutagenesis of
menB, menF, and menG in Synechococcus sp. PCC 7002 results in the complete
interruption of MQ-4 biosynthesis and demonstrates that the predicted pathway indeed
functions in this cyanobacterium. Phylogenetic analysis of MenA (dihydroxynaphthoate
phytyltransferase) showed that the MenA sequences of the plant Arabidopsis thaliana
and Oryza sativa group with cyanobacterial sequences, whereas MenB
(dihydroxynaphthoate-CoA synthase) and MenD (2-succinyl-6-hydroxy-2,4-
cyclohexadiene-1-carboxylate synthase) do not. This leads to the hypothesis that PhyQ
biosynthesis is a result of mosaic evolution derived from a non-cyanobacterial MQ

34
pathway together with a cyanobacterial-type MenA, possibly derived from
endosymbiosis.
ABBREVIATIONS
Chl a chlorophyll a
DHNA dihydroxynaphthoate
HPLC high-performance liquid chromatography
MQ-4 menaquinone-4
PCR polymerase chain reaction
PhyQ phylloquinone
PQ plastoquinone-9
PS I Photosystem I

35
INTRODUCTION
Phylloquinone (PhyQ, 2-methyl-3-phytyl-1,4-naphthoquinone), also known as
vitamin K1, is synthesized only in oxygenic phototrophs such as cyanobacteria, algae, and
higher plants. PhyQ plays a crucial role in photosynthesis as a cofactor in Photosystem I
by mediating electron transfer between the primary acceptor A0 (Chl a) and the terminal
acceptor Fx (4Fe-4S cluster) after photoexcitation of the primary donor P700 (Golbeck,
1994, 2003; Brettel and Leibl, 2001). Two molecules of PhyQ are found within the
protein environments of the PS I complex, which is composed of 12 protein subunits, 96
Chl a molecules, 22 β-carotenes, 3 [4Fe-4S] clusters, and 4-5 structural lipids (Jordan et
al., 2001). PS I is a light-driven cytochrome c6/plastocyanin:ferredoxin/flavodoxin
oxidoreductase and is responsible for providing electrons that are ultimately used for the
generation of NADPH, the reducing power for cellular anabolism. The quantum
efficiency of this enzyme is largely dependent on the stability of a charge-separated state
between P700+ and FX-, and the presence of PhyQ ensures an optimal rate of transfer
between these cofactors.
Biosynthesis of PhyQ has been studied by isotope tracer and direct enzymatic
activity assays in higher plants (see a review by Pennock and Threlfall, 1981). The results
have indicated that PhyQ biosynthesis resembles menaquinone (MQ) biosynthesis in E.
coli (see reviews by Meganathan 1996, 2001). MQ biosynthesis occurs through eight
enzymatic steps as summarized in Fig 2.1. The first committed step is the isomerization
of chorismate to isochorismate by isochorismate synthase (MenF), followed by the

36
condensation of isochorismate and thiamine-pyrophosphate-succinic semialdehyde by 2-
succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate (SHCHC) synthase (MenD).
Dehydration of SHCHC is catalyzed by o-succinylbenzoate (OSB) synthase (MenC) and
the subsequent CoA thioesterification by OSB-CoA synthetase (MenE); the ring
cyclization reaction is catalyzed by 1,4-hydroxynaphthoyl-CoA synthase (MenB), and
deesterification by 1,4-hydroxynaphthoyl-CoA thioesterase (MenH) results in 1,4-
dihydroxy-2-naphthoate (DHNA). DHNA polyprenyltransferase (MenA) catalyzes the
condensation of DHNA and polyprenyl diphosphate, yielding 2-polyprenyl-1,4-
naphthoquinone (demethylmenaquinone). The final methylation at the C2 position is
carried out by 2-polyprenyl-1,4-naphthoquinone methyltransferase (MenG) with S-
adenosyl-methinone as a methyl donor, resulting in the production of MQ. PhyQ and MQ
have the same general molecular structure except that PhyQ has a partially saturated, C20
phytyl substituent and MQ found in other bacteria has an unsaturated polyprenyl
substituent typically consisting of 30 to 50 carbon units (Collins and Jones, 1981).
The targeted inactivation of the menA, menB, menD, and menE homologs in
Synechocystis sp. PCC 6803 was shown to result in the complete interruption of PhyQ
biosynthesis. This demonstrated that a MQ-like pathway is responsible for PhyQ
biosynthesis in this organism (Johnson et al., 2000; Johnson et al., 2002). The following
questions remained to be answered: is this MQ-like PhyQ biosynthesis pathway
ubiquitous among cyanobacteria and higher plants; what is its evolutionary origin; how
has it evolved among the various oxygenic phototrophs? In this study, MQ biosynthesis
was investigated in 14 cyanobacteria whose genomes have been or are currently being

37
sequenced. The results indicate that the MQ-like pathway is conserved among most,
although not all, of the cyanobacteria and higher plants. A detailed analysis of the gene
arrangements and phylogenetic reconstruction lead to the hypothesis that the PhyQ
biosynthetic pathway has evolved from the MQ pathway in both cyanobacteria and
higher plants.

38
RESULTS
As of April 26, 2004, complete genomic sequences of 8 cyanobacteria are
available: Synechocystis sp. PCC 6803 (Kaneko et al., 1996), Nostoc PCC 7120 (Kaneko
et al., 2001), Thermosynechococcus elongatus BP-1 (Nakamura et al., 2002ab),
Gloeobacter violaceus (Nakamura Y et al., 2003), Prochlorococcus marinus MED4 and
Prochlorococcus marinus MIT9313 (Rocap et al., 2003), Prochlorococcus marinus
SS120 (Dufresne et al., 2003), Synechococcus sp. WH8102 (Palenik et al., 2003); and
incomplete sequences exist for Synechococcus sp. PCC 7002 (Jürgen Marquardt, Tao Li,
Jindong Zhao, and Donald A. Bryant, unpublished), Nostoc punctiforme (Gene Bank
accession number: NZ_AAAY00000000), Trychodesmium erythraeum (Gene Bank
accession number: NZ_AABK00000000), Synechococcus elongatus sp. PCC 7942 (Gene
Bank accession number: NZ_AADZ01000001), Anabaena variabilis ATCC 29413 (Gene
Bank accession number: NZ_AAEA01000001), and Crocosphaera watsonii WH 8501
(Gene Bank accession number: NZ_AADV01000004). Similarity searches with the
MenA through MenH protein sequences of E. coli as queries were performed against
each genome, and the results are summarized in Table 2.1. Of 14 cyanobacteria
examined, 11 were shown to possess the complete set of 8 Men protein homologs.
Similarities based on amino acid sequences were typically in the range of 30-70%,
although the MenB sequences were very highly conserved among a wide range of
organisms including cyanobacteria, the γ-proteobacterium E. coli, the green sulfur
bacterium Chlorobium tepidum, the Gram-positive bacterium Bacillus subtilis, and the
archeon Halobacterium sp. NRC-1 (similarity >70%). In the genome of G. violaceus,

39
homologs of only MenA and MenG were detected. When the Synechocystis sp. PCC
6803 MenB sequence was used to search the G. violaceus genome database, the best hit
was Gll2549 with only 38 % similarity. Further database searching has revealed that
Gll2549 is highly similar to 6-oxo camphor hydrolase in Rhodococcus sp. NCIMB 9784
(69%). When Synechocystis sp. PCC 6803 MenC, MenD, MenE, and MenF sequences
were used to search the G. violaceus database, the best hits were Gll3099 (35%), Gll2804
(44%), Glr1146 (45%), and Gll0757 (43%), respectively. These ORFs also showed
higher similarities to other hypothetical proteins: Gll3099 is similar to chloromuconate
cycloisomerase in Clostridium acetobutylicum (57%); Gll2804 is similar to acetolactate
synthase in Synechocystis sp. PCC 6803 (Sll1981, 86%); Glr1146 is similar to a long-
chain fatty-acid CoA ligase in Bacillus cereus (54%); and Gll0757 is similar to
anthranilate synthase component I in Synechocystis sp. PCC 6803 (Slr1979, 66%). No
homologs of MenD (A. variabilis) or MenC and MenF (C. watsonii) have been identified
to date, but these genomes are not yet completely sequenced.
Some of the men genes involved in PhyQ biosynthesis are found to form clusters
in several cyanobacterial genomes. A cluster composed of menF, menA, menC, and menE
is conserved in T. erythraeum, N. punctiforme, Nostoc sp. PCC 7120, Synechococcus sp.
WH 8102, and all three Prochlorococcus species (Fig 2.2). In a separate region of the
genome, menD and menB are also found to form a cluster. The observed gene
arrangements in these cyanobacteria are similar to parts of the gene clusters found in E.
coli, C. tepidum, B. subtilis and Halobacterium sp. NRC-1 (Fig 2.2). Combined with the
conservation in the amino acid sequences among Men proteins, the conserved

40
arrangements of the men genes are strong indications that PhyQ biosynthesis in
cyanobacteria evolved from the MQ pathway of other bacteria.
Given that oxygenic photosynthesis is believed to have originated in
cyanobacteria and that PhyQ is associated exclusively with PS I, one may expect that the
common ancestor of higher plants and algae acquired the entire PhyQ biosynthetic
pathway from cyanobacteria upon endosymbiosis. Interestingly, the men gene
arrangement in Arabidopsis thaliana does not resemble that in cyanobacteria (Fig 2.2).
Lack of conserved gene arrangements does not necessarily indicate the absence of an
evolutionary relationship; however, it is sufficient to cast doubt about it. Thus the
evolutionary relatedness between cyanobacterial and higher plant PhyQ biosynthetic
genes was evaluated further. As shown in Table 2.1, A. thaliana contains the whole set of
men genes in its nuclear chromosomes, which indicates that PhyQ biosynthesis in A.
thaliana is similar to that in cyanobacteria and to MQ synthesis in other prokaryotes. In a
phylogenetic tree based on the MenA sequences, the two higher plants, A. thaliana and
Oryza sativa, form a clade within a domain composed entirely of cyanobacteria, which
suggests that MenA in higher plants is evolutionarily more related to MenA in
cyanobacteria than to MenA of other eubacteria (Fig 2.3A). On the contrary, in trees
based on the MenB and MenD sequences, these two higher plants form a clade outside
the cyanobacterial domain. This suggests that MenB and MenD in higher plants are more
closely related to their homologs in other eubacteria than to those in cyanobacteria (Fig
2.3B and C). Trees based on the MenC, MenE, and MenF sequences showed the topology
similar to those observed for the MenB and MenD sequences; however, the detailed
branching orders within the cyanobacterial domain were variable probably due to low
sequence conservation of these proteins (< ~50%). The observed variation in the

41
phylogenetic relationships therefore suggests that not all men genes in higher plants
originated from cyanobacteria.
By comparative genome analyses and phylogenetic reconstructions, it was so far
hypothesized that PhyQ biosynthesis in cyanobacteria and higher plants evolved from
that of MQ, which suggests that early cyanobacteria synthesized and utilized MQ for the
PS I complexes. This hypothesis was supported by the discovery that G. violaceus and
Synechococcus sp. PCC 7002 synthesize MQ-4 instead of PhyQ. Fig 2.4A shows the
HPLC profile of the solvent extract obtained from the PS I complexes isolated from
Synechocystis sp. PCC 6803 wild type. PhyQ, which typically elutes at 22.4 min, was
shown to be present. In the HPLC profiles of the solvent extracts obtained from the PS I
complexes isolated from Synechococcus sp. PCC 7002 wild type and from the G.
violaceus whole cells, no such peak was present as judged by the absence of UV-
absorbing compounds at this retention time (Fig 2.4B and C). Further examination of the
chromatograms led to the discovery of a new peak at ca. 14 min that had an intense UV-
absorption with maxima at 248, 263, 270, and 332 nm (Fig 2.5B and C), which coincide
well with the absorption spectrum of phylloquinone obtained from Synechocystis sp. PCC
6803 (Fig 2.5A). This spectrum is characteristic of a 1,4-naphthoquinoid compound with
two alkyl substitutions at the C2 and C3 positions (Dunphy and Brodie, 1971). The
absence of PhyQ in G. violaceus was further confirmed by the absence of the PhyQ
absorption in the eluate at 22.5 min (Fig 2.5C). The same results were obtained for
Synechococcus sp. PCC 7002. Mass spectrometric analysis of the solvent extracts of PS I
complex isolated from Synechococcus sp. PCC 7002 showed that the compound eluting
at ca. 14 min has a m/z of 444, as opposed to a m/z of 450 for PhyQ (Figure 2.6). This
difference is best explained by the presence of a geranylgeranyl substituent with four
unsaturated isoprenoid units. Thus, Synechococcus sp. PCC 7002 synthesizes MQ-4

42
instead of PhyQ. The absence of PhyQ was confirmed by the absence of any component
with a m/z of 450 at 22.5 min (Fig 2.6). The solvent extracts from the PS I complexes
isolated from Synechococcus sp. PCC 7002 revealed the presence of 2.3 MQ-4 molecules
per 100 Chl a molecules, demonstrating that MQ-4 plays a role as the A1 cofactor in the
PS I complexes in this cyanobacterium. Similar quantitative analysis for G. violaceus has
not yet performed.
Targeted insertional inactivation of menB, menF, and menG in Synechococcus sp.
PCC 7002 was performed to verify the involvement of these gene products in the MQ-4
biosynthesis (Fig 2.7). The menF and menG mutations were introduced into the wild
type, whereas the menB mutation was introduced into the rubA mutant for the purpose of
generating PS I complexes that contain plastoquinone-9 but lack iron-sulfur clusters for
future studies concerning electron transfer kinetics and thermodynamics (Y. Sakuragi, B.
Zybailov, G. Shen, R. Balasubramanian, B. A. Diner, I. Karygina, Y. Pushkar, D. Stehlik,
D. A. Bryant and J. H. Golbeck, manuscript in preparation). The full segregation of the
mutated alleles from the respective wild-type alleles was analyzed by PCR as shown in
Fig 2.8. In the rubA mutant, PCR using the designed primers that are targeted to the
menB region resulted in a product of 1.0 kb, which is expected based on the restriction
map, as shown in Fig 2.7. Likewise, in the wild type, PCR using the designed primers
that are targeted to the menF and menG regions resulted in products of 1.7 kb, and 1.8 kb,
respectively. No products with corresponding sizes were detected for the mutants; instead
products with larger sizes were detected. In the menB rubA and the menF mutants, the
size of the products were ca. 2.1 kb and 2.8 kb, respectively. The difference between the
PCR products from the wild type and the mutants corresponds to the size of the 1.1-kb
gentamicin-resistance cartridge derived from pMS266. In the menG mutant a product of
2.2 kb was observed. The difference between the PCR products from the wild type and

43
the menG mutant is 0.3 kb, which corresponds to the difference between the 1.1-kb
gentamicin-resistance cartridge derived from pMS266 and the 0.8-kb deletion in the N-
terminal region (Fig 2.7C). These results show that the menB rubA double mutant and
menF and menG single mutants are free of the respective wild-type alleles and therefore
homozygous in the menB::GmR, menF::GmR , and menGB::GmR alleles, respectively.
When the pigment extracts from the PS I complexes isolated from the menB rubA
and the menG mutants were analyzed, no peak with UV-absorption was detected at ca. 14
min, demonstrating that the menB and menG homologs are indeed required for MQ-4
synthesis in Synechococcus sp. PCC 7002 (Fig 2.9). In extracts of PS I complexes from
the menF mutant, a small peak was detected at this retention time; however, the
absorption properties of the component eluted at this retention time were significantly
different from those of MQ-4 with an absorption maximum at 292 nm (Fig 2.10B). This
component is virtually absent in the wild type and its chemical nature has not yet been
determined. These results confirm that the interruption of the menF gene also results in
the complete loss of MQ-4 and that MenF is required for MQ-4 synthesis in
Synechococcus sp. PCC 7002. In the menF and the menB rubA mutants, a peak at 35.1
min was detected, which is absent in the extracts of PS I complexes in the wild type (Fig
2.9). The component eluted at this retention time showed absorption properties with a
single peak at 256 nm (Fig 2.10C), which is characteristic of PQ-9 (Crane and Dilley,
1963). Mass spectrometric analysis of the component has shown a m/z of 748, which is
characteristic of PQ-9. These results are in good agreement with a previous study in
which the interruption of PhQ biosynthesis in Synechocystis sp. PCC 6803 resulted in the
incorporation of PQ-9 in PS I (Johnson et al., 2000). Likewise, no MQ was detectable in
the chromatogram of PS I complexes isolated from the menG mutant (Fig 2.9D). A new
peak was detected at 11.9 min, which showed a strong UV-absorption with a maximum at

44
248 nm and a shoulder at 265 nm. Because the loss of a methyl group is expected to
lower the hydrophobicity and size of a molecule, it was expected that
demethylphylloquinone would elute earlier than MQ-4 in the reverse phase HPLC
analysis. Therefore, it is highly plausible that the component eluting at 11.9 min is due to
demethylmenaquinone. Combined with the sequence similarities to E. coli counterparts,
these results confirm that menB, menG, and menF encode dihydroxynaphthoic acid-CoA
synthase, 2-geranylgeranyl-1,4-naphthoquinone methyltransferase, and isochorismate
synthase, respectively, in Synechococcus sp. PCC 7002. Together with the previous
identification of menA, menB, menD, menE, and menG in Synechocystis sp. PCC 6803, it
is concluded that PhyQ/MQ-4 biosynthesis in cyanobacteria requires MenF, MenD,
MenE, MenB, MenA, and MenG.

45
DISCUSSION
Comparative genome analyses were conducted on 14 cyanobacteria, E. coli, B.
subtilis, Halobacterium sp. NRC-1 and A. thaliana. Based on the results, it was
concluded that PhyQ biosynthesis in cyanobacteria is similar to the MQ biosynthesis
pathway in E. coli. Supporting this hypothesis is the discovery that G. violaceus and
Synechococcus sp. PCC 7002 synthesize MQ-4 instead of PhyQ and that Synechococcus
sp. PCC 7002 utilizes it as the A1 cofactor in PS I complexes. G. violaceus is thought to
be one of the earliest branching lineages among the cyanobacteria (Nelissen et al., 1995).
The fact that this organism synthesizes MQ-4 is a strong indication that PhyQ
biosynthesis probably evolved from MQ biosynthesis pathway in other bacteria. It is
therefore highly plausible that the early cyanobacteria synthesized MQ-4, and PhyQ
biosynthesis evolved from the MQ-4 biosynthesis in cyanobacteria. This is consistent
with the fact that the occurrence of PhyQ biosynthesis is restricted to cyanobacteria and
to algae and higher plants, which are evolutionarily related to cyanobacteria. A recent
study has shown that the red alga Cyanidium caldarium also synthesizes MQ-4 and
utilizes it in its PS I complexes (Yoshida et al., 2004). Therefore, MQ-4 synthesis may be
a widely occurring phenomenon among oxygenic phototrophs.
Based on phylogenetic reconstructions, it was concluded that not all men genes in
higher plants are of cyanobacterial origin. It is believed that plastids, and hence the ability
to perform oxygenic photosynthesis, in eukaryotic phototrophs were acquired from a
cyanobacterium upon endosymbiosis (Margulis, 1970). Therefore, the presence of a gene

46
with strong similarity to the cyanobacterial menA in the plant nuclear chromosome is
probably a result of gene transfer from the plastids. This gene is presumably derived from
the endosymbiotic cyanobacterium and would have been transferred to a host nuclear
chromosome during endosymbiotic interaction (Douglas, 1998; Delwiche and Palmer,
1997; Martin et al., 1998). On the other hand, the presence of the non-cyanobacterial-type
menB and menD in the plant nuclear chromosome suggests they originated in other
organisms. One possibility is that menB and menD were already present in the eukaryotic
host cells before the endosymbiotic cyanobacterium was incorporated into them. It is
believed that mitochondria in eukaryotic cells originated from a proteobacterium,
possibly in the α-subdivision (Olsen et al., 1994; Andersson et al., 1998; Martin and
Müller, 1998; Emelyanov 2001; Horner et al., 2001; Emelyanov, 2003), through an
endosymbiosis that preceded the acquisition of plastids. Given that some, although not
all, proteobacteria synthesize both ubiquinone and MQ (Collins and Jones, 1981), it is
possible that the endosymbiotic proteobacterium brought the ability to synthesize MQ
into the eukaryotic host cells. Another possibility is that menB and menD genes were
acquired via horizontal gene transfer from bacteria to plant cells. Interestingly, the men
genes in the plastid genomes of the red algae Cyanidium caldarium and Cyanidioschyzon
merolae form a cluster (menD-menF-menB-menA-menC-menE), which is very similar to
the cluster found in a variety of prokaryotes (Fig 2.2). Phylogenetic analyses show that
neither the MenA nor the MenB sequences of the red algae Cyanidioschzon merolae
forms a cluster with the cyanobacterial sequences (Fig 2.3). These observations therefore
support the occurrence of horizontal gene transfer from bacteria other than cyanobacteria
to these eukaryotic phototrophs.

47
Targeted mutagenesis of the menB, menF, and menG homologs in Synechococcus
sp. PCC 7002, encoding for DHNA synthase, isochorismate synthase, and 2-methyl-1.4-
naphthoquinone methyltransferase, respectively, resulted in the expected interruption of
PhyQ biosynthesis. This confirms that the pathway responsible for MQ-4 synthesis in
Synechococcus sp. PCC 7002 is similar to that in other bacteria.
PhyQ mediates electron transfer between A0 and FX of PS I during the charge
separation process upon photoexcitation of P700. The optimal rate of electron transfer
from A0 to FX is ensured by the thermodynamically downhill arrangement of these
cofactors. The molecular structures of PhyQ and MQ-4 are identical except for the level
of saturation of the C3 polyprenyl side chain, and thus the redox potentials of the two
quinones are also expected to be identical. Therefore, it is not surprising that MQ-4 can
function at the A1 site in the PS I complexes. The prenylation reaction of DHNA in
Synechococcus sp. PCC 7002, G. violaceus, and C. caldarium is likely to be different
from that in PhyQ-synthesizing species either by i) higher substrate specificity of the
MenA protein for geranylgeranyl diphosphate compared to phytyl diphosphate; and/or ii)
substantially higher intracellular concentration of geranylgeranyl diphosphate than phytyl
diphosphate in these cells.
The whole-genome analysis of G. violaceus seems to suggest that menB, menC,
menD, menE, and menF homologs are absent in this organism. ORFs that show similarity
to the proteins encoded by these men genes are more related to other proteins that do not
play roles in MQ-4 biosynthesis. One example is the MenB-like protein Gll2549, which
shows high similarity to 6-oxo camphor hydrolase. This enzyme is a member of the

48
crotonase superfamily that catalyzes the cleavage of carbon-carbon bonds. Therefore, it is
possible that Gll2549 participates in the PhyQ pathway and catalyzes the ring cyclization
reaction of o-succinylbenzoate. However, MenB is a very highly conserved enzyme
among a wide variety of organisms (Table 2.1), and the absence of its homolog together
with four other Men proteins in G. violaceus suggests the presence of an alternative
pathway that can lead to the synthesis of DHNA. Alternatively, it was also possible that
G. violaceus does not synthesize PhyQ, MQ-4 or related compounds at all. Previous
studies have shown that PhyQ-less mutants recruit PQ into the A1 site of PS I, and,
despite a slightly lower catalytic efficiency, as measured by the flavodoxin reduction rate,
the cells are able to grow photoautotrophically (Johnson et al., 2000). However, HPLC
analyses performed in this study demonstrated that MQ-4 is present in the whole cell
extracts of G. violaceus (see above). This result largely rules out the possibility of PQ-9
dependent PS I function in this organism. It also suggests that a part of the MQ-4
biosynthetic pathway in this organism is very different from those in the rest of the
oxygenic phototrophs analyzed in this study.
In this study, a comparative genomic approach was shown to be useful in
predicting the entire biosynthetic pathway for a secondary metabolite. Combined with
reverse genetics, one can now genetically modify the quinone species in PS I in
cyanobacteria. The advantage of this approach is that one can study the biochemical and
biophysical properties of PS I, as well as the physiology of the cells, by using a
biologically intact system. Further examples are provided in the following chapters.

49
SUMMARY
Synechococcus sp. PCC 7002 and G. violaceus were found to synthesize MQ-4
instead of PhyQ. Targeted inactivation of the menF, menB, and menG genes in
Synechococcus sp. PCC 7002 demonstrated that these genes are required for MQ-4
biosynthesis. The PS I complexes isolated from the menB rubA and menF mutants
contained PQ-9, while the PS I complexes isolated from the menG mutant contained an
UV-absorbing compound, which may be demethyl-MQ-4. Comparative genome analyses
of 14 cyanobacteria and four eukaryotic phototrophs suggested that PhyQ biosynthesis in
cyanobacteria evolved from MQ biosynthesis in bacteria and that this pathway in higher
plants may have developed as a result of mosaic evolution.

50
MATERIALS AND METHODS
Comparative Genome Analyses
The Men protein sequences of E. coli were used as queries to detect the presence
of homologs in cyanobacteria using the Blast-P search method. Retrieved sequences were
named after their locus tags in the genomes. Sequence similarities between the homologs
were obtained based on pairwise alignment by BLOSUM30 using ClustalW. Multiple
amino acid sequence alignments were also generated by BLOSUM30 using ClustalW.
For the phylogenetic reconstructions based on the MenA, MenB, and MenD sequences,
unambiguous regions in the alignments were removed. The resulting “trimmed” multiple
alignments were subjected to phylogenetic reconstruction by the Parsimony method using
PAUP*4.0 beta. The bootstrap values are results of 10,000 repetitions.
Generation of Mutant and Verification of Segregation
Wild type and mutant strains of Synechococcus sp. PCC 7002 were grown as
described previously (Frigaard et al., 2004). A DNA fragment containing the menB
coding region was amplified from the isolated genomic DNA by PCR using the primer
B1 (5’-CGTCGGGACACATCTTTCAC-3’) and the primer B2 (5’-
CGGTATCTACTCTACCAAATTGGG-3’). The resulting fragment was digested with
HindIII and BamHI, and inserted into the HindIII-BamHI sites of pUC19 (pUC19menB).
A fragment containing the menF coding region was amplified by PCR using the primer
F1 (5’-TTTGGGCAGCATCCCGGGTAAGCTGG-3’) that contains the engineered KpnI
cleavage site and the primer F2 (5’-GGAACCTATGAAGTTAGTCTGCGTTTCG-3’).
The resulting fragment was digested with KpnI and HindIII, and inserted into the KpnI-

51
HindIII fragment of the pUC19 cloning vector (pUC19menF). A fragment containing the
menG coding region was amplified by PCR using the primer G3 (5’-
GATGGTATTGTCGTCAACACCGC-3’) and the primer G4 (5’-
CACCGCCGCTATGGTACCAGGCG-3’). The resulting fragment was digested
(pUC19menG). The accC1 gene, derived from the plasmid pMS266 and conferring
gentamycin resistance, was inserted into the unique PstI site and SmaI cleavage sites in
the plasmids pUC19menB and pUC19menF, respectively. The accC1 gene was inserted
into the EcoRV sites of pUC19menG, resulting in a deletion of 740 bp encompassing the
N-terminal and the upstream region of the menG gene. The menB::accC1 construct was
used to transform the rubA mutant of Synechococcus sp. PCC 7002 as previously
described (Shen et al., 2002). The menF::accC1 construct was used to transform a psaB-
strain. After achieving complete segregation of the menF::accC1 locus, the wild type
psaB gene was introduced into the menF::accC1 psaB mutant to generate the
menF::accC1 single mutant by homologous recombination at the psaB locus. The
menG::accC1 construct was introduced into the wild-type strain. The full segregation of
each allele was tested by PCR using the primers B1 and B2 for the menB::accC1
transformants, the primers F1 and F2 for the menF::accC1 transformant, and the primers
G1 (5’- CGTTTCAATTTCCCAGGCAAAGTC-3’) and G2 (5’-
CCGAGGAAGTAAACCGCATATTC-3) for the menG::accC1 transformant.

52
Isolation of Thylakoid Membranes and PS I Particles
Thylakoid membranes were isolated from cells in the late-exponential growth
phase as described previously (Shen et al., 1993). Cells were broken at 4° C by three
passages through a French pressure cell at 124 MPa. The thylakoid membranes were
resuspended and solubilized for 2 h at 4° C in the presence of 1% (w/v) n-dodecyl-β-D-
maltoside. PS I complexes were separated from other membrane components by
centrifugation on 5-20% (w/v) sucrose gradients with 0.05% n-dodecyl-β-D-maltoside in
the buffer. Further purification was achieved by a second centrifugation on sucrose
gradients in the buffer in the absence of n-dodecyl-β-D-maltoside. Trimeric PS I
complexes were used in these studies (Golbeck, 1995).
Quinone and Chlorophyll Analysis
Isolated PS I was either lyophilized or dried under N2 gas before extraction with
acetone:methanol (7:2 v/v). Pigments and quinones were extracted with cold
acetone:methanol (7:2 v/v) after brief ultrasonication. Alternatively, 20 µL of the PS I
preparations were extracted with 400 µL of acetone:methanol. After centrifugation and
filtration through a PTFE filter membrane with a 0.2-µm pore size (Whatman
International Ltd., Maldstone, UK), the extract in the organic phase was directly injected
into an Agilent Technology 1100 series HPLC system (Agilent Technology, Palo Alto,
CA, USA) equipped with a SUPELCO (Sigma-Aldrich Corp., St. Louis, MO, USA)
Discovery® C18 column (25 cm x 4.6 mm, 5 µm). Analyses were carried out using the
following protocol: 80%A/20%B from 0 to10 min, a linear change to 20%A/80%B in 40
min, and 20%A/80%B for 5 min, where solvent A and solvent B are 100% methanol and
100% isopropanol, respectively. The flow rate was 0.75 ml min-1. Detection of eluates
was performed with a diode array detector (Agilent 1100 series). The chlorophyll a,

53
phylloquinone, and plastoquinone contents of each sample were determined based on
integrated peak areas and their molar absorption coefficients, which are 17.4 mM-1 cm-1
at 618 nm, 18.9 mM-1 cm-1 at 270 nm (Dunphy and Brodie, 1971), and 15.2 mM-1 cm-1 at
254 nm (Crane and Dilley, 1963), respectively. The absorption coefficient of chlorophyll
a at 618 nm was calculated from the ratio of the absorption peaks at 618 nm and 666 nm
in methanol-isopropanol (6:4, v/v) and from its absorption coefficient at 666 nm in
methanol (MacKinney, 1941). The peak assignments were confirmed by mass
spectrometry using atmospheric-pressure chemical ionization with a Quattro II time-of-
flight mass spectrometer (Micromass, Beverly, MA, USA) operated in the negative-ion
mode.

54
REFERENCES
Andersson SG, Zomorodipour A, Andersson JO, Sicheritz-Ponten T, Alsmark UC,
Podowski RM, Naslund AK, Eriksson AS, Winkler HH, Kurland CG (1998) The genome
sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396: 133-140
Brettel K, Leibl W (2001) Electron transfer in Photosystem I. Biochim Biophys Acta
1507: 100-114
Collins MD and Jones D (1981) Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiological Reviews 45: 316-354
Crane F.L., and Dilley R.A (1963) Determination of coenzyme Q (ubiquinone). In: Glick
D (ed), Methods of Biochemical Analysis, 4: 279-306
Delwiche CF, Palmer JD (1997) The origin of plastids and their spread via secondary
endosymbiosis. In Bhattacharya (ed), Origins of Algae and Their Plastids. pp 53-86.
Springer-Verlag, New York
Douglas SD (1998) Plastid evolution: origins, diversity, trends. Curr Opin Genet Dev 8:
655-661
Dufresne A, Salanoubat M, Partensky F, Artiguenave F, Axmann IM, Barbe V, Duprat S,
Galperin MY, Koonin EV, Le Gall F, Makarova KS, Ostrowski M, Oztas S, Robert C,
Rogozin IB, Scanlan DJ, Tandeau de Marsac N, Weissenbach J, Wincker P, Wolf YI,
Hess WR (2003) Genome sequence of the cyanobacterium Prochlorococcus marinus

55
SS120, a nearly minimal oxyphototrophic genome. Proc Natl Acad Sci USA 100: 9647-
9649
Dunphy PJ, Brodie AF (1971) The structure and function of quinones in respiratory
metabolism. Methods Enzymol 18: 407-461
Emelyanov VV (2001) Evolutionary relationship of Rickettsiae and mitochondria.
FEBS Lett 501: 11-18
Emelyanov VV (2003) Common evolutionary origin of mitochondrial and richettsial
respiratory chains. Arch Biochem Biophys 420: 130-141
Frigaard N.-U, Sakuragi Y., and Bryant D. A. (2004) Gene inactivation in the
cyanobacterium Synechococcus sp. PCC 7002 and the green sulfur bacterium
Chlorobium tepidum using in vitro-made DNA constructs and natural transformation. In:
R. Carpentier (ed), Photosynthesis Research Protocols, Methods in Molecular Biology,
Vol, 274. Humana Press, Totowa. In press
Golbeck JH (1994) Photosystem I in cyanobacteria. In: Bryant DA (ed), The Molecular
Biology of Cyanobacteria, pp 319-360. Kleuwer Academic Publishers, Dordrecht
Golbeck, J. H. (1995) In: Song PS (ed), in CRC Handbook of Organic Photochemistry
and Photobiology, Vol. 1, pp 1407-1419. CRC Press, Bocca Raton, Florida
Golbeck JH (2003) The binding of cofactors to Photosystem I analyzed by spectroscopic
and mutagenic methods. Annu Rev Biophys Biomol Struct 32: 237-256

56
Horner DS, Embley TM (2001) Chaperonin 60 phylogeny provides further evidence for
secondary loss of mitochondria among putative early-branching eukaryotes. Mol Biol
Evol 18: 1970-1975
Johnson TW, Shen G, Zybailov B, Folling D, Reategui R, Beauparlant S, Vassiliev IR,
Bryant DA, Jones AD, Golbeck JH, Chitnis PR (2000) Recruitment of a foreign quinone
into the A1 site of Photosystem I: I. Genetic and physiological characterization of
phylloquinone biosynthetic pathway mutants in Synechocystis sp. PCC 6803. J Biol
Chem 275: 8523-8530
Johnson TW, Naithani S, Zybailov B, Jones AD, Golbeck JH, Chitnis P (2002) The
menD and menE homologus code for 2-succinyl-6-hydroxyl-2,4-cyclohexadiene-1-
carboxylate synthase and O-succinylbenzoic acid-CoA ligase in the phylloquinone
biosynthetic pathway of Synechocystis sp. PCC 6803. Biochim Biophys Acta 45220: 1-10
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauß N (2001) Three-dimensional
structure of cyanobacterial Photosystem I at 2.5 Å resolution. Nature 411: 896-899
Kaneko T, Nakamura Y, Wolk CP, Kuritz T, Sasamoto T, Watanabe A, Iriguchi M,
Ishikawa M, Kawashima M, Kimura T, Kishida Y, Kohara M, Matsumoto M, Matsuno
A, Muraki A, Nakazaki N, Shimpo S, Sugimoto M, Takazawa M, Yamada M, Yasuda M,
Tabata S (2001) Complete genomic sequence of the filamentous nitrogen-fixing
cyanobacterium Anabaena sp. strain PCC 7120. DNA Res 8: 205-213
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y, Miyajima N, Hirosawa
M, Sugiura M, Sasamoto S, Kimura T, Hosouchi T, Matsuno A, Muraki A, Nakazaki N,

57
Naruo K, Okumura S, Shimpo S, Takeuchi C, Wada T, Watanabe A, Yamada M, Yasuda
M, Tabata S (1996) Sequence analysis of the genome of the unicellular cyanobacterium
Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and
assignment of potential protein-coding regions. DNA Res 3: 109-136
MacKinney G (1941) Absorption of light by chlorophyll solutions. J Biol Chem 140:
315-322
Margulis L (1970) Origin of eukaryotic cells. Yale University Press, New Haven
Martin W, Müller M (1998) The hydrogen hypothesis for the first eukaryote. Nature 392:
370-341
Martin W, Stoebe B, Goremykin V, Hansmann S, Hasegawa M, Kowallik KV (1998)
Gene transfer to the nucleus and the evolution of chloroplasts. Nature 393: 162-165
Meganathan R (1996) Biosynthesis of the isoprenoid quinones-menaquinone (vitamin
K2) and ubiquinone (coenzyme Q). In: Escherichia coli and Salmonella-Cellular and
Molecular Biology, Vol, 2, pp 642-656. ASM press, Washington DC
Meganathan R (2001) Biosynthesis of menaquinone (vitamin K2) and ubiquinone
(coenzyme Q): a perspective on enzymatic mechanisms. Vitam horm 61: 173-218
Nakamura Y, Kaneko T, Sato S, Ikeuchi M, Katoh H, Sasamoto S, Watanabe A, Iriguchi
M, Kawashima K, Kimura T, Kishida Y, Kiyokawa C, Kohara M, Matsumoto M,
Matsuno A, Nakazaki N, Shinpo S, Sugimoto M, Takeuchi C, Yamada M, Tabata S

58
(2002) Complete genome structure of the thermophilic cyanobacterium
Thermosynechococcus elongatus BP-1. DNA Res 9: 2-130
Nakamura Y, Kaneko T, Sato S, Mimuro M, Miyashita H, Tsuchiya T, Sasamoto S,
Watanabe A, Kawashima K, Kishida Y, Kiyokawa C, Kohara M, Matsumoto M,
Matsuno A, Nakazaki N, Shimpo S, Takeuchi C, Yamada M, Tabata S. Complete
genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks
thylakoids DNA Res 10: 137-145
Nelissen B, van de Peer Y, Wilmotte A, de Wachter R (1995) An early origin of plastids
within the cyanobacterial divergence is suggested by evolutionary trees based on
complete 16 S rRNA sequences. Mol Biol Evol 12: 1166-1173
Olsen GJ, Woese CR, Overbeek R (1994) The winds of (evolutionary) change: breathing
new life into microbiology. J Bacteriol 176:1-6
Palenik B, Brahamsha B, Larimer FW, Land M, Hauser L, Chain P, Lamerdin J, Regala
W, Allen EE, McCarren J, Paulsen I, Dufresne A, Partensky F, Webb EA, Waterbury J.
(2003) The genome of a motile marine Synechococcus. Nature 424: 1037-1042
Pennock JF, Threlfall DR (1981) Biosynthesis of ubiquinone and related compounds. In:
Porter JW and Spurgeon SL (eds), Biosynthesis of Isoprenoid Compounds. Vol 2, pp
191-303 A Wiley-Interscience Publication, New York
Rocap G, Larimer FW, Lamerdin J, Malfatti S, Chain P, Ahlgren NA, Arellano A,
Coleman M, Hauser L, Hess WR, Johnson ZI, Land M, Lindell D, Post AF, Regala W,

59
Shah M, Shaw SL, Steglich C, Sullivan MB, Ting CS, Tolonen A, Webb EA, Zinser ER,
Chisholm SW (2003) Genome divergence in two Prochlorococcus ecotypes reflects
oceanic niche differentiation. Nature 424: 1042-1047
Shen GZ, Boussiba S, Vermaas WFJ (1993) Synechocystis sp PCC 6803 strains lacking
Photosystem I and phycobilisome function. Plant Cell 5: 1853-1863
Shen G, Zhao J, Reimer SK, Antonkine ML, Cai Q, Weiland SM, Golbeck JH, Bryant
DA (2002) Asembly of Photosystem I; I. Inactivation of the rubA gene encoding a
membrane-associated rubredoxin in the cyaobacterium Synechococcus sp. PCC 7002
causes a loss of Photosystem I activity. J Biol Chem 277: 20343-20354
Thornber JP, Alberte RS, Hunter FA, Shiozawa JA, Kan K-S (1976) The organization of
chlorophyll in the plant photosynthetic unit. Brookhaven Symposia in Biology 28: 132-
148
Yoshida E, Nakamura A, Watanabe (2003) Reversed-phase HPLC determination of
chlorophyll a’ and naphthoquinones in Photosystem I of red algae: existence of two
menaquinone-4 molecules in Photosystem I of Cyanidium caldarium. Anal Sci 19: 1001-
1005

60
Tab
le 2
.1: W
hole
gen
ome
anal
yses
for
the
MQ
/Phy
Q b
iosy
nthe
sis e
nzym
es in
14
cyan
obac
teri
a, E
. col
i, B.
subt
ilis,
C.
tepi
dum
, and
Hal
obac
teriu
m sp
. NR
C-1
. Sim
ilarit
y to
Syn
echo
cyst
is sp
. PC
C 6
803
prot
eins
are
show
n in
par
enth
eses
(%).
The
sim
ilarit
y is
bas
ed p
airw
ise
alig
nmen
t of t
he e
ntire
sequ
ence
s con
stru
cted
by
BLO
SUM
30. (
Tabl
e co
ntin
ues o
n ne
xt p
age)
S.
680
3 S.
700
2 N
osto
c T.
ery
N
. pun
A.
var
C
. wat
S.
elo
T.
elo
G
. vio
Men
A
Slr1
518
b
(67%
) A
lr003
3 (7
0%)
Tery
1070
(7
3%)
Npu
n060
4 (7
0%)
Ava
r026
348
(68
%)
a Cw
at14
3901
(3
4%)
Selo
0206
47
(66%
) Tl
r219
6 (7
0%)
Gll1
578
(53%
)
Men
B
Sll1
127
b (8
9%)
All2
347
(91%
) Te
ry10
85
(83%
) N
pun2
498
(91%
) A
var4
1360
1 (9
1%)
Cw
at15
9701
(9
1%)
Selo
1873
01
(89%
) Tl
l245
8 (8
8%)
-
Men
C
Sll0
409
b (5
2%)
Alr0
034
(54%
) Te
ry10
67
(54%
) N
pun0
605
(52%
) A
var0
2638
2 (5
4%)
- Se
lo02
0304
(5
0%)
Tlr1
174
(55%
) -
Men
D
Sll0
603
b (6
2%)
Alr0
312
(61%
) Te
ry10
75
(61%
) N
pun2
496
(59%
) -
Cw
at13
1201
(6
1%)
Selo
0271
8 (6
1%)
Tll0
130
(56%
) -
Men
E Sl
rl049
2 b
(44%
) A
lr003
5 (5
6%)
Tery
1066
(5
6%)
Npu
n060
6 (5
4%)
Ava
r026
381
(52%
) C
wat
0260
26
(52%
) Se
lo02
305
(45%
) Tl
l122
1 (5
0%)
-
Men
F Sl
r081
7 b
(54%
) A
ll003
2 (5
8%)
Tery
al71
(5
7%)
Npu
n060
3 (6
0%)
Ava
r026
437
(58%
) -
Selo
0214
74
(53%
) Tl
l121
3 (5
4%)
-
Men
G
Slr1
653
b (7
0%)
Alr5
252
(72%
) Te
ry33
21
(63%
) N
pun3
758
(65%
) a A
var2
8710
1 (4
8%)
Cw
at17
9201
(6
8%)
Selo
0203
83
(66%
) Tl
l237
3 (6
3%)
Gll0
127
(53%
)
Men
H
Sll1
916
b (4
3%)
All0
111
(68%
) Te
ry19
86
(46%
) N
pun0
061
(55%
) A
var5
1370
1 (6
5%)
Cw
at02
4832
(4
5%)
Selo
0200
62
(52%
) Tl
r206
6 (4
3%)
-
S. 6
803,
Syn
echo
cyst
is sp
. PC
C 6
803;
S.7
002,
Syn
echo
cocc
us sp
. PC
C 7
002;
Nos
toc,
Nos
toc
sp. P
CC
712
0; N
. pun
, Nos
toc
punc
tifor
me;
T.e
lo, T
. elo
ngat
us
BP-
1; T
. ery
, Tri
chod
esm
ium
ery
thra
eum
; G.v
io, G
. vio
lace
us P
CC
742
1; A
.var
, Ana
baen
a va
riab
ilis A
TCC
294
13; S
. elo
, Syn
echo
cocc
us e
long
atus
sp.
7942
; C.w
at, C
roco
spha
era
wat
soni
i WH
8501
. a N-te
rmin
al se
quen
ce w
as tr
unca
ted.
b No
locu
s tag
s are
ava
ilabl
e.

61
Tabl
e 2.
1 co
ntin
ued
S. 8
102,
Syn
echo
cocc
us sp
. WH
8102
; P.M
ED4,
Pro
chlo
roco
ccus
mar
inus
MED
4; P
.MIT
, Pro
chlo
roco
ccus
mar
inus
MIT
9313
; P.S
S120
, Pro
chlo
roco
ccus
mar
inus
SS1
20; A
rabi
dops
is, A
rabi
dops
is t
halia
na; E
. col
i, Es
cher
ichi
a co
li; C
. tep
, Chl
orob
ium
tepi
dum
; B. s
ub, B
acill
us su
btili
s. a N
-term
inal
sequ
ence
was
trun
cate
d.
S.
810
2 P.
MED
4 P.
MIT
P.
SS12
0 Ar
abid
opsi
s E.
col
i C
. tep
B.
sub
Men
A
Synw
2307
(5
6%)
Pmm
0176
(5
8%)
Pmt2
059
(55%
) Pr
o020
1 (5
5%)
At1
g606
00
(55%
) B
3930
(4
7%)
CT1
1511
(4
4%)
Bsu
3849
0 (4
1%)
Men
B
Synw
0998
(8
0%)
Pmm
0608
(7
7%)
Pmt0
405
(79%
) Pr
o105
3 (7
9%)
At1
g605
50
(78%
) B
2262
(7
8%)
CT1
846
(77%
) B
su30
75
(81%
)
Men
C
Synw
2306
(3
9%)
Pmm
0175
(4
1%)
Pmt2
058
(43%
) Pr
o020
0 43
%)
At1
g689
00
(46%
) B
2261
(3
8%)
CT1
847
(35%
) B
su30
780
(35%
)
Men
D
Synw
0997
(4
6%)
Pmm
0607
(4
6%)
Pmt0
406
(47%
) Pr
o105
4 (4
6%)
At1
g688
90
(45%
) B
2264
(4
3%)
CT1
839
(44%
) B
su30
77
(49%
)
Men
E Sy
nw23
05
(35%
) Pm
m01
74
(34%
) Po
mt2
057
(36%
) Pr
o019
9 (3
5%)
At3
g489
90
(43%
) B
2260
(4
6%)
CT1
848
(39%
) B
su30
74
(39%
)
Men
F Sy
nw23
08
(42%
) Pm
m01
77
(48%
) Pm
t206
0 (4
7%)
Pro0
202
(47%
) A
t1g7
4710
(5
1%)
B22
65
(44%
) C
T183
8 (4
4%)
Bsu
3078
(4
7%)
Men
G
Synw
1674
(5
6%)
Pmm
0431
(5
1%)
Pmt0
276
(37%
) Pr
o042
7 (5
7%)
At1
g233
60
(50%
) B
3833
(4
7%)
CT0
462
(52%
) B
su22
750
(45%
)
Men
H
Synw
0681
(3
8%)
Pmm
1628
(3
5%)
Pmt0
128
(38%
) Pr
o179
0 (4
0%)
At5
g385
20
(43%
) B
2263
(3
5$)
CT1
845
(31%
) B
su30
76
(34%
)
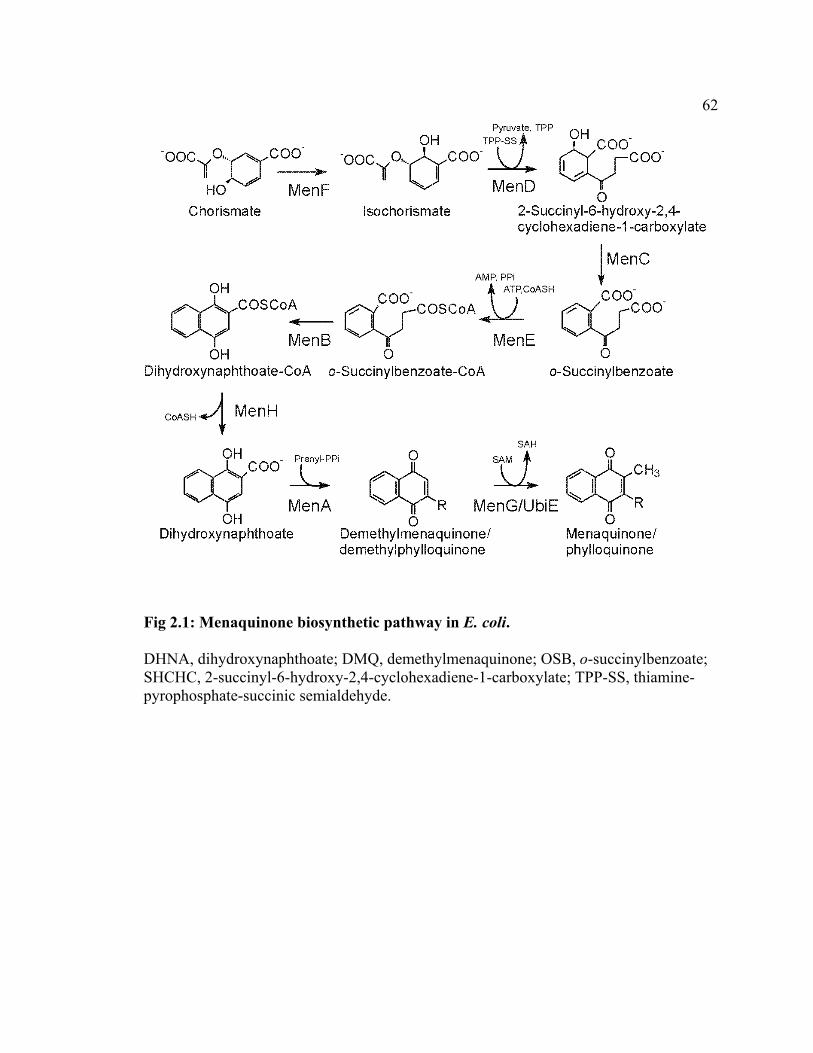
62
Fig 2.1: Menaquinone biosynthetic pathway in E. coli.
DHNA, dihydroxynaphthoate; DMQ, demethylmenaquinone; OSB, o-succinylbenzoate; SHCHC, 2-succinyl-6-hydroxy-2,4-cyclohexadiene-1-carboxylate; TPP-SS, thiamine-pyrophosphate-succinic semialdehyde.
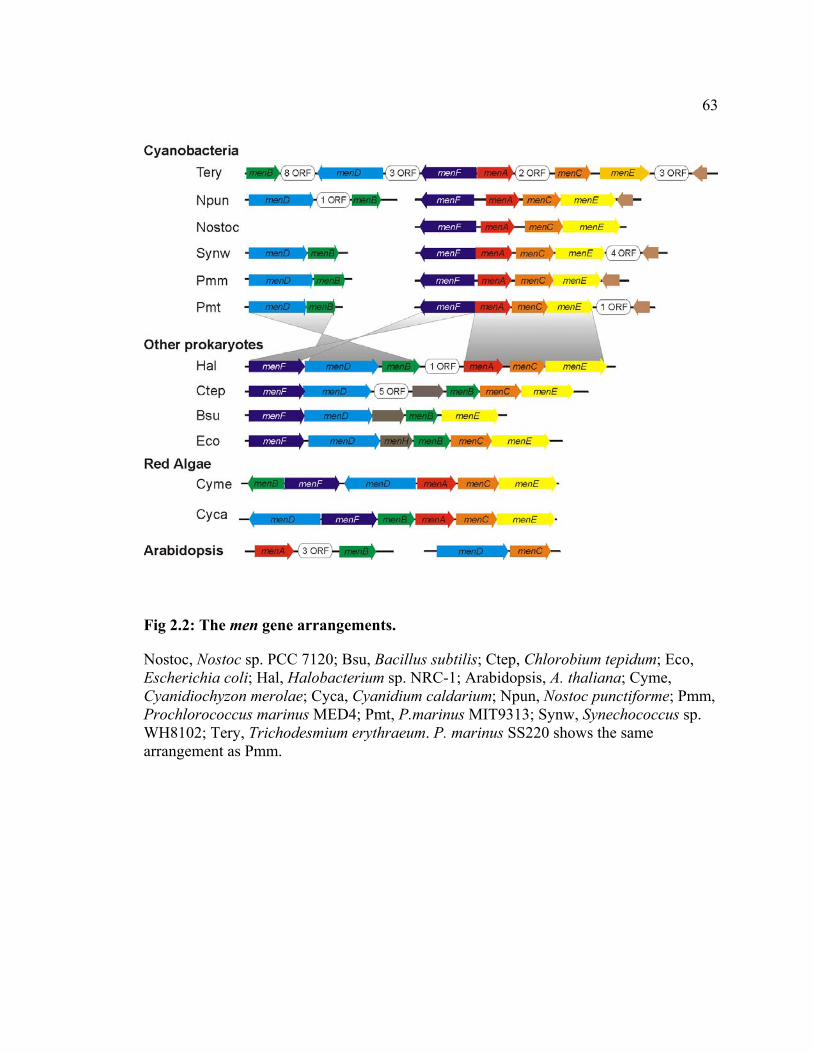
63
Fig 2.2: The men gene arrangements.
Nostoc, Nostoc sp. PCC 7120; Bsu, Bacillus subtilis; Ctep, Chlorobium tepidum; Eco, Escherichia coli; Hal, Halobacterium sp. NRC-1; Arabidopsis, A. thaliana; Cyme, Cyanidiochyzon merolae; Cyca, Cyanidium caldarium; Npun, Nostoc punctiforme; Pmm, Prochlorococcus marinus MED4; Pmt, P.marinus MIT9313; Synw, Synechococcus sp. WH8102; Tery, Trichodesmium erythraeum. P. marinus SS220 shows the same arrangement as Pmm.
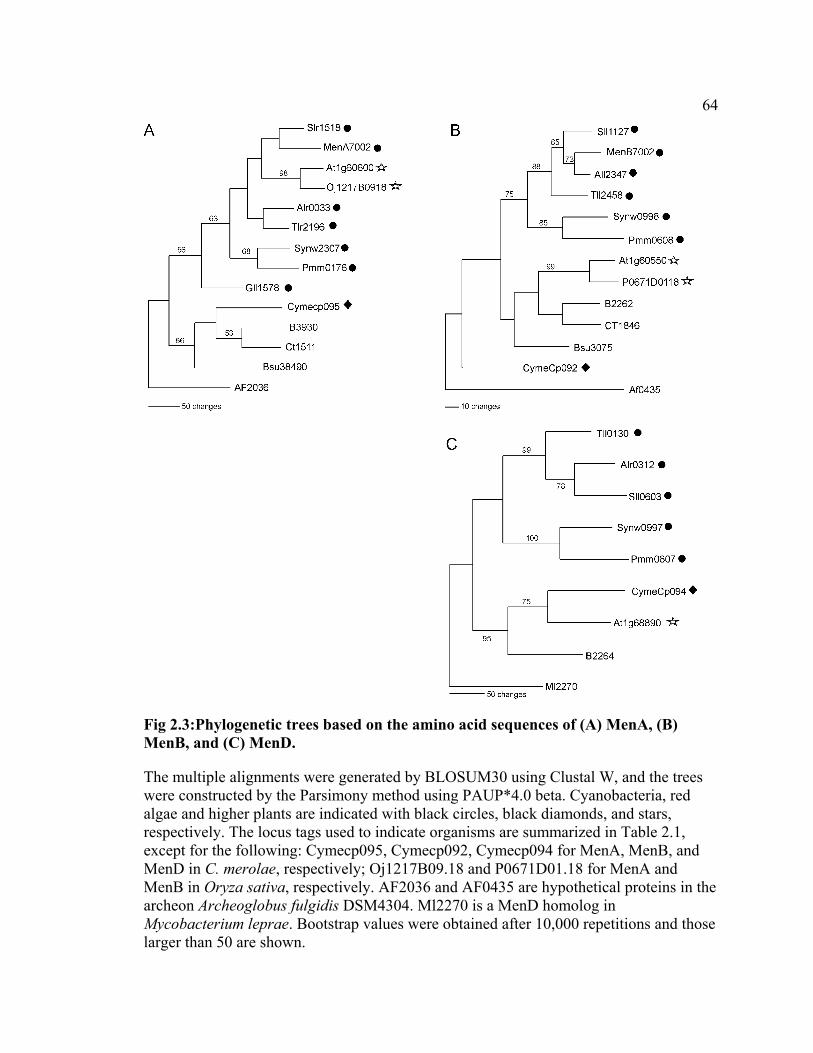
64
Fig 2.3:Phylogenetic trees based on the amino acid sequences of (A) MenA, (B) MenB, and (C) MenD.
The multiple alignments were generated by BLOSUM30 using Clustal W, and the trees were constructed by the Parsimony method using PAUP*4.0 beta. Cyanobacteria, red algae and higher plants are indicated with black circles, black diamonds, and stars, respectively. The locus tags used to indicate organisms are summarized in Table 2.1, except for the following: Cymecp095, Cymecp092, Cymecp094 for MenA, MenB, and MenD in C. merolae, respectively; Oj1217B09.18 and P0671D01.18 for MenA and MenB in Oryza sativa, respectively. AF2036 and AF0435 are hypothetical proteins in the archeon Archeoglobus fulgidis DSM4304. Ml2270 is a MenD homolog in Mycobacterium leprae. Bootstrap values were obtained after 10,000 repetitions and those larger than 50 are shown.

65
Fig2.4: HPLC profiles of solvent extracts from the PS I complexes and whole cells. Chromatograms of reverse-phase HPLC analysis of solvent extracts obtained from (A) PS I complexes isolated from the wild-type Synechocystis sp. PCC 6803 (B) PS I complexes isolated from wild-type Synechococcus sp. PCC 7002, and (C) whole cells of the wild-type G. violaceus. Chromatograms were recorded at 270 nm. Chl a, chlorophyll a; β-car, β-carotene; MQ-4, menaquinone-4.

66
Fig 2.5: Absorption spectra of solvent-extracted components analyzed by HPLC.
Components eluting (A) at 22.5 min from the PS I complex of the wild-type Synechocystis sp. PCC 6803, (B) at 14.5 min from the PS I complex of the wild-type Synechococcus sp. PCC 7002, and (C) at 14.5 min from the whole cell of the wild-type G. violaceus (solid line) (see Fig 2.4). A component eluting at 22.5 min in G. violaceus is also shown by dotted line and does not appear to be a quinone.
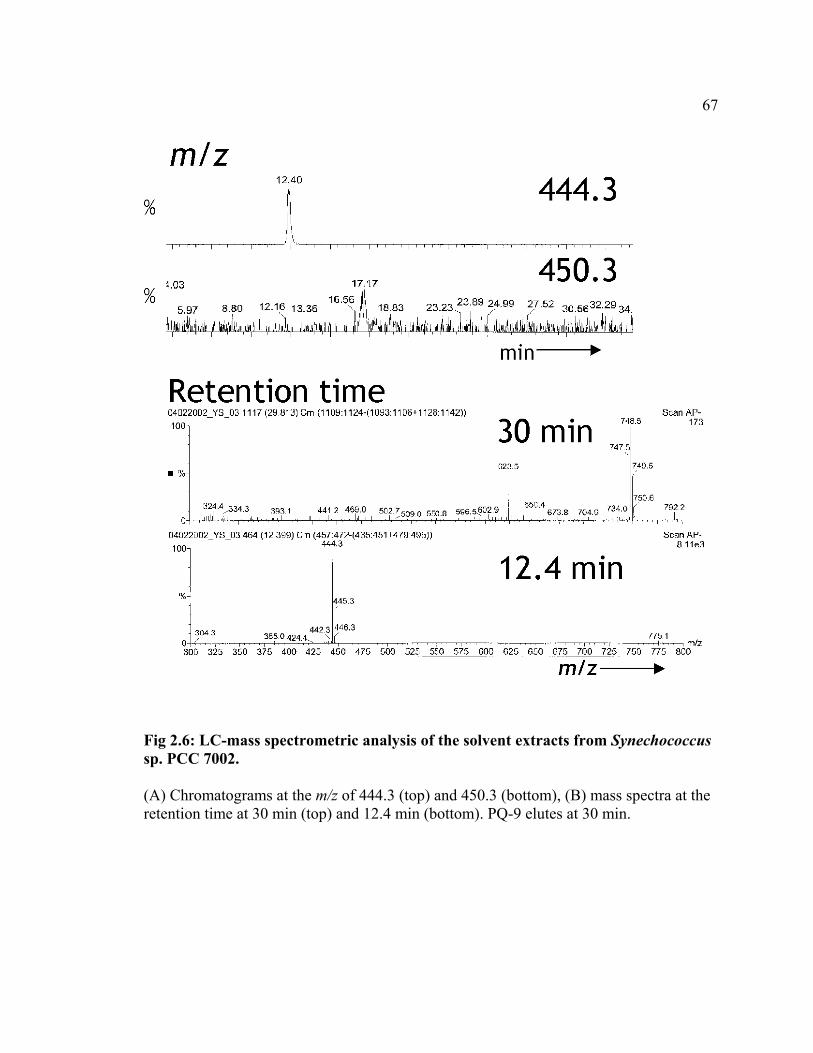
67
Fig 2.6: LC-mass spectrometric analysis of the solvent extracts from Synechococcus sp. PCC 7002. (A) Chromatograms at the m/z of 444.3 (top) and 450.3 (bottom), (B) mass spectra at the retention time at 30 min (top) and 12.4 min (bottom). PQ-9 elutes at 30 min.
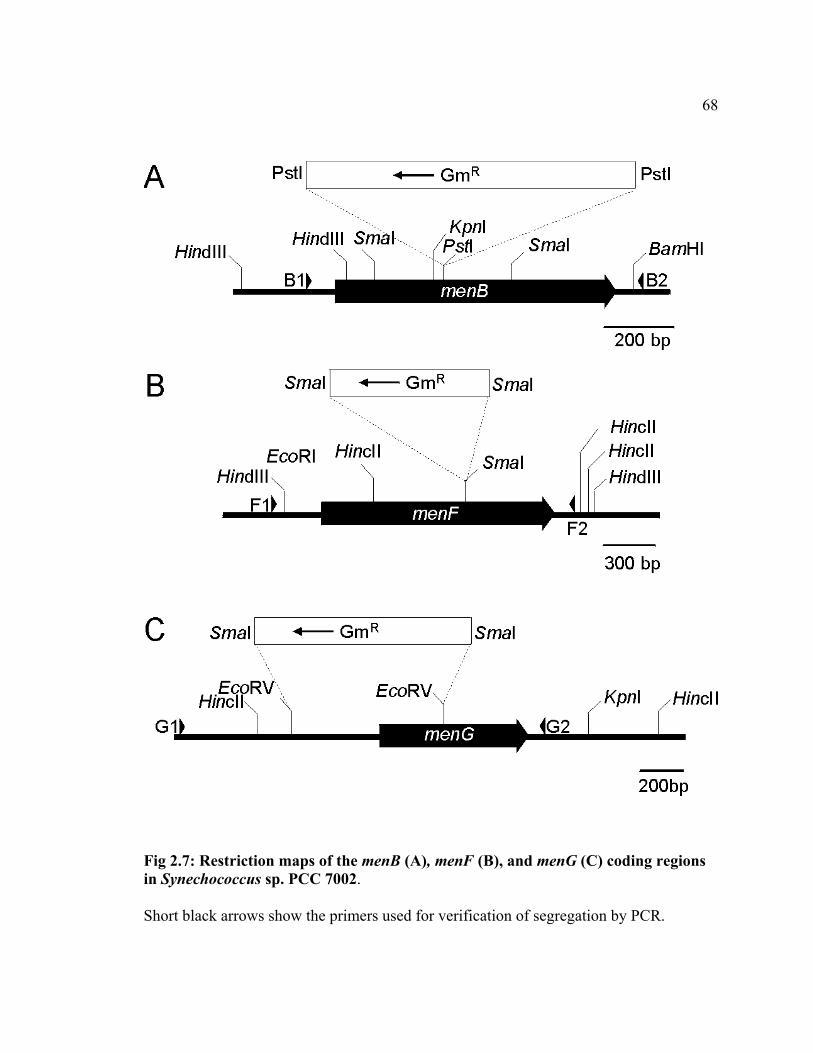
68
Fig 2.7: Restriction maps of the menB (A), menF (B), and menG (C) coding regions in Synechococcus sp. PCC 7002. Short black arrows show the primers used for verification of segregation by PCR.
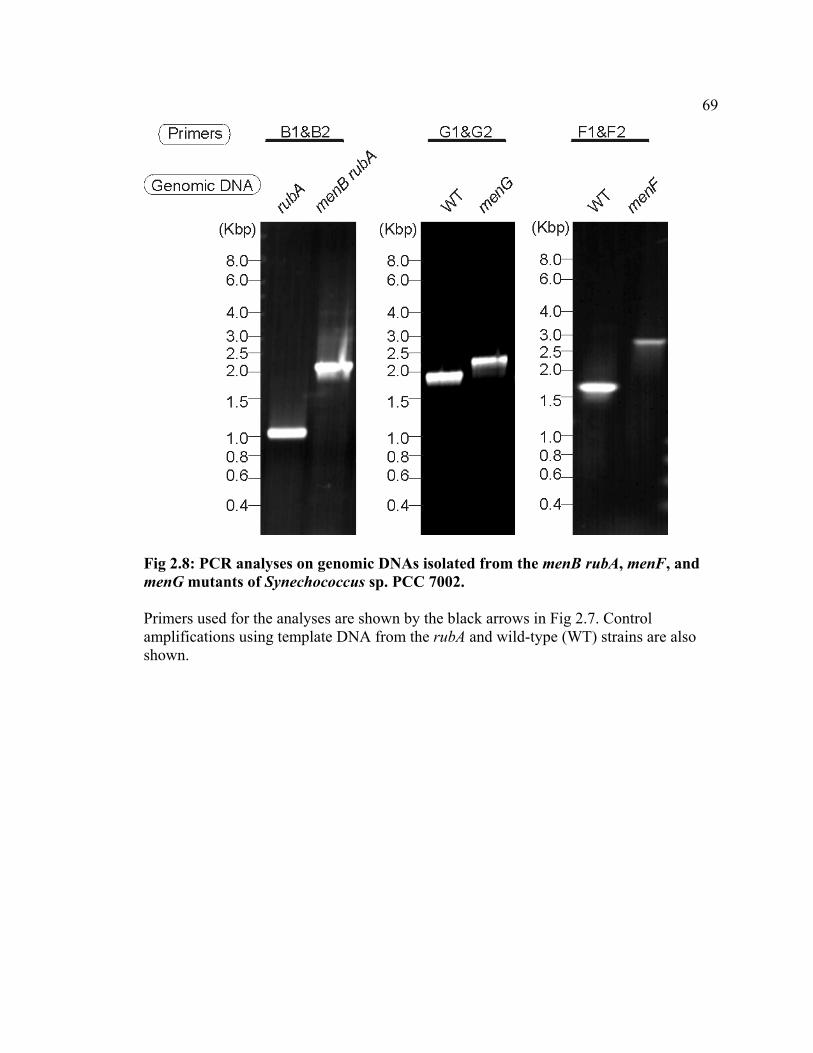
69
Fig 2.8: PCR analyses on genomic DNAs isolated from the menB rubA, menF, and menG mutants of Synechococcus sp. PCC 7002. Primers used for the analyses are shown by the black arrows in Fig 2.7. Control amplifications using template DNA from the rubA and wild-type (WT) strains are also shown.

70
Fig 2.9: Reverse-phase HPLC analyses of the solvent extracts from PS I complexes. (A) wild type, (B) the menB rubA mutant, (C) the menF mutant, and (D) the menG mutant of Synechococcus sp. PCC 7002. The retention time for MQ-4 is indicated by the black arrows.
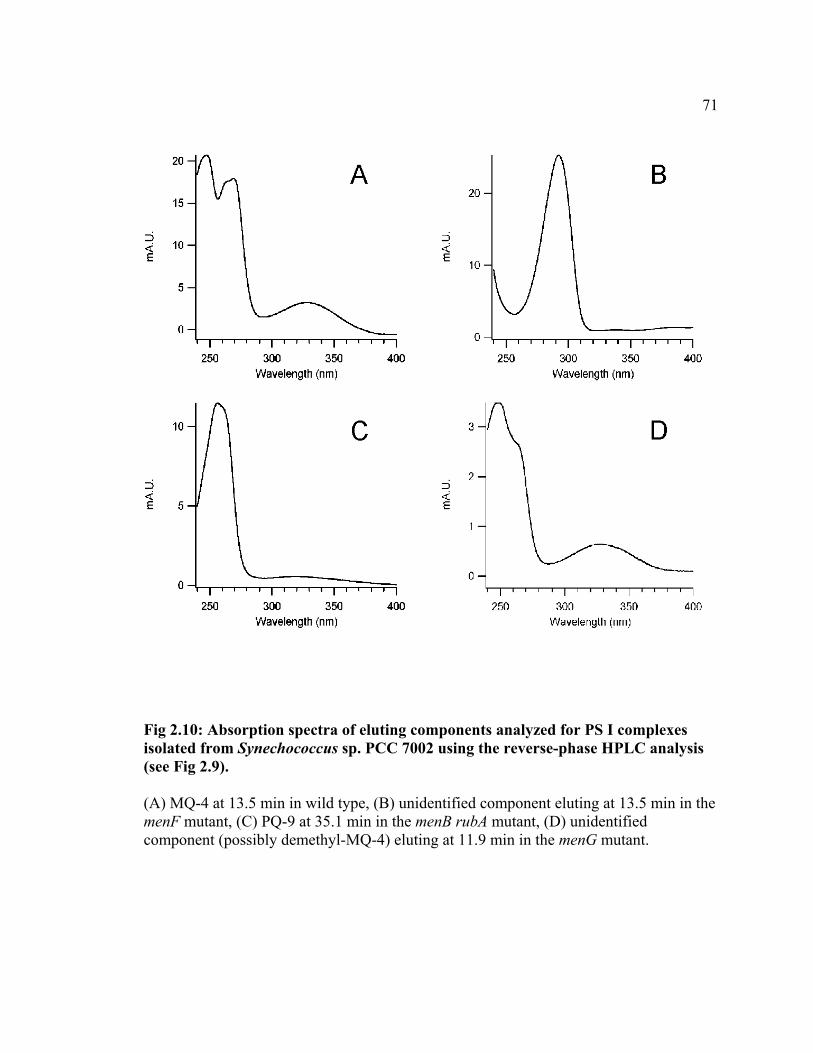
71
Fig 2.10: Absorption spectra of eluting components analyzed for PS I complexes isolated from Synechococcus sp. PCC 7002 using the reverse-phase HPLC analysis (see Fig 2.9). (A) MQ-4 at 13.5 min in wild type, (B) unidentified component eluting at 13.5 min in the menF mutant, (C) PQ-9 at 35.1 min in the menB rubA mutant, (D) unidentified component (possibly demethyl-MQ-4) eluting at 11.9 min in the menG mutant.

72
Chapter 3
Recruitment of a Foreign Quinone into the A1 Site of Photosystem I.
Interruption of menG in the Phylloquinone Biosynthetic Pathway of Synechocystis sp.
PCC 6803 Results in the Incorporation of 2-Phytyl-1,4-Naphthoquinone and
Alteration of the Equilibrium Constant between A1 and FX
Publication:
Yumiko Sakuragi, Boris Zybailov, Gaozhong Shen, Parag R. Chitnis, Art van der Est,
Robert Bittl, Stephan Zech, Dietmar Stehlik, John H. Golbeck and Donald A. Bryant.
(2002) Biochemistry 41: 394-405

73
ABSTRACT
A gene (menG), encoding a methyltransferase was identified in Synechocystis sp. PCC
6803 as responsible for transferring the methyl group to 2-phytyl-1,4-naphthoquinone
(demethylphylloquinone) in the biosynthetic pathway of phylloquinone, the secondary
electron acceptor in Photosystem I (PS I). Mass spectrometric measurements showed that
targeted inactivation of the menG gene prevents the synthesis of phylloquinone and leads
to the accumulation of 2-phytyl-1,4-naphthoquinone in PS I complexes. Growth rates of
the wild-type and the menG mutant strains under photoautotrophic and photomixotrophic
conditions were virtually identical. The chlorophyll a content in whole cells of the menG
mutant was similar to that in the wild type when the cells were grown at a light intensity of
50 µE m-2 s-1 but slightly lower when grown at 150 µE m-2 s-1. Low temperature chlorophyll
fluorescence emission measurements showed a larger increase in the ratio of PS II to PS I
in the menG mutant strain relative to the wild type as the light intensity was elevated from
50 µE m-2 s-1 to 300 µE m-2 s-1. CW EPR studies at 34 GHz and transient EPR studies at
multiple frequencies showed that the quinone radical in the menG mutant has the same
overall linewidth as in the wild type, but the pattern of hyperfine splittings showed two
lines in the low-field region consistent with the presence of an aromatic proton at ring
position 3. The spin polarization pattern indicated that 2-phytyl-1,4-naphthoquinone is in
the same orientation as phylloquinone, and out-of-phase, spin-echo modulation
spectroscopy showed the same P700+ to Q- center-to-center distance in the two samples.
Transient EPR studies indicated that forward electron transfer from Q- to FX is slowed from

74 290 ns in the wild-type to 600 ns in the menG mutant. The redox potential of
2-phytyl-1,4-naphthoquinone was estimated to be 50 to 60 mV more oxidizing than
phylloquinone in the A1 site, which translates to a lowering of the equilibrium constant
between Q-/Q and FX-/FX by a factor of ca. 10. The kinetics of the P700+ [FA/FB]-
backreaction increased from 80 msec in the wild-type to 20 msec in the menG mutant
strain, thus supporting a thermally-activated uphill electron transfer through the quinone
rather than a direct route for charge recombination between [FA/FB]- and P700+.

