Journal of Chemical Ecology, Vol. 16, No. 4, 1990 CUTICULAR HYDROCARBONS OF EIGHT SPECIES NORTH AMERICAN CONE BEETLES, Conophthorus HOPKINS 1 OF MARION PAGE, 2 LORI J. NELSON, 2 MICHAEL I. HAVERTY, 2 and GARY J. BLOMQUIST 3 2pacific Southwest Forest and Range Experiment Station Forest Service, U.S. Department of Agriculture Berkeley, California 94701 3Department of Biochemistry University of Nevada/Reno Reno, Nevada 89557-0014 (Received March 13, 1989; accepted June 26, 1989) Abstract--A study to determine the degree of similarity and/or diversity among eight of the 15 described species of Conophthorus is reported. Cu- ticular hydrocarbons were evaluated for C. conicolens, C. ponderosae, C. cembroides, C. edulis, C. radiatae, C. coniperda, C. resinosae, and C. banksianae. Seventy-eight individual and isomeric mixtures of hydrocarbons were identified by gas chromatography-mass spectrometry, including n- alkanes, alkenes, atkadienes, 2- or 4-methylalkanes, 3-methylalkancs, and single-component and isomeric mixtures of internally branched mono-, di-, and trimethylalkanes. Differences in alkenes and mono-, di-, and trimeth- ylalkanes can be used easily to separate the eight species, Conophthorus con- icolens and C. ponderosae contain the most complex blends. Hydrocarbon patterns in three geographically separated populations of C. ponderosae, each from a different host, are qualitatively identical with the exception of a homologous series of 3,7-dimethylalkanes from adults collected from Pinus lambertiana cones. The latter could comprise a sibling species. Hydrocarbon mixtures of two eastern species, C. resinosae and C. banksianae, are quali- tatively identical, supporting the suspicion that C. banksianae may not be a valid species. Closely related C. cembroides and C. edulis have similar com- binations of hydrocarbons except for a unique and abundant alkene (C27:~) in C. edulis and two dimethyhexacosanes in C. cembriodes. JColeoptera: Scolytidae. 1173

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Chemical Ecology, Vol. 16, No. 4, 1990
CUTICULAR HYDROCARBONS OF EIGHT SPECIES NORTH AMERICAN CONE BEETLES,
Conophthorus HOPKINS 1
OF
M A R I O N P A G E , 2 L O R I J. N E L S O N , 2 M I C H A E L I. H A V E R T Y , 2 and
G A R Y J. B L O M Q U I S T 3
2pacific Southwest Forest and Range Experiment Station Forest Service, U.S. Department of Agriculture
Berkeley, California 94701
3Department of Biochemistry University of Nevada/Reno Reno, Nevada 89557-0014
(Received March 13, 1989; accepted June 26, 1989)
Abstract--A study to determine the degree of similarity and/or diversity among eight of the 15 described species of Conophthorus is reported. Cu- ticular hydrocarbons were evaluated for C. conicolens, C. ponderosae, C. cembroides, C. edulis, C. radiatae, C. coniperda, C. resinosae, and C. banksianae. Seventy-eight individual and isomeric mixtures of hydrocarbons were identified by gas chromatography-mass spectrometry, including n- alkanes, alkenes, atkadienes, 2- or 4-methylalkanes, 3-methylalkancs, and single-component and isomeric mixtures of internally branched mono-, di-, and trimethylalkanes. Differences in alkenes and mono-, di-, and trimeth- ylalkanes can be used easily to separate the eight species, Conophthorus con- icolens and C. ponderosae contain the most complex blends. Hydrocarbon patterns in three geographically separated populations of C. ponderosae, each from a different host, are qualitatively identical with the exception of a homologous series of 3,7-dimethylalkanes from adults collected from Pinus lambertiana cones. The latter could comprise a sibling species. Hydrocarbon mixtures of two eastern species, C. resinosae and C. banksianae, are quali- tatively identical, supporting the suspicion that C. banksianae may not be a valid species. Closely related C. cembroides and C. edulis have similar com- binations of hydrocarbons except for a unique and abundant alkene (C27: ~) in C. edulis and two dimethyhexacosanes in C. cembriodes.
J Coleoptera: Scolytidae.
1173

1174 PAGE ET AL.
Key Words--Cuticular lipids, chemotaxouomy, methyl-branched hydrocar- bons, mass spectra, Pinus species, Coleoptera, Conophthorus, Scolytidae, insect integument.
INTRODUCTION
Beetles of the genus Conophthorus Hopkins (family Scolytidae) attack and kill female cones of Pinus species during the second year of cone development. In severe infestations up to three-fourths of the cones can be destroyed (Furniss and Carolin, 1977). Such sizable reductions in seed crops adversely affect nat- ural and managed reforestation. Most species of the genus are monophagous; the most dramatic exception is Conophthorus ponderosae Hopkins. This poly- phagous species reportedly breeds in cones of 8-13 species of Pinus and ranges from northern British Columbia to southern Mexico (Hedlin et al., 1980; Wood, 1982).
Wood (1982) synonomized C. ponderosae with five other species of Con- ophthorus: C. scopulorum Hopkins, C. contortae Hopkins, C. monticolae Hop- kins, C. flexilis Hopkins, and C. lambertianae Hopkins. Previous species diagnoses were generally made on the basis of host, geography, and morpho- logical features. "Taxonomic characters in Conophthorus species are so few and poorly developed that a search for non-conventional approaches is needed in order to find a means for the accurate identification of species" (S. L. Wood, personal communication). Wood (1982) found the collected material difficult to interpret. Perhaps C. ponderosae is in the process of speciation or it comprises two or more sibling species awaiting discovery. Wood (1982) candidly admits " . . . material from five-needle pines could be different."
Future control methods for Conophthorus will likely rely on mate-finding and host-acceptance behaviors, which are presumably under genetic control and are assumed to be species specific. Thus proper identification and understanding of the biology of Conophthorus species are fundamental to development of sound pest management programs. Since many of the Conophthorus species are dif- ficult to separate morphologically, we decided to evaluate cuticular hydrocar- bons as another set of taxonomic characters.
In this paper we report the identification of the hydrocarbon components in the cuticular wax of eight species of Conophthorus. This is the first step in determining the degree of similarity and/or diversity of cuticular hydrocarbon patterns among the species of this genus.
METHODS AND MATERIALS
Adult cone beetles characterized in this paper were collected as overwin- tering adults from infested cones or twig terminals. C. ponderosae was col- lected from three different species of pine: sugar pine, Pinus lambertiana

CUTICULAR HYDROCARBONS OF CONE BEETLES 1175
Dougl.; ponderosa pine, P. ponderosae Dougl. ex Laws.; and western white pine, P. monticola Dougl. All other Conophthorus were collected from a single host: C. conicolens Wood from Chihuahua pine, P. leoiphylla var. chihuahua (Engelm.) Shaw; C. cembroides Wood from Mexican pinyon pine, P. cem- broides Zucc.; C. edulis Hopkins from pinyon pine, P. edulis Engelm.; C. coniperda (Schwartz) from eastern white pine, P. strobus L.; C. radiatae Hop- kins from Monterey pine, P. radiata D. Don; C. banksianae McPherson from Jack pine, P. banksiana Lamb.; and C. resinosae Hopkins from red pine, P. resinosa Ait.