75
ABBREVIATIONS
bp basepair(s)
Chl chlorophyll
CW continuous wave
ENDOR electron nuclear double resonance
EPR electron paramagnetic resonance
HEPES 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid
HPLC high performance liquid chromatography
kb kilobase
MS mass spectrometry
NQ naphthoquinone
ORF open reading frame
PCR polymerase chain reaction
PhyQ phylloquinone
PQ-9 plastoquinone-9
PS photosystem
Tricine N- [2-hydroxy-1,1-bis(hydroxymethylethyl)]glycine

76
INTRODUCTION
Photosystem I (PS I) in cyanobacteria is a membrane-integral pigment-protein
complex consisting of 12 protein subunits (PsaA through PsaF, PsaI through PsaM, and
PsaX) and a variety of cofactors, including 96 molecules of chlorophyll a (Chl a), two
molecules of 2-phytyl-3-methyl-1,4-naphthoquinone (phylloquinone, PhyQ), three
[4Fe-4S] clusters, and about 22 molecules of β-carotene (Jordan et al., 2001). After the
absorption of a photon by one of the antenna molecules, charge separation takes place
between a special pair of Chl a molecules (P700) and a Chl a monomer (A0). The
charge-separated state is stabilized on the donor side by electron transfer from
plastocyanin or cytochrome c6 to P700+, and on the acceptor side by electron transfer to
PhyQ in the A1 site, through the iron-sulfur clusters at FX, FA and FB, and ultimately to
ferredoxin or flavodoxin. This light-driven enzyme therefore functions as cytochrome
c6/plastocyanin:ferredoxin/flavodoxin oxidoreductase that ultimately provides reducing
power to the cell in the form of NADPH.
Two molecules of PhyQ are present per P700 in PS I complexes of Synechocystis
sp. PCC 6803 and Synechococcus sp. PCC 7942 (Biggins and Mathis, 1988; Malkin, 1986;
Schoeder and Lockau, 1986). Both quinones have been located on the 2.5 Å-resolution
electron density map of Synechococcus elongatus PS I complexes (Jordan et al., 2001), and
there is a good agreement with EPR studies that provide distance (Bittl et al., 1997) and
orientation (Kamlowski et al., 1998; Zech et al., 2000) information for the EPR-active
quinone. For example, the same orientation of the carbonyl oxygen bonds of PhyQ, tilted

77 ca. 30 degrees from the membrane plane, is deduced from the orientation of the magnetic
gxx [(Zech et al., 2000) and earlier references therein] and the electron density map (Jordan
et al., 2001). PhyQ is structurally related to menaquinone-9 (2-
nonaprenyl-3-methyl-1,4-naphthoquinone). Both have 3-methyl-1,4-naphthoquinone core
structures with a poly-isoprenoid chain at the C2 ring position: menaquinone has an
unsaturated C45 isoprenoid chain, while PhyQ has a partially saturated C20 phytyl chain.
Because of their similarities in structure, it is likely that the PhyQ biosynthetic pathway is
similar to the menaquinone biosynthetic pathway in Escherichia coli (Johnson et al., 2000,
see also Chapter 2). The biosynthetic pathway of PhyQ is interesting for two reasons.
Firstly, the genes and enzymes of this pathway have not yet been completely characterized
in cyanobacteria. Secondly, knowledge of this pathway should enable one to devise
biological and chemical strategies to modify the quinone present in the A1 site.
A pathway of PhyQ biosynthesis in Synechocystis sp. PCC6803 was recently
proposed and included the genes menA, menB, menC, menD, menE, menF, and menG
(Johnson et al., 2000) (see Fig 2.1). Inactivation of the menA and menB genes, predicted to
encode 1,4-dihydroxy-2-naphthoic acid phytyltransferase and 1,4-dihydroxy-2-naphthoate
synthase, respectively, prevented the synthesis of PhyQ and led to the incorporation of
plastoquinone-9 (PQ-9) into the A1 site of PS I (Johnson et al., 2000). PQ-9 is the
secondary quinone acceptor in Photosystem II (PS II) and additionally functions as the
molecule that shuttles electrons to the cytochrome b6f complex. It was reported that PQ-9 is
present in the A1 site of PS I with the same orientation and asymmetric spin-density
distribution as PhyQ in wild type PS I complexes (Zybailov et al., 2000). PQ-9 is a

78 benzoquinone with a C45 alkyl tail and is structurally and functionally only distantly
related to PhyQ. The fact that PQ-9 could be inserted into the A1 site and could function in
lieu of PhyQ indicates that the binding site is not highly specific for PhyQ. Rather, the
redox properties of the quinone appear to be conferred largely by the protein environment
of the A1 site rather than by the identity of the quinone (Semenov et al., 2000).
The object of this study is the protein encoded by ORF sll1653, which was
originally annotated as gerC2, a spore germination protein in Bacillus subtilis. This ORF is
homologous to menG, which codes for a methyltransferase enzyme in E. coli and other
bacteria. In Synechocystis sp. PCC 6803, this gene is anticipated to encode a
2-phytyl-1,4-naphthoquinone (2-phytyl-1,4-NQ) methyltransferase enzyme (Johnson et
al., 2000). 2-phytyl-1,4-NQ differs from PhyQ only by the absence of the methyl group at
ring position C3 and by a higher redox potential (Depew and Wan, 1988). The relationship
between the molecular structure of this quinone and the axes of the molecular g-tensor is
depicted in Fig 3.1.
This study has two aims. The first is to confirm that sll1653 codes for the
methyltransferase responsible for synthesizing 2-phytyl-3-methyl-1,4-NQ from
2-phytyl-1,4-NQ. Indeed, it was found that inactivation of ORF sll1653 resulted in the
synthesis of PS I complexes containing 2-phytyl-1,4-NQ in the A1 site. The second is to
study the roles of the methyl group in quinone binding and electron transfer in PS I. Here, it
was shown that 2-phytyl-1,4-NQ is present in the A1 site with the same orientation and
asymmetric spin-density distribution as PhyQ in native PS I. It was also reported that

79 2-phytyl-1,4-NQ functions in forward electron transfer with some attendant alteration of
the PS I activity and electron-transfer kinetics.

80
RESULTS
Analysis of the Genotype of the menG Mutant Strain
As shown in Fig 3.2A, the menG gene (ORF sll1653) was inactivated by insertion
of a 1.3-kb DNA cassette, encoding the aphII gene and conferring resistance to kanamycin,
in the unique KpnI site within the coding sequence. Full segregation of the menG and
menG::aphII alleles was confirmed by PCR amplification and by Southern
blot-hybridization analysis of the genomic DNA. For PCR analysis, the primers flanking
the menG coding sequence (Fig 3.2A small arrows) were used to amplify the DNA
fragments from the wild-type and the menG mutant genomic DNAs. As expected, a 1.2-kb
fragment was amplified by PCR for wild type (Fig 3.2B). For the menG::aphII mutant
strain a 2.5-kb fragment was amplified by PCR; no DNA fragment with a size equal to that
expected from the wild-type menG allele was detected. This result indicates that the menG
mutant is homozygous for the menG::aphII gene interruption. To verify the menG
mutation further, the genomic DNA isolated from the wild-type and the menG mutant
strains were digested with restriction enzymes BglII and HindIII (see in Fig 3.2A) for the
Southern-blot hybridization analysis. Hybridization was performed using the PCR product
from the wild type genome as a probe. A single band with the size of 2.3 kb was observed
for the wild type strain (Fig 3.2C). For the menG mutant, two bands with the sizes of 2.7 kb
and 1.5 kb were observed due to the presence of the BglII site in the Kmr cartridge. All of
these results confirmed that the menG::aphII allele has fully segregated from the wild-type
gene.

81 High Performance Liquid Chromatography/Mass Spectrometry Analysis
To determine whether the menG gene is involved in PhyQ biosynthesis,
solvent-extracted quinone fractions from PS I complexes isolated from the wild-type and
the menG mutant strains were subject to orthogonal acceleration/time-of-flight mass
spectrometry. Pigments and quinones extracted from the isolated PS I trimers were
separated by reverse-phase HPLC, and the molecular mass of each eluate was determined
by mass spectrometry. In the wild-type complexes, PhyQ was detected by its characteristic
retention time, its UV absorption spectrum, and its m/z of 450 (Fig 3.3A). In PS I from the
menG mutant strain, no compound absorbing at 270 nm eluted at this retention time, nor
was a m/z of 450 detected. Rather, a UV-absorbing component eluted near the retention
time of Chl a that had a m/z of 436, which corresponds to the mass of 2-phytyl-1,4-NQ (Fig
3.3B).
Growth Rate under Photoautotrophic and Photomixotrophic Conditions
As indicated by HPLC and mass spectrometry, mutation of the menG gene
resulted in the replacement of PhyQ by 2-phytyl-1,4-NQ in PS I complexes. To measure
the effectiveness of 2-phytyl-1,4-NQ in supporting photosynthesis, growth rates of the
wild-type and menG mutant strains were compared. The results are summarized in Table
3.1. Growth of the wild-type and menG mutant strains were virtually the same under low
(50 µE m-2 s-1) and moderate (150 µE m-2 s-1) illumination conditions, in either the presence
or absence of glucose (Table 3.1). When cells were grown photoautotrophically, doubling
times of both the wild-type and the menG mutant strains were ca. 10 h under low light
intensity conditions and ca. 8 h under moderate light intensity conditions. When cells were

82 grown photomixotrophically, doubling times of the wild-type and the menG mutant strains
were both ca. 8 h under low light intensity conditions, and ca. 6 h under the moderate light
intensity conditions. The observed differences in the growth rates between the wild-type
and the menG mutant strains were all within the range of error. These results suggest that
under these illumination regimes, replacement of PhyQ with 2-phytyl-1,4-NQ has no
significant effect on growth.
PS II/PS I Ratio Measured by 77 K Fluorescence Emission Spectra
The relative ratios of PS II to PS I were compared by measuring 77 K fluorescence
emission spectra of wild-type and the menG mutant cells grown at various light intensities.
The results are summarized in Fig 3.4. When cells were grown under low-light conditions,
which is typical for Synechocystis sp. PCC 6803 (Fig 3.4A), the emission amplitude from
PS I at 721 nm was much greater than that from PS II at 695 nm. When cells were grown at
higher light intensities, an obvious decrease at 721 nm was found in the emission spectra
of the menG mutant cells when compared to wild-type cells (Fig 3.4B and 3.4C). In
wild-type cells the emission peak at 721 nm due to Chl a associated with PS I decreased
approximately 10% in cells grown at a light intensity of 300 µE m-2 s-1 relative to cells
grown at a light intensity of 50 µE m-2 s-1, while emission peaks at 685 and 695 nm due to
Chl a associated with PS II remained nearly constant. However, the peak amplitude at 721
nm decreased substantially in the menG mutant cells grown at higher light intensities. The
amplitude of this peak in cells grown at a light intensity of 300 µE m-2 s-1 was less than
70% that for cells grown at 50 µE m-2 s-1. The PS II/PS I ratio is slightly higher in the
menG mutant than in wild type when grown at 50 µE m-2 s-1 and significantly higher when

83
grown at 300 µE m-2 s-1. These results suggest that there may be either a defect in the
assembly of PS I complexes or an accelerated degradation of PS I complexes in the menG
mutant cells grown at high light intensities.
Chlorophyll Content of Whole Cells Grown at Various Light Intensities
The chlorophyll contents of wild-type and the menG mutant cells grown under
various light intensities were also determined (Table 3.2). When cells were grown under
low or moderate light intensities (50 or 150 µE m-2 s-1), the chlorophyll content of the
menG mutant cells was similar to that of the wild-type cells. However, when the light
intensity was increased to 300 µE m-2 s-1, the chlorophyll content in the wild-type cells
decreased to approximately 70% of the initial value (Table 3.2). A significantly lower
value, approximately 58% of the value obtained at lower light intensities, was observed for
menG mutant cells grown at a light intensity of 300 µE m-2 s-1. This value is only 80% that
of the wild-type strain under the same conditions, and this decrease can be correlated with
the loss of fluorescence emission from PS I observable in the decreased emission at 721 nm
in Fig 3.4. In combination, the results from Table 3.2 and Fig 3.4 suggest that content of PS
I complexes is lower in the menG mutant strain when cells were grown at high light
intensity. This difference could be due to a defect either in biogenesis or in stability of the
PS I complexes in the mutant strain.
Polypeptide Composition of Isolated PS I Trimers
To analyze whether the interruption of the menG gene has any effect on the PS I
subunit composition, PS I trimers were analyzed by SDS-polyacrylamide gel
electrophoresis. Polypeptide bands were visualized by silver staining. All of the eleven

84 polypeptides detected in the wild-type, PsaA though F and PsaI, J, L and M, were found to
be present in the menG mutant strain (Fig 3.5). No additional polypeptides were observed
in PS I complexes isolated from the menG mutant strain. These results indicate that the
replacement of PhyQ by 2-phytyl-1,4-NQ does not affect the subunit composition of the
PS I complexes in the menG mutant strain.
EPR spectroscopy of Q- and P700+ Q-
Fig 3.6A shows continuous wave EPR spectra measured at 34 GHz (Q-band) of
the photoaccumulated semiquinone radical (Q-) in PS I complexes isolated from the
wild-type (solid line, top) and the menG mutant strains (solid line, bottom). The PhyQ
anion radical is characterized (Fig 3.6A, solid line) by principal g-tensor values (Zybailov
et al., 2000) and the principal hyperfine A-tensor for the three protons in the methyl group
at the C2 position (Rigby et al., 1996) as summarized in Table 3.3. The hyperfine splitting
due to the remaining aromatic hydrogen atoms and the methylene group of the phytyl chain
at the C3 position remain unresolved and are included in the inhomogeneous line
broadening using linewidths of 3.0, 6.0, and 4.0 G for each of the three g-tensor
components (Fig 3.6B, dotted line). The deviations of the experimentally obtained
spectrum (Fig 3.6B, solid line) from the simulated spectrum (Fig 3.6A, dotted line) seen in
the mid- and high field regions are due to contamination by A0-.
The Q-band spectrum of the photoaccumulated semiquinone radical in the PS I
complexes isolated from menG mutant strain is shown in Fig 3.6B (solid line). The
spectrum has about the same overall width as that of wild type, which indicates that the
g-tensor of the semiquinone radical is about the same as that of the PhyQ anion radical. The

85 pattern of hyperfine splitting, however, is different in the menG mutant sample. The usual
four hyperfine lines are missing and instead, only two lines are present in the low-field
region. Such properties are expected if the semiquinone radical in the A1 site is replaced
with a 2-phytyl-1,4-NQ that lacks the methyl group at ring position 3 (see below). Indeed,
a satisfactory simulation of the spectrum of the menG mutant strain was obtained by
modifying the wild-type simulation, with the A-tensor of the methyl group replaced by that
of an aromatic C-H fragment with the same tensor components evaluated from the spin
polarized spectra described in the next section (Fig 3.6B, dotted line).
Transient EPR Spectroscopy
Fig 3.7 compares transient, spin-polarized EPR spectra (X-, Q-, W-band) of the
transient charge separated P700+ Q- state in the PS I complexes isolated from the wild-type
(dashed line) and the menG mutant strains (solid line). The two sets of spectra coincide
quite well in the high-field region, which is dominated by the P700+ contribution, but are
quite different in the low-field region. The W-band spectrum indicates a slightly larger
g-anisotropy for the menG mutant than that of the wild type PS I complex. The best fit
yields gxx = 2.00633 (menG) versus 2.00622 (wild type) while gyy = 2.00507 and gzz =
2.00218 remain unchanged from the wild type (Zech et al., 2000) as summarized in Table
3.3.
Closer inspection reveals that the main difference in the low field region concerns
the partially resolved hyperfine pattern. It is sufficiently resolved to permit an initial,
qualitative analysis directly from the spectra. For wild-type, the characteristic hyperfine
pattern due to the 1:3:3:1 quartet associated with a methyl group in ring position 2 is

86 particularly obvious in the Q-band spectrum. The fact that it is partially resolved results
from the increased spin density at the carbon ring position to which the methyl group is
attached (specific to the semiquinone radical ion in the A1 site). The components of the
axially symmetric hyperfine tensor have been determined by ENDOR to be A11 = 12.8
MHz and A⊥ = 9.0 MHz (see Rigby et al., 1996, for summary and further references; see
also Kamlowski et al., 1998; Zech et al., 2000). In the menG mutant, the CH3 group is
expected to be replaced by an aromatic C-H fragment. Indeed, the quartet hyperfine pattern
due to the CH3 group is missing in the menG mutant spectrum. Instead it is replaced by a
doublet (menG), the center position of which is shifted down field with respect to the
center of the quartet (wild type). Such a shift is expected with an altered orientation of the
dominant hyperfine principal axis with respect to the molecular axis frame. As concluded
from the observed shift direction, the dominant hyperfine axis will point more in the
direction of the gxx(Q) tensor axis in the menG mutant while it points more in the direction
of the gyy tensor axis in the wild type. Since the dominant hyperfine axis A11 of the CH3
hyperfine tensor is usually almost parallel to the C-CH3 bond axis, its direction is closer to
(about 30 degrees off) the gyy tensor axis. In contrast, the direction of the dominant
hyperfine axis of the aromatic C-H fragment (which is in-plane and perpendicular to the
C-H bond, see Carrington and MacLachlan, 1967) is closer to (about 30 degrees off) the
gxx(Q) tensor axis. The associated downfield shift of the center of the hyperfine doublet is
therefore in agreement with the experimental result.
To prepare for a quantitative simulation of the spectra, the following findings
were taken into account. The spin polarization pattern shows that 2-phytyl-1,4-NQ has the

87 same orientation in the A1 site as native PhyQ (Fig 3.7). The same P700 to A1
center-to-center distance has been verified by out-of-phase, spin-echo modulation
spectroscopy (data not shown). An independent determination of the hyperfine tensor
components of the C-H fragment has been obtained from pulsed ENDOR spectroscopy of
the same P700+ Q- state in the menG mutant (data not shown). The principal hyperfine
tensor components are given in Table 3.3. While the two large components, A11 and A22 are
clearly resolved, the third component is part of the line manifold in the center part of the
ENDOR spectrum and cannot be evaluated. With the reasonable assumption that the spin
density of the relevant carbon ring position does not change with respect to wild type
(corresponding to Aiso = -10.3 MHz with the appropriate McConnell relation) one can
estimate the third hyperfine principal component to be A33 = -3.6 MHz.
Although the axis associated with the largest hyperfine component is expected to
be in-plane and perpendicular to the C-H bond, this direction can be influenced by
significant spin densities on the neighboring molecular atoms, such as the spin density that
is known to reside on the carbonyl groups of the semiquinone radical. Therefore,
simulations of the Q-band spectrum (Fig 3.8) were carried out for various angles between
the largest principal axis A11 and the gxx (Q) axis (in-plane along the carbonyl bond
direction). The A11 axis is perpendicular to the C-H bond direction when this angle is 30°.
Indeed, of the simulations shown, the one for this value (Fig 3.8, bottom, long dashed
curve) comes closest to the experimental spectrum (Fig 3.8, top). A slightly smaller angle
than 30 degrees improves the simulation result to some degree. This indicates a weak
influence from spin density on neighboring atoms. The only significant spin density is

88 expected for the oxygen atom of the carbonyl group in ring position 1, as concluded from
the alternating spin density distribution due to asymmetric H-bonding to the opposite
carbonyl group in position 4 (see Kamlowski et al., 1998; Zech et al., 2000, and references
therein). However, the oxygen atom is already quite distant from the proton spin of the C-H
fragment, and thus the deviation of the C-H fragment hyperfine tensor from the (3:2:1)
ratio of the undisturbed C-H fragment is small, as indeed is observed here.
Kinetics of Forward Electron Transfer from A1- to FX
Forward electron transfer kinetics from the quinone to the iron sulfur clusters in
PS I complexes isolated from the wild-type and the menG mutant are compared in Fig 3.9
and Fig 3.10, which show spin-polarized EPR transients and decay-associated spectra,
respectively. The transients in Fig 3.9 labeled a, b and c were taken at the corresponding
field positions indicated with arrows in Fig 3.10. In wild-type PS I complexes, two
consecutive spectra are observed as electron transfer from A1- to FX occurs. At early times
(< ~1 µs) an emission/absorption/emission (EAE) pattern due to P700+ A1- is observed;
this pattern changes to an emissive spectrum due to P700+ in the state P700+ FeS- at later
times (> ~1 µs). The identity of the iron-sulfur cluster partner to P700+ in the state giving
the late spectrum requires some discussion. It has been shown that electron transfer
proceeds via FX (van der Est et al., 1994), which suggests the spectrum is due to P700+ FX-.
However, there is evidence (Leibl et al., 1995) that electron transfer from FX- to FA/FB is
faster than the transfer from A1 to FX. In this case, the late spectrum would be due to P700+
FA- and/or P700+ FB
-. Calculations (Kandrashkin et al., 1998) show the polarization
patterns corresponding to P700+ FX-, P700+ FA
-, and P700+ FB- show only minor

89 differences. Thus, the state giving the late spectrum P700+ FeS- is labeled P700+ FeS- and
the identity of the iron sulfur cluster(s) is left open. In the kinetic traces (Fig 3.9), the
electron transfer is seen most clearly at field position b. The positive (absorptive) signal at
early times (< ~50 ns) is due to P700+ Q-, while the negative (emissive) signal at late times
(< ~1 µs) is due to P700+ FeS-. The electron transfer dominates the decay of the early signal
while relaxation of the spin polarization causes the decay of the late signal. At positions a
and c the intensity of the late signal is weak and thus the transients decay primarily with the
electron transfer rate. As can be seen at all three field positions shown in Fig 3.10, the rate
of electron transfer is considerably slower in the mutant than in the wild type sample. For
the mutant PS I complexes, a fit of the entire data set as described in (van der Est, 1994)
yields an average electron transfer lifetime of 600 ± 100 ns. This compares with a value of
290 ± 70 ns for the wild-type complexes.
Kinetics of Charge Recombination Measured by Optical Spectroscopy
The reduction of P700+ was monitored at 811 nm after a brief laser flash in PS I
trimers isolated from the wild-type and the menG mutant strains (Fig 3.11). Because no
terminal electron acceptors such as ferredoxin or flavodoxin were present, the reduction
rate of P700+ represents the rate of charge recombination between [FA/FB]- to P700+. The
kinetics of P700+ reduction in the wild-type PS I complexes were fitted by one stretched
exponent with (1/e) lifetimes of 0.70, 30.7, and 94.7 ms (Fig 3.11A). The latter two values
are similar to the two lifetime components found previously for the P700-FA/FB complexes
from Synechocystis sp. PCC 6803 and are attributed to charge recombination between
[FA/FB]- and P700+ (Vassiliev et al., 1997). The first value is probably due to charge

90 recombination between P700+ and FX
- in damaged PS I complexes that lack an intact PsaC
subunit. The kinetics of P700+ reduction in PS I complexes from the menG mutant (Fig
3.11B) were also fitted by two exponentials with (1/e) lifetimes of 10.3 and 29.3 ms, These
values are attributed to charge recombination between [FA/FB]- and P700+ and are ca. 3
times faster than the wild-type rates. Even though the reason for the heterogeneity in P700+
reduction is not understood, it is noteworthy that the two kinetic phases attributed to charge
recombination between [FA/FB]- and P700+ are equally slowed in the menG mutant
samples.
Steady-State Rates of Flavodoxin Reduction
Steady-state rates of flavodoxin reduction as a function of light intensity were
measured for PS I complexes isolated form the wild-type and menG mutant strains to
assess the relative efficiencies of forward electron transfer. The rates at saturating light
intensity were determined by treating light as a substrate in a Michaelis-Menten kinetic
analysis (Fig 3.12). The maximal rate of flavodoxin reduction was found to be 1527 ± 247
µmol (mg Chl a)-1 h-1 for the wild-type PS I complexes and 1845 ± 399 µmol (mg Chl a)-1
h-1 for the menG mutant PS I complexes. Assuming that 100 Chl a molecules are present
per P700 in all PS I complexes, these maximal rates of electron transfer correspond to 37.9
± 6.2 e- PS I-1 s-1 in the wild-type PS I complexes and 45.8 ± 8.4 e- PS I-1 s-1 in the menG
mutant PS I complexes. Although the value obtained for the PS I complexes from the
menG mutant was slightly higher than that for the wild-type, this difference is within the
margin of error and therefore is not statistically significant (Fig 3.12). Hence, the
steady-state rate of electron transfer for the PS I complexes of the menG mutant, which

91 contain 2-phytyl-1,4-NQ, was virtually identical to the steady-state rate of electron transfer
for the PhyQ-containing PS I complexes of the wild-type.

92
DISCUSSION
The menG gene, encoding the 2-phytyl-1,4-NQ (demethylphylloquinone)
methyltransferase, was identified by targeted inactivation of ORF sll1653 in Synechocystis
sp. PCC 6803. This open reading frame was originally assigned as gerC2, which encodes
the spore germination protein C2 in Bacillus species and other spore-former bacteria
(Kaneko et al., 1996). In fact, the deduced amino acid sequence for ORF sll1653 shows
only 27% sequence identity to the menG gene product of E. coli. Targeted inactivation of
ORF sll1653 was accomplished by insertion of a kanamycin resistance cartridge in the
coding region, which was confirmed by PCR amplification of the region containing the
ORF and by Southern-blot hybridization analysis. The presence of 2-phytyl-1,4-NQ in PS I
complexes was confirmed by high performance liquid chromatography and mass
spectrometry, and its function in electron transfer was demonstrated by CW and transient
EPR spectroscopy. The orientation and distance of 2-phytyl-1,4-NQ relative to P700 was
confirmed to be the same as for PhyQ in wild-type PS I complexes by transient EPR,
out-of-phase, spin-echo modulation, and pulsed ENDOR spectroscopy. It can be safely
concluded that ORF sll1653 in Synechocystis sp. PCC 6803 is menG; that it encodes
2-phytyl-1,4-NQ methyltransferase, the enzyme required for the last step of the PhyQ
biosynthetic pathway; and that 2-phytyl-1,4-NQ functionally replaces PhyQ at the A1 site
in PS I complexes.
Despite the absence of PhyQ and the presence of 2-phytyl-1,4-NQ in PS I
complexes, the growth rates of the wild-type and menG mutant cells were identical. As
observed for many other cyanobacteria (Fujita, 1997), the ratio of PS II to PS I is known to

93 change in Synechocystis sp. PCC 6803 as a function of the light intensity during cell
growth (G. Shen, J. H. Golbeck, and D. A. Bryant, unpublished observation). An
interesting observation was that the change in the ratio of PS II to PS I was more
pronounced in the menG mutant strain than in the wild type as the light intensity was
increased from 50 µE m-2 s-1 to 300 µE m-2 s-1. In wild-type cells, the PS II to PS I ratio
increased by a factor of 1.1, while in the menG mutant cells the PS II to PS I ratio increased
by a factor of 2 as estimated from the amplitudes of the fluorescence emission spectra at 77
K. It is noteworthy that the overall chlorophyll content in the wild-type and menG mutant
cells were similar at light intensities of 50 and 150 µE m-2 s-1 and differed only at 300 µE
m-2 s-1. Therefore, the difference in the PS II to PS I ratio between wild type and the menG
mutant strain can be primarily attributed to a change in the content of PS I, i.e. the number
of PS I complexes declines in the menG mutants to a greater extent than in the wild-type
cells as the light intensity is raised. By way of comparison, the menA and menB mutant
strains, which contain plastoquinone-9 in the A1 site, have been observed to grow
significantly more slowly and can be grown only under very low light intensity conditions
(< 20 µE m-2 s-1). The ratio of PS II to PS I is already greater than 2:1 at low light intensity,
and might be expected to be even greater as the light intensity is increased (the cells,
however, do not survive). Thus, it appears that Synechocystis sp. PCC 6803 must exceed
ca. 2:1 ratio of PS II to PS I before light becomes toxic to the growing cells.
To understand further the behavior of 2-phytyl-1,4-NQ in electron transfer, one
must consider the redox potential of this quinone in the A1 site. In aqueous solution, the
E(Q/Q-)7 of 2-methyl-1,4-NQ is –203 mV vs. NHE (normal hydrogen electrode) and the

94 E(Q/Q-)7 of 2,3-dimethyl-1,4-NQ is –240 mV vs. NHE (Swallow, 1982). The E(Q/Q-)7 of
2-phytyl-3-methyl-1,4-NQ (PhyQ) in aqueous solution (containing 5 M 2-propanol and
2M acetone) is reported to be –170 mV. The presence of the second alkyl group, ortho to
the first, therefore lowers the redox potential of the quinone by ca. 37 mV. However, the A1
site is highly hydrophobic (Iwaki and Itoh, 1991, 1994), and reduction potentials of
quinones in organic solvent are probably more relevant to their properties in PS I
complexes. In dimethylformamide (DMF), the E1/2 value for 2-methyl-1,4-NQ has been
reported as –650 mV vs. SCE and the E1/2 value for 2,3-dimethyl-1,4-NQ has been reported
to be –746 mV (Prince et al., 1983) vs. SCE (standard calomel electrode). The lower redox
potential for the dimethyl quinone is logical chemically; a methyl group is an electron
donor and should therefore destabilize the semiquinone radical in either aqueous solution
or in organic solvent. A prenyl substituent on the quinone ring is not as effective as a
second alkyl group at electron donation and lowers the E1/2 value by only 50 to 60 mV
(Prince et al., 1983). For example the E1/2 of menaquinone-2
(2-all-trans-polyprenyl-3-methyl-1,4-NQ) is –746 mV vs. SCE (to the best of our
knowledge, the reduction potential of 2-phytyl-1,4-NQ in DMF is not available). Using the
redox potentials for 2-methyl-1,4-NQ and 2,3-dimethyl-1,4-NQ as models for
2-phytyl-1,4-NQ and PhyQ (both differ by the addition of one methyl group), the addition
of a second methyl group ortho to the first should lower the redox potential by ca. 96 mV.
If one further employs the concept of an ‘acceptor number’ that was utilized for PQ-9 in the
menA and menB mutants (Semenov et al., 2000), the redox potential of 2-phytyl-1,4-NQ in
the A1 site would be 61 mV more oxidizing than PhyQ. An acceptor number corrects the

95 E1/2 value for the degree of the nucleophilic (donor) or electrophilic (acceptor) properties of
the solvent (Gutmann, 1976) and was employed initially by Itoh (Iwaki and Itoh., 1994) to
estimate the redox potential of PhyQ in the A1 site of PS I. It is a dimensionless number that
expresses the acceptor properties of a solvent relative to SbCl5.
The estimated redox potential of 2-phytyl-1,4-NQ in the A1 site can now be
compared to estimates based upon the experimentally measured rate of electron transfer.
Forward electron transfer is ca. 3-times slower when 2-phytyl-1,4-NQ rather than PhyQ
occupies the A1 site. According to the Moser-Dutton formulation (Moser et al., 1992), the
rate of electron transfer in proteins depends on the Gibbs free energy between the
donor-acceptor pair, the edge-to-edge distance between the donor-acceptor pair, and the
reorganization energy. The distance between the 2-phytyl-1,4-NQ anion radical and P700+
was experimentally determined here to be the same as the center-to-center distance
between the PhyQ anion radical and P700+. The orientation of 2-phytyl-1,4-NQ relative to
the membrane plane was also shown to be identical to that of PhyQ. Hence, the
edge-to-edge distance between 2-phytyl-1,4-NQ and FX is likely to be the same as the
distance between PhyQ and FX. It seems reasonable to assume similar reorganization
energies when 2-phytyl-1,4-NQ and PhyQ occupy the A1 site, given that the only
difference between the two quinones is the presence or absence of a single methyl group.
Using the values of 11.3 Å for the edge-to-edge distance between the quinone and FX
(Klukas et al., 1999), and a value of 0.7 eV for the reorganization energy, a (1/e) lifetime of
ca. 290 ns for Q- in the wild-type translates to an 81 mV difference in Gibbs free energy
between Q-/Q and FX-/FX. Retaining the same values for the distance and reorganization

96 energy, a (1/e) lifetime of 600 ns for Q- in the menG mutant translates to a 28 mV
difference in the Gibbs free energy between Q-/Q and FX-/FX. According to this analysis,
the addition of methyl group to 2-phytyl-1,4-NQ lowers the redox potential of the quinone
in the A1 site by 53 mV. This value is only a rough estimate, especially given the
uncertainties in the edge-to-edge distance between the cofactors and given the assumed
(unchanged) reorganization energy of the site. Nevertheless, this value agrees quite well
with the difference in redox potentials of 2-methyl-1,4-NQ and 2,3-dimethyl-1,4-NQ in
organic solvents, especially after correction for the site’s “acceptor number”. The
replacement of PhyQ by 2-phytyl-1,4-NQ would therefore lower the equilibrium constant
between Q-/Q and FX-/FX from 27 to 3, a factor of ca. 10. Estimating the absolute redox
potential of 2-phytyl-1,4-NQ in the A1 site is somewhat more difficult because the redox
potential of A1 is not known with certainty. Three values have been published for the redox
potentials of the Q-/Q couple [-810 mV vs. NHE (Vos and Gorkom, 1990), -800 mV vs.
NHE (Chamrovoskii and Cammack, 1983) and -754 mV vs. NHE (Iwaki and Itoh, 1994)].
If one uses the lower redox potentials, the replacement of PhyQ by 2-phytyl-1,4-NQ would
therefore raise the redox potential of the Q-/Q couple to ca. –750 mV, a value that is
probably still lower than the redox potential of the FX-/FX couple.
The kinetics of reduction of P700+ after a single flash represents dissipative
charge recombination between P700+ and terminal electron acceptors [FA/FB]- in the
absence of an acceptor such as ferredoxin or flavodoxin. It is not yet clear whether electron
transfer occurs directly from [FA/FB]- to P700+, or whether the electron travels backward

97 by a thermally activated, uphill electron transfer through the quinone. The latter
presupposes that each redox pair (Q-/Q ↔ FX-/FX; FX
-/FX ↔ FB-/FB; FB
-/FB ↔ FA-/FA) is
described by an equilibrium constant that can be determined from the midpoint potentials
of the acceptors. The large distance between P700 and the terminal iron-sulfur clusters [46
Å center-to-center distance from P700 to FA (Schubert et al., 1998)] argues against the
direct recombination mechanism, although experimental data to support thermally
activated uphill electron transfer have been lacking. If the charge recombination between
[FA/FB]- and P700+ were to occur through backward electron transfer, then a change in the
equilibrium constant between Q-/Q and FX-/FX should logically affect the rate. The
equilibrium constant between Q-/Q and FX-/FX would be correspondingly higher as the
redox potential of Q-/Q were driven more negative, thereby resulting in a depopulation of
reduced quinone. Conversely, the equilibrium constant between Q-/Q and FX-/FX would be
correspondingly lower as the redox potential of Q-/Q were driven more positive, thereby
resulting in a repopulation of reduced quinone. Consequently, the back-reaction kinetics in
the mutant would be faster than in wild-type PS I complexes.
The latter description is precisely the behavior observed. The forward
electron-transfer kinetics from Q- to FX becomes correspondingly slower as PhyQ [1/e
lifetime of 200 ns (van der Est et al., 1994)], 2-phytyl-1,4-NQ (1/e lifetime of 600 ns), and
PQ-9 [1/e lifetime of 15 µs (Semenov et al., 2000)] occupy the A1 site. Conversely, the
charge recombination kinetics between [FA/FB]- and P700+ become more rapid as PhyQ
[1/e lifetime of 80ms (Vassiliev et al., 1997)], 2-phytyl-1,4-NQ (1/e lifetime of 20 ms) and
PQ-9 [1/e lifetime of 3 ms (Semenov et al., 2000)] occupy the A1 site. The trend among the

98 wild type, menA or menB, and menG mutants therefore strongly supports the backward,
through-carrier route of electron transfer for charge recombination. Since the redox
potential of the quinone in the A1 site influences the rate of charge recombination from the
terminal electron acceptors to P700+ as well as the forward electron transfer between the
quinone and the terminal iron-sulfur clusters, this suggests that charge recombination
occurs by thermally activated, uphill electron transfer through the secondary quinone
acceptor in PS I.
Finally, although the kinetics of electron transfer within PS I are altered when
PhyQ is replaced by 2-phytyl-1,4-NQ, these changes do not affect the growth rate of the
organism. An explanation for this apparent discrepancy is that the slower rate of forward
electron transfer from A1- to FX is still not the rate-limiting step in the overall electron
transfer reaction to flavodoxin (or probably to ferredoxin). Indeed, the maximal rate of
flavodoxin reduction of the PS I complexes isolated from the menG mutant was virtually
identical to that observed for wild-type PS I complexes. Analysis of the rate of electron
donation from PS I to flavodoxin is complicated by the two-electron reduction using this
acceptor. Since the potentials at pH 7 for the oxidized flavodoxin/flavodoxin semiquinone
and the flavodoxin semiquinone/hydroquinone couple are –212 and –436 mV (Schubert et
al., 1998), respectively, it is likely that only the latter couple is physiologically relevant.
Studies show that the reduction of flavodoxin semiquinone is biphasic; the fast, first-order
phase corresponds to electron transfer within a preformed complex, but the slower phase is
concentration-dependent, with a second-order rate constant of 1.7 x 108 M-1 s-1. Given the
nearly wild-type electron-transfer rates from cytochrome c6 to flavodoxin in the menG

99 mutant, it is clear that forward electron transfer continues to out-compete the faster back
reaction rate in the mutant. Hence, PS I complexes are remarkably robust and are tolerant
to changes caused by replacement of wild-type components with “suboptimal” ones. This
robustness still allows for a remarkable level of functionality of the bound electron-transfer
cofactors under suboptimal conditions.

100
SUMMARY
The menG gene, encoding the 2-phytyl-1,4-NQ (demethylphylloquinone)
methyltransferase, was identified in Synechocystis sp. PCC 6803. The PS I complexes
isolated from the menG mutant accumulated 2-phytyl-1,4-NQ, which functioned as the A1
cofactor in the electron transfer. The rate of forward electron transfer from A1 to the
iron-sulfur clusters was slowed by a factor of two, while the rates of the P700+ [FA/FB]-
backreaction increased by a factor of 3 to 4. These results were explained by the lowering
of the equilibrium constant between Q-/Q and FX-/FX by a factor of ~10. Despite the defect
in the forward electron transfer, the PS I complexes containing 2-phytyl-1,4-NQ showed
catalytic activity similar to those of PS I complexes containing phylloquinone, which
demonstrates that PS I complexes are remarkably robust and can function with presumably
“suboptimal” electron transfer cofactors.

101
MATERIALS AND METHODS
Generation of the menG Mutant Strain of Synechocystis sp. PCC 6803
To generate the construct for inactivation of the menG gene, a 1.2-kb DNA
fragment containing the sll1653 ORF was amplified through PCR from the genome of the
Synechocystis sp. PCC 6803. For cloning convenience, a new EcoRI site was created in the
3'-end primer with one nucleotide change. The PCR-amplified DNA fragment was cloned
into pUC19 vector and confirmed by sequencing. To inactivate the menG gene, a 1.3-kb
kanamycin-resistance cartridge encoded by the aphII gene was inserted into the KpnI site
in the middle of the menG gene. The resulting plasmid was linearized by digestion with
EcoRI and was used to transform the Synechocystis sp. PCC 6803 wild-type cells. The
transformation and the transformant screening were carried out as described (Shen et al.,
1993). After several rounds of restreaking to single colonies, full segregation of the
menG::aphII from the menG alleles was verified by PCR and Southern blot-hybridization
analyses.
Growth of Synechocystis sp. PCC 6803
Wild-type Synechocystis sp. PCC 6803 was grown in medium B-HEPES. This
medium is prepared by supplementing BG-11 medium (Stainer et al., 1971) with 4.6 mM
of HEPES [4-(2-hydroxyethyl)piperazine-N’-(2-ethnesulfonic acid)]-KOH and 18 mg L-1
ferric ammonium citrate. The menG mutant cells were grown in medium B-HEPES
supplemented with 50 µg mL-1 kanamycin. The growth temperature was maintained at 32o
C and the light intensity was adjusted to 50, 150, and 300 µE m-2 s-1 by adding fluorescent

102 lamps or by shielding with paper as required. For photomixotrophic growth, the medium
was supplemented with 5 mM glucose. Growth of the cells was monitored by measuring
the absorbance at 730 nm using a Cary-14 spectrophotometer that had been modified for
computerized data acquisition by On-Line Instruments, Inc (Bogart, GA). Cells from a
liquid starter culture in the exponential phase (OD730nm < 0.7) were adjusted to the same
initial cell density (OD730nm = 0.05) for growth-curve measurements. The liquid cultures
were bubbled with 1% (v/v) CO2 balanced with air.
DNA Isolation, PCR, and Southern Blotting
Chromosomal DNA from Synechocystis sp. PCC 6803 wild-type and mutant
strains was isolated as previously described (Shen, et al., 1993). PCR primers used to
amplify the menG DNA fragment for evaluation of the menG alleles were positioned as
shown in the Fig 3.2. The sizes of the PCR products from the wild type and menG mutant
strains were determined by agarose gel electrophoresis. For Southern blot-hybridization
analysis, the chromosomal DNAs were subjected to restriction enzyme digestion, agarose
gel electrophoresis, and capillary transfer to nitrocellulose membranes. Hybridization
probes were generated by labeling the wild-type menG PCR fragment with the
random-primed DNA labeling kit (Boehringer-Mannheim, Indianapolis, IN).
Hybridization conditions and detection were as previously described (Shen and Bryant
1995)

103 Chlorophyll Extraction and Analysis
Chlorophyll was extracted from whole cells, thylakoid membranes and PS I
particles with 100% methanol, and its concentration was determined
spectrophotometrically as described (MacKinney, 1941).
Isolation of Thylakoid Membranes and PS I Particles
Thylakoid membranes were isolated from cells in the late exponential growth
phase as described previously (Shen et al., 1993). Cells were broken at 4°C by three
passages through a French pressure cell at 124 MPa. The thylakoid membranes were
resuspended and solubilized for 2 h at 4°C in the presence of 1% (w/v)
n-dodecyl-β-D-maltoside. PS I complexes were separated from other membrane
components by centrifugation on 5-20% (w/v) sucrose gradients with 0.05%
n-dodecyl-β-D-maltoside in the buffer. Further purification was achieved by a second
centrifugation on sucrose gradients in the buffer in the absence of
n-dodecyl-β-D-maltoside. Trimeric PS I complexes were used in these studies (Golbeck,
1995).
SDS-Polyacrylamide Gel Electrophoresis Analysis
Methods used for SDS-polyacrylamide gel electrophoresis were identical to those
previously described (Shen and Bryant, 1995). A Tricine/Tris discontinuous buffer system
was used to resolve the polypeptide composition of the PS I complexes prepared from the
wild type and the menG mutant strains of Synechocystis sp. PCC 6803 (Schägger and van

104 Jagow, 1987) The separating gel contained 16% acrylamide and 6 M urea, and the resolved
proteins were visualized by silver staining (Blum et al., 1987)
Analysis of Quinones by High Performance Liquid Chromatography and Mass Spectrometry
For quinone extraction, PS I particles isolated from the wild-type and the menG
mutant strains were exchanged into distilled water by dialysis for 4 h and lyophilized. The
pigments were extracted from the lyophilized samples with acetone:methanol, 1:1 (v/v) at
4 °C, vacuum dried at room temperature in the dark, and resuspended in 100% methanol.
The extracts were separated by reverse-phase high performance liquid chromatography
(HPLC) using a C18 column as previously described (Johnson et al., 2000). Eluates were
analyzed using atmospheric-pressure chemical ionization with a Micromass Quattro II
mass spectrometer operated in the negative-ion mode.
Flavodoxin Photoreduction
Steady-state rates of flavodoxin reduction were measured for the PS I trimers
isolated from the wild-type and the menG mutant strains as previously described (Jung et
al., 1995). Cyanobacterial flavodoxin was overexpressed in E. coli and purified as
described (Fillat et al., 1991). PS I trimers were resuspended to a final concentration of 5
µg Chl mL-1 in 25 mM Tris-HCl buffer, pH 8.3, containing 50 mM MgCl2, 15 µM
cytochrome c6, 15 µM flavodoxin, 6.0 mM sodium ascorbate, and 0.05 %
n-dodecyl-β-D-maltoside. Measurements were made by monitoring the rate of change in
the absorption at 467 nm using a modified Cary 219 spectrophotometer fitted with a 465
nm interference filter on the surface of the photomultiplier. The 4-sided clear cuvette was

105 illuminated from two sides using high intensity, red light-emitting diodes (LS1, Hansatech
Ltd., Norfolk, UK). Initial rates of the reaction were recorded under various light
intensities and plotted as a function of the light intensity to assess the relative efficiency of
forward electron transfer to flavodoxin.
77 K Fluorescence Emission Spectra
Fluorescence emission spectra were measured at 77 K using an SLM 8000C
spectrofluorometer as previously described (Shen and Bryant, 1995). Cells from the
exponential phase of growth were harvested and resuspended in 25 mM HEPES/NaOH,
pH 7.0, containing 60% glycerol. Samples were adjusted to equal cell density (OD730nm =
1.0) prior to freezing in liquid nitrogen. The excitation wavelength was 440 nm, the
excitation slit width was 4 nm, and the emission slit width was 2 nm. Each spectrum shown
is the average of four spectra.
Q-band EPR of Photoaccumulated PS I Complexes
Photoaccumulation experiments were performed using a Bruker ER300E
spectrometer and an ER 5106-QT resonator equipped with an opening for in-cavity
illumination similar to that described in (Yang et al., 1998). Low temperatures were
maintained with an ER4118CV liquid nitrogen cryostat and an ER4121 temperature
controller. The microwave frequency was measured with a Hewlett-Packard 5352B
frequency counter and the magnetic field was measured with a Bruker ER035M NMR
gaussmeter. The pH of the sample was adjusted to 10.0 with 1.0 M glycine buffer, and
sodium dithionite was added to a final concentration of 50 µM. After incubation for 10 min
in the dark, the sample was placed into the resonator and the temperature was adjusted to

106 205 K. The sample was illuminated for 40 min with a 20 mW He-Ne laser operating at 630
nm. A dark background spectrum was subtracted from the photoaccumulated spectrum.
EPR spectral simulations were carried out on a 466 MHz Power Macintosh G3 computer
using a Windows 3.1 emulator (SoftWindows 3.0, Insignia Solutions, UK) and SimFonia
software (Bruker Analytik GMBH, Germany).
Time-Resolved EPR Spectroscopy at Multiple Frequencies
Transient EPR spectra as well as pulsed EPR/ENDOR data were obtained using
the same instrumentation described earlier (Zybailov et al., 2000; Bittl et al., 1997).
Flash-Induced Absorbance Changes at 811 nm
The transient absorbance changes were measured with a home-built, double-beam
spectrophotometer described in (Vassiliev et al., 1997). The sample was prepared at a
chlorophyll concentration of 7 µM in 50 mM Tris buffer, pH 8.3. Measurements were
made at pH 8.3 in 50 mM Tris buffer in the presence of 2 mM sodium ascorbate and 5 µM
2,6-dichlorophenolindophenol. The samples were measured in a quartz cuvette with a
1-cm path for the measuring light. The excitation beam was provided by a
frequency-doubled Nd-YAG laser (532 nm) with the flash energy attenuated to ca. 20 mJ
using the Q-switch delay and neutral density filters. The measuring beam was provided by
a 5 mW semiconductor laser operating at 811 nm. Kinetic analysis was performed using a
non-linear regression algorithm in Igor Pro (WaveMetrics Inc., Lake Oswego, OR)
(Vassiliev et al., 1997). All kinetic constants are reported as 1/e lifetimes (τ).

107 X-band Transient EPR Spectroscopy at Room Temperature
Room-temperature, transient EPR experiments were carried out using a modified
Bruker ESP 200 equipped with a home-built broad-band amplifier (bandwidth >500 MHz)
for direct detection experiments. Excitation was provided by a Continuum YAG/OPO laser
system operating at 680 nm and 1 Hz. The EPR signals were digitized using a LeCroy
LT322 500 MHz digital oscilloscope and transferred to a PC for storage and analysis. The
samples were measured using a flat cell and a Bruker rectangular resonator fitted with a
piece of rough-surfaced glass in front of the window to provide optimal illumination. The
response time of the system in direct detection mode is limited by the bandwidth of the
resonator and is estimated as ~50 ns, and the decay of the spin polarization limits the
accessible time range to times shorter than a few microseconds. The same set-up can also
be used with field modulation and lock-in detection. In this mode the response time is ~50
µs but the sensitivity is much higher, and the charge recombination can be monitored in the
millisecond time range. In both modes of operation, full time/field data sets are collected
and analyzed to determine the lifetimes of the species and their decay-associated spectra as
described in detail (van der Est et al., 1994; Semenov et al., 2000).

108
REFERENCES
Biggins, J, Mathis P (1988) Functional role of vitamin K in Photosystem I of the
cyanobacterium Synechocystis 6803. Biochemistry 27: 1494-500
Bittl, R, Zech SG, Fromme P, Witt HT, Lubitz W (1997) Pulsed EPR structure analysis of
Photosystem I single crystals: localization of the phylloquinone acceptor. Biochemistry
36: 12001-12004
Bock CH, Van der Est AJ, Brettel K, Stehlik, D (1989) Nanosecond electron transfer
kinetics in Photosystem I as obtained from transient EPR at room temperature. FEBS Lett.
247: 91-96
Brettel K (1997) Electron transfer and the arrangements of the redox cofactor in
Photosystem I. Biochim Biophys Acta 1318: 322-373
Carrington A, MacLachlan AD (1967) Introduction to Magnetic Resonance with
Applications to Chemistry and Chemical Physics, Hamper and Row, New York
Depew MC, Wan JKS (1988) in Patai S, Rappoport (eds), The Chemistry of the Quinonoid
Compounds. Vol. 2, pp. 963-1018, John Wiley and Sons, New York
Fillat M, Borrias W, Weisbeek P (1991) Isolation and overexpression in Escherichia coli
of the flavodoxin gene from Anabaena PCC 7119. Biochem J 280: 187-191

109 Golbeck, J. H. (1995) In Song PS (ed), in CRC Handbook of Organic Photochemistry and
Photobiology. Vol. 1, pp 1407-1419. CRC Press, Boca Raton, FL
Gutmann V (1976) Solvent effects on the reactivities of organometallic compounds.
Coord. Chem. Rev 18: 225-255
Iwaki, M., and Itoh, S. (1991) Structure of the phylloquinone-binding (QΦ) site in green
plant Photosystem I reaction centers: the affinity of quinones and quinonoid compounds
for the QΦ site. Biochemistry 30: 5347-5352
Iwaki, M, Itoh S (1994) Reaction of reconstituted acceptor quinone and dynamic
equilibration of electron transfer in the photosystem I reaction center. Plant Cell Physiol
35: 983-993
Johnson TW, Shen G, Zybailov B, Folling D, Reategui R, Beauparlant S, Vassiliev IR,
Bryant DA, Jones AD, Golbeck JH, Chitnis PR (2000) Recruitment of a foreign quinone
into the A1 site of Photosystem I: I. Genetic and physiological characterization of
phylloquinone biosynthetic pathway mutants in Synechocystis sp. PCC 6803. J Biol Chem
275: 8523-8530
Johnson TW, Zybailov B, Jones AD, Bittl R, Zech S, Stehlik D, Golbeck JH, Chitnis PR
(2001) Recruitment of a foreign quinone into the A1 site of Photosystem I: in vivo
replacement of plastoquinone-9 by media-supplemented naphthoquinones in
phylloquinone biosynthetic pathway mutants of Synechocystis sp. PCC 6803. J Biol Chem
276: 39512-39521

110 Jung, YS, Yu L, Golbeck JH (1995) Reconstitution of iron-sulfur center FB results in
complete restoration of NADP+ photoreduction in Hg-treated Photosystem I complexes
from Synechococcus sp. PCC 6301. Photosynth Res 46: 249-255
Kamlowski A, Altenberg-Greulich B, van der Est A, Zech SG, Bittl R, Fromme P, Lubitz
W, and Stehlik D (1998) The quinone acceptor A1 in Photosystem I: binding site, and
comparison to QA in purple bacteria reaction centers. J Phys Chem B 102: 8278-8287
Kandrashkin Y, Salikhov K, van der Est A, Stehlik D (1998) Electron spin polarization in
consecutive spin-correlated radical pairs: Application to short-lived and long-lived
precursors in type 1 photosynthetic reaction centers. Appl Magn Res 15: 417-447
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y, Miyajima N, Hirosawa
M, Sugiura M, Sasamoto S, Kimura T, Hosouchi T, Matsuno A, Muraki A, Nakazaki N,
Naruo K, Okumura S, Shimpo S, Takeuchi C, Wada T, Watanabe A, Yamada M, Yasuda
M, Tabata S (1996) Sequence analysis of the genome of the unicellular cyanobacterium
Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and
assignment of potential protein-coding regions. DNA Res 3: 109-136
Klukas O, Schubert WD, Jordan P, Krauss N, Fromme P, Witt HT, Saenger W (1999)
Localization of two phylloquinones, QK and QK', in an improved electron density map of
Photosystem I at 4-Å resolution. J Biol Chem 274: 7361-7367
Leibl W, Toupance B, and Breton J (1995) Photoelectric characterization of forward
electron transfer to iron-sulfur centers in Photosystem I. Biochemistry 34: 10237-10244

111 MacKinney G (1941) Absorption of light by chlorophyll solutions. J Biol Chem 140:
315-322
Malkin, R. (1986) On the function of two vitamin K1 molecules in the PS I electron
acceptor complex. FEBS Lett 208: 343-346
Moser CC, Keske JM, Warncke K, Farid RS, Dutton PL (1992) Nature of biological
electron transfer. Nature 355: 796-802
Prince RC, Dutton PL, Bruce JM (1983) Electrochemistry of ubiquinones : Menaquinones
and plastoquinones in aprotic solvents. FEBS Lett 160: 273-276
Rigby SEJ, Evans MCW, Heathcote P (1996) ENDOR and special triple resonance
spectroscopy of A1 of Photosystem 1. Biochemistry 35: 6651-6656
Schägger H, van Jagow G (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal
Biochem 166: 368-379
Schoeder HU, Lockau W (1986) Phylloquinone copurifies with the large subunit of
Photosystem I. FEBS Lett 199: 23-27
Semenov AY, Vassiliev IR, van der Est A, Mamedov MD, Zybailov B, Shen G, Stehlik D,
Diner BA, Chitnis PR, Golbeck JH (2000) Recruitment of a foreign quinone into the A1
site of Photosystem I: Altered kinetics of electron transfer in phylloquinone biosynthetic

112 pathway mutants studied by time-resolved optical, EPR, and electromagnetic techniques. J
Biol Chem 275: 23429-23438
Shen GZ, Bryant DA (1995) Characterization of a Synechococcus sp. starin PCC 7002
mutant lacking Photosystem I. Protein assembly and energy distribution in the absence of
the Photosystem I reaction center core complex. Photosynth Res 44: 41-53
Shen GZ, Boussiba S, Vermaas WFJ (1993) Synechocystis sp PCC 6803 strain lacking
Photosystem I and phycobilisome function. Plant Cell 5: 1853-1863
Swallow A J (1982) Physical Chemistry of semiquinones. In: Trumpower BL (ed),
Functions of Quinones in Energy Conserving Systems, pp. 59-72, Academic Press, New
York
Van der Est A, Bock C, Golbeck J, Brettel K, Sétif P, Stehlik D (1994) Electron transfer
from the acceptor A1 to the iron-sulfur centers in Photosystem I as studied by transient EPR
spectroscopy. Biochemistry 33: 11789-11797
Vassiliev IR, Jung YS, Mamedov MD, Semenov AY, Golbeck JH (1997) Near-IR
absorbance changes and electrogenic reactions in the microsecond-to-second time domain
in Photosystem I. Biophys J 72: 301-315
Vos MH, Van Gorkom HJ (1990) Thermodynamic and structural information on
photosynthetic systems obtained from electroluminescence kinetics. Biophys J 58:
1547-55

113 Yang F, Shen G, Schluchter WM, Zybailov BL, Ganago AO, Vassiliev IR, Bryant DA,
Golbeck JH (1998) Deletion of the PsaF polyptptide modifies the environment of the
redox-active phylloquinone (A1). Evidence of unidirectionality of electron transfer in
Photosystem I. J Phys Chem B (1998) 102: 8288-8299
Zech SG, Hofbauer W, Kamlowski A, Fromme P, Stehlik D, Lubitz W, Bittl R (2000) A
structural model for the charge separated state P700+A1
- in Photosystem I from the
orientation of the magnetic interaction. J Phys Chem B 104: 9728-9739
Zybailov B, van der Est A, Zech SG, Teutloff C, Johnson TW, Shen G, Bittle R, Stehlik D,
Chitnis PR, Golbeck JH (2000) Recruitment of a foreign quinone into the A1 site of
Photosystem I: II. Structural and functional characterization of phylloquinone biosynthetic
pathway mutants by electron paramagnetic resonance and electron-nuclear double
resonance spectroscopy. J Biol Chem 275: 8531-8539
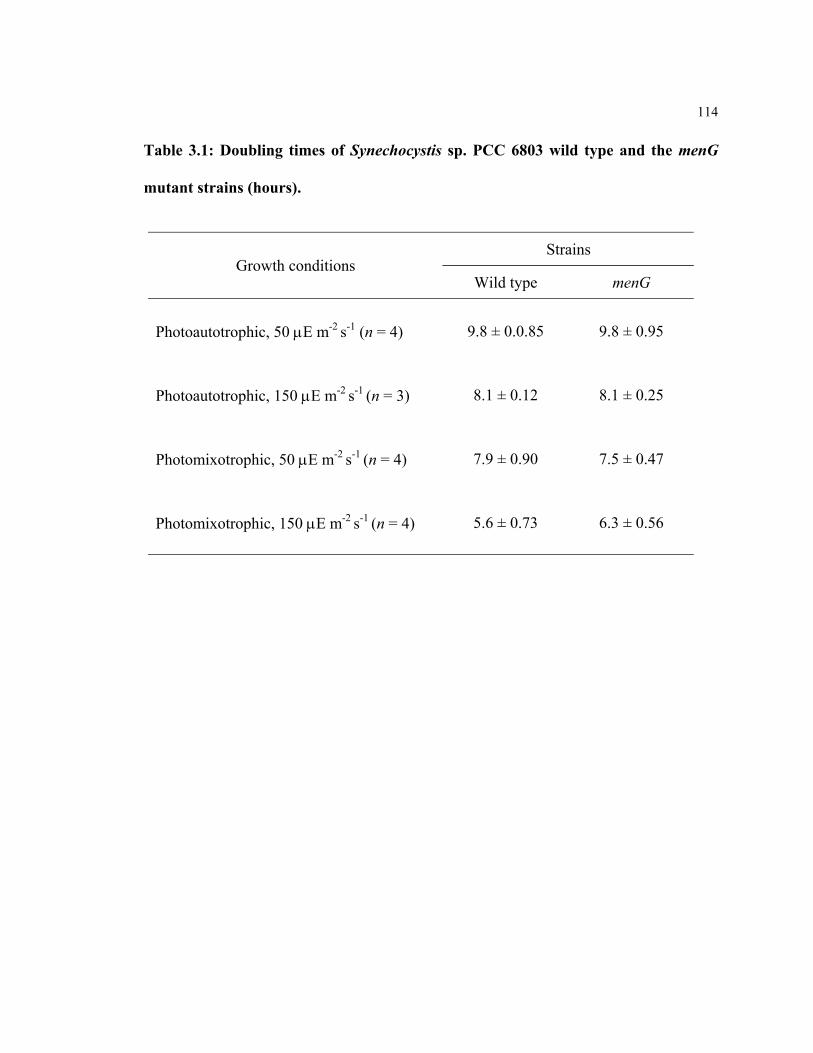
114
Table 3.1: Doubling times of Synechocystis sp. PCC 6803 wild type and the menG
mutant strains (hours).
Strains Growth conditions
Wild type menG
Photoautotrophic, 50 µE m-2 s-1 (n = 4) 9.8 ± 0.0.85 9.8 ± 0.95
Photoautotrophic, 150 µE m-2 s-1 (n = 3) 8.1 ± 0.12 8.1 ± 0.25
Photomixotrophic, 50 µE m-2 s-1 (n = 4) 7.9 ± 0.90 7.5 ± 0.47
Photomixotrophic, 150 µE m-2 s-1 (n = 4) 5.6 ± 0.73 6.3 ± 0.56

115
Table 3.2: Chlorophyll content in cells grown photoautotrophically at 32° C under
various illumination conditions.
Chlorophylls were extracted from whole cells with methanol. The absorbance coefficient
of 82 (µg/ml) –1 at 666 nm was used to calculate the chlorophyll content in 1 ml of the
cultures with an optical density of 1.0 at 730 nm.
µg Chlorophyll / OD730nm (n = 3) Light intensity
(µE m-2 s-1) Wild type menG
50 3.55 ± 0.24 3.40 ± 0.14
150 3.65 ± 0.21 3.40 ± 0.29
300 2.48 ± 0.34 1.98 ± 0.15

116
Table 3.3: Magnetic parameters of PhyQ (A1-) and 2-phytyl-1,4-NQ (Q-) in the A1
binding site.
g-tensors a
gxx gyy gzz
A1- b 2.00622 2.00507 2.00218
Q- 2.00633 2.00507 2.00218
Hyperfine coupling principal values (MHz)
A11 A22 A33
A1- b 12.8 9.0 9.0
Q- -15.5 ± 0.5 -11.8 ± 0.5 <5
a the principal axes of the g-tensors are shown in Fig 3.1. b Zech et al., (2000) J. Phys. Chem.
B 104, 9728-9739 and references therein.
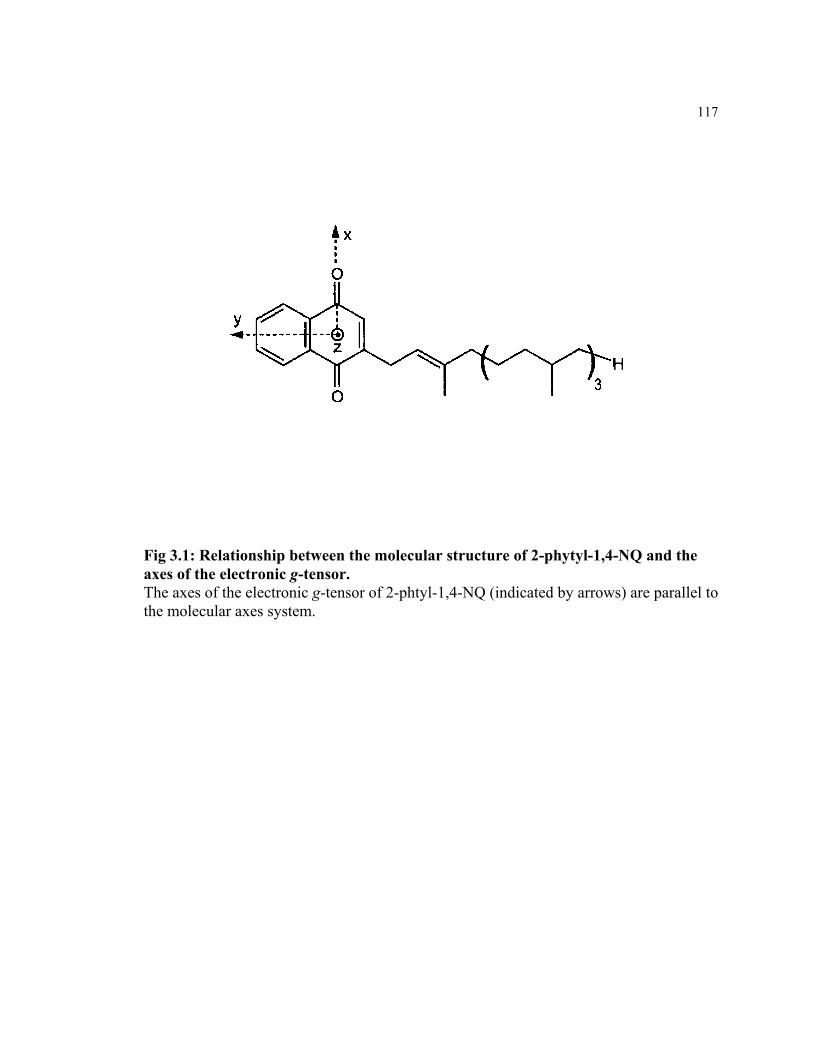
117
Fig 3.1: Relationship between the molecular structure of 2-phytyl-1,4-NQ and the axes of the electronic g-tensor. The axes of the electronic g-tensor of 2-phtyl-1,4-NQ (indicated by arrows) are parallel to the molecular axes system.
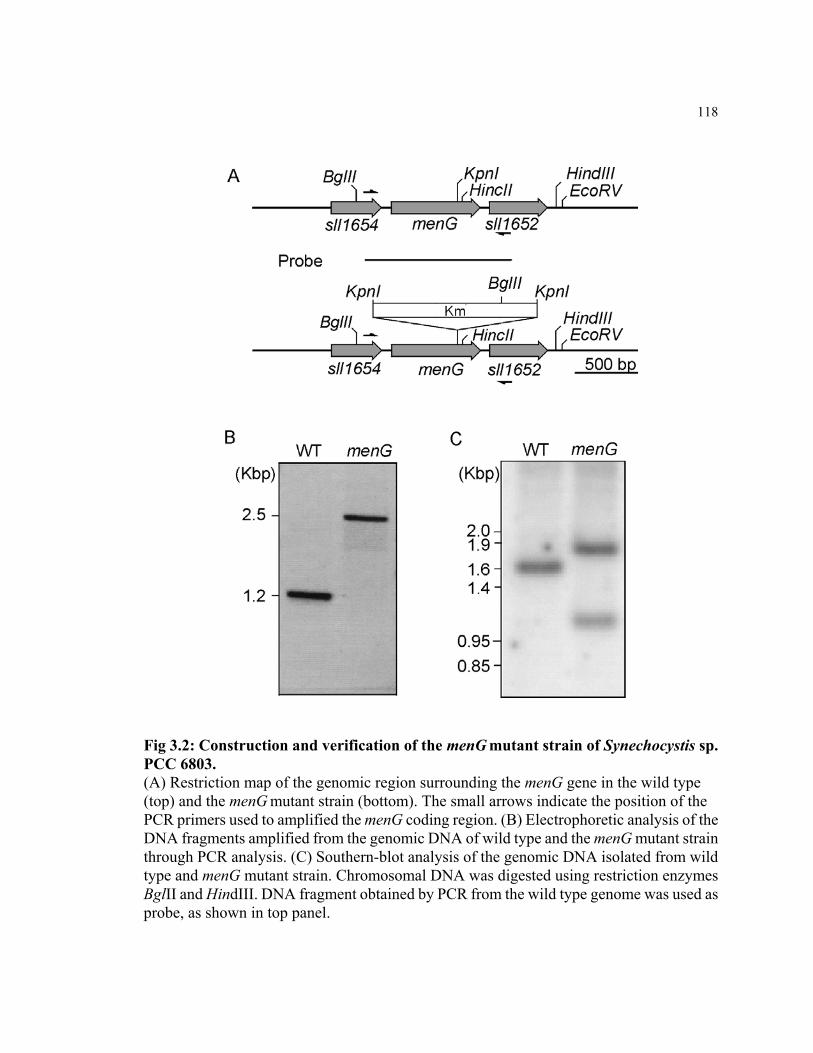
118
Fig 3.2: Construction and verification of the menG mutant strain of Synechocystis sp. PCC 6803. (A) Restriction map of the genomic region surrounding the menG gene in the wild type (top) and the menG mutant strain (bottom). The small arrows indicate the position of the PCR primers used to amplified the menG coding region. (B) Electrophoretic analysis of the DNA fragments amplified from the genomic DNA of wild type and the menG mutant strain through PCR analysis. (C) Southern-blot analysis of the genomic DNA isolated from wild type and menG mutant strain. Chromosomal DNA was digested using restriction enzymes BglII and HindIII. DNA fragment obtained by PCR from the wild type genome was used as probe, as shown in top panel.

119
Fig 3.3: Mass spectra recorded for pigments and quinone extracts from the PS I complexes isolated from the wild-type and menG mutant strains of Synechocystis sp. PCC 6803. (A) Wild type, and (B) the menG mutant.

120
Fig 3.4: 77 K Fluorescence emission spectra from whole cells of Synechocystis sp. PCC 6803. Chlorophyll fluorescence emission spectra were measured for the whole cells of wild type (solid lines) and the menG mutant strain (dotted lines) grown under three different light intensities, (A) 50 µE m-2 s-1, (B) 150 µE m-2 s-1, (C) 300 µE m-2 s-1. Each spectrum was recorded at the same cell density and presented as the average of four measurements.

121
Fig 3.5: SDS-PAGE analysis of PS I complexes isolated from the wild-type and menG mutant strains of Synechocystis sp PCC 6803. The equal amount of Chl a (5 µg) is loaded for each lane. Polypeptides were visualized after silver staining.

122
Fig 3.6: Photoaccumulated Q-band EPR spectra and simulations of PS I complexes from Synechocystis sp. PCC 6803.
Wild type (solid line), and the simulated spectrum (dotted line). B, menG mutant strain (solid line), and the simulated spectrum (dotted line). See text for details of the parameters of the simulation.

123
Fig 3.7: Transient spin-polarized EPR spectra (X-, Q- and W-) of the charge separated P700+ Q- state in PS I trimers. Wild-type spectra (dashed lines) and the menG mutant (solid lines). The frequencies are from bottom to top: 9, 35, and 95 GHz.

124
Fig 3.8: Simulations of the Q-band spectrum of the menG mutant carried out for various angles between the largest principal axis A11 and the gxx axis. Top: experimental spectrum as in Fig 3.4 (middle). Bottom: simulations as described and with the parameters specified in the text.
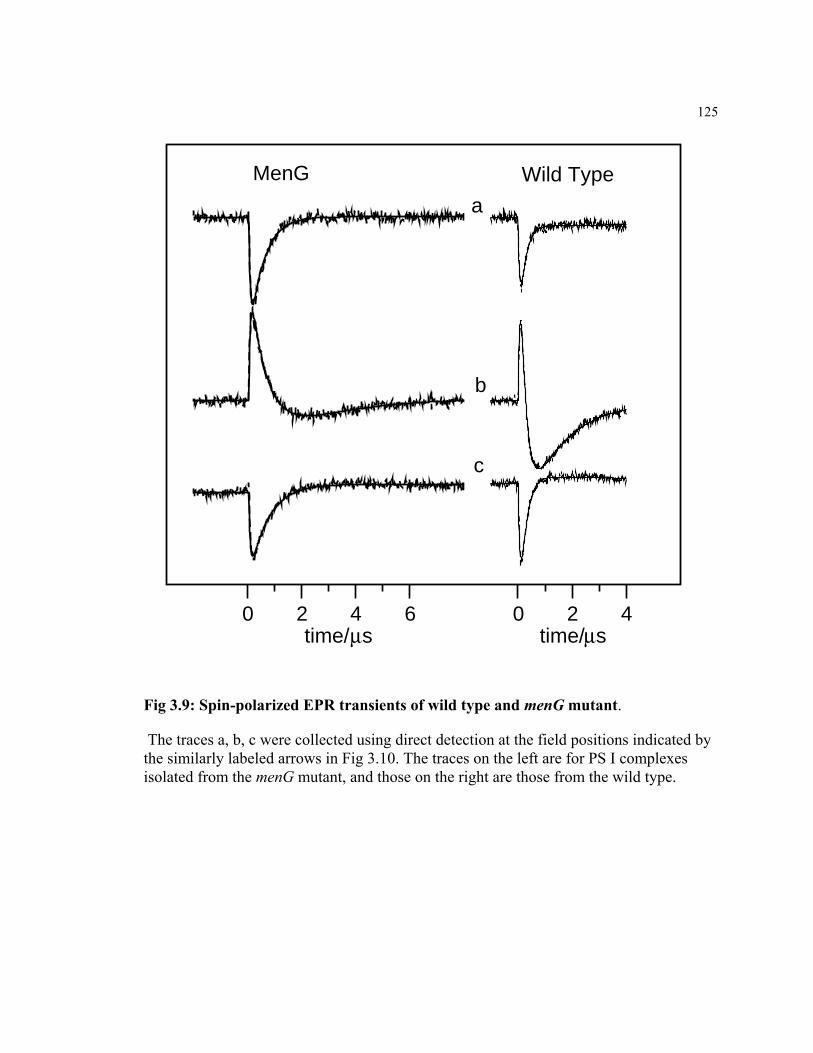
125
0 2 40 2 4 6
a
b
c
MenG Wild Type
time/µstime/µs
Fig 3.9: Spin-polarized EPR transients of wild type and menG mutant.
The traces a, b, c were collected using direct detection at the field positions indicated by the similarly labeled arrows in Fig 3.10. The traces on the left are for PS I complexes isolated from the menG mutant, and those on the right are those from the wild type.
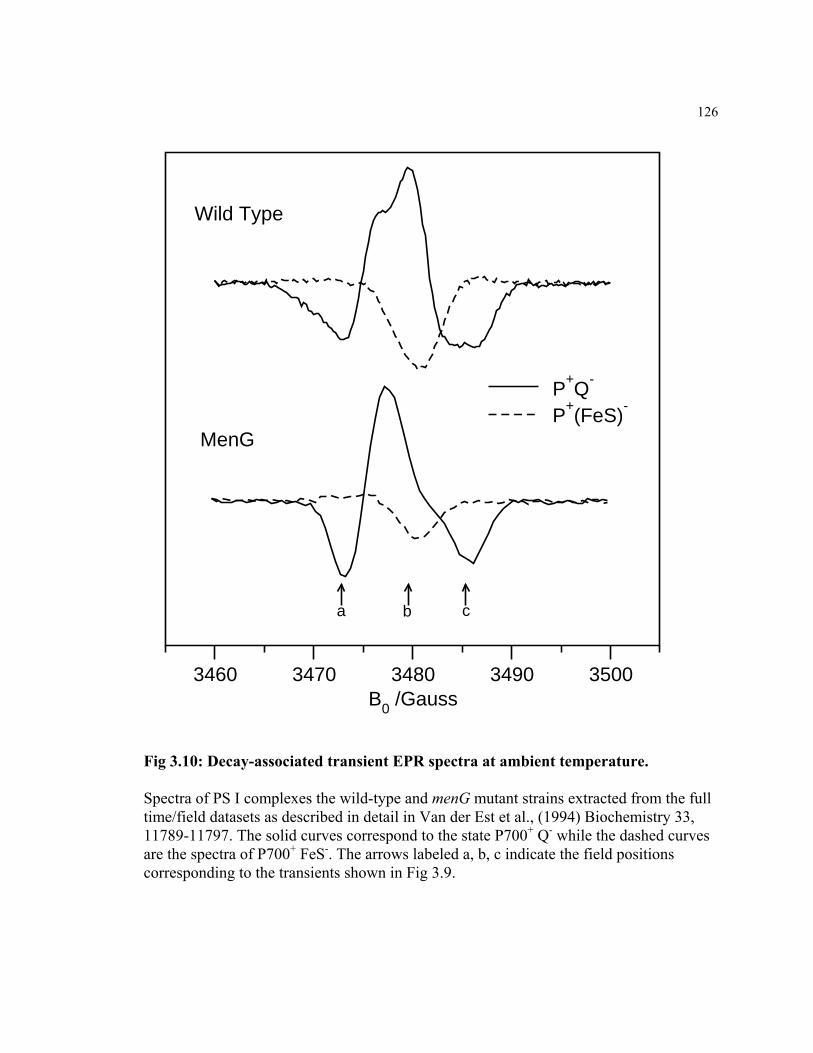
126
3460 3470 3480 3490 3500B0 /Gauss
a b c
P+Q
-
P+(FeS)
-
Wild Type
MenG
Fig 3.10: Decay-associated transient EPR spectra at ambient temperature. Spectra of PS I complexes the wild-type and menG mutant strains extracted from the full time/field datasets as described in detail in Van der Est et al., (1994) Biochemistry 33, 11789-11797. The solid curves correspond to the state P700+ Q- while the dashed curves are the spectra of P700+ FeS-. The arrows labeled a, b, c indicate the field positions corresponding to the transients shown in Fig 3.9.
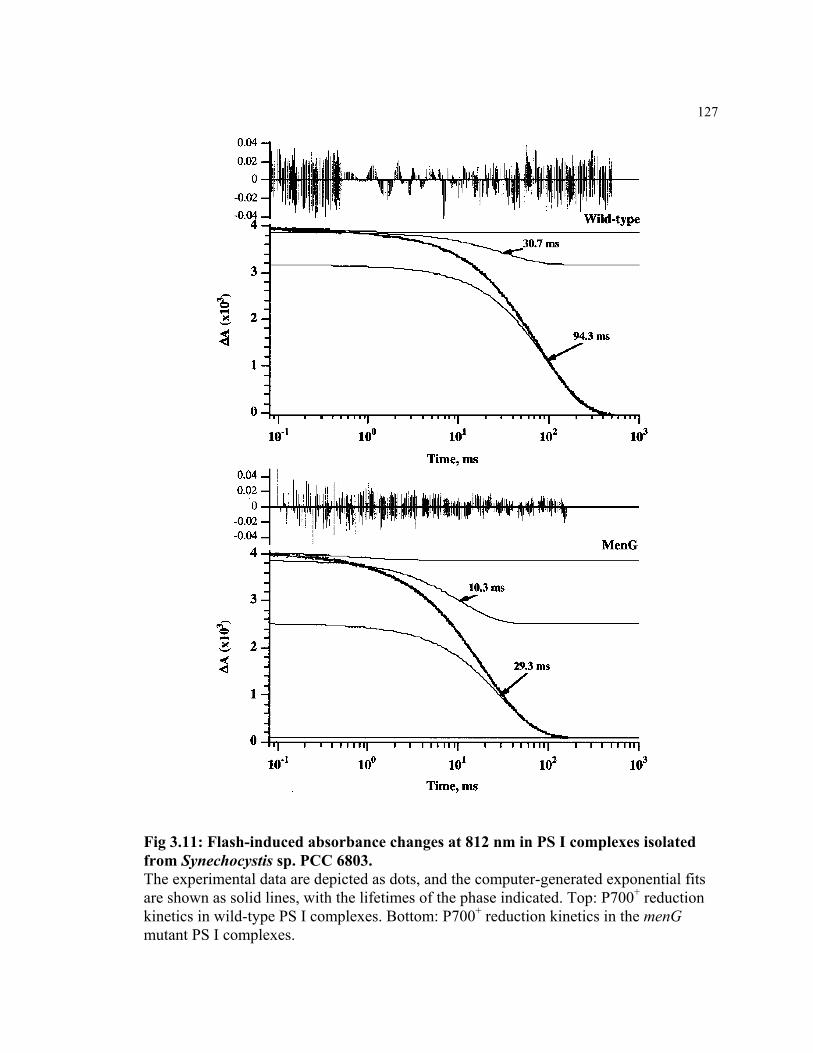
127
Fig 3.11: Flash-induced absorbance changes at 812 nm in PS I complexes isolated from Synechocystis sp. PCC 6803. The experimental data are depicted as dots, and the computer-generated exponential fits are shown as solid lines, with the lifetimes of the phase indicated. Top: P700+ reduction kinetics in wild-type PS I complexes. Bottom: P700+ reduction kinetics in the menG mutant PS I complexes.
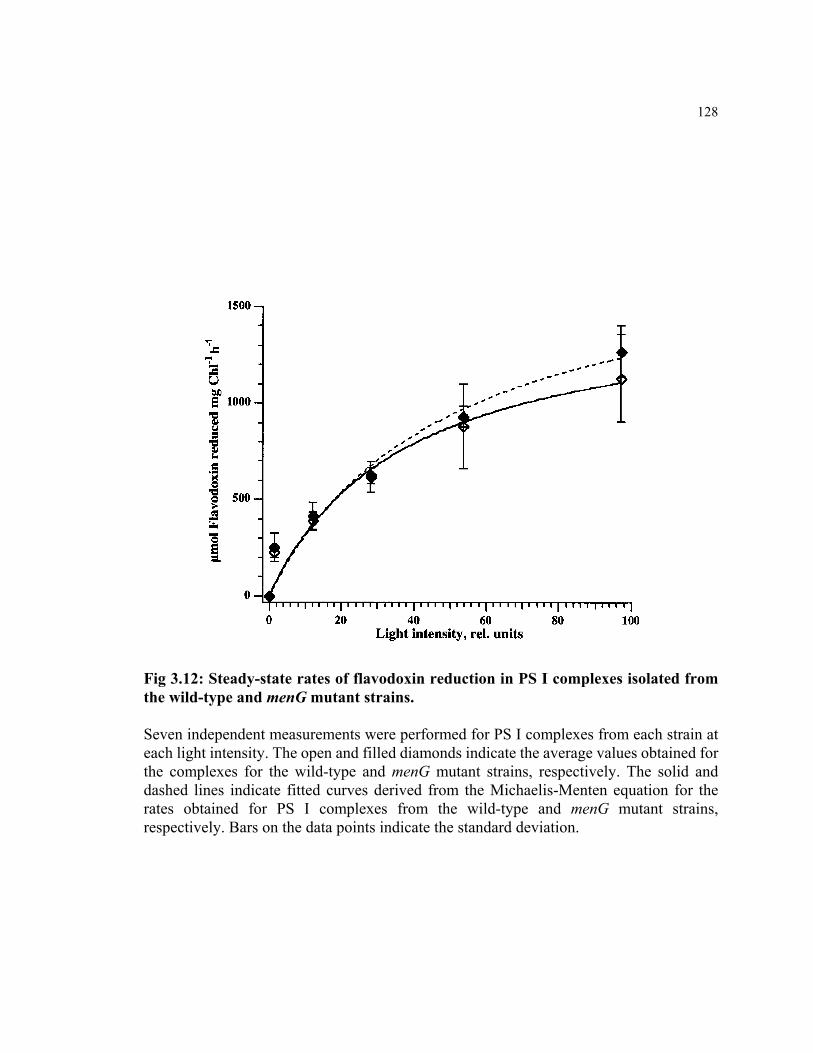
128
Fig 3.12: Steady-state rates of flavodoxin reduction in PS I complexes isolated from the wild-type and menG mutant strains. Seven independent measurements were performed for PS I complexes from each strain at each light intensity. The open and filled diamonds indicate the average values obtained for the complexes for the wild-type and menG mutant strains, respectively. The solid and dashed lines indicate fitted curves derived from the Michaelis-Menten equation for the rates obtained for PS I complexes from the wild-type and menG mutant strains, respectively. Bars on the data points indicate the standard deviation.

129
Chapter 4
Physiological Characterization of Tocopherol Biosynthesis Mutants in the
Cyanobacterium Synechocystis sp. PCC 6803: Demonstration of a Conditionally
Lethal Phenotype in the Presence of Glucose at pH 7.0
Publications:
Zigang Cheng, Scott Sattler, Hiroshi Maeda, Yumiko Sakuragi, Donald A. Bryant, Dean
DellaPenna (2003) Highly divergent methyltransferases catalyze a conserved reaction in
tocopherol and plastoquinone synthesis in cyanobacteria and photosynthetic eukaryotes.
Plant Cell 15: 2343-2356
Yumiko Sakuragi, Hiroshi Maeda, Dean DellaPenna and Donald A. Bryant, manuscript
in preparation.

130
ABSTRACT
The phenotypes of three tocopherol mutant populations, slr1736, sll0418 and slr0089,
previously selected in the presence of glucose in the cyanobacterium Synechocystis sp.
PCC 6803, have been reexamined. When cultured on solid media containing glucose, the
mutant populations showed extremely heterogeneous colony morphologies, which
suggested that the strains were genotypically inhomogeneous. Newly isolated tocopherol
mutant strains were unable to grow in the presence of glucose at pH values below 7.2.
Therefore, it was concluded that “authentic” slr1736, sll0418, and slr0089 mutants are
glucose-sensitive and are not able to grow in the presence of glucose. Further
characterization of the “authentic” mutants revealed that the chlorophyll a,
phylloquinone, and plastoquinone contents decreased when cells were grown in the
presence of glucose, and Photosystem II (PS II) activity was completely lost after 24 h.
However, levels of the D1 and PsbO proteins remained nearly constant, and the levels of
the sodB transcripts (encoding the superoxide dismutase) were virtually identical between
the wild-type and mutant strains before and after transfer to medium containing glucose.
These results indicate that the loss of the D1 and PsbO proteins is not responsible for the
loss of PS II activity and strongly support the conclusion that the glucose-induced
phenotype is not directly associated with oxidative stress. The authentic mutants
exhibited chlorosis when grown in the presence of glucose. The relative phycobiliprotein
contents decreased to nearly one-third of the wild-type level, and the concomitant
accumulation of the nblA (encoding a protein essential for the degradation of
phycobilisomes) transcript was detected. These results indicate that the authentic mutants

131
experience macronutrient starvation when grown in the presence of glucose. Further
growth analyses revealed that the glucose-induced growth defects, PS II inactivation, and
the macronutrient starvation responses occur at pH below 7.2. At pH 7.0 in the presence
of glucose, the PS II activity was completely lost, and the growth ceased after 24 h. The
results further demonstrate that α-tocopherol plays an essential role for the growth and
survival of Synechocystis sp. PCC 6803 in the presence of glucose and suggest that α-
tocopherol plays a role in the regulation of macronutrient metabolism in cyanobacteria.

132
ABBREVIATIONS
Chl a chlorophyll a
DCMU 3-(3,4-dichlorophenyl)-1,1-dimethyl-urea
HEPES 4-(2-hydroxyethyl)-piperazine-N’-(2-ethanesulfonic acid)
HGA homogentisic acid
HPP 4-hydroxyphenylpyruvate
HPPD 4-hydroxyphenylpyruvate dioxygenase
HPT homogentisate phytyltransferase
MPBQ 2-methyl-6-phytyl-1,4-benzoquinol
MPBQ MT 2-methyl-6-phytyl-1,4-benzoquinol methyltransferase
PBP phycobiliprotein
PCR polymerase chain reaction
PS II Photosystem II
RT-PCR reverse transcriptase-polymerase chain reaction

133
INTRODUCTION
α-Tocopherol (vitamin E) is a lipid-soluble, organic molecule that is synthesized
only by oxygen-evolving phototrophs, including some cyanobacteria and all green algae
and plants (Threlfall and Whistance, 1971; Collins and Jones, 1981). The conservation of
α-tocopherol synthesis during the evolution of photosynthetic organisms suggests that the
molecule performs one or more critical functions. In plants, α-tocopherol is synthesized
and localized in the plastids (Soll et al., 1980, 1985), and it is particularly abundant in the
thylakoid membranes (Fryer, 1992), in which it is thought to afford protection against
various oxidative stresses (Munné-Bosch and Lenor, 2000). Because α-tocopherol is also
an essential component of animal diets, most of our knowledge of tocopherol function
has been obtained from studies in these systems. Studies in whole animals, animal cell
cultures and artificial membranes have shown that tocopherols scavenge and quench
various reactive oxygen species and lipid oxidation byproducts, which would otherwise
propagate lipid peroxidation chain reactions in membranes (Kamal-Eldin and Appelqvist,
1996). In addition to these antioxidant functions, other “non-antioxidant” functions
related to modulation of signaling and transcriptional regulation in mammals have also
been reported (Chan et al., 2001; Azzi et al., 2002; Ricciarelli et al., 2002). In
photosynthetic organisms, tocopherol functions have not yet been rigorously defined, but
it is believed that they are likely to include some or all of the functions reported in
animals, as well as other possible functions specific to photosynthetic organisms.

134
The biosynthetic pathway of α-tocopherol is summarized in Fig 4.1. It was
elucidated in plants in the mid-1980’s and during the past five years, most of the genes
encoding the enzymes of this pathway have been identified in both plants and
cyanobacteria (Threlfall and Whistance, 1971; d’Harlingue and Camara, 1985; Norris et
al., 1998; Shintani and DellaPenna, 1998; Collakova and DellaPenna, 2001; Schledz et
al., 2001; Porfirova et al., 2002; Savidge et al., 2002; Shintani et al., 2002; Dähnhardt et
al., 2002; Collakova and DellaPenna, 2003; Koch et al., 2003; Cheng et al., 2003).
Briefly, homogentisic acid (HGA) is the aromatic head-group precursor for all
tocopherols, and HGA is produced from 4-hydroxyphenylpyruvate (HPP) by the enzyme
HPP dioxygenase (Norris et al., 1998; Dähnhardt et al., 2002). The committed step in
tocopherol synthesis is the condensation of HGA and phytyl-pyrophosphate by the
enzyme homogentisate phytyltransferase to produce 2-methyl-6-phytyl-1,4-benzoquinol
(MPBQ) (Collakova and DellaPenna, 2001; Schledz et al., 2001; Savidge et al., 2002;
Collakova and DellaPenna, 2003). The ring of MPBQ is methylated by MPBQ
methyltransferase to yield 2,3-dimethyl-5-phytyl-1,4-benzoquinol (Soll et al., 1985;
Shintani et al., 2002; Cheng et al., 2003), which is cyclized by tocopherol cyclase to yield
γ-tocopherol (Soll et al., 1985; Porfirova et al., 2002; Sattler et al., 2003). Finally, a
second ring methylation by γ-tocopherol methyltransferase yields α-tocopherol (Soll et
al., 1980; d’Harlingue and Camara, 1985; Shintani and DellaPenna, 1998, Koch et al.,
2003). Genes encoding HPP dioxygenase (slr0090), homogentisate phytyltransferase
(slr1736), MPBQ methyltransferase (sll0418) and γ-tocopherol methyltransferase
(slr0089) have been identified in Synechocystis sp. PCC 6803 (Shintani and DellaPenna,

135
1998; Shintani et al., 2002; Collakova and DellaPenna, 2001; Dähnhardt et al., 2002,
Cheng et al., 2003). The remaining enzyme, tocopherol cyclase, has recently been shown
to be the product of slr1737 in Synechocystis sp. PCC 6803 (Sattler et al., 2003).
In the cyanobacterium Synechocystis sp. PCC 6803, the tocopherol biosynthetic
pathway has been studied genetically and homozygous mutants of the slr1736, sll0418,
and slr0089 genes, encoding homogentisate phytyltransferase, MPBQ methyltransferase,
and γ-tocopherol methyltransferase, respectively, have been obtained (Shintani and
DellaPenna, 1998; Shintani et al., 2002; Cheng et al., 2003; Collakova and DellaPenna,
2001). As expected from the biosynthetic pathway (Fig 4.1), the slr0089 mutant
accumulates only γ-tocopherol (Shintani and DellaPenna, 1998), the sll0418 mutant
accumulates 30% of the wild-type level of α-tocopherol and a small amount of β-
tocopherol (Cheng et al., 2003), and the slr1736 mutant does not accumulate any
tocopherols (Collakova and DellaPenna, 2001). In light of the established antioxidant
activity of α-tocopherol in animal systems, one would expect that the loss or reduction of
α-tocopherol would lead to a detectable phenotypic difference between the wild-type and
mutant strains. Intriguingly, however, the tocopherol-deficient slr1736 mutant grew
similarly to the wild-type strain both in the absence and in the presence of glucose at a
moderate light intensity (110 µE m-2 s-1) (Collakova and DellaPenna, 2001). A limited
comparative analysis of the photosynthetic activity of the wild-type and slr1736 mutant
strains failed to reveal any differences. This led the authors to conclude that α-tocopherol
is dispensable for the survival of Synechocystis sp. PCC6803 under the conditions tested
(Collakova and DellaPenna, 2001).

136
In this study, it is demonstrated that the “authentic” tocopherol mutants are
glucose-sensitive and that they cannot maintain growth in the presence of glucose at pH
7.2 or lower. Mutant populations obtained in previous studies probably contained
mixtures of secondary suppressor mutations that accumulate during cultivation in the
presence of glucose, although their exact nature has yet to be identified. The glucose-
sensitive phenotype of the “authentic” mutants was characterized, and the data indicate
that α-tocopherol is essential for the survival of Synechocystis sp. PCC 6803 in the
presence of glucose at pH 7.2 or lower.

137
RESULTS
Extremely Heterogeneous Colony Morphologies in Tocopherol Mutants Previously
Isolated in the Presence of Glucose
When maintained on solid media containing 5 mM glucose, three tocopherol
mutants, slr1736::aphII, sll0418::aphII, and slr0089::aphII that had been isolated in
previous studies in the presence of glucose, showed much larger variations in colony size
and pigmentation than what is typically observed for the wild-type strain (Fig 4.2 E-H).
The larger colonies exhibited more intense blue-green pigmentation, which is
characteristic of wild-type colonies, while the smaller colonies appeared more chlorotic
and were yellow-green in color. Generally such non-homogenous colony morphology is
indicative of heterogeneous genotypes within a given population. However, PCR analysis
of genomic DNA isolated from three mutant strains showed that the targeted, mutated
loci were homozygous and that no corresponding wild-type allele could be detected in
any of the three mutant strains. These results eliminate the possibility that the observed
colony heterogeneity (Fig 4.2) was due to incomplete segregation of the mutant loci or to
contamination of the cultures with wild-type cells. When the tocopherol mutants were
maintained in the absence of glucose, more uniform colony morphologies were observed
(Fig 4.2 A-D). Although the size of colonies showed somewhat larger variation as
compared to wild type, all colonies showed intense blue-green pigmentation. Therefore it
was concluded that the heterogeneity of colony morphologies observed for the tocopherol
mutants is a specific response to growth in the presence of glucose.

138
The growth characteristics of two cell lines, one derived from a small colony and
the other derived from a large colony of the slr0089 mutant selected in the presence of
glucose, were analyzed in the absence and in the presence of glucose. In the absence of
glucose, the growth behavior of both cell lines was virtually indistinguishable from that
of wild type (Fig 4.3 A and B). Upon transfer to media containing glucose, however, the
cell line derived from the small colony ceased growth within 24 h and did not recover
during the time course of measurements (Fig 4.3 C). The cell line derived from the large
colony grew somewhat more slowly than the wild-type strain, but it was able to maintain
growth during the course of the experiment (Fig 4.3 D). The dramatic phenotypic
difference between the two cell lines strongly suggested that they were genotypically
distinct, and that the mutant populations previously obtained by selection in the presence
of glucose (Shintani and DellaPenna, 1998; Collakova and DellaPenna, 2001; Shintani et
al., 2002) were genotypically inhomogeneous.
Tocopherol Mutants Are Glucose Intolerant
“Authentic” tocopherol mutant strains were constructed by transforming an
immotile, glucose-tolerant wild-type strain of Synechocystis sp. PCC 6803 with genomic
DNAs isolated from the slr1736, sll0418, and slr0089 mutants selected in the presence of
glucose. Because the presence of glucose was found to be unfavorable for some
populations of these tocopherol mutants, kanamycin-resistant transformants were selected
in the absence of glucose. Segregation of each mutant allele from the corresponding wild-
type allele was tested by PCR analysis using specifically designed oligonucleotide
primers (Fig 4.4). Products anticipated for the wild-type slr1736, sll0418, and slr0089

139
genes were 1.3 kbp, 1.0 kbp, and 1.2 kbp, respectively. No such product was observed for
any of the mutant strains; instead, a product ~1.3 kbp larger was detected in each mutant
line. This difference in size corresponds to the aphII-containing, kanamycin-resistance
cartridge originally used for the insertional inactivation of each gene. It was therefore
concluded that each mutant strain is homozygous and that the slr1736::aphII,
sll0418::aphII, and slr0089::aphII alleles in each mutant line segregated completely in
the absence of glucose.
The newly isolated slr1736, sll0418, and slr0089 mutants produced uniform
colonies and grew similarly to the wild type in the absence of glucose under moderate
light intensity (< 50 µE m-2 s-1; Fig 4.5 A). In the presence of glucose, however, all three
mutants showed severe growth defects and were incapable of maintaining growth after 24
h (Fig 4.5 B and C). This growth phenotype of the mutant strains selected in the absence
of glucose is very similar to that observed for the cell line derived from the small colonies
of the mutants selected in the presence of glucose (see Fig 4.3 C), but sharply contrasts
with the growth behavior of the cell lines derived from the large colonies (Fig 4.3 D).
These results clearly demonstrate that “authentic” slr1736, sll0418, and slr0089 mutants
are glucose sensitive. The data also suggest that α-tocopherol plays an essential role for
the survival of Synechocystis sp. PCC 6803 in the presence of glucose.
When cells from these “authentic” mutants grown in the presence of
glucose for 72 h were transferred to fresh medium containing no glucose, no
growth was detected even after 3 weeks of incubation. These results indicated that
these “authentic” mutants lost viability during cultivation in the presence of

140
glucose for 72 h. The pH of the cultures grown in the presence of glucose
typically shifted from the original value (pH 8.0) to a value in the range of 7.0 to
7.3. Preconditioned media prepared from cultures of the “authentic” mutants
grown in the presence of glucose supported the normal growth of the wild-type
cells. These results show that the glucose toxicity of the mutants is not the result
of products excreted from the cells in the presence of glucose.
To test whether the growth light intensity was responsible for the observed failure
of the “authentic” tocopherol mutants to grow in the presence of glucose, the “authentic”
slr1736 mutant was grown in the presence of glucose at various light intensities. Under
both weak (< 5 µE m-2 s-1) and strong (300 µE m-2 s-1) illumination conditions, however,
the presence of glucose in the growth medium resulted in the death of the “authentic”
slr1736 mutant (Fig 4.6 A and B). This indicates that light intensity is unlikely to be a
contributing factor in the observed glucose toxicity of the tocopherol mutants. The effects
of inhibitors of the electron transport chain on the glucose toxicity of the slr1736 mutant
were also studied. Addition of a sublethal concentration of DCMU (3-(3,4-
dichlorophenyl)-1,1-dimethyl-urea) to the growth medium led to a small recovery of
growth for the slr1736 mutant in the presence of glucose (Fig 4.6 C). However,
increasing the DCMU concentration further did not improve the growth of the slr1736
mutant. Addition of potassium cyanide in various concentrations did not rescue the
slr1736 mutant from the glucose toxicity (Fig 4.6 D).
The effects of other organic carbon sources were analyzed on the growth of the
“authentic” slr1736 mutant. The presence of 5 mM pyruvate, succinate, and fumarate,

141
which are intermediates of the TCA cycle, did not cause growth defects in the slr1736
mutant. (Fig 4.7 A-D). The presence of 15 mM glutamate did not affect the growth of the
slr1736 mutant (Fig 4.7 E). The presence of ornithine, however, resulted in growth
defects similar to those caused by glucose (Fig 4.7 F). The results suggested that the
toxicity induced by glucose is caused by one or more metabolic intermediate that
accumulate during glucose and ornithine catabolism. Ornithine is an intermediate in the
citrulline cycle, in which as much as 20 % of CO2 fixation can take place (Tabita, 1987).
On the other hand, glucose is metabolized through the glycolytic pathway or oxidative
pentose phosphate pathway. Intermediates of these pathways are also shared with the
Calvin-Benson cycle (reductive pentose phosphate pathway), in which the major CO2
fixation occurs (Tabita, 1987). The fact that both glucose and ornithine caused lethal
effects on the tocopherol mutants implies that α-tocopherol is perhaps involved in carbon
metabolism by coordinating inorganic and organic carbon metabolic balance in the cells.
Chlorophyll a, Phylloquinone and Plastoquinone Content
The complete loss of, or a decrease in, tocopherol content led to small changes in
the pigmentation of the “authentic” tocopherol mutants grown in the absence of glucose.
The chlorophyll a (Chl a), phylloquinone, and plastoquinone contents were
approximately 10%, 10-20 %, and 3-30% lower, respectively, in these tocopherol
mutants than in the wild type (Fig 4.8). The slr1736 mutant cells contained nearly the
same amount of Chl a and quinones as wild type, while the sll0418 and slr0089 mutant
cells contained somewhat lower amounts. These differences, however, did not seem to
affect the photosynthetic electron transport activity; the rates oxygen evolution observed

142
for both whole-chain electron transport and the Photosystem II (PS II)-dependent electron
transport for the three tocopherol mutants were very similar to, or perhaps somewhat
higher than, those of wild type (Fig 4.9, see below).
When grown in the presence of glucose, however, the mutants exhibited more
dramatic changes in the contents of Chl a, phylloquinone, and plastoquinone. The Chl a
and phylloquinone contents were largely unchanged by 24 h after transfer from medium
without glucose to medium containing glucose and decreased significantly after 48 to 72
h to as low as 50% of the corresponding wild-type level. Under these conditions, the Chl
a as well as phylloquinone level of wild type was largely unaffected during the course of
measurements (Fig 4.8 A and B). Because phylloquinone is associated exclusively with
the Photosystem I complexes in cells, the parallel decreases in the Chl a and
phylloquinone contents may imply a decrease in the Photosystem I content in the cells.
In contrast, following transfer from a medium without glucose to a medium
containing glucose, the plastoquinone content gradually decreased in the wild-type strain
(Fig 4.8 C); by 48 h, the plastoquinone content of wild type was only half of that found in
the absence of glucose. An even more severe decrease was observed in the mutants, in
which the plastoquinone content decreased to ~50 % of the wild-type level by 24 h and as
low as 15 % by 72 h after transfer to medium containing glucose (Fig 4.8 C). Recent
studies have shown that the redox state of the plastoquinone pool in the thylakoid
membranes modulates the expression levels of photosynthetic genes and affects the
cellular contents of components of the photosynthetic apparatus, including the
phycobilisomes and Photosystem I (Alfonso et al., 2000; Li and Sherman, 2000). This

143
raises the question as to whether the dramatic decrease in the plastoquinone content of the
“authentic” mutants could lead to a change in its redox state, which in turn could be
responsible for the decrease in the phycobiliprotein content and PS II activity (see
below), and ultimately cell death. The redox state of the plastoquinone pool can be
modified by various growth conditions including the intensity of illumination applied
during cell growth. Low light intensity (< 5 µE m-2 s-1) typically causes reduction of the
plastoquinone pool while high light intensity (300 µE m-2 s-1) causes its oxidation
(Cooley and Vermaas, 2001). However, incubation under either of these light conditions
did not lead to recovery of growth in the “authentic” tocopherol mutants in the presence
of glucose (Fig 4.6 A and B). Similarly, addition of various concentrations of potassium
cyanide, which promotes reduction of the plastoquinone pool, did not alleviate the
glucose sensitivity of these mutants (Fig 4.6 D), and addition of DCMU, which causes
oxidation of the plastoquinone pool, did so only to a small extent (Fig 4.6 C). These
combined results therefore suggest that the changes in the plastoquinone pool size, and its
possibly altered redox state, are not the primary cause of the glucose toxicity observed for
the authentic tocopherol mutants in the presence of glucose.
Photosystem II Activity
Given the significant reduction in the Chl a, phylloquinone, and plastoquinone
contents in response to the presence of glucose, whole-chain and PS II-dependent
electron transport was assessed in the mutants (Fig 4.9). In cells grown in the absence of
glucose, the whole-chain and the PS II-dependent oxygen evolution rates of all three
mutants were similar to, or even slightly higher, than that of the wild type. Likewise,

144
somewhat higher O2-evolving activity (the whole-chain or the PS II-dependent) was
detected for the wild type when grown in the presence of glucose. In contrast, no O2-
evolving activity (the whole-chain and the PS II-dependent) was detected for the
tocopherol mutants when grown in the presence of glucose.
Loss of PS II activity is often seen when cells are exposed to oxidative stress
caused by exposure to extremely high light, to low temperature, or to an environment
containing a high salt concentration (Allakhverdiev et al., 1999; Hideg et al., 2000; see a
review by Aro et al., 1993, and references therein), and this phenomenon is known as
photoinhibition. Under such conditions, the D1 protein, a subunit that forms a part of the
PS II core structure (Ferreira et al., 2004), rapidly degrades, and, as a result, the activity
of PS II is lost. The possibility that the observed PS II inactivation is related to
photoinhibition in the tocopherol mutant in response to glucose was tested by
immunological assay. When cells were grown in the absence of glucose, the amount of
immunologically detectable D1 protein was virtually indistinguishable between the wild-
type and the mutant strains (Fig 4.10). After transfer to medium containing glucose, the
amount of immunologically detectable D1 protein was only slightly reduced in the
mutants compared to wild type, even though by 24 h a complete loss of PS II-mediated
oxygen evolution activity was observed (Fig 4.9). PsbO, or the 33 kDa protein, is a
subunit that forms a peripheral domain of PS II known as the oxygen-evolving complex
(Ferreira et al., 2004). This complex is responsible for the catalysis of water splitting and
the production of oxygen. The immunologically detectable PsbO protein was also largely
unaffected in response to glucose in both wild-type and three tocopherol mutant strains.

145
These results demonstrate that the loss of PS II activity in the tocopherol mutants in the
presence of glucose is not associated with structural damage that leads to a loss of the D1
protein and PsbO, and they suggest that oxidative stress is not the cause of the PS II
inactivation. The expression of the sodB gene, encoding superoxide dismutase, which is
known to accumulate several-fold in response to various reactive oxygen species
(Ushimaru et al., 2002), was analyzed by RT-PCR (Fig 4.11). The amounts of the sodB
transcript detected for the wild type and the slr1736 mutant were very similar when cells
were grown in the absence of glucose (Fig 4.11, shown at 0 h) and in the presence of
glucose (Fig 4.11, at 4, 8, and 24 h). The combined results therefore strongly suggest that
the loss of α-tocopherol does not cause oxidative stress in response to glucose and that
the inactivation of PS II is unrelated to oxidative stress.
Relative Phycobiliprotein Content
The relative content of phycobiliproteins (PBPs), the primary protein components
of the light-harvesting antennae known as phycobilisomes, was determined in response to
the transfer of the wild type and the “authentic” tocopherol mutants to medium
containing glucose (Fig 4.12 A). In the absence of glucose, all three tocopherol mutants
contained 20-25% less PBP than the wild type. After 24 h of growth in the presence of
glucose, the PBP content of wild-type cells decreased to ~60% of the level found in the
absence of glucose, while the PBP content of the three tocopherol mutants decreased
even more sharply to 18-26% of the level in their respective controls obtained for the
cells grown in the absence of glucose. Time-course analyses revealed that the reduction
of PBPs initiates by 4 h after transfer to medium containing glucose (Fig 4.12 B). In

146
comparing the decreased PBP levels of the wild type and the three tocopherol mutants
during growth in the presence of glucose, it is obvious that the loss of PBPs is much more
severe in the three mutants than in the wild type.
These data are consistent with the pale, chlorotic (yellow-green) coloration of the
mutant colonies in the presence of glucose. The mutants had significantly lower
absorption in the orange-red region of their visible spectra, and as a result were less blue-
green in coloration than control cells. This bleaching, due to a decrease in the cellular
content of PBPs, is well known to be associated with nutrient starvation, and can be
caused by carbon (Miller and Holt, 1977), nitrogen (Allen and Smith, 1969; Foulds and
Carr, 1977; Wood and Haselkorn, 1980; Yamanaka and Glazer, 1980; Stevens et al.,
1981; Elmorjani and Herdman, 1987), phosphorous (Ihlenfeldt and Gibson, 1975), sulfur
(Schmidt et al., 1982; Jensen and Rachlin, 1984; Warner et al., 1986), or iron limitation
(Sherman and Sherman, 1983). The nblA gene, encoding a factor essential for the
controlled degradation of phycobilisomes, is known to accumulate in Synechocystis sp.
PCC 6803 cells in response to nitrogen deprivation (Richaud et al., 2001). RT-PCR
analyses showed that, in the wild-type samples, the nblA transcript was barely detectable
when cells were grown in the absence of glucose (Fig 4.11, shown at 0 h) and that a small
induction occurred by 4 h after transfer to medium containing glucose. In the slr1736
mutant sample, the wild-type level of the nblA transcript was detected in the absence of
glucose and a dramatic increase followed by 4 h after transfer to medium containing
glucose. This correlates well with the observed behavior of PBP contents in the
tocopherol mutants (Fig 4.12, also see above) and suggests that the slr1736 mutant is

147
experiencing nutrient limitation in response to glucose. Because the BHEPES medium
used in culturing these mutants contains an appropriate mixture of all micronutrients and
macronutrients to support the growth of Synechocystis sp. PCC 6803 (Stainer et al., 1971;
see Materials and Methods), the observed macronutrient starvation response of the
mutants in the presence of glucose is most likely to be due to an impairment in an
essential biosynthetic or regulatory process, which involves carbon, nitrogen, and
glucose.
Involvement of CO2 in the Glucose Toxicity
The observed glucose-sensitive phenotype of the “authentic” tocopherol mutants
resembled that of the icfG mutant reported in a previous study (Beuf et al., 1994). The
icfG gene encodes a putative protein phosphatase similar to RsbU (anti anti-sigma factor
B) in Bacillus subtilis (Price, 2000; Woodbury et al., 2004). This mutant showed a
glucose sensitive-phenotype only at low CO2 [air level, 0.035% (v/v)] levels. The
presence of succinate, fumarate, pyruvate, and glutamate did not cause lethality at low
CO2, while the presence of ornithine caused a lethal effect. The striking behavioral
resemblance between the icfG mutant and the “authentic” tocopherol mutants in their
response to various carbon sources suggested a possible involvement of CO2 in the
glucose toxicity observed for the tocopherol mutants. Fig 4.13 shows the growth curves
of wild type and the “authentic” slr1736 mutant in the presence of glucose in air and 3 %
(v/v) CO2. The slr1736 mutant grew similarly to the wild type in air even in the presence
of glucose, and in fact, the growth of the two strains was virtually indistinguishable under
these conditions. When cells were grown in the presence of glucose at 3% (v/v) CO2,

148
however, the slr1736 mutant exhibited severe growth defects as described above.
Therefore, these results demonstrated that the glucose toxicity occurs only at high CO2
levels. This led to the hypothesis that α-tocopherol is involved in the global regulation of
organic and inorganic carbon metabolism in Synechocystis sp. PCC 6803, possibly
through a signal transduction cascade involving IcfG.
Involvement of pH in Glucose Toxicity
The observed dependence of the glucose toxicity on the CO2 concentration was,
however, questioned after discussion with Prof. Aaron Kaplan of the Hebrew University,
Israel. In the presence of 3% (v/v) CO2, the growth medium was found to acidify from
the initial pH 8.0 to about pH 7 (see above). Acidification of cultures can be caused by a
high concentration of CO2, such as 3% (v/v), since the in organic carbon (Ci ) supplied
into medium as dissolved CO2 is rapidly hydrated and reaches equilibrium with HCO3-.
Thus, the possibility that the high-CO2 requirement of the glucose toxicity was actually
because of medium acidification was tested directly. The wild-type and the “authentic”
tocopherol mutants were grown at various pH values at 1% CO2 (v/v) in medium
BHEPES40, which is a modified BHEPES medium supplemented with 40 mM HEPES
for higher pH buffering capacity (see Materials and Methods). After culturing for 2 days
in this medium at 1% (v/v) CO2, the pH remained near the original value within a
deviation of ≤ 0.05 pH unit. In the absence of glucose, both the wild type and the
tocopherol mutant strains grew well at all pH values tested between 6.8 and 8.0, and no
significant difference was observed among the four strains (Fig 4.14). This establishes
that the loss of α-tocopherol does not affect the growth rates of the cells at all pH values

149
between 6.8 and 8.0 in the absence of glucose. When the cells were transferred to media
containing glucose, the wild type grew faster as compared to the growth in the absence of
glucose (Fig 4.14), which confirms that the glucose is taken up and metabolized in the
wild-type cells. The tocopherol mutants grew similarly to wild type at pH 8.0 and 7.6
(Fig 4.14). However, they grew very poorly or not at all at pH 7.2, 7.0, and 6.8. In
comparing the pH-independent growth phenotype in the absence of glucose and the pH-
dependent growth in the presence of glucose, it is obvious that the observed glucose
toxicity of the tocopherol mutants occurs at pH 7.2 or lower. Therefore, the glucose-
sensitive phenotype characterized in detail in the earlier section of this chapter is also due
to the lowering of the pH caused by the acidification of the growth media during growth
in the presence of 3% (v/v) CO2.
Fig 4.15 shows that growth curves of the wild-type and the “authentic” slr1736
mutant cultures at various pH values in air and 1% (v/v) CO2. The slr1736 mutant grew
similarly to wild type at pH 8.0 and 7.6 both in air and 1% (v/v) CO2 (Fig 4.15 A and B).
At pH 7.2, the slr1736 mutant grew slightly slower than wild type during the initial 20 h
and grew even more slowly after 20 h (Fig 4.15 C). At pH 7.0 and 6.8, the mutant grew
substantially slower than wild type during the first 20 h and then stopped growing after
20 h (Fig 5.2 D and E). The growth behavior of the slr1736 mutant was virtually
indistinguishable either when grown with air or with 1% (v/v) CO2 (Fig 4.15 A-E). These
results demonstrate it is the pH of the medium and not the extracellular CO2
concentration that causes the glucose-sensitive growth phenotype of the slr1736 mutant.

150
It is noteworthy that the pH-dependent glucose toxicity occurs after 20 h, which
suggests that the cells were accumulating a toxic substance during the initial 20 h. 3-O-
methyl-glucose is known to be transported into the cells through the glucose transporter;
however, it is not phosphorylated and therefore is not metabolized further (Beuf et al.,
1994). Both the wild type and the slr1736 mutant grew well in the presence of this
glucose analogue in the medium at pH 7.0 (Fig 4.16, top). The growth rates were very
similar to those in the absence of glucose (based on visual inspection). The fact that the
slr1736 mutant was able to grow similarly to wild type in the presence of this glucose
analog suggests that the cause of the glucose toxicity is not the presence of glucose per se
that accumulates inside the cells. The presence of 5 mM pyruvate, succinate, or fumarate
in growth medium at pH 7.0 caused the tocopherol mutant to grow slightly slower yet
very similarly to wild type (Fig 4.16, top). These results strongly suggest that the
causative agent for the glucose toxicity at pH 7.0 in the tocopherol mutants is likely to be
metabolites accumulated either during glucose metabolism through the glycolytic
pathway or the pentose phosphate pathway. The glucose toxicity was observed when
cells were grown at RT (ca. 25 °C). The growth experiments described so far were
conducted under constant pH values; in other words, the cells were grown at pH 7.0 in
the absence of glucose before being transfer to medium containing glucose at pH 7.0 in
order to assess the effect of glucose on the growth of these cells. Likewise, the cells were
grown at pH 8.0 in the absence of glucose before transferred to medium containing
glucose at pH 8.0. Fig 4.16 (bottom) shows a similar, yet somewhat different experiment.
In this experiment, the cells were initially grown at pH 8.0 in the absence of glucose and
then were transferred to medium containing glucose at pH 7.0. The purpose of this

151
experiment was to test whether the incubation at pH 7.0 (in the absence of glucose)
“sensitizes” the tocopherol mutant cells to glucose or its by-product. The results showed
that the tocopherol mutants were unable to grow after transfer from medium at pH 8.0
(containing no glucose) to medium containing glucose at pH 7.0. This suggested that the
pH value of the medium before transfer to medium containing glucose has nothing to do
with the observed glucose toxicity in the tocopherol mutant cells.
Photosynthetic Activity
Fig 4.17 shows the activities of PS II measured for cells grown at various pH
values in the absence and the presence of glucose. In the absence of glucose, PS II
activities of the slr1736 cells (filled squares) were higher those that of wild type (filled
circles) at all pH values. Although the reason for this higher activity for the slr1736
mutant is not known, this result is reproducible, as can be also seen in Fig 4.9. In the
presence of glucose, the PS II activities of the wild-type cells were higher than those in
the absence of glucose at all pH values (open circles). On the contrary, no PS II activity
was detectable for the slr1736 mutant at pH 6.8 and 7.0. Higher activity was observed as
the pH value was increased, and at pH 8.0 the PS II activity was virtually
indistinguishable from that of wild type. Therefore, the inactivation of the PS II is also
dependent on the presence of glucose, the pH of the surrounding environment, and the
absence of tocopherols.
The PBP contents
When the cells were grown at pH 7.0 in the absence of glucose and with 1% CO2
(v/v), the PBP contents of the wild type and the “authentic” tocopherol mutants were

152
virtually indistinguishable (Fig 4.18A). When cells were grown in the presence of
glucose at pH 7.0, however, the PBP contents in the tocopherol mutants decreased to
about one-third of the wild type level (Fig 4.18A). This is consistent with the results
described in Fig 4.12, and demonstrates that the tocopherol mutants are experiencing
macronutrient starvation in response to glucose at pH 7. The effect of pH on the PBP
content was analyzed for the wild-type and the slr1736 mutant cells grown in the
presence of glucose (Fig 4.18B). At all pH values tested, the PBP content in the wild-type
cells remained constant. The PBP content in the slr1736 mutant cells was significantly
lower, less than 40 % of the wild-type level at pH 7.0 and below, which confirms the
reproducibility of the pH-dependent macronutrient starvation response in the tocopherol
mutant as described above. As the pH of the medium increased, higher PBP contents
were detected. At pH 8.0, the PBP content of the slr1736 mutant was 70% of the wild-
type level. The observed reduction of the PBP content over a wide range of pH values
strongly indicates that the presence of glucose alone induces a macronutrient starvation
response in the tocopherol mutants and that the macronutrient starvation response is
severely exacerbated at pH values below 7.2.

153
DISCUSSION
In this study, it was determined that the previously reported Synechocystis sp.
PCC 6803 mutants of genes slr1736, sll0418, and slr0089, encoding homogentisate
phytyltransferase, MPBQ methyltransferase and γ-tocopherol methyltransferase,
respectively, are probably genotypically heterogeneous mixtures of secondary suppressor
mutants. These mutants with defects in tocopherol biosynthesis were originally selected
and maintained in the presence of glucose, and although the targeted genes were
completely disrupted, a variety of colony growth morphologies have now been observed,
which is consistent with genetic heterogeneity. When these mutant lines were
reconstructed by a second round of transformation, followed by selection and
maintenance in the absence of glucose, the resulting mutant lines (here denoted as
“authentic” mutants to distinguish them from the original mutants previously selected in
the presence of glucose) were also found to be completely disrupted at each respective
locus, but the observed colony morphology and growth characteristics were now uniform.
It was found that the “authentic” mutants are highly sensitive to the presence of glucose
at pH values less than 7.2, and that the growth of each mutant strain ceased within 24 h
following transfer to medium containing glucose under those conditions. These results
clearly show that α-tocopherol is conditionally essential for the survival of Synechocystis
sp. PCC 6803 in the presence of glucose. It is likely that the selection and culturing of the
original transformants in the presence of glucose caused the selection of secondary
suppressor mutations that permitted survival under these otherwise suboptimal or lethal
conditions. Although the exact nature of the suppressor mutations has yet to be

154
determined, one can propose a few possibilities that enable the glucose-sensitive cells to
become glucose tolerant: inactivation of the glucose transporter, alteration of flux through
the glycolytic and/or oxidative pentose phosphate pathways, or the alteration of the
regulatory pathway(s) that controls glucose metabolism (Beuf et al., 1994, Hihara et al.,
1997, also see below).
The glucose toxic phenotype was found to be dependent on the pH of medium and
occurs at pH ~7.2 or lower. PS II inactivation and the reduction of PBP contents were
also found to be pH-dependent when glucose is present in the medium and α-tocopherol
synthesis is interrupted. Similar pH-dependent growth defects and reduction of
photosynthetic activities were observed for the cotA mutant (Katoh et al., 1996). CotA is
involved in light-induced proton extrusion, and its absence has been shown to lead to
severe growth defects at pH 7.0 or lower. Therefore, it is possible that the glucose
toxicity observed for the “authentic” tocopherol mutants are related to defects in proton
extrusion. However, a simple comparison between the cotA mutant and the tocopherol
mutants is not appropriate, because of the significant phenotypic differences between
them. The cotA mutant shows pH-dependent growth defects in the absence of glucose,
whereas the tocopherol mutant does not under the same conditions. Growth rate of the
cotA mutant is slower than wild type (60-70% of the wild-type level) at pH 8.0 in the
absence of glucose, whereas the tocopherol mutants grow similarly to wild type under the
same conditions (visual inspection). Therefore, the pH-dependent glucose toxicity of the
tocopherol mutants seems to be very complex and involves one or more factors that are
associated with the pH homeostasis of the cells. Further studies are required to resolve

155
the molecular mechanism of the pH-dependent glucose toxicity in the tocopherol
mutants.
Trebst and co-workers reported that the herbicide-mediated loss of α-tocopherol
led to the complete loss of PS II activity with concomitant degradation of the D1 protein
following high light treatment in the green alga Chlamydomonas reinhardtii (Trebst et
al., 2002). This was certainly not the case in the tocopherol mutants of Synechocystis sp.
PCC 6803, as the level of the D1 protein remained similar to the wild type even when the
PS II activity was completely lost. The growth light intensity (50 µE m-2 s-1) used in this
study was less than one half of the saturating intensity, and this value seems unlikely to
promote a photoinhibitory oxidative stress. In fact, the glucose toxicity observed for these
mutants is light-independent, since it occurs over a wide range of light intensity, from < 5
to 300 µE m-2 s-1, as described in the Results. Given that the transcript level of the sodB
gene is largely unaffected in the mutants under these lethal conditions, it is highly
unlikely that the inactivation of PS II and the death of the cells are directly associated
with an oxidative stress. Furthermore, the deleterious effects of glucose on the slr1736,
sll0418, and slr0089 mutants were virtually indistinguishable despite the varying
compositions and contents of tocopherols in each mutant. Should α-tocopherol function
solely as an antioxidant, an increasing susceptibility to glucose with decreasing
tocopherol content would reasonably be expected among the various tocopherol mutants,
but this is not observed. Therefore, it appears most likely that α-tocopherol plays a role
other than, or at least in addition to, that as a bulk antioxidant in the survival of
Synechocystis sp. PCC 6803 in the presence of glucose. Taking all of these considerations

156
together, it was concluded that the observed metabolic perturbation, PS II inactivation,
and cell death of the tocopherol mutants grown in the presence of glucose at pH values
below 7.2 are unlikely to be directly associated with the antioxidant activity of α-
tocopherol in Synechocystis sp. PCC 6803. Moreover, it appears most likely that α-
tocopherol plays a role other than as a bulk antioxidant in the survival of Synechocystis
sp. PCC 6803 in the presence of glucose.
PBPs are the major constituent of the light harvesting antennae in cyanobacteria,
and they can constitute nearly half of the total soluble protein (Grossman et al., 1994).
When cyanobacteria are subjected to macronutrient deprivation conditions for carbon,
nitrogen, phosphorus, sulfur, and iron, the synthesis of PBPs ceases and their active
degradation begins (Allen and Smith, 1969; Ihlenfeldt and Gibson, 1975; Foulds and
Carr, 1977; Yamanaka and Glazer, 1980; Wood and Haselkorn 1980; Stevens et al.,
1981; Schmidt et al., 1982; Sherman and Sherman, 1983; Jensen and Rachlin, 1984;
Elmorjani and Herdman, 1987). It is generally believed that the degradation of these
proteins provides amino acids for energy production and maintenance protein synthesis,
while reducing the potential for photooxidative stress due to light stress at PS II.
Therefore, it is highly likely that the observed, dramatic decrease in the PBP contents in
the “authentic” tocopherol mutants during growth in the presence of glucose represents a
response to a perceived state of nutritional deprivation. Supporting this interpretation is
the observation that transcription of the nblA gene, known to accumulate in response to
nitrogen deprivation (Richaud et al., 2001), also accumulated in the tocopherol-deficient
mutant in response to glucose. The combined results hence suggest that the complete loss

157
of, or a substantial decrease of, α-tocopherol perturbs metabolic processes related to
nitrogen, carbon, and sulfur, and that this disruption ultimately leads to phycobilisome
degradation and cell death.
Recent studies in mammalian systems have demonstrated the involvement of α-
tocopherol in modulating the production of signaling molecules and pathways (Chan et
al., 2001; Azzi et al., 2002; Ricciarelli et al., 2002). For example, α-tocopherol has been
shown to bind to phospholipase A2 specifically at the substrate-binding pocket and act as
a competitive inhibitor, thereby decreasing the release of arachidonic acid for eicosanoid
synthesis (Chandra et al., 2002). α-Tocopherol has also been suggested to modulate the
phosphorylation state of protein kinase Cα in rat smooth-muscle cells, possibly via
phosphorylation of protein phosphatase 2A (Ricciarelli et al., 1998). α-Tocopherol is also
directly involved in transcriptional regulation in animals, including the expression of the
genes encoding liver collagen αI, α-tocopherol transfer protein, and α-tropomyosin
collagenase (Yamaguchi et al., 2001; Azzi et al., 2002). Therefore, it seems possible that,
in addition to acting as a bulk lipid-soluble antioxidant, α-tocopherol could also play a
regulatory role in Synechocystis sp. PCC 6803, possibly in the regulation of metabolic
processes such as those involved in carbon and nitrogen metabolism.
Two glucose-tolerant mutants reported previously appear to be relevant to the
observations reported here. The pmgA (sll1986) and icfG (slr1680) loci have been shown
to be determinants for glucose sensitivity (Hihara and Ikeuchi, 1997; Beuf et al., 1994).

158
Inactivation of the pmgA locus has been shown to result in a glucose-sensitive phenotype
and enhanced photoautotrophic growth capability (Hihara and Ikeuchi, 1997).
Inactivation of the icfG locus, on the other hand, results in a glucose-sensitive phenotype
under low CO2 conditions (Beuf et al., 1994). The “authentic” tocopherol mutants, whose
phenotypes are characterized in this chapter, are glucose-sensitive both under high and
low CO2 environments but only at pH values less than 7.2. The amino acid sequences
deduced from the pmgA and icfG coding regions show weak similarity to RsbT/W (a
serine/threonine protein kinase) and RsbU/X (a protein phosphatase and a negative
regulator of sigma factor B) in Bacillus subtilis. These proteins form a signal transduction
network with other Rsb proteins and regulate the activity of sigma factor B through
phosphorylation/dephosphorylation interactions upon perception of environmental stress
(Price 2000; Woodbury et al., 2004). Homologous protein sequences of other Rsb
proteins are also found in the Synechocystis sp. PCC 6803 genome: Slr2031 (RsbU/X),
Ssr1600 (RsbS/V), Slr1856 (RsbS/V), and Slr1861 (RsbT/W). It is therefore plausible
that an Rsb-like pathway is present in this organism and may participate in the regulation
of glucose metabolism. In fact, studies have shown that recombinant Slr1861
phosphorylates Slr1856 and IcfG dephosphorylates the phosphorylated Slr1856 (Shi et
al., 1999). Whether the role of α-tocopherol is related to this hypothetical pathway is an
open question and is a subject of continuing investigation.

159
SUMMARY
The tocopherol mutants (slr1736, sll0418, and slr0089), which had been
previously isolated in the presence of glucose, were shown to be phenotypically and
genotypically heterogeneous populations. Newly isolated “authentic” tocopherol mutants
were glucose-sensitive and were not able to grow in the presence of glucose after 24 h.
This glucose toxicity was shown to occur when the mutants were grown at pH values
below 7.2. Under these conditions, the PS II activities were significantly reduced or not
detected at all in the tocopherol mutants.

160
MATERIALS AND METHODS
Growth Conditions and Strains
The slr1736, sll0418, and slr0089 mutants were generated by targeted insertion
with the aphII gene, which confers kanamycin resistance, as described previously
(Shintani and DellaPenna, 1998; Collakova and DellaPenna, 2001; Shintani et al., 2002).
The wild-type strain used for transformation and further analysis was a glucose-tolerant
strain of Synechocystis sp. PCC 6803 (Williams, 1988). Medium BHEPES, pH 8.0, was
used for selection, maintenance, and growth measurements of wild type and mutants.
This medium is prepared by supplementing BG-11 medium (Stainer et al., 1971) with 4.6
mM HEPES [4-(2-hydroxyethyl)piperazine-N’-(2-ethnesulfonic acid)]-KOH and 18 mg
L-1 ferric ammonium citrate. Medium BEHEPS40, modified medium BEHEPS after
supplementing with 40 mM HEPES for higher buffer strength, was used for pH-
dependent or pH-controlled growth analyses. Wild-type cells were maintained on solid B-
HEPES medium containing 1.5 % (w/v) agar and 5 mM glucose, and the “authentic”
tocopherol mutants were maintained on solid medium BHEPES containing 1.5 % (w/v)
agar, 50 µg kanamycin ml-1, and importantly, no glucose. Prior to all growth
measurements and preparations for subsequent analyses, cells were transferred from solid
to liquid medium BHEPES and allowed to grow exponentially for several generations in
the absence of glucose. For determination of growth characteristics, late-exponential
phase cultures were diluted in fresh liquid medium BHEPES to an OD730 nm of
approximately 0.05 cm-1. The diluted cultures were grown at 32 °C with continuous

161
bubbling with air containing 1% or 3% (v/v) CO2. Growth was monitored by measuring
the optical density at 730 nm. For growth in the presence of glucose, the medium was
supplemented with 5 mM glucose. The growth light intensity was 50 µE m-2 s-1, unless
otherwise noted. For growth measurements in the presence of inhibitors of the electron
transport chain, the following concentrations were used: 0.03 µM DCMU (3-(3,4-
dichlorophenyl)-1,1-dimethyl-urea); 2, 20, or 200 µM potassium cyanide. Pre-
conditioned medium was prepared from a culture of an authentic tocopherol mutant that
had been grown in the presence of glucose for 72 h. The cells were removed by
centrifugation at 8000 × g, and the supernatant was filtered through a sterile 0.22 µm
pore-size membrane. The filtrate was then mixed with five-fold strength medium
BHEPES in a four-to-one ratio (v/v) to achieve a normal, one-fold-strength medium.
Isolation of Tocopherol Mutants
Synechocystis sp. PCC 6803 wild type was transformed with genomic DNA
extracted from the slr1736, sll0418, and slr0089 mutants selected in the presence of
glucose as previously described (Williams, 1988). Segregation of mutant alleles from the
wild type allele was carried out in the absence of glucose and in the presence of 50 µg
kanamycin ml-1. Oligonucleotide primers used for PCR analysis were as follows; slr1736
forward primer (5’-GGCTTCTCCTACCCGGAATTCTACTTCCTG-3’), slr1736 reverse
primer (5’-GCTTTCTAAGTGTACATCTAGACTCCGCCA-3’), sll0418 forward primer
(5’-ATGCCCGAGTATTTGCTTCTGCC-3’), sll0418 reverse primer (5’-
GCACTGCTTTGAACATACCGAAG-3’), slr0089 forward primer (5’-

162
TCTACCGGAAATTGCCAACTACCA-3’), and slr0089 reverse primer (5’-
CCTAGGAGATTGTGGACTTCAA-3’).
Oxygen Evolution and Consumption Measurements
Cells grown in the absence and in the presence of glucose for 24 h were harvested
by centrifugation at 8,000 × g at room temperature and resuspended in a 25-mM HEPES
buffer, pH 7.0, to obtain an optical density of 1.0 cm-1 at 730 nm. For whole-chain
oxygen evolution and consumption measurements, the cell suspensions were incubated in
the dark in the presence of 10 mM NaHCO3 for at least 15 min prior to measurements.
For the Photosystem II (PS II)-dependent oxygen evolution measurements, 1 mM 1,4-
benzoquinone and 0.8 mM K3Fe(CN)6 were added instead of NaHCO3 and measurements
were performed immediately. The excitation light intensity was approximately 3 mE m-2
s-1. The oxygen concentration was measured polarographically with a Clark-type
electrode as previously described (Sakamoto and Bryant, 1998).
Quantification of Pigments and Quinones by High Performance Liquid Chromatography
Chl a in whole cells was extracted with methanol, and its molar content per unit
optical density at 730 nm was determined using the molar absorption coefficient 82 L g-1
cm-1 (MacKinney, 1941). For quinone analyses, cells were harvested by centrifugation at
8,000 × g, at 4 °C for 6 min, washed in 1 ml of 50 mM Tris-HCl buffer, pH 8.0, and
centrifuged at 18,000 × g for 1 min to obtain tight cell pellets. Pigments and quinones
were extracted by ultrasonication in 400 µl of cold acetone-methanol (7:2, v/v) in the

163
dark. High performance liquid chromatography (HPLC) analysis was performed with a
Agilent 1100 system equipped with a diode array detector (Agilent Technologies,
Wilmington, DE, USA). One hundred microliters of extract was mixed with 10 µl of 1 M
ammonium acetate and injected onto a NovaPak C18 column (3.9 × 300 mm, 4 µm
packing, Waters, Milford, MA). Analytes were eluted by a linear gradient program
consisting of solvent A (100% methanol) and solvent B (100% isopropanol) using the
following protocol: [100% A] for 10 min, [100% A] to [20% A : 80% B] in 20 min, [20%
A: 80% B] for 5 min, and [20% A : 80% B] to [100% A] for 5 min. The flow rate was
0.75 ml min-1. The Chl a, phylloquinone, and plastoquinone contents of each sample
were determined based on integrated peak areas and their molar absorption coefficients,
which are 17.4 mM-1 cm-1 at 618 nm, 18.9 mM-1 cm-1 at 270 nm (Dunphy and Brodie,
1971), and 15.2 mM-1 cm-1 at 254 nm (Crane and Dilley, 1963), respectively. The
absorption coefficient of Chl a at 618 nm was calculated from the ratio of the absorption
peaks at 618 nm and 666 nm in methanol-isopropanol (6:4, v/v) and from its absorption
coefficient at 666 nm in methanol (MacKinney, 1941). The quinone content per unit
optical density at 730 nm was calculated by multiplying the molar ratio of quinone to Chl
a by the molar Chl a content per optical density at 730 nm.
Estimation of Relative Content of Phycobiliproteins
Cells were harvested by centrifugation at 8000 × g for 6 min, and pellets were
resuspended in the 25 mM HEPES buffer, pH 7.0, to obtain 2 ml of the cell suspensions
with an optical density of 0.5 cm-1 at 730 nm. One milliliter of this suspension was heated

164
at 100 °C for 1 min. A modification was made in the method previously described (Zhao
and Brand, 1989) for estimation of the relative PBP content. The absorbance at 635 nm
and 730 nm were recorded for unheated and heated samples, and the data were then
inserted into the following equation: the relative PBP content = (∆OD635nm –
∆OD730nm)/OD730nm·unheated, where ∆ indicates ODunheated sample minus ODheated sample
SDS-Polyacrylamide Gel Electrophoresis and Immunoblotting of the D1 protein
Cells were grown in the absence of glucose to mid-exponential phase or in the
presence of glucose for 24 h, and harvested by centrifugation at 8,000 × g at 4 °C for 6
min. The resulting cell pellets were immediately frozen in liquid N2 and stored at –80 °C
until being used. Frozen cells were thawed on ice and resuspended in the 25 mM HEPES
buffer, pH 7.0, at an optical density of 100 cm-1 at 730 nm. An equal volume of glass
beads was added to the cell suspension, which was allowed to stand on ice for 5 min.
Using a home-built bead-beater, the samples were vigorously shaken 4 times for 30 s at 4
°C with 30 s intervals on ice. An aliquot (10 µl) of each sample was mixed with an equal
volume of loading buffer, and the mixture was incubated at 65 °C for 20 min, and applied
onto a discontinuous SDS-polyacrylamide gel with 10% (w/v) acrylamide in the
separating gel as described (Schägger and van Jagow, 1987). Electrophoresis was carried
out at 50 V for 12 h. The antibodies raised against the amino acids 234-242 of the D1
protein of Synechocystis sp. PCC 6803 were a kind gift from Prof. Eva-Mari Aro of the
University of Turku, Finland. The antibodies raised against the PsbO protein of
Synechocystis sp. PCC 6803 were a kind gift from Prof. Robert Burnap of the Oklahoma

165
State University, USA. After electrophoresis, proteins were transferred to a nitrocellulose
membrane. Proteins were detected by immunoblotting by using an enhanced
chemiluminescence reagents and analysis system (Amersham Biosciences, Piscataway,
NJ) according to manufacturer’s specifications.
Isolation of total RNA and RT-PCR analyses
Total RNA was isolated from cells using the Mini-to-Midi RNA isolation kit
(Invitrogen Corp., Carlsbad, CA) followed by an RNase-free DNase treatment, and
repurification by using the same kit. The RNAs obtained were adjusted to a final
concentration of 50 ng mL-1 in RNase free double distilled water and store at -80°C until
being used. RT-PCR reactions were carried out by using the One-Step RT-PCR kit
(Qiagen Inc., Valencia, CA) for detecting the rnpB transcript. The reaction volume was
25 µL, and 4 ng µL-1 of the total RNAs was used. For detecting the nblA and sodB
transcripts, 400 ng of total RNA was first converted to cDNA by the RT reaction using
M-MLV reverse transcriptase (Promega, Madison, WI). The reaction volume was 20 µL
for each sample. The resulting cDNA was diluted with 80 µL of TE buffer, and stored at
–20 °C until being used. Two microliters of cDNA stock were used for PCR (final
volume, 25 µL) using Hot-Start Taq polymerase (Qiagen Inc., Valencia, CA). RNasin
(Promega, Madison, WI), an RNase inhibitor, was present in one-step RT-PCR reactions
and during the cDNA synthesis.

166
ACKNOWLEDGEMENTS
The authors thank Prof. Eva-Mari Aro (University of Turku) for kindly providing the D1
antibodies, Prof. Robert Burnap (Oklahoma State University) for kindly providing the
PsbO antibodies, and Prof. Aaron Kaplan, Hebrew University for discussion, critical
comments, and suggestions.

167
REFERENCES
Alfonso M, Perewoska I, Kirilovsky D (2000) Redox control of psbA gene expression in
the cyanobacterium Synechocystis PCC 6803. Involvement of the cytochrome b6/f
complex. Plant Physiol 122: 505-515
Allakhverdiev SI, Nishiyama Y, Suzuki I, Tasaka Y, Murata N (1999) Genetic
engineering of the unsaturation of fatty acids in membrane lipids alters the tolerance of
Synechocystis to salt stress. Proc Natl Acad Sci USA 96: 5862-5867
Allen MM, Smith AJ (1969) Nitrogen chlorosis in blue-green algae. Arch Mikrobiol 69:
114-120
Aro E-M, Girgin T, Andersson B (1993) Photoinhibition of Photosystem II. Inactivation,
protein damage and turn over. Biochim Biophys Acta 1143: 113-134
Azzi A, Ricciarelli R, Zingg JM (2002) Non-antioxidant molecular functions of α-
tocopherol (vitamin E). FEBS Lett 519: 8-10
Beuf L, Bedu S, Durand MC, Joset F (1994) A protein involved in co-ordinated
regulation of inorganic carbon and glucose metabolism in the facultative
photoautotrophic cyanobacterium Synechocystis PCC 6803. Plant Mol Biol 25: 855-864
Chan SS, Monteiro HP, Schindler F, Stern A, Junqueira VB. (2001) α-Tocopherol
modulates tyrosine phosphorylation in human neutrophils by inhibition of protein kinase
C activity and activation of tyrosine phosphatases. Free Radic Res 35: 843-856

168
Chandra V, Jasti J, Kaur P, Betzel C, Srinivasan A, Singh TP (2002) First structural
evidence of a specific inhibition of phospholipase A2 by α-tocopherol (vitamin E) and its
implications in inflammation: crystal structure of the complex formed between
phospholipase A2 and α-tocopherol at 1.8 Å resolution. J Mol Biol 320: 215-222
Cheng Z, Sattler S, Maeda H, Sakuragi Y, Bryant DA, DellaPenna D (2003) Highly
divergent methyltransferases catalyze a conserved reaction in tocopherol and
plastoquinone synthesis in cyanobacteria and photosynthetic eukaryotes. Plant Cell 15:
2343-2356
Collins MD, Jones D (1981) Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiol Rev 45: 316-354
Collakova E, DellaPenna D (2001) Isolation and functional analysis of homogentisate
phytyltransferase from Synechocystis sp. PCC 6803 and Arabidopsis. Pant Physiol 127:
1-12
Collakova E, DellaPenna D (2003) Homogentisate phytyltransferase activity is limiting
for tocopherol biosynthesis in Arabidopsis. Plant Physiol 131: 632-642
Cooley JW, Vermaas WFJ (2001) Succinate dehydrogenase and other respiratory
pathways in thylakoid membranes of Synechocystis sp. strain PCC 6803: capacity
comparisons and physiological function. J Bacteriol 183: 4251-4258
Crane FL, Dilley RA (1963) Determination of coenzyme Q (ubiquinone). Methods
Biochem Anal 11: 279-306

169
d’Harlingue A, Camara B (1985) Plastid enzymes of terpenoid biosynthesis. J Biol Chem
260: 15200-15203
Dähnhardt D, Falk J, Appel J, van der Kooij TAW, Schulz-Friedrich R, Krupinska K
(2002) The hydroxyphenylpyruvate dioxygenase from Synechocystis sp. PCC 6803 is not
required for plastoquinone biosynthesis. FEBS Lett 523: 177-181
Dunphy PJ, Brodie AF (1971) The structure and function of quinones in respiratory
metabolism. Methods Enzymol 18: 407-461
Elmorjani K, Herdman M (1987) Metabolic control of phycocyanin degradation in
cyanobacterium Synechocystis PCC 6803: a glucose effect. J Gen Microbiol 133: 1685-
1694
Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S (2004) Architecture of the
photosynthetic oxygen-evolving center. Science 303: 11831-1838
Foulds IJ, Carr NG (1977) A proteolytic enzyme degrading phycocyanin in
cyanobacterium Anabaena cylindrica. FEMS Microbiol Lett 2: 117-119
Fryer MJ (1992) The antioxidant effects of thylakoid vitamin E (α-tocopherol). Plant Cell
Environ 15: 381-392
Grossman AR, Schaefer MR, Chiang GG, Collier JL (1994) The responses of
cyanobacteria to environmental conditions, light and nutritions. In: Bryant DA (ed), The
Molecular Biology of Cyanobacteria. pp 641-675. Kluwer, Dordrecht

170
Hideg E, Kalai T, Hideg K, Vass I (2000) Do oxidative stress conditions impairing
photosynthesis in the light manifest as photoinhibition? Phil Trans R Soc Lond B 355:
1511-1516
Hihara Y, Ikeuchi M (1997) Mutation in a novel gene required for photomixotrophic
growth leads to enhanced photoautotrophic growth of Synechocystis sp. PCC 6803.
Photosynth Res 53: 243-252
Ihlenfeldt MJA, Gibson J (1975) Phosphate utilization and alkaline phosphatase activity
in Anacystis nidulans (Synechococcus). Arch Microbiol 102: 23-28
Jensen TE, Rachlin JW (1984) Effect of varying sulfur deficiency on structural
components of a cyanobacterium Synechococcus leopoliensis: a morphometric study.
Cytobios 41: 35-46
Kamal-Eldin A, Appelqvist L (1996) The chemistry and antioxidant properties of
tocopherols and tocotrienols. Lipids 31: 671-701
Katoh A, Sonoda M, Katoh H, Ogawa T (1996b) Absence of light-induced proton
extrusion in a cotA-less mutant of Synechocystis sp. strain PCC 6803. J Bacteriol 178:
5452-5455
Koch M, Lemke R, Heise K, Mock H (2003) Characterization of α-tocopherol
methyltransferase from Capsicum annuum L and Arabidopsis thaliana. Eur J Biochem
270: 84-92

171
Li H, Sherman L (2000) A redox-responsive regulator of photosynthesis gene expression
in the cyanobacterium Synechocystis sp. PCC 6803. J Bacteriol 182: 4268-4277
MacKinney G (1941) Absorption of light by chlorophyll solutions. J Biol Chem 140:
315-322
Miller LS, Holt SC (1977) Effect of carbon dioxide on pigment and membrane content in
Synechococcus lividus. Arch Microbiol 115: 185-198
Munné-Bosch S, Leonor A (2000) The function of tocopherols and tocotrienols in plants.
Critic Rev Plant Sci 21: 31-57
Norris SR, Shen X, DellaPenna D (1998) Complementation of the Arabidopsis pds1
mutation with the gene encoding p-hydroxyphenylpyruvate dioxygenase. Plant Physiol
117: 1317-1323
Porfirova S, Bergmüller E., Tropf S, Lemke R, Dörmann P (2002) Isolation of an
Arabidopsis mutant lacking vitamin E and identification of a cyclase essential for all
tocopherol biosynthesis. Proc Natl Acad Sci USA 99: 12495-12500
Price CW (2000) Protective function and regulation of the general stress response in
Bacillus subtilis and related Gram-positive bacteria. In: Storz G, Hengge-Aronis R (eds)
Bacterial Stress Response. pp 179-197. ASM Press, Washington DC.
Ricchiarelli R, Tasinato A, Clément S, Özer NK, Boscoboinik D, Azzi A (1998) α-
Tocopherol specifically inactivates cellular protein kinase Cα by changing its
phosphorylation state. Biochem J 334: 243-249

172
Ricciarelli R, Zingg JM, Azzi A (2002) The 80th anniversary of vitamin E: Beyond its
antioxidant properties. Biol Chem 383: 457-465
Richaud C, Zabulon G, Joderr A, Thomas JC (2001) Nitrogen and sulfur starvation
differentially affects phycobilisomes degradation and expression of the nblA gene in
Synechocystis strain PCC 6803. J Bacteriol 183: 2989-2994
Sakamoto T, Bryant DA (1998) Growth at low temperature causes nitrogen limitation in
the cyanobacterium Synechococcus sp. PCC 7002. Arch Microbiol 169: 10-19
Sattler SE, Cahoon EB, Coughlan SJ, DellaPenna D. (2003) Characterization of
tocopherol cyclases from higher plants and cyanobacteria: evolutionary implications for
tocopherol synthesis and function. Plant Physiol 132: 2184-2195
Savidge B, Weiss JD, Wong YH, Lassner NW, Mitsky TA, Shewmaker CK, Post-
Beittenmiller D, Valentin HE (2002) Isolation and characterization of homogentisate
phytyltransferase genes from Synechocystis sp. PCC 6803 and Arabidopsis. Plant Physiol
129: 321-332
Schledz M, Seidler A, Beyer P, Neuhaus G (2001) A novel phytyltransferase from
Synechocystis sp. PCC 6803 involved in tocopherol biosynthesis. FEBS Lett 499: 15-20
Schmidt A, Erdle I, Kost H. (1982) Changes of C-phycocyanin in Synechococcus 6301 in
relation to growth on various sulfur compounds. Z Naturforsch Teil C 37: 870-876

173
Schägger H, van Jagow G (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal
Biochem 166: 368-379
Sherman DM, Sherman LA (1983) Effect of iron deficiency and iron restoration on
ultrastructure of Anacystis nudulans. J Bacteriol 156: 393-401
Shi L, Bischoff KM, Kennelly PJ (1999) The icfG gene cluster of Synechocystis sp. strain
PCC 6803 encodes an Rsb/Spo-like protein kinase, protein phosphatase, and two
phosphoproteins. J Bacteriol 181: 4761-4767
Shintani D, Cheng Z, DellaPenna D (2002) The role of 2-methyl-6-phytyl-benzoquinone
methyltransferase in determining tocopherol composition in Synechocystis sp. PCC 6803.
FEBS Lett 511: 1-5
Shintani D, DellaPenna D (1998) Elevating the vitamin E content of plants through
metabolic engineering. Science 282: 2098-2100
Soll J, Kemmerling M, Schultz G. (1980) Tocopherol and plastoquinone synthesis in
spinach chloroplasts subfractions. Arch Biochem Biophys 204: 544-550
Soll J, Schultz G, Joyard J, Douce R, Block MA (1985) Localization and synthesis of
prenylquinones in isolated outer and inner envelope membranes from spinach
chloroplast. Arch Biochem Biophys 238: 290-299
Stainer RY, Kunisawa R, Mandel M, Cohen-Bazire G (1971) Purification and properties
of unicellular blue-green algae (Order Chroococcales). Bact Rev 35: 171-205

174
Stevens SE, Blkwill DL, Paone DAM (1981) The effects of nitrogen limitation on the
ultrastructure of the cyanobacterium Agmenellum quadruplicatum. Arch. Microbiol.
130:204-212
Tabita R (1987) Carbon dioxide fixation and its regulation in cyanobacteria. In: Fay P,
Baalen G (eds), The Cyanobacteria. pp 95-117. Elsevier Science Publishers, Amsterdam
Threlfall DR, Whistance GR (1971) Biosynthesis of isoprenoid quinones and
chromanols. In: Goodwin TW (ed) Aspects of Terpenoid Chemistry and Biochemistry,
pp 357-404. Academic Press, London
Trebst A, Depka B, Höllander-Czytko H (2002) A specific role for tocopherol and of
chemical singlet oxygen quenchers in the maintenance of Photosystem II structure and
function in Chlamydomonas reinhardtii. FEBS Lett 516: 156-160
Ushimaru T, Nishiyama Y, Hayashi H, Murata N (2002) No coordinated transcriptional
regulation of the sod-kat antioxidative system in Synechocystis sp. PCC 6803. J Plant
Physiol 159: 805-807
Wanner G, Henkelmann A, Schmidt A, Kost H (1986) Nitrogen and sulfur starvation of
the cyanobacterium Synechococcus 6301. An ultrastructural, morphological and
biochemical comparison. Z Naturforsch Teil C 41: 741-750
Williams JGK (1988) Construction of specific mutations in Photosystem-II
photosynthetic reaction center by genetic engineering methods in Synechocystis 6803.
Methods Enzymol 167: 766-778

175
Wood NB, Haselkorn RH (1980) Control of phycobiliporotein proteolysis and heterocyst
differentiation in Anabaena. J Bacteriol 141: 1375-1385
Woodbury RL, Luo T, Grant L, Haldenwang WG (2004) Mutational analysis of RsbT, an
activator of the Bacillus subtilis stress response transcription factor, σB. J Bacteriol 186:
2789-2797
Yamaguchi J, Iwamoto T, Kida S, Masushige S, Yamada K, Esashi T (2001) Tocopherol-
associaated protein is a ligand-dependent transcriptional activator. Biochem Biophys Res
Commun 285: 295-299
Yamanaka G, Glazer AN (1980) Dynamic aspects of phycobilisome structure.
Phycobilisome turnover during nitrogen starvation in Synechococcus sp. Arch Microbiol
124: 39-47
Zhao J, Brand JJ (1989) Specific bleaching of phycobiliproteins from cyanobacteria and
red algae at high temperature in vivo. Arch Microbiol 152: 447-452
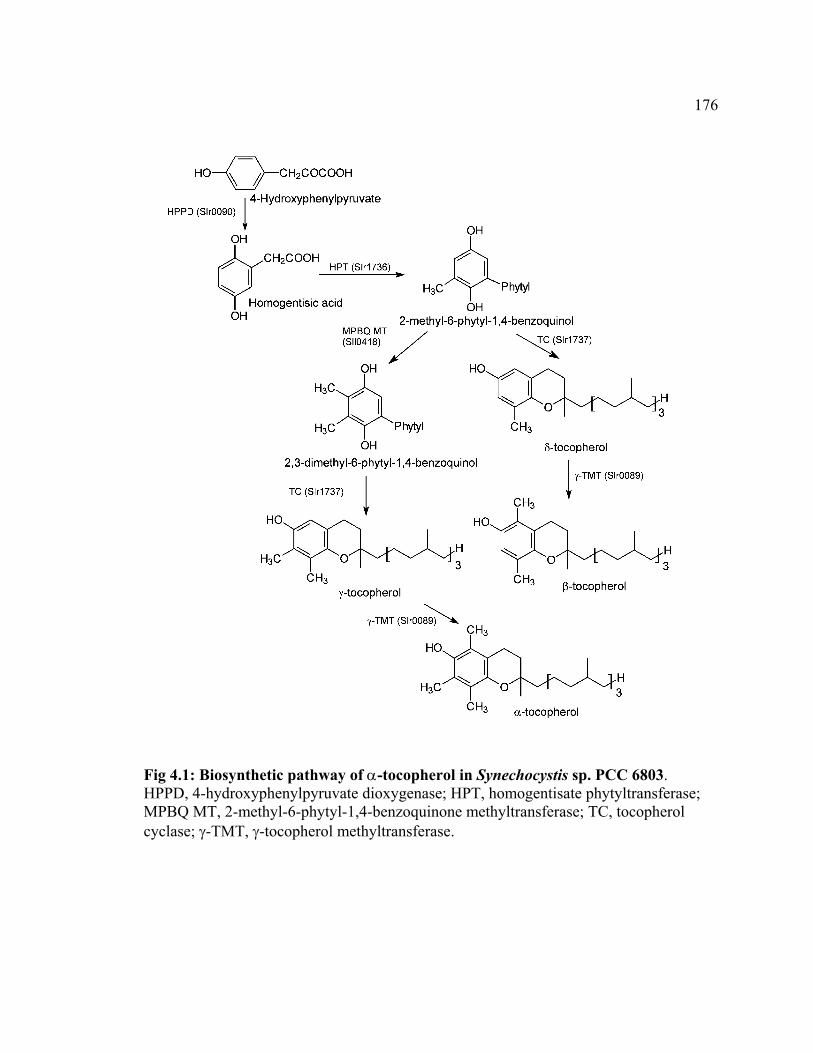
176
Fig 4.1: Biosynthetic pathway of α-tocopherol in Synechocystis sp. PCC 6803. HPPD, 4-hydroxyphenylpyruvate dioxygenase; HPT, homogentisate phytyltransferase; MPBQ MT, 2-methyl-6-phytyl-1,4-benzoquinone methyltransferase; TC, tocopherol cyclase; γ-TMT, γ-tocopherol methyltransferase.