Pine cones infested with Conophthorus are not always easy to collect. Cones of sugar pine, western white pine, and eastern white pine fall to the ground at the end of the summer. Overwintering, adult Conophthorus remain in the cones on the ground and emerge as parent adults in the spring. Infested cones of these species are relatively easy to collect; cones are simply picked up off the ground and placed in a collection bag. Cones of Monterey pine, pon- derosa pine, and Chihuahua pine are difficult to collect. Even after they are infested, they are retained in the crown of the tree for one or more seasons. By the time trees present their infested cones to the biologist (i.e., drop them on the ground), the beetles have already emerged. Cones are usually borne in the upper portions of the crown of trees > 20 m in height. Biologists must resort to the use of a pole pruner to cut the cones down, a rifle to shoot the cones down, or we must collect from sexually mature, small trees or downed, mature trees in logging operations. Cones of the pinyon pines are relatively easy to collect; the trees are short (< 6 m in height) and bushy, and trees with cones usually have them distributed throughout the crown.
Hydrocarbon mixtures of Conophthorus reported in this paper represent data obtained from adult beetles that emerged from cones collected from the following locations: sugar pine from the Eldorado National Forest near Placer- ville, California; ponderosa pine from the Sierra National Forest ca. 40 km south of Yosemite National Park, California; western white pine from Sand- point, Idaho; Chihuahua pine and Mexican pinyon pine cones from the Chirp cahua Mountains in southern Arizona; pinyon pine from Prescott, Arizona and the Chiricahua Mountains; eastern white pine from Murphy, North Carolina and Petawawa, Ontario, Canada; Monterey pine from Albany and Aptos, Cal- ifornia; jack pine from Aubrey Falls, Ontario, Canada, and red pine from Little Rapids, Ontario, Canada.
Infested cones from the western pine species were collected by three of the authors; cones from the eastern species were supplied by colleagues. In addi- tion, we have examined cuticular hydrocarbons from museum specimens. Some of these beetles were collected before 1920; hydrocarbon patterns of these spec- imens are, remarkably, qualitatively identical to those collected and extracted by the authors. Hydrocarbon patterns of many of the Conophthorus species presented in this paper are representative of additional beetle collections made by the authors and of museum specimens.

1176 PAGE ET AL.
Beetles were allowed to emerge from the cones or twigs. One to three days after emergence beetles were frozen and held at - 2 0 ~ until their hydrocarbons were extracted. Beetles were removed from the freezer and allowed to warm to ambient temperature. Cuticular lipids were then extracted by immersing 15-50 beetles as a group once in 10 ml of hexane for 10 min. Sexes were not separated; males and females were extracted together. Extracted beetles were prepared as voucher specimens and submitted to the U.S. National Museum, Smithsonian Institution, Washington, D.C. After beetles were extracted, hydrocarbons were separated from other components by pipetting the 10-ml extract and an addi- tional 8 ml of hexane through 3 cm of activated BioSil-A in Pasteur pipet mini- columns (Blailock et al., 1976). All hydrocarbon extracts were evaporated to dryness under a stream of nitrogen and redissolved in 30/zl of hexane for anal- ysis.