177
Fig 4.2: Colony morphologies of three tocopherol mutants previously isolated in the presence of glucose. Cells were grown in the absence of glucose (A-D) and in the presence of glucose (E-H). Wild type (A, E), the slr1736 mutant (A, E), the sll0418 mutant (C, G), and the slr0089 mutant (D, H), on solid BHEPES medium.

178
Fig 4.3: Growth curves of wild type and cell lines derived from small (A, C) and large colonies (B, D) of tocopherol mutants selected in the presence of glucose (see Fig 4.2) at 32 °C, 50 µE m-2 s-1 with 3% (v/v) CO2. The data shown for each strain are averages of at least three independent cultures, and standard error bars are shown. Growth in the absence of glucose (A, B) and in the presence of glucose (C, D). (A, C) Wild type (filled circles) and a cell line derived from a small colony slr0089 mutant selected in the presence of glucose (open circles) (see Fig 4.2). (B, D) Wild type (filled circles) and cell lines derived from large colonies the slr0089 (open circles), sll0418 (open triangles), and slr1736 mutants (open squares) selected in the presence of glucose (see Fig 4.2).
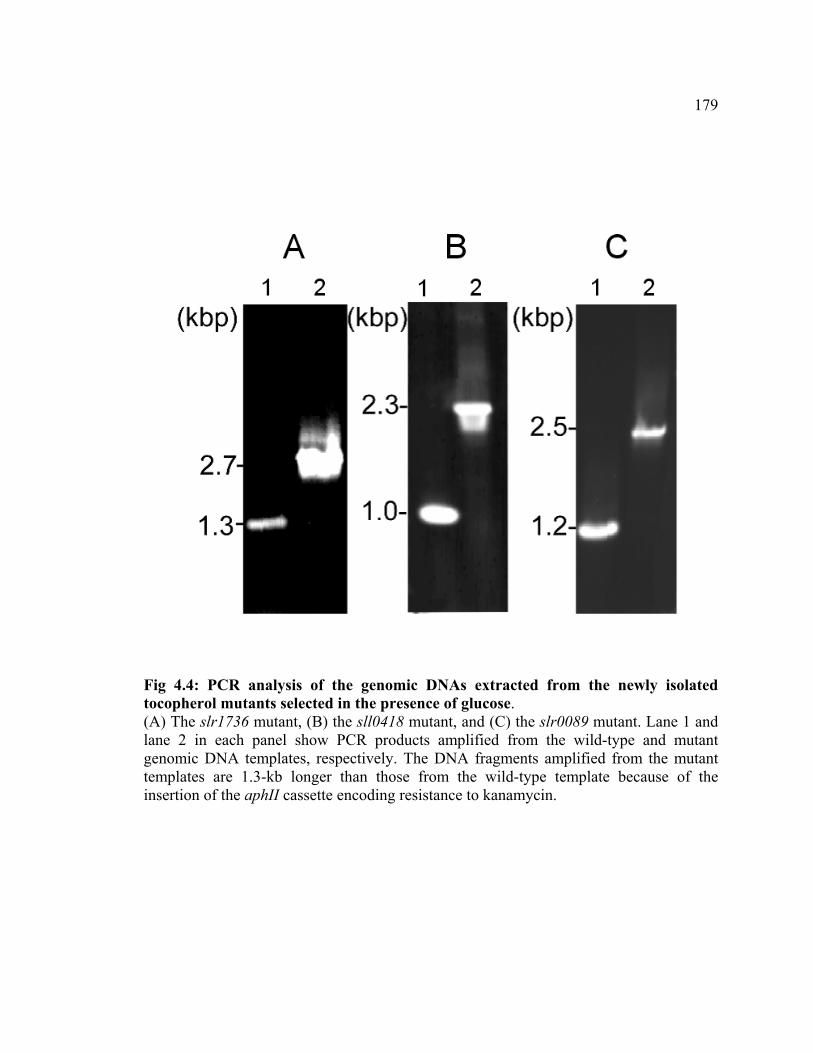
179
Fig 4.4: PCR analysis of the genomic DNAs extracted from the newly isolated tocopherol mutants selected in the presence of glucose. (A) The slr1736 mutant, (B) the sll0418 mutant, and (C) the slr0089 mutant. Lane 1 and lane 2 in each panel show PCR products amplified from the wild-type and mutant genomic DNA templates, respectively. The DNA fragments amplified from the mutant templates are 1.3-kb longer than those from the wild-type template because of the insertion of the aphII cassette encoding resistance to kanamycin.
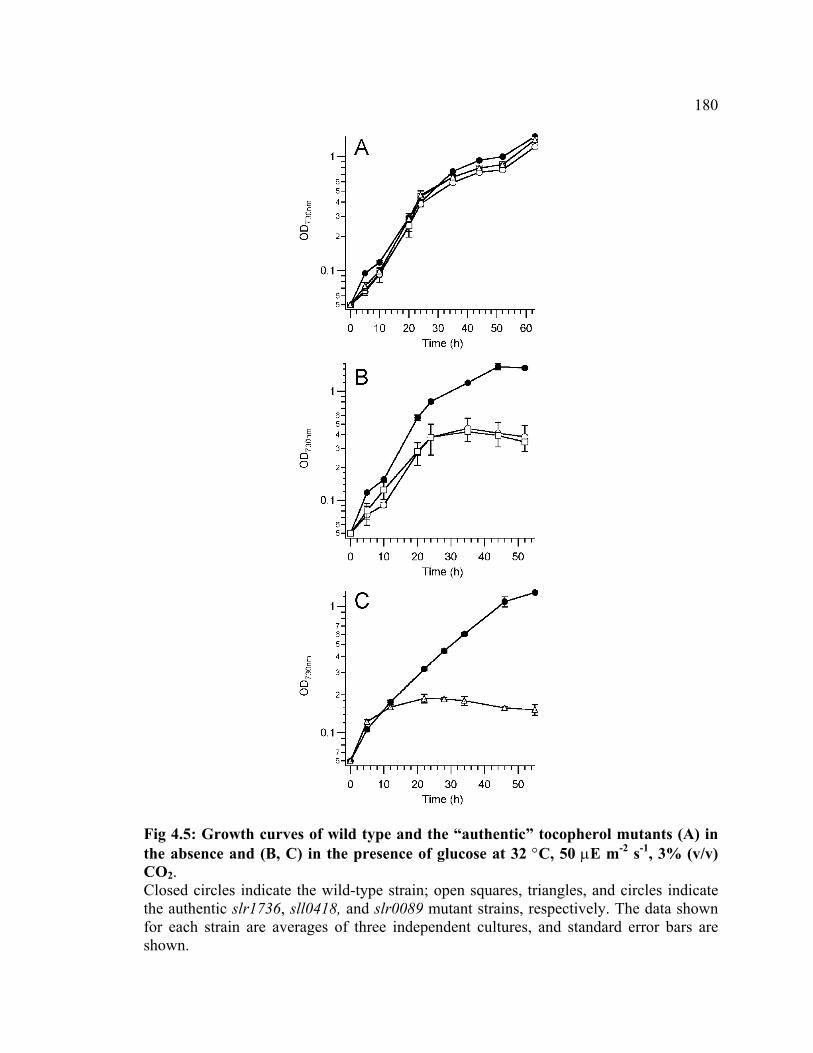
180
Fig 4.5: Growth curves of wild type and the “authentic” tocopherol mutants (A) in the absence and (B, C) in the presence of glucose at 32 °C, 50 µE m-2 s-1, 3% (v/v) CO2. Closed circles indicate the wild-type strain; open squares, triangles, and circles indicate the authentic slr1736, sll0418, and slr0089 mutant strains, respectively. The data shown for each strain are averages of three independent cultures, and standard error bars are shown.
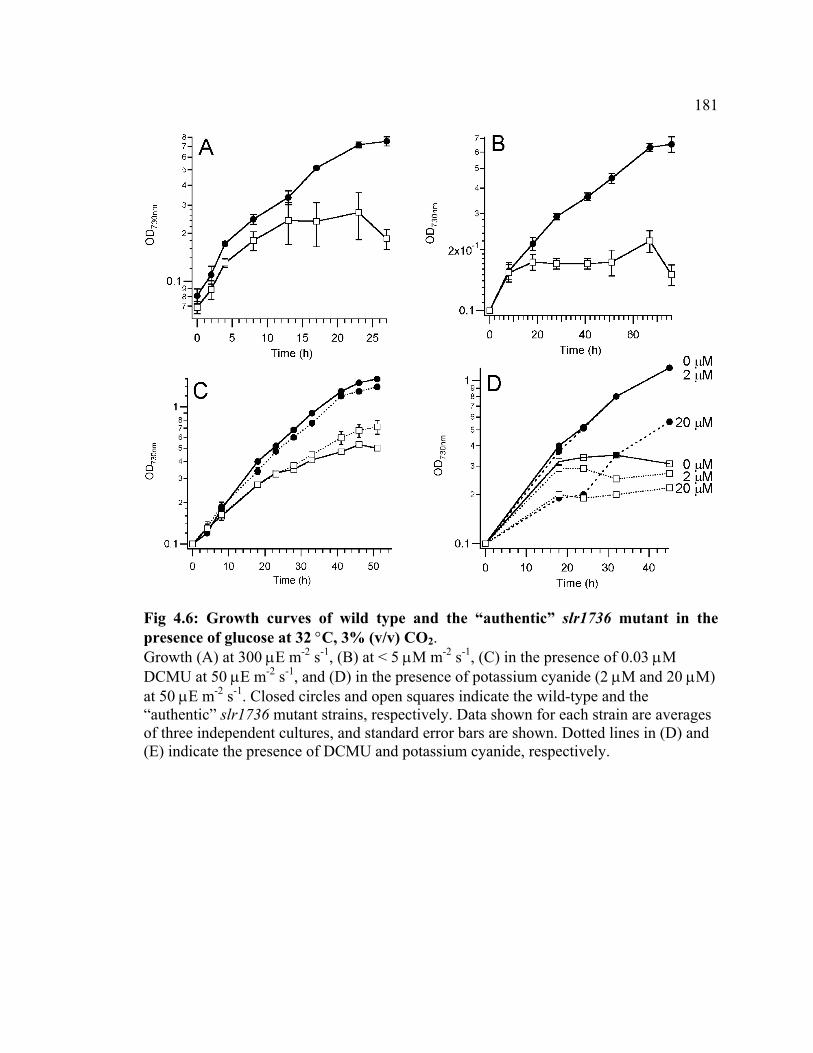
181
Fig 4.6: Growth curves of wild type and the “authentic” slr1736 mutant in the presence of glucose at 32 °C, 3% (v/v) CO2. Growth (A) at 300 µE m-2 s-1, (B) at < 5 µM m-2 s-1, (C) in the presence of 0.03 µM DCMU at 50 µE m-2 s-1, and (D) in the presence of potassium cyanide (2 µM and 20 µM) at 50 µE m-2 s-1. Closed circles and open squares indicate the wild-type and the “authentic” slr1736 mutant strains, respectively. Data shown for each strain are averages of three independent cultures, and standard error bars are shown. Dotted lines in (D) and (E) indicate the presence of DCMU and potassium cyanide, respectively.
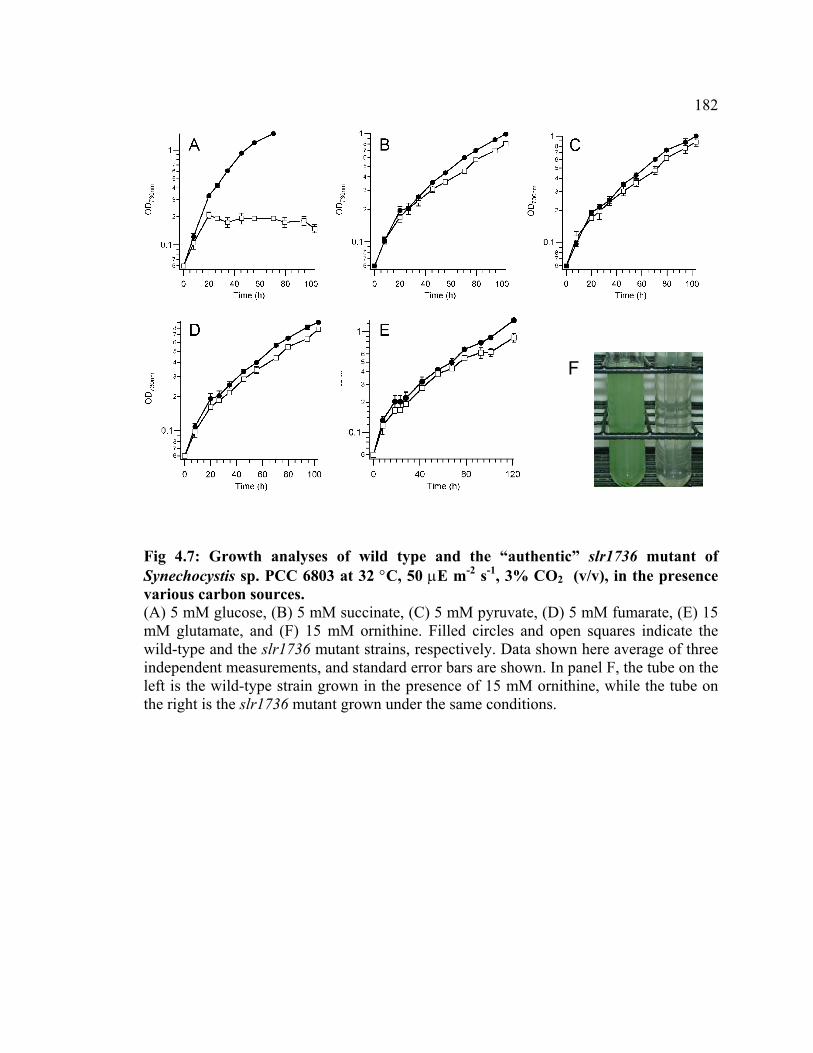
182
Fig 4.7: Growth analyses of wild type and the “authentic” slr1736 mutant of Synechocystis sp. PCC 6803 at 32 °C, 50 µE m-2 s-1, 3% CO2 (v/v), in the presence various carbon sources. (A) 5 mM glucose, (B) 5 mM succinate, (C) 5 mM pyruvate, (D) 5 mM fumarate, (E) 15 mM glutamate, and (F) 15 mM ornithine. Filled circles and open squares indicate the wild-type and the slr1736 mutant strains, respectively. Data shown here average of three independent measurements, and standard error bars are shown. In panel F, the tube on the left is the wild-type strain grown in the presence of 15 mM ornithine, while the tube on the right is the slr1736 mutant grown under the same conditions.
F
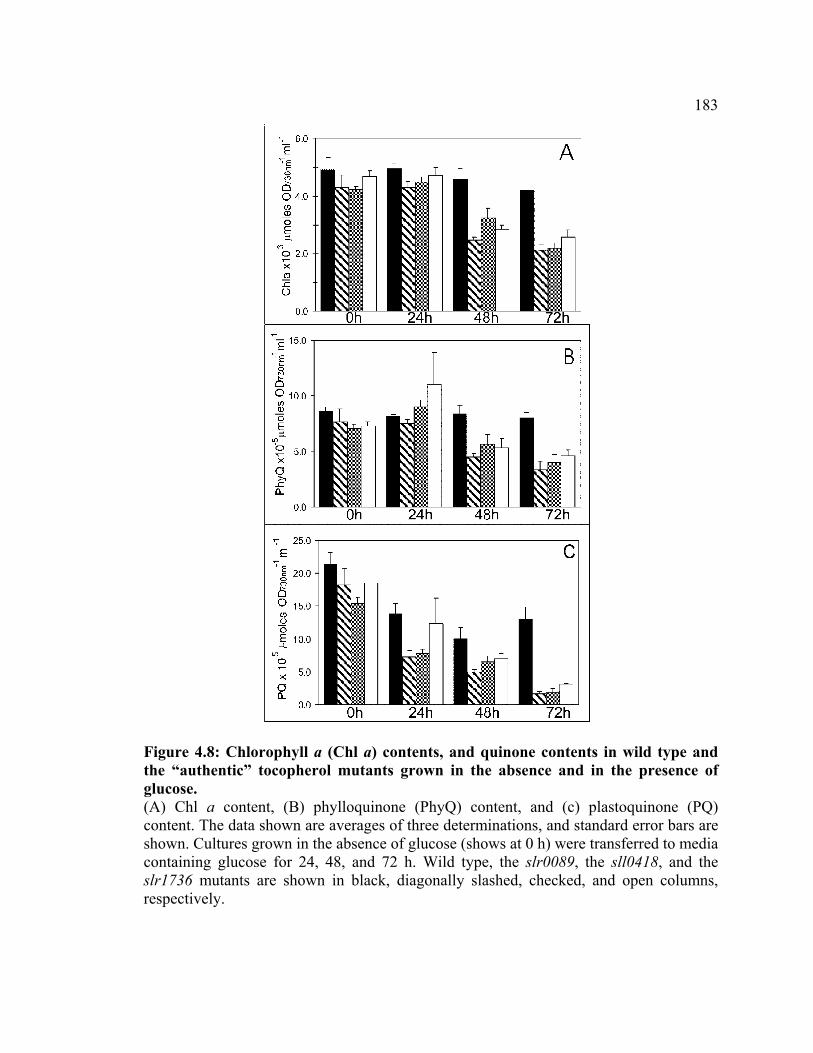
183
Figure 4.8: Chlorophyll a (Chl a) contents, and quinone contents in wild type and the “authentic” tocopherol mutants grown in the absence and in the presence of glucose. (A) Chl a content, (B) phylloquinone (PhyQ) content, and (c) plastoquinone (PQ) content. The data shown are averages of three determinations, and standard error bars are shown. Cultures grown in the absence of glucose (shows at 0 h) were transferred to media containing glucose for 24, 48, and 72 h. Wild type, the slr0089, the sll0418, and the slr1736 mutants are shown in black, diagonally slashed, checked, and open columns, respectively.
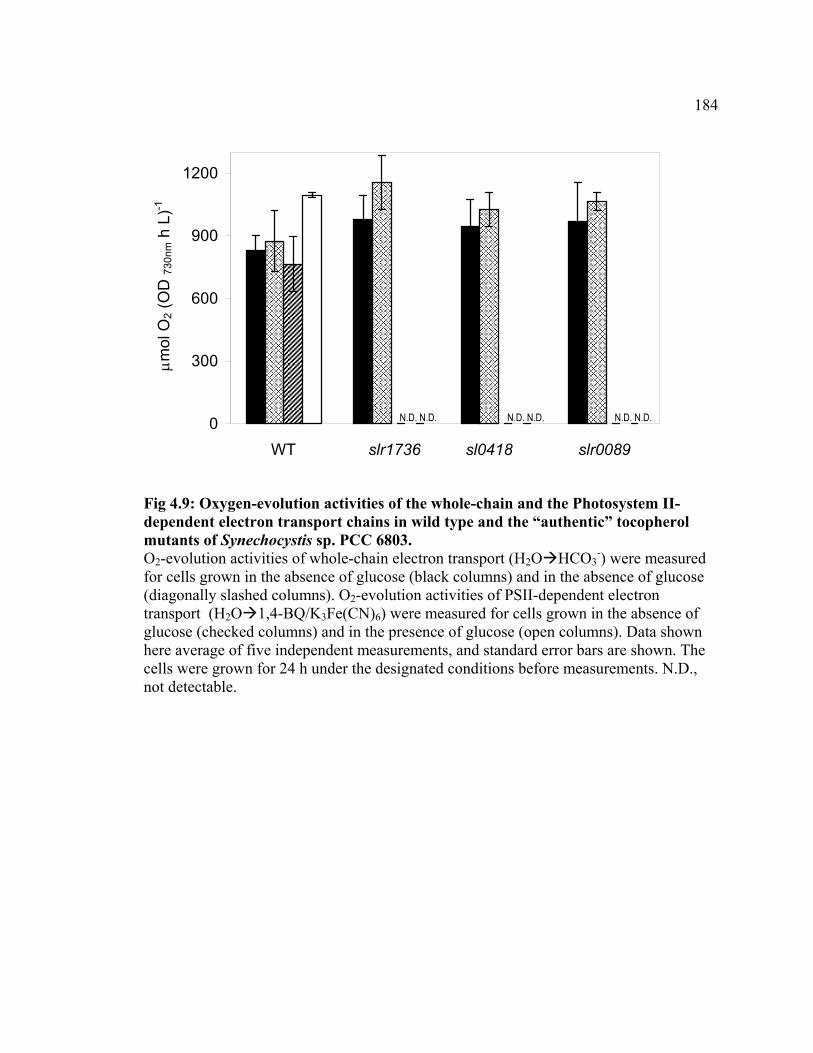
184
������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������������������������������
���������������������������������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������
����������������������������������������������������������������������������������������
0
300
600
900
1200
Fig 4.9: Oxygen-evolution activities of the whole-chain and the Photosystem II-dependent electron transport chains in wild type and the “authentic” tocopherol mutants of Synechocystis sp. PCC 6803. O2-evolution activities of whole-chain electron transport (H2O HCO3
-) were measured for cells grown in the absence of glucose (black columns) and in the absence of glucose (diagonally slashed columns). O2-evolution activities of PSII-dependent electron transport (H2O 1,4-BQ/K3Fe(CN)6) were measured for cells grown in the absence of glucose (checked columns) and in the presence of glucose (open columns). Data shown here average of five independent measurements, and standard error bars are shown. The cells were grown for 24 h under the designated conditions before measurements. N.D., not detectable.
WT slr1736 sl0418 slr0089
N.D. N.D.
µmol
O2 (
OD
730n
m h
L)-1
N.D. N.D. N.D. N.D.

185
Fig 4.10: Immunoblotting analysis for the D1 and PsbO proteins in whole cells of wild type and three “authentic” tocopherol mutants grown in the absence of glucose (Photoautotrophic) and in the presence of glucose (Photomixotrophic) at 32 °C, 50 µE m-2 s-1, with 3% (v/v) CO2. Proteins from equal amount of cells (10 µL of cell suspension with OD730 nm =100) was loaded in each lane. After electrophoresis, proteins were transferred to a nitrocellulose membrane. Proteins were detected by immunoblotting as described in the Materials and Methods.

186
Fig 4.11: Time-dependent RT-PCR analysis of the sodB and nblA transcripts in wild type and the “authentic” slr1736 mutant of Synechocystis sp. PCC 6803 The cells were initially grown in the absence of glucose (shown at 0 h) and transferred to media containing 5mM glucose and grown for 4, 8, 24 h at 3% CO2 (v/v), 32°C, 50 µE m-2 s-1. rnpB, encoding the catalytic RNA subunit of RNase P, was used as control.
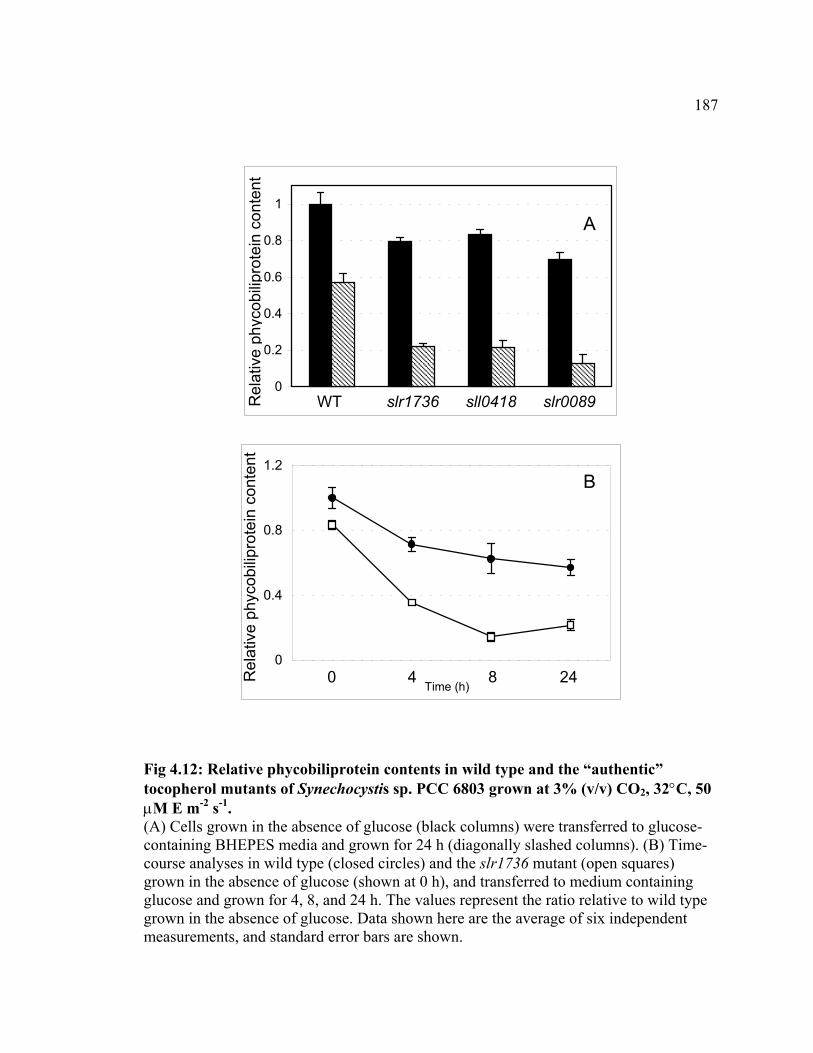
187
��������������������������������������������������������������������������������
����������������������������������������
����������������������������������������
��������������������
0
0.2
0.4
0.6
0.8
1
0
0.4
0.8
1.2
Fig 4.12: Relative phycobiliprotein contents in wild type and the “authentic” tocopherol mutants of Synechocystis sp. PCC 6803 grown at 3% (v/v) CO2, 32°C, 50 µM E m-2 s-1. (A) Cells grown in the absence of glucose (black columns) were transferred to glucose-containing BHEPES media and grown for 24 h (diagonally slashed columns). (B) Time-course analyses in wild type (closed circles) and the slr1736 mutant (open squares) grown in the absence of glucose (shown at 0 h), and transferred to medium containing glucose and grown for 4, 8, and 24 h. The values represent the ratio relative to wild type grown in the absence of glucose. Data shown here are the average of six independent measurements, and standard error bars are shown.
Rel
ativ
e ph
ycob
ilipro
tein
con
tent
WT slr1736 sll0418 slr0089
Rel
ativ
e ph
ycob
ilipro
tein
con
tent
0 4 8 24 Time (h)
B
A
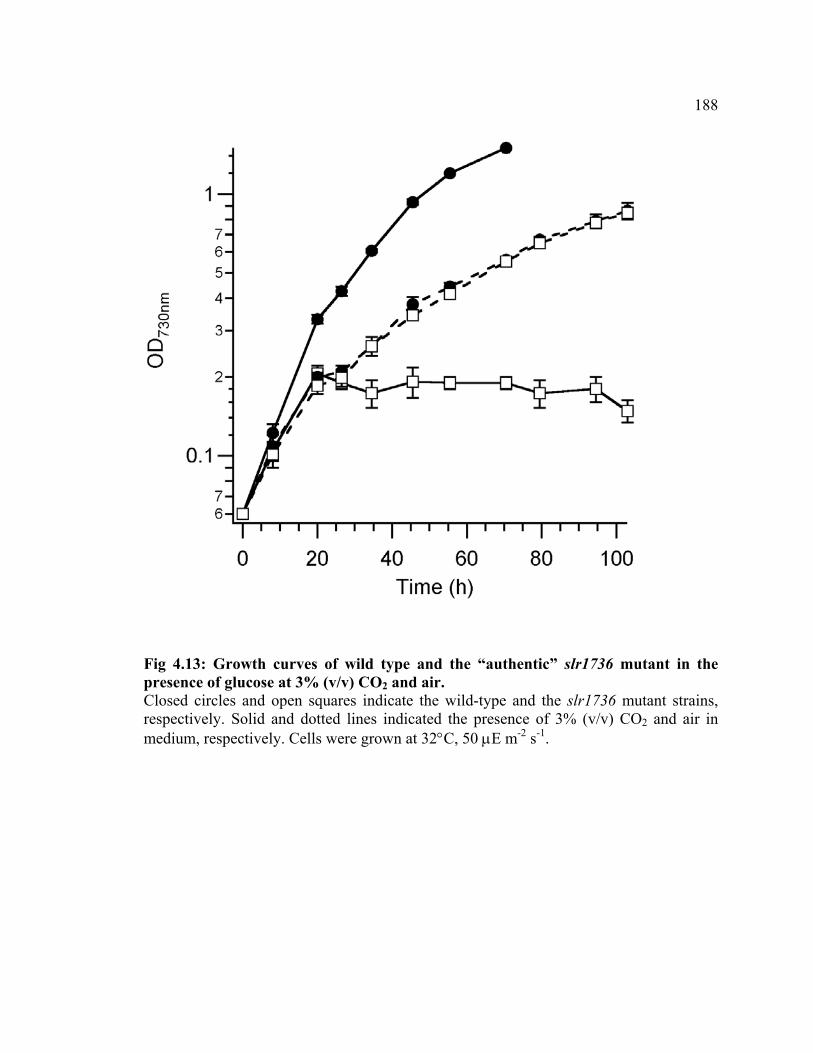
188
Fig 4.13: Growth curves of wild type and the “authentic” slr1736 mutant in the presence of glucose at 3% (v/v) CO2 and air. Closed circles and open squares indicate the wild-type and the slr1736 mutant strains, respectively. Solid and dotted lines indicated the presence of 3% (v/v) CO2 and air in medium, respectively. Cells were grown at 32°C, 50 µE m-2 s-1.
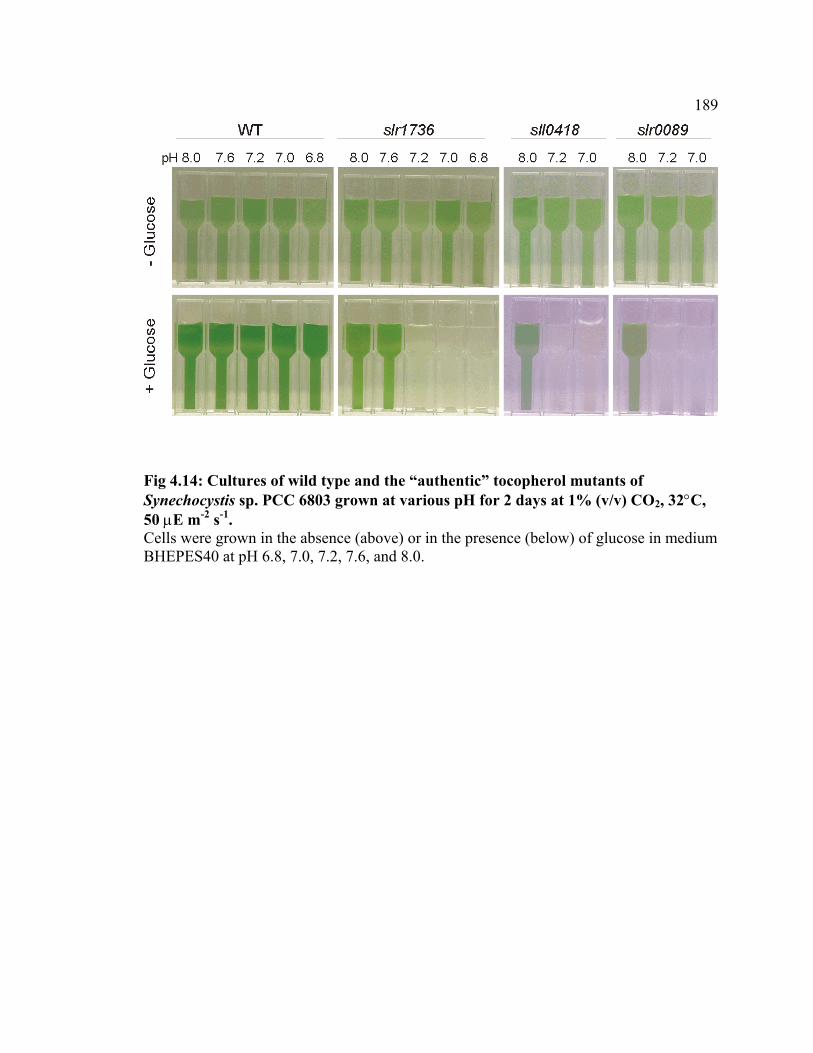
189
Fig 4.14: Cultures of wild type and the “authentic” tocopherol mutants of Synechocystis sp. PCC 6803 grown at various pH for 2 days at 1% (v/v) CO2, 32°C, 50 µE m-2 s-1. Cells were grown in the absence (above) or in the presence (below) of glucose in medium BHEPES40 at pH 6.8, 7.0, 7.2, 7.6, and 8.0.
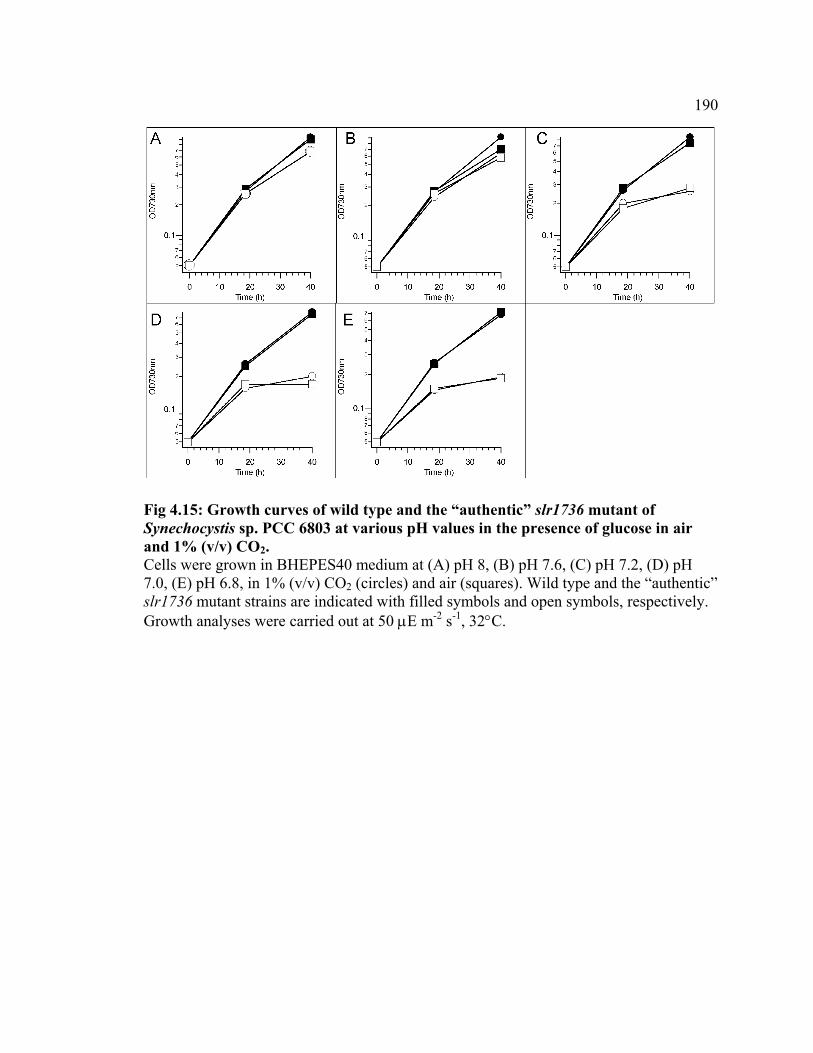
190
Fig 4.15: Growth curves of wild type and the “authentic” slr1736 mutant of Synechocystis sp. PCC 6803 at various pH values in the presence of glucose in air and 1% (v/v) CO2. Cells were grown in BHEPES40 medium at (A) pH 8, (B) pH 7.6, (C) pH 7.2, (D) pH 7.0, (E) pH 6.8, in 1% (v/v) CO2 (circles) and air (squares). Wild type and the “authentic” slr1736 mutant strains are indicated with filled symbols and open symbols, respectively. Growth analyses were carried out at 50 µE m-2 s-1, 32°C.
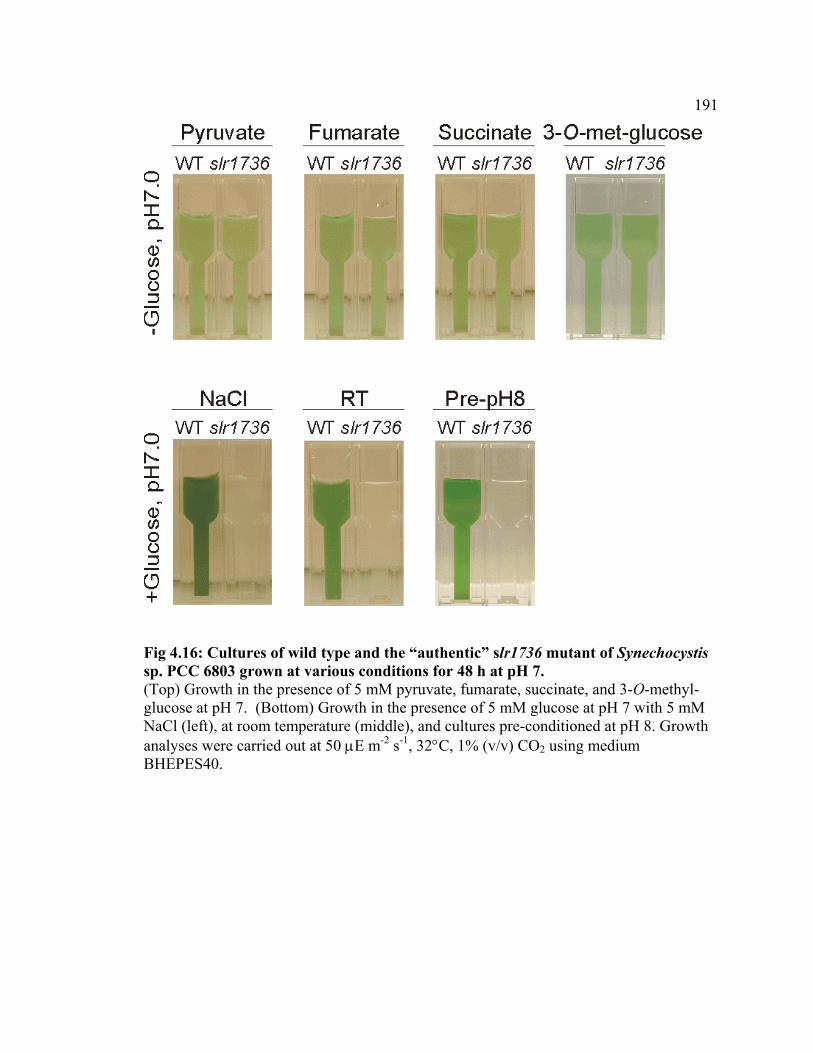
191
Fig 4.16: Cultures of wild type and the “authentic” slr1736 mutant of Synechocystis sp. PCC 6803 grown at various conditions for 48 h at pH 7. (Top) Growth in the presence of 5 mM pyruvate, fumarate, succinate, and 3-O-methyl-glucose at pH 7. (Bottom) Growth in the presence of 5 mM glucose at pH 7 with 5 mM NaCl (left), at room temperature (middle), and cultures pre-conditioned at pH 8. Growth analyses were carried out at 50 µE m-2 s-1, 32°C, 1% (v/v) CO2 using medium BHEPES40.
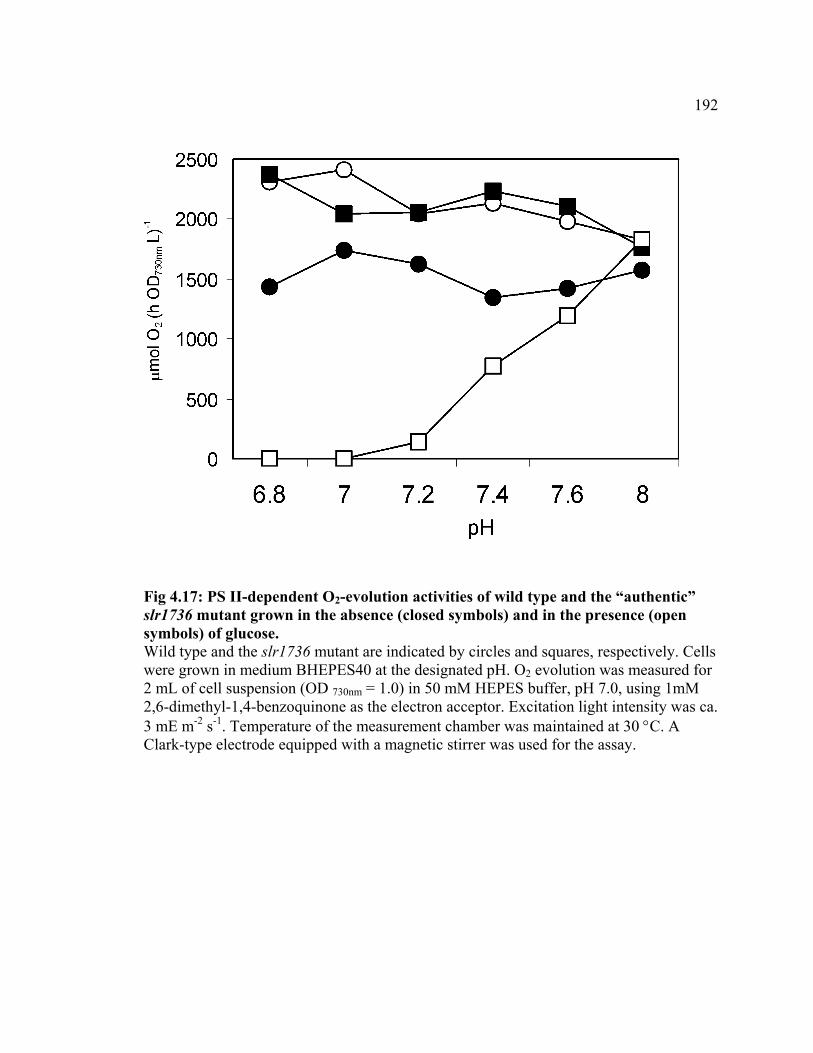
192
Fig 4.17: PS II-dependent O2-evolution activities of wild type and the “authentic” slr1736 mutant grown in the absence (closed symbols) and in the presence (open symbols) of glucose. Wild type and the slr1736 mutant are indicated by circles and squares, respectively. Cells were grown in medium BHEPES40 at the designated pH. O2 evolution was measured for 2 mL of cell suspension (OD 730nm = 1.0) in 50 mM HEPES buffer, pH 7.0, using 1mM 2,6-dimethyl-1,4-benzoquinone as the electron acceptor. Excitation light intensity was ca. 3 mE m-2 s-1. Temperature of the measurement chamber was maintained at 30 °C. A Clark-type electrode equipped with a magnetic stirrer was used for the assay.
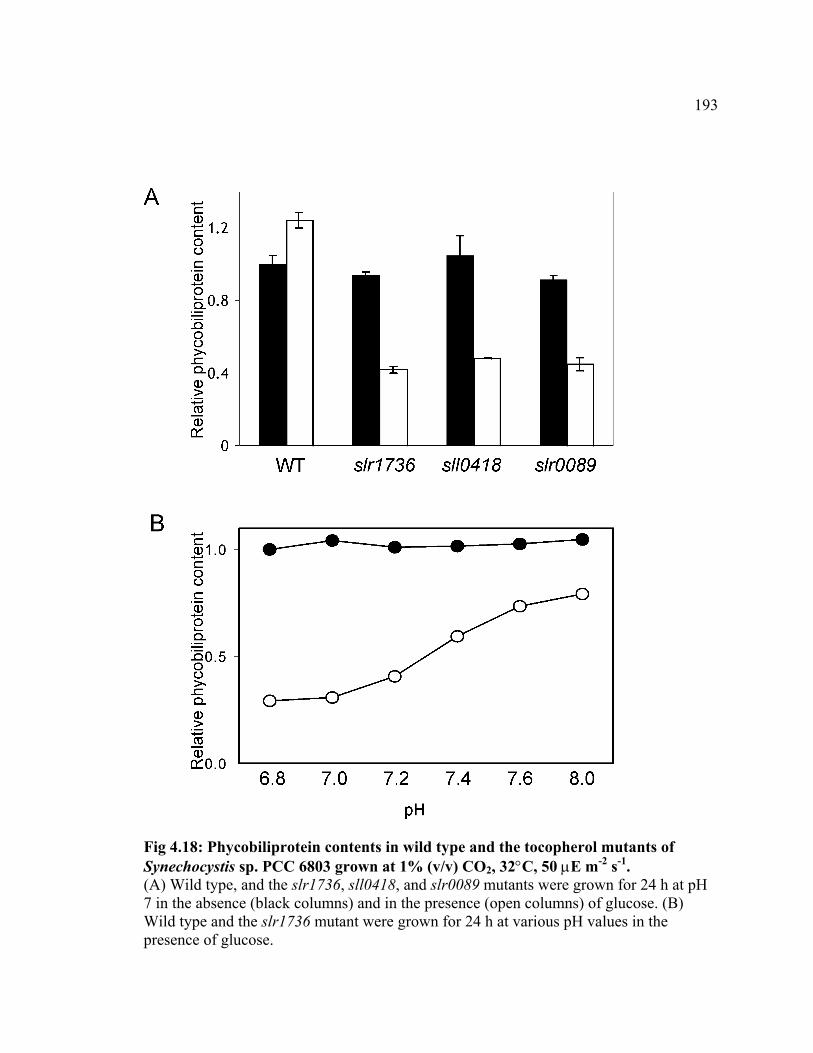
193
Fig 4.18: Phycobiliprotein contents in wild type and the tocopherol mutants of Synechocystis sp. PCC 6803 grown at 1% (v/v) CO2, 32°C, 50 µE m-2 s-1. (A) Wild type, and the slr1736, sll0418, and slr0089 mutants were grown for 24 h at pH 7 in the absence (black columns) and in the presence (open columns) of glucose. (B) Wild type and the slr1736 mutant were grown for 24 h at various pH values in the presence of glucose.

194
Chapter 5
Transcriptional Regulation of Metabolic and Sigma Factor Genes in the Vitamin E-
Deficient Mutant of Synechocystis sp. PCC 6803
Publication:
Yumiko Sakuragi, Dean DellaPenna, Donald A. Bryant, manuscript in preparation

195
ABSTRACT
Transcriptional regulation of metabolic genes and sigma factor genes was studied in the
vitamin E (α-tocopherol)-deficient mutant (slr1736) of Synechocystis sp. PCC 6803.
When glucose was present in growth medium at pH 8.0, the transcripts of nblA (a factor
essential for the controlled degradation of PBP), sbpA (the periplasmic sulfate binding
protein), and sigB, sigC, sigD, sigE, sigH, sigI (alternative sigma factors) were shown to
accumulate in the tocopherol-less mutant cells. These results indicate that the α-
tocopherol mutant responds to the presence of glucose as though nitrogen, carbon, and
sulfur were limiting in the environment. When glucose is present in the growth medium
at pH 7.0, the tocopherol mutant ceased growth after 20 h. Under these conditions, the
amounts of the sigA (the principal sigma factor), rbcL (the RuBisCO large subunit), and
ccmK1 and ccmL (carboxysome shell proteins) transcripts diminished after 4 h. Because
the amounts of these transcripts remained unaltered when cells were grown in the
absence of glucose or in the presence of glucose pH 8.0, it was concluded that the pH-
dependent, glucose-induced misregulation of the sigA and rbcL are largely responsible
for the glucose-induced lethality of the tocopherol mutants under these conditions. Thin-
section electron micrographs revealed the absence of carboxysomes in the tocopherol
mutant grown in the presence of glucose at pH 7.0. The transcription of slr2031 (a
putative protein phosphatase that shows sequence similarity with RsbU and that is
essential under sulfur- and nitrogen-limiting conditions) and ndhF3, ndhR, sbtA
(inorganic carbon uptake mechanisms) were shown to be constitutively down-regulated
under all conditions tested in the tocopherol mutant. This result demonstrates that α-

196
tocopherol is involved in the transcriptional regulation of these metabolic genes in
Synechocystis sp. PCC 6803. Thin-section electron micrographs of the tocopherol-less
mutant showed that a large number of glycogen granules accumulated in the cells when
grown even in the absence of glucose. The targeted interruption of the pmgA gene (a
putative anti-sigma factor homologous to RsbW) resulted in a similar pH-dependent,
glucose-induced lethal phenotype. Based on these results, it is concluded that α-
tocopherol acts as a global regulatory molecule in carbon, sulfur, and nitrogen
metabolism in Synechocystis sp. PCC 6803.

197
ABBREVIATIONS
CCM inorganic carbon concentrating mechanisms
DCMU 3-(3,4-dichlorophenyl)-1,1-dimethyl-urea
HEPES 4-(2-hydroxyethyl)-piperazine-N’-(2-ethanesulfonic acid)
PBP phycobiliprotein
PS II Photosystem II
RT-PCR reverse transcriptase-polymerase chain reaction
RuBisCO ribulose-1,5-bisphosphate carboxylase/oxygenase

198
INTRODUCTION
α-Tocopherol is synthesized only in oxygenic phototrophs including
cyanobacteria, green algae and higher plants (Threlfall and Whistance 1971; Collins and
Jones, 1981). Given that α-tocopherol constitutes an essential component of our diet,
most of our knowledge concerning the roles and functions of this molecule has been
obtained from studies in animal systems. Based on results gathered in animals, animal
cell cultures, and artificial membranes, it has been shown that tocopherols function as
antioxidants and scavenge or quench various reactive oxygen species and lipid oxidation
byproducts that would otherwise propagate lipid peroxidation chain reactions in
membranes (Kamal-Eldin and Appelqvist, 1996). This antioxidant role of α-tocopherol
has now been demonstrated in eukaryotic phototrophs owing to studies in the past several
years. The herbicide-mediated interruption of the α-tocopherol biosynthesis pathway has
been shown to render PS II more susceptible to oxidative stress under extremely high
light illumination in the green alga Chlamydomonas reinhardtii (Trebst et al., 2002). The
presence of α-tocopherol has been shown to suppress the propagation of lipid
peroxidation during seedling germination in Arabidopsis thaliana (Sattler et al., 2004).
Thus, there is little doubt that α-tocopherol functions as a bulk antioxidant in the
eukaryotic phototrophs as well as in animals. The antioxidant role of α-tocopherol is also
being investigated in the cyanobacterium Synechocystis sp. PCC 6803, and the results
obtained so far support an antioxidant role of α-tocopherol in this organism (H. Maeda,
Y. Sakuragi, D. A. Bryant, and D. DellaPenna, personal communication)

199
Perhaps the most novel and exciting functions of α-tocopherol have been
uncovered by various research groups during the last few years. In addition to the
antioxidant functions, other “non-antioxidant” functions related to modulation of
signaling and transcriptional regulation in mammals have also been reported (Chan et al.,
2001; Azzi et al., 2002; Ricciarelli et al., 2002). For example, α-tocopherol has been
shown to bind to phospholipase A2 specifically at the substrate-binding pocket and to act
as a competitive inhibitor, thereby decreasing the release of arachidonic acid for
eicosanoid synthesis (Chandra et al., 2002). α-Tocopherol is also directly involved in
transcriptional regulation in animals, including the expression of the genes encoding liver
collagen αI, α-tocopherol transfer protein, and α-tropomyosin collagenase (Yamaguchi
et al., 2001; Azzi et al., 2002). The presence of α-tocopherol resulted in a modulation of
the phosphorylation state of protein kinase Cα in rat smooth-muscle cells, possibly via
phosphorylation of protein phosphatase 2A (Ricciarelli et al., 1998). None of these non-
antioxidant functions of α-tocopherol has yet been experimentally demonstrated in
oxygenic phototrophs.
In the previous studies described in Chapter 4, the role(s) of α-tocopherol in
cyanobacteria was investigated by using three mutants, slr1736, sll0418, and slr0089,
deficient in enzymes involved in the α-tocopherol biosynthetic pathway (see Fig 4.1).
The tocopherol mutants are able to grow similarly to wild type at pH values between 6.8
and 8.0 in the absence of glucose (control conditions), under which conditions the PS II
activities and the phycobiliprotein (PBP) contents were comparable to those of wild type.
After transfer to a medium containing glucose at pH 8.0, the tocopherol mutants also

200
grew similarly to wild type and the PS II activity remained unaltered, although the PBP
content decreased by ~30 % (permissive conditions). After transfer to medium containing
glucose at pH 7.0, however, the tocopherol mutants ceased growth by 24 h, and the PS II
activity was completely lost. Under these conditions the PBP content was reduced by ≥
60 % (lethal conditions). The involvement of oxidative stress in the PS II inactivation
was largely ruled out, because the levels of the D1 protein and the sodB transcript were
comparable between wild type and the tocopherol mutants under these conditions. Based
on the observed reduction in the PBP content and the accumulation of the nblA transcript,
which is known to accumulate in response to nutrient limitation (Collier and Grossman,
1994; Richaud et al., 2001), it was suggested that the tocopherol mutants are experiencing
macronutrient starvation. This led to the hypothesis that α-tocopherol plays a role as a
regulatory molecule in macronutrient metabolism in Synechocystis sp. PCC 6803.
In this chapter, a study of the transcriptional regulation of metabolic genes and
sigma factor genes in a tocopherol-deficient mutant (slr1736) is described. Of 39 genes
initially analyzed, the amounts of the transcripts of 10 genes were shown to respond
either to the presence of glucose alone or to the presence of glucose and pH in the
medium, whereas the amounts of the transcripts of 4 genes were shown to be
constitutively down regulated upon loss of α-tocopherol. Based on these observations, it
is concluded that α-tocopherol is involved in transcriptional regulation of some key
metabolic genes and plays a role in the global coordination of macronutrient metabolism
in Synechocystis sp. PCC 6803.

201
RESULTS
Transcription of nblA and sbpA
Given the significant reduction of the PBPs in response to glucose and lowered
pH (see Chapter 4), the nblA transcript was analyzed in wild type and slr1736 mutant
cells grown at pH 7.0 and pH 8.0 (Fig 5.1). In the absence of glucose, the wild-type strain
barely accumulated detectable nblA transcripts both at pH 7.0 and pH 8.0, and only a
small increase followed in response to glucose in the growth medium. This appeared
somewhat contradictory to the observed behavior of the PBP content in the wild type, in
which the PBP content increased by 20% after transfer to medium containing glucose
when cells were grown at pH 7.0 (see Chapter 4, Fig 4.18). Furthermore, the medium
used for culturing these cells contains a growth-rate saturating concentration of nitrate
(ca. 18 mM) (Stainer 1971; see Materials and Methods). Thus, it is inconceivable that
these cells were actually limited by the availability of nitrogen. The presence of glucose
in the medium might lead to an increase in the intracellular carbon pool and, as a result,
facilitate the faster growth of this organism. These “carbon-saturated”, fast-dividing cells
perhaps are not capable of metabolizing nitrogen at an equivalent rate, and thus respond
as though they are limited by nitrogen in their environment.
The slr1736 mutant accumulated a higher amount of nblA transcript in the
absence of glucose as compared to the wild-type control (Fig 5.1, shown at 0 h), which
suggests that the slr1736 mutant is “perceiving” nitrogen limitation even in the absence
of glucose. In response to glucose, the amount of the nblA transcripts increased
dramatically in the slr1736 mutant after 4 h when cells were grown in medium at pH 7.0

202
(Fig 5.1). Similar although more moderate increases were observed when cells were
grown in the presence of glucose at pH 8.0. These results indicate that the slr1736 mutant
is “perceiving” a severe nitrogen limitation at pH 7.0 and moderate nitrogen limitation at
pH 8.0. This is consistent with the dramatic reduction of PBP contents at these pHs (ca.
40% of the wild-type level at pH7.0 and ca. 70% of the wild-type level at pH 8.0).
The sbpA transcript, encoding the periplasmic sulfate binding protein, is known to
accumulate in response to sulfate limitation (Laudenbach et al., 1991). No detectable
sbpA transcripts were present in wild-type cells grown at pH 7.0 and pH 8.0 in the
absence or presence of glucose (Fig 5.1). In the slr1736 mutant cells grown at pH 7.0, a
small yet discernible amount of the sbpA transcripts was detected in the absence of
glucose, and these transcripts dramatically increased by 8 h after transfer to medium
containing glucose. When the slr1736 mutant cells were grown at pH 8.0, no sbpA
transcripts were detectable in the absence of glucose, and a gradual increase was
observed in 24 h after transfer to a medium containing glucose. These results indicate that
the slr1736 mutant is “perceiving” sulfur limitation even in the absence of glucose at pH
7.0, and the response is exacerbated after transfer to a medium containing glucose. It is
noteworthy that the amounts of the sbpA and nblA transcripts accumulated at pH 7.0 and
8.0 coincided well with the degree of reduction of the PBP contents at these pHs in the
presence of glucose (See Chapter 4, Fig 4.18, and Fig 5.1). The combined results hence
demonstrate that the reduction of the PBP content is associated with “perceived” nitrogen
and sulfur limitation in the slr1736 mutant.

203
The cpcA transcripts, encoding the phycocyanin α-subunit, which is a subunit of
the most abundant PBPs, were found to decrease gradually in the tocopherol mutant in
response to glucose when the slr1736 mutant cells were grown at pH 7.0. At pH 8.0 its
level remained largely unaltered both in wild type and the slr1736 mutant before and
after addition of glucose. Therefore, it was concluded that the mechanism of the
reduction of the PBP content at pH 7.0 and 8.0 in the slr1736 mutant grown in the
presence of glucose are different. At pH 8.0, the reduction of the PBP content primarily
involves active degradation of the PBP proteins, while at pH 7.0 it involves both the
active degradation, as characterized by the accumulation of the nblA transcript, and the
transcriptional down regulation of the genes encoding PBPs, which further supports the
conclusion that the cells are perceiving nutrient limitation and responding to it (Grossman
et al., 1994; also see references therein)
Transcription of Alternative Sigma Factor Genes
The Synechocystis sp. PCC 6803 genome encodes eight σ70-type alternative sigma
factors (SigB through I) in addition to the principal sigma factor (SigA), and they play
roles as transcription factors that are activated in response to various kinds of stresses
(Caslake et al., 1997; Gruber and Bryant, 1998; Huchauf et al., 2000; Muro-Pastor et al.,
2001; Imamura et al., 2003a, b). Fig 5.2 shows the results of the RT-PCR analysis of
sigB, sigC, sigD, sigE, sigH, and sigI in wild type and the slr1736 mutant in the absence
of glucose and in the presence of glucose. SigB has been suggested to be involved in a
variety of stress responses, including dark response (Imamura et al., 2003a) and heat-
shock response (Huchauf et al., 2000) in Synechocystis sp. PCC 6803, and carbon and

204
nitrogen starvation responses in Synechococcus sp. PCC 7002 (Caslake et al., 1997).
When cells were grown at pH 7.0, the amount of the sigB transcripts was largely
unaltered in wild type before and after addition of glucose to the medium (Fig 5.2). In the
slr1736 mutant, the amount of the sigB transcript was somewhat higher in the absence of
glucose as compared to the wild-type control, followed by a large increase by 4 h after
transfer to medium containing glucose. A similar response was observed for the slr1736
mutant grown at pH 8.0. Therefore the amount of the sigB transcripts responded to
glucose alone and was not influenced by the pH of the medium. Similar accumulation
patterns of transcripts were also observed for sigC, sigH, and sigI. SigH is involved in the
heat-shock response (Hackauf et al., 2000), while SigC, which is related to SigE in
Synechococcus sp. PCC 7002, is likely to be involved in the post-exponential phase of
growth (Gruber and Bryant, 1998). The role of SigI has not yet been elucidated. The
results suggest that the slr1736 mutant is experiencing a mixture to stresses in response of
glucose.
Transcription of sigE is under the control of NtcA, a global nitrogen regulator,
which binds to its characteristic binding site located in the upstream flanking sequence of
the sigE transcription initiation site and induces transcription of sigE in response to
nitrogen deprivation (Muro-Pastor et al., 2001). Interestingly, the amount of the sigE
transcripts increased in response to glucose in wild type both at pH 7.0 and 8.0 (Fig 5.2).
This is consistent with the pattern of the accumulation observed for the nblA transcripts
(Fig 5.1, see above). Although the reason for this glucose-induced nitrogen limitation
response has not yet been defined, it is possible that this is due to an altered intracellular

205
carbon-nitrogen balance caused by increased carbon flux induced by the glucose
metabolism (see also above). In the slr1736 mutant, the amount of the sigE transcripts
increased more dramatically than for the wild type in response to glucose. This glucose-
induced accumulation of the sigE transcripts occurred both at pH 7.0 and pH 8.0, which
is consistent with the results obtained for the nblA transcripts (Fig 5.1, see above) and
which indicates that the slr1736 mutant is “perceiving” a nitrogen limitation in response
to glucose.
In contrast to sigE, the amount of the sigD transcripts was largely unaffected by
the presence of glucose in wild type (Fig 5.2). In the tocopherol mutant, the amount of
the sigD transcript was lower than that of wild type in the absence of glucose and
increased by several-fold in response to glucose (by visual inspection). SigD is highly
expressed during exposure to high light (Imamura et al., 2003a,b). It has been suggested
that this light-dependent behavior of sigD expression is regulated by the redox state of PS
II (Imamura et al., 2003a). Because the growth conditions used in this study employ
continuous illumination of the cultures with a moderate light intensity (50 µE m-2 s-1),
and because the PS II activity is nearly at the wild-type level for the slr1736 mutant in the
absence of glucose at pH 7.0 and 8.0 or in the presence of glucose at pH 8.0, the observed
alteration of the sigD transcription can not be explained simply by the previously
suggested light-responsive role of SigD.
Taking these results together, it was firmly concluded that the transcription of
these alternative sigma factor genes is modulated in the slr1736 mutant in response to
glucose and that pH plays little role in the glucose-induced response of the transcription

206
of the alternative sigma factors. This indicates that α-tocopherol plays a role in regulating
the stress response related at least to carbon, nitrogen and sulfur metabolism in
Synechocystis sp. PCC 6803.
Transcription of sigA and Ci Genes
The sigA gene encodes the principal sigma factor, which is responsible for the
transcription of genes encoding proteins essential for growth and reproduction in
eubacteria including cyanobacteria (Gruber and Bryant, 1997; Caslake and Bryant, 1996).
The amount of the sigA transcripts was analyzed for wild type and the slr1736 mutant
grown at pH 7.0 and 8.0 (Fig 5.3). At pH 7.0, the sigA transcripts in wild type remained
constant after addition of glucose to medium, which is consistent with the fact that the
wild-type cells grow normally under these conditions. The sigA transcript level in the
slr1736 mutant at pH 7.0 was virtually identical to that for the wild type in the absence of
glucose, which is also consistent with the fact that the slr1736 mutant grows similarly to
wild type in the absence of glucose at pH 7.0. The level of the sigA transcripts remained
unaltered for 4 h when the slr1736 mutant was transferred to medium containing glucose
at pH 7.0. However, after 8 h the sigA transcript level significantly diminished. Such a
dramatic reduction in the amount of the sigA transcript is likely to cause a reduction of
the content of SigA available for transcription of house-keeping genes, which would in
turn severely impair the growth capacity of the cells. Indeed, the tocopherol mutants are
unable to grow in the presence of glucose at pH 7.0 (see Chapter 4, Fig 4.14 and Fig
4.15). In contrast, when the slr1736 mutant was grown in the presence of glucose at pH
8.0, the amount of the sigA transcript was virtually unaltered and was very similar to that

207
in the wild type control. This is consistent with the fact that the slr1736 mutant grew
similarly to wild type under these conditions (see Chapter 4, Fig 4.14 and Fig 4.15). The
direct correlation between the amounts of the sigA transcripts and the growth behavior of
the slr1736 mutant in the absence and in the presence of glucose at pH 7.0 and 8.0
strongly indicate that the pH-dependent glucose toxicity is caused by the pH-dependent,
glucose-induced alteration of the sigA transcript level in the slr1736 mutant.
Accompanying the reduction of the amounts of the sigA transcript, the amounts of
the transcripts of carboxysome-related genes (rbcL, ccmK1, ccmL), a carbon-
concentrating mechanism (CCM) gene (ndhF4), and sigG were also decreased in a pH-
and glucose-dependent manner in the slr1736 mutant (Fig 5.3). rbcL encodes the large
subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), which is
responsible for the fixation of CO2 and is essential for the survival of the cells, because
this gene cannot be completely inactivated in cyanobacteria (Pierce et al., 1989). ccmK1
and ccmL encode carboxysome shell proteins, which form the outer shell of
carboxysomes and encapsulate RuBisCO within the carboxysomal structure (Price et al.,
1993). ndhF4 encodes a subunit of a multi-subunit complex that is a low-affinity
constitutive CO2 uptake transporter (Shibata et al., 2001). The reduction in the levels of
the carboxysome transcripts most likely leads to the reduction in the amount of
carboxysomes in the cells; as a result, the ability to fix CO2 would be severely impaired
(Price et al., 1993).

208
Intracellular Structures
Thin-section electron micrographs of wild-type and slr1736 mutant cells grown at
pH 7.0 in the absence and the presence of glucose were recorded. Fig 5.4 shows the
electron micrographs of wild type grown in the absence (Fig 5.4A) and in the presence of
glucose (Fig 5.4B). In the wild-type cells grown in the absence of glucose, the space
between the thylakoid membranes was filled with electron-opaque material, which most
likely represents the protein-rich phycobilisomes (Fig 5.9A). In the cytoplasm, electron-
opaque polyhedral bodies are visible, which are characteristic of carboxysomes. Slightly
larger electron-transparent bodies with round or oval shapes are most likely due to poly-
β-hydroxybutyrate (Stainer, 1988). In the wild-type cells grown in the presence of
glucose (Fig 5.4B), on the other hand, the intermembrane space is filled with small
electron-transparent objects with irregular shapes, which are characteristic of glycogen
granules. Glycogen granules serve as general carbohydrate reserves (Stainer, 1998). The
cellular content of glycogen accounts for 10-20 % of the dry cell weight when cells are
growing photoautotrophically and exponentially, whereas it accounts for up to 60 % of
dry cell weight when cells are subjected to nitrogen-limiting conditions (Stainer, 1988;
Smith 1982). That few glycogen granules are observed in the wild-type cells grown in the
absence of glucose is consistent with the fact that these cells grow in a balanced manner
and optimally under these conditions. The observed accumulation of glycogen in the
wild-type cells grown in the presence of glucose most likely reflects a carbon-saturated,
or nitrogen-limited metabolic state of the cells.

209
Fig 5.5A shows an electron micrograph of an slr1736-mutant cell grown in the
absence of glucose at pH 7.0. The intracellular structures, such as the presence of
thylakoid membranes, carboxysomes, and poly-β-hydroxybutyrate, are very similar to
those of the wild-type control grown under the same conditions (Fig 5.4 A). However, in
the slr1736 mutant, the intermembrane space was filled with glycogen granules. This
clearly contrasts with the situation in the wild-type cells, in which only a small number of
glycogen granules was detectable (Fig 5.4A). Rather, this feature of the slr1736 mutant
cell resembles that of the wild-type cell grown in the presence of glucose (Fig 5.4B). This
suggests that the slr1736 mutant accumulates an unusually large amount of carbohydrate
even in the absence of glucose. Therefore, the slr1736 mutant cells seem either to be
experiencing nitrogen limitation, or to be performing CO2 fixation or glycogen synthesis
much more efficiently than the wild-type cells. The former is not likely to be the case as
the mutant cells grow similarly to wild type under these conditions. Therefore, it is most
likely that the mutant is overproducing photosynthate. In either case, it is obvious that the
“metabolic state” of the slr1736 mutant is different from that of the wild type even in the
absence of glucose.
When the slr1736 mutant was grown in the presence of glucose, the appearance of
the intracellular structures changed dramatically (Fig 5.5B). The intermembrane space is
no longer electron-opaque, which is consistent with the significant reduction in the PBP
content. No carboxysomes were observed in the majority of the cells, which supports the
results that the transcripts of the carboxysome genes are substantially reduced in the
slr1736 mutant under these conditions. The number of layers formed by the thylakoid

210
membranes, however, does not seem to be affected significantly. Interestingly, new oval
structures with an electron-opaque appearance are visible between the thylakoid
membranes; these structures were absent in wild type grown in the absence and in the
presence of glucose and in the slr1736 mutant grown in the absence of glucose.
Constitutive Alteration of the CCM Genes and slr2031 Expression
Given the alteration of the “metabolic state” in the slr1736 in the absence of
glucose, further transcription analyses were carried out in order to understand the
involvement of α-tocopherol in macronutrient metabolism. Of 39 genes analyzed for
their transcript level, 4 genes (ndhF3, ndhR, sbtA, slr2031) showed constitutive down-
regulation both in the presence and in the absence of glucose (Fig 5.6). ndhF3, ndhR, and
sbtA encode components of high-affinity CO2 uptake transporters (Klughammer et al.,
1999; Shibata et al., 2001, 2002), a repressor of the ndhF3 operon, and the sodium-
dependent bicarbonate transporter, respectively, and all participate in inorganic carbon
concentrating mechanisms (CCM) under limited inorganic carbon environments. The
transcripts of these genes were detected in wild type both in the absence and the presence
of glucose at pH 7.0 and 8.0. On the contrary, the transcript levels for these genes were
significantly reduced in the slr1736 mutant both in the presence and in the absence of
glucose both at pH 7.0 and 8.0. Thus, the constitutive alteration of these CCM genes is
directly linked to the loss of α-tocopherol.
These CCM genes are known to be upregulated in response to low inorganic
carbon environments, and are hardly detectable under high CO2 conditions as analyzed
by Northern-blot hybridization (Figge et al., 2001). This raises a question as to whether

211
the detection of the CCM genes in this study [at 1% (v/v) CO2] is due to an artifact such
as contamination with genomic DNA during the RNA isolations. The RNA preparation is
carried out through three steps, including isolation, RNase-free DNase treatment, and
finally purification; thus, it is unlikely that genomic DNA is present in the preparation.
Indeed, PCR analyses on the same RNA templates used for RT-PCR analyses using
primers targeted to the rnpB gene resulted in no product formation, which confirms that
the RNA preparations are free of genomic DNA contamination. Currently, we do not
have an explanation other than attributing the discrepancy to a difference in the
sensitivity of the detection methods. Typically one-step RT-PCR analyses using the
Qiagen One-Step RT-PCR kit (Qiagen inc., Valencia, CA) requires 50 pg µL-1 to 5 ng
µL-1 of total RNA in order to detect the target transcripts, whereas typical Northern-blot
hybridization reactions require several micrograms of RNA. Indeed, when more sensitive
methods were employed, other groups have also detected the presence of the ndhF3 and
sbtA transcripts in RNA preparations obtained from cells grown at high CO2 level [3-5%
(v/v) CO2] (Shibata et al., 2002; McGinn et al., 2003).
slr2031 encodes a putative protein phosphatase (similar to RsbU) involved in the
regulation of SigB in B. subtilis. In wild-type cells, the amount of slr2031 transcripts
were low yet detectable in the absence of glucose, and transcript levels substantially
increased by 8 h in response to glucose both at pH 7.0 and pH 8.0 (Fig 5.6). In the
slr1736 mutant, the slr2031 transcript is hardly detectable in the absence of glucose, and
only a slight induction was observed in response to glucose at pH 7.0. At pH 8.0, the
slr2031 transcript behaved similarly at pH 7.0, although a moderate increase was

212
observed in the presence glucose by 24 h. In Synechocystis sp. PCC 6803, Slr2031 has
been shown to be somehow involved in sulfur and nitrogen metabolism, and its absence
is deleterious under sulfur- and nitrogen-limiting conditions (Huchauf et al., 2000). An
independent study has shown that the presence of Slr0231 is also essential when the cotA
gene is inactivated (Katoh et al., 1995). CotA has been suggested to be involved in light-
induced proton extrusion and CO2 transport in Synechocystis sp. PCC 6803 (Katoh et al.,
1996a,b). Thus, it is possible that Slr2031 is involved in the transcriptional regulation of
genes that are involved in regulating processes including carbon, nitrogen, and sulfur
metabolism. The fact that the expression of slr2031, as well as ndhF3, ndhR, and sbtA, is
constitutively down-regulated in the absence of α-tocopherol demonstrates that α-
tocopherol plays a role in the transcriptional regulation of metabolic genes in
Synechocystis sp. PCC 6803.
Is the significant reduction of slr2031 transcription directly related to the cause of
the pH-dependent glucose toxicity of the tocopherol mutant? In order to answer this
question, an slr2031 disruption mutant was generated by insertion of the aadA gene
cassette, conferring spectinomycin resistance, into the unique MfeI site of the slr2031
coding region. Complete segregation of the mutant allele from the wild-type allele was
demonstrated by PCR analysis (Fig 5.7). Using the primers designed to target and
amplify a 0.9-kb N-terminal portion of the slr2031 loci resulted in the amplification of a
0.9-kb DNA fragment when the genomic DNA isolated from wild type was used as the
template. No 0.9-kb product was detectable when the genomic DNA isolated from the
slr2031::addA transformant was used as the template. Instead, a product of 2 kb was

213
detected. The size difference between the two products corresponds to the size of the
addA gene cassette, which demonstrates that the slr2031::addA transformant is
homozygous and free from the slr2031 wild-type allele.
A preliminary growth analysis was carried out at pH 7.0 and pH 8.0. The slr2031
mutant (slr2031::aadA) was able to grow both at pH 7.0 and pH 8.0 in the absence of
glucose. When the cells were transferred to medium containing glucose, the slr2031
mutant was able to grow similarly to wild type both at pH 7.0 and pH 8.0 (Fig 5.8 A and
B). An slr1736 slr2031 double mutant behaved similarly to the slr1736 mutant at both
pHs in the presence of glucose (Fig 5.8 A and B). Therefore, it was concluded that the
constitutive down-regulation of slr2031 transcription is not the direct cause of the pH-
dependent glucose toxicity observed for the tocopherol mutant.
As mentioned in Chapter 4, the pmgA mutant of Synechocystis sp. PCC 6803 is
unable to grow in the presence of glucose, which indicates that PmgA plays a role in the
regulation of photomixotrophic growth (growth on CO2 and glucose as the carbon source)
(Hihara and Ikeuchi, 1997). To test whether the role of PmgA is related to the role of α-
tocopherol, a pmgA inactivation mutant was constructed by inserting the accC1 gene
cassette, conferring gentamicin resistance, into the unique SpeI site within the pmgA
coding region. The complete segregation of the mutant allele from that of wild type was
demonstrated by PCR analysis (Fig 5.7). PCR using the primers designed to target and
amplify a 0.8-kbp of the pmgA locus resulted in the amplification of a 0.8-kbp fragment
when genomic DNA isolated from wild-type cells was used as the template. No 0.8-kb
product was detectable for the PCR product when genomic DNA isolated from the

214
pmgA::aacC1 transformant was used as the template. Instead, a product of 1.9 kb was
detected. The size difference between the two products corresponds the size of the aacC1
gene cassette, which demonstrates that the pmgA::aacC1 transformant is homozygous
and free from the pmgA wild-type allele.
A preliminary growth analysis was carried out in the absence of glucose and in
the presence of glucose at pH 7.0 and pH 8.0. The results showed that the pmgA mutant
grew similarly to wild type in the absence of glucose at pH 7.0 and at pH 8.0 (by visual
inspection). In the presence of glucose, the pmgA mutant grew similarly to wild type in
medium at pH 8.0; however, its growth was found to be severely impaired in the presence
of glucose at pH 7.0. This apparent similarity in the growth behavior between the
tocopherol and pmgA mutants in response to glucose and pH suggests that α-tocopherol
and PmgA play roles in the same regulatory pathway. PmgA shows similarity to
RsbW/RsbT, protein serine/threonine kinase, in B. subtilis (Price CW, 2000; Woodbury
et al., 2004). RsbW functions as an anti-SigB factor, while RsbT functions as an activator
of RsbU, which in turns activates SigB in the signal transduction cascade that regulates
the activity SigB. They act together with other protein phosphatases and kinases
including RbsU, an Slr2031 homologue (Price CW, 2000; Woodbury et al., 2004). More
than 6 Rsb-like proteins were found to be conserved in the genome of Synechocystis sp.
PCC 6803, and PmgA, Slr2031, and Ssr1600 are also found to be present in the genomes
of Thermosynechococcus elongatus BP-1, Nostoc sp. PCC 7120, Nostoc punctiforme,
Trichodesmium erythraeum, Synechococcus sp. PCC 7002, Prochlorococcus marinus sp.
MED4, P. marinus sp. MIT9313, P. marinus sp. SS120, and Synechococcus sp.

215
WH8102. Thus, it is highly plausible that a pathway similar to the Rsb pathway in B.
subtilis is present in cyanobacteria and regulates the balance between carbon, nitrogen
and sulfur metabolism.

216
DISCUSSION
The potential involvement of α-tocopherol in macronutrient metabolism was
investigated. RT-PCR analyses performed on the cells grown at pH 7.0 and 8.0 in the
absence and in the presence of glucose demonstrated that the transcripts of the alternative
sigma factors accumulate in the slr1736 mutant in response to glucose, which indicated
that the slr1736 mutant is experiencing a variety of stresses when grown in the presence
of glucose (Caslake et al., 1997; Gruber and Bryant, 1998; Huchauf et al., 2000; Muro-
Pastor et al., 2001; Imamura et al., 2003a,b). The nblA and sigE transcripts, known to
accumulate in response to nitrogen limitation (Mulo-Pastor, 2001; Richaud 2001), also
accumulated in response to glucose in the slr1736 mutant. The sbpA transcript, known to
accumulate in response to sulfur limitation (Laudenbach and Grossman, 1992), also
accumulated in response to glucose in the slr1736 mutant. In combination with the results
obtained for the PBP content as described in Chapter 4, these results demonstrate that
slr1736 mutant is experiencing macronutrient starvation when grown in the presence of
glucose, and that α-tocopherol is somehow influencing carbon, nitrogen, and sulfur
metabolism in Synechocystis sp. PCC 6803.
When cells were grown at pH 7.0 in the presence of glucose, the level of
transcripts of sigA, ndhF4, cpcA, and carboxysome genes (rbcL, ccmK1, ccmL) decreased
substantially. These genes were constitutively expressed, and their products play
housekeeping roles for the survival of cyanobacteria. Thus, it is possible that the
reduction of the ndhF4, and cpcA, and carboxysome transcripts is caused by the reduction
of the cellular content of SigA (Pierce et al., 1989; Caslake and Bryant, 1996; Gruber and

217
Bryant, 1997). Because the functions of SigA and RuBisCO are absolutely indispensable
for the survival of cyanobacteria, the observed pH-dependent, glucose-induced
transcriptional misregulation of sigA and rbcL is a likely cause of the cell death of the
tocopherol mutant in the presence of glucose in medium at pH 7.0.
The mechanism of the pH-dependent, glucose-induced misregulation of sigA
transcription has yet to be determined. A study has suggested that sigA transcription is
regulated by the activity of the photosynthetic electron transport chain, and in the
presence of DCMU or in the dark the amount of sigA transcripts is rapidly lowered
(Tuominen et al., 2003). Interestingly, the activity of PS II, and thus the activity of the
photosynthetic electron transport chain, is reduced in a pH-dependent manner in the
tocopherol mutant in response to glucose, and PS II activity was completely lost at pH
7.0 (see Chapter 4, Fig 4.17). The PS II content in the tocopherol mutant under these
conditions appeared to be largely unaltered based on the contents of the D1 and PsbO
proteins (see Chapter 4, Fig 4.10). Thus, the inactivation of PS II is thought to be related
either to the loss of electron transfer cofactors or to the generation of metabolites that
inhibit the activity of PS II. Decades of studies and recent X-ray crystallographic
structures of the PS II complexes from the cyanobacterium Synechococcus elongatus
have confirmed the presence of one bicarbonate molecule in the vicinity of the non-heme
iron located between the two bound plastoquinone molecules (QA and QB) (Blubaugh and
Govindjee, 1986; Petrouleas et al., 1994; Ferreira et al., 2004, see also reviews by
Blubaugh and Govindjee, 1988; Govindjee and van Rensen, 1993; van Rensen et al.,
1999, and references therein ). It has been demonstrated that this bicarbonate molecule is

218
displaced in vitro by other molecules such as formate, NO, glycolate, glyoxylate, and
oxalate (Petrouleas et al., 1994). Glycolate is a metabolite generated by the oxygenase
activity of RuBisCO, which occurs when the intracellular CO2 concentration is
substantially reduced. Although, there is no evidence suggesting a significant reduction in
the intracellular Ci pool in the tocopherol mutant, this possibility needs to be examined
nevertheless.
Perhaps the most exciting outcome of the RT-PCR analyses is the discovery that
the expression of slr2031 and CCM genes (ndhF3, ndhR, sbtA) is constitutively down-
regulated in the tocopherol mutant in the presence and absence of glucose both at pH 7.0
and 8.0. This direct effect of the loss of α-tocopherol on gene expression is a strong
indication that α-tocopherol is involved in transcriptional regulation in cyanobacteria.
Because the function of these products are directly related to macronutrient metabolism,
this further leads to the conclusion that α-tocopherol is involved in the regulation of the
macronutrient metabolism as a transcriptional regulator in cyanobacteria. It is noteworthy
that, in mammalian systems, it has been demonstrated that α-tocopherol is directly
involved in transcriptional regulation of the genes encoding liver collagen αI, α-
tocopherol transfer protein, and α-tropomyosin collagenase (Yamaguchi et al., 2001;
Azzi et al., 2002). The mechanisms of these transcriptional regulation processes are not
completely understood; however, it is a reasonable assumption that it involves a
modulation of a signal transduction cascade that regulates the expression of these genes.
Indeed, α-tocopherol has been suggested to modulate the cell signaling pathway by
modulation of the phosphorylation state of protein kinase Cα in rat smooth-muscle cells,

219
possibly via phosphorylation of protein phosphatase 2A (Ricciarelli et al., 1998). In light
of the observed involvement of α-tocopherol in cell signaling in the animal system, it is
intriguing to recognize the phenotypic similarity between the tocopherol mutant and the
pmgA mutant in Synechocystis sp. PCC 6803. As mentioned earlier, the pmgA gene has
been demonstrated to be a determinant of glucose tolerance in this organism (Hiahara and
Ikeuchi, 1997), and its deduced amino acid sequence shows similarity to a
serine/threonine protein kinase, which has been characterized as the anti-sigma factor
(RsbW) or the activator for the anti-RsbW (RsbT) in the regulatory pathway of SigB in B.
subtilis (Price, 2000; Woodbury et al., 2004). Thus, it is highly plausible that α-
tocopherol plays a regulatory role through this Rsb-like pathway in Synechocystis sp.
PCC 6803 as summarized in Fig 5.9. This is the first time, to the best of my knowledge,
that the involvement of α-tocopherol in transcriptional regulation has been
experimentally demonstrated in the oxygenic phototrophs. This newly discovered role of
α-tocopherol in Synechocystis sp. PCC 6803 thus strongly indicates that α-tocopherol
plays a regulatory role in addition to a role as a bulk antioxidant in some cyanobacteria.
The next question is whether such a regulatory role is evolutionarily
conserved among green algae and plants. Recent studies have shown that a mutant
in maize, originally isolated because of its deficiency in sucrose transport through
plasmodesmata, is also deficient in tocopherol cyclase (Porfirova et al., 2002;
Sattler et al., 2003). This suggests that α-tocopherol plays a role in sugar
metabolism in maize. It is therefore highly plausible that α-tocopherol has similar
regulatory functions in macronutrient metabolism in plants.

220
SUMMARY
It was shown that a α-tocopherol mutant responds to the presence of glucose as
though nitrogen and sulfur were limiting in the environment. This conclusion is based on
the accumulation of stress-responsive gene transcripts. When glucose is present in growth
medium at pH 7.0, the amounts of the sigA and rbcL transcripts diminished after 4 h.
Because the amount of these transcripts remained unaltered when cells were grown in the
absence of glucose or in the presence of glucose pH 8.0, it was concluded that the pH-
dependent glucose-induced misregulations of the sigA and rbcL are largely responsible
for the glucose-induced lethality of the tocopherol mutants under these conditions. The
transcription of metabolic genes (slr2031, ndhF3, ndhR, and sbtA) is shown to be
constitutively down regulated under all conditions tested in the tocopherol mutant. This
result demonstrates that α-tocopherol is involved in transcriptional regulation of these
metabolic genes in Synechocystis sp. PCC 6803. Thin-section electron micrographs of the
tocopherol mutant showed that large amounts of glycogen granules accumulated in the
cells. The targeted interruption of the pmgA gene (a putative anti sigma factor
homologous to RsbW) resulted in the similar pH-dependent glucose-induced lethal
phenotype. Based on theses results, it was concluded that α-tocopherol is involved as a
regulatory molecule in the global regulation of carbon, sulfur, and nitrogen metabolism in
Synechocystis sp. PCC 6803.

221
MATERIALS AND METHODS
Strains and Growth Conditions
The wild-type strain used for transformation and further analyses was a glucose-
tolerant stain of Synechocystis sp. PCC 6803 (Williams 1988). Medium BHEPES, pH
8.0, was used for maintenance of the wild type and mutant strains. This medium is
prepared by supplementing BG-11 medium (Stainer et al., 1971) with 4.6 mM of HEPES
[4-(2-hydroxyethyl)piperazine-N’-(2-ethnesulfonic acid)]-KOH and 18 mg L-1 ferric
ammonium citrate. Medium BHEPES40 is prepared by supplementing the medium
BHEPES with 40 mM HEPES. Prior to all growth measurements and the preparation of
cells for subsequent analyses, cells were transferred from solid to liquid medium
BHEPES and allowed to grow exponentially for several generations in the absence of
glucose. For the determination of growth characteristics, late-exponential phase cultures
were diluted in fresh liquid medium BHEPES40, pH 7.0 or 8.0, to an OD730 nm of
approximately 0.05 cm-1. The diluted cultures were grown at 32 °C with continuous
bubbling with air containing 1% (v/v) CO2. For growth in the presence of glucose, the
medium was supplemented with 5 mM glucose. The light intensity was 50 µE m-2 s-1,
unless otherwise noted. For isolation of RNA, late-exponential phase cultures were
diluted in fresh liquid medium BHEPES40 to an OD730 nm of approximately 0.3 cm-1 in a
total volume of 80 mL. The diluted cultures were grown under the same conditions as
described above.

222
Generation of the slr2031 and the pmgA Mutants
The 5’-terminal portion of the slr2031 locus was amplified by PCR using primers
designed to target this genomic region [slr2031-F, 5’-
AGGAGTTGGTGGCTAAGCTTTACCGGGAACAAA-3’, including an engineered
HindIII cleavage site (underlined); slr2031-R, 5’-
TGACAAAGCGATGGGAATTCTCCAGGTCGGCA-3’, including an engineered
EcoRI cleavage site (underlined)]. The pmgA locus was amplified by PCR using primers
designed to target this genomic region [pmgA-F, 5’-
TTCTCTGTGCCGAAAGCTTCTATG-3’, including an engineered HindIII cleavage site
(underlined); pmgA-R, 5’-CACCATGGTGGCGAATTCAGCC-3’, including an
engineered EcoRI cleavage (underlined)]. The amplified DNA fragments were digested
with HindIII and EcoRI and ligated with pUC19 obtained after digestion of pUC19 with
HindIII and EcoRI. An EcoRI fragment of pSRA2, containing the aadA gene conferring
spectinomycin resistance, was inserted into the unique MfeI site within the 5’ slr2031
coding region. An XbaI fragment of pMS266, containing the accC1 gene conferring
gentamicin resistance, was inserted into the unique SpeI site within the pmgA coding
region. The resulting constructs were linearized after digestion with EcoRI and used to
transform wild-type Synechocystis sp. PCC 6803 cells. Selection of transformants was
carried out on solid medium BHEPES, pH 8.0, in the presence of 50 µg mL-1
spectinomycin or in the presence of 20 µg mL-1 gentamicin at 27 °C, under a moderate
light intensity (~50 µE m-2 s-1). PCR analyses were carried out using the same sets of
primers used for the cloning.

223
Isolation of Total RNA and RT-PCR Analyses
Total RNA was isolated from cells using the Mini-to-Midi RNA isolation kit
(Invitrogen corp., Carlsbad, CA) followed by an RNase-free DNase treatment and
repurification by using the same kit. The final concentration of the purified RNA
preparations was 50 ng mL-1 in RNase free double-distilled water; the RNA samples were
stored at -80°C until being used. RT-PCR reactions were carried out by using the One-
Step RT-PCR kit (Qiagen Inc., Valencia, CA) for detecting the rnpB, spbA, sigC, sigE,
sigI, ndhF4, ndhF3, ndhR, sbtA and slr2031 transcripts. The reaction volume was 25 µL,
and 4 ng µL-1 of the RNA preparation was used for each reaction. For detecting the other
transcripts, 400 ng of total RNA was first converted to cDNA by a reverse transcription
reaction using M-MLV reverse transcriptase (Promega, Madison, WI). The reaction
volume was 20 µL for each sample. The resulting cDNA was diluted with 80 µL of TE
buffer, and stored at –20 °C until being used. Two microliters of cDNA stock were used
for each PCR reaction (final volume, 25 µL) using Hot-Start Taq polymerase (Qiagen
Inc., Valencia, CA). RNasin (Promega, Madison, WI), an RNase inhibitor, was present
in the one-step RT-PCR reactions and during the cDNA synthesis.
Transmission Electron Microscopy
Cells grown in the absence or in the presence of glucose for 24 h were harvested
and immediately fixed in a 2.5 % glutaraldehyde solution prepared in a 0.1 M cacodylate
buffer, pH 7.4, over night at 4°C. The fixed cells were washed in the 0.1 M cacodylate
buffer for three times at room temperature, and subjected to the second fixation in a 1%
osmium tetroxide solution prepared in the 0.1 M cacodylate buffer for 1. 5 h at room

224
temperature. The cells were washed twice in the 0.1 M cacodylate buffer and then once in
double distilled water. After incubation in a 2 % uranyl acetate solution for 2 h at room
temperature, the cells were washed in the following concentration of ethanol: 50% (v/v),
70% (v/v), 90% (v/v), 95% (v/v) ethanol in water followed by two washes in 100% (v/v)
ethanol. Final washes were performed three times in 100% ethanol (EM grade) and then
three times in acetone at room temperature before infiltration. The washed cells were
incubated in the presence of the following concentration of the resin Spurr (Spurr 1969)
over night at room temperature: 50% (v/v) and 72 % (v/v) in acetone and twice in 100%
(v/v) Spurr. The samples were then polymerized at 60°C over night. Thin sections (ca.
50-60 nm thickness) were stained with 2 % (v/v) uranyl acetate before examination under
a JEM 1200 EXII transmission electron microscope (Joel USA Inc., Peabody, MA,
USA).

225
REFERENCES
Azzi A, Ricciarelli R, Zingg JM (2002) Non-antioxidant molecular functions of α-
tocopherol (vitamin E). FEBS Lett 519: 8-10
Blubaugh DJ, Govindjee (1986) Bicarbonate, not CO2, is the species required for the
stimulation of Photosystem II electron transport. Biochim Biophys Acta 848: 147-151
Blubaugh DJ, Govindjee (1988) The molecular mechanism of the bicarbonate effect at
the plastoquinone reductase site of photosynthesis. Photosynth Res 19: 85-128
Caslake L, Bryant DA (1996) The sigA gene encoding the major sigma factor of RNA
polymerase from the marine cyanobacterium Synechococcus sp. strain PCC 7002:
cloning and characterization. Microbiology 142: 347-357
Caslake L, Gruber TM, Bryant DA (1997) Expression of two alternative sigma factors of
Synechococcus sp. strain PCC 7002 is modulated by carbon and nitrogen stress.
Microbiology 143: 3807-3818
Chan SS, Monteiro HP, Schindler F, Stern A, Junqueira VB. (2001) α-Tocopherol
modulates tyrosine phosphorylation in human neutrophils by inhibition of protein kinase
C activity and activation of tyrosine phosphatases. Free Radic Res 35: 843-856
Chandra V, Jasti J, Kaur P, Betzel C, Srinivasan A, Singh TP (2002) First structural
evidence of a specific inhibition of phospholipase A2 by α-tocopherol (vitamin E) and its

226
implications in inflammation: crystal structure of the complex formed between
phospholipase A2 and α-tocopherol at 1.8 Å resolution. J Mol Biol 320: 215-222
Collier JL, Grossman AR (1994) A small polypeptide triggers complete degradation of
light-harvesting phycobiliproteins in nutrient-deprived cyanobacteria. EMBO J 13: 1039-
1047
Collins MD, Jones D (1981) Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiol Rev 45: 316-354
Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S (2004) Architecture of the
photosynthetic oxygen-evolving center. Science 303: 11831-1838
Figge RM, Cassier-Chauvat C, Chauvat F, Cerff R (2001) Characterization and analysis
of an NAD(P)H dehydrogenase transcriptional regulator critical for the survival of
cyanobacteria facing inorganic carbon starvation and osmotic stress. Mol Microbiol 39:
455-468
Govindjee, van Rensen JJS (1993) Photosystem II reaction center and bicarbonate. In:
Deisenhofer J, Norris JR (eds), The photosynthetic reaction center, Vo1 1, pp 357-389.
Academic Press. Location.
Grossman AR, Schaefer MR, Chiang GG, Collier JL (1994) The responses of
cyanobacteria to environmental conditions, light and nutritions. In: Bryant DA (ed), The
Molecular Biology of Cyanobacteria. pp 641-675. Kluwer Academic Pubisher, Dordrecht

227
Gruber T, Bryant DA (1997) Molecular systematic studies of eubacteria, using sigma 70-
type sigma factors of group 1 and group 2. J Bacteriol 179: 1734-1747
Gruber T, Bryant DA (1998) Characterization of the alternative σ-factors SigD and SigE
in Synechococcus sp. strain PCC 7002. SigE is implicated in transcription of post-
exponential-phase-specific genes. Arch Microbiol 169: 211-219
Hihara Y, Ikeuchi M (1997) Mutation in a novel gene required for photomixotrophic
growth leads to enhanced photoautotrophic growth of Synechocystis sp. PCC 6803.
Photosynth Res 53: 243-252
Huckauf J, Nomura C, Forchhammer K, Hagemann M (2000) Stress responses of
Synechocystis sp. strain PCC 6803 mutants impaired in genes encoding putative
alternative sigma factors. Microbiology 146: 2877-2889
Imamura S, Asayama M, Takahashi H, Tanaka K, Takahashi H, Shirai M (2003a)
Antagonistic dark/light-induced SigB/SigD, group 2 sigma factors, expression through
redox potential and their roles in cyanobacteria. FEBS Lett 554: 357-362
Imamura S, Yohihara S, Nakano S, Shiozaki N, Yamada A, Tanaka K, Takahashi H,
Asayama M, Shirai M (2003b) Purification, characterization, and gene expression of all
sigma factors of RNA polymerase in a cyanobacterium. J Mol Biol 325: 857-872
Kamal-Eldin A, Appelqvist L (1996) The chemistry and antioxidant properties of
tocopherols and tocotrienols. Lipids 31 :671-701

228
Kaplan A, Scherer S, Lerner M (1989) Nature of the light-induced H+ efflux and Na+
uptake in cyanobacteria. Plant Phyiol. 89: 1220-1225
Katoh A, Lee K-S, Fukuzawa H, Ohyama K, Ogawa T (1995) A putative CO2 transporter
in Synechocystis PCC 6803 driven by NADPH dehydrogenase-mediated Photosystem I
cyclic electron flow. In: Mathis P (ed.) Photosynthesis: from Light to Biosphere, Vol. II,
pp 807-812. Kluwer Academic Pubisher, Dordrecht
Katoh A, Lee K-S, Fukuzawa H, Ohyama K, Ogawa T (1996a) cemA homologue
essential to CO2 transport in the cyanobacterium Synechocystis PCC 6803. Proc Natl
Acad Sci USA 93: 4006-4010
Katoh A, Sonoda M, Katoh H, Ogawa T (1996b) Absence of light-induced proton
extrusion in a cotA-less mutant of Synechocystis sp. strain PCC 6803. J Bacteriol 178:
5452-5455
Klughammer B, Sültemeyer D, Badger MR, Price GD (1999) The involvement of
NAD(P)H dehydrogenase subunits, NdhD3 and NdhF3, in high-affinity CO2 uptake in
Synechococcus sp. PCC 7002 gives evidence for multiple NHD-1 complexes with
specific roles in cyanobacteria. Mol Microbiol 32: 1305-1315
Laudenbach DE, Grossman AR (1991) Characterization and mutagenesis of sulfur-
regulated genes in a cyanobacterium: evidence for function in sulfate transport. J
Bacteriol 173: 2739-2750

229
Lockau W and Pfeffer S (1982) A cyanobacterial ATPase distinct from the coupling
factor of photophosphorylation. Z Naturforsch Teil C 37: 658-664
McGinn PJ, Price GD, Maleszka R, Badger MR (2003) Inorganic carbon limitation and
light control the expression of transcripts related to the CO2-concentrating mechanisms in
the cyanobacterium Synechocystis sp. strain PCC6803. Plant Physiol 132: 1-12
Muro-Pastor AM, Herrero A, Flores E (2001) Nitrogen-regulated group 2 sigma factor
from Synechocystis sp. strain PCC 6803 involved in survival under nitrogen stress. J
Bacteriol 183: 1090-1095
Ogawa T and Kaplan A (1987) The stoichiometry between CO2 and H+ fluxes involved
in the transport of inorganic carbon in cyanobacteria. Plant Physiol 83: 888-891
Petroulesas V, Deligiannakis Y, Diner BA (1994) Binding of carboxylate anions at the
none-heme Fe (II) of PS II. II. Competition with bicarbonate and effects on the QA/QB
electron transfer rate. Biochim Biophys Acta 1188: 271-277
Pierce J, Carlson TJ, Williams JG (1989) A cyanobacterial mutant requiring the
expression of ribulose bisphosphate carboxylase from a photosynthetic anaerobe. Proc
Natl Acad Sci USA 86: 5753-5757
Porfirova S, Bergmüller E, Tropf S, Lemke R, Dörmann P (2002) Isolation of an
Arabidopsis mutant lacking vitamin E and identification of a cyclase essential for all
tocopherol biosynthesis. Proc Natl Acad Sci USA 99: 12495-12500

230
Price GD, Howitt SM, Harrison K, Badger MR (1993) Analysis of a genomic DNA
region from the cyanobacterium Synechococcus sp. strain PCC7942 involved in
carboxysome assembly and function. J Bacteriol 175: 2871-2879
Price CW (2000) Protective function and regulation of the general stress response in
Bacillus subtilis and related Gram-positive bacteria. In: Storz G, Hengge-Aronis R (eds),
Bacterial Stress Response. pp 179-197. ASM Press, Washington DA
Ricchiarelli R, Tasinato A, Clément S, Özer NK, Boscoboinik D, Azzi A (1998) α-
Tocopherol specifically inactivates cellular protein kinase Cα by changing its
phosphorylation state. Biochem J 334: 243-249
Ricchiarelli R, Zingg JM, Azzi A (2002) The 80th anniversary of vitamin E: Beyond its
antioxidant properties. Biol Chem 383: 457-465
Richaud C, Zabulon G, Joderr A, Thomas J-C (2001) Nitrogen and sulfur starvation
differentially affects phycobilisomes degradation and expression of the nblA gene in
Synechocystis strain PCC 6803. J Bacteriol 183: 2989-2994
Sattler SE, Cahoon EB, Coughlan SJ, DellaPenna D (2003) Characterization of
tocopherol cyclases from higher plants and cyanobacteria: evolutionary implications for
tocopherol synthesis and function. Plant Physiol 132: 2184-2195
Sattler S, Gillilanda LU, Magallanes-Lundback M, Pollard M, DellaPenna D (2004)
Vitamin E is essential for seed longevity and preventing lipid peroxidation during
germination. Plant Cell 16: 1419-1432

231
Shibata M, Ohkawa H, Kaneko T, Fukuzawa H, Tabata S, Kaplan A, Ogawa T (2001)
Distinct constitutive and low-CO2-induced CO2 uptake systems in cyanobacteria: genes
involved in and their phylogenetic relationship with homologous genes in other
organisms. Proc Natl Acad Sci USA 98: 11789-11794
Shibata M, Katoh H, Sonoda M, Ohkawa H, Shimoyama M, Fukuzawa H, Kaplan A,
Ogawa T (2002) Genes essential to sodium-dependent bicarbonate transport in
cyanobacteria. J Biol Chem 277: 18658-18664
Smith AJ (1982) Modes of cyanobacterial carbon metabolism. In: Carr NG, Whitton BA
(eds) The Biology of Cyanobacteria. Blackwell Scientific Publications, Oxford
Sonoda M, Kitano K, Katoh A, Katoh H, Ohkawa H, Ogawa T (1997) Size of cotA and
identification of the gene product in Synechocystis sp. strain PCC 6803. J Bacteriol 179:
3845-3850
Stainer RY, Kunisawa R, Mandel M, Cohen-Bazire G (1971) Purification and properties
of unicellular blue-green algae (Order Chroococcales). Bact Rev 35: 171-205
Stainer G (1988) Fine structgure of cyanobacteria. Method Enzymol 167: 157-172
Threlfall DR, Whistance GR (1971) Biosynthesis of isoprenoid quinones and
chromanols. In: Goodwin TW (ed) Aspects of Terpenoid Chemistry and Biochemistry,
pp. 357-404. Academic Press, London

232
Trebst A, Depka B, Höllander-Czytko H (2002) A specific role for tocopherol and of
chemical singlet oxygen quenchers in the maintenance of Photosystem II structure and
function in Chlamydomonas reinhardtii. FEBS Lett 516: 156-160
Tuominen I, Tyystjarvi E, Tyystjarvi T (2003) Expression of primary sigma factor (PSF)
and PSF-like sigma factors in the cyanobacterium Synechocystis sp. strain PCC 6803. J
Bacteriol 185: 1116-1119
van Rensen JJS, Xu C, Govindjee (1999) Role of bicarbonate in Photosystem II, the
water-plastoquinone oxido-reductase of plant photosynthesis. Physiol Plant 105: 585-592
Williams JGK (1988) Construction of specific mutations in Photosystem-II
photosynthetic reaction center by genetic engineering methods in Synechocystis 6803.
Meth Enzymol 167: 766-778
Woodbury RL, Luo T, Grant L, Haldenwang WG (2004) Mutational analysis of RsbT, an
activator of the Bacillus subtilis stress response transcription factor, σB. J Bacteriol 186:
2789-2797
Yamaguchi J, Iwamoto T, Kida S, Masushige S, Yamada K, Esashi T (2001) Tocopherol-
associaated protein is a ligand-dependent transcriptional activator. Biochem Biophys Res
Commun 285: 295-299

233
Fig 5.1: Time-course RT-PCR analysis of the cpcA, nblA, and sbpA transcripts in wild type and slr1736 mutant grown at pH 7.0 and pH 8.0. Cell were grown in the absence of glucose (shown at 0 h) and in the presence of glucose for 4, 8, and 24 h, at 1% (v/v) CO2, 32 °C at 50 µE m-2 s-1
.

234
Fig 5.2: Time-course RT-PCR analysis of alternative sigma factor transcripts in wild type and slr1736 mutant grown at pH 7.0 and pH 8.0. Cell were grown in the absence of glucose (shown at 0 h) and in the presence of glucose for 4, 8, and 24 h, at 1% (v/v) CO2, 32 °C at 50 µE m-2 s-1
.

235
Fig 5.3: Time-course RT-PCR analysis of transcripts for sigA, sigG and genes involved in Ci uptake and fixation in wild type and slr1736 mutant grown at pH 7.0 nd pH 8.0. Cell were grown in the absence of glucose (shown at 0 h) and in the presence of glucose for 4, 8, and 24 h, at 1% CO2 (v/v), 32 °C at 50 µE m-2 s-1
.
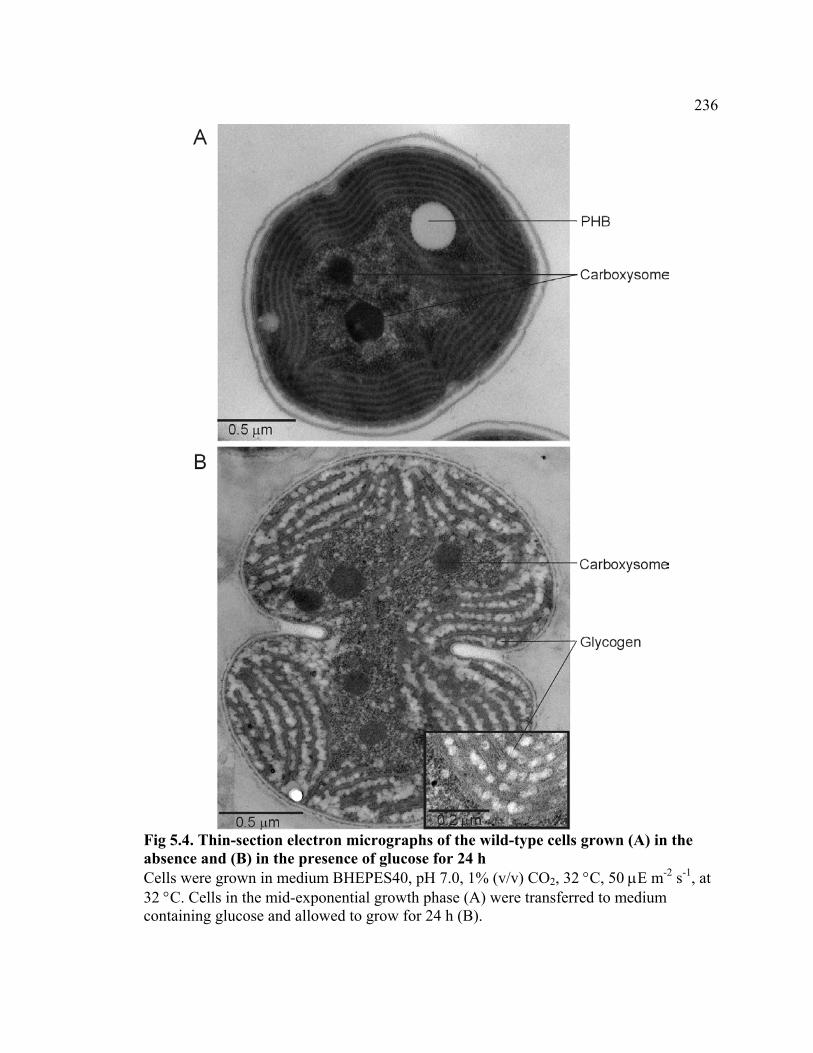
236
Fig 5.4. Thin-section electron micrographs of the wild-type cells grown (A) in the absence and (B) in the presence of glucose for 24 h Cells were grown in medium BHEPES40, pH 7.0, 1% (v/v) CO2, 32 °C, 50 µE m-2 s-1, at 32 °C. Cells in the mid-exponential growth phase (A) were transferred to medium containing glucose and allowed to grow for 24 h (B).
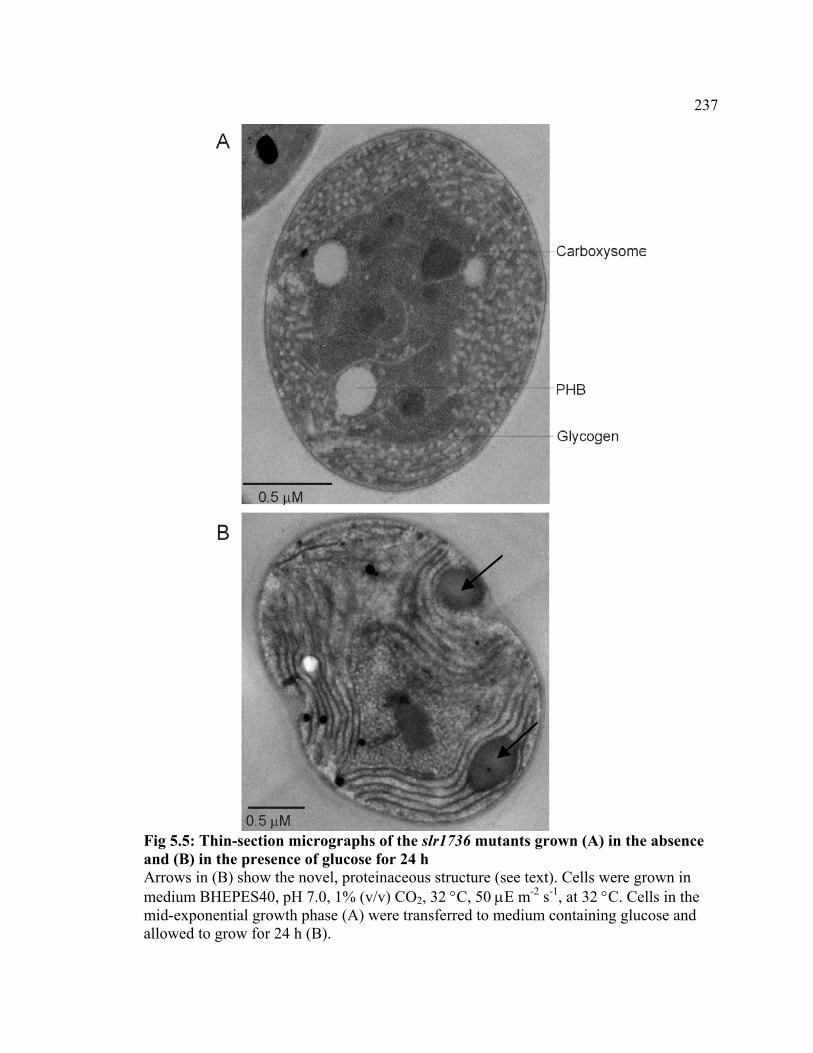
237
Fig 5.5: Thin-section micrographs of the slr1736 mutants grown (A) in the absence and (B) in the presence of glucose for 24 h Arrows in (B) show the novel, proteinaceous structure (see text). Cells were grown in medium BHEPES40, pH 7.0, 1% (v/v) CO2, 32 °C, 50 µE m-2 s-1, at 32 °C. Cells in the mid-exponential growth phase (A) were transferred to medium containing glucose and allowed to grow for 24 h (B).

238
Fig 5.6: Time-course RT-PCR analyses of the slr2031 and various CCM gene transcripts in wild type and slr1736 mutant grown at pH 7.0 and pH 8.0, 1% (v/v) CO2, 32 °C at 50 µE m-2 s-1. Cell were grown in the absence of glucose (shown at 0 h) and in the presence of glucose for 4, 8, and 24 h.
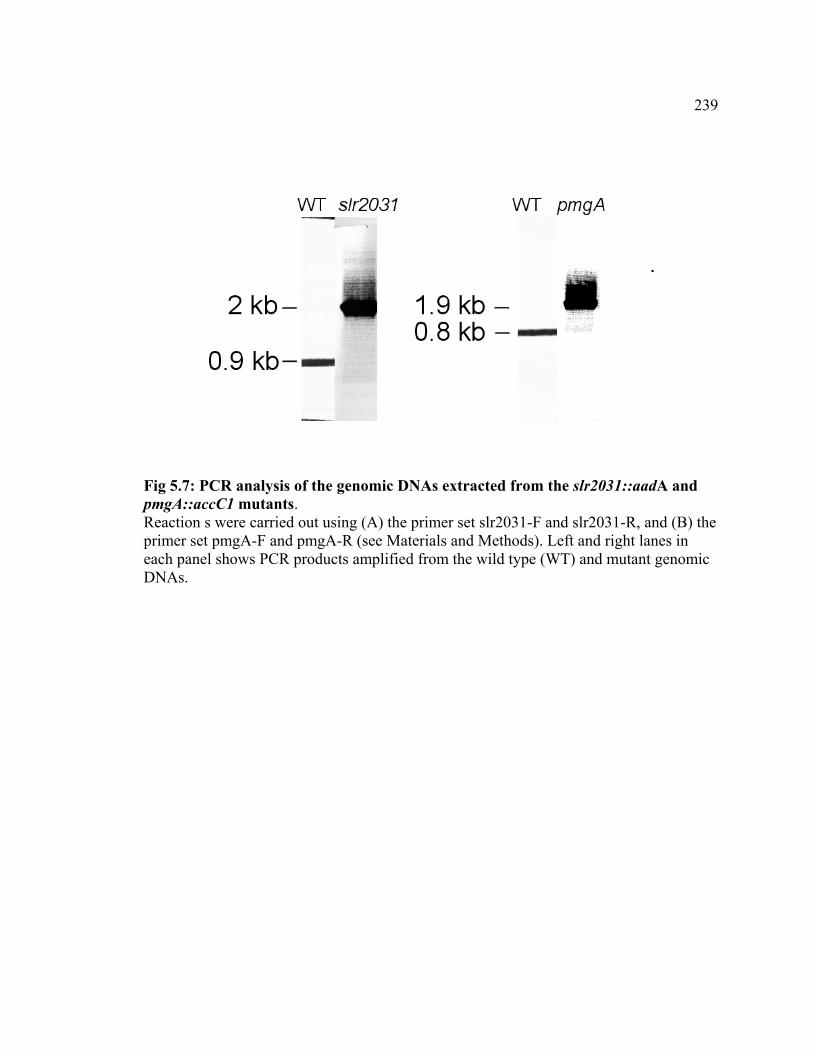
239
Fig 5.7: PCR analysis of the genomic DNAs extracted from the slr2031::aadA and pmgA::accC1 mutants. Reaction s were carried out using (A) the primer set slr2031-F and slr2031-R, and (B) the primer set pmgA-F and pmgA-R (see Materials and Methods). Left and right lanes in each panel shows PCR products amplified from the wild type (WT) and mutant genomic DNAs.

240
Fig 5.8: Cultures of wild type, slr1736, slr2031, and pmgA mutants of Synechocystis sp. PCC 6803 grown in the presence of glucose (A) at pH 8.0 and (B) at pH 7.0 Cell were grown at 1% (v/v) CO2, 32 °C at 50 µE m-2 s-1
.

241
Fig 5.9: Summary and a model of the non-antioxidant role of α-tocopherol in Synechocystis sp. PCC 6803 A Rsb-like regulatory pathway, which involves PmgA, appears to exist in Synechocystis sp. PCC 6803 and possibly in other cyanobacteria (see text). In this model, α-tocopherol either directly or indirectly interacts with the Rsb-like pathway, possibly via the function of PmgA. Through such interaction, it is hypothesized that α-tocopherol affects the transcriptional regulation of some metabolic genes, which may be required for photomixotrophic growth of this organism.

242
Chapter 6
Comparative Genome Analyses and Genetic Studies of the Plastoquinone
Biosynthesis Pathway in Cyanobacteria
Publication:
Yumiko Sakuragi and Donald A. Bryant, manuscript in preparation

243
ABSTRACT
The biosynthetic pathway of plastoquinone-9 (PQ-9) was studied by means of
comparative genome analyses in 14 cyanobacteria. The presence of genes homologous to
ubiA, ubiC, ubiD, ubiE, ubiH, and ubiX involved in ubiquinone-8 (UQ) biosynthesis in
Escherichia coli was detected in cyanobacterial genomes, which suggests that PQ-9
biosynthesis in cyanobacteria occurs from 4-hydroxybenzoate through a pathway similar
to that for the UQ biosynthesis in proteobacteria. Attempted insertional inactivation of
the ubiA, ubiX, and ubiH homologs in Synechocystis sp. PCC 6803 resulted in an
incomplete segregation of the alleles, indicating that the products of these genes are
essential for the survival of this cyanobacterium. Of the several homologs of ubiE genes
found in the cyanobacterial genomes, only two are conserved among 14 species.
However, inactivation mutagenesis of these two conserved ubiE homologs, sll1653 and
sll0418, as well as others, including sll0829, slr1039, sll0487, slr0407, and slr1618 had
little effect on the cellular PQ-9 content. These results therefore demonstrate that these
UbiE homologs are probably not involved in PQ-9 biosynthesis in cyanobacteria. After
exhaustive database searching, the presence of ubiC homologs, which are closely related
to the plastid ycf21 gene, was detected in many cyanobacterial genomes. Taking these
observations together, a hypothetical PQ-9 biosynthetic pathway in cyanobacteria is
proposed.

244
ABBREVIATIONS
PQ-9 plastoquinone-9
UQ-8 ubiquinone-8
MSBQ 2-methyl-6-solanyl-1,4-benzoquinone
MPBQ 2-methyl-6-phytyl-1,4-benzoquinone
ORF open reading frame
4-HBA 4-hydroxybenzoate
PCR polymerase chain reaction
HPLC high performance liquid chromatography
SAM S-adenosyl-L-methionine

245
INTRODUCTION
Plastoquinone (PQ-9, 2-3-dimethyl-5-solanyl-1,4-benzoquinone) is an isoprenoid
quinone that is only synthesized in oxygenic phototrophs including cyanobacteria, algae,
and higher plants (Threlfall and Whistance, 1971; Collins and Jones, 1981). It serves as a
one-electron carrier (QA) as well as a two-electron carrier (QB) within Photosystem II and
stabilizes the charge separation between P680+ and QB- upon photoexcitation (Barry et
al., 1994; Diner and Babcock, 1994; Bricker and Ghanotakis, 1996; Britt, 1996). PQ-9
also shuttles electrons from NADH dehydrogenase to cytochrome b6f complex, therefore
serving as an essential membrane-associated, lipid-soluble electron carrier both in
photosynthetic and respiratory electron transport chains.
Studies of PQ-9 biosynthesis have been carried out since the 1960’s in higher
plants by using isotopic tracers and direct enzyme activity assays (see a review by
Threlfall and Whistance, 1970) as well as by genetic methods (Collakova and
DellaPenna, 2001; Shintani et al., 2002; Cheng et al., 2003). The combined results have
shown that PQ is synthesized from homogentisate after two enzymatic reactions:
condensation of solanyl diphosphate and homogentisate by a solanyl transferase resulting
in 2-methyl-6-solanyl-1,4-benzoquinone (MSBQ), and a methylation reaction at the C3
position of the benzoquinoid ring system by MSBQ methyltransferase. This MSBQ
methyltransferase is also responsible for methylation of 2-methyl-6-phytyl-1,4-
benzoquinone (MPBQ) in α-tocopherol biosynthesis, and therefore it is a bifunctional
enzyme in higher plants (Shintani et al., 2002; Cheng et al., 2003). Given the frequent
similarity of biochemical pathways in plants and cyanobacteria, it might be assumed that

246
the same pathway functions in cyanobacteria. However, recent studies (Dänhardt et al.,
2002; Cheng et al., 2003) showed that neither the synthesis of homogentisate nor the
presence of MPBQ methyltransferase activity is required for PQ-9 biosynthesis in the
cyanobacterium Synechocystis sp. PCC 6803. These results suggested that PQ-9
biosynthesis in cyanobacteria is different from that in higher plants and that the PQ-9
biosynthetic pathways convergently evolved in cyanobacteria and higher plants (Cheng et
al., 2003).
How is PQ synthesized in cyanobacteria? A preliminary inspection of the whole
genome sequence of Synechocystis sp. PCC 6803 has shown the presence of ORFs that
show sequence similarity to enzymes responsible for ubiquinone-8 (UQ-8, 2,3-
dimethoxy-5-methyl-6-octaprenyl-1,4-benzoquinone) biosynthesis. UQ-8 and PQ-9 are
both members of the alkyl-substituted benzoquinone family and contain a 1,4-
benzoquinoid ring with a polyprenyl side chain (45 carbon units for PQ-9 and 40 carbon
units for UQ-8) at the C6 position of the benzoquinoid ring system. UQ-9 biosynthesis
occurs as follows (Fig 6.1, see a review by Meganathan, 2001, and references therein).
The first committed step of the pathway is the aromatization of chorismate to 4-
hydroxybenzoate (4-HBA), which is catalyzed by chorismate-pyruvate lyase (UbiC).
Prenylation by 4-HBA octaprenyltransferase (UbiA) and decarboxylation by 3-
octaprenyl-4-HBA decarboxylase (UbiD and UbiX) results in the synthesis of 2-
octaprenyl phenol, which is converted to UQ after three hydroxylation reactions that
alternate with three methylation reactions. A hydroxyl group is first introduced by 2-
octaprenyl-phenol monooxygenase (UbiB) (at the position ortho to the C1 OH), which is

247
subsequently methylated by an O-methyltransferase (UbiG) resulting in 2-methoxy-6-
octaprenyl phenol. Secondly a hydroxyl group is introduced by a monooxygenase (UbiH)
at the position para to the C1 hydroxyl group, resulting in the synthesis of 2-methoxy-6-
octaprenyl-1,4-benzoquinol. UbiE, a C-methyltransferase, delivers a methyl group to a
position ortho to the C6 prenyl side chain, resulting in the synthesis of 2-methoxy-5-
methyl-6-octaprenyl-1,4-benzoquinol. Further hydroxylation by a monooxygenase (UbiF)
and methylation by UbiG at the C3 position of the ring system results in UQ-8. In the
structure of PQ-9, the two methoxy groups are replaced by two methyl groups (see Fig
6.1), and no additional methyl group is present at the ring position ortho to the prenyl
side chain. Therefore, it is possible that the PQ-9 biosynthesis occurs by a pathway that is
similar to the UQ-8 biosynthesis pathway in proteobacteria, at least in that part of the
pathway that leads to the formation of the prenyl benzoquinone.
To investigate the PQ-9 biosynthesis pathway in cyanobacteria, comparative
genome analyses using 14 cyanobacterial genome sequences were performed in
combination with a reverse genetic approach. The results suggest that the PQ-9
biosynthesis occurs by a pathway similar to that for the UQ-8 pathway in proteobacteria.

248
RESULTS
Comparative Genome Analyses
As of April 26, 2004, largely completed genomic sequences of 14 cyanobacteria
are available: eight complete sequences exist for Synechocystis sp. PCC 6803 (Kaneko et
al., 1996), Nostoc sp. PCC 7120 (Kaneko et al., 2001), Thermosynechococcus elongatus
BP-1 (Nakamura et al., 2002a,b), Gloeobacter violaceus (Nakamura et al., 2003),
Prochlorococcus marinus MED4 and Prochlorococcus marinus MIT9313 (Rocap et al.,
2003), Prochlorococcus marinus SS120 (Dufresne et al., 2003), Synechococcus sp.
WH8102 (Palenik et al., 2003); and incomplete sequences exist for Synechococcus sp.
PCC 7002 (J. Marquardt, T. Li, J. Zhao, and D. A. Bryant, unpublished), Nostoc
punctiforme (Gene Bank accession number: NZ_AAAY00000000), Trichodesmium
erythraeum (Gene Bank accession number: NZ_AABK00000000), Synechococcus
elongatus PCC 7942 (Gene Bank accession number: NZ_AADZ01000001), Anabaena
variabilis ATCC 29413 (Gene Bank accession number: NZ_AAEA01000001), and
Crocosphaera watsonii WH8502 (Gene Bank accession number: NZ_AADV01000004).
Database searches using the E. coli UbiA, UbiB, UbiC, UbiD, UbiE, UbiF, UbiG, UbiH,
and UbiX protein sequences against these genomes revealed the presence of homologs
for UbiA, UbiD, UbiX, UbiE, and UbiH. Similarities between the sequences of E. coli
Ubi proteins and the cyanobacterial homologs were about 50% for UbiA (Table 6.1), 30-
50% for UbiH (Table 6.2), 60% for UbiD (Table 6.3), and 50% for UbiX (Table 6.4).
Multiple UbiE/UbiG homologues were detected, and the number varied depending on the
species (Table 6.5). The observed conservation of these Ubi proteins among

249
cyanobacteria and between cyanobacteria and E. coli suggests that the part of the UQ-8
biosynthesis pathway involving prenylation, decarboxylation and the introduction of a
hydroxyl group is probably functioning in cyanobacteria. The only exception to this is P.
marinus MED4, in which no UbiD and UbiX homologs were detected. Reverse-phase
HPLC analyses of the whole cell extracts confirmed that P. marinus MED4 synthesizes
about two molecules of PQ-9 per 100 chlorophyll a molecules. Similar value was
obtained for the whole cell extracts from Synechocystis sp. PCC 6803. Therefore, it is
safely concluded that P. marinus MED4 synthesizes PQ-9 and it is suggested that the
carboxylation reaction is probably catalyzed by yet unknown enzymes.
UQ-8 biosynthesis in E. coli involves eight enzymes and two of these were absent
in cyanobacteria: UbiB, and UbiF. The absence of UbiB and UbiF, the monooxygenases
that introduce hydroxyl groups into ring system (which are subsequently methylated by
UbiG), is consistent with the absence of the methoxy substituents in PQ-9. In
combination with the results described above, it was hypothesized that biosynthesis of
PQ-9 in cyanobacteria occurs as shown in Fig. 6.2. In this pathway, 4-hydroxybenzoate is
converted by UbiA to 2-solanyl-4-hydroxybenzoate, which is decarboxylated by
UbiD/UbiX giving rise to 2-solanyl phenol. It is proposed that 2-solanyl phenol is first
hydroxylated by UbiH, followed by two methylation reactions performed by one or more
of the UbiE homologs, although it is entirely possible that one or both of the methylation
reactions occurs before hydroxylation, as is the case for O-methylation by UbiG, which
precedes hydroxylation by UbiH during UQ-8 synthesis (see Meganathan, 2001, and Fig
6.1).

250
UbiC, chorismate lyase, is responsible for the first committed step in the pathway
(Fig 6.1). The ubiC gene in some β-proteobacteria and γ-proteobacteria, including
Neisseria meningitis Z2491, Pseudomonas aeruginosa PA01, Salmonella enterica subsp.
enterica serovar Typhi, Shigella flexneri str. 301 setotype 2a, Yersinia pestis C092 and E.
coli, is located immediately upstream of the ubiA gene based on genome analyses
performed on these organisms. In cyanobacteria, no ubiC homolog was found in the
vicinity of the ubiA gene, and no obvious ubiC homologs were found in their genomes.
These observations appeared to indicate that homologs of this gene were absent in the
cyanobacteria, and that synthesis of 4-hydroxybenzoate occurs through an alternative
pathway (see discussion). After exhaustive database searching, however, ORFs that
showed very weak similarity to the UbiC sequences of E. coli (< 30 %) were detected
(Table 6.6). These ORFs were highly conserved among cyanobacteria (50 –80%
similarity) and showed amino acid sequence similarity to Ycf21 proteins, which are also
found in the plastid genomes of eukaryotic phototrophs including Porphyra purpurea and
Cyanophora paradoxa. Fig 6.3 shows an alignment of sequences of the UbiC protein in
E. coli and the Yfc21 homologs in the cyanobacteria and eukaryotic phototrophs. In the
alignment, a glutamate residue at the position 155 (UbiC) was found to be conserved
among the Ycf21 homologs in both the cyanobacteria and eukaryotic phototrophs. This
residue constitutes a portion of the catalytic site and is hydrogen-bonded to the hydroxyl
oxygen of 4-hydroxybenzoate as previously shown in the 1.4-Å resolution X-ray crystal
structure of the UbiC protein (Gallagher et al., 2001). An arginine residue at the position
79, which has also been shown to constitute a portion of the active site, was conserved
among many cyanobacteria, although not all. Therefore, it is possible that the Ycf21

251
homologs catalyze the conversion of chorismate to 4-hydroxybenzoate in the PQ-9
biosynthesis in cyanobacteria.
Mutagenesis of ubiA, ubiX, and ubiH
An attempt was made to inactivate the ORFs slr0926 (ubiA), slr1099 (ubiX), and
slr1300 (ubiH) in Synechocystis sp. PCC 6803 by insertion of the aphII gene conferring
kanamycin resistance. These attempts, however, failed to lead to homozygous mutants
and resulted in incomplete segregation of the alleles under photoautotrophic conditions
even under selective conditions in the presence of 50 µg mL-1 kanamycin (data not
shown). Addition of glucose to growth medium, which provides electrons to the electron
transport chains through NADH (thereby bypassing Photosystem II), likewise did not
lead to complete segregation. Addition of various quinoid compounds to the growth
medium either to inhibit the Photosystem II activity (atrazine) or to compensate the loss
of PQ-9 (2,6-dimethyl-1,4-benzoquinone, PhyQ, UQ-0, UQ-1, and UQ-2) did not lead to
complete segregation (data not shown). Merodiploid mutants were unstable both under
photoautotrophic and photomixotrophic conditions under illumination at a normal light
intensity (20 - 50 µE m-2 s-1), as no further growth was observed after several transfers on
solid media. These results indicates that the ubiA, ubiX, and ubiH genes are indispensable
for the survival of Synechocystis sp. PCC 6803, which is consistent with the anticipated
essential role of plastoquinone in cyanobacteria.

252
Phylogenetic Reconstruction with Ubi proteins of Cyanobacteria and Proteobacteria
Evolutionary relationships between the Ubi homologues of cyanobacteria and
proteobacteria (E. coli, Rhodopseudomonas palustris, Rhodospirillum rubrum and
Rhodobacter sphaeroides) were analyzed by constructing phylogenetic trees based on
deduced amino acid sequences and by comparing the results to the phylogenetic tree
based on 16S ribosomal RNA (rRNA) (Fig 6.4). 16S rRNA is universally present and
highly conserved among prokaryotes and thus is often used as a molecular marker to
reflect the evolutionary clock (Woese, 1987). Phylogenetic trees based on the nucleotide
sequence of 16S rRNA showed that the cyanobacteria and proteobacteria formed
independent groups, which is consistent with previous results (Woese, 1987). Within the
cyanobacteria, two major clades were observed: clade A consisted of Synechocystis sp.
PCC 6803, Synechococcus sp. PCC 7002, Nostoc sp. PCC 7120, N. punctiforme, and T.
erythraeum, and clade B consisted of Synechococcus sp. WH8102 and the three
Prochlorococcus species (Fig 6.4). Phylogenetic trees based on the amino acid sequences
of UbiA, UbiD, and UbiX showed similar branching orders and topologies to the tree for
16S rRNA. The trees were supported by high bootstrap values (Fig 6.4), suggesting that
the ubiA, ubiD, and ubiX genes have evolved through vertical heritage from the last
common ancestor of both cyanobacteria and the proteobacteria. The topology of the tree
based on the UbiH sequences appeared different, however. Within the group formed by
all cyanobacteria, the proteobacteria formed a cluster with clade A, which may indicate
the lateral transfer of the ubiH gene between these two groups of organisms. However,
the bootstrap value assigned to the branch connecting a node where cyanobacteria in
clade A and clade B bifurcate and a node where the proteobacteria deviate from the

253
cyanobacteria in clade A is only 52, which only poorly supports the likeliness of this
branching order. Alternatively, it is possible that the out-group used to construct the tree
(Mlr2092, a putative UbiH homologue in Mesorhizobium loti) may be distorting the
topology and thereby gives rise to an artifact. This happens if the sequence of the out-
group is more closely related to the cyanobacterial sequences than to the proteobacterial
sequences. As shown in Table 6.2, the amino acid sequence of Mlr2092 that was used as
an out-group is equally distantly related to any of the UbiH sequences analyzed. The
replacement of Mlr2092 by another protein sequence, NahG (salicylate hydroxylase in
Pseudomonas putida ) or Bsu1050 (a putative monooxygenase in Bacillus subtilis), did
not alter the topology of the tree. Therefore, it is possible that the proteobacteria form a
group within the cyanobacteria and a lateral exchange of ubiH took place between
proteobacteria and the cyanobacteria in clade A. This evolutionarily close relationship of
UbiH homologs between the two groups of organisms provides further support for the
involvement of these proteins in the PQ-9 biosynthesis in cyanobacteria.
UbiE
Database search of the genomes of cyanobacteria showed the presence of multiple
UbiE homologs (Table 6.5, Fig 6.5). There are two ORFs that are conserved in all 14
cyanobacteria: these are homologs of Sll1653 and Sll0418 of Synechocystis sp. PCC
6803. Note that the incomplete genome sequence of N. punctiforme only contained a
homolog of Slr0089, which encodes γ-tocopherol methyltransferase. Whether a homolog
of Sll0418 is present in this organism is yet to be determined. Homologs of Sll0829 and
Sll0487 are conserved in all cyanobacteria except P. marinus MED4 and P. marinus

254
SS120. Slr1618 and Slr0407 seem to be specific to Synechocystis sp. PCC 6803 and C.
watsonii, or Synechocystis sp. PCC 6803, A. variabilis, and G. violaceus, respectively
(Table 6.5). Amino acid sequences of all these homologs contain the conserved S-
adenosyl-L-methionine binding motif (data not shown). UbiE in E. coli is a bifunctional
enzyme and is responsible for the C-methylation reactions in both UQ-8 and
menaquinone biosyntheses (Lee et al., 1997). Phylogenetic analyses conducted on the
UbiE homologues of cyanobacteria showed that Sll1653 forms a cluster with the E. coli
UbiE (Fig 6.5). Furthermore, as described in Chapter 2 and Chapter 3 (also see Sakuragi
et al., 2002), this ORF is responsible for transferring a methyl group to 2-prenyl-1,4-
naphthoquinone in phylloquinone and menaquinone-4 biosyntheses in cyanobacteria.
Therefore, it was expected that Sll1653 would be responsible for at least one of the
methylation reactions in PQ-9 synthesis. Whole-cell extracts from the sll1653 mutant
were analyzed by reverse-phase HPLC. PQ-9 eluted at 36 min using the current analytical
system (see “Materials and Methods”, see also Chapter 2). Whereas the chemically
synthesized 2-methyl-6-solanyl-1,4-benzoquinone (a kind gift from Prof. DellaPenna in
the Michigan State University, see Shintani et a., 2000), a predicted intermediate in the
PQ-9 synthesis, eluted at 33 min as shown in Fig 6.6A. The HPLC profile of the whole-
cell solvent extracts from the sll1653 mutant showed a peak at 35.5 min. The absorption
spectrum of the component eluting at this retention time showed a maximum at 256 nm
(Fig 6.6B, solid line), which is characteristic of PQ-9 (Dunphy and Brodie, 1971) and is a
few nanometers red shifted as compared to the absorption maximum observed for the
MPBQ standard (Fig 6.6B, dotted line). The results therefore demonstrate that PQ-9 is
present in the sll1653 mutant cells. The contents of PQ-9 were analyzed based on the

255
integrated peak area in the HPLC chromatograms and were expressed as relative to the
cellular contents of chlorophyll a (Fig 6.7). The results showed that the PQ-9 content in
the sll1653 mutant is comparable to that in wild type. Therefore, it was concluded that
Sll1653 is not involved in the PQ-9 biosynthesis or that a redundant activity conferred by
another methyltransferase(s) is present.
An in vitro study has previously demonstrated that recombinant Sll0418, MPBQ
methyltransferase, can catalyze methylation of MSBQ and lead to the synthesis of PQ-9
(Shintani et al., 2002). Because this ORF is also highly conserved among the
cyanobacteria (Table 6.5, Fig 6.5), it was predicted that Sll0418 is either responsible for
transferring of one or two of the methyl groups in the PQ-9 biosynthesis or that it
provides a redundant activity with Sll1653. Targeted inactivation and complete
segregation of the mutated allele was performed and confirmed by PCR (see Chapter 3).
The sll0418 mutant was, however, capable of synthesizing the wild-type level of PQ-9 as
shown by HPLC analysis on the whole-cell lipid extracts (Fig 6.7).
Double and triple mutants of the UbiE homologs were constructed in
Synechocystis sp. PCC 6803 in further attempt to identify methyltransferase(s) involved
in the PQ-9 biosynthesis. Combinations of mutated loci are as follows: double mutations
were constructed for sll1653 and sll0418, sll1653 and slr0407, sll1653 and sll0829,
sll0418 and sll0829, sll0418 and slr0407, and sll0418 and slr0089; and triple mutations
were constructed for of sll1653, sll0418 and sll0829; and sll1653, slr0407, and sll0829.
Antibiotic resistance cartridges were used to insertionally inactivate these genes (Fig 6.8).
The full segregation of the mutated alleles from the respective wild-type alleles was

256
analyzed by PCR as shown in Fig 6.9. PCR using the designed primers (Table 6.7) that
are targeted to the sll0829, sll01653, sll0418, slr0407, slr0089, slr1618 resulted in
products of 0.84 kbp, 0.3 kbp, 0.92 kbp, 0.94 kbp, 1.2 kbp, and 1.8 kbp, respectively (Fig
6.9, lane 1), which are expected based on the restriction maps as shown in Fig 6.8. No
product with corresponding wild-type size was detected for the mutants (Fig 6.9, lanes 2);
instead products with larger sizes were detected in all cases. The difference between the
mutants and the wild-type corresponds either to the size of antibiotic resistance cartridges
or to the size of antibiotic resistance cartridges minus the deleted regions (Fig 6.8).
Therefore, it was concluded that all the mutants are free of respective wild-type alleles
and were homozygous in the allele for which the drug cartridges had been introduced.
Reverse-phase HPLC analyses on whole-cell lipid extracts showed that none of
the double and triple mutants showed a significant difference from the wild type in the
amount of PQ-9 accumulated in the cells (Fig 6.7). The fact that the sll1653 sll0418
double mutant did not affect the PQ-9 content strongly indicates that Sll1653 and
Sll0418, despite their conservation among all the cyanobacteria and their predicted
functioning in PhyQ/MQ synthesis (or the demonstration of function in vitro PQ-9
synthesis), do not play major roles in the PQ-9 biosynthesis. This is further supported by
the construction of the sll1635 sll0418 double mutant, which also synthesized a wild-type
level of PQ-9 (Fig 6.7). The possibility that another UbiE homolog is providing an
additional activity to Sll01653 and/or Sll0418 was tested by analyzing the PQ-9 contents
in the sll0418 slr1618, sll0418 slr0407, sll0418 sll0829, sll1653 sll0829, and sll1653
slr0407 double mutants by reverse-phase HPLC. The results showed that all of the double

257
mutants contained essentially the wild-type level of PQ-9, which indicates that Sll0829,
Slr0407, and Slr1618 are not involved in the PQ-9 biosynthesis. This is further supported
by the result that the two triple mutants, sll1653 sll0418 sll0829 and sll1653 slr0407
sll0829, contained a wild-type level of the PQ-9 in the whole-cell solvent extracts.
Participation of Slr0089 in the PQ synthesis was also ruled out as the sll0418 slr0089
double mutant synthesized the wild-type level of PQ-9. Slr1039 and Slr0487 are two
hypothetical proteins with an S-adenosyl-L-methionine binding motif, and they are well
conserved among most of the cyanobacteria (Fig 6.5, Table 6.5). Targeted inactivation of
these ORFs, however, did not affect the PQ-9 biosynthesis. In combination, these results
strongly suggest that the UbiE homologs do not play a major role in the PQ-9
biosynthesis in Synechocystis sp. PCC 6803.

258
DISCUSSION
Database searches of the whole genome sequences of 14 cyanobacteria (8
complete and 6 incomplete) revealed the presence of the UbiA, UbiC, UbiD, UbiE, UbiH,
and UbiX homologs, which are proteins involved in the UQ-8 biosynthetic pathway in
proteobacteria. Attempted insertional inactivation of the ubiA, ubiH, and ubiX genes in
Synechocystis sp. PCC 6803 resulted in the generation of unstable merodiploids, which
indicates that these genes are essential for the survival of this organism and which is
consistent with the presumed essential role of PQ-9 in cyanobacteria. Based on these
findings, it is proposed that the PQ biosynthesis occurs in cyanobacteria as shown in Fig
6.2. In this proposal, UbiA catalyzes the condensation of 4-hydroxybenzoate and
solanyldiphosphate resulting in 3-solanyl-4-hydroxybenzoate, which is decarboxylated by
UbiD/UbiX resulting in 2-solanyl phenol. Hydroxylation by UbiH is expected to take
place at the C4 position of the ring system, resulting in 2-solanyl-1,4-benzoquinol.
Whether the two methylation reactions at the C2 and C3 position take place before or after
the hydroxylation is an open question. These methylation reactions, however, are not
catalyzed by the UbiE homologs (Sll1653, Sll0418, Sll0829, Slr0407, Sll0487, Slr1618
and Slr1039) in Synechocystis sp. PCC 6803 as demonstrated by insertional mutagenesis
and subsequent HPLC analyses. Phylogenetic analyses have indicated that the presence
of the UbiA, UbiD, and UbiX homologs in cyanobacteria are the result of vertical
heritage from the common ancestor of cyanobacteria and proteobacteria, suggesting a
possible common evolutionary origin of the UQ-8 and PQ-9 biosyntheses. UbiE
(Sll1653) is probably inherited from the common ancestor and evolved in cyanobacteria

259
to be specific to the methylation reaction in the PhyQ/MQ biosynthesis, while another
methyltransferase(s) has been recruited to function for the PQ-9 biosynthesis.
The methyltransferase(s) that function in the PQ-9 biosynthesis is/are still a
mystery. The fact that mutants of 7 UbiE homologs contained wild-type levels of PQ-9
suggests i) that a SAM (S-adenosyl-L-methonine)-dependent methyltransferase(s) that is
only very distantly related to UbiE is responsible for these reactions, or ii) that a SAM-
independent methyltransfrase(s) is responsible for the reactions. Of the few SAM-
independent methyltransferases detected in the Synechocystis sp. PCC 6803 genome,
none is conserved among cyanobacteria except for slr0212, which is annotated as
encoding the methionine synthase and uses 5-methyltetrahydrofolate as the methyl donor
([EC 2.1.1.13], see http://www.chem.qmul.ac.uk/iubmb/enzyme/EC2/1/1/13.html). This
ORF is, however, highly conserved among other bacteria and eukaryotes and is present in
the genome as a single copy. Therefore, this gene product most likely functions as
methionine synthase and is unlikely to be involved in the PQ biosynthesis. When the
genome was searched for SAM-binding proteins, ORFs sll1407, slr1115, slr0303,
slr1071, sll1693, slr1815 were detected. Of these, slr0303 is the only ORF that is
conserved among all the cyanobacteria. The possible involvement of this hypothetical
protein in PQ-9 biosynthesis needs to be tested.
The presence of conserved Ycf21 homologs in cyanobacteria, which are weakly
similar to UbiC in E. coli, suggested that 4-hydroxybenzoate derives from chorismate by
the action of chorismate-lyase activity possibly conferred by these Ycf21 homologs. This
was thought to be a highly plausible hypothesis because catalytic residues of the UbiC

260
protein are largely conserved among the Ycf21 homologs in cyanobacteria. However, the
poor amino acid sequence similarity between the Ycf21 homologs and UbiC proteins
leaves a possibility that an alternative pathway exists that leads to the synthesis of 4-
hydroxybenzoate from a compound other than chorismate. Genome analyses have
revealed that many organisms that synthesize UQ lack UbiC, including higher plants,
animals, and even some proteobacteria such as Agrobacterium tumefaciens, Brucella
melitensis, Rhodopseudomonas palustris, Chromobacterium violaceum, and
Xanthomonas axonopodis. There are three pathways that are known to lead to the
synthesis of the intermediate 4-hydroxybenzoate in these organisms: i) the UbiC-
dependent chorismate pathway in E. coli and some β- and γ-proteobacteria; ii) the 4-
hydroxyphenyllactate pathway in rat (see review by Meganathan, 2001, and references
therein); and iii) the aromatic amino acid pathway in higher plants (see a review by
Threlfall and Whistance, 1971, and references therein). In UQ biosynthesis, 4-
hydroxybenzoate is synthesized from the aromatic amino acid tyrosine or phenylalanine
via p-coumarate (Loscher and Heide, 1994). It has also been reported that the
cyanobacterium Anacystis nidulans converts the aromatic amino acid tyrosine to 4-
hydroxybenzoate, and that this conversion may be carried out by thylakoid-bound
enzyme complexes consisting of tyrosine ammonia-lyase and 4-hydroxybenzoate
synthase (Löffelhardt, 1976). Therefore, it is plausible that a similar aromatic amino acid
pathway may exist in other cyanobacteria and could provide 4-hydroxybenzoate for PQ-9
biosynthesis as proposed in Fig 6.10.

261
Lastly, the fact that the sll0418 mutation did not affect the cellular PQ content in
Synechocystis sp. PCC 6803 indicates that Sll0418 is only involved in α-tocopherol
biosynthesis. However, this ORF is conserved in nearly all cyanobacteria whose genome
sequence is so far available, including those that are predicted not to be capable of α-
tocopherol synthesis (Synechococcus sp. WH 8102 and three Prochlorococcus species,
see Chapter 5). What is the function of the Sll0418 homologs in these cyanobacteria? Is it
possible that the Sll0418 homologs are involved in PQ biosynthesis in these
cyanobacteria, given that the recombinant Sll0418 can catalyze the methylation of MSBQ
and generate PQ-9. Genetic analyses of the role of Sll0418 needs to be performed in these
other organisms.

262
SUMMARY
The PQ-9 biosynthesis pathway was studied by means of comparative genome
analyses in 14 cyanobacteria. The presence of UbiA, UbiD, UbiE, UbiH, and UbiX
homologs, which are required for UQ-8 biosynthesis in E. coli, was detected in the
cyanobacteria. The presence of Ycf21 homologs, which show weak similarity to UbiC,
was detected after exhaustive database searching in cyanobacteria. The results suggest
that PQ-9 synthesis occurs similarly to UQ-8 biosynthesis pathway. Targeted inactivation
of six UbiE homologs (methyltransferases), however, did not affect PQ-9 biosynthesis in
Synechocystis sp. PCC 6803. This suggested that enzymes that are not evolutionarily
related to UbiE in E. coli catalyze the methylation reactions in PQ-9 biosynthesis in
cyanobacteria.