Gas chromatography-mass spectrometry (GC-MS) analyses were per- formed on Hewlett Packard 5890 gas chromatograph equipped with a Hewlett Packard 5970B Mass Selective Detector, which was interfaced with a Hewlett Packard Chemstation computer. The GC-MS system was equipped with a fused silica capillary column (30 m • 0.2 mm ID, HP-1) and operated in split mode (with a split ratio of 20 : 1). Each mixture was analyzed by a temperature pro- gram from 200~ to 320 ~ at 3~ with a final hold of 20 min. Electron impact (El) mass spectra were obtained at 70 eV. Retention times and mass spectra of extracted n-alkanes were compared with external standards. Alkenes and methyl-branched alkanes were tentatively identified by calculating their equivalent chain lengths (ECL) (Nelson and Sukkestad, 1970; Jackson and Blomquist, 1976). Mass spectra of alkenes and methlyalkanes were interpreted as described by Nelson et al. (1972), Nelson (1978), Pomonis et al. (1978), and Blomquist et al. (1987).
RESULTS
Data presented in this report are the results of pooled samples (mixed sexes), from numerous cones, from several trees of the same species, from any location. In preliminary studies with other scolytid beetles, Dendroctonus brev- icomis LeConte and D. frontalis Zimmerman, we determined that the hydro- carbon blends of pupae and callow adults are incomplete when compared to adults. However, hydrocarbon mixtures of fully tanned, emerged adults are fixed and do not change over the short lifetime of the beetles. In early, unpub- lished studies of C. ponderosae, we determined that there were no qualitative differences within beetles collected from the same host from several disparate geographical locations. Furthermore, we examined groups of males and females of C. banksianae and C. resinosae and found no qualitative differences in their hydrocarbon profiles. Therefore, we feel justified in presenting results of pooled samples of each species of Conophthorus.

CUTICULAR HYDROCARBONS OF CONE BEETLES 1177
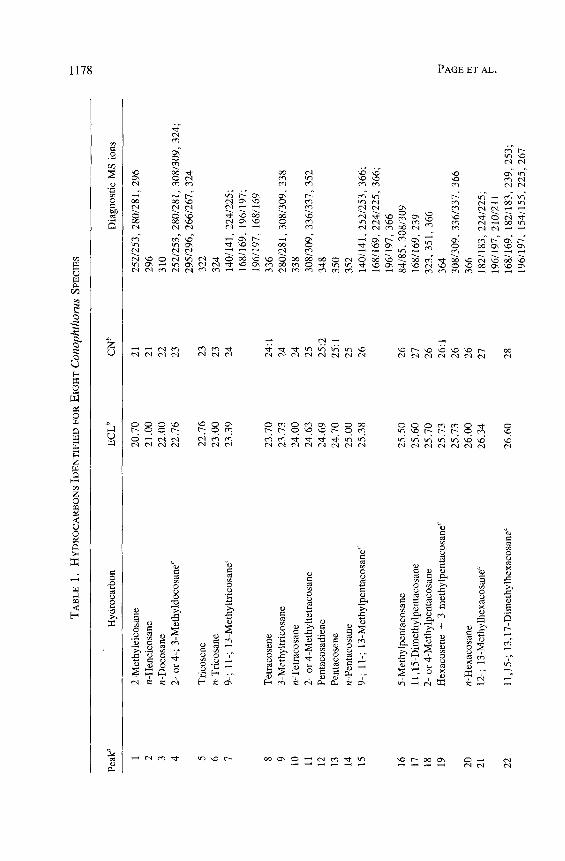
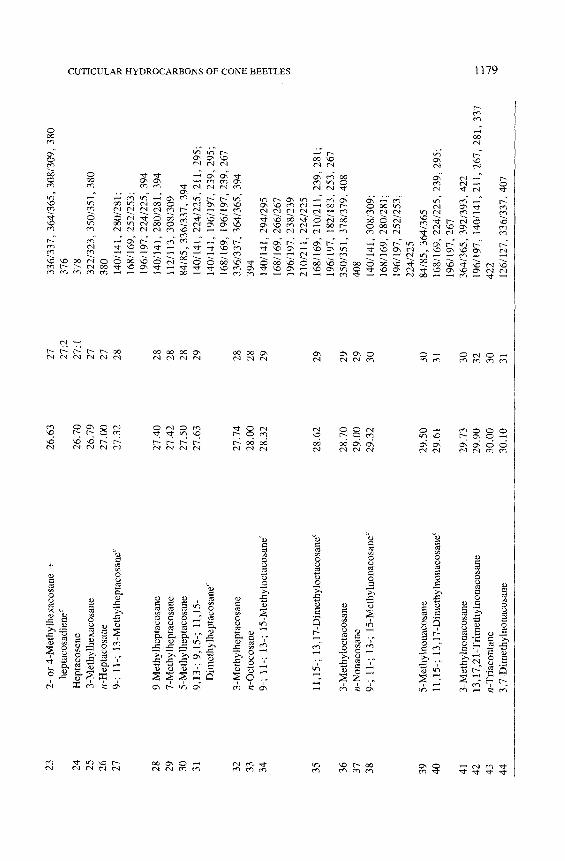
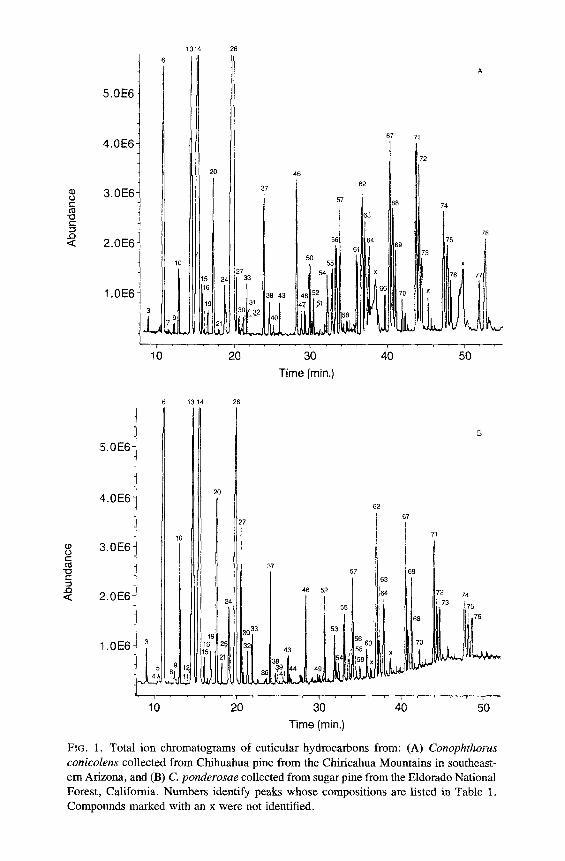
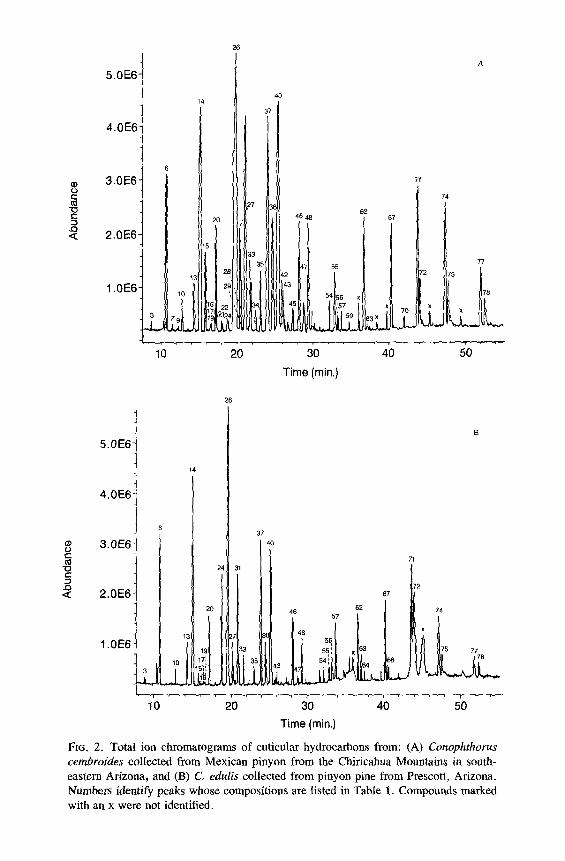
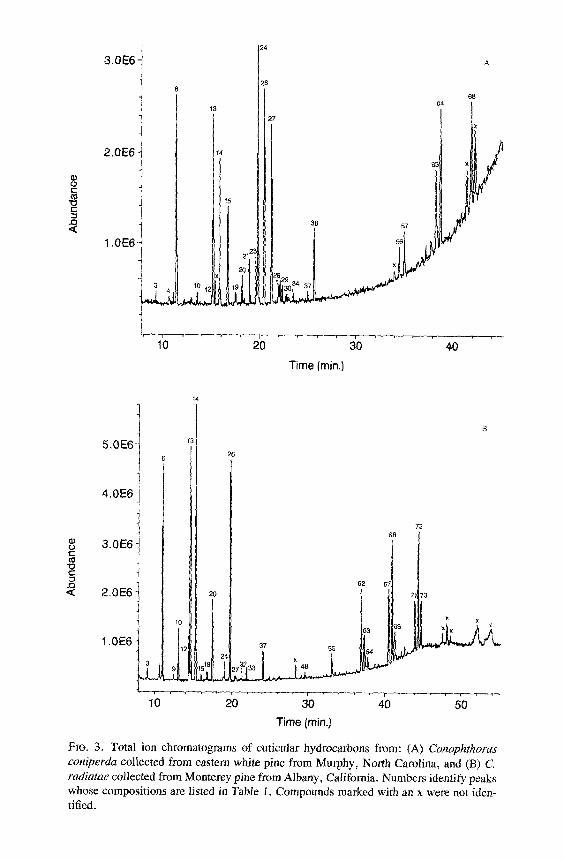
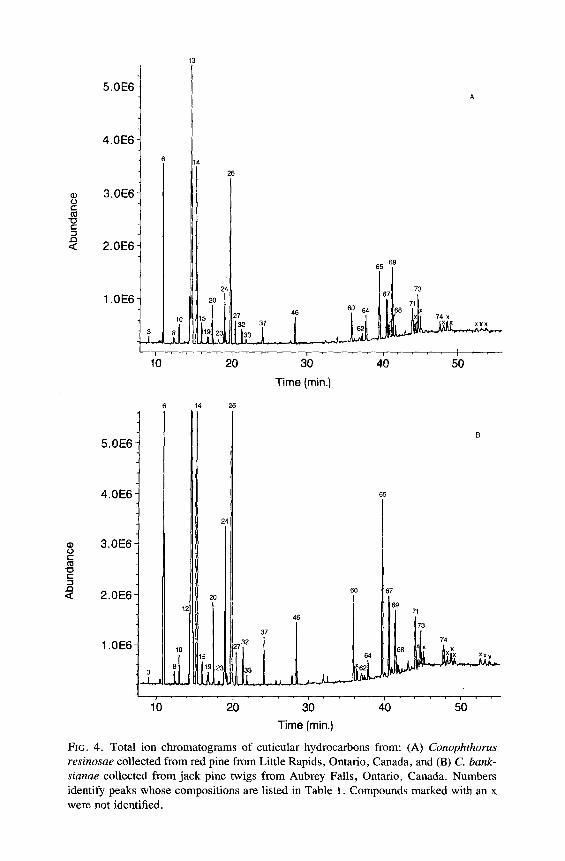
All of the major hydrocarbon components (mean percent ___ 0.5 % of the total hydrocarbon mixture) in the cuticular wax of eight species of Conophtho- rus were characterized (Table 1). Hydrocarbon components consist of homol- ogous series of n-alkanes, alkenes, alkadienes, 2- or 4-methylalkanes, 3- methylalkanes, and single-component and isomeric mixtures of internally branched monomethylalkanes, dimethylalkanes, and trimethylalkanes. Unique and abundant species-specific hydrocarbon components were identified. Monomethyl-, dimethyl-, trimethylalkanes and unsaturated components can easily be used to separate the eight species of Conophthorus (Table 2). GC-MS total ion chromatograms for each species are displayed; closely related species, as inferred from Wood's (1982) key, are paired on the same figure (Figures 1- 4).