263
MATERIALS AND METHODS
Growth Conditions
The wild-type strain used for transformation and further analysis was a glucose-
tolerant stain of Synechocystis sp. PCC 6803 (Williams, 1988). Medium BHEPES, pH
8.0, was used for selection, maintenance, and growth measurements of wild type and
mutants. This medium is prepared by supplementing BG-11 medium (Stainer et al., 1971)
with 4.6 mM of HEPES [4-(2-hydroxyethyl)piperazine-N’-(2-ethanesulfonic acid)]-KOH
and 18 mg L-1 ferric ammonium citrate. The wild-type and ubiA, ubiH, and ubiX mutant
strains were grown on solid medium BHEPES containing the following additions: 5 mM
glucose alone; 5 mM glucose and various concentrations (5 µM, 50 µM, 500 µM, 1 mM,
and 10 mM) of ubiquinone-0 (2,3-dimethoxy-5-methyl-1,4-benzoquinone), ubiquinone-1
(2,3-dimethoxy-5-methyl-6-(3-methyl-2-butenyl)-1,4-benzoquinone), ubiquinone-2 (2,3-
dimethoxy-5-methyl-6-geranyl-1,4-benzoquinone), or phylloquinone; 5 mM glucose and
5 µM atrazine (2-chloro-4-ethylamino-6-isopropylamino-1,3,5-triazine); or 5 mM
glucose, 5 µM ubiquinone-2, and 20 µM 2,6-di-O-methyl-β-cyclodextrin. The cells on
these solid media were grown at room temperature at moderate light intensity (ca. 50 µE
m-2 s-1) or at low light intensity (ca. 5 µE m-2 s-1).
Constructions of Single and Double Methyltransferase Mutants
DNA fragments containing the sll0829, sll0418, slr0407, slr1618, slr1039, and
slr0089 genes were amplified from isolated genomic DNA by PCR using a set of forward
and reverse primers (see Table 6.7 for sequences and Fig 6.8 for positions). The

264
constructions of the insertional inactivation of these alleles are described in Table 6.8.
The aacC1 gene, conferring gentamicin resistance, derived from the pMS266 plasmid
(Becker et al., 1995) and the aadA gene, conferring spectinomycin resistance, derived
from the pSRA2 plasmid (Frigaard et al., 2004) were inserted into the selected site within
these gene coding regions (see Table 6.8 and Fig 6.8). The resulting p829Gm and
p1039Sp plasmids were linearized and used to transform the wild-type Synechocystis sp.
PCC 6803 to obtain the homozygous sll0829::accC1 and slr1039::aadA mutants.
Genomic DNA isolated from the sll0418::aphII mutant (Chapter 4) and the
sll1653::aphII mutant (Chapter 3) were used to transform the homozygous
sll0829::accC1 mutant to generate the sll0418::aphII sll0829::accC1 and sll1653::aphII
sll0829::accC1 double mutants. The linearized p418Sp and p407Sp were used to
transform the homozygous sll1653::aphII sll0829::accC1 double mutant to generate the
sll1653::aphII sll0418::aadA sll0829::accC1 and sll1653::aphII slr0407::aadA
sll0829::accC1 triple mutants. The linearized p418Sp was used to transform the
homozygous sll1653::aphII mutant to generate the sll0418::aadA sll1653::aphII double
mutant. The linearized p1618Sp, p407Sp, and p89Gm were used to transform the
homozygous sll0418::aphII mutant to generate the sll0418::aphII 1618::aadA,
sll0418::aphII slr0407::aadA, and sll0418::aphII slr0089::accC1 double mutants. A
homozygous sll0487::aphII mutant was constructed in a previous study (W. M.
Schluchter, G. Shen, and D. A. Bryant, unpublished results). Complete segregation of the
each mutated alleles from the corresponding wild-type allele was confirmed by PCR.
Primer used for the PCR analyses are summarized in Table 6.7.

265
Quinone and Chlorophyll Analysis
Cells were harvested from photoautotrophically grown cultures and recovered in a
tight pellet. Pigments and quinones were extracted with 400 µL of cold acetone:methanol
(7:2 v/v) after brief ultrasonication. After centrifugation and filtration through a PTFE
filter membrane with a 0.2-µm pore size (Whatman International Ltd., Maldstone, UK),
the extract in the organic phase were directly injected into an Agilent Technology 1100
series HPLC system (Agilent Technology, Palo Alto, CA, USA) equipped with
SUPELCO (Sigma-Aldrich Corp., St. Louis, MO, USA) Discovery® C18 column (25 cm
x 4.6 mm, 5 µm). Analyses were carried out using the following protocol: 80%A/20%B
from 0 to10 min, a linear change to 20%A/80%B in 40 min, and 20%A/80%B for 5 min,
where solvent A and solvent B are 100% methanol and 100% isopropanol, respectively.
The flow rate was 0.75 ml min-1. Detection of eluates was performed with a diode array
detector (Agilent 1100 series). The chlorophyll a, phylloquinone, and plastoquinone
contents of each sample were determined based on integrated peak areas and their molar
absorption coefficients, which are 17.4 mM-1 cm-1 at 618 nm, 18.9 mM-1 cm-1 at 270 nm
(Dunphy and Brodie, 1971), and 15.2 mM-1 cm-1 at 254 nm (Crane and Dilley, 1963),
respectively. The absorption coefficient of chlorophyll a at 618 nm was calculated from
the ratio of the absorption peaks at 618 nm and 666 nm in methanol-isopropanol (6:4,
v/v) and from its absorption coefficient at 666 nm in methanol (MacKinney, 1941).

266
REFERENCES
Barry BA, Boerner RJ, de Paula JC (1994) The use of cyanobacteria in the study of the
structure and function of Photosystem II. In: Bryant DA (ed) Molecular Biology of
Cyanobacteria. pp 217-257. Kluwer Academic Publishers, Dordrecht
Becker A, Schmidt M, Jäger W, Pühler A (1995) New gentamicin-resistance and lacZ
promoter-probe cassettes suitable for insertion mutagenesis and generation of
transcriptional fusions. Gene 162: 37-39
Bonner CA, Jensen RA (1995) Novel features of prephenate aminotransferase from cell
cultures of Nicotiana silvestris. Arch Biochem Biophys 238 237-246
Bricker TM, Ghanotakis DF (1996) Introduction to oxygen evolution and the oxygen-
evolving complex. In: Ort DR, Yocum CF (eds) Oxygenic Photosynthesis: The Light
Reactions. pp 113-136. Kluwer Academic Publishers, Dordrecht
Britt RD (1996) Oxygen evolution. In: Ort DR, Yocum CF (eds) Oxygenic
Photosynthesis: The Light Reactions. pp 137-164. Kluwer Academic Publishers,
Dordrecht
Cheng Z, Sattler S, Maeda H, Sakuragi Y, Bryant DA, DellaPenna D (2003) Highly
divergent methyltransferase catalyzes a conserved reaction in tocopherol and
plastoquinone synthesis in cyanobacteria and photosynthetic prokaryotes. Plant Cell 15:
2343-2356

267
Collakova E, DellaPenna D (2001) Isolation and functional analysis of homogentisate
phytyltransferase from Synechocystis sp. PCC 6803 and Arabidopsis. Plant Physiol 127:
1-12
Collins MD and Jones D (1981) Distribution of isoprenoid quinone structural types in
bacteria and their taxonomic implications. Microbiological Reviews 45: 316-354
Crane FL, Dilley RA (1963) Determination of coenzyme Q (ubiquinone). Methods
Biochem Anal 11: 279-306
Diner BA, Babcock GT (1996) Structure, dynamics, and energy conversion efficiency in
Photosystem II. In: Ort DR, Yocum CF (eds) Oxygenic Photosynthesis: The Light
Reactions. pp 213-247. Kluwer Academic Publishers, Dordrecht
Dufresne A, Salanoubat M, Partensky F, Artiguenave F, Axmann IM, Barbe V, Duprat S,
Galperin MY, Koonin EV, Le Gall F, Makarova KS, Ostrowski M, Oztas S, Robert C,
Rogozin IB, Scanlan DJ, Tandeau de Marsac N, Weissenbach J, Wincker P, Wolf YI,
Hess WR (2003) Genome sequence of the cyanobacterium Prochlorococcus marinus
SS120, a nearly minimal oxyphototrophic genome. Proc Natl Acad Sci USA 100: 9647-
9649
Dähnhardt D, Falk J, Appel J, van der Kooij TAW, Schulz-Friedrich R, Krupinska K
(2002) The hydroxyphenylpyruvate dioxygenase from Synechocystis sp. PCC 6803 is not
required for plastoquinone biosynthesis. FEBS Lett 523: 177-181

268
Dunphy PJ, Brodie AF (1971) The structure and function of quinones in respiratory
metabolism. Methods Enzymol 18: 407-461
Frigaard N-U, Li H, Milks KJ, Bryant DA (2004) Nine mutants of Chlorobium tepidum
each unable to synthesize a different chlorosome protein still assemble functional
chlorosomes. J Bacteriol 186: 646-653
Gallagher DT, Mayhew M, Holde MJ, Howard A, Kim K-J, Vilker VL (2001) The
crystal structure of chorismate lyase shows a new fold and a tightly retained product.
Proteins 44: 304-311
Kaneko T, Nakamura Y, Wolk CP, Kuritz T, Sasamoto T, Watanabe A, Iriguchi M,
Ishikawa M, Kawashima M, Kimura T, Kishida Y, Kohara M, Matsumoto M, Matsuno
A, Muraki A, Nakazaki N, Shimpo S, Sugimoto M, Takazawa M, Yamada M, Yasuda M,
Tabata S (2001) Complete genomic sequence of the filamentous nitrogen-fixing
cyanobacterium Anabaena sp. strain PCC 7120. DNA Res 8: 205-213
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y, Miyajima N, Hirosawa
M, Sugiura M, Sasamoto S, Kimura T, Hosouchi T, Matsuno A, Muraki A, Nakazaki N,
Naruo K, Okumura S, Shimpo S, Takeuchi C, Wada T, Watanabe A, Yamada M, Yasuda
M, Tabata S (1996) Sequence analysis of the genome of the unicellular cyanobacterium
Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and
assignment of potential protein-coding regions. DNA Res 3: 109-136

269
Lee PT, Hsu AY, Ha HT, Clarke CF (1997) A C-methyltransferase involved in both
ubiquinone and menaquinone biosynthesis: Isolation and identification of Escherichia
coli ubiE gene. J Bacteriol 179: 174-1754
Löffelhardt W (1976) Formation of benzoic acid and p-hydroxybenzoic acid in the blue
green algae Anacystis nidulans: a thylakoid-bound enzyme complex analogous to the
chloroplast system. Z Naturforsch C 31: 693-699
Loscher R and Heide L (1994) Biosynthesis of p-hydroxybenzoate from p-coumarate and
p-coumaroyl-coenzyme A in cell-free extracts of Lithospermum erythrorhizon cell
cultures. Plant Physiol 106: 271-279
MacKinney G (1941) Absorption of light by chlorophyll solutions. J Biol Chem 140:
315-322
Meganathan R (2001) Biosynthesis of menaquinone (vitamin K2) and ubiquinone
(coenzyme Q): a perspective on enzymatic mechanisms. Vitam Horm 61: 173-218
Nakamura Y, Kaneko T, Sato S, Ikeuchi M, Katoh H, Sasamoto S, Watanabe A, Iriguchi
M, Kawashima K, Kimura T, Kishida Y, Kiyokawa C, Kohara M, Matsumoto M,
Matsuno A, Nakazaki N, Shinpo S, Sugimoto M, Takeuchi C, Yamada M, Tabata S
(2002) Complete genome structure of the thermophilic cyanobacterium
Thermosynechococcus elongatus BP-1. DNA Res 9: 2- 130
Nakamura Y, Kaneko T, Sato S, Mimuro M, Miyashita H, Tsuchiya T, Sasamoto, S,
Watanabe A, Kawashima K, Kishida Y, Kiyokawa C, Kohara M, Matsumoto M,

270
Matsuno A, Nakazaki N, Shimpo S, Takeuchi C, Yamada M, Tabata S (2003) Complete
genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks
thylakoids. DNA Res 10: 137-145
Palenik B, Brahamsha B, Larimer FW, Land M, Hauser L, Chain P, Lamerdin J, Regala
W, Allen EE, McCarren J, Paulsen I, Dufresne A, Partensky F, Webb EA, Waterbury J.
(2003) The genome of a motile marine Synechococcus. Nature 424: 1037-1042
Pennock JF and Threlfall DR (1981) Biosynthesis of ubiquinone and related compounds.
In: Porter JW and Spurgeon SL (eds) Biosynthesis of Isoprenoid Compounds. Vol 2, pp
19-303. John Wiley & Sons, New York
Rocap G, Larimer FW, Lamerdin J, Malfatti S, Chain P, Ahlgren NA, Arellano A,
Coleman M, Hauser L, Hess WR, Johnson ZI, Land M, Lindell D, Post AF, Regala W,
Shah M, Shaw SL, Steglich C, Sullivan MB, Ting CS, Tolonen A, Webb EA, Zinser ER,
Chisholm SW (2003) Genome divergence in two Prochlorococcus ecotypes reflects
oceanic niche differentiation. Nature 424: 1042-1047
Shintani D, Cheng Z, DellaPenna D (2002) The role of 2-methyl-6-phytyl-benzoquinone
methyltransferase in determining tocopherol composition in Synechocystis sp. PCC 6803.
FEBS Lett 511: 1-5
Stainer RY, Kunisawa R, Mandel M, Cohen-Bazire G (1971) Purification and properties
of unicellular blue-green algae (Order Chroococcales). Bact Rev 35: 171-205

271
Threlfall DR and Whistance GR (1971) Biosynthesis of isoprenoid quinones and
chromanols. In: Goodwin TW (ed) Aspects of Terpenoid Chemistry and Biochemistry.
Vol. 12, pp 357-404. Academic Press, New York
Williams JGK (1988) Construction of specific mutations in Photosystem-II
photosynthetic reaction center by genetic engineering methods in Synechocystis 6803.
Methods Enzymol 167: 766-778
Woese CR (1987) Bacterial evolution. Microbiol.Reviews 51: 221-271

272 ab
le 6
.1: A
min
o ac
id se
quen
ce si
mila
ritie
s bet
wee
n U
biA
hom
olog
s in
14 c
yano
bact
eria
and
E. c
oli (
expr
esse
d as
%)
Org
anis
m
Locu
s ta
g 68
03
7002
N
ost
Npu
n Te
ry
Telo
n 81
02
Selo
Av
ar
Cw
at
MED
M
IT
S120
G
loe
Eco
Syne
choc
ystis
680
3 Sl
r092
6 10
0
Sy
nech
ococ
cus
7002
* O
RF
76
100
N
osto
c 71
20
Alr3
553
79
77
100
N. p
unct
iform
e *
Npu
n433
9 80
76
89
10
0
T. e
ryth
raeu
m*
Tery
4006
76
74
77
76
10
0
T.
elo
ngat
us B
P-1
Tlr0
402
67
64
66
66
65
100
Sy
nech
ococ
cus
8102
Sy
nw18
47
54
53
55
55
56
53
100
S. e
long
atus
794
2 *
Selo
0212
51
71
71
73
73
69
68
56
100
A.
var
iabi
lis 2
9413
* Av
ar02
0964
80
78
80
80
77
69
57
74
10
0
C
. wat
soni
i 850
1 *
Cw
at09
8001
83
76
98
89
79
68
54
72
80
10
0
P. m
arin
us M
ED4
Pmm
0456
53
52
54
51
55
55
64
57
54
55
10
0
P.
mar
inus
MIT
Pm
t931
3 54
54
54
55
56
57
75
57
57
57
67
10
0
P. m
arin
us S
S120
Pr
o045
5 56
54
56
56
57
53
70
55
58
59
68
75
10
0
G
. vio
lace
us 7
421
Glr3
428
71
70
70
70
69
67
52
71
73
72
53
53
56
100
E.
col
i K-1
2 U
biA
51
52
55
53
53
51
48
51
56
52
48
46
50
51
100
68
03, S
ynec
hocy
stis
sp. P
CC
680
3; 7
002,
Syn
echo
cocc
us sp
. PC
C 7
002;
Nos
t, N
osto
c sp
. PC
C 7
120;
Npu
n, N
osto
c pu
nctif
orm
e; T
ery,
Tri
chod
esm
ium
er
ythr
aeum
; Tel
on, T
herm
osyn
echo
cocc
us e
long
atus
BP-
1; 8
102,
Syn
echo
cocc
us sp
. WH
8102
; Sel
o, S
ynec
hoco
ccus
elo
ngat
us P
CC
794
2; A
var,
Anab
aena
va
riab
ilis A
TCC
294
13; C
wat
, Cro
cosp
haer
a w
atso
nii W
H85
01; P
roch
loro
cocc
us m
arin
us M
ED4;
MIT
, Pro
chlo
roco
ccus
mar
inus
MIT
9313
; S12
0,
Proc
hlor
ococ
cus m
arin
us S
S120
; Glo
e, G
loeo
bact
er v
iola
ceus
PC
C 7
421;
Eco
, E. c
oli.
Pairw
ise
alig
nmen
ts w
ere
gene
rate
d w
ith B
LOSU
M30
usi
ng
Clu
stal
W.
* Inco
mpl
ete
geno
me
sequ
ence
s.

273 T
able
6.2
: Am
ino
acid
sequ
ence
sim
ilari
ties b
etw
een
Ubi
H h
omol
ogs i
n 14
cya
noba
cter
ia a
nd E
. col
i (ex
pres
sed
as %
) U
biH
Lo
cus
tag
6803
70
02
Nos
t N
pun
Tery
Te
lon
8102
Se
lo
Avar
C
wat
M
ED
MIT
S1
20
Glo
e Ec
o Sy
nech
ocys
tis 6
803
Slr1
300
100
Syne
choc
occu
s 70
02*
OR
F 64
10
0
Nos
toc
7120
Al
l427
4 63
65
10
0
N
. pun
ctifo
rme *
N
pun3
352
62
64
91
100
T.
ery
thra
eum
* -
- -
- -
- -
T.
elo
ngat
us B
P-1
Tlr2
289
56
57
57
58
- 10
0
Syne
choc
occu
s 81
02
Synw
0816
42
42
40
41
-
39
100
S. e
long
atus
794
2 *
Selo
0213
43
63
64
65
64
- 43
45
10
0
A. v
aria
bilis
294
13*
Avar
0252
18
66
65
98
90
- 58
42
65
10
0
C
. wat
soni
i 850
1 *
- -
- -
- -
- -
- -
-
P. m
arin
us M
ED4
Pmm
0834
37
38
35
36
-
36
48
41
36
- 10
0
P.
mar
inus
MIT
Pm
t081
2 35
43
42
44
-
42
63
49
45
- 47
10
0
P. m
arin
us S
S120
Pr
o090
6 42
38
36
39
-
36
50
42
37
- 50
41
10
0
G
. vio
lace
us 7
421
Gll2
827
31
32
33
33
- 29
30
38
37
-
27
33
31
100
E.
col
i U
biH
47
52
47
49
-
45
40
49
49
- 38
42
40
29
10
0
Mes
orhi
sobi
um l
oti*
Mlr2
092
33
31
33
34
- 32
26
33
32
-
28
31
27
29
29
68
03, S
ynec
hocy
stis
sp. P
CC
680
3; 7
002,
Syn
echo
cocc
us sp
. PC
C 7
002;
Nos
t, N
osto
c sp
. PC
C 7
120;
Npu
n, N
osto
c pu
nctif
orm
e; T
ery,
Tri
chod
esm
ium
er
ythr
aeum
; Tel
on, T
herm
osyn
echo
cocc
us e
long
atus
BP-
1; 8
102,
Syn
echo
cocc
us sp
. WH
8102
; Ava
r, An
abae
na v
aria
bilis
ATC
C 2
9413
; Sel
o,
Syne
choc
occu
s elo
ngat
us P
CC
794
2; C
wat
, Cro
cosp
haer
a w
atso
nii W
H85
01; P
roch
loro
cocc
us m
arin
us M
ED4;
MIT
, Pro
chlo
roco
ccus
mar
inus
MIT
9313
; S1
20, P
roch
loro
cocc
us m
arin
us S
S120
; Glo
e, G
loeo
bact
er v
iola
ceus
PC
C 7
421;
Eco
, E. c
oli.
Pairw
ise
alig
nmen
ts w
ere
gene
rate
d w
ith B
LOSU
M30
usi
ng
Clu
stal
W. *
Inco
mpl
ete
geno
me
sequ
ence
s.* U
sed
as a
n ou
t-gro
up fo
r phy
loge
netic
reco
nstru
ctio
n (F
ig 6
.3).

274 T
able
6.3
: Am
ino
acid
sequ
ence
sim
ilari
ties b
etw
een
Ubi
D h
omol
ogs i
n 14
cya
noba
cter
ia a
nd E
. col
i (ex
pres
sed
as %
)
Locu
s ta
g 68
03
7002
N
ost
Npu
n Te
ry
Telo
n 81
02
Selo
Av
ar
Cw
at M
ED4
MIT
S1
22
Glo
e Ec
o Sy
nech
ocys
tis 6
803
Sl09
36
100
Syne
choc
occu
s 70
02* O
RF
92
100
N
osto
c 71
20
Alr1
108
93
92
100
N. p
unct
iform
e*
Npu
n188
7 93
92
98
10
0
T. e
ryth
raeu
m*
- -
- -
- -
T. e
long
atus
BP-
1 Tl
r222
5 88
87
88
87
-
100
Sy
nech
ococ
cus
8102
Syn
w10
06
81
79
81
80
- 81
10
0
S.
elo
ngat
u 79
42*
Selo
0226
40
89
89
90
90
- 88
82
10
0
A. v
aria
bilis
294
13*
Avar
4035
01
93
92
98
97
- 87
81
90
10
0
C
. wat
soni
i 850
1*
- -
- -
- -
- -
- -
-
P. m
arin
us M
ED4
- -
- -
- -
- -
- -
- -
P. m
arin
us M
IT
Pmt0
397
78
78
80
79
- 79
90
80
80
-
- 10
0
P. m
arin
us S
S120
Pr
o104
7 79
77
79
79
-
79
85
80
80
- -
88
100
G. v
iola
ceus
742
1 G
ll301
0 85
83
84
84
-
84
77
85
84
- -
76
78
100
E.
col
i K-1
2 U
biD
60
60
60
60
-
59
58
61
61
- -
59
59
59
100
6803
, Syn
echo
cyst
is sp
. PC
C 6
803;
700
2, S
ynec
hoco
ccus
sp. P
CC
700
2; N
ost,
Nos
toc
sp. P
CC
712
0; N
pun,
Nos
toc
punc
tifor
me;
Ter
y, T
rich
odes
miu
m
eryt
hrae
um; T
elon
, The
rmos
ynec
hoco
ccus
elo
ngat
us B
P-1;
810
2, S
ynec
hoco
ccus
sp. W
H81
02; S
elo,
Syn
echo
cocc
us e
long
atus
PC
C 7
942;
Ava
r, An
abae
na
vari
abili
s ATC
C 2
9413
; Cw
at, C
roco
spha
era
wat
soni
i WH
8501
; Pro
chlo
roco
ccus
mar
inus
MED
4; M
IT, P
roch
loro
cocc
us m
arin
us M
IT93
13; S
120,
Pr
ochl
oroc
occu
s mar
inus
SS1
20; G
loe,
Glo
eoba
cter
vio
lace
us P
CC
742
1; E
co, E
. col
i. Pa
irwis
e al
ignm
ents
wer
e ge
nera
ted
with
BLO
SUM
30 u
sing
C
lust
alW
. * In
com
plet
e ge
nom
e se
quen
ces.

275 T
able
6.4
: Am
ino
acid
sequ
ence
sim
ilari
ties b
etw
een
Ubi
X h
omol
ogs i
n 14
cya
noba
cter
ia a
nd E
. col
i (ex
pres
sed
as %
) O
rgan
ism
Lo
cus
tag
6802
70
02
Nos
t N
pun
Tery
Te
lon
8102
Se
lo
Avar
C
wat
M
ED4
MIT
S1
20
Glo
e Ec
o
Syne
choc
ystis
680
3 Sl
r109
9 10
0
Syne
choc
occu
s 70
02*
OR
F 81
10
0
Anab
aena
712
0 Al
r124
1 85
83
10
0
N. p
unct
ofor
me*
N
pun3
020
87
84
92
100
T. e
ryth
raeu
m*
(Ter
y298
0)
(64)
-
- -
-
T. e
long
atus
BP-
1 Tl
r222
5 80
78
80
79
-
100
Syne
choc
occu
s 81
02
Synw
1201
69
67
66
68
-
67
100
S. e
long
atus
794
2*
Selo
0211
96
79
76
78
77
- 78
64
10
0
A. v
aria
bilis
294
13*
(Ava
r334
101)
(9
1)
- -
- -
- -
- -
C. w
atso
nii 8
501*
-
- -
- -
- -
- -
- -
P. m
arin
us M
ED4
- -
- -
- -
- -
- -
- -
P. m
arin
us M
IT93
13
Pmt0
767
70
65
64
66
- 68
84
64
-
- -
100
P. m
arin
us S
S120
Pr
o098
9 71
66
66
65
-
68
78
66
- -
- 78
10
0
G. v
iola
ceus
742
1 G
ll052
9 78
77
80
80
-
78
77
74
- -
- 68
68
10
0
E. c
oli
Ubi
X 52
52
50
52
-
54
48
52
- -
- 50
55
52
10
0
68
03, S
ynec
hocy
stis
sp. P
CC
680
3; 7
002,
Syn
echo
cocc
us sp
. PC
C 7
002;
Nos
t, N
osto
c sp
. PC
C 7
120;
Npu
n, N
osto
c pu
nctif
orm
e; T
ery,
Tri
chod
esm
ium
er
ythr
aeum
; Tel
on, T
herm
osyn
echo
cocc
us e
long
atus
BP-
1; 8
102,
Syn
echo
cocc
us sp
. WH
8102
; Sel
o, S
ynec
hoco
ccus
elo
ngat
us P
CC
794
2; A
var,
Anab
aena
va
riab
ilis A
TCC
294
13; C
wat
, Cro
cosp
haer
a w
atso
nii W
H85
01; P
roch
loro
cocc
us m
arin
us M
ED4;
MIT
, Pro
chlo
roco
ccus
mar
inus
MIT
9313
; S12
0,
Proc
hlor
ococ
cus m
arin
us S
S120
; Glo
e, G
loeo
bact
er v
iola
ceus
PC
C 7
421;
Eco
, E. c
oli.
Pairw
ise
alig
nmen
ts w
ere
gene
rate
d w
ith B
LOSU
M30
usi
ng
Clu
stal
W. *
Inco
mpl
ete
geno
me
sequ
ence
s. Th
e N
-term
inal
107
and
62
resi
dues
wer
e tru
ncat
ed in
the
tery
2980
and
Ava
r334
101
sequ
ence
s wer
e 10
7 an
d 62
re
sidu
es a
s com
pare
d to
the
Slr1
099
sequ
ence
.
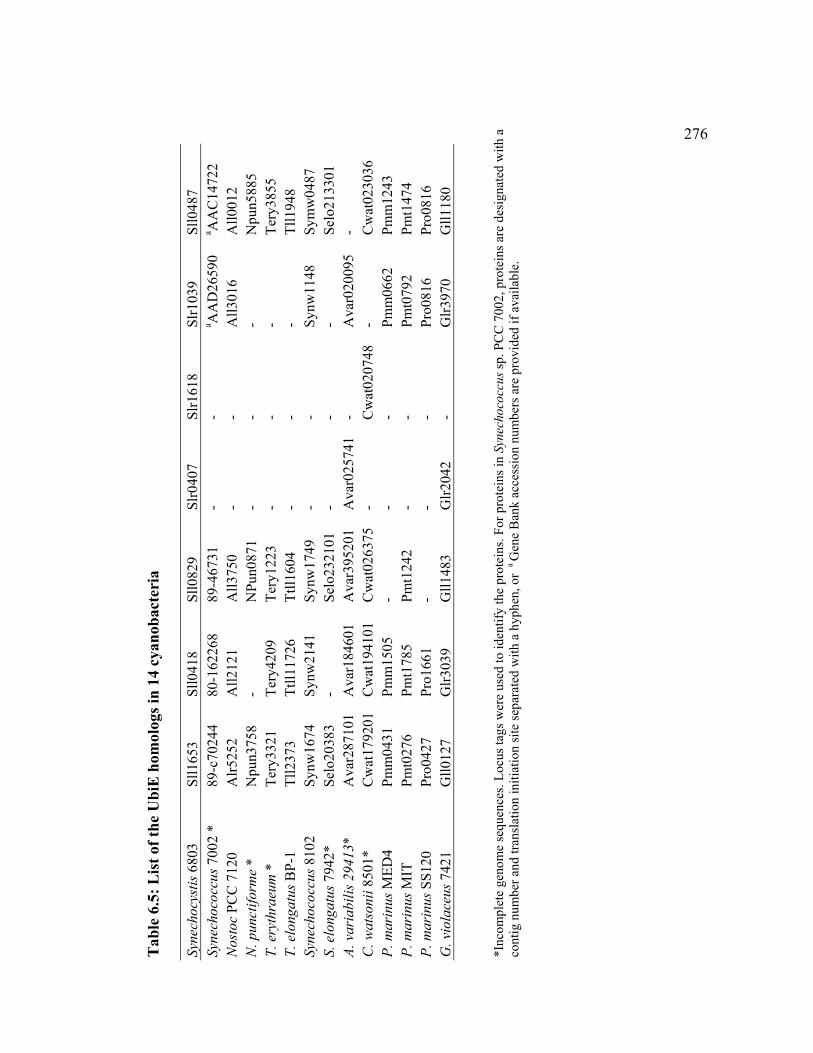
276 T
able
6.5
: Lis
t of t
he U
biE
hom
olog
s in
14 c
yano
bact
eria
Syne
choc
ystis
680
3 Sl
l165
3 Sl
l041
8 Sl
l082
9 Sl
r040
7 Sl
r161
8 Sl
r103
9 Sl
l048
7 Sy
nech
ococ
cus 7
002
* 89
-c70
244
80-1
6226
8 89
-467
31
- -
a AA
D26
590
a AA
C14
722
Nos
toc
PCC
712
0 A
lr525
2 A
ll212
1 A
ll375
0 -
- A
ll301
6 A
ll001
2 N
. pun
ctifo
rme
* N
pun3
758
- N
Pun0
871
- -
- N
pun5
885
T. e
ryth
raeu
m *
Te
ry33
21
Tery
4209
Te
ry12
23
- -
- Te
ry38
55
T. e
long
atus
BP-
1 Tl
l237
3 Tt
ll117
26
Ttll1
604
- -
- Tl
l194
8 Sy
nech
ococ
cus 8
102
Synw
1674
Sy
nw21
41
Synw
1749
-
- Sy
nw11
48
Sym
w04
87
S. e
long
atus
794
2*
Selo
2038
3 -
Selo
2321
01
- -
- Se
lo21
3301
A.
var
iabi
lis 2
9413
* A
var2
8710
1 A
var1
8460
1 A
var3
9520
1 A
var0
2574
1 -
Ava
r020
095
- C
. wat
soni
i 850
1*
Cw
at17
9201
C
wat
1941
01
Cw
at02
6375
-
Cw
at02
0748
-
Cw
at02
3036
P.
mar
inus
MED
4 Pm
m04
31
Pmm
1505
-
- -
Pmm
0662
Pm
m12
43
P. m
arin
us M
IT
Pmt0
276
Pmt1
785
Pmt1
242
- -
Pmt0
792
Pmt1
474
P. m
arin
us S
S120
Pr
o042
7 Pr
o166
1 -
- -
Pro0
816
Pro0
816
G. v
iola
ceus
742
1 G
ll012
7 G
lr303
9 G
ll148
3 G
lr204
2 -
Glr3
970
Gll1
180
*Inc
ompl
ete
geno
me
sequ
ence
s. Lo
cus t
ags w
ere
used
to id
entif
y th
e pr
otei
ns. F
or p
rote
ins i
n Sy
nech
ococ
cus s
p. P
CC
700
2, p
rote
ins a
re d
esig
nate
d w
ith a
co
ntig
num
ber a
nd tr
ansl
atio
n in
itiat
ion
site
sepa
rate
d w
ith a
hyp
hen,
or
a G
ene
Ban
k ac
cess
ion
num
bers
are
pro
vide
d if
avai
labl
e.

277
Table 6.6: Ycf21/UbiC homologs in cyanobacteria and eukaryotic phototrophs
Organism Locus tag
Synechocystis sp. PCC 6803 Sll1797
Crocosphaera watsonii WH 8501 Cwat020398
Nostoc sp. PCC 7120 All0938
Nostoc punctiforme Npun5760
Anabaena variabilis ATCC 29413 Avar303301
Trichodesmium erythraeum Tery3876
Thermosynechococcus elongatus Tll1562
Synechococcus elongatus sp. PCC 7942 Selo236201
Synechococcus sp. WH8102 Synw2384
Prochlorococcus marinus sp. MIT9313 Pmt2179
Prochlorococcus marinus sp. MED4 Pmm1676
Prochlorococcus marinus sp. SS120 Pro1837
Porphyra purpurea (red algae) PopuCp195
Cyanophora paradoxa (a photoautotrophic protist) CypaCp147

278
Table 6.7: Sequences of primers used for PCR analyses Lower case letters indicate heterologous nucleotides that are introduced in order to engineer restriction enzyme cleavage sites, which are indicated with under lines. Primer Sequence
sll0829-F 5’-GGTCCGCCAAGATTTAGCAGTAAG-3’
sll0829-R 5’-AGATTGGGACAGTAATCGAAGGC-3’
sll0418-F1 5’-CGACTGAGGAAACGGTTGAaTTCCCGCACC-3’ (EcoRI cleavage site).
Used for cloning the sll0418 coding region
sll0418-R1 5’-CTTACGTGGCAATTTAAGCTTGAGTGGCGT-3’
Used for cloning the sll0418 coding region
sll0418-F2 5’-ATGCCCGAGTATTTGCTTCTGCC-3’
Used for verification of segregation
sll0418-R2 5’-GCACTGCTTTGAACATACCGAAG-3’
Used for verification of segregation
slr0089-F 5’-TCTACCGGAAATTGCCAACTACCA-3’
slr0089-R 5’-CCTAGGAGATTGTGGACTTCAA
slr0407-F 5’-CTCTGGAAACATTTGAAAgCTTTATCCG-3’ (HindIII cleavage site)
slr0407-R 5’-TACGTTACTATTCCGCCGAaTTCCTCT-3’ (EcoRI cleavage site)
slr1618-F 5’-TCTATTCTGGATATGCTGGCACA-3’
slr1618-R 5’-TTGTTTGCCTTCGGCTTTAGC-3’
slr1039-F 5’-GGGTGTTTACAATAACAGGGC-3’
slr1039-R 5’-ACTGGTGGGAGGTTGGTGCC-3’

279
Table 6.8: Insertional inactivation constructs of the methyltransferase mutants in Synechocystis sp. PCC 6803. HindIII (blunt) indicates that the 3’-overhand generated after digestion with HindIII is filled by Klenow to yield a blunt end.
Target allele
(Construct)
Restriction digestions
Site of insertion in pUC19 Insertion of drug cartridge
sll0829 (p829Gm) HindIII, HincII HindIII, HincII aacC1 between the BxtXI
sites
sll0418 (p418Sp) EcoRI, HindIII EcoRI, HindIII aadA at the unique SmaI
site
slr0407 (p407Sp) EcoRI, HindIII EcoRI, HindIII aadA between tow HincII
sites
slr1618 (p1618Sp) HindIII, HincII HindIII, HincII aadA at the unique NheI
site
slr1039 (p1039Sp) EcoRI, HindIII EcoRI, HindIII aadA at the unique MfeI
site
slr0089 (p89Gm)
HindII (blunt), KpnI
HindII (blunt), KpnI
aacC1 between the two PstI sites
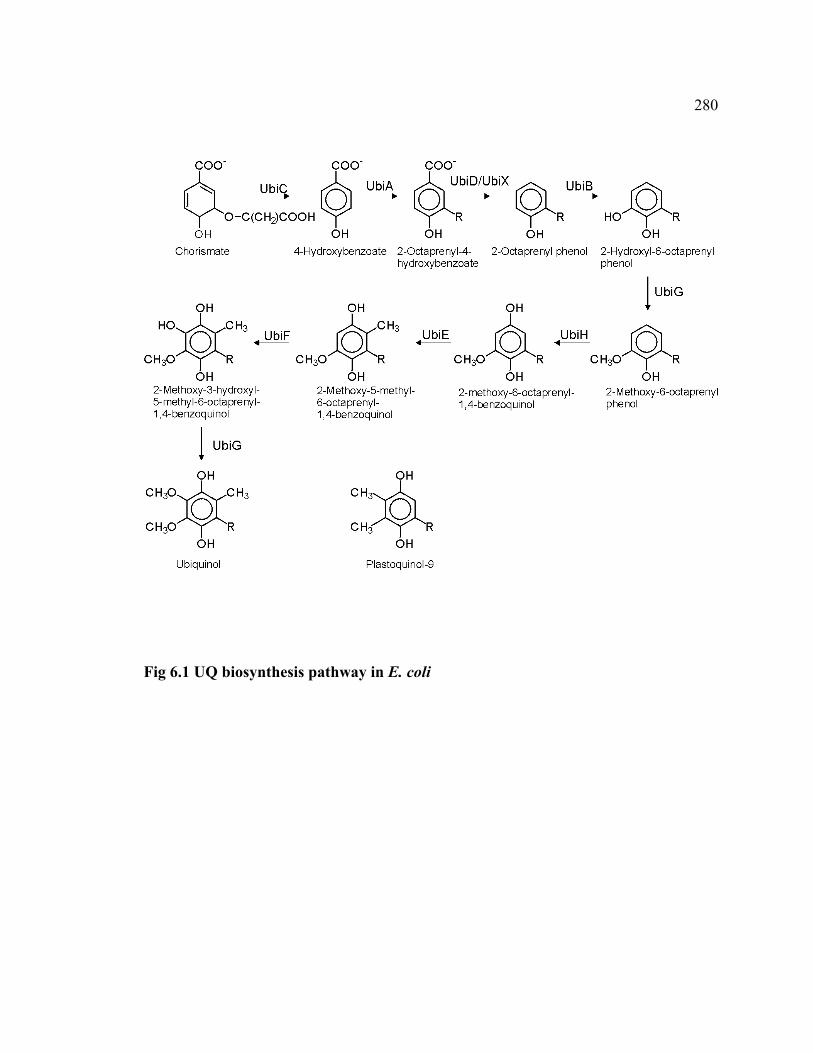
280
Fig 6.1 UQ biosynthesis pathway in E. coli

281
Fig 6.2. Hypothetical PQ-9 biosynthesis pathway in cyanobacteria The question mark indicates the possibility that an alternative route exists for 4-hydroxybenzoate synthesis.

282
Fig 6.3: An alignment of the Ycf21/UbiC homologs in cyanobacteria and eukaryotic phototrophs. Two catalytic residues (arginine at the position 76 and glutamate at position 155) of the UbiC protein are shown by black arrows.
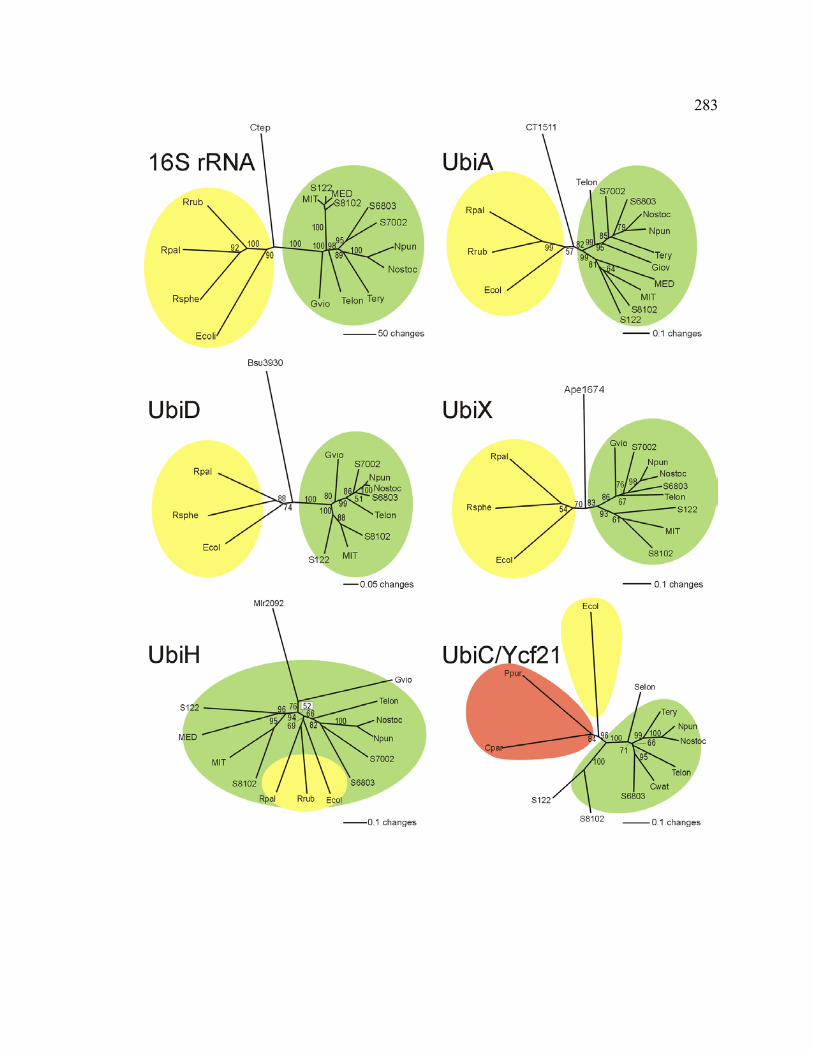
283

284
Fig 6.4: Phylogenetic analyses based on nucleotide sequences of 16S rRNA and amino acid sequences of Ubi proteins in cyanobacteria and proteobacteria. Multiple alignments were generated with BLOSUM30 using ClustalW. Phylogenetic reconstruction was carried out using the Neighbor Joining method using PAUP* 4.0 beta. Bootstrap values were obtained after 1000 times repetitions. Ctep, Chlorobium tepidum; Cpar, Cyanophora paradoxa; Cwat, Crochosphaera watsonii WH 8501; Ecol, E. coli; Gvio, G. violaceus PCC 7421; Nostoc, Nostoc sp. PCC 7120; Npun, N. punctiforme; PMM, P. marinus MED4; PMT, P. marinus MIT 9313; S122, P. marinus SS120; Ppur, Porphyra purpurea; Rpal, Rps. palustris; Rrub, Rs. rubrum; Rsphe, Rb. sphaeroides; S6803, Synechocystis sp. PCC 6803; S7002, Synechococcus sp. PCC 7002; Synw, Synechococcus sp. WH 8102; Selon, Synechococcus elongatus PCC 7942; Tery, T.. erythraeum; Telon, T. elongatus. Aeropyrum pernix K1 (locus tag: Ape1674), C. tepidum (locus tag: CT1511), Bacillus halodurance (locus tag: Bh3930), and Mesorhizobium loti (locus tag: Mlr2092) were used as out-groups.
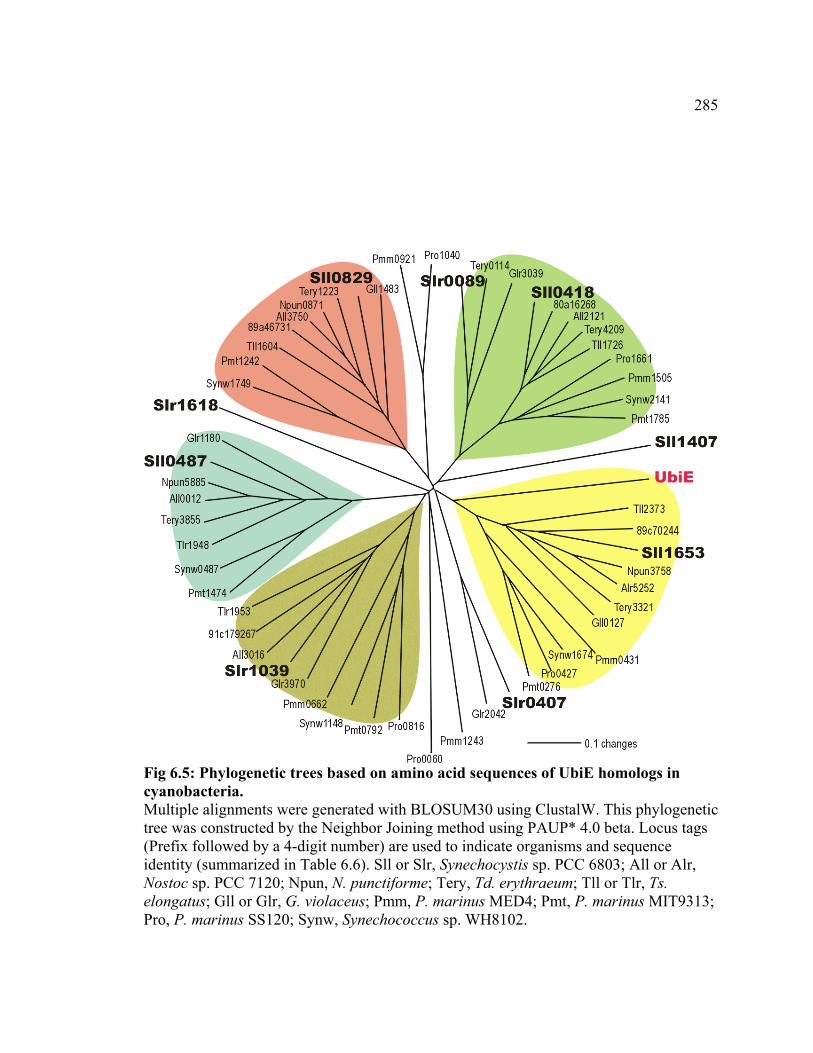
285
Fig 6.5: Phylogenetic trees based on amino acid sequences of UbiE homologs in cyanobacteria. Multiple alignments were generated with BLOSUM30 using ClustalW. This phylogenetic tree was constructed by the Neighbor Joining method using PAUP* 4.0 beta. Locus tags (Prefix followed by a 4-digit number) are used to indicate organisms and sequence identity (summarized in Table 6.6). Sll or Slr, Synechocystis sp. PCC 6803; All or Alr, Nostoc sp. PCC 7120; Npun, N. punctiforme; Tery, Td. erythraeum; Tll or Tlr, Ts. elongatus; Gll or Glr, G. violaceus; Pmm, P. marinus MED4; Pmt, P. marinus MIT9313; Pro, P. marinus SS120; Synw, Synechococcus sp. WH8102.

286
Fig 6.6: HPLC analysis of the sll1653 mutant of Synechocystis sp. PCC 6803 and the 2-phytyl-1,4-benezoquinone (MPBQ) standard. (A) Chromatograms of the whole-cell extract of the sll1653 mutant of Synechocystis sp. PCC 6803 (solid line) and the MPBQ standard (dotted line). (B) Absorption spectra of PQ-9 eluting at 35.5 min in the sll1653 mutant sample (dotted line) and of MPBQ eluting at 33 min.
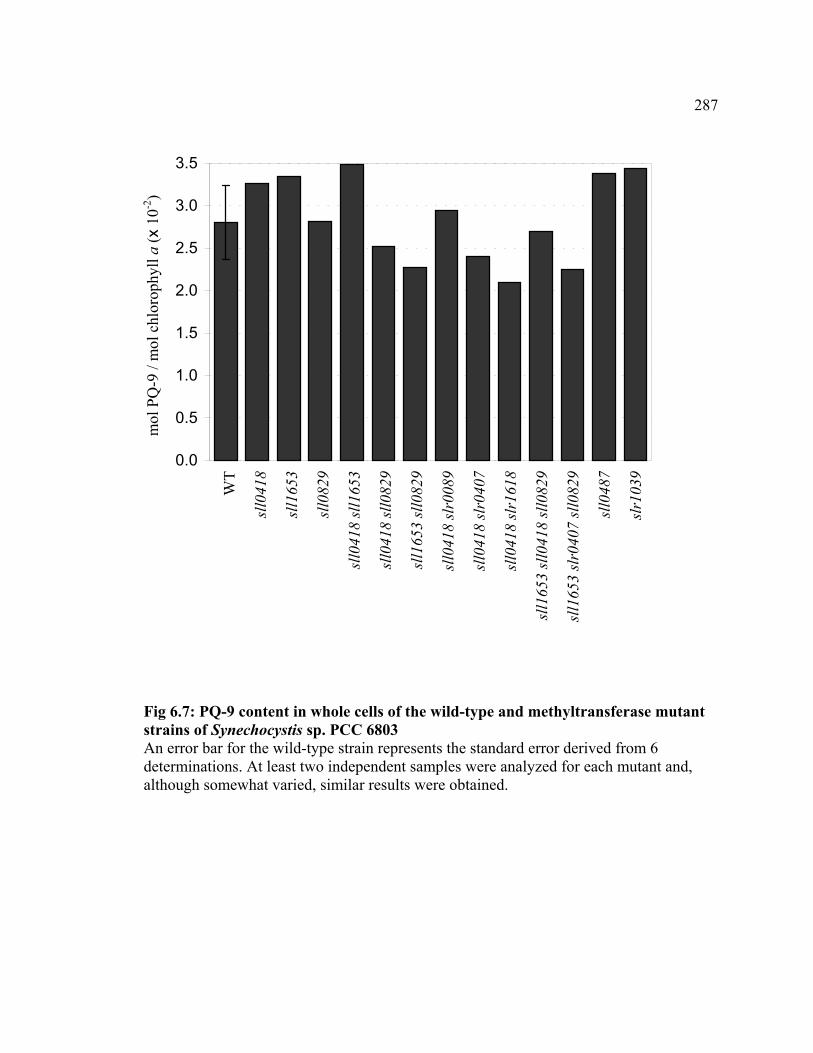
287
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Fig 6.7: PQ-9 content in whole cells of the wild-type and methyltransferase mutant strains of Synechocystis sp. PCC 6803 An error bar for the wild-type strain represents the standard error derived from 6 determinations. At least two independent samples were analyzed for each mutant and, although somewhat varied, similar results were obtained.
mol
PQ
-9 /
mol
chl
orop
hyll
a (x
10-2
)
WT
sll0
418
sll1
653
sll0
829
sll0
418
sll1
653
sll0
418
sll0
829
sll1
653
sll0
829
sll0
418
slr0
089
sll0
418
slr0
407
sll0
418
slr1
618
sll1
653
sll0
418
sll0
829
sll1
653
slr0
407
sll0
829
sll0
487
slr1
039

288
Fig 6.8: Restriction maps of coding regions and their flanking sequences for the UbiE homologs in Synechocystis sp. PCC 6803. Primers used for PCR for both cloning and PCR verifications are shown by black arrows. F1 and R1 primers were used for cloning the sll0418 coding region, while F2 and R2 primers were used for verification of segregation.
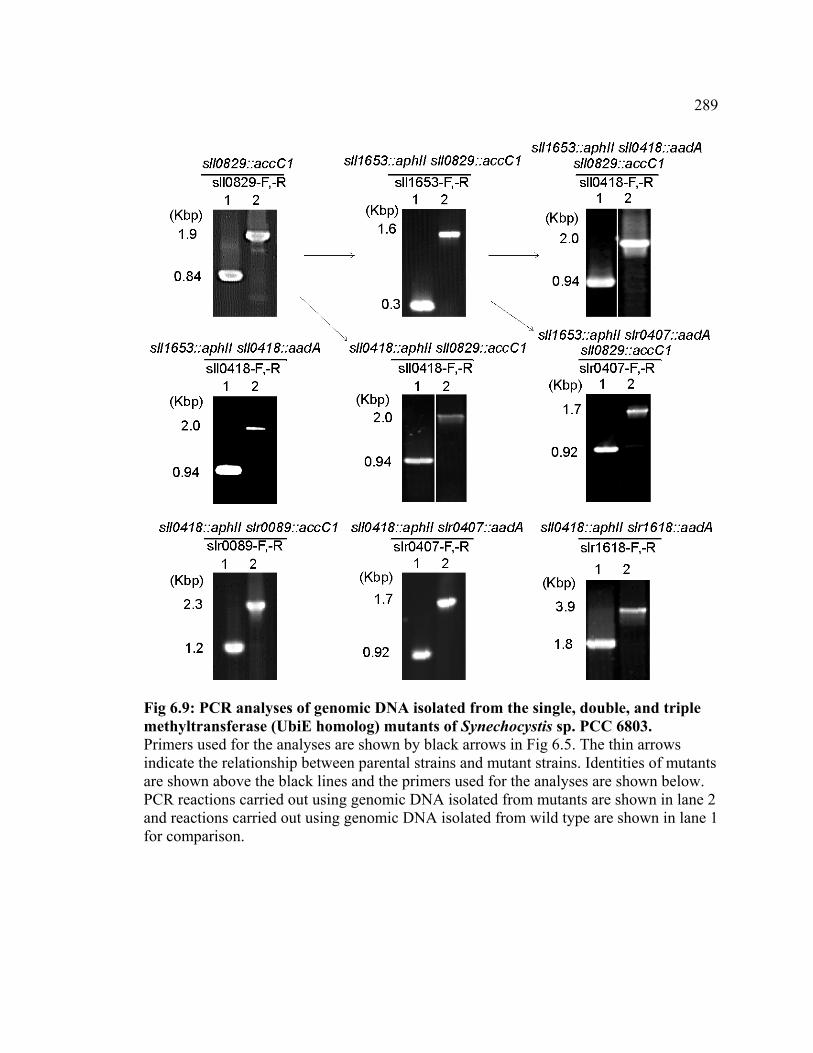
289
Fig 6.9: PCR analyses of genomic DNA isolated from the single, double, and triple methyltransferase (UbiE homolog) mutants of Synechocystis sp. PCC 6803. Primers used for the analyses are shown by black arrows in Fig 6.5. The thin arrows indicate the relationship between parental strains and mutant strains. Identities of mutants are shown above the black lines and the primers used for the analyses are shown below. PCR reactions carried out using genomic DNA isolated from mutants are shown in lane 2 and reactions carried out using genomic DNA isolated from wild type are shown in lane 1 for comparison.

290
Fig 6.10: Aromatic amino acid biosynthesis and an alternative proposal for 4-hydroxybenzoate synthesis in cyanobacteria. CM, chorismate mutase; PD, prephenate dehydratase; PPAT, prephenate amino transferase; ADH, arogenenate dehydrogenase; TAL, tyrosine amino-lyase; PAT, phenylalanine aminotransferase; PAL, phenylalanine amino-lyase; CH, chorismate hydratase. The pathways leading to tyrosine and phenylalanine biosynthesis in cyanobacteria are as previously described (Bonner et al., 1995).

291
Appendix
A.1. Amino acid sequence alignments of phylloquinone (Men) and plastoquinone (Ubi)
biosynthetic enzymes.
A.1.1. MenA homologs in cyanobacteria and other organisms
A.1.2. MenB homologs in cyanobacteria and other organisms
A.1.3. MenD homologs in cyanobacteria and other organisms
A.1.4. UbiA homologs in cyanobacteria and other organisms
A.1.5. UbiD homologs in cyanobacteria and other organisms
A.1.6. UbiH homologs in cyanobacteria and other organisms
A.1.7. UbiX homologs in cyanobacteria and other organisms
A.1.8. UbiE homologs in Synechocystis sp. PCC 6803
A.1.9. Sll1653 homologs in cyanobacteria
A.1.10. Sll0418 homologs in cyanobacteria
A.1.11. Sll0829 homologs in cyanobacteria
A.1.12. Sll0487 homologs in cyanobacteria
A.1.13 Slr1039 homologs in cyanobacteria
A.2. Photosystem I (PS I) complexes
A.2.1. Polypeptide composition of PS I complexes isolated from the
menG, menF, and rubA menB mutants of Synechococcus sp. PCC
7002.
A.2.2. Illustration of genetically engineered PS I in cyanobacteria

292
A.1.1. An amino acid sequence alignment of MenA homologs in cyanobacteria and other
organisms
Slr1518MenA7002Alr0033Tlr2196Gll1578Synw2307Pmm0176At1g60600OJ1217B0918CymeCp095B3930Bsu38490CT1511AF2036
1 32
1 31
1 35
1 27
1 22
1 31
1 22
1 6
1 80
1 20
1 25
1 35
1 28
1 20
M T E S S P L A P S T A P A T R K L W L A A I K P P M Y T V A V
M L T E N P P T S S P K A T R R L W L A A I K P P I Y T V A V
M L N C L P Q L M T T K Q I S H P K I K L W M A A I K P P M Y S V A I
M T S S S E S I D R S R L W W A A I K L P L Y S V A V
M S G S P L R P W L N S F N P L I Y T A S V
M V E P Q A V A S R Y A D R R L L W K A A I K W P M Y S V A V
M N E R N K S L W K Q A I K W P L Y S V A I
M Y S V A L
M P L A G I A L A P L F V S H L A P P H H R R S S V A S A A A A A A R R R P R A V Q C S A T A T A A S G E G A G D D V E L S R G T L L W R A A K L P I Y S V A L
M N K I V T L F L A S R P K T L L V S F
M T E Q Q I S R T Q A W L E S L R P K T L P L A F
M N Q T N K G E G Q T A P Q K E S M G Q I L W Q L T R P H T L T A S F
M S V S S S S Q L S A F Q A W M L A I R P K T L P A G A
M E K L K A L I E V T K P K Q T F L L M
M P L A G I A L A P L F V S H L A P P H H R R S S V A S A A A A A A R R R P R A V Q C S A M S L W A I K P P Y . V A .
Slr1518MenA7002Alr0033Tlr2196Gll1578Synw2307Pmm0176At1g60600OJ1217B0918CymeCp095B3930Bsu38490CT1511AF2036
33 104
32 103
36 107
28 99
23 94
32 104
23 94
7 78
81 152
21 84
26 104
36 108
29 107
21 90
V P I T V G S A V A Y G L T G Q W H G D V F T I F L L S A I A I I A W I N L S N D V F D S D T G I D V R - - - - - - - - K A H S V V N L T G N R N L V F L I S N
T P V I V T T A A A Y G E F G V F N P V R F G L F L G A A I C L I A W M N L S N D V F D A D T G I D V N - - - - - - - - K A T S V V N L T G D R P L V F G I A L
M P I W V G G A V A F A G N K V F N L A V F F T F I V A A I L I L A W E N I S N D V F D S E T G I D V N - - - - - - - - K H H S L V N L T K K K T L V F W L G N
M P I W L G T A V A I A K T G R G H W W P F G L F L T A A V L I L A W L N L S N D V F D A E T G I D R H - - - - - - - - K Y H S V V N L T G K K Q L I F W L S N
I P I L V G T A F A W W Q G G V F D L Q R F A L T L A G L V F V H A W I N L T N D V F D W D T G V D R N - - - - - - - - K L N S L V R T T G S R S Q V L W V G N
M P V L L A A G W R I G A V G S L R W T Q L F G F L L A A I L L L L W E N L S N D L F D A D T G V D T T G - - - - - - - K P H S V V N L T G R R D R V A L A S L
L P V F I S G A Y T L N S F K N V K I Y N L I A F T I A A I L I L I W E N L T N D L F D S E T G I D E F - - - - - - - - K F H S I V N L V R S K T I V S I T A Y
V P L T V G A S A A Y L E T G L F L A R R Y V T Q L L S S I L I I T W L N L S N D V Y D F D T G A D K N - - - - - - - - K M E S V V N L V G S R T G T L A A A I
V P L T V G S A C A Y H H V G S F F G K R Y F V L L V A S V L V I T W L N L S N D V Y D S D T G A D K N - - - - - - - - K K E S V V N I V G S R T M T Q Y A A N
C C W L S V S S L Q H A F - - - - S I P A L L L A I G - - - - L Q I W A N W V N D Y L D F M N G I D H A - - - - - - - - L R Q G P T R W T S R L T P S F M K I I
A A I I V G T A L A W W Q - G H F D P L V A L L A L I T A G L L Q I L S N L A N D Y G D A V K G S D K P D R I G P L R G M Q K G V I T Q Q E M K R A L I I T V V
V P V L L G T V L A M F Y - V K V D L L L F L A M L F S C L W I Q I A T N L F N E Y Y D F K R G L D T A E S - - - - - - V G I G G A I V R H G M K P K T I L Q L
M P V V I G A A L A A A S - G V F K P L P A L V A L I C A L G I Q I A T N F I N E I Y D F R K G A D T A E R L G P T R T V A A G I I T E Q T M I R V S I V L G V
I T F L V S Y I V A R G G - A D F N F V I A A I S M F L A I S G T T A I N M W L D R - D I D A I M P R T - - - - - - - - R K R P V P A G I L K P S E C A A F G A
. P . . V G . A . A . G F . . . . L . . A I L . . . W N L . N D . F D . T G . D R G P R K S V V N . . . . . . . . .
Slr1518MenA7002Alr0033Tlr2196Gll1578Synw2307Pmm0176At1g60600OJ1217B0918CymeCp095B3930Bsu38490CT1511AF2036
105 176
104 174
108 179
100 171
95 162
105 177
95 169
79 154
153 231
85 146
105 177
109 181
108 174
91 157
F F L L A G V L G L M S M S W R A Q D - W T V L E L I G V A I F L G Y T Y Q G P P F R L G Y L G L G E L I C L I T F G P L A I A A A Y Y S Q S Q S - - - - - - -
G F L G L G I T G I A L I S Y L Q Q D - W M V F N L L L L A T A I A Y T Y Q G P P F R L G Y L G I G E F V C F L A Y W I T G Q G V F Y S - Q A Q T - - - - - - -
L C L I S G L L G I L A I A T W Q Q D - P T V I G I I L L C C G L G Y M Y Q G P P F R L G Y Q G L G E I L C F F A F G P L G V S A G Y Y S Q T Q T - - - - - - -
L C L L L G L L G I A A I S W L Q Q D - G T V L G L V L L C C A L G Y S Y Q G P P F R L G Y L G L G E P I C F I C F G P L A I A A A Y Y S Q V Q T - - - - - - -
I F L S I G Y V - - - - C F W A L Q D - W W L L A F G S L G I F L G Y A Y Q G P P F R L A Y K G L G E W I S S S C F G P I A V L T A T R A Q T G S - - - - - - -
A S L G S G L A L M A W L A W C S S - - V L V L A L V L L C C G L G Y I Y Q G P P F R L G Y R G L G E P L C W L A F G P C A T A A A L M V L E P Q - - - - - G A
T S L L I G L V V I A I I S I S T S - - I N V M L L V G A C C F L G Y L Y Q G P P F R L G Y Q G L G E P L C W L A F G P F A Y A A A L I A L N P S D - - - I Y M
T S L A L G V S G L V W T S L N A S N - I R A I L L L A S A I L C G Y V Y Q C P P F R L S Y Q G L G E P L C F A A F G P F A T T A F Y L L L G S - - - S S E M R
I S L L F G F M G L F W A F A Q A G D - A R F I L S V T C A I I C G Y V Y Q C P P F R L S Y R G L G E P L C F A A F G P L A T T A F Y F S N S S R N I S S G T A
L C L W C C L L F V C A S Y - - - - - - F K M K I L V G I L I L I A W F Y S - - - - - A W L G F A G E W L S F F A C G P L A T Y L F S S I Y Q V S - - - - - - -
L I C L S G L A L V A V A C H T L A D F V G F L I L G G L S I I A A I T Y T V G N R P Y G Y I G L G D I S V L V F F G W L S V M G S W Y L Q A H T - - - - - - -
A L A S Y G I A I L L G V Y I C A S S S W W L A L I G L V G M A I G Y L Y T G G P L P I A Y T P F G E L F S G I C M G S V F V L I S F F I Q T D K - - - - - - -
S V F V L G L Y L V A I G G - - - - - - W P I L L I G V L S L L F A W A Y T G G P F P I A Y S G L G D V F V F I F F G L V A V G G T Y Y V Q A L S - - - - - - -
A I F A I G Q V L A F L V S L E F G L V V F F G L F F D I V V Y T I L L K R K S P Y S I V L G G F A G A M P A L G - G W V A V Q G F T L - - - - - - - - - - - -
. L . . G . . . . . . . D . . . L . . . . . . . G Y . Y Q G P P F R L . Y G L G E . C . . . F G P . A . . . . . Q . I S S
Slr1518MenA7002Alr0033Tlr2196Gll1578Synw2307Pmm0176At1g60600OJ1217B0918CymeCp095B3930Bsu38490CT1511AF2036
177 252
175 250
180 255
172 247
163 238
178 256
170 248
155 234
232 311
147 223
178 253
182 257
175 249
158 231
- - - F S W N L L T P S V F V G I S T A I I L F C S H F H Q V E D D L A A G K K S P I V R L G T K L G S Q V L T L S V V S L Y L I T A I G V L C H Q A P W Q - T
- - - L T L N A V W A S A W V A L T T S I I L F C S H F H Q A E D D L A A G K K S P I V R L G T Y Q G A Q V L T Y S L G A V L V L T C L L V A V G T W S P W - M
- - - W S I T S L A A S V I V G I V T S L V L F C S H F H Q V D D D I K A G K R S P I V R L G T A N G A K V L T W F T A S I Y P F S L L F V L L G F F P V W - T
- - - F S A S I W P V A I L N G L T T T L I L F C S H F H Q V A D D L A A G K R S P V V R L G T A R S A Q L V Y G A C G L F Y G V L V A S V L W G L L P W P - T
- - - W E L G A L L A G I A V G S W T A T I L Y A H H F Y Q Y E D D R E F D K R T P V V R W G P E L A A K R F W W V I S P S Y V V L L L A V I G G F L P W T - V
N V I P W A T A W M L G A G P A V A T S L V L F C S H F H Q V S E D S A H G K R S P V V R L G T A R A A A L I P W L V A L T L A L E W L P M C R G D W P L T - V
I S I P W K E S L L L G S G S S L A T T L V L F C S H F H Q I K E D K E H G K N S P L V L L G A K K G A K I I P W I V F I I Y V F Q L F L I I N G F I P I L - C
H L P L S G R V L S S S V L V G F T T S L I L F C S H F H Q V D G D L A V G K Y S P L V R L G T E K G A F V V R W T I R L L Y S M L L V L G L T R I L P L P C T
L L P L S K T V I A S S I L V G L T T T L I L F C S H F H Q I D G D R A V G K M S P L V R I G T K T G S R L V T L G V V T L Y V L L A A F G M S K S L P S A C T
- - F W Q W D L L L L S I I N G S W S A A L L M A N N M R D R I S D T H S P K K S T C V W F G Q D W A S M Q Y Q M F I L M C I W C L F A W L I F R P S W F W - L
- - - L I P A L I L P A T A C G L L A T A V L N I N N L R D I N S D R E N G K N T L V V R L G E V N A R R Y H A C L L M G S L V C L A L F N L F S L H S L W - G
- - - I N M Q S I L I S I P I A I L V G A I N L S N N I R D I E E D K K G G R K T L A I L M G H K G A V T L L A A S F A V A Y I W V V G L V I T G A A S P W - L
- - - L P M E V L V A A A A P G A F S V C I L L V N N I R D I D T D R K V G K M T L P A R I G A P A A R A L Y V A L V V L A Y L V P - F Y M I S T G Y S L W - C
- - - - - P G F I I A A I V L L W I P S H I W Y I S M H Y E E D Y R M A N I P M Y P L V - V G M E R A S W A I V F A T A A M L V L A A S L Y V L L P L G I F Y L
. . . . . . . G . T . . I L F C S H F H Q . . . D . G K . S P . V R L G . . . . . . . . . Y . . . . . . . . . P C
Slr1518MenA7002Alr0033Tlr2196Gll1578Synw2307Pmm0176At1g60600OJ1217B0918CymeCp095B3930Bsu38490CT1511AF2036
253 307
251 317
256 309
248 298
239 295
257 310
249 305
235 287
312 346
224 269
254 308
258 311
250 307
232 281
L L I I A S L P W A V Q L I R H V G Q Y H D Q P E Q V S N C K F I A V N L H F F S G M L M A A G Y G W A G L G
L L T F A S A P F A L S L I N H V R L N H D Q P D K V S N C K F F A V K F H F A S G L L L A I A Y V L S Y Y N L E F L S V T G L G Y N
L L S W L S L P Y A F K L C R H V Q H N H N Q P E K V S N C K F I A V A V H F W S C L L L G L G F V I A G V
L L A L S S L P W A I Y L C Q R M A Q F H S V P E Q I K N S K F I A V Q L H F W S S L L L G L G Y L L
L A A L A T I P L A V S L V S F V R E N A T N P A V V V G A L P L A A R F H L V S G A L L T L G L L A G T L L P R
L L S G L G L P A G L A L I R L M Q R C H D Q P D R I S G S K F L A L R F Q A W N G L G L A L G L A L G R L
V L F L I S F P Q S L K L I N L L K Y S Y N K P E A I K N C K F I A I K F Q T L N G I G L I A G F I I N Y L I Y K
L M C F L T L P V G N L V S S Y V E K H H K D N G K I F M A K Y Y C V R L H A L L G A A L S L G L V I A R
V L C A L T L P V G K W V V D Y V L K N H E V T S D L T Y N L D A N L
L M M C V F L P Y A L R L C M M S L T T P T - - A L L Y P T L S F I F R Y C F L F S F C A S Y S
W L F L L A A P L L V K Q A R Y V M R E M D P - V A M R P M L E R T V K G A L L T N L L F V L G I F L S Q W A A
F V V F L S V P K P V Q A V K G F V Q N E M P - M N M I V A M K S T A Q T N T F F G F L L S I G L L I S Y F R
L L S L L S I P L A I G M V R T L Y A S E G - - Q A L N A V L A G T G K V L T V H G L L F S L G L V I P N I I S I F R P
V I S T S A V A F F L Y K A V K F A L S P D R - V K A R K M Y K L A S M T L G L V Y F S L L L G V F L
L L . . . . P . . L . . P . . A . . . G . L L . L G . . F V T G L G Y N
The multiple alignment was generated by BLOSUM30 using Clustal W. The locus tags used to indicate organisms are summarized in Table 2.1, except for the following: Cymecp095 in Cyanidiochyzon merolae and OJ1217B09.18 in Oryza sativa. AF2036, a hypothetical protein in the archaeon Archeoglobus fulgidis DSM4304, was used as an out-group in phylogenetic construction as shown in Fig 2.3.

293
A.1.2. An amino acid sequence alignment of MenB homologs in cyanobacteria and other
organisms
Sll1127MenB7002All2347Tll2458Synw0998Pmm0608At1g60550P0671D0118CymeCp092B2262CT1846Bsu3075AF0435
1 5
1 8 1 7
1 8
1 18
1 14
1 67
1 64
1 0
1 21
1 27
1 6
1 0
M D W H I
M E S L A W Q AM Q I D W Q S
M D S V H W R Q
M A E L R Q V L P G D P A A A W T A
M K I L P G E T N S N W T E
M A D S N E L G S A S R R L S V V T N H L I P I G F S P A R A D S V E L C S - - - A S S M D D R F H K V H G E V P T H E V V W K K T D F F G
M D A A G R R L A R V T A H L L P S S L P L P L A S A P T L A P S P A A S P A S D S Y R R V H G D V P S E P P E W R A A T D E S
M I Y P D E A M L Y A P V E W H D C S E G - - - -
M P V E P G Q R R F S S T T I E I S D M S T V N W I T
M A E W K T
M A D S N E A R R L V T H L P A L S P A A S . W
Sll1127MenB7002All2347Tll2458Synw0998Pmm0608At1g60550P0671D0118CymeCp092B2262CT1846Bsu3075AF0435
6 70
9 73
8 72
9 73
19 84
15 80
68 132
65 126
1 56
22 79
28 88
7 66
1 55
- - - A K H Y D D I L Y Y K A G - - G I A K I V I N R P H K R N A F R P Q T V F E L Y D A F C N A R E D N R I G V V L L T G A G P H S D G K
- - - V K A Y E D I L Y Q K L D - - G I A K I T I N R P H K R N A F R P Q T I F E M Y E A F C D A R E D Q D I G V I L L T G A G P H T D G K
- - - T K T Y E D I L Y D K A D - - G M A K I T I N R P H K R N A F R P K T V F E L Y D A F C D A R E D T S I G V V L F T G A G P H T D G K
- - - V H N Y T D I L Y H K T T - - G I A K I T I N R P H K R N A F R P Q T I V E M S Q A F E D A R N D P T I G V V L L T G A G P H T D G K
- - - W G T Y T D I L V D R C S E - G I A R V A I N R P A K R N A F R P Q T V V E L C D A F S R I R D D R S I G V V L F T G V G P A A D G G
- - - W K S Y E D I L F H N S G D - G I A R I A I N R P E K R N A F R P K T V C E M L D A F N L V R N D E K I G V V L L T G A G P D K K G V
E G D N K E F V D I I Y E K A L D E G I A K I T I N R P E R R N A F R P Q T V K E L M R A F N D A R D D S S V G V I I L T G K G - - - - - T
- - - G K G F V D I L Y D K A V G E G I A K I T I N R P D R R N A F R P L T V K E L M R A F E D A R D D S S I G V I I L T G K G - - - - - T
M D I K Y E K - K E - G I A T I T I N R P H V L N A F R P Q T V K Q M L E A M I D A R W D D D I G V V I L T G E G - - - - - T
- - - - - - F E D I R Y E K S T D - G I A K I T I N R P Q V R N A F R P L T V K E M I Q A L A D A R Y D D N I G V I I L T G A G - - - - - D
- - - A G E Y S D I L Y H K T E E - G I A K I T I N R P E R R N A F R P Q T V D Q M I E A L Q D A R N D S Q I G V I I L T G A G - - - - - D
- - - K R T Y D E I L Y E T Y N - - G I A K I T I N R P E V H N A F T P K T V A E M I D A F A D A R D D Q N V G V I V L A G A G - - - - - D
M G E R V K L E L D G - - E I A V A T L N R P E K L N A L D T K T R M E L A E V I E G I - - E E V A R V L I I T G S G - - - - - -
E G D K Y . D I L Y . K E G I A K I T I N R P K R N A F R P Q T V E M . A F D A R . D I G V . . L T G A G P H D G
Sll1127MenB7002All2347Tll2458Synw0998Pmm0608At1g60550P0671D0118CymeCp092B2262CT1846Bsu3075AF0435
71 139
74 142
73 141
74 142
85 153
81 149
133 201
127 195
57 125
80 149
89 157
67 135
56 123
Y A F C S G G D Q S V R G E G G - Y I D D Q G T P R L N V L D L Q R L I R S M P K V V I A L V A G Y A I G G G H V L H L V C D L T I A A D N
Y A F C S G G D Q S V R G E G G - Y V D Q A G V P R L N V L D L Q K L I R S M P K V V I A L V A G Y A I G G G H V L H L L C D L T I A A D N
Y A F C S G G D Q S V R G Q A G - Y V D D A G I P R L N V L D L Q R L I R S M P K V V I A L V A G Y A I G G G H V L H L I C D L T I A A D N
Y A F C A G G D Q S I R G E A G - Y L D E Q G V A R L N V L D L Q R Q I R S L P K V V I A L V A G Y A I G G G H V L H L V C D L T I A A E N
F A F C S G G D Q S V R G D G G - Y V G D D G L P R L N V L D L Q R I I R S L P K V V I A L V A G Y A M G G G Q V L H L L C D L S L A A D N
F S F C S G G D Q S I R G Q N G - Y E D D E G T Q R L N V L E L Q R L I R T L P K V V I A L V P G F A I G G G Q V L Q L V C D L S I A S E N
K A F C S G G D Q A L R T Q D G - Y A D P N D V G R L N V L D L Q V Q I R R L P K P V I A M V A G Y A V G G G H I L H M V C D L T I A A D N
Q S F C S G G D Q A L R D A D G - Y V D F D S F G R L N V L D L Q V Q I R R L P K P V I A M V A G Y A V G G G H V L H M V C D L T I A A D N
R A F C T G G D Q K V R G T K G - Y K D E N G K Q S L N V L Q L Q R E I R T I P K P V I A K V A G Y A I G G G Q I L Q M L C D L T I A A D N
K A F C S G G D Q K V R G D Y G G Y K D D S G V H H L N V L D F Q R Q I R T C P K P V V A M V A G Y S I G G G H V L H M M C D L T I A A D N
L A F C S G G D Q K I R G N A G - Y A D E K G V N K L N V L D F Q R D I R T C P K P I I A M V A G Y A I G G G H V L H M L C D L T I A A E N
K A F C S G G D Q K V R G H G G - Y V G D D Q I P R L N V L D L Q R L I R V I P K P V V A M V S G Y A I G G G H V L H I V C D L T I A A D N
K A F A A G A D I N E L L Q R D A I K A F E A T K L G - - T D L F S R I E E L E I P V I A A V N G Y T L G G G C E L A M A C D I R I A S E K
A F C S G G D Q V R G . G Y . D . G . R L N V L D L Q R I R . . P K P V I A . V A G Y A I G G G H V L H . . C D L T I A A D N
Sll1127MenB7002All2347Tll2458Synw0998Pmm0608At1g60550P0671D0118CymeCp092B2262CT1846Bsu3075AF0435
140 209 143 212
142 211
143 212
154 223
150 219
202 271
196 265
126 195
150 219
158 227
136 205
124 193
A I F G Q T G P K V G S F D G G F G S S Y L A R I V G Q K K A R E I W Y L C R Q Y S A Q E A E R M G M V N T V V P V D R L E E E G I Q W A KA I F G Q T G P K V G S F D G G F G A S Y M A R I V G Q K K A R E I W F L C R Q Y D A Q Q A L Q M G L V N Q V V P I A Q L E A E G V Q W A R
A I F G Q T G P K V G S F D G G F G A S Y L A R I V G Q K K A R E I W F L C R Q Y D A Q Q A L E M G L V N C V V P I E Q L E A E G I K W A G
A I F G Q T G P K V G S F D A G F G A S Y L A R I V G Q K K A R E I W F L C R Q Y T A Q E A L A M G L V N A V V P V E A L E A E G I R W A Q
A V F G Q T G P K V G S F D G G F G A G Y L A R V V G Q R K A R E I W F L C R R Y G A D E A L R M G L V N A V V P L D Q L E A E G V R W A R
A I F G Q T G P R V G S F D A G F G S S Y L A R L V G Q R K A R E I W F L C R K Y N S K E A L E M G L V N A I T K I E E L E A E G V I W A R
A I F G Q T G P K V G S F D A G Y G S S I M S R L V G P K K A R E M W F M T R F Y T A S E A E K M G L I N T V V P L E D L E K E T V K W C R
A I F G Q T G P K V G S F D A G Y G T S I M S R L V G P K K A R E M W F L S R F Y T A D E A D R M G L V N V V V P L A D L E R E T V K W C R
A I F G Q A G P R V G S F D G G Y G A S Y M A R I V G Q K R A R E I W F L C R Q Y S A Q E A L Q M G W I N A V V P L E E L D Q H T K A W A K
A I F G Q T G P K V G S F D G G W G A S Y M A R I V G Q K K A R E I W F L C R Q Y D A K Q A L D M G L V N T V V P L A D L E K E T V R W C R
A R F G Q T G P R V G S F D G G W G A S Y M A R L V G Q K K A R E I W Y L C R Q Y N A Q E A L D M G L V N T V V P L E K L E E E T I Q W C R
A I F G Q T G P K V G S F D A G Y G S G Y L A R I V G H K K A R E I W Y L C R Q Y N A Q E A L D M G L V N T V V P L E Q L E E E T I K W C E
A K F G Q P E I N L A I I P G A G G T Q R L P R L V G L G M A K K L V L T G E I I D A Q T A L R I G L V E E V V E H E R L M E R A K E V A A
A I F G Q T G P K V G S F D G G . G A S Y L A R I V G Q K K A R E I W F L C R Q Y A Q E A L M G L V N V V P L E L E E . . . W A R
Sll1127MenB7002All2347Tll2458Synw0998Pmm0608At1g60550P0671D0118CymeCp092B2262CT1846Bsu3075AF0435
210 275
213 279
212 277
213 278
224 289
220 285
272 337
266 331
196 263
220 285
228 293
206 271
194 256
E I L S K S P L A I R C L K A A F N A D C D G - - Q A G L Q E L A G N A T L L Y Y M T E E G S E G K Q A F L E K R P P D F S Q Y P W L P
E V L S K S P I A I R C L K A A L N A D C D G - - Q A G L Q E L A G N A T M L Y Y M T E E G R E G K Q A F L E K R S P N F R Q Y P W L P N
E I L E K S P I A I R C L K A A F N A D C D G - - Q A G L Q E L A G N A T L L Y Y M T E E G A E G K Q A F L E K R P P N F R D F P W L P
E I L S K S P L A I R C L K A A F N A D C D G - - Q A G L Q E L A G N A T L L Y Y L T Q E S A E G K T A F L E K R P P N F Q Q F P W R P
E V L Q H S P T A I R C L K A A F N A E T D G - - L A G I Q E L A G N A T H L F Y R T D E A L E G R N A F L E K R P P D F S E T G W L P
E I L R N S P T A I R I L K A S F N A E C D G - - I A G I Q E L S G Y T T Q L F Y S T D E A K E G R D A F L E K R P P D F S D Y G W T P
E I L R N S P T A I R V L K A A L N A V D D G - - H A G L Q G L G G D A T L L F Y G T E E A T E G R T A Y M H R R P P D F S K F H R R P
K I L R N S P T A I R V L K S A L N A A D D G - - H A G L Q E L G G N A T L I F Y G T E E A K E G K N A Y M E R R R P D F S K F P R K P
Q I L E H S P T A L S M L K A A L N A D C D G - - Q A G L Q E L A G L A T H L F Y G T E E A Q E G L N A F L E K R N P K F R R R K A L D E N
E M L Q N S P M A L R C L K A A L N A D C D G - - Q A G L Q E L A G N A T M L F Y M T E E G Q E G R N A F N Q K R Q P D F S K F K R N P
E I L A N S P L A I R C L K A A L N A D C D G - - Q A G L Q E L A G N A T L L Y Y M S E E G Q E G R N A F V E K R K P D F S K F P K R P
E M L E K S P T A L R F L K A A F N A D T D G - - L A G I Q Q F A G D A T L L Y Y T T D E A K E G R D S F K E K R K P D F G Q F P R F P
K I I E K S P L A V K V A K K A L N A S I N M P L K E G L R Y E A S L F A L L F S S - E D A K E G M R A F L E K R K P E F R G R
E I L . S P A I R C L K A A L N A D C D G P L Q A G L Q E L A G N A T L L F Y T E E A E G . . A F L E K R P D F S F P P N
The multiple alignment was generated by BLOSUM30 using Clustal W. The locus tags used to indicate organisms are summarized in Table 2.1, except for the following: Cymecp092 in Cyanidiochyzon merolae and P0671D01.18 in Oryza sativa. AF0435, a hypothetical protein in the archaeon Archeoglobus fulgidis DSM4304, was used as an out-group in phylogenetic construction as shown in Fig 2.3.

294
A.1.3. An amino acid sequence alignment of MenD homologs in cyanobacteria and other
organisms
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
1 0
1 0
1 0
1 0
1 0
1 80
1 0
1 0
1 0
M R S S F L V S N P P F L P S L I P R Y S S R K S I R R S R E R F S F P E S L R V S L L H G I R R N I E V A Q G V Q F D G P I M D R D V N L D D D L V V Q V C V
M R S S F L V S N P P F L P S L I P R Y S S R K S I R R S R E R F S F P E S L R V S L L H G I R R N I E V A Q G V Q F D G P I M D R D V N L D D D L V V Q V C V
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
1 0
1 0
1 0
1 0
1 0
81 160
1 0
1 0
1 0
T R T L P P A L T L E L G L E S L K E A I D E L K T N P P K S S S G V L R F Q V A V P P R A K A L F W F C S Q P T T S D V F P V F F L S K D T V E P S Y K S L Y
T R T L P P A L T L E L G L E S L K E A I D E L K T N P P K S S S G V L R F Q V A V P P R A K A L F W F C S Q P T T S D V F P V F F L S K D T V E P S Y K S L Y
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
1 0
1 0
1 0
1 0
1 0
161 240
1 0
1 0
1 0
V K E P H G V F G I G N A F A F V H S S S V D S N G H S M I K T F L S D E S A M V T A Y G F P D I E F N K Y S T V N S K D G S S Y F F V P Q I E L D E H E E V S
V K E P H G V F G I G N A F A F V H S S S V D S N G H S M I K T F L S D E S A M V T A Y G F P D I E F N K Y S T V N S K D G S S Y F F V P Q I E L D E H E E V S
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
1 0
1 0
1 0
1 0
1 0
241 320
1 0
1 0
1 0
I L A V T L A W N E S L S Y T V E Q T I S S Y E K S I F Q V S S H F C P N V E D H W F K H L K S S L A K L S V E E I H P L E M E H M G F F T F S G R D Q A D V K
I L A V T L A W N E S L S Y T V E Q T I S S Y E K S I F Q V S S H F C P N V E D H W F K H L K S S L A K L S V E E I H P L E M E H M G F F T F S G R D Q A D V K
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
1 65
1 71
1 65
1 45
1 69
321 397
1 64
1 64
1 64
M V D F T N P N T L A A S V L V E T L F R L G L Q Q A V I C P G S R S S P L T V A L A R H G - - - G I D C V V S L D E R S A S F F A L G
M S N L M P I A Y K N I N Q L W A Y I F T E T L K R L G L A Y A V I C P G S R S T P L A V A F A Q Q A P - - N I E A I S I L D E R S A A F F A L G
M I L T E N L N V L W A S V L F E T L Y R L G L R T V V L S P G S R S G P L A I A A A A H P - - - H L E A L P I L D E R S A A F F A L G
M T R L V L C P G S R S G P L A A A A G L L A A R G D L A L T T A I D E R S A A F L A L G
M T S H I E C Q N F F R S L Q L L D L F T K I G V K N L I L C P G S R S G P L A I A A G E L N K R R V L N V F N S I D E R S A G F H S L G
E L L N R E T E V S N F L R D E A N I N A V W A S A I I E E C T R L G L T Y F C V A P G S R S S H L A I A A A N H P - - - L T T C L A C F D E R S L A F H A I G
M L L F C S F S L I D W S E L I I K L L A H Y G F Q Y K F I A P G A R S S L L A R A A L L H G - - - - - N C V V H F D E R S L A F A A L G
M S V S A F N R R W A A V I L E A L T R H G V R H I C I A P G S R S T P L T L A A A E N S - - - A F I H H T H F D E R G L G H L A L G
M N P S T T Q A R V V V D E L I R G G V R D V V L C P G S R N A P L A F A L Q D A D H A G R I R L H V R I D E R T A G Y L A I G
E L L N R E T M N N W A . . . E L R . G . . V . C P G S R S . P L A . A A A R G . . . . D E R S A A F A L G
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
66 135
72 141
66 135
46 115
70 139
398 467
65 144
65 134
65 134
H G K R - - - - - - - - - - T G Q P V A L V C T S G T A A A N F L P A I I E A H Y S Q V P L L V L T G D R P P R L R H C R A G Q T I D Q T K L Y G H Y P Q W Q T
I A K A - - - - - - - - - - T N R P V A I V C T S G T A G A N F Y P A V I E A Q E S R V P L L L L T A D R P P E L R D C H S G Q T I D Q M K L Y G S Y P N W Q A
L V Q Q - - - - - - - - - - Q G R P V A L L C T S G T A A A N F Y P A I I E A S L S H L P L I V L S A D R P P E L R F C Q A G Q A I D Q V H L Y G H A V R H Y R
M A T A - - - - - - - - - - G G R A V A V V T T S G T A V A N L L P A A V E A D R S T Q P L L L L T A D R P A R L K N C G A N Q T V N Q E Q F L Q P V C R W L G
I S T A - - - - - - - - - - S G D V S L V V T T S G T A V G N L L P A A I E A D K S C K S I V F I T A D R P L R L K N C G S N Q T V N Q E E F L N S V C R S N L
Y A K G - - - - - - - - - - S L K P A V I I T S S G T A V S N L L P A V V E A S E D F L P L L L L T A D R P P E L Q G V G A N Q A I N Q I N H F G S F V R F F F
V G K A S Q I Q T S K T N A S K T K A I L I V T S G T A V G N L W P A V M E A S L S Q T S L I L L T A D R P Y E L R D T S A N Q T L D Q V K L F N G Y V R W Q F
L A K V - - - - - - - - - - S K Q P V A V I V T S G T A V A N L Y P A L I E A G L T G E K L I L L T A D R P P E L I D C G A N Q A I R Q P G M F A S H P T H S I
L A V A - - - - - - - - - - A G A P V C V A M T S G T A V A N L G P A V V E A N Y A R V P L I V L S A N R P Y E L L G T G A N Q T M E Q L G Y F G T Q V R A A I
. A K A S Q I Q T S K T N A . G P V A . . T S G T A V A N L . P A . I E A S . P L . L L T A D R P P E L . C G A N Q T I . Q . G . R . .
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
136 206
142 211
136 204
116 186
140 209
468 539
145 205
135 203
135 210
E L A L P E P N M D Y C H Y L R Q T A L H S W Q K C F W P G L - - - - - - - G V V H L N C P F D E P L V P L E H A R - - E S L Q H L A E E F S K E H F Y R G L T
E L A L P V S D M G M L A Y L R Q T L V H S W Y R M Q A P T P - - - - - - - G P V H L N I P F R D P L A P I P D G - - - T D L S Y L L A K F H P E E F F A G I T
E L S L P - - E L P L L P Y L R Q T L C H S W Q T A L W P D P - - - - - - - G P V H L N I P L R D P L D L R S Q A N - - F H G E L P K G F F D Q V Q P F V P P R
H G A P E G L A S Q P P Q A L F D L A H E A W R H C H G R P P - - - - - - - G P V Q L N L P F E E P L H G A P E D Q - - L S L Q V S G L Q R S A P A P S S D S P
S T N L N G I H E N N D D D I L K I V E T V K K Q I L Q - S P - - - - - - - G P I H L N I A F E K P L D I S L K N K - - K K N F E V F E R F Y L N K T Y K F L K
N L P P P - T D L I P V R M V L T T V D S A L H W A T G S A C - - - - - - - G P V H L N C P F R D P L D G S P T N W S S N C L N G L D M W M S N A E P F T K Y F
D V P P P - H H E L D A D W L A S T L A Y A A F K A S D - - - - - - - - - - G P V H L N L M F R E P F A S Y P I P - - - - - - - - M P K K L V E Y V P A S T A T
S L P R P - T Q D I P A R W L V S T I D H A L G T L H A - - - - - - - - - - G G V H I N C P F A E P L Y G E M D D T G L S W Q Q R L G D W W Q D D K P W L R E A
S L G L A E D A P E R L D S L N A T W R S A T C R V L V A A T G S R T A N A G P V Q F D I P L R E P L I P D P E P - - - - H G A I T P Q G R P G G K P W T Y I P
L L P . . L T . A . . . P G S R T A N A G P V H L N . P F R E P L . P . . . P . .
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
207 279
212 274
205 259
187 240
210 274
540 609
206 253
204 257
211 254
T F T T M N R G E A I S L P V N F S I F S F P S P Q L D F S P L G L I L V G V M P G G E T P S - - L L T D I L A I A R G L G Y P V L C D A L C S L R N - - - - -
D T T P L P H H S P L S I P P - - - - - - - - - - E W L Q S Q R G I I I A G V A Q P Q Q P Q E - - Y C R A I A R L S Q T L Q W P V L A E G L S P V R N - - - - -
V V T S L P W Q T - - - - - - - - - - - - - - - - - W Q Q M Q R G L I I A G P S H G V D P L A - - E A A A I D R L S R F L Q W P V L A D A L S S A R - - - - - -
- L G A A P Q L D - - - - - - - - - - - - - - - - - P N Q - - A G V V V A G P W R G L A P A L P A Y Q Q A L L Q W L A R S G W P L L A D P L A A I P - - - - - -
N D N Q N K N I Q F S E K F F K R - - - - - - - - - I N L S N P G I I I V G P Y Q G S T K D L F S F N S A L K K L Q E I T G W P V F V D P V S G V S - - - - - -
Q V Q S H K S D G V T T G Q I T - - - - - E I L Q V I K E A K K G L L L I G A I H T E D E I W - - - - - A S L L L A K E L M W P V V A D V L S G V R L R K L F K
G F E W S L D W Q - - - - - - - - - - - - - - - - - - Q K - - - G L F I V A Q A M D D E S F H - - - - - L L L Q L A Q Q W S W P I L V E V F C H A R - - - - - -
P R L E S E K Q R D - - - - - - - - - - - - - - W F F W R Q K R G V V V A G R M S A E E G K - - - - - - K V A L W A Q T L G W P L I G D V L S Q T G - - - - - -
Q V T F D Q P L E - - - - - - - - - - - - - - - - - I D L S A D T V V I A G H G A G V H P N - - - - - - - - - - - - L T - E L P I V A E P T A P F G - - - - - -
S I F S S G . . I A G G . A . . L . L W P V L A D . L S . R N R K L F K

295 Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
280 355
275 350
260 332
241 312
275 347
610 688
254 326
258 329
255 318
- - Y D D G Q T A L I T N Y D F L I R C P R W A E Q L V - P E Q I I Q I G E L P T S K A L R H W L G - T I D C P R Y I L N F H G E N L D P L Q G Q T I Y L S A S
- - Y A D F N P Y L I S T Y D L I L R N Q Q L A T R L A - P D M V I Q I G D M P T S K E L R N W I D - T H Q P R R W V I D P S D Q N L D P L H G R T T H L R I R
- - - - - G L P H G I S H Y D L L L R D A H L R E Y L R - P E A V I Q L G P L P T S K A L R E W L S - A C D P L I W C L D P T G D N N N P L H G R C Q M L A I A
- - - - A D C P G Q L D G W E L Q L D R L Q L A P - - - - G S P V L R L G P L P A S R R L E A W L Q R Q T G P Q L L I S E G E P R G L D P L G L A D Q W S G G L
- - - - A E L K G L V E N W E L I L K - - K N K N I I Q - C D Q I L R F G P L S S S N Y L E D F L L N F E G L Q I L V K E N N I R K L D P I K K S L E Y D F G I
P F V E K L T H V F V D H L D H A L F S D S V R N - L I E F D V V I Q V G S R I T S K R V S Q M L E K C F P F A Y I L V D K H P C R H D P S H L V T H R V Q S N
- - - - - Y Q Y K H S C V I A Y H E T I C H K T M P F P D A N I I I Q F G E R L I S N H L I T I M K - - K V N Q Y V V V S S S T V R V D P L K A V T T R I C V T
- - - - - - - - Q P L P C A D L W L G N A K A T S E L Q Q A Q I V V Q L G S S L T G K R L L Q W Q A S C E P E E Y W I V D D I E G R L D P A H H R G R R L I A N
- - - - - - - - - P Y R P Y P L H P L A L P L L H - - - - P K Q V I M L G R P T L H R P V S A L L A - - - D P Q V P V Y A L T T G P C W P D V S G N S Q A A G T
P F Y . . D L . L . . L . V I Q . G T S K L W L . . . L D P L . . . .
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
356 430
351 425
333 404
313 383
348 427
689 764
327 401
330 398
319 386
V A Q V A E Y I H N - - Q G F I P D A E Q K N Y A H - - - S W L E K Q R Q S Q T I I I S A L A D A H T P L M V A Q L A H C L P P Q T N L F V A N S L P V R W L E
V E E L G Y Q G V E - - E D K S S V S E Y L Q L W C - - - N A E T K V R V N V D E T L D K M E D L V E C K A A W L L S Q M L P P E T P L F I A N S M P V R D V E
P Q A V D C P - - - - - P D P L P P N P Y L K D W Q - - - D Q D Q R V R E Q L K R T F E A I D W F S E V K L I Y H L P Q W L P S Q T A I F V A S S M P V R D V E
A A W W A E Q N Q D - - F P G A S T P E Q L M P Q S - - - G I A S L L R - - - - Q R L P L Q G A V N E P A L A H W L P L L L P P Q L P V M L A A S S P V R D W L
S N F V N Q L L G A F L N H K K K S K P L I N L G Q V L I K E G A K I K E I L K E Q L F S N T E I T E Y K L A N F V P K I W P E N Y P I M L S A S S P I R D W L
I V Q F A N C V L K S R F P W R R S K L H G H L Q A - - - - L D G A I A R E M S F Q I S A E S S L T E P Y V A H M L S K A L T S K S A L F I G N S M P I R D V D
N V Q S L L K F W F K T L R P T T K P Q W L S W W Q - - - H L N F C V G Y K L R Q Y F A Q E I K L S E P G V L R M C F Q Y A K P - - L I F V G N S M P I R D A N
I A D W L E L H P A E - - - - K R Q P W C V E I P R - - - - - - - L A E Q A M Q A V I A R R D A F G E A Q L A H R I C D Y L P E Q G Q L F V G N S L V V R L I D
R A V T T G T - - - - - - - - - P H P A W L Q R C A - - - E M N Q H A I S T V R S Q L A A H P L I T G L H V A A A V A D A L R P G D Q L V L G A S N P V R D M A
. . . L V L I . . . . . . . . E . A L L P P . L F . . N S P V R D . .
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
431 481
426 475
405 454
384 433
428 477
765 831
402 454
399 448
387 448
F F W P A N G D H - - - - - - - - - - - - - - - - - H R I F V N R G A N G I D G T L S T A M G I A H R S R G - - - - - - - - - - - - E T V L L T G D L S L L H D
F F W K P N N L R - - - - - - - - - - - - - - - - - V R S H F N R G A N G I D G T L S T A L G I S H R Q Q - - - - - - - - - - - - - S S V L I T G D L A L L H D
S V W Q G S D R R - - - - - - - - - - - - - - - - - H R F Y F N R G A N G I D G T L S T A L G V A H R G Q - - - - - - - - - - - - - P T L L I T G D L A C L H D
I W G G L Q A Q N - - - - - - - - - - - - - - - - - R R C F S F R G A S G I D G T L S L A M G L A M E Q G - - - - - - - - - - - - - P M V L V T G D L A L L H D
T F S E N A T L T - - - - - - - - - - - - - - - - - R E C F S F R G A S G I D G T L S L A L G I A R I T K - - - - - - - - - - - - - P L L L V T G D L A L I H D
M Y G C S S E N S S H V V D M M L S A E L P C Q W - I Q V T G N R G A S G I D G L L S S A T G F A V G C K K - - - - - - - - - - - - R V V C V V G D I S F L H D
H F V E C H H L H D P - - - - - - - - - - - - - - - V H V F A N R G V S G I D G N L A T S M G V S Y A L Q E - - - - - - - - - - - - P L M C I L G D L A F L H D
A L S Q L P A G Y - - - - - - - - - - - - - - - - - - P V Y S N R G A S G I D G L L S T A A G V Q R A S G K - - - - - - - - - - - - P T L A I V G D L S A L Y D
L V G L S T D G I - - - - - - - - - - - - - - - - - - Q V R S N R G V A G I D G T V S T A I G A A L A Y E R S P G Y A G S T D R P A R T V A L I G D L A F V H D
. . V V D M M L S A E L P C Q W . . . N R G A S G I D G T L S T A . G . A . . S P G Y A G S T D R P A P . V L . T G D L A . L H D
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
482 557
476 551
455 530
434 507
478 554
832 894
455 523
449 522
449 528
S N G F L N Q S Q - - M R G N L T I I L L N N N G G G I F Q T L P I A Q C E D V - - F E T Y F A T P Q G V D F G Q L C R T Y G V E H K I I T N L W D L K E Q W P
T N G F L I R N K - - F V G H L T I I L I N N N G G G I F E M L P I A K F E P P - - F E E F F G T P Q D I D F A Q L C T T Y N V Q H E L I H S W V H L Q Q R L N
T N G W L I T P Q - - F Q G C L T V L L I N N R G G G I F E H L P I R Q F D P P - - F E A F F A T P Q Q V D F S Y L A A A Y G I P Y H C L K D W A D V E A Q L R
S N G W L H G A Q - - G N P P L L V L L I D N Q G G G I F Q Q L P I Q S K Q - - - - F D R L F A M P Q R V N P L A L A A A H G I D G R Q V A C L E D L P E A L E
I N G F L I E N A - - I E L N L T I L L I N N N G G N I F N N L Y K N N L D E E - E L K K L F I M P K S I N W E N L A K G Y Q V P I K N V S D L N K L R E A F E
T N G L A I L K Q R I A R K P M T I L V I N N R G G G I F R L L P I A K K T E P S V L N Q Y F Y T A H D I S I E N L C L A H G
L T S L H L L R Q - - L T K H I W V I V I N N Q G G G I F E L I P N V F N H - - - - - - - - Y F H H H D H - F E H V A K Q F D F H Y Y A V N N W Y E L A L H L Q
L N A L A L L R Q - - V S A P L V L I V V N N N G G Q I F S L L P T P Q S E - - - - R E R F Y L M P Q N V H F E H A A A M F E L K Y H R P Q N W Q E L E T A F A
S S G L L I G P T E P T P Q Q L K I V V S N D N G G G I F E L L E Q G D P R L S A V S S R I F G T P H D V D V G A L C R A Y H V E G R Q I E V D K L R E A L D E
. N G . L I . Q . . L T I . L I N N N G G G I F L P I . . V . . F . T P Q V F L A . Y . . . . L
Sll0603Alr0312Tll0130Synw0997Pmm0607At1g68890CymeCp094B2264ML2270
558 595
552 587
531 562
508 545
555 588
895 894
524 561
523 556
529 556
S N N S - S P I R V L E I I G D R H Q E A Q W L K S L Q A Q F C C A D S L I Q
P L P N - T G I R V L E L R T N R K I D A Q W R R D N L S N F A A D N I I
Q T P W - P K I R L L E F R S D R H R N A Q W R Q Q V L A H L G G
W G L A Q G R P A L L R L A T D R E A D A R L R T Q L R S A A Q N A E P L L
W S L S M Q K S V I I K V D I N V E N E M K G R N L I L K K I L T S
N T K G - - - A N L I E I Q T N Q L E N A T L H K K L T Q Y T N L F S V S F H S D
D A W R T P T T T V I E M V V N D T D G A Q T L Q Q L L A Q V S H L
P G V G - - - M R V L E V K A D R S S L R Q L H A A I T A T L
. L E . R A Q . . . S D
The multiple alignment was generated by BLOSUM30 using Clustal W. The locus tags used to indicate organisms are summarized in Table 2.1, except for the following: Cymecp094 in Cyanidiochyzon merolae. ML2270, a MenD homolog in Mycobacterium leprae, was used as an out-group in phylogenetic construction as shown in Fig 2.3.

296
A.1.4. An amino acid sequence alignment of UbiA homologs in cyanobacteria and other
organisms
S6803S7002A7120NpuncTeryTelonS8102PmarMED4PmarMIT9313PmarSS122GvioRpalRrubEcolCT1511
10 20 30 40 50 60 70
M V A Q T P S S P P L W L T I I Y L L R W H K - P A G R L I L M I P A L W A V C L A A Q G L P P L - - - - P L L G T I A L G T L A T S G LM V S E P T W K K I I Y L L R W D K - P A G R L I L I I P A L W A V F L A A E A K P D W - - - - K L V L V I I L G A I T T S A A
M L E T H Q - E P I W L T I F R L L R W H K - P E G R L I L M I P A L W A V F L A A A G K P P L - - - - P L V G V I V L G T L A T S A AL R M P E R P Q - P P V W L V I I R L L R W H K - P E G R L I L M I P A L W A V F L A A S G K P P L - - - - P L V G V I I L G T L A T S A A
M T Q A A S P E S T L M K V I R L L R W D K - P A G R F I L M I P A L W A V F L A T H A M P P I - - - - P L V G V I V V G T L A T S A AN S Q V S R T S V S T W V A I A Q L L R W H K - P A G R L I L L I P A L W S L T L A S E G L P S L - - - - K L L L I I T M G A I A T S A A
M D S I R E R L G P W L E L L R W T K - P T G R L I L L I P A G W S L W L S P S A P P G L - - - - D L L L Q I V V G G L A V S G AM I K K D Q N S K I N I L F K L L R W N K - P T G R L I L L I P A G W S L Y L T P E S N P S I - - - - Y M L L K I L I G G L L V S G LM T S R S F S N G L R Q W L E L L R W H K - P S G R L I L L V P A G W S L W L T P D A P P L A - - - - G L V S L I V V G G L M V S G AM K S F L L T K K L L N I F H L L R W N K - P T G R L I L L I P A G W S L W L T P N A P P S K - - - - E L V T L I I A G A I S V S G A
M S T A P T D W R A K A D A V F R L L R W D K - P A G R L I L M I P A L W G V F A A A G G S P S P - - - - L L V L V M V V G T I A T S A AN W V D N H A P L W T R P Y L R L A R F D R - P I G S W L L L M P C L W S A A L A A G M A H D L S R L P L F C A L F F I G S F V M R G A
D G W V D R M A P G V I R P Y L K L M R L D R - P I G T W L L L F P C W W S Q S M A A Q G W P D L - - - - W L A A L F A A G A L V M R G AM E W S L T Q N K L L A F H R L M R T D K - P I G A L L L L W P T L W A L W V A T P G V P Q L - - - - W I L A V F V A G V W L M R A A
M S V S S S S Q L S A F Q A W M L A I R P K T L P A G A M P V V I G A A L A A A S G - V F K P L P - - - - - - A L V A L I C A L G I Q I AL . . . . L L R W K L P . G R L I L L I P A L W . . . L A . P L S R L P L . . . I . . G . L . S . A
S6803S7002A7120NpuncTeryTelonS8102PmarMED4PmarMIT9313PmarSS122GvioRpalRrubEcolCT1511
80 90 100 110 120 130 140
G C V V N D L W D - R D I D P Q V E R T K - Q R P L A A R A L S V Q V G I G V A L V A L L C A A G L A F Y L T - - - - P L S F W L C V A A VG C V V N D L W D - R D I D P Q V E R T K - V R P L A S K A L T V K V G I I T A I A F F L C A A G L A F Y F N P W T N P L S F W L C V A A VG C V V N D L W D - R D I D P E V E R T R - D R P L A A R T L S I K V G I A V G I I A L F C A A I L A Y Y L N - - - - S L S F W L S V A A VG C V V N D L W D - R D I D P E V E R T R - D R P L A S R A L S V K V G I V V A I V S L A C A A I L A F Y L N - - - - P L S F W L C V A A VG C V V N D L W D - R D I D S E V E R T R - D R P L T S R A L T I Q T G I I I V I I A F A C A G V I A L Y L N - - - - P L S F W L S V A A VG C I V N D C W D - R N I D P H V Q R T Q - N R P L A S R R L S L G V A L G L M V I A L L C A W G L T F Y L T - - - - P L G Y W L A V A A VG C I A N D L W D - R R F D G R V E R T K - Q R P L A R G A I R P T S A L V L L I V L L S L S L A V V L S L D E A S R Q L C L L L S I G A LG C V A N D I W D - K R I D Q K V L R T K - N R P L A A N K I S T K T A Y L I L I F L I I C S F F L T L S L P E N G R L L S L S L A F F A LG C I A N D L W D - R R I D R Q V E R T R - E R P L A K G S I R I C T A V G L L I V L L L L S L L V V A S L P T P S Q R L C L A L A I L A LG C I A N D L W D - R K I D S K V K R T K - N R P L A N G S L S I V E A F S L L I F L L I I S L K V I L L L P T P S Q N L S L K L A A M A LG C V I N D L W D - R D I D P L V E R T R - A R P L A S R E L A V P V A V V T F A V S L L C A W G L T F F L N - - - - P F T F W L C V A A VG C T W N D I T D - R D L D D K V E R T R - S R P I P A G Q V T A K Q A A V F M V L L C L I G L V V L L Q F N - - - - S F A I A T G I S S LG C T F N D I V D - R D F D A Q V A R T A - A R P I P S G A V S R K K A V V F L G A Q L L V G L A V L L S L N - - - - G F S I W L G V A S LG C V V N D Y A D - R K F D G H V K R T A - N R P L P S G A V T E K E A R A L F V V L V L I S F L L V L T L N - - - - T M T I L L S I A A LT N F I N E I Y D F R K G A D T A E R L G P T R T V A A G I I T E Q T M I R V S I V L G V S V F V L G L Y L V A I G G W P I L L I G V L S LG C V . N D L W D F R D I D V E R T . P R P L A . L . . A . . . . I . . L L C . . . L . L . L N L S . W L V A A L
S6803S7002A7120NpuncTeryTelonS8102PmarMED4PmarMIT9313PmarSS122GvioRpalRrubEcolCT1511
150 160 170 180 190 200 210
P V I V A Y P G A K R V F P V P Q L V L S I A W G F A V L I S W S A V T G D L T - - D A T W V L W G A T V F W T L G F D T V Y A - - M A D RP V I I A Y P L A K R F F P V P Q L V L A I A W G F A V L I S W S A V T Q N L T - - A A T W L L W G A T V F W T L G F D T I Y A - - L S D RP V I L L Y P G A K R V F P V P Q L V L S I A W G F A V L I S W S A V T Q N I S - - Q A T W L L W G A T L L W T L G F D T V Y A - - M S D RP V I V L Y P G A K R V F P V P Q L V L S I A W G F G V L I S W S A V T Q T I T - - Q P T W L L W G A T V L W T L G F D T V Y A - - M S D KP V I I C Y P L A K R F F P V P Q L V L S I A W G F A V L I S W S A A V A K L D - - S A T W I L W G A T I V W T L G F D T V Y A - - M S D RP V I L L Y P L A K R V L P I P Q L V L A V A W G F A V L I P W A A V Q G R L T - - L T T A G L W A A V V M W T L A F D T V Y A - - M P D RP A I L L Y P S A K R W F A Y P Q A V L A F C W G F A V L I P W A A A E R S L T L Q P A L I G C W L A T L L W T F G F D T V Y A - - M A D RP L I L I Y P S A K R W F K Y P Q F I L S I C W G F A V V I P W A A N E G N I N - S I V L L F C W L A T I F W T F G F D T V Y A - - L A D KP P I L L Y P S A K R W F A Y P Q A V L A L C W G F A V L I P W A A S Q A N L K G G W P L L G C W L A T L M W T F G F D T V Y A - - M A D RV P I L I Y P S S K R W F R Y P Q V L L A F C W G F A V L I P W A A S E H S L N G G F P L L T C W L A T M V W T F G F D T V Y A - - M A D KP V I A L Y P G A K R V F P V P Q L V L S L A W G F A A L I S W S A V T G G L E - - T P A W W L W G A T V L W T L G F D T V Y A - - L S D RI I V A A Y P F M K R I T Y W P Q F V L G L A F S W G A L M G F A G T F E R L D - - L V A F A L Y A G S I A W V I G Y D T V Y A - - H Q D AI L V F G Y P F M K R I T Y W P Q A W L G L T F T Y G A L M G W A A V R G S L D - - P A P L L L Y A A C F F W T L H Y D T I Y A - - H Q D KA L A W V Y P F M K R Y T H L P Q V V L G A A F G W S I P M A F A A V S E S V P - - L S C W L M F L A N I L W A V A Y D T Q Y A - - M V D RL F A W A Y T G G P F P I A Y S G L G D V F V F I F F G L V A V G G T Y Y V Q A L S L P M E V L V A A A A P G A F S V C I L L V N N I R D IP . I . . Y P . A K R F . P Q L V L . . A W G F A V L I W A A V L . . . L W . A T . . W T L G F D T V Y A N N M D R
S6803S7002A7120NpuncTeryTelonS8102PmarMED4PmarMIT9313PmarSS122GvioRpalRrubEcolCT1511
220 230 240 250 260 270 280
E D D R R I G V N S S A L F F G Q Y V G E A V G I F F A L T I G C L F Y L G M I L M L N P L Y W - L S L A I A I V G W V I Q Y I Q L S A P TD D D L K I G I N S S A I F F G K F A P E A V G I F F V L T A T C L A I L G K T L N I N L L I Y G I A W A I A V G L W F K Q Y L K I R K P NE D D Q R I G V N S S A L F F G D Y A P L A I G I F F L G T I I C L T W L G V A I H L H L A F W - I T L G I A S V G W I W Q V L R L R Q P QE D D R R I G V N S S A L F F G N Y A P V A I G I F F A G T I L L L A W L G L L I N L H F A F W - I S L A V A T I G W V W Q S L R L R Q R DE D D Q R L G I N S S A L F F G K Y A N N A V G I C F F L T T I F L G L L G I K M E L H T G F W - I T L G I A T V F W F I Q Y K K L G Q E DP D D R R L G V N S S A L F F G A Y A P L A I A L F Y A L T V V F L M M V G V R A G L T W R F W - L A L A V T T Y F W L R Q S L Q I Y T H ER D D A L I G L N S S A L S L G N R A V V T V R A C Y V L T V V A L G V A A A S A G V H P L F W I F W L G A S L L M Q I S C Q S L N - H R NK Y D I Q I G V N S S A V N L A Y N T K K T I Q I C Y F L T S S F L A I C A F I N Q L N F V F W P I W L I T G F L M Q K D I L K I F P E S KR D D A N L A L K S S A L S L G S H A L K T V A F S Y A L A C T F L A S S A V S A G V G W A F W P F W L I A S I G M Q K E T W T L R - G S ND D D K K L G L Q S S A L S L G Q K A L K S V F I C Y A L T S T L I A F S A L T K D I G I I F W P F W L I A S I G M Q K E V F S L K - E L EE D D L K I G I N S S A I F F G R F T P E A I A A F F A G A A L C L A V V G Y Q L E L G S I F Y - V C L I V A T L F W W R E Y R S L V T G DE D D L M V G I K S T A L L F G E N T K L A L V L L F G V A V S L I G V A L K L A D A G W F A W - I G L A A F A A H L V Q Q I V R V D I H DE D D L L V G V K S S A L A L G S N T R P A L V V F S T L M M A L V A A A G W Q A G L S W P F W - A L L G L P A A H L A W Q I W R V D I H DD D D V K I G I K S T A I L F G Q Y D K L I I G I L Q I G V L A L M A I I G E L N G L G W G Y Y W S I L V A G A L F V Y Q Q K L I A N R E RD T D R K V G K M T L P A R I G - - A P A A R A L Y V A L V V L A Y L V P F Y M I S T G Y S L W C L L S L L S I P L A I G M V R T L Y A S E. D D . . I G . N S S A L F G . A A . . I . . . L T . . L A . . G . L . . F W . L . . . . . . Q . .
S6803S7002A7120NpuncTeryTelonS8102PmarMED4PmarMIT9313PmarSS122GvioRpalRrubEcolCT1511
290 300 310 320 330 340 350
P - - - - - - - - E P K L Y G Q I F G Q N V I I G F V L L A G M L L G W LL - - - - - - - - P R K A Y N R L F E E N V T I G F I L L S G M I S S F I FL - - - - - - - - P N P A Y G E M F R Q N V W I G F I V L A G M I F G S LL - - - - - - - - P N S V Y G K M F R Q N V W I G F I L L A G M I A G S F- - - - - - - - - I L K P Y A Q I F R Q N V W I G F I L L A G M I T G L I FR R D T Q T T T L P I Q T Y G R Y F N E N V W L G F L L W V G M W C P R PA - - - - - - - - T M A S F G L H F R R Q V Q L G S L L L L G L I V S R G L S GQ - - - - - - - - S I K K I G D H F K N Q S I Y G G L I L L G F I I A SQ - - - - - - - - P I T T Y G Q H F Q H Q V L L G A M L L L G L I L G R I SA - - - - - - - - N T N K F G K H F R N Q V F L G G L I L F G L I L G N F- - - - - - - - - D P A R F N R I F N E N V T V G F V I L T G L I G G T L IS - - - - - - - - - - P L C L K L F K S N R D A G L L L F G G L V A D A V S R S M AS - - - - - - - - - - P V C L M I F K S N R F V G W L L L A A I V A G R A P G G VE - - - - - - - - - - - A C F K A F M N N N Y V G L V L F L G L A M S Y W H FG Q - - - - - - A L N A V L A G T G K V L T V H G L L F S L G L V I P N I I S I F R P
D T Q T T T . . G F . N V . G . . L L . G . I . G . P
The multiple alignment was generated by BLOSUM30 using Clustal W. S6803, Synechocystis sp. PCC 6803; S7002, Synechococcus sp. PCC 7002; A7120, Nostoc sp. PCC 7120; Npun, Nostoc punctiforme; Tery, Trichodesmium erythraeum; Telon, Thermosynechococcus elongatus BP-1; S8120, Synechococcus sp. WH8102; PmarMED4, Prochlorococcus marinus MED4; PmarMIT9313, P. marinus MIT9313; PmarSS122, P. marinus SS120; Gvio, Gloeobacter violaceus PCC7421; Rpal, Rhodopseudomonas palustris; Rrub, Rhodospirillum rubrum; Ecol, E. coli; CT1511, Chlorobium tepidum.

297
A.1.5. An amino acid sequence alignment of UbiD homologs in cyanobacteria and other
organisms
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
10 20 30 40 50 60 70 80
M A R D L R G F I Q L L E T R G Q L R R I T A E V D P D L E V A E I S N R M L Q A G G P G L L F E N V K - - - - - G S P F P V A V N L M G T V EM A R D L R A F I K L V E E R G Q L R R I K A L V D S D L E I A E I A N R M L Q V G G P A L L F E N V R - - - - - G T S V P V V V N L L G T V EM A R D L R G F I K I L E E R G Q L Q R I S A L V D P D L E I A E I S N R M L Q K G G P G L L F E N V K - - - - - G A S F P V A V N L M G T V EM A R D L R G F I K I L E E K G Q L R R I S A L V D P E L E I A E I S N R M L Q K G G P G L L F E N V K - - - - - G A S F P V A V N L M G T V EM T K D L R H Y L Q L L E Q R Q Q L R R I T V P V D P D L E M A E I C N R L L A A G G P A L L F E N V I - - - - - G S P Y P V A I N L L G T L E
M A L F R S G P A T R D L R G F L Q L L D Q R G Q L K R I T A A V D P D L E L A A I A D R V L S Q G G P A L L F E N V I - - - - - G S S M P V A V N T L G T V EM P L L R P G P A S Q D M R D F L A L L E Q R G Q L R R I T A P V E P D L E L A A I T D R V L G L G G P A L L F E N V I - - - - - G S S M P V A V N L M G T L EM N V F Q N G V G S Q D M R D F L S L L E K K G Q L R R I S K P V D P D L E L A A I S D R V L S M G G P A L L F E N V I - - - - - G S S M P V A I N L L G T I E
M G R D L R T F L Q L L E S R G Q L R R I S A P V E R D L E V A E I A N R L L A A G G P A L L F E N V K - - - - - G S P F P V A V N L L G T V EM L S R V K P P F P D L R A F S G Y L E S R G Q L H R I R K P V S V V H D L T E I H R R V L H A G G P A L L I E N P I K A D G T P S E M P I L V N L F G T V E
M R S L P I F P D L R A F L D W C A R K G D L S R I A E P V S L R H E T T A V A S K I L R G G G P V L R F E S - - - L R D T S A T M P L V A N L F G T R EM D A M K Y N D L R D F L T L L E Q Q G E L K R I T L P V D P H L E I T E I A D R T L R A G G P A L L F E N - - - - P - K G Y S M P V L C N L F G T P K
M Y Q N L Q E C I N D L E K H G H L I R I R E E V D P Y L E M A A I H L K V Y E A G G P A L L F E N V K - - - - - G S N Y Q A V S N L F G T M EM L R P M . D L R . F L L L E R G Q L R R I . A P V D P D L E . A E I R . L . G G P A L L F E N V K T G S S P V A V N L G T V E
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
90 100 110 120 130 140 150 160
R I C W A M N M D H P L E L E D L G K K L A L L Q Q P K P P K K I S Q A I D F G K V L F D V L K A K P G - R N F F P P C Q E V V I D G E N L D L N Q I P L I R PR I C W A M K M E K P E E L E D L G R K L A M L Q Q P K P P K K I S Q A I D F G K V L F D V L K A K P G - G S F F P P C Q Q V V V E Q E E L D L G L I P M L R PR I C W A M N M Q H P Q E L E T L G K K L S M L Q Q P K P P K K I S Q A I D F G K V L F D V L K A K P G - R D F F P A C Q Q V V V Q G D D V D L N T L P L I R PR I C W A M N M Q H P E E L E T L G K K L S M L Q Q P K P P K K I S Q A I D F G K V L F D V V K A K P G - R D F F P A C Q Q V V L Q G N D L D L N K L P L I R PR V C W A M N M D D P L E L E T L G E K L G K L Q Q P K P P K T L S Q A L D F G K I L F D V V R A K P S - R D L L P P C Q Q V V I T A P D L D L R Q L P L I R PR V V W S M G L E R A E Q L E D L G S R L A L L Q Q P R P P K G L S E T K Q F A R V F W D L V K A K P D - R D L T P P C R Q Q I F K G D A V N L Y N I P L I R PR V V W S M G L D N A Q Q L E D L G T R L A L L Q Q P R P P K G L Q E T R Q F A S V F W D L I K A R P D - L D L T P P C H Q Q V L R G D A L N L D N L P L I R PR V V W S M G L S S Q D E L E G L G K K L S Y L Q Q P E P P K G I K K T I E F G G I L W D L L Q A R P D - L D L T P P C H Q R L L K G E D V N L D K L P L I R PR V V W S M G L E E Q Q Q L E A L G E K L G K L Q K P R P P R S L Q D A L E F A P L L F D V I K A R P G R V L F A P E C Q Q V V L E G K A V D L D R I P M I R PR V A W G L G - I L P E N L S R L G E A L A E M R E P A P P Q S L T D A L S K L P M A K A A L A M R P K - L A K S A P V Q E V V L T G D A V D L G R L P V Q I PR V A A G L G - L G L E Q I P E L G A F L A A L R A P A P V A G M R D A L S R W P Q L Q A A L N T R A K - I V R S A E A Q E V V H E G A A V D L G M I P V P T CR V A M G M G Q E D V S A L R E V G K L L A F L K E P E P P K G F R D L F D K L P Q F K Q V L N M P T K - R L R G A P C Q Q K I V S G D D V D L N R I P I M T CR S K F I F R - - - - - Q T W Q S A E N V V A L R N - D P M S A L K H P F A H A R T A L A A S K A L P L K K S R L P A G F E E I T I S D - - - - - - L P L I Q HR V . W . M G . L E L G K L A L Q Q P . P P K . . A . . F . . . L . D V L K A . P . P P C Q Q V V . G . V D L L P L I R P
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
170 180 190 200 210 220 230 240
Y P G D A G K I I T L G L V I T K D C - - - - - E T G T P N V G V Y R L Q L Q S - - - - - K T T M T V H W L S V R G G A R H L R K A A E Q G - - K K L E V A I AY P K D A G K N I T L G L V I T K D C - - - - - E T G V P N V G V Y R L Q L Q S - - - - - K N T M T V H W L S V R G G A R H L R K A A E N G - - K K L E V A I AY P S D A G K I I T L G L V I T K D C - - - - - E T G T P N V G V Y R L Q L Q S - - - - - K N T M T V H W L S V R G G A R H L R K A A E R G - - K K L E I A I AY V G D A G K I I T L G L V I T K D C - - - - - E T G T P N V G V Y R L Q L Q S - - - - - K N T M T V H W L S V R G G A R H L R K A A E R G - - K K L E V A I AY P K D A G K I M T L G L V I T K D C - - - - - E T G I P N V G I Y R L Q L Q S - - - - - P T T M T V H W L S V R G G A R H L R K A A A Q G - - K K L E V A V AW P G D A G G V I T L G L V I T K D P - - - - - E T G V P N V G V Y R L Q R Q S - - - - - V N T M T V H W L S V R G G A R H L R K A A A M G - - K K L E V A V AW P G D A S G V I T L G L V I T K D P - - - - - E T G V P N V G V Y R L Q R Q S - - - - - P K T M T V H W L S V R G G A R H L R K A A A M G - - Q K L E V A I AW P G D A G K I I T F G L V I T K D P - - - - - E T N T P N V G V Y R L Q Q Q S - - - - - I N T M T V H W L S V R G G A R H L R K A S A M G - - K K L E V A I AY A G D A G R I L T F G L V I T H D P - - - - - E D G T P N V G I Y R L Q Q Q S - - - - - R D T M T V H W L S V R G G A R H L R K A A E R G - - Q K L P V A I AW P G E P A P L I T W G L V F T K P P - - - P G A H G T D N V G V Y R M Q V L G - - - - - K D R L I M R W L A H R G G A K H H H Q W K A D K - - R E M P V A I VW P G D A G P L V T W P V V L T R P H G T S A E E T L H Y N A G V Y R A Q V I G - - - - - R D R L I M R W L A H R G G A G H C R S W M R A G - - E P M P V A L AW P E D A A P L I T W G L T V T R G P H K - - - E - - R Q N L G I Y R Q Q L I G - - - - - K N K L I M R W L S H R G G A L D Y Q E W C A A H P G E R F P V S V AW P D D G G A F I T L P Q V Y S E D P D K - - P G I M N A N L G M Y R V Q L T G N E Y E L D Q E V G L H Y Q I H R G I G V H Q T K A N Q K G - - E P L K V S I FW P G D A G . . I T L G L V I T K D P K S E T G P N V G V Y R L Q L Q S N E Y E L . . T M T V H W L S V R G G A R H L R K A A G P G K K L E V A I A
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
250 260 270 280 290 300 310 320
L G V D P L I I M A A A T P I P V D L S E W L F A G L Y G G S G V A L A K C K T V D L E V P A D S E F V L E G T I T P G E M L P D G P F G D H M G Y Y G G V E DV G I D P I L I M A A A T P I P V D L S E W L F A G L Y G G S G V A L T K C K T V D L E V P A D A E F I L E G T I T P G E M L P D G P F G D H M G Y Y G G V E DL G V D P L I I M A A A T P I P V D L S E W L F A G L Y G G S G V Q L A K C K T V D L E V P A D S E F V L E G T I T P G E I L P D G P F G D H M G Y Y G G V E DL G V D P L I I M A A A T P I P V D L S E W L F A G L Y G G S G V Q L A K C K T V D L E V P A D S E F V L E G T I T P G E V L P D G P F G D H M G Y Y G G V E DV G V H P L I I M A A A T P I P V D L S E W L F A G L Y G G G G I H L A K C K T L D L E V P A Q S E F V L E G T I T P G E V L P D G P C G D H M G Y Y G G V E DI G V H P L L V M A A A T P I P V Q L S E W L F A G I Y A G E G V R L T P C K T L D L Q V P S H S E V V L E G T I T P G E V L P D G P F G D H M G F Y G G V E DI G V H P L L I M A A A T P I P V Q L S E W L F A G L Y A G E G V R L T G C K T L D L K V P S H S E V V L E G T I T P G E E L E D G P F G D H M G F Y G G V E SI G V H P L I L M A A A T P I P V T L S E W L F A G L Y A G K G V R L S N C K T V N L Q V P S H S E I V L E G T I A P G E E M N D G P F G D H M G F Y G G I E KV G V D P L L V M A A A T P I P V E L S E W V F A G L Y A G E G V Q L A Q C R T S D L L V P A H S E F V L E G T I T P G E V L P D G P F G D H M G Y Y G G V E DI G A D P S M I L S A V L P L P E T V S E I K F A G L L G G E R P S L T P C Q T I P I S V P A D A E I V L E G F V S P T E T A P E G P Y G D H T G Y Y N A V E EL G C D P A L L L A A A L P L P E Q V S E L T F S G V L R G A R T P L V A G R T V P L M V P A T A E I V V E G W V H P G D M A P E G P F G D H T G Y Y N S V E DL G A D P A T I L G A V T P V P D T L S E Y A F A G L L R G T K T E V V K C I S N D L E V P A S A E I V L E G Y I E Q G E T A P E G P Y G D H T G Y Y N E V D SV G G P P A H S L S A V M P L P E G L S E M T F A G L L S G R - - R F R Y S Y V D G Y C I S H D A D F V I T G E I P P G D T K P E G P F G D H L G Y Y S L I H D. G V D P L . I M A A A T P I P V L S E W L F A G L Y . G G V L . C K T . D L V P A S E F V L E G T I T P G E L P D G P F G D H M G Y Y G G V E D
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
330 340 350 360 370 380 390 400
S P L V R F Q C L T H R K N P V Y L T T F S G R P P K E E A M M A I A L N R I Y T P I L R Q Q V S E I T D F F L P M E A L S Y K A A I I S I D K A Y P G Q A K RS P L V R F H C L T H R K D P I Y L A T F S G R P P K E E A M M A I A L N R I Y T P I L R Q Q V S E I V D F F L P M E A L S Y K A A I I S I E K A Y P G Q A R RS P L I R F Q C M T H R K D P I Y L T T F S G R P P K E E A M M A I A L N R I Y T P I L R Q Q V S E I V D F F L P M E A L S Y K A A I I S I D K A Y P G Q A R RS P L I R F E C M T H R K N P I Y L T T F S G R P P K E E A M M A I A L N R I Y T P I L R Q Q V S E I V D F F L P M E A L S Y K A A I I S I D K A Y P G Q A R RS P V I H F H C L T H R R N P I Y L T T F S G R P P K E E A M I A L A L N R I Y T P I L R Q Q V P E I V D F F L P M E A L S Y K A A I I S I D K A Y P G Q A R RS P L V R F H C M T Q R R D P V F L T T F S G R P P K E E A M L A I A L N R I Y T P I L R Q Q I P E I T D F F L P M E A L S Y K L A V I S I D K A Y P G Q A K RS P L V R F H C V T Q R R D P I F L T T F S G R P P K E E A M L A I A L N R I Y T P I L R Q Q V S E I V D F F L P M E A L S Y K L A V I A I D K S Y P G Q A K RS P L I R F H C I T S R K D P I Y L T T F S G R P P K E E A M L A I A L N R I Y T P I L R R Q I N E I T D F F L P M E A L S Y K L A I I S I D K S Y P G Q A R RS P L V R I H T V T H R H D P I Y H T T F S G R P P K E E A M I A I A L N R I Y T P L L R A Q V P E V R D F F L P M D A L S Y K L A V L S I K K A Y P G Q A R RF P V M R I T A I T M R R H P I Y L S T Y T G R P P D E P S R L G E A F N D V F L P V A R R Q F P E I V D L W L P P E A C S Y R I A V A S I K K R Y P G Q A R RF P V L R V S A I T H R K D P L Y L T T H T G R P P D E P S V I G E V F N D L A M P V F R Q Q I P E V K D L Y L P P A A C S Y R I A I V S I D K R Y P G Q A R RF P V F T V T H I T Q R E D A I Y H S T Y T G R P P D E P A V L G V A L N E V F V P I L Q K Q F P E I V D F Y L P P E G C S Y R L A V V T I K K Q Y A G H A K RF P V M K V H K V Y A K Q G A I W P F T V V G R P P Q E D T S F G A L I H E L T G D A V K L E I P G V K E V H A V D A A G V H P L L F A I G S E R Y T P Y Q K VS P L . R F H C . T H R . D P I Y L T T F S G R P P K E E A M . A I A L N R I Y T P I L R Q Q V P E I V D F F L P M E A L S Y K . A I I S I D K A Y P G Q A R R
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
410 420 430 440 450 460 470 480
A A L A F W S A L P - - - - - - - Q F T Y T K F V I V V D K S I N I R D P R - - - Q V V W A I S S K V D P V R D V F I L P E T P F D S L D F A S E K I G L G G RA A L A F W S A L P - - - - - - - Q F T Y T K F V I V V D K D V N I R D P R - - - Q V V W A I S S K V D P V R D V F I L P D T P F D T L D F A S E K I G M G G RA A L A F W S A L P - - - - - - - Q F T Y T K F V I V V D K D I N I R D P R - - - Q V V W A I S S K V D P T R D V F I L P N T P F D T L D F A S E K I G L G G RA A L A F W S A L P - - - - - - - Q F T Y T K F V I V V D K D I N I R D P R - - - Q V V W A I S S K V D P V R D V F I L P N T P F D T L D F A S E K I G L G G RA A L A F W S A L P - - - - - - - Q F T Y T K F V I V V D K E I N I R D P R - - - Q V V W A I S S K V D P S R D V F I L G E T P F D S L D F A S E K I G L G G RA A M A F W S A L P - - - - - - - Q F T Y T K F V V V V D K H I N V R D P R - - - Q V V W A I A A Q V D P Q R D L F T L A D T P F D S L D F A S E Q L G L G G RA A M A F W S A L P - - - - - - - Q F T Y T K F V V V V D A N I N V R D P R - - - Q V V W A I A A Q V D P Q R D L F V L E N T P F D S L D F A S E H L G L G G RA A M A F W S A L P - - - - - - - Q F T Y T K F V V V V D S T I N V R D P R - - - Q V V W A I S S L V D P Q R D L L V M E N T P F D S L D F A S E N I G L G G RA A M A F W T A L A - - - - - - - Q F T Y T K F V I V V D E D I D I R D P S - - - Q V V W A I A S K V D P E R D L F V I P R T P F D T L D F A C E R I G L G A RL M M G L W S M L P - - - - - - - Q F S Y T K L L I I V D D D V D V R D W A - - - D V M W A V S T R C D T S R D M V S I S D T P I D Y L D F A S P K S G L G G KV M M A L W G M L A - - - - - - - Q F S Y T K M V I V V D E D I N P R D W D - - - D V A W A M A T R M D P S R D V V L L E K T P M D Y L D F A S P E P G L A G KV M M G V W S F L R - - - - - - - Q F M Y T K F V I V C D D D V N A R D W N - - - D V I W A I T T R M D P A R D T V L V E N T P I D Y L D F A S P V S G L G S KK Q P A E L L T I A N R I L G T G Q L S L A K Y L F I T A E Q D K P L D T H K E E E F L T Y L L E R I D L H R D I H F Q T N T T I D T L D Y S G T G L N T G S KA A M A F W S A L P N R I L G T G Q F T Y T K F V I V V D D I N . R D P R K E E Q V V W A I S S . V D P R D . F . L T P F D . L D F A S E . I G L G G R
S6803S7002A7120NpuncTelonS8102PmarMIT9313PmarSS122GvioRpalRspheEcolBH3930
490 500 510 520 530 540 550 560
M G I D A T T K I P P E T D H E W G E V L E - - - - - S D P A M A E Q V S Q R W A E Y G L G D I N L T E V N P N L F G Y D VM G I D A T T K I P P E T D H E W G E V L E - - - - - S D P K I A E R C D R R W A E Y G L A D I D L T A V D P N K F G Y E M NM G I D A T T K I P P E T D H E W G A P L E - - - - - S D A D V A A M V E R R W A E Y G L A D L Q L G E V D P N L F G Y D M KM G I D A T T K I P P E T D H E W G S P L E - - - - - S D P D V A A M V E R R W A E Y G L A E L Q L G E V D P N L F G Y D M KM G I D A T T K I P P E T D H P W G D P L T - - - - - S D P E V A R R V T E R W Q E Y G L G D I D L T A V D A T R F G Y E L D P A F R W RL A I D A T T K V G P E K N H D W G E P L S - - - - - R P A D L E E R V S A R W S E L G L D G L G Q D E P D P S L F G Y A L D R L I Q G L K T S PM A I D A T T K I G P E K R H E W G E P L S - - - - - R D A D L E S R V D A R W Q E L G L E D L G S E E P D P S L F G Y V M E S L C R Q A M V K K TL A I D A T T K V G P E K N H E W G N P L I - - - - - H S S N L S K K I D E R W N E L G L S D L K P N H A D P S L F G Y V I D E I I K F N Q I T K SL G I D A T T K I P P E T D H A W G E P L R - - - - - A D P A T A E Q V S K R W A E Y G L G D I P L G G V D P R A W G Y S G KL G I D A T N K I G T E T E R E W G K V L E - - - - - M D K D V I A R V D A M W T S L G L S P E H Q P A A G Q R R L I RI G I D A T N K I G P E T H R E W G E V M T - - - - - Q S P E A E A F A D R L I A K L R I G AM G L D A T N K W P G E T Q R E W G R P I K - - - - - K D P D V V A H I D A I W D E L A I F N N G K S AV V I A A Y G D K K R E L C Q E V P E L F K G L Q T F Q N P R L V M P G V V A L E G P S F T S Y E R A Q E E F R A F S E T I T Q N G E L P S C P M IM G I D A T T K I P P E T . H E W G E P L G L Q T F D P . . . . V . R W E G L . D . V D P L F G Y

298 The multiple was generated by BLOSUM30 using Clustal W. S6803, Synechocystis sp. PCC 6803; S7002, Synechococcus sp. PCC 7002; A7120, Nostoc sp. PCC 7120; Npun, N. punctiforme; Telon, T. elongatus BP-1; S8120, Synechococcus sp. WH8102; PmarMIT9313, P. marinus MIT9313; PmarSS122, P. marinus SS120; Gvio, G. violaceus PCC7421; Rpal, R. palustris; Rsphe, Rhodobacter sphaeroides; Ecol, E. coli; BH3930, Bacillus halodulans.

299
A.1.6. An amino acid sequence alignment of UbiH homologs in cyanobacteria and other
organisms
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
10 20 30 40 50 60 70 80
M T F V A A S I T D N Q F D V A I A G G G V V G L V L A A G L R H - - - T G L K IM E K S H L G F D V I I V G G G I V G V T L A A C L K R - - - T N L R I
M A L T Q L T Q T I S P Q P T P P E L R K Y D Y D L V I V G G G I V G L T L A S A L K D - - - S G L N IM A L T Q L E Q P L S P Q P T P P D L R G Y E Y D L V I V G G G I I G L T L A S A L K D - - - S G L S I
M M T S A F A P S E Q S L V A S P Q D L P A V D V A I V G A G I V G L S L A C A L R N - - - S G L A IM A T S S T T F H I L G A G P T G S L A A I A L A S - - - T G C S V
M N N Y L N F K I V G S G P S G L L L A I S L A K - - - L N F N IM V D H L N R F H S A A K G S L P V D I R V K I L G A G P T G T L L A L A L A R - - - L G N S V
M S N L H I K V I G A G P S G S L L A L S L V G - - - N S N A VM A F N N R N T L A A E R G K A A L Q A D C I I I G G G P A G A V L S L L L A R - - - Q G V S V
M P R G H A G D F P P P A L S V R N R P G N S G L V L E T C F A Q L R S G R K D H G A R V R N M T A P R S I V I G G G A F A G L A L A L A L R Q G - - L G P E VM E Y G V S E P L L R G L A A G D P P S A T G P V T G S A D K V A D V L I V G G G L V G G T L A C A L A E - - - K G V S V
M S V I I V G G G M A G A T L A L A I S R L S H G A L P VM Q A Q D D Q Q K G K D M T T A A A G P T E V D V A I I G A G M A G T T L A T L L G N - - - A G R K V
M P R G H A G D F P P P A L S V R N R D . . I . G G G . G . L A . A L S H G . V
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
90 100 110 120 130 140 150 160
A I I E A L P K E Q A L T K - - - P Q A Y A I S L L S G K I L A G L G V W E N I K D S I G H F E R I Q I S D N D Y R G T V P F A K E D V D E - - - - - L A L G HA I V E A Q P L D V V T Q R - - - Q R A Y A M S L L S Q R L F Q E L G V W E V I A A R S G K Y R N I Q L S D E D F S G V V R F S R Q D L K T - - - - - D F L G FL L I E A K V A S A A V A K - - - G Q A Y A V H L L S A L I Y Q G I G I W D K I L P Q I A K Y R Q V R L S D A D Y P G V V E F E S T D L G T - - - - - P E L G YL L I E A K V A S A A V A K - - - G Q A Y A V H Q L S A L I Y Q G I G V W D K I L P Q I A K Y R R V R L S D A D Y P N V V E F T T D D I H T - - - - - A E L G YA L I E A T P Y R R E N T K - - - G Q A Y A L H Q V S R Y F F E E I G V W D K L M P H V Q P F E V V Q L S D E R F P L T V H F S P A D L G T - - - - - E A L G YV L T D P L T R K E L L S R - - - S R A Y A I T H S S R R L L T D L N L W T S L Q G S L T A F S F L D L R D S A C G G R V V F G L D D L P N S N G R H E A I G WY I T D L L T R E K L I N K - - - D K T Y A I T H S T R K I L S K F N L W S K L K S Y L Y K F D S L S I S D S V T S D F T I L T T S D L D K D I C S L D T I G WT L C D P L T V E E L P A R - - - S R A Y A L T H S S R R L L Q S L D L W A S L I P H L A P F T R L R L D D Q E L N R H T Y F E V D D L S S Q N Q L H G A V G WS I Y E A K S K E Q I L L R - - - D R T Y A I N H S S R R L L Q R I G I W D D L N E F M I P F D S L S L E D Y S V N Q R L V L Y N K D L V N L N S S Y K E V G WI L F E A Q K D F D R D F R - - - - - G D S L H A V V M E L M D D L R L T E R L L E H K H S K L K T L T M V S A V G A A R V A D F S R L P T - - - - H H N Y M TP V I V A D P A L A L R P S - R D P R A T A I V A A C R R L F E A L G V W D E V A P T A Q P I I D M V V T D S H L E D A T R P V F L T F A G E V Q P G E P F A HV V I D G E D P E A L L A A G Y D G R C S A I A L A C Q R L L D T I G L W D L L G G E S Q P I L D I R V V D G G S P L F L H Y A Q A E A Q G - - - - - - P M G YH L I E A T A P E S H A H P G F D G R A I A L A A G T C Q Q L A R I G V W Q S L A D C A T A I T T V H V S D R G H A G F V T L A A E D Y Q L - - - - - A A L G QA L I D P H R V H H D E F R - - - - - A E K I G A D Q M Q L F E K L G L D K M V M P L V T P F T D I D V F R L G - - - - - Q F F A R E - - K - - - - - - K W E Y. L I E A . . . G D R A Y A . . . S . L . G . W . L . . . . . . D . V F D L N . G .
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
170 180 190 200 210 220 230 240
V A E H P V I L Q A L E N C V E Q C P R I A W F R P A E L I S F T A G E N H K Q V T L Q Q E G R - - - - E I T L Q T K L L V A A D G A R S H T R S L A G I Q T KS G Q H Q V V L E T L Q A S L R G Y G N I R W F C P A Q V T A I H Y E S E Q A I V E L S S E T Q G - - - Q I K I Q G A L V V G A D G P R S L V R Q G A S I K T RV A E H Q A L L E P L Q E F V Q N C P N V T Y L C P A E V V S T N Y Q P N E V V I A V K I A D Q - - - - L N T I R S K L L V A A D G S R S P I R Q A A G I K T HV A E H Q A L L Q P L Q E F V Q D C P N V T Y L C P A E V V N T Q Y Q K D I V A V D I K V A D Q - - - - L Y T V R S K L L I A A D G A R S P I R Q A A G I K T SV G E H S A I A E T L Q S V L Q S A G N V T F Y C P W R V V T N E V H G D R A Q L T L I S G D P G E P K V A H L A A R L V V A A D G G K S P L R Q Q M G I E P KI L D H Q P L M K L L M D R L H Q H H R V E L A L G - C T A P T P G I D D - - - - - L - - - - - - - - - - - - - - - - - I V A A D G P R S T T R S Q W D I G C WV V K H S D L M N V F F D E V D N V D N I F F K S S L D L S F N D I K F D - - - - - - - - - - - - - - - - - - - - - - F E F V S T G A N P N H K K N R N F I D II L D H R P L M E L L I D R L Q T S P K V L L N L G R F D D H H L D G H D - - - - - L - - - - - - - - - - - - - - - - - V V A A D G P R S L T R Q A W G I N T WT L D H K L L M E Y L F Y K I S K S S N I N I K F S S K D N D A N A L Y D - - - - - - - - - - - - - - - - - - - - - - Y T F A A D G T N S R Y R D L W K F K S YM V A Q K D F L N F M V A E A K R Y A N F R V F M A T S V Q R L I E E E G E I R G V V F N G P D G - - - P Q E A R C A L T V G A D G R S S R T R Q M A G I D L VM V E N R Y L V E A L A K R A E A E G V E L R A T P - - V T S Y D A R T E A I G V T L G D G S - - - - - - - V I D A S L L V A A D G A K S K L R Q R A G I A T YM V E N R L L R Q A I L T R L G R L P A A T L L A P A R M T A L R R D L D G V S A T L S D G Q - - - - - - - T V R A R L V V G A D G R R S Q V R E S A G I G I RV V E L H N V G Q R L F A L L R K A P G V T L H C P D R V A N V A R T Q S H V E V T L E S G E - - - - - - - T L T G R V L V A A D G T H S A L A T A C G V D W QA F S Y G A L I N G L R D A L P P Q V P I T I G K V A E V S T G P N R Q R - - - L V L A D G - - - - - - - A V I D A R L L V V A T G Y S E L V R R A I G V E R I. . . H . L . L . . P V . . . L G G E P K . L . V A A D G . S . R . . G I
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
250 260 270 280 290 300 310 320
G W K Y W Q S C V A F T I Q H Q A P D N T T A F E R F C D T G P - - - - - - - - M G I L P L P G D R A Q I V W T M - - - P H H K A H T L V N L P E A D F I T E LG W K Y W Q S C V T C V V E H D A P R N D V A F E R F W G T G P - - - - - - - - M G V L P L T E N R C Q I V W T N - - - P H A E A Q R L K E L P P E V F L E Y LG W K Y W Q S C I V A F V K P E K P H N D T A Y E R F W P S G P - - - - - - - - F A I L P L P E N R C R I V W T A - - - P H D E A K A L C A L N D E E F L R E LG W K Y W Q S C I V A F V K P E K P H N N T A Y E R F W S S G P - - - - - - - - F A I L P L P E N R C R I V W T A - - - P H E E A K A L C A L D D E Q F L A E LG W Q Y P Q S C V V A T L D V A H P Q P V I A Y E R F W P T G P - - - - - - - - M G V L P L P R N R Y R V V W T L - - - P H P E A E A V A A L D D R S F L A T LK F R Y R Q G C L T S K I A L R G V K P G M A Y E L F R P E G P - - - - - - - - F A I L P L G D G I F Q V V W S A - - - P W T Q C Q R R A D L P T P E F L D E LR K S Y N Q S C L T F E V L V R G N V P R R A Y E I F R K E G P - - - - - - - - L A L L P L D K N K Y Q I I W T A - - - T T S K S M D R L N S S K S F L L D N LN H P Y Q N G C L T A K V L L R G A D P L M A Y E L F R A E G P - - - - - - - - L A V L P M G G E V F Q M V W S G - - - P L R R C Q E R A G L S P S C F L D H LR F K Y N Q E C I T F K A L L R T P F T H R A Y E I F R K D G P - - - - - - - - M A I L P M A N D L Y Q I V L S M - - - P P L K S D Y L L N L T T S H F L D T VK F S Q - A V D V L W F R L P R Y P H E P E G V L Y R V G Q G A - - - - - - - - V M G Q A D R S D V W Q I S Y I I R K G T Y Q K L R A Q G I E G L R R S V T T LG W D Y D Q S G I V V T V E H E R D H N G C A E E H F L P A G P - - - - - - - - F A I L P L T G R R S S L V W S E - - - S R R D A E R I V A L P E K E F Q R E LT L G Y G Q T A I V L T V E H E R S H R G C A V E H F L P A G P - - - - - - - - F A I L P M P G N R S S L V W T E - - - R S D L V P G L L A L P A E H F Q A E LQ E P Y E Q L A V I A N V A T S V A H E G R A F E R F T Q H G P - - - - - - - - L A M L P M S D G R C S L V W C H - - - P L E R R E E V L S W S D E K F C R E LE L S K A H S L S M G F D L A I A P R - D C A Y R A V T C Y G K R A S D R V A Y L T V F P I G D K M R A N M F V Y R T V A E Q W T R D F R A D P Q K - M L C E L
. Y Q S C . . . V P . A Y E . F G P R A S D R V A Y A . L P L R . V W . R P . . . . L F L E L
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
330 340 350 360 370 380 390 400
R Q R I G D R L G E F H L I N A R R L F P V Q L M Q S D C Y V - Q P R L A L V G D A A H C C H P V G G Q G L N L G I R D G A A L A Q V I A T A H - S Q G E D W GE Q H T G P Q L G R L K L L G D R Q V F P V Q L M Q G D R Y V - D H R L A L V G D A A H C C H P V G G Q G L N L G I R D A A A L A Q T L I E A Q - Q R G Q D L GS Q R F G Q Q M G K L E L L G D R F V F Q V Q L M Q S D R Y V - L P R L A L V G D A A H N C H P V G G Q G L N L G I R D V A A L A Q V I Q K A H - Q A G E D I GT R R Y G D Q M G K L E L L G D R F V F Q V Q L M Q S D R Y V - L P R L A L I G D A A H N C H P V G G Q G L N L G I R D A A A L A Q V I Q T A H - Q A G K D I GQ P Y L D P Q M A E I T A V G D R F V F P A Q F M Q V N S Y A - A D R F V V I G D A A H R C H P V G G Q G L N L G L R D V W N L A Q Q I L A S - - - P L E E I GA A V L P A G I E P D L L L D T P R A F S Q Q W S M A R R L S - R G R G V L L G E A G H R C H P V G G Q G L N L C W R D V A S L R N L A E H G G - - - - - - - TS T I L P D N F K L D Q I N S D I N I F P V S L S F R L P I F N F K K V V F V G D S F H T F H P V G G Q G L N T C W R D V N V I Y D I F N R N - - F P V S N R AA A V L P D G L Q P D V L L D R P A A F P L Q L A F A F R L Q - R G R G V L V G E S G H R C H P V G G Q G L N L C W R D V S T L M N L V K N V D E - - - G K L PA T Y L P I G M E I D T L I N K P Q K F S L S L N V P I K L Q - D Y N R F L I G E S L H S L H P V G G Q G L N L S I R D I D D I M N L L T S S N - - - - - - - -V P E L A D R V G L L K D W S Q A S L L S V E S G Y V R R W Y - R P G L L L I G D A A H V M S P F G G V G I S Y A I Q D A V A A A N V L A G P L R - - - S G R VE K R F G L R L G D I K P L D K P K S F P L G Y F V A Q S F I - A P R L A L I G D A A H V I H P I A G Q G L N M G L R D V A A L A E V V V D A A R - L G I D P GE R R F G D H L G W V R P V G P R F S Y R L T L Q A A N R Y V - D H R L A L V G D A A H G M H P V A G Q G M N Y G L R D V A V L A E R L V A A Q R - L G L D P GQ S A F G W R L G K I T H A G K R S A Y P L A L T H A A R S I - T H R T V L V G N A A Q T L H P I A G Q G F N L G M R D V M S L A E T L T Q A Q - E R G E D M GM P E I A T Q C G N F E V A S P V E V R Q V N L T T T Q G H R - R D G V V F I G D A F V T T C P T P G V G I G R V M V D V D Q L H S V H I P R - - - - - - - W L
. . . G . . . . F . . L . R . . N R . . L . G D A A H . H P V G G Q G L N L G . R D V . . L A . . . . R G D G
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
410 420 430 440 450 460 470 480
S L A V L K R Y E H W R K P E N - W L I L G F T D L L D R F F S S H W L P A I A L R R F G L E V L R L V P P A K K L A L R L M T G L L G R K P - - - Q L A T G QS L A V L K R Y E R W R K R E N - L V I L G F T D F L N R L F S N R I W P V V W V R R L G L I V M A K F N P L R L F A L K L M T G Q K G R Q P - - - R I L AN I R V L K R Y E S W R Q K E N - L A I L G F T D M L D R M F S N Q F P P L V F L R R L G L W F L R S V P V L K T F M L K L M I G L K G K T P - - - E L A K RD I Q I L K G Y E R W R K L E N - L T I L G F T D L L D R M F S N N F L P V V V V R R L G L W F L Q R V P L L K V F M L K L M I G L K G R T P - - - E L A K RD R R H L M H F H R Q R W W Q N - V L T L A F T D F L N R L F S N T G W P L V A V R R C G L G L L C L L P P L K R L L L R F M A G L L L P L P G D R E F P R G RS Q R L A R R Y G R S R W A D L - L M V G L A T D L L V R L F S N Q Q P W L L P F R R I G L K A M A R F S W L R R I S L L A M T D G P T Q L L - - K P L P DL N I F K Y K Y Y F I R C L D I - I S T I V V T D S L I Q F F A N N K F F L L P F R K F S F L L L N K F I F M R R I I L N H M T K S L I Y S S I KV E K L P Q V Y A R K R I F D L - V L V G L F T D L I V R F F S S R N I L L L M V R L P L M F L L A R L S L L R Q L L L K A M T D G P I T V I - - R L L P E- - - - L L Q Y K I C R Y I D I - Y T T S F L T H M L I V I F S S N N I I L K V F K L S L F T I L R K S I L F R K I V L S L M T D G I P H I F KG V G H L A A V Q R R R E L P T R L S Q Y Y Q A M I Q D Q L L A T A A N E G I P P K P T L L R R L L R L P G L R E L P T R L L A F G L W P A R L R I K N S I P PQ V D V L E R Y Q R W R R F D T - V A M G V A T N G L N F L F S N N S P L L R G I R D I G L G L V D R L P P L K G A F I R Q A A G L T G E I P - - - R L L K G EA P A L L A E Y E A L R R P D N - L L M L A I T D A L V R L F S N D I A P V A L A R R L G I G A V E R M G P L K R L F M R H A M G T L K L G P E P P R L M R G VD Y G V L C R Y Q Q R R Q S D R - E A T I G V T D S L V H L F A N R W A P L V V G R N I G L M T M E L F T P A R D V L A Q R T L G W V A RE T P G M A A D K I G A F Y D D P V K V A A D K A G M R V S F Y A K Q I T T E T G L E W R V R R L R N N A A R Q L M I I G R K M R H L G P R R - - - E P G M A
. L . Y . R D . . . . T D . L R . F S N L . . R . G L . L . L . . . L . . M G . . . P L
S6803S7002A7120NpuncTelonS8102PmarMED4PmarMI9313PmarSS122GvioRpalRrubEcolMlr2092
490 500 510 520 530 540 550 560
S L V S Q
N I H RA LP L
L Q

300 The multiple alignment was generated by BLOSUM30 using Clustal W. S6803, Synechocystis sp. PCC 6803; S7002, Synechococcus sp. PCC 7002; A7120, Nostoc sp. PCC 7120; Npun, N. punctiforme; Telon, T. elongatus BP-1; S8120, Synechococcus sp. WH8102; PmarMED4, P. marinus MIT9313; PmarMIT9313, P. marinus MIT9313; PmarSS122, P. marinus SS120; Gvio, G. violaceus PCC7421; Rpal, R. palustris; Rrub, R. rubrum; Ecol, E. coli; Mlr2092, Mesorhizobium loti.