The n-alkane composition in all species, except C. conicolens, is a contin- uous series from C2l (n-heneicosane) to C31 with C23, C25, and C27 predomi- nating. In C. conicolens the n-alkane series includes small quantities of C32, C33, C35, and C37. All species have measurable quantities of n-alkanes of even- numbered chain length from C22 to C28; C24 and C26 are the most abundant.
Alkenes were identified by their retention times and diagnostic mass ions. The molecular ion of an alkene is 2 mass units less ". an the molecular ion of the corresponding n-alkane. Alkadienes produce a molecular ion which is 4 mass units less. No structural information, position of double bond(s), or ste- reochemistry was obtained on alkenes. Each Conophthorus species contains C25:1 (pentacosene) and C~7:1 �9 C. ponderosae populations possess alkenes from C23:1 to C35:1. The major alkene in C. conicolens, C. ponderosae, C. cem- broides, C. radiatae, C. resinosae, and C. banksianae is C25: 1, whereas C27:1 is most abundant in C. edulis and C. coniperda. C. resinosae and C. banksianae both contain significant amounts of C35:1 and C37 : 1" Other alkenes are present in very small or trace amounts and occur ~fly as a shoulder of the peak of a 3-(Me) Cn (3-methylalkane). The hydrocarbons from C. ponderosae, C. coni- perda, C. resinosae, and C. banksianae contain the only alkadienes, C25:2 (pentacosadiene, peak 12) and C27:2 (peak 23). The mass spectra of peaks 12 and 23 showed the characteristic molecular ions, m/z 348 and 376, respectively, which are 4 ~ mass units less than the molecular ions of C25 and C27, respecti~ .ly.
All Conophthorus species, except C. conicolens, C. cembroides, and C. edulis, contain mixtures of hydrocarbons with both 2-(Me) Cn and 3-(Me) C,. It is difficult to distinguish 2-(Me) from 4-(Me) Cn (Blomquist et al., 1987). Peak 23 (Table 1) from C. coniperda illustrates this situation and is interpreted as 2- or 4-(Me) C26. The high-abundance M-43 ion at m/z 337 is consistent with 2-(Me) C26. The M-71 : M-72 ion pair from this spectrum, and others, varied in intensity, indicating the possible presence of 4-(Me) C26. Therefore, all are designated as either 2- or 4-(Me) Cn (Tables 1 and 2). The 3-(Me) Cn were identified by their retention times and spectra, which gave a strong (M-29) + ion and a weaker (M-57) § ion.
TA
BL
E 1
. H
YD
RO
CA
RB
ON
S ID
EN
TIF
IED
FO
R E
IGH
T C
onop
htho
rus
SPE
CIE
S
Peak
~ H
ydro
carb
on
EC
L b
CN
b D
iagn
osti
c M
S i
ons
,..,..1
1 2-
Met
hyle
icos
ane
2 n-
Hen
eico
sane
3
n-D
ocos
ane
4 2-
or
4-;
3-M
ethy
ldoc
osan
e ~
5 T
rico
sene
6
n-T
rico
sane
7
9-;
11-;
13-
Met
hylt
rico
sane
"
8 T
etra
cose
ne
9 3-
Met
hylt
fico
sane
10
n-
Tet
raco
sane
11
2-
or
4-M
ethy
ltet
raco
sane
12
P
enta
cosa
dien
e 13
P
enta
cose
ne
14
n-P
enta
cosa
ne
15
9-;
11-;
13-
Met
hylp
enta
cosa
ne'
16
17
18
19
20
21
22
5-M
ethy
lpen
taco
sane
ll
,15-
Dim
ethy
lpen
taco
sane
2-
or
4-M
ethy
lpen
taco
sane
H
exac
osen
e +
3-m
ethy
lpen
taco
sane
"
n-H
exac
osan
e 12
-; 1
3-M
ethy
lhex
acos
ane ~
11,1
5-;
13,1
7-D
imet
hylh
exac
osan
e C
20.7
0 21
2
52
/25
3,2
80
/28
1,2
96
21
.00
21
296
22.0
0 22
31
0 22
.76
23
252/
253,
280/
281,
308/
309,
32
4;
295/
296,
266/
267,
32
4 22
.76
23
322
23.0
0 23
32
4 23
.39
24
140/
141,
224/
225;
16
8/16
9, 1
96/1
97;
196/
197,
168
/169
23
.70
24:1
33
6 23
.73
24
280/
281,
308
/309
, 33
8 24
.00
24
338
24.6
3 25
30
8/30
9, 3
36/3
37,3
52
24.6
9 25
:2
348
24.7
0 25
:1
350
25.0
0 25
35
2 25
.38
26
140/
141,
252
/253
, 36
6;
168/
169,
224
/225
, 36
6;
196/
197,
366
25
.50
26
84/8
5, 3
08/3
09
25.6
0 27
16
8/16
9, 2
39
25.7
0 26
32
3, 3
51
,36
6
25.7
3 26
:1
364
25.7
3 26
30
8/30
9, 3
36/3
37,
366
26.0
0 26
36
6 26
.34
27
182/
183,
224/
225;
19
6/19
7, 2
10/2
11
26.6
0 28
16
8/16
9, 1
82
/18
3,2
39
,25
3;
196/
197,
1
54
/15
5,2
25
,26
7
> ~q
>
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
2- o
r 4-
Met
hylh
exac
osan
e +
he
ptac
osad
iene
c
Hep
taco
sene
3-
Met
hylh
exac
osan
e n-
Hep
taco
sane
9-
; 11
-; 1
3-M
ethy
lhep
taco
sane
"
9-M
ethy
lhep
taco
sane
7-
Met
hylh
epta
cosa
ne
5-M
ethy
lhep
taco
sane
9,
13-;
9,1
5-;
11,1
5-
Dim
ethy
lhep
taco
sane
"
3-M
ethy
lhep
taco
sane
n-
Oct
ocos
ane
9-;
11-;
13-
; 15
-Met
hylo
ctac
osan
d
11,1
5-;
13,1
7-D
imet
hylo
ctac
osan
e ~
3-M
ethy
loct
acos
ane
n-N
onac
osan
e 9-
; 11
-; 1
3-;
15-M
ethy
lnon
acos
ane
~
5-M
ethy
lnon
acos
ane
11,1
5-;
13,1
7-D
imet
hyln
onac
osan
e'
3-M
ethy
lnon
acos
ane
13,1
7,21
-Tri
met
hyln
onac
osan
e n-
Tri
acon
tane
3,
7-D
imet
hyln
onac
osan
e
26.6
3
26.7
0 26
.79
27.0
0 27
.32
27.4
0 27
.42
27.5
0 27
.63
27.7
4 28
.00
28.3
2
28.6
2
28.7
0 29
.00
29.3
2
29.5
0 29
.61
29.7
3 29
.90
30.0
0 30
.10
27
27:2
27
:1
27
27
28
28
28
28
29
28
28
29
29
29
29
30
30
31
30
32
30
31
336/
337,
36
4/36
5,30
8/30
9,
380
376
378
322/
323,
350/
351,
380
380
140/
141,
280/
281;
16
8/16
9, 2
52/2
53;
196/
197,
224/
225,
39
4 14
0/14
1,28
0/28
1,39
4 11
2/11
3,30
8/30
9 84
/85,
336/
337,
394
140/
141,
224/
225,
211,
295;
14
0/14
1,
196/
197,
239,
295;
16
8/16
9,
196/
197,
239,
267
33
6/33
7,
364/
365,
394
394
140/
141,
294
/295
16
8/16
9, 2
66/2
67
196/
197,
238/
239
210/
211,
224/
225
168/
169,
210/
211,
239,
281;
19
6/19
7,
182/
183,
253,
267
350/
351,
378/
379,
408
408
140/
141,
308
/309
; 16
8/16
9, 2
80/2
81;
196/
197,
252/
253;
22
4/22
5 84
/85,
364/
365
168/
169,
224
/225
,239
, 29
5;
196/
197,
267
364/
365,
39
2139
3, 4
22
196/
197,
1
40
/14
1,2
11
,26
7,2
81
,33
7
422
126/
127,
336
/337
,407
,q
> m
o �9 g �9
�9
2:
m
m
M
.-.3
TA
BL
E 1
. C
on
tin
ued
Peak
a H
ydro
carb
on
EC
L b
CN
b D
iagn
ostic
MS
ions
45
1 l, 1
5-;
13,1
7-D
imet
hyltr
iaco
ntan
e c
30.6
0 32
16
8/16
9,23
8/23
9,23
9,30
9;
196/
197,
210/
211,
267,
281
46
n-H
entr
iaco
ntan
e 31
.00
31
436
47
7-;
9-;
11-M
ethy
lhen
tria
cont
ane"
31
.40
32
112/
113,
364
/365
; 14
0/14
1,33
6/33
7;
168/
169,
308
/309
48
11
,15-
; 13
,17-
Dim
ethy
lhen
tria
cont
ane
c 31
.64
33
168/
169,
252/
253,
239,
32
3;
196/
197,
224/
225,
267,
295
49
Doc
otri
acon
tene
+
31
.71
32:1
44
8 3-
met
hyltr
iaco
ntan
e"
32
421/
422,
392/
393,
450
50
ll,1
5,19
-Tri
met
hylh
entr
iaco
ntan
e 31
.90
34
168/
169,
19
6/19
7, 2
67,
239,
309
, 33
7 51
D
ocot
riac
onta
ne
32.0
0 32
45
0 52
3,
7-D
imet
hylh
entr
iaco
ntan
e 32
.14
33
126/
127,
364
/365
,435
53
T
ritr
iaco
nten
e 32
.75
33:1
46
2 54
n-
Tri
tria
cont
ane
33.0
0 33
46
4 55
9-
; ii
-; 1
3-;
15-M
ethy
ltritr
iaco
ntan
e c
33.3
4 34
14
0/14
1 36
4/36
5;
168/
169
336/
337;
19
6/19
7. 3
08/3
09;
224/
225
252/
253
56
11,1
9-;
11,1
5-;
11,2
1-;
13,1
7-;
13,1
9-
33.5
7 35
16
8/16
9 2
24
/22
5,2
95
,35
1;
Dim
ethy
ltritr
iaco
ntan
e c
168/
169
28
0/2
81
,23
9,3
51
; 16
8/16
9.
196/
197,
323,
351;
19
6/19
7. 2
52/2
53,2
67,
323;
19
6/19
7. 2
24/2
25,2
95,
323
57
11,1
5,21
-;
13,1
7,21
-;
13,1
7,23
- 33
.86
36
168/
169,
19
6/19
7,23
9, 2
95,
337,
T
rim
ethy
ltritr
iaco
ntan
e C
36
5;
196/
197,
267,
337;
19
6/19
7,
168/
169,
267,
337,
36
5 t-

58
59
60
61
62
63
64
65
66
67
68
69
70
71
3,7-
Dim
ethy
ltri
tria
cont
ane
11-;
12-
Met
hylt
etra
cont
ane c
Pent
atri
acon
tene
Pe
ntat
riac
onta
ne
9-;
11-;
13-
Met
hylp
enta
tria
cont
ane'
ll,2
3-;
13,1
9-;
13,2
1-;
15,1
9-
Dim
ethy
lpen
tatr
iaco
ntan
e "
Hep
tatr
iaco
nten
e 11
,15,
23-;
13,
17,2
3-
Tri
met
hylp
enta
tria
cont
ane'
Hep
tatr
iaco
ntan
e 9-
; 11
-; 1
3-M
ethy
lhep
tatr
iaco
ntan
e"
13,2
1-;
15,2
1-
Dim
ethy
lhep
tatr
iaco
ntan
e"
13,1
7,21
-; 1
3,17
,23-
T
rim
ethy
lhep
tatr
iaco
ntan
e 11
-; 1
2-;
13-M
ethy
loct
atri
acon
tane
c
9-;
11-;
13-
; 15
- M
ethy
lnon
atri
acon
tane
"
34.0
8 35
12
6/12
7, 3
92/3
93,4
63
34.3
0 35
16
8/16
9, 3
50/3
51;
i82/
I83,
336/
337
34.7
0 35
:1
490
> 35
.00
35
492
35.3
4 36
14
0/14
1,39
2/39
3;
168/
169,
364
/365
; 19
6/19
7, 3
36/3
37
35.6
0 37
16
8/16
9,22
4/22
5, 3
51,3
79;
196/
197,
252/
253,
295,
35
1;
196/
197,
323
,351
; ~ O
22
4/22
5,25
2/25
3,29
5,32
3 36
.70
37:1
51
8 O
z
36.7
8 38
16
8/16
9, 1
96/1
97,2
39,
323,
365,
39
3;
196/
197,
267,
295,
365
37.0
0 37
52
0 37
.30
38
140/
141,
406/
407;
16
8/16
9, 3
92/3
93;
196/
197,
364/
365
37.5
1 39
19
6/19
7,25
2/25
3,32
3,37
9;
224/
225,
252/
253,
323,
35
1 37
.90
40
196/
197,
252
/253
, 26
7, 3
23,3
37,
393
196/
197,
224/
225,
267,
323,
365,
393
38.3
0 39
16
8/16
9,40
6/40
7;
182/
183,
392/
393;
19
6/19
7, 3
78/3
79
39.3
0 40
14
0/14
1,44
8/44
9;
168/
169,
420/
421;
19
6/19
7,39
2/39
3;
224/
225,
364/
365
TA
ttL
E 1
. C
on
tin
ued
Peal
~ H
ydro
carb
on
EC
L b
CN
b D
iagn
osti
c M
S io
ns
72
13,1
7-;
13,2
1-;
15,1
9-
39.6
0 41
19
6/19
7,33
6/33
7,26
7,40
7;
Dim
ethy
lnon
atri
acon
tane
~ 19
6/19
7,28
0/28
1,32
3,40
7;
224/
225,
308/
309,
295,
379
73
13,1
7,23
-;
13,1
7,25
- 39
.90
42
196/
197,
252/
253,
267,
36
5, 3
51,
Tri
met
hyln
onat
riac
outa
ne c
421;
19
6/19
7,22
4/22
5,26
7,
351,
393
,421
74
11
-; 1
3-;
15-M
ethy
lhen
tetr
acon
tane
" 41
.30
42
168/
169,
448/
449;
i9
6/19
7, 4
20/4
21 ;
224/
225,
392/
393
75
13,1
7-;
13,2
1-
41.6
0 43
19
6/19
7,36
4/36
5,26
7,43
5 D
imet
hylh
ente
trac
onta
ne
196/
197,
308
/309
, 32
3,43
5 76
13
,17,
21-T
rim
ethy
lhen
tetr
acon
tane
41
.90
44
196/
197,
308/
309,
267,
33
7,37
9,44
9 77
11
-; 1
3-;
15-M
ethy
ltri
tetr
acon
tane
C
43.3
0 44
16
8/16
9, 4
76/4
77;
196/
197,
448/
449;
22
4/22
5,42
0/42
1 78
13
,17-
Dim
ethy
ltri
tetr
acon
tane
43
.60
45
196/
197,
392/
393,
267,
463
b3
aPea
k nu
mbe
rs r
efer
to p
eaks
ide
ntif
ied
in F
igur
es 1
-4.