301
A.1.7. An amino acid sequence alignment of UbiX homologs in cyanobacteria and other
organisms
S6803S7002A7120NpunTelonS8102PmarMIT9313PmarSS122GvioEcolRpalRspheApe1647
10 20 30 40 50 60
M A Q P L I L G V S G A S G L I Y A V R A I K H L L A A D - Y T I E L VM D R P L T V G I S G A S G L I Y A V R T L K Y L L E A N - L T V D V V
M S Q H N K P L I I G V S G A S G L I Y A V R A L K Y L L T A D - Y E I E L VM S Q K S K P L I L G V S G A S G L I Y A V R A I K F L L E A D - Y S I E L V
M I A V V T S S S L P I V L G V T G A S G L I Y A V R A I K F L L Q A N - Y P I Q L VM H P Y V L A V T G A S A Q P L A E R T L Q L L L Q A G - R S V H L VM D P Y V L A V S G A S A Q P L A E R S L Q L L L E N N - R D V H L IM S S I V L A I T G A S A Q I L A E R S I D L L L R C N - Q K L D V I
M Q T Q A P S S R P I V L G V A G A S G L I Y A V R T V R F L L A A G - H A V D L VM K R L I V G I S G A S G A I Y G V R L L Q V L R D V T D I E T H L V
M S T Q H R I V V G I S G A S G A A V G L R V V E L L A S V D - C E V H L VM G R G H D A V A R G R S L R R P A D R E A E D R S M R V V L G V S G A S G A A L A A A C A R H L S A L G - A E I D L V
M S C R P S S V S I A V T G S S G V R V A L R L L E A L K G E V - E I R G I IM G R G H D A V A R G R S L R R P . P . V L G V S G A S G . I Y A V R . . . L L A D . . L V
S6803S7002A7120NpunTelonS8102PmarMIT9313PmarSS122GvioEcolRpalRspheApe1647
70 80 90 100 110 120
A S R A S Y Q V W Q A E Q N I Q M P G E P S A Q A E F W R S Q A G V E K G G K L I C H R W G D V G A T I A S G S Y R C AA S K A S F M V W Q A E N G I T M P A D P D H Q A K F W R D Q A G V P E K G I L R C H R W G D V G A G I A S G S Y R T KA S K S T Y M V W Q A E Q E T R M P S E P K Q Q E Q F W R E Q A G V A L S G K L R C H P W S D V G A N I A S G S F R T LA S K S T Y M V W Q S E Q E I R M P P E P T Q Q E Q F W R E Q A G V A L S G K L R C H P W G D V G A G I A S G S F A T LA S K A A H Q V W R A E Y G L T L P T Q P D K Q E L F W R E Q A Q V W - E G C L T C H R P T D V A A A I A S G S F R T LL S R G A Y A V F Q A E Q G V Q V P V N P E R Q A S F W R E R L N C S - E G E L F C H R W N D Q S V G I A S G S F R T SF S H G A L Q V W K A E R G I E V P V D P S D Q E K F W R D H L Q V K - N G N L T C H R W N D Q S A C I A S G S F R T RI S K G A Y E V W K S E M N V N I P A E G A K Q A T F W R N R L K N N - Q G E I I C H K W N D N S A T I A S G S F R T KA S R A S F M V W R E E M G T A M P V E P A E Q E R F W R E Q S G E S - G G K L R C H A F A N L A A S I A S G S Y R T RM S Q A A R Q T L S L E T - D F S L R E V Q A L A - - - - - - - - - - - - - - D V T H D A R D I A A S I S S G S F Q T LV S K A A R R T I A Y E V G P N A L D H A A D L V - - - - - - - - - - - - - - H R F H K I E D V G A C I A S G S F R T SI S K A G E R T L V E E I G L E A A D A L R A Q A - - - - - - - - - - - - - - T R V H A I M N V G A A I A S G S A P V AL T R G A E E V A R Y E E G I E P E D L R R L L R - - - - - - - - - - - - S Y A P L Y M E N D M S S P L A S S S N Q P D. S K A A . V W . E G . P . P Q . F W R E Q A G V G L C H . W D V . A I A S G S F R T
S6803S7002A7120NpunTelonS8102PmarMIT9313PmarSS122GvioEcolRpalRspheApe1647
130 140 150 160 170 180
G M V V L P C S M S T V A K L A V G M S S D L L E R A A D V Q I K E G K P L V V V P R E T P L S L I H L R N L T S L A EG M L V I P C S M S T V A K I A N G L S S D L L E R A A D V S I K E G R K V V V V P R E T P F S L I H L R N L T Q L A EG M I V I P C S M S T V A K L A G G L S S D L L E R A A D V H L K E G R K L V I V P R E T P F S L I H L R N L T T L A EG M I I I P C S M S T V A K L A V G L S S D L L E R A A D V Q L K E G R K L V I V P R E T P F S L I H L R N L T S L A EG M V I L P C S M S T V A K L A A G L S S D L L E R V A D V H L K E H R P L V L V P R E T P F S L I H L Q N L T Q L A MG M L I V P C S M G T A G R I Q A G V A M D L I E R C A D V H L K E G R P L L I A P R E M P F N L I H L R N L T A L A EG M A I V P C S M G T I G R I N A G V S I D L I E R C A D V H L K E G R P L V I A P R D M P W S I I H L R N L T S L A EG M V I V P C S M G T I G R I A S G S S I N L I E R C A D V H L K E G R P L I I S P R E S P F N L I H L R N M T T L C EG M L V I P C S M S T V G K L A A G L S S D L L E R A A D V Q L K E G R P L V L V P R E T P F S L I H L R N L T A L A EG M V I L P C S I K T L S G I V H S Y T D G L L T R A A D V V L K E R R P L V L C V R E T P L H L G H L R L M T Q A A EG M I V A P C S I R T L S A I A H G D L D N L L V R A A D V Q L K E R R R L V L M L R E T P L H L G H I R S M A Q A T EG M I V A P C S M R S L A A I A H G L D D N L L T R A A S V Q L K E R R R L V L L A R E S P L T L A H L R N M T A V T EA M A I V P A S M K T V G L I A R G I P S S L P A R A A L A V L R L G R R L V V A P R E T P L G V V E L E N M L A I A RG M . I . P C S M T V . . I A . G . S S D L L E R A A D V L K E G R P L V . . P R E T P F S L I H L R N L T L A E
S6803S7002A7120NpunTelonS8102PmarMIT9313PmarSS122GvioEcolRpalRspheApe1647
190 200 210 220 230 240
A G V R I V P A I P A W Y H Q P Q S V E D L V D F V V A R A L D Q L A I D C V P L Q R W Q G G M E G EV G V K I V P A I P A W Y H Q P Q T I H D L V D F V V A R A L D Q F E I D C V P L K R W K E E A A S Q K S E NT G V R I V P A I P A W Y H N P Q T I E D L V D F V V A R T L D Q L D I D C V P L K R W E G G K H N SV G V R I V P A I P A W Y H N P Q T I E D L V D F V V A R A L D Q L D I D C I P I Q R W E G R RA G A R I V P A I P A W Y H R P Q T I E D L V D F V I A R A L D Q L G L D C V P L Q R W Q G P L SA G A R I A A P I P A W Y T Q P R T L E E M V D F I V I R L L D G F E D D L A P L Q R W T G P I KA G A K I A P P I P A W Y T K P N T I E E M V D F L V V R L F D L F D E R L A P I N R W N G P S QA G A K I I P C I P A W Y S K P K D L E E V I D F M V V R L Y D L F D L N M K D I N R W K G NA G A R I V P A I P A W Y Q Q P K T I E D L V D F V I G R A L D Q L D I D H S L F E R W H P D TI G A V I M P P V P A F Y H R P Q S L D D V I N Q T V N R V L D Q F A I T L P E D L F A R W Q G AI G A I V A P L S P A F Y Q R P R S I S E M V D H M A L R A I G L L G L N D L E L S A P E W S D D T P P T C P K PM G G I V A P P V P A F Y L R P E S L A A A V E Q I A A R A V D L L G L G R P Q A Q A W EM G G I V V P L T L S F Y I K P S S V E D L V D F A A G K V L D A L G V K V D V Y R R W R G P E E G D. G A . I V P . I P A W Y . P . T I E D L V D F . V . R A L D Q L . . D . P . . R W G K P
The multiple alignment was generated by BLOSUM30 using Clustal W. S6803, Synechocystis sp. PCC 6803; S7002, Synechococcus sp. PCC 7002; A7120, Nostoc sp. PCC 7120; Npun, N. punctiforme; Telon, T. elongatus BP-1; S8120, Synechococcus sp. WH8102; PmarMIT9313, P. marinus MIT9313; PmarSS122, P. marinus SS120; Gvio, G. violaceus PCC7421; Rpal, R. palustris; Rsphe, R. sphaeroides; Ecol, E. coli; Ape1647, Aeropyrum pernix K1.

302
A.1.8. An amino acid sequence alignment of UbiE homologs in Synechocystis sp. PCC
6803
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
10 20 30 40 50 60
M S N S L L T Q P T Q E S V Q Q I F A R I A P - - - - - - Q Y D D L N T F LM P E Y L L L P A G L I S L S L A I A A G L Y L L T A R G Y Q S S D S V A N A Y D Q W T E D G I L E Y Y W G D H I H L G
M A T I F R T W S Y Q Y P W V Y A L V S - - - - - - - R L A T L N - - -M L S N S D Q H R A V Q A L Y N T Y P F P P E - - - - - - P L L Q E P P - -M L E R I L E P E V M D T E A E A M D Y D A M - - - - - - D F Q A V N - - -
M V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S - - - - - - K Y D V M N D L MM P E Y L L L P A G L M L . . . . . . D G I L E Y . N L
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
70 80 90 100 110 120
S F G Q H - - - H I W K A M A V K - - - - - - - W S G V S - - - P G D R L L D V C C G S G D L A F Q G A K V V G T R G KH Y G D P P V A K D F I Q S K I D N V H A M A Q W G G L D T L P P G T T V L D V G C G I G G - - - S S R I L A K D Y G N- V G G E - - - E R F H Q L P L E - - - - - - - N L A I S - - - P G Q K V L D L C C G G G - - - - Q A T V Y L A Q S G A- P G Y N - - - W R W Q W T A A H N - - - - - F C L G R R P A N Q K V R I L D A G C G T G V G T - E Y L V H L N P E A E- L A - - - - - - - F A Q R A C Q - - - - - - - - L G P S - - - - K A T V L D A G T G T A R I P - I L L A Q L R P A W QS F G I H - - - R L W K R F T I D - - - - - - - C S G V R - - - R G Q T V L D L A G G T G D L T A K F S R L V G E T G KS G P V A . . N V H A M A . G . P G V L D . . C G . G . . . G
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
130 140 150 160 170 180
V V G L D F C A E L L A I A A G K H K S K - Y A H L P M Q W L Q G D A L A L P F S D N E F D G A T M G Y G L R N V G - -N V T G - I T I S P Q Q V K R A T E L T P - P - D V T A K N A V D D A M A L S N P D G S F D V V W S V E A G P H M P - -T V V G - L D A S P K A L G R A K I N - - - - - V P Q A T Y V Q G L A E D L P F G E G E F D L V H T S V A L H E M T P AV H A V D I S E G A L A V A Q T R L Q K S G V V C D R V H F H H L S L E N L A H L P G Q F D Y I N S V G V L H H L P - -I T A I D F A R S M L A I A K E N V I A A - G C E T Q I C L E F V D A K Q L P Y A N G S F D G V I A N S L C H H L P - -V V L A D I N E S M P K M G R E K L R N I - G V I G N V E Y V Q A N A E A L P F P D N T F D C I T I S F G L R N V T - -. V . . D . S A . . . . . G . . . . A L P . . G F D . . . L . . P A
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
190 200 210 220 230 240
N I P Q A L T E L Q R V L K P G K K V A I L D F H Q - - - - - - - - - - - - - P G N - - A L A A N F Q R W Y L A - - - -D K A V N A K E L L R V V K P G G I L V V A D W N Q R D D R Q V P L N F W E K P V M - - R Q L L D Q W S H P A N - - - -Q L Q S I I S G V H R V L K P G G I F A L V D L H R - - - - - - - - - - - - - P - - - - - - - S N W L F W P P L - - - -D P V A G I Q A V A E K L A P G G L F H I F V Y A E I G R W E I Q L M Q K A I A I L Q G K K R G D Y Q D G V A V G R E IR P L D F L R E V K R V L K P H G F L L I R D L I R - - - - - - - - - - - - - P E S P - E K L A D F V T A I G S - - - -D K D K A L R S M Y R V L K P G G R L L V L E F S K - - - - - - - - - - - - - P I I - - E P L S K A Y D A Y S F - - - -
. . . R V L K P G G . . . . D . L P . G L . . . G R E I
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
250 260 270 280 290 300
- - - - - - - - N V V V P M A K Q W R - - - - - - - - L T E E Y A Y L Q P S L D R F P T G P K Q V Q F A L E V G F - - A- - - - - - - - A S I E G N A E N L E A - - - - - - - T G L V E G Q V T T A D W T V P T L P A W L D T I W Q G I I R P Q- - - - - - - - A I F M G L F E T E T - - - - - - - - - - - A W Q L I N T D L G S L L D Q A G - F T V V R K H L Y - - AF A S L P E Y N R L V K R E K E R W S L E N H R D E S F A D M Y V H P Q E T D Y N I D T L F E L I D S A G L E F L G F S- - - - - - - - D Y N D Q Q R K L F A D S L H - - - - A S L T I A E I Q D Y L Q H L G W V G D D G P N L E L H L Y Q - S- - - - - - - - H V L P R I G S L V A N - - - - - - - D A D S Y R Y L A E S I R M H P D Q D T L K A M M Q D A G F - - EF A S L P E Y N . . H R D E S . . . . . . . . . . .
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
310 320 330 340 350 360
K A V H Y P I A - - - - - - - - - - A G L M G V L V A E KG W L Q Y G I R - - - - - - - - - - G N I K S V R E V P T I L L M R L A N G - - V G L C R F G M F K A V R K N A T Q A NG G S L Q V I Q - - - - - - - - - - A R A N K T V NN P D Y W Q L D R L L G K A P D L M E R A K D L S E K E R Y R L I E L L D P E I T H Y E F F L A K P P I Q I S D W T D DS D R H W T L E - - - - - - - - - - K K F Q S S T I V Q RS V D Y Y N L T - - - - - - - - - - A G V V A L H R G Y K F
. . R L L G K A P D L M . . . . . L L E I F
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
370 380 390 400 410 420
Q T L L K A L P N L H P C L T G W P S R S L F D Y N Y Q P L E L T E E E F K F L Q A V E A Q T E I Q A N V E K V L T D L
Q T L L K A L P N L H P C L T G W P S R S L F D Y N Y Q P L E L T E E E F K F L Q A V E A Q T E I Q A N V E K V L T D L
Sll1653Sll0418Sll0829Sll0487Slr1039UbiE
430 440 450 460 470 480
G D R S I D L A L V R S L Q T R Q L L T L G N S
G D R S I D L A L V R S L Q T R Q L L T L G N S
The multiple alignment was generated by BLOSUM30 using Clustal W. Sequences with “Sll” and “Slr” prefixes indicate the UbiE homologs of Synechocystis sp. PCC 6803. UbiE indicate the E. coli sequence.

303
A.1.9. An amino acid sequence alignment of Sll1653 homologs in cyanobacteria Sll165389-c70244Alr5252Npun3758Tery3321Tll2373Synw1674Pmm0431Pmt0276Pro0427Gll0127UbiE
10 20 30 40 50 60
M S N S L L T Q P - T Q E S V Q Q I F A R I A P Q Y D D L N T F L S F G Q H H I W K A M A V K W S GM S K P N A A T E I Q G I F N R I A P Q Y D A L N E W I S L G Q H R I W K K M A V K W S GM T N - - - - - K I R A I F D R I A P V Y D Q L N D W L S L G Q H R I W K E M A I K W T GM T N - - - - - E V Q S I F N R I A P V Y D Q L N D W L S L G Q H R I W K E M A V K W S AM T D D - - S I E V K A I F N K I A P V Y D Q L N D L L S L G I H K V W K Q M A V R W S EM T N - - - - - - I Q A L F E R I A P L Y D R L N D Q L S F G L H H V W K Q M A V D W L EM K P G - D P A A V E Q L F D S V A S R Y D Q L N D L L S L G L H R Q W K R Q L Q C L L RM R Y K - K T N E V K S I F N N I S C S Y D F L N S L L S L G L H K Y W K K T L V D L L K
M K P Q - D P I A I K E L F D S V S K N Y D F L N D L F S L G F H R I W K R Q L L T W L KM Q G S - - - Q E I Q G I F N A I A P V Y D Q I N D R M S F G L H R V W K K M A I R W T D
M V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S K Y D V M N D L M S F G I H R L W K R F T I D C S GM V D K S Q E T T H M V I F I A P Y D L N D L S L G H R . W K M A V W .
Sll165389-c70244Alr5252Npun3758Tery3321Tll2373Synw1674Pmm0431Pmt0276Pro0427Gll0127UbiE
70 80 90 100 110 120
V S P G D R L L D V C C G S G D L A F Q G A K V V G T R G K V V G L D F C A E L L A I A A G K H K S K Y A H L P - M Q WV T G G D L A L D V C C G S G D L T L L L A K T V G V Y G K T V G I D F A S Q Q L A I A A Q K Q N R L C P H L D - I Q WA K P G D T C L D L C C G S G D L A L R L A R R V G S T G Q V S G V D F S A N L L E T A K Q R A Q S Q Y P Q P N - I S WA K S G N T A L D L C C G S G D L A L R L A R R V G A T G Y V Y G V D F S C N L L E T A K E R S Q K Q Y P Q P A - I A WP D Y G S T C L D L C C G S G D L A Y M L A K H V G N T G H V F G V D F S R E Q L A I A R N R E P P L L N K I S P I H WL P Q G A T A L D L C C G T G D L T R L L A R R V G R Q G R V V G L D F A A A P L A I A R Q R S D - - - - H Y P Q I E WP A A G E T W L D L C C G T G D L A L E L A R R V R P G G S V L G L D A A A A P L E L A R H R Q S R Q - - P W L N V V FP I N G E N W A D L C C G T G D L S F L I F K K V R P N G S V T G I D S A K E I L N I A K E K S K L I - - E N K F I S W
M A R R R A A V E - - P W L P L S WP A S G E K W I D L C C G T G D L S L P L A R L V R P F G S V I G V D F S R A Q I N C A H K R S L K E - - P W L P I S WC K P G D T V L D L C C G T G D L A F L L A R R V G V A G T V W G V D F A A A Q L A Q A R R R D - - - - - R E G R V R WV R R G Q T V L D L A G G T G D L T A K F S R L V G E T G K V V L A D I N E S M P K M G R E K L R N I G - V I G N V E Y
G . T L D L C C G T G D L A L A R . V G G V G . D F L A . R P I W
Sll165389-c70244Alr5252Npun3758Tery3321Tll2373Synw1674Pmm0431Pmt0276Pro0427Gll0127UbiE
130 140 150 160 170 180
L Q G D A L A L P F S D N E F D G A T M G Y G L R N V G N I P Q A L T E L Q R V L K P G K K V A I L D F H Q P G - N A LQ E G D A L A L P Y P D N H F D G A T M G Y G L R N V T D I P Q A L R E L Q R V L K P G K K V A I L D F H R P D - N P GV E A N V L D L P F K D N Q F D A A T M G Y G L R N V T D I P R S L Q E L H R V L K P N A K A A I L D F H R P N - N Q QV E A D V L N L P F D D N Q F D A A T M G Y G L R N V K D I P R S L Q E L H R V L K P G A K A A I L D F H R P S - N P QV E A N A L S L P F P H N Y F D C A T M G Y G L R N V S N I P L C L Q E L Y R V L K P N A K A A I L D M H C P S - S S TL Q G D A L A V P F A P Q T F Q G I T I G Y G L R N V V D I P Q A L R E M F R L L V P G G R A A I L D F S H P Q - T S AQ Q G D A L S T G L P D A S A D G I V M A Y G L R N L A D P V V G L K E M A R L L K P G R L A G V L D F N R L T T A G PE M Q D I L E I D E D L K N Y D G I C M S Y G L R N L V N V E E G L K K V F N L L S D T G R A G F L D F N H S K F N S IL Q G D A L D T G L P S H R F D G A V M A Y G L R N L A D P G A G L I E L R R L L R P G A R A G V L D F N R M G E G S LL N E D V L K A G L P S S T F D G V V M A Y G L R N L S S P E A G L K E I H R L L K P G A R A G V L D F N H T I E G S KL E A D A L D L P F A D D S F D A V T Q G F G L R N V V D I P A C F A Q L R R V L K P G G R A V I L D L H A P A - D A GV Q A N A E A L P F P D N T F D C I T I S F G L R N V T D K D K A L R S M Y R V L K P G G R L L V L E F S K P I I E P L. . D A L L P F D . F D G . T M G Y G L R N V D I P . L E L R V L K P G . . A . I L D F . P .
Sll165389-c70244Alr5252Npun3758Tery3321Tll2373Synw1674Pmm0431Pmt0276Pro0427Gll0127UbiE
190 200 210 220 230 240
A A N F Q R W Y L A N V V V P M A K Q W R L T E E Y A Y L Q P S L D R F P T G P K Q V Q F A L E V G F A K A V H Y P I AK Q M F Q Q W Y L D R F V V P L A E R F Q L T P E Y A Y I Q P S I E R F P T G K T Q E K L A K E A G F R N A I H Y G I VF R T F Q Q W Y L D S I V V P L A D R L G V K E E Y A Y I S P S L D R F P I G K E Q V E I A L K V G F T S A T H Y P I AL R A F Q Q L Y L N S F V V P V A N Y L G L K E E Y A Y I S P S L D R F P I G K E Q I E L A R Q V G F A V A T H Y P I AI R W F Q K W Y L D T V V V P V A S N L G L T K E Y A Y I S P S L T K F L T G K D Q V A L A Y K L G F V N A K H Y P I IL E Q F Q Q W Y L Q Q W V V P T A R H Y G L A A E Y D Y L W P S I Q A F P T P P T L C A L I Q Q A G F E R V K H Y P L LA A D F Q R F Y L R R L V V P V A S A A G L K E E Y A Y L E E S L K R F P D G T A Q E R L A L E A G F A E A R H H T L VA D L F Q K I Y L R F V V V P I S K I F R L S K E Y S Y I E K S I K N F P K G N E L M L I A K G V G F K K V E Y R T I FA A R F Q R F Y L R R L V V T V A A Q V G L R E H Y A Y L E A S L Q Q F P K G E V Q E R L A R D A G F A A A S H R P L AN S F F Q K F Y L R N F V V P I A S K M G L K E E Y A Y L E E S L K K F P V G L I Q E Q I A I N V G F Q E A N Y K T L AW R A F Q R W Y L E G W V T A L G R A Q G L E A E Y A Y I A P S L E R F P T G D E Q V R L A Q Q A G F S Q A S H Y P L IS K A Y D A Y S F H V L P R I G S L V A N D A D S Y R Y L A E S I R M H P D Q D T L K A M M Q D A G F E S V D Y Y N L T. F Q . . Y L . V V P . A G L . E Y A Y . P S L . F P G Q L A . G F A H Y P . .
Sll165389-c70244Alr5252Npun3758Tery3321Tll2373Synw1674Pmm0431Pmt0276Pro0427Gll0127UbiE
250 260 270 280 290 300
A G L M G V L V A E KG D M M G V L V A T K NN G M M G V L I I S KN G M M G V L V V S K FG G M M G I L V I T KG G L M A I T V A Q KA G Q M G M L L L K RF N Q M G I L I L E KA G Q M G A L L L T AG G Q M G A L L L R AG G T V G V L V A QA G V V A L H R G Y K F. G M G . L . . K F
The multiple alignment was generated by BLOSUM30 using Clustal W. Locus tags are used to indicate each organism (see also Table 6.5): Sll1653, Synechocystis sp. PCC 6803; Alr5252, Nostoc sp. PCC 7120; Npun3758, N. punctiforme; Tery3321, T. erythraeum; Tll2373, T. elongatus BP-1; Synw1674, Synechococcus sp. WH8102; Pmm0431, P. marinus MED4; Pmt0276, P. marinus MIT9313; Pro0427, P. marinus SS120; Gll0127, G. violaceus PCC7421. 89-c70244 indicates a tentative locus in Synechococcus sp. PCC 7002.

304
A.1.10. An amino acid sequence alignment of Sll0418 homologs in cyanobacteria Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
10 20 30 40 50 60
M P E Y L L L P A G L I S L S L A I A A G L Y L L T A R G Y Q S S D S V A N A Y D Q W T E D G I L E Y Y W G D H I HM V Y H V R P K H A L F L A F Y C Y F S L L T M A S A T I A S A D L Y E K I K N F Y D - - D S S G L W E D V W G E H M H
M N I L L L A L G I S A T I L A I A L I S Y L L T A R R Y Q S A D T V A D S Y D E W T E D G I L E Y Y W G E H I HM S W L F S T L V F F L T L L T A G I A L Y L I T A R R Y Q S S N S V A N S Y D Q W T E D G I L E F Y W G E H I H
M T N E V Q S I F N R I A P V Y D - - - - - - - Q L N D W L S L G QM Y L Y Y A L G F I L L L V A L G I V I Y L I T P R S Y E S S N T V A N S Y D D W T Q D G I L E F Y W G E H I H
M S H L G L I L V I T V V A L V L L G L A L Y L L F P R K Y E S A R S V A E S Y D N W T K D G I L E F Y W G E H I HM L A G L L L L T G A A G A T A L L I W L Q R D R R Y H S S D S V A A A Y D A W T D D Q L L E R L W G D H V H
M S I F L I S S L V I F L T L L F S S L I L W R I N T R K Y I S S R T V A T A Y D S W T Q D K L L E R L W G E H I HM I L A G T T L T S A V V I W T Q R D R R Y K S S A S V A S A Y D A W T N D Q L L E R L W G E H V H
M I A F L L P I V S L L V L S L I F L W L F N D R K Y K S S E S V S S A Y D S W T N D R L L E K L W G E H I HM G E R T V L N E N I R R F Y D - - A S S G L W E E V W G E H M H
M V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S K Y D V M N D L M S F G IM V M L L . . . . . . . . . . L . T R . Y S S . . V A . Y D W T D G . L E W G E H I H
Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
70 80 90 100 110 120
L G H Y G D P P V A K D - F I Q S K I D N V H A M A Q W G G L D T L P P G T T V L D V G C G I G G S S R I L A K D Y G NH G Y Y G P H G T Y R I D R R Q A Q I D L I K E L L A W A V P Q N S A K P R K I L D L G C G I G G S S L Y L A Q Q H Q AL G H Y G N P P R A K N - F L K A K A D F V H E M V R W G G L D Q L P P G T K V L D V G C G I G G S S R I L A R D Y G FL G H Y G S P P Q R K D - F L V A K S D F V H E M V R W G G L D K L P P G T T L L D V G C G I G G S S R I L A R D Y G FH R I W K - - - - - - - - - - - - - - - - - E M A V K W S A A K - - - S G N T A L D L C C G S G D L A L R L A R R V G AL G H Y G S P P R R K D - F L Q A K A D F V H E M V K W G G L D K L P R G T T V L D V G C G I G G S S R I L A K E Y E FL G H Y G L P P R P K D - F R Q A K V D F V H E M V R W A G L D R L P P G T T V L D V G C G I G G S S R I L A R D Y G FL G H Y G N P P G S V D - F R Q A K E A F V H E L V R W S G L D Q L P R G S R V L D V G C G I G G S A R I L A R D Y G LL G F Y P - L N K N I D - F R E A K V Q F V H E L V S W S G L D K L P R G S R I L D V G C G I G G S S R I L A N Y Y G FL G Y Y G K P P S T R D - F R A A K Q D F V H E L V Q W S G L A Q L P R G S R V L D V G C G I G G S A R I L A R D Y N FL G Y Y E N S Y K T K D - F R Q A K I D F V H K L A H W S G L S T L P K G S R I I D I G C G I G G S S R I L A K D Y G FH G H W E V G E A D K D - R R V A Q V D L V V R L L D W A G - - - I D R A E S I V D V G C G I G G S S L F L A E R F G AH R L W K R - - - - - - - - - - - - - - - - - F T I D C S G V R - - - R G Q T V L D L A G G T G D L T A K F S R L V G EL G H Y G P P K D D F R Q A K . D F V H E . V . W . G L D L P . G . . V L D V G C G I G G S S R I L A R D Y G F
Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
130 140 150 160 170 180
N - - V T G I T I S P Q Q V K R A T E L T P P - - - D V T A K N A V D D A M A L S N P D G S F D V V W S V E A G P H M PE - - V M G A S L S P V Q V E R A G E R A R A L G L G S T C Q F Q V A N A L D L P F A S D S F D W V W S L E S G E H M PD - - V T G I T I S P K Q V E R A T Q L T P P - - - G L T A K F A V D D A M N L S F A D G S F D V V W S V E A G P H M PA - - V T G I T I S P Q Q V Q R A Q E L T P Q - - - E L N A Q F L V D D A M A L S F P D N S F D V V W S I E A G P H M PT G Y V Y G V D F S C N L L E T A K E R S Q K Q Y P Q P A I A W V E A D V L N L P F D D N Q F D A A T M G Y G L R N V KE - - V T G V T I S P K Q V Q R A T E L T P Q - - - G V T A K F Q V D D A L A L S F P D N S F D V V W S I E A G P H M PH - - V T G I T I S P E Q V R R A R E L T P A - - - E L N V R F Q L D D A L A L S F P D A S F D V V W S I E A G P H M PD - - V L G V S I S P A Q I R R A T E L T P A - - - G L S C R F E V M D A L N L Q L P D R Q F D A V W T V E A G P H M PN - - V T G I T I S P A Q V K R A K E L T P Y - - - E C K C N F K V M D A L D L K F E E G I F D G V W S V E A G A H M ND - - V L G I T I S P A Q V K R A S Q L T P E - - - G M T C Q F Q V M D A L D L K L A N G S F D A V W S V E A G P H M PD - - V V G I T I S S E Q V K R A N Q L T P K - - - E L K C H F E I M N A L N L K F E D G S F D G V W S V E A G P H I LR - - V E G I T L S P V Q C K R A A E R A R E H H L D G R A H F Q V A D A H R M P F A D G R F D L V W S L E S G E H M AT G K V V L A D I N E S M P K M G R E K L R N I G V I G N V E Y V Q A N A E A L P F P D N T F D C I T I S F G L R N V T
G V G I T I S P Q V . R A E L T P . . . F V D A L L F D . S F D . V W S . E A G P H M P
Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
190 200 210 220 230 240
D K A V N A K E L L R V V K P G G I L V V A D W N Q R - - D D R Q V P L N F W E K P V M R Q L L D Q W S H P A N A S I EN K A Q F L Q E A W R V L K P G G R L I L A T W C H R P I D P G N G P L T A D E R R H L Q A I Y D V Y C L P Y V V S L PD K A I F A Q E L L R V L K P G G K L V V A D W N Q R - - D D R Q I P L N W W E K P V M T Q L L D Q W A H P K F S S I ED K A I F A K E L M R V L K P G G I M V L A D W N Q R - - D D R Q K P L N F W E K P V M Q Q L L D Q W S H P A F S S I ED I P R S L Q E L H R V L K P G A K A A I L D F H R P - - - - - - - - S N P Q L R A F Q Q L Y L N S F V V P - V A N Y LD K V K Y G S E M M R V L K P G G I L V V A D W N Q R - - D D R Q K P L N Y W E K P V M R Q L L D Q W S H P A F S S I ED K Q Q F A K E L L R V L K P G G I L V V A D W N Q R - - D D R Q Q P L N F W E R L I M R Q L L D Q W A H P A F A S I ED K Q R F A D E L L R V L R P G G C L A A A D W N R R - - A P K D G A M N S T E R W V M R Q L L N Q W A H P E F A S I SN K T K F A D Q M L R T L R P G G Y L A L A D W N S R - - D L Q K Q P P S M I E K I I L K Q L L E Q W V H P K F I S I ND K Q R Y A D E L L R V L R P K G V L A V A D W N R R - - D Y E D G E M T S L E R W V M R Q L L D Q W A H P E F A S I KN K Q L F A D E M L R V L R P G G V L A V A D W N R R - - D Y A K K E I G F L N S L V L K Q L L N Q W S H P D F A T I YD K A Q F L R E C H R V L R P G G R F V F V T W C C R - - - - - H G A L D A R D Q K W L G A I Y R I Y H L P Y I L S I ED K D K A L R S M Y R V L K P G G R L L V L E F S K P - - - - - - - - - - - - I I E P L S K A Y D A Y S F H V L P R I GD K F A E L L R V L K P G G L . V A D W N R P I D P L N E . V M . Q L L D Q W H P F . S I
Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
250 260 270 280 290 300
G N A E N L E A T G L V E G Q V T T A D W T V P T L P A W L D T I W Q G I I R P Q G W L Q Y G I R G N I K S V R E V P TD Y E A I A R E C G - - F G E I K T A D W S V A V A P F W D R V I E S A F - D P R V L W A L G Q A G - P K I I N A A L CG F S E L L E E T G L V Q D Q V I N A D W S Q E T L P S W L H T I W V G A I R P W G W L Q Y G L P G F I K S V R E I P TG F S E L L A A T G L V E G E V I T A D W T K Q T L P S W L D S I W Q G I V R P E G L V R F G L S G F I K S L R E V P TG L K E E Y A Y I S - - - - - - P S L D R - - - - - - - - - - - - - - - - - F P - - - - - I G K E Q - I E L A R Q V G FG F S E Q I A E T G L V E G E V A T A D W T Q E T L P S W F E S I W Q G I V R P K G L I K F G F S G F I K S L R E V P TG F A E A L A A T G L V A G E V M T A D W T Q E T L P S W L D S I W Q G I V R P E G L I R F G L P G L V K S L R E V P TG F R A N L E A S P H Q R G L I S T G D W T L A T L P S W F D S I A E G L R R P W A V L G L G P K A V L Q G L R E T P TE F S S I L I N N K N S S G Q V I S S N W N S F T N P S W F D S I F E G M R R P N S I L S L G P G A I I K S I R E I P TG F R R N L L H S P F A C G T V E S D D W T R S I L P S W N D S I L E G F R R P G A V L G L G P A A L V K G F R E I P TG F R N N L S D S I Y S A G R V E T D D W T K Y T I P S W N D S I I E G I R R P N V F F D L G L G S F F K A I R E V P TS Y T Q L L G E T G - - F S G I R T T D W S D R V A R F W S L V I D S A L - E P A V L W K V I A Q G - P T V I K G A L AS L V A N D A D S Y - - - - - - R Y L A E S I R M H P - - - - - - - - - - - D Q - - - - - - - - D T L K A M M Q D A G FG F L . . G L V G V T . D W T T L P S W D S I G . R P . . . . G G . K . R E . P T
Sll0418Slr008980a16268All2121Npun3758Tery4209Tll1726Synw2141Pmm1505Pmt1785Pro1661Glr3039UbiE
310 320 330 340 350 360
I L L M R L A N G V G L C R F G M F K A V R K N A T Q A NL R L M K W G Y E R G L V R F G L L T G I K P L VI L L M R L A F G T G L C R F G M F Q A V R G E G N P V T K N R T T Q Q A I S R NL L L M R L A F G T G L C R F G M F R A L R A D T V R S S A E Q T S A I K V A Q KA V A T H Y P I A N G M M G V L V V S K FM L L M R L G F G A G L C R F G M F R A V K S N S V P V S T E T A T T E V A N AF L L M R I A F G M G L C R F G M F R A V R A E I P A V S L E P A P Q V N CL L L M H W A F A T G L M Q F G V F R L S RI L L M D W A F K K G L M E F G V Y K C R GI L L M R W A F A H G L M Q F G V F R S R DI V L M R W A F H T G L M Q F G V F R S R GM Q L M R R S Y A R G L V R F G V F A A Q K A E GE S V D Y Y N L T A G V V A L H R G Y K F. L L M R . A F . G L . R F G . F . . . V S E

305 The multiple alignment was generated by BLOSUM30 using Clustal W. Locus tags are used to indicate each organism (see also Table 6.5): Sll0418 and Slr0089, Synechocystis sp. PCC 6803; All2121, Nostoc sp. PCC 7120; Npun3758, N. punctiforme; Tery4209, T. erythraeum; Tll1726, T. elongatus BP-1; Synw2141, Synechococcus sp. WH8102; Pmm1505, P. marinus MED4; Pmt1785, P. marinus MIT9313; Pro1661, P. marinus SS120; Gll3039, G. violaceus PCC7421. 80a16268 indicates a tentative locus in Synechococcus sp. PCC 7002.

306
A.1.11. An amino acid sequence alignment of Sll0829 homologs in cyanobacteria
Sll082989a46731All3750Npun0871Tery1223Tll1604Synw1749Pmt1242Gll1483UbiE
10 20 30 40 50 60
M A T I F R - T W S Y Q Y P W V Y A L V S - - - - R L A T L N V G G E E R F H Q L P L E N L A IM A T F L R - T L S Y R Y Q W L Y D T I S - - - - R L A A L T V G G E T R F R N L A L Q G L T GM A T I F R - D L S Y R Y Q W L Y D S I S - - - - R V A A L T V G G E A R F R K L A L Q G L T IM A T I L R - D W S Y R Y Q W L Y D S I S - - - - R L A A L S V G G E A R F R Q L A L Q A L T IM A T I L R - D W S Y R Y Q W L Y D G I S - - - - R L A S L S V G G E T R F R Q L A L E G L I IM A T I L R - T W S Y Q S P W L Y D T I S - - - - A L A A I A V G G S D R L H R L A W Q D L N LM S S F L R - P L A Y R H R W I Y D L V T - - - - A V S S L S V G G V A R L R G L G L E A L G PM T S P L R - T F A Y E H R W F Y D L V T - - - - T I S A L S V G G V K R L R S L G L T A L Q D
M E A P A W N N A L A T I L R - D W S F A C A P L Y E A I T - - - - S G A A V L A G G E R H F R Q A A W Q N V S LM V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S K Y D V M N D L M S F G I H R L W K R F T I D C S GM V D M A T I L R E . S Y W L Y D I S S K Y D . A A L . V G G E R F R L A L . L .
Sll082989a46731All3750Npun0871Tery1223Tll1604Synw1749Pmt1242Gll1483UbiE
70 80 90 100 110 120
S P - - G Q K V L D L C C G G G Q A T V Y L A Q S G A T - - - V V G L D A S P - - K A L G R A K I N - - - - V P Q A T YD K - - S L K I L D L C C G A G Q T T Q F L T Q Y S D D - - - V T G L D I S P - - L A I E R A K K N - - - - V P Q A N YQ A - - N T S V L D V C C G S G Q A T Q L L V K S S Q N - - - V T G L D A S P - - L S L Q R A R Q N - - - - V P E A N YH S - - D T Q V L D L C C G S G Q T T Q F L V K I S Q N - - - V T G L D A S P - - K S L Q R A R L N - - - - V P E A S YE E - - N T Q I L D L C C G S G Q G T N F L A K Y S Q S - - - V T G L D A S P - - L S I N R A K K N - - - - V P S A K YP R - - T A V V L D L C C A H G I V T Q A L T Q A F D Q - - - V T G L D A A P - - K A I A R A R E R - - - - V P Q A T YH L N P D A A V L D L C C G S G E A A A P W L E A G Y R - - - V T G L D I S P - - R A L A L A A Q R - - - - H P A M T RK I S K G A P V L D L C C G S G E T A A P W I E A G F A - - - V T G L D L S P - - K A L A L A A E R - - - - T P Q L Q CS A - - A S S V L D L C C G P G G A T R Y L A A T G A R - - - V T G L D R S E - - G S L R H A R A R - - - - V P G A H FV R - R G Q T V L D L A G G T G D L T A K F S R L V G E T G K V V L A D I N E S M P K M G R E K L R N I G V I G N V E Y
V L D L C C G . G . T L . T G K V T G L D . S P S M L R A . N I G V V P A Y
Sll082989a46731All3750Npun0871Tery1223Tll1604Synw1749Pmt1242Gll1483UbiE
130 140 150 160 170 180
V Q G L A E D L P F G E G E F D L V H T S V A L H E M T P A Q L Q S I I S G V H R V L K P G G I F A L V D L H R P S N WV V A P A E K M P L P D Q Q F D L V H T S A A L H E M T P T Q L S Q I F Q E V Y R V L K P G G I F T F I D L H Q P T N PV E A F A E K M P F P D N Q F D I V H T S A A L H E M E P Q Q L R E I I Q E V Y R V L K P G G V F T L V D F H T P T N PV E A F A E E M P F T D N L F D V V H I S V A L H E M Q P Q Q L R K I I D E V Y R V L K P G G I F T L V D F H A P T N PV E G F A E D M P F S S N Q F D L V H T S A A L H E M N Y E Q L R Q I I Q E V Y R V L K P S G I F T F V D F H S P T N PV Q A F A E K M P F A D A T F D L V H T S M A L H E M T A A Q R D A I L A E V W R I L K P G G W F A L I D F H R P Q V PV E G L A E D P P L A D G S F A A I Q L S V A L H E F P R S D R E A V L R S C L R L L Q P G G W L V V V D L H - P A G PI E G M A E K P P L N T S Q F A A I Q I S L A L H E F S S E E R Q Q V L K A C M R L L Q P G G W L V L I D L H - P A G PV Q G R A E A M P F A G A S F D L V H T S V A L H E M E P T Q R R T I I R E V L R V L K P G G T F A L I D Y H R P T N PV Q A N A E A L P F P D N T F D C I T I S F G L R N V T D K - - D K A L R S M Y R V L K P G G R L L V L E F S K P I I EV . A E M P F D F D . V H T S . A L H E M Q L I . E V . R V L K P G G F . L . D . H P . N P
Sll082989a46731All3750Npun0871Tery1223Tll1604Synw1749Pmt1242Gll1483UbiE
190 200 210 220 230 240
L F W P P L A I F M G L F E T E T A W Q L I N T D L G S L L D Q A G F T V V R - - - - - K H L Y A G G S L Q V I Q A R AF F V P S L Y T F M F L F E T E T A W Q L I K T N L A E K L T T T G F E I H K - - - - Q - E Q Y A G G S L Q M I Q S R KI F W P G L T V F L L L F E T E T A W Q L L K T D L A G L L T E I G F E A G - - - - - Q S I L Y A G G S L Q V I Q A K KI L W P G I S L F L L L F E T E T A W Q L L K T D L A G L L T E T G F D V S - - - - - K P I L Y A G G S L Q V I Q A K KL F W P G F A M F L W L F E T E T S W K F I N T N I L E L L T T I G F N L V D T K L P L P I L Y A G G S L Q V I Q V Q KL L W P G I A L F F W L F E T E T A W Q L L Q T D L A E Q L R L Q G F T V E R - - - - - Q T Y H L G R S L Q V L H G R KW L Q L P Q Q L F C A L F E T D T A T A M L E D D L P A Q L K Q L G F S A V N - - - - - Q E L L A G Q A L Q R I T A T RC L K L P Q Q L F C A L F E T E T A L T M L Q A D L P K Q L R E M G Y S T I E - - - - - Q E L L A G R A L Q R I T A R LL Y W P G L A L F L W V F E T H T A W E L L A T D L P G L L A E C G F S T L E - - - - - Q Q L L A G G S L Q T V Q A R KP L S K A Y D A Y S F H V L P R I G S L V A N D A D S Y R Y L A E S I R M H P D Q D T L K A M M Q D A G F E S V D Y Y N. W P . . . F L F E T E T A W L L T D L . L G F . . L A G G S L Q I Q A . K
Sll082989a46731All3750Npun0871Tery1223Tll1604Synw1749Pmt1242Gll1483UbiE
250 260 270 280 290 300
N K T V NP G S E P L K N
P C A N DS A A S T SPPL T A G V V A L H R G Y K FP A H R G Y K F
The multiple alignment was generated by BLOSUM30 using Clustal W. Locus tags are used to indicate each organism (see also Table 6.5): Sll0829, Synechocystis sp. PCC 6803; All3750, Nostoc sp. PCC 7120; Npun0871, N. punctiforme; Tery1223, T. erythraeum; Tll1604, T. elongatus BP-1; Synw1749, Synechococcus sp. WH8102; Pmt1242, P. marinus MIT9313; Gll1483, G. violaceus PCC7421. 89a46731 indicates a tentative locus in Synechococcus sp. PCC 7002.

307
A.1.12. An amino acid sequence alignment of Sll0487 homologs in cyanobacteria Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
10 20 30 40 50 60
M L S N - - - S D Q H R A V Q A L Y N T Y P F P P E P L L Q E P P P G Y N W R W Q W T A A H N F CM S D P - - - Q A V S S A V A K L Y D T Y P F P P E P I L D E P P P G Y N W R W N W L A A H S F CM S D S - - - Q T V S A A V A K L Y N T Y P F P P E A L L D E P P P G Y N W R W N W L A A H N F CM K D R - - - I N I S S A V E G L Y D T F P F P P D P L T D E A P P G Y N W R W S W P V A Y S F CM T S A - - - A E I T A A V R Q L Y N T Y P F P P E P L L D E P P P G Y N W R W S W P A A Y S F CM A E P G N R D A A T P V V S A F Y D R F P F P G D P L Q D G P P P G Y N W R W C H Q S V L A A V
M S S V A Q P - - C D E A T P V V S A F Y D R F P Y P G D P L Q D G P P P G Y N W R W S V D M A Y A V CM D I T R A V Q S L Y E R Y P F P P D P L S D K P P P G W N W R W S Y S F A H S F C
M V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S K Y D V M N D L M S F G I H R L W K R F T I D - - CM V D K S Q E T M . . A V L Y . T Y P F P P D P L D E P P P G Y N W R W W . A F C
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
70 80 90 100 110 120
L G R R P - - - - A N Q K V R I L D A G C G T G V G T E Y L V H L N P E - A E V H A V D I S E G A L A V A Q T R L Q K ST G R K P - - - - A K Q D I R I L D A G C G S G V G T E Y L V H L N P Q - A Q V V G I D L S A G T L A V A K V R C Q R ST G Q K P - - - - Q K Q D I R I L D A G C G T G V S T E Y L V H L N P Q - A S V V G I D L S T G A L D V A K E R C Q R ST G Q K P - - - - K K Q D I R I L D A G C G T G S S T D Y L V H L N P Q - A S V V G I D L S S G A L R V A K E R C S R ST G R Y P - - - - S Q L D V A I L D A G C G T G V G T E Y L A H L N P Q - A K I T A L D L S E A A L A I A C E R C R R SH G A V P - - - A G T V R P R I L D A G C G T G V S T D Y L C H L N P G - A E V L G I D I S E G A L A V A R E R L Q R ST G S V V R K C A G S E P L K I L D A G C G T G V S T D Y L A H L N P G - A E I L A V D I S L G A L E V A R E R L Q R SR G W R P - - - - E P R P I R I L D A G C G T G V S T E Y L A V Q N P E - A Q V T A I D I S E A S L A L A K R R C A G HS G V R R - - - - - - - G Q T V L D L A G G T G D L T A K F S R L V G E T G K V V L A D I N E S M P K M G R E K L R N IT G . P R K C A . R I L D A G C G T G V S T E Y L . H L N P T A V . . I D I S E G A L A V A . E R C Q R S
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
130 140 150 160 170 180
G V V C D - - R V H F H H L S L E N L A H L P G Q F D Y I N S V G V L H H L P D P V A G I Q A V A E K L A P G G L F H IG A N - - - - R V E F H H L S L Y D V E Q L P G E F D L I N C V G V L H H L P D P I R G I Q A L A K K L A P G G L M H IG A N - - - - R V E F H H L S L F D V E Q L P G E F D L I N C V G V L H H T S D P I R G I Q A L A Q K L A P G G L M H IG A S - - - - R V D F H H L S I Y D V G R L E G E F K F I N C V G V L H H L P D P I R G I Q N L A L K L A P G G I M H IG A T - - - - N V Q F H H L S L E E V A Q L G Q T F Q M I N C V G V L H H L P E P Q R G I Q A L A D V L A P G G I V H IG A A A Q V S Q L R Q E Q R S L L D L E S E - G P F D Y I N S V G V L H H L D Q P E S G L R S L A G R L A P D G L L H LG G N D Q - A H V R I E N Q S L L E L E D E - G Q F D Y I N S V G A L H H L R Q P E A G L K A L A S R L K P G A L L H LA N V - - - - - - Q F A Q M S L T D A G D L E G Q F D L I N C V G V L H H L A D P K A G L A A L A A K L A P G G L L H IG V I G - - - N V E Y V Q A N A E A L P F P D N T F D C I T I S F G L R N V T D K D K A L R S M Y R V L K P G G R L L VG A Q V . V F H H L S L D . L G F D I N C V G V L H H L D P . G I Q A L A K L A P G G L . H I
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
190 200 210 220 230 240
F V Y A E I G R W E I Q L M Q K A I A I L Q G K K R G D Y Q D G V A V G R E I F A S L P E Y N R L V K R E K E R W S L EF V Y G E L G R R E I Q L M Q K A I A L L Q N E K Q G D Y R D G V Q V G R K I F A S L P E N N R L V K R E K E R W A M EF V Y G E L G R W E I Q L M Q K A I A L L Q G D K K G D Y R D G V Q V G R Q I F A S L P E N N R I V K Y E K Q R W S L EF V Y A E L G R W E I R L M Q Q A I A I L Q G N E R G N Y K D G V K I G R Q L F A S L P E N N R I V K Y E K E R W S W EF V Y G E Y G R W E I K L M Q Q A I A L L Q G S D R H N Y R E G V A I G R Q L F A A L P E N N R L K K R E Q E R W S L EF L Y A D A G R W E I H R T Q Q A L T L L D - - - V G T G R E G L R L G R E L L A S L P E G N R L A R H H C E R W A V DF L Y A E A G R W E I H R I Q R F L S S L G - - - V G T D E Q G L R L A R Q L F E I L P E H N R L R L N H E Q R W A I DF V Y G E L G R W E I R L M Q E A I R L L R - D G R D P L A D G L K V G R A L F E A L P E D N R L K S A D R - R W V L EL E F S K P - - - I I E P L S K A Y D A Y S - - - - - - - - - - - - - - - - - F H V L P R I G S L V A N D A D S Y R Y LF V Y . E . G R W E I . L M Q . A I A L L Q G . G Y . D G V . G R L F A S L P E N R L V K E . E R W . E
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
250 260 270 280 290 300
N H R D E S F A D M Y V H P Q E T D Y N I D T L F E L I D S A G L E F L G F S N P D Y W Q L D R L L G K A P D L M E R AN Q R D E C F A D M Y V H P Q E V D Y N I D T L F E L I D A S G L E F I G F S N P S F W N L D R L L G K A P E L I E R SN H K D E H F A D M Y V H P Q E I D Y N I E T L F E L I D A S G L E F I G F S N P S F W D L E R L L S K A P E L V E R AN K K D E C F A D M Y V H P Q E I H Y N I D T L F E L I E A S G L E F L G F S N W E Y W D L G R L L G K S P E L I E R AN Q R D E C F A D M Y V H P Q E I D Y T I A S L F E L I R A S G L E F I G F S N P Q V W Q L E R L L G S Q P E L L Q R AC A A D A N F A D M Y L H P Q E T S Y D L Q R L F A F I E A A D L H F A G F S N A E V W D P A R L L N G - - E L L E R AC V A D V N F A D M Y L H P Q E T S Y N L E R L F A F V A S A D L E F V G F S N P K V W D P V R L L Q G - - E L L E M AN H H D T C F V D M Y V Q V Q E I R Y T I P T L F D L I A S S G L G F L G F S N P G F W R L E R L V G K A P W L L E R AA E S - - - - - - I R M H P D Q - - - - - D T L K A M M Q D A G F E S V D Y Y N - - - - - - - - L T A G - - - - - - - VN . D E F A D M Y V H P Q E . Y N I . T L F E L I . A S G L E F . G F S N P . W L . R L L G K A P E L . E R A
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
310 320 330 340 350 360
K D L S E K E R Y R L I E L L D P E - I T H Y E F F L A K P P I Q I S D W T D D Q T L L K A L P N L H P C L T G W P S RE K L S D R Q R Y R L I E L L D P E - V T H Y E F F L G R P P I K K A D W S N D N A L L Q A I P E L N P C I D G F P S QK E L S D R Q L Y R L I E L L D P E - V T H Y E F F L G R P P L V K T D W S D D K A L L A A I P E L N P C I E G F P S QI D L N D Q E R Y R L I E L L D P Q - I S H Y E F F L G R P P L Q K V D W S H D E L L K A A I P E L S P C I Q G W P S EQ Q L D P I Q Q Y Q L I E C L D P E A M T H F E F F L A K P P L P R H D W A A D R E L L A A I P W R H P C I D G W P G AQ A L P Q R Q Q W L L V E Q L D P N - I S H F E F F L S S Q P V Q P A S W S - D A A L Q A A K G L R Q P C L W G E P - DL A L P Q R Q Q W Q L I E Q L D P D - I S H F E F F L S K G P Q A S L G W T D D Q E L L A A T G K L S I C L W G W P G QA T L S D K E R F R L I E L L D P V - V A H Y E F F L G A P P L E R L E L D D D - R L D A A L P V R S P Y L Y P W P E RV A L H R G - - - - - - - - Y K F
L . . Q Y R L I E L L D P . A . . H Y E F F L . . P P . . . D W . D D L L A A . P L P C . G W P S
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
370 380 390 400 410 420
S L F D Y N Y Q P L E L T E E E F K F L Q A V E A Q T E - - I Q A N V E K V L T D L G D R S I D L A L V R S L Q T R Q LC I F N Y D Y Q I V N L S A L E F E F L Q K C D G - - - - - - N S T V A E I L V S V Q - - - L S L D E V R S L I Q Q Q LC F F N Y N Y Q I V N L S E P E F E F M Q K C D A - - - - - - N S T V A E I L A D V Q - - - L G L D E V R K L L Q Q Q LN I F D G D Y R L V N L S R P E Y E F L E A C T Q G V S N K S Q S T V G E I L T D V P - - - V G V E T V R S L Q S R L LC V F N C H Y E V I H L N Q A E Q E F L N A A S G - - - - - - K A T V A E L L A Q Q P D - - F N L A E V R S L L A R H LP I L D R N M Q P L Q L S D A E R Q L L R R V D E Q - - - - - P N T P L G A M A E - - - - - - - P A V I R D L A A R Q LS L L D F E M A P L E L S V E Q V E L L K A I E A T - - - - - P D T A L G C L P L G W D S K R I S S L A R D L Q R R Q VQ V F D H E Y E L V E L S E A E V A F L Q R C N G - - - - - - R Q T L G Q L R G D I A Q - - - A R I R E L L E H F L L Q
. F D Y . . L S E E F L . . . N K T V . . L . . . D . V R L R Q L
Sll0487All0012Npun5885Tery3855Tlr1948Synw0487Pmt1474Glr1180UbiE
430 440 450 460 470 480
L T L G N SV M L I P G A K G Y A SI L L A P AI L L S P N S G Y HL L L G V K EL L L K AL L L S P G N V A S TL A P P P G L L S S K M R V
L L L P R V
The multiple alignment was generated by BLOSUM30 using Clustal W. Locus tags are used to indicate each organism (see also Table 6.5): Sll0487, Synechocystis sp. PCC 6803; All0012, Nostoc sp. PCC 7120; Npun5885, N. punctiforme; Tery3855, T. erythraeum; Tlr1948, T. elongatus BP-1; Synw0487, Synechococcus sp. WH8102; P. marinus MIT1474; Glr1180, G. violaceus PCC7421.

308
A.1.13. An amino acid sequence alignment of Slr1039 homologs in cyanobacteria Slr103991c179267All3016Tlr1953Synw1148Pmm0662Pmt0792Pro0816Glr3970UbiE
10 20 30 40 50 60
M L E R I L E P E - - - - V M D T E A E A M D Y D A M D F Q A V N L - - A F A Q R A C Q L G P S K - - - - - - - - - -M A L T R I L E P E - - - - V M D D P A E A I A Y D A M D F R A V N Q - - D F A D L V V T T Y S Q E T - - - - - - - - -
M H R Q L E P E - - - - V M D S W E E A S E Y D A M D F T E V N N - - A F A E E A V A C G L S E H - - - - - - - - -M P L P R I L E P E - - - - V M E T P E T A A A Y D A M D F T A V N T - - A F C D D L A A V L P R V D Q P - - - - - - -
M Q R L P E P E - - - - L M V D P A Q V L A Y A A A D F S G G D Q - - R T L D R I E A L L S S A S G R A A - - - - -M E R T V E P E - - - - L M E R E D Q V D S Y A K A D F S E G E N - - N L I S Q I N H Y L I K N N I Y L S - - - - -
M P E P E - - - - L M D D P L Q A R A Y A E A D F S C G D E - - A L I Q R L Q E Y L I S L C K I P S - - - - -M Q R V T E P E - - - - L M L A P D Q V Q A Y A E A D F S S S E A - - S M L S E V D L L I R D R G M H I D - - - - -
M L E R V L E P E - - - - V M D S Q A E A E A Y D A M D H G E V N A - - R F V A D V L A L G P A D G - - - - - - - - -M V D K S Q E T T H F G F Q T V A K E Q K A D M V A H V F H S V A S K Y D V M N D L M S F G I H R L W K R F T I D C S
M M R . . E P E H F G F . M . A A Y A D F . V K Y . . . T I D C S
Slr103991c179267All3016Tlr1953Synw1148Pmm0662Pmt0792Pro0816Glr3970UbiE
70 80 90 100 110 120
- - - - - A T V L D A G T G T A R I P I L L A Q L R P A - W Q I T A I D F A R S M L A I A K E N V I - - - A A G C E T Q- - - - - A F V L D L G T G T A Q I P I L L G Q Q R P Q - W Q I K G T D L A Q S M L A L G Q K N V V - - - A A G L T A Q- - - - - G L V L D A G T G T A R I P V L I C Q K R P Q - W Q L V A I D M A E N M L Q I A T Q H V Q - - - Q S G L Q E H- - - - - L K V L D V G T G N G R I L Q L L H Q R Y P H - W Q L T G I D L S P A M L A I A R H H S - - - - - - - - - P Q- - S D P A V I L D L G C G P G N I S L P L A K R F P E - S Q V I G V D G S R A M L Q V A R D R - - - - - A N Q Q G L S- - E K - E L I V D L G C G P G N I S E K L S T K W P N - A N V I G I D G S K E M I R I A E L N K K N S L N R S R L K N- - P G - S M I V D L G C G P G N I C E R L Q R L W P E - V M V L G I D G A Q A M L D H A L Q R Q K A - - M A A E L K R- - R K - K L I V D L G C G P G N I T Q R L A D Q W P E - A K V L G L D D S P E M L M Y A R K K Q K E - - K I S F L E R- - - - - - P W L D L G T G P A H L P V L L A S E R P D - I R I T A V D L A A P M L A I A R R R V E - - - A A G L A G RG V R R G Q T V L D L A G G T G D L T A K F S R L V G E T G K V V L A D I N E S M P K M G R E K L R - - - N I G V I G NG V . L D L G G G I . L P T . . . G . D . M L . A . . S L . G .
Slr103991c179267All3016Tlr1953Synw1148Pmm0662Pmt0792Pro0816Glr3970UbiE
130 140 150 160 170 180
I C L E F V D A K Q L P Y A N G S F D G V I A - - - - N S L C H H L P R P L D F L R E V K R V L K P H G F L L I R D L II E L V Y A D A K N L P W P D Q S F D V I I S - - - - N S L I H H L P D P L P C F Q E M R R L L K P Q G N I I V R D L FI R L E L V D A K R L P Y E D G I F D L V V S - - - - N S L V H H L P D P L P F F A E I K R V C K P Q G G I F I R D L LL N F I E G D A K T L P F A A A S F D V V I S - - - - N S L V H H L A D P Q P A L G E M L R V L R P Q G T L F I R D L CI D L R C S T L Q D L A L E P V D L I V S - - - - - - N S L L H H L H E P G L L W G L T R E L A A P G C R V L H R D L RL R Y I C A D I K S L K S S D I S F E K N I S L L V S N S L I H H I T Y L D D F F N C I K R L S S D L T I N F H K D L KL T Y R C S N L S S L V N Q C V D L K C S A A L V V S N S L L H H L H D P N L L W Q V T K Y L A A P G A F V F H R D L RI T Y K K I N I S Q I A N G D F P F R E C A D L V V S N S L I H H I H D P L I F I K A L K N I S K H G A I H F H R D L RI T L V H G D A K A P A L G A A R F A C V F S - - - - N S L V H H L P Q P A P F W Q A C A R L L A P G G V L F V R D L AV E Y V Q A N A E A L P F P D N T F D C I T I - - - - S F G L R N V T D K D K A L R S M Y R V L K P G G R L L V L E F SI . D A K L . F . . . S L V V S N S L . H H L D P . . . R . . . P . G . F . R D L
Slr103991c179267All3016Tlr1953Synw1148Pmm0662Pmt0792Pro0816Glr3970UbiE
190 200 210 220 230 240
R P E S P E K L A D F V T A I G S D Y N D Q - Q R K L F A D S L H A S L T I A E I Q D Y L Q H L G W V G D D G P N L E LR P D S T A A I D Q I V A A A E G P G F D A R Q R Q L F W D S L H A A F T V P E I E A I A T E A G L T K - - - - - - - AR P E D E A T M N A L V A S I G N E Y D D Y - Q K K L F R D S L H A A L T L D E V N Q L I I T A G L T G - - - - - - - VR P Q T L A E L R A L V E Q Y A G Q D T P Q - Q Q K L F A D S L H A A L T L R E M R H L L R T L P Q S P Q - - - - - R WR P A A - L A E V H R L Q Q L H C F D A P A V L I Q D F C A S L V A A F T P E E V Q Q Q L A L A E L D G - - - - - - - LR P I D - E Q S A L Y L K E K C G E K Y N E I L T N D Y Y A S L K A S Y T S K E L K D F I F E N K L S S - - - - - - - LR P L S - T K H A I A L Q Q K H M Q E A P S I L I R D Y L A S L H A A F T L E E V Q S Q L V H A G L D H - - - - - - - LR P S S - L E E A L A I K Q K H L P T A P S V M E K D F I A S L K A A Y T S R E I S Q Y L K D S E L N S - - - - - - - FR P D S L S R R D A L V E Q Y A A G C D D D - Q R R L F A E S L Q A A L I P A E V A S Q A R A A G L T G - - - - - - - AK P I I - E P L S K A Y D A Y S F H V L P R I G S L V A N D A D S Y R Y L A E S I R M H P D Q D T L K A M M Q - - - - DR P . . . . . . F . S L . A A . T . E . . . L P N L .
Slr103991c179267All3016Tlr1953Synw1148Pmm0662Pmt0792Pro0816Glr3970UbiE
250 260 270 280 290 300
H L Y Q S S D R H W T L E K K F Q S S T I V Q RQ V Y Q S S E R H W T L I F P Q P NE I Y Q S S D R H W T A K R S W T NQ A I L T S D R H W T V V A Q RT V E M E D D R Y L V V S G L V DE V F E E G D Q Y L V I Y G K VH V I E V E D R Y L D V F G T IK V V E V D D R Y L D V F G V I IQ V A M S S D R H W T L I R R G A VA G F E S V D Y Y N L T A G V V A L H R G Y K F
V . D R . . .
The multiple alignment was generated by BLOSUM30 using Clustal W. Locus tags are used to indicate each organism (see also Table 6.5): Slr1039, Syechocystis sp. PCC 6803; All3016, Nostoc sp. PCC 7120; Tlr1953, T. elongatus BP-1; Synw1148, Synechococcus sp. WH8102; Pmm0662, P. marinus MED4; Pmt0792, P. marinus MIT9313; Pro0816, P. marinus SS120; Glr3970, G. violaceus PCC7421. 91a179267 indicates a tentative locus in Synechococcus sp. PCC 7002.

309
A.2.1. Polypeptide composition of isolated PS I complexes isolated from the wild-type,
menG, menF, rubA, rubA menG, and rubA menB mutants of Synechococcus sp. PCC
7002
M and T indicate monomeric and trimeric PS I complexes isolated from the respective wild type (WT) and mutant strains. SDS-PAGE gel containing 16% acrylamide and 6 M urea were used to resolve the polypeptides (electrophoresis at 80 mV for 12 h at room temperature). Small blue, yellow, and red arrows indicate polypeptide bands that are not detectable in the wild-type PS I trimeric forms. The polypeptide bands were visualized after silver staining.

310
Presence of all the polypeptides was confirmed in the PS I complexes isolated
from menF and menG mutants. Immunoblotting analysis showed that PsaA and PsaB
polypeptides were present in the PS I complexes isolated from the rubA menB and rubA
menB double mutants, while PsaC, PsaD, and PsaE were absent. The absence of the latter
three polypeptides is expected as a result of the inactivation of the rubA gene, as
previously reported [Shen G, Antonkine ML, van der Est A, Vassiliev IR, Brettel K, Bittl
R, Zech SG, Zhao J, Stehlik D, Bryant DA, Golbeck JH (2002) J Biol Chem 277: 20355-
20366]. In the rubA menB mutant, no PsaL was detectable when the isolated PSI
complexes were analyzed, while it was detectable when thylakoid membranes was
analyzed. This suggests that PsaL was lost during the preparation of PS I complexes. The
absence of PsaL is also visible in the SDS-PAGE gel (a band immediately below PsaF is
missing). It has been reported that PsaL is required for the formation of trimeric forms of
PS I complexes [Chitnis VP and Chitnis PR (1993) FEBS Lett 336: 330-334]. Indeed,
after the first sucrose-density gradient centrifugation, most of the chlorophyll (an thus, PS
I complexes) was collected in the PS I monomeric fraction from the rubA menB mutant.
311
A.2.2. Illustration of genetically engineered PS I in cyanobacteria.
Strain names are shown below the respective PS I complexes. WT, wild-type strains of Synechocystis sp. PCC 6803 (containing PhyQ) and Synechococcus sp. PCC 7002 (containing MQ-4); DMQ, demethylmenaquinone-4; DphQ, demethylphylloquinone; DMPBQ, 2,3-dimethyl-6-phytyl-1,4-benzoquinone; PQ, plastoquinone-9; AQ, 9,10-anthraquinone. The PS I core is shown in green, while the PsaC subunit is shown in blue. P700 and A0 are indicated by purple circles, A1 is indicated by red diamonds, and [4Fe-4S] (FX, FA, FB) clusters are indicated by yellow squares. All the PS I complexes shown here have been generated, except for the DMPBQ-containing complexes, which are expected to be produced in a menF cyclase (in α-tocopherol biosynthetic pathway) double mutant that is yet to be constructed. For constructions of the mutants, see Chapter 2 and 3. The rubA menG mutant in Synechococcus sp. PCC 7002 was generated by transforming the rubA mutant with the menG::accC1 construct (see the construction of the rubA menB double mutant described in Chapter 2). The PS I complex devoid of quinone and [4Fe-4S] clusters has been recently generated by treating the PS I complexes isolated from the rubA menB mutant with AQ followed by extensive washing (Y. Sakuragi, B. Zybailov, G. Shen, R. Balasubramanian, B.A. Diner, I. Karygina, Y. Pushkar, D. Stehlik, D.A. Bryant, and J.H. Golbeck, unpublished).

CURRICULUM VITAE
YUMIKO SAKURAGI Date of Birth: April 2, 1973 Nationality: Japanese Place of Birth: Tokyo, Japan
EDUCATION
1996 B. S., Biology, Department of Biology, Tokyo Metropolitan University, Japan
1999 M. S., Biology, Department of Biology, Tokyo Metropolitan University, Japan
2004 Ph.D. in Biochemistry, Microbiology, and Molecular Biology, Department of Biochemistry and Molecular Biology, The Pennsylvania State University, USA
HONORS
1996 Danish Government Scholarship, Denmark
1999 Braddock Graduate Fellowship, The Pennsylvania State University
2000 First place in the competition for poster presentation, American Association for Microbiology, Allegheny Branch Meeting, Pennsylvania, USA
2001 Invited speaker at the 7th Cyanobacterial Workshop, California, USA
2002 Center for Biomolecular Structure & Function Travel Award, The Pennsylvania State University
2003 Second place in the competition for oral presentation in Environmental Engineering, Ecology and Soil Science, 6th Annual Environmental Chemistry Symposium, The Pennsylvania State University
2003 Invited speaker at the 11th International Symposium on Phototrophic Prokaryotes, Tokyo, Japan
2003 Alumni Association Dissertation Award, The Pennsylvania State University
PROFESSIONAL MEMBERSHIPS 2000 American Society of Microbiology
2002-2004
American Association for the Advancement of Science
Related Documents