bEC
L =
eq
uiva
lent
cha
in l
engt
h, C
N =
ca
rbon
num
ber.
C
An
isom
eric
mix
ture
or
two
or m
ore
com
pone
nts
coel
ute
in t
his
peak
. ~3
,-q
t"
TA
BL
E
2.
HY
DR
OC
AR
BO
NS
ID
EN
TIF
IED
FR
OM
EIG
HT
S
PE
CIE
S O
F E
IGH
T
SP
EC
IES
O14
Co
no
ph
tho
rus
A
Pea
k
No
.
Con
opht
horu
s sp
ecie
s b
Hy
dro
carb
on
E
CL
C
oni
Cpo
n C
cem
C
edu
Cra
d C
con
Cre
s C
ban
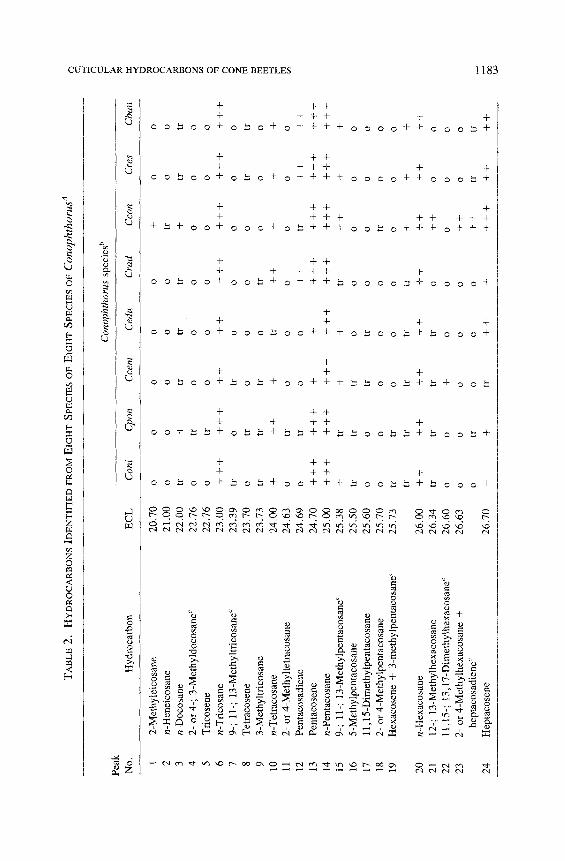
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
2-M
eth
yle
ico
san
e 2
0.7
0
o o
o o
o +
o
n-H
enei
cosa
ne
21
.00
o
o o
o o
tr
o
n-D
oco
san
e 2
2.0
0
tr
+ tr
tr
tr
+
tr
2- o
r 4-
; 3
-Met
hy
ldo
cosa
ne
c 2
2.7
6
o tr
o
o o
o o
Tri
cose
ne
22
.76
o
tr
o o
o o
o
n-T
fico
san
e 2
3.0
0
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+
9-;
11-;
1
3-M
eth
ylt
rico
san
e "
23
.39
tr
o
tr
o o
o o
Tet
raco
sen
e 2
3,7
0
o tr
o
o o
o tr
3-M
eth
ylt
rico
san
e 2
3.7
3
tr
tr
tr
o tr
o
o
n-T
etra
cosa
ne
24
.00
+
+ +
+ tr
+
+ +
+
2- o
r 4
-Met
hy
ltet
raco
san
e 2
4,6
3
o tr
o
o o
o o
Pen
taco
sad
ien
e 2
4.6
9
o tr
o
o +
+ tr
+
+
Pen
taco
sen
e 2
4,7
0
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+
n-P
enta
cosa
ne
25
.00
+
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
9-;
11-;
1
3-M
eth
ylp
enta
cosa
ne'
2
5.3
8
+ tr
+
+ tr
+
+ +
5-M
eth
ytp
enta
cosa
ne
25
.50
tr
tr
tr
o
o o
o
11,1
5-D
imet
hylp
enta
cosa
ne
25
.60
o
o tr
tr
o
o o
2- o
r 4
-Met
hy
lpen
taco
san
e 2
5,7
0
o o
o o
o tr
o
Hex
aco
sen
e +
3-m
eth
ylp
enta
cosa
ne"
2
5.7
3
tr
tr
o o
o o
o
tr
tr
tr
tr
tr
+ +
n-H
exac
osa
ne
26
.00
+
+ +
+ +
+ +
+ +
+ +
+ +
+
12-;
1
3-M
eth
ylh
exac
osa
ne
26
.34
tr
tr
tr
tr
o
+ +
o
11
,15
-;
13
,17
-Dim
eth
ylh
exac
osa
ne'
: 2
6.6
0
o o
+ o
o o
o
2- o
r 4
-Met
hy
lhex
aco
san
e +
26
.63
o
o o
o o
+ +
o
hep
taco
sad
ien
e"
o tr
o
o o
+ +
tr
Hep
taco
sen
e 2
6.7
0
+ +
tr
+ +
+ +
+ +
+ +
O
O
tr
o o ++
+
O tr
O + o ++
+ +
+
++
+
+ o o o o + ++
o o o tr
+H
ta
~
TA
BL
E
2.
Co
nti
nu
ed
4~
Pea
k
No
.
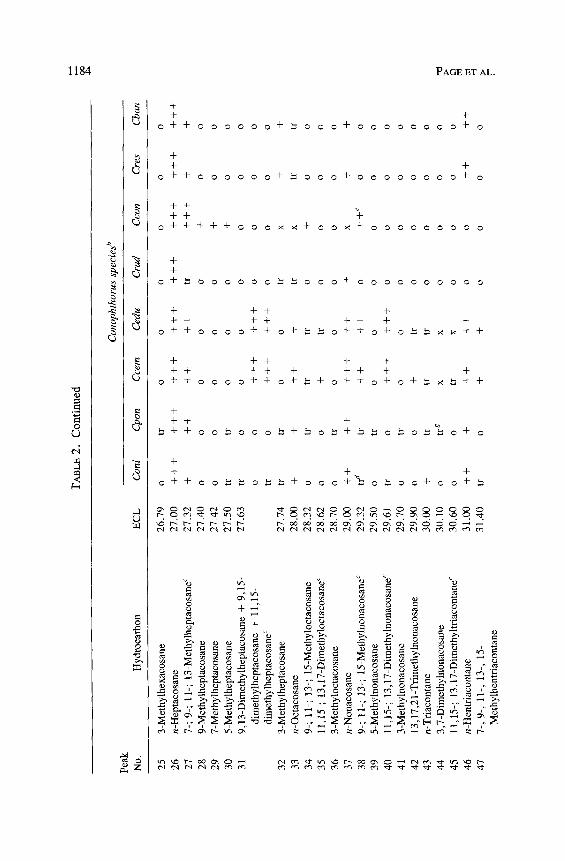
Con
opht
horu
s sp
ecie
s b
Hy
dro
carb
on
E
CL
C
oni
Cpo
n C
cem
C
edu
Cra
d C
con
Cre
s C
ban
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
3-M
eth
ylh
exac
osa
ne
26
.79
o
tr
n-H
epta
cosa
ne
27
.00
+
+ +
+ +
+
7-;
9-;
11-;
1
3-M
eth
ylh
epta
cosa
ne
c 2
7.3
2
+ +
+
9-M
eth
ylh
epta
cosa
ne
27
.40
o
o
7-M
eth
ylh
epta
cosa
ne
27
.42
o
o
5-M
eth
ylh
epta
cosa
ne
27
.50
tr
tr
9,1
3-D
imet
hy
lhep
taco
san
e +
9,1
5-
27
.63
tr
o
dim
eth
ylh
epta
cosa
ne
+ 1
1,1
5-
o o
dim
eth
ylh
epta
cosa
ne
c tr
o
3-M
eth
ylh
epta
cosa
ne
27
.74
tr
tr
n-O
ctac
osa
ne
28
.00
+
+
9-;
11-;
13
-;
15
-Met
hy
loct
aco
san
e 2
8.3
2
o tr
11
,15
-;
13
,17
-Dim
eth
ylo
ctac
osa
ne"
2
8.6
2
o o
3-M
eth
ylo
ctac
osa
ne
28
.70
o
tr
n-N
on
aco
san
e 2
9.0
0
+ +
+ +
9-;
1l-
; 13
-;
15
-Met
hy
lno
nac
osa
ne
c 2
9.3
2
tr ~
tr
5-M
eth
yln
on
aco
san
e 2
9.5
0
o tr
11
,15
-;
13
,17
-Dim
eth
yln
on
aco
san
e C
2
9.6
1
tr
o
3-M
eth
yln
on
aco
san
e 2
9.7
0
o tr
13
,17
,2 l
-Tri
met
hy
lno
nac
osa
ne
29
.90
o
o
n-T
riac
on
tan
e 3
0.0
0
+ tr
3,7
-Dim
eth
yln
on
aco
san
e 3
0.1
0
o tr
e
11
,15
-;
13
,17
-Dim
eth
ylt
riac
on
tan
e c
30
.60
o
o
n-H
entr
iaco
nta
ne
31
.00
+
+ +
7-,
9-,
11-,
13
-,
15-
31
.40
tr
o
Met
hy
lhen
tria
con
tan
e
0 0
0 0
0 0
++
+
++
+
++
+
++
+
++
+
++
+
++
+
+
tr
++
+
+ +
o o
o +
o o
o o
o +
o o
o o
o +
o o
o o
o o
o o
++
+
++
+
o o
o o
++
+
++
+
o o
o o
o o
tr
x +
+
++
+
tr
x tr
tr
tr
tr
o +
o o
+ tr
o
o o
o
o o
o o
o o
++
+
++
+
x +
+ +
+
++
o
++
e o
0
o o
o o
o o
++
+
++
+
o o
o o
o o
o o
o o
+ tr
o
o o
o
tr
tr
o o
o o
x x
o o
o o
tr
x o
o o
o
++
+
+
o o
++
+
+
+ +
o o
o o
> m
>
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
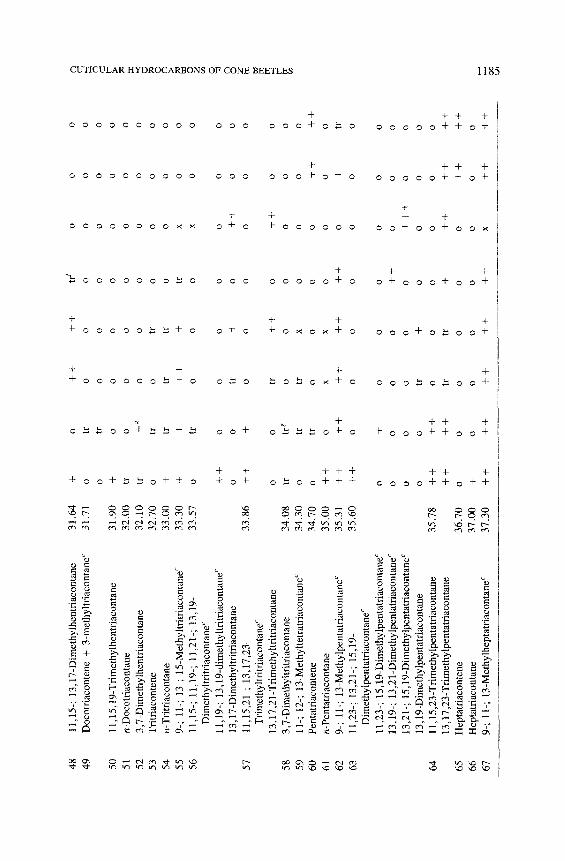
11,1
5-;
13,1
7-D
imet
hylh
entr
iaco
ntan
e 31
.64
Doc
otri
acon
tene
+
3-m
ethy
ltri
acon
tanC
31
.71
+ O
o tr
o
tr
11,1
5,19
-Tfi
met
hylh
entf
iaco
ntan
e 31
.90
+ o
n-D
ocot
riac
onta
ne
32.0
0 tr
o
3,7-
Dim
ethy
lhen
tria
cont
ane
32.1
0 tr
+
g T
ritr
iaco
nten
e 32
.70
o tr
n-
Tri
tria
cont
ane
33.0
0 +
tr
9-;
11-;
13
-;
15-M
ethy
ltri
tria
cont
ane"
33
.30
+ +
11,1
5-;
11
,19
-; 1
1,2
1-;
13
,19-
33
.57
o tr
D
imet
hylt
ritr
iaco
ntan
e'
11,1
9-;
13,1
9-di
met
hylt
ritr
iaco
ntan
e c
+ +
o 13
,17-
Dim
ethy
ltri
tria
cont
ane
o o
11,1
5,21
-;
13,1
7,23
- 33
.86
++
+
Tri
met
hylt
ritr
iaco
ntan
e c
13,1
7,21
-T
rim
ethy
ltri
tria
cont
ane
o o
3,7
-Dim
ethy
ltri
tria
cont
ane
34.0
8 tr
tr
g 11
-;
12-;
13-
Met
hylt
etra
tria
cont
anC
34
.30
o tr
P
enta
tria
cont
ene
34.7
0 o
tr
n-P
enta
tria
cont
ane
35.0
0 +
+ o
9-;
11-;
13
-Met
hylp
enta
tria
cont
anC
35
.31
+ +
+ +
11,2
3-;
13,2
1-;
15,1
9-
35.6
0 +
+
o D
imet
hylp
enta
tria
cont
ane
~ 11
,23-
; 15
,19-
Dim
ethy
lpen
tatf
iaco
ntan
C
o +
13,1
9-;
13,2
1-D
imet
hylp
enta
tria
cont
anC
o
o 13
,21-
; 15
,19-
Dim
ethy
lpen
tatr
iaco
ntan
C
o o
13,1
9-D
imet
hylp
enta
tria
cont
ane
o o
11,1
5,23
-Tri
met
hylp
enta
tria
cont
ane
35.7
8 +
+ +
+ 13
,17,
23-T
rim
ethy
lpen
tatr
iaco
ntan
e +
+ +
+ H
epta
tria
cont
ene
36.7
0 o
o H
epta
tria
cont
ane
37.0
0 +
o 9-
; 11
-;
13-M
ethy
lhep
tatr
iaco
ntan
C
37.3
0 +
+ +
+
++
0 0 0 0 0 o tr
++
0 0 tr
0 tr
o tr
0 x ++
0 O
O
O tr
O tr
O
O ++
++
O
O
O
O
O tr
tr
+ o O + o ++
0 X
0 x ++
0 0 0 0 + 0 tr
0 0 ++
t/
O
O
O
o O
O
O tr
O
O
O
o O
O
O
O
O ++
O
0 ++
0 0 0 + 0 0 ++
0 0 0 0 0 0 0 0 x x 0 ++
0 ++
O
o O
O
O
O
O
O ++
+
O
O ++
O
O
X
O
O
O
O
O
O
O
O
O
O
O
O
O
0 0 0 ++
0 + 0 O
O
O
O
O ++
++
O ++
O
o O ++
O tr
O
O
O
O
O
O ++
++
O ++
0 >
0 z o O
Z m
,..1 7-
oo
L
/I
TA
BL
~ 2
. C
on
tin
ued
Peak
N
o.
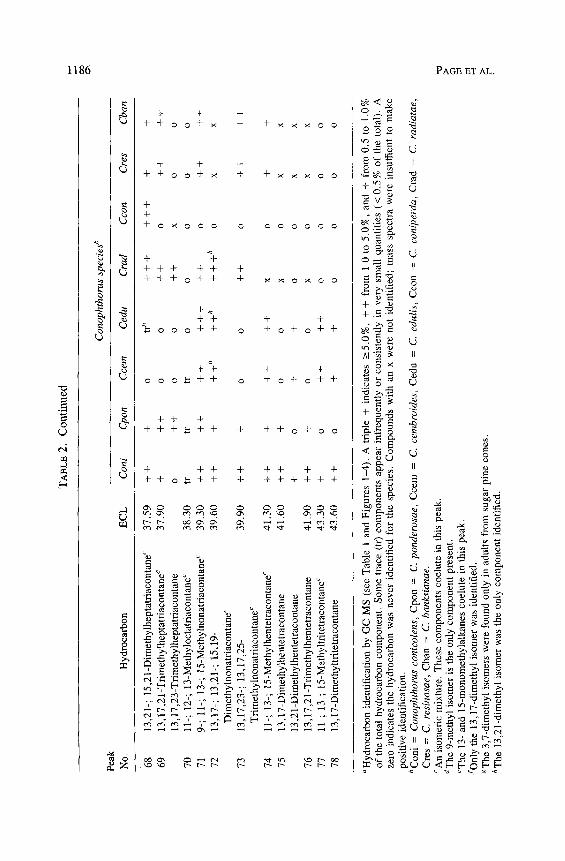
Con
opht
horu
s sp
ecie
s ~
Hyd
roca
rbon
E
CL
C
oni
Cpo
n C
cem
C
edu
Cra
d C
con
Cre
s C
ba
n
68
69
70
71
72
73
74
75
76
77
78
13
,21
-;
15
,21
-Dim
eth
ylh
epta
tria
con
tan
e"
37
.59
+
+ +
o tr
j~
+ +
+ +
+ +
+ +
13
,17
,21
-Tri
met
hy
lhep
tatf
iaco
nta
ne
c 3
7.9
0
+ +
+ o
o +
+ o
+ +
+ +
13
,17
,23
-Tri
met
hy
lhep
tatr
iaco
nta
ne
o +
+ o
o +
+ x
o o
11-;
12
-;
13
-Met
hy
loct
atri
aco
nta
ne
' 3
8.3
0
tr
tr
tr
o o
o o
o
9-;
11-;
13
-;
15
-Met
hy
lno
nat
riac
on
tan
e"
39
.30
+
+ +
+ +
+ +
+ +
+ +
o +
+ +
+
13,1
7-;
13
,21
-;
15,1
9-
39
.60
+
+
+ +
+h
+
+n
+
++
h
o x
x
Dim
eth
yln
on
atri
aco
nta
ne
~
13
,t7
,23
-;
t3,1
7,2
5-
39
.90
+
+
+ o
o +
+
o +
+
++
Tri
met
hy
lno
nat
riac
on
tan
e ~
11%
13
-;
15
-Met
hy
lhen
tetr
aco
nta
ne'
4
1.3
0
+ +
+ +
+ +
+ x
o +
+
13
,17
-Dim
eth
ylh
ente
trac
on
tan
e 4
1.6
0
+ +
+ o
o x
o x
x
13,2
l-D
imet
hy
lhen
tetr
aco
nta
ne
+ o
+ +
o o
x x
13
,17
,21
-T
rim
eth
ylh
ente
trac
on
tan
e 4
1.9
0
+ +
+ o
o x
o x
x
11-;
13
-;
15
-Met
hy
ltfi
tetr
aco
nta
ne"
4
3.3
0
+ o
+ +
+ +
o o
o o
13
,17
-Dim
eth
ylt
rite
trac
on
tan
e 4
3.6
0
+ +
o +
+ o
o o
o
aHyd
roca
rbon
ide
ntif
icat
ion
by G
C-M
S (
see
Tab
le 1
and
Fig
ures
1-4
). A
tri
ple
+ in
dica
tes
_>5.
0%,
+ +
from
1.0
to
5.0%
, an
d +
from
0.5
to
1.0%
of
the
tot
al h
ydro
carb
on c
ompo
nent
. S
ome
trac
e (t
r) c
ompo
nent
s ap
pear
inf
requ
entl
y or
con
sist
entl
y in
veD
' sm
all
quan
titi
es (
< 0
.5 %
of
the
tota
l).
A
zero
ind
icat
es t
he h
ydro
carb
on w
as n
ever
ide
ntif
ied
for
the
spec
ies.
Com
poun
ds w
ith
an x
wer
e no
t id
enti
fied
; m
ass
spec
tra
wer
e in
suff
ient
to
mak
e po
siti
ve i
dent
ific
atio
n.
bCon
i =
Con
opht
horu
s co
nieo
lens
, C
pon
=
C.
po
nd
ero
sae,
C
cem
=
C.
cem
broi
des,
C
edu
=
C.
edul
is,
Cco
n =
C
. co
nipe
rda,
C
rad
=
C.
radi
atae
, C
res
=
C.
resi
nosa
e, C
ban
=
C.
bank
sian
ae.
CA
n is
omer
ic m
ixtu
re.
The
se c
ompo
nent
s co
elut
e in
thi
s pe
ak.
aThe
9-m
ethy
l is
omer
is
the
only
com
pone
nt p
rese
nt.
~The
13-
and
15-
mon
omet
hyla
lkan
es c
oelu
te i
n th
is p
eak.
:O
nly
the
13,1
7-di
met
hyl
isom
er w
as i
dent
ifie
d.
gThe
3,7
-dim
ethy
l is
omer
s w
ere
foun
d on
ly i
n ad
ults
fro
m s
ugar
pin
e co
nes.
hT
he 1
3,21
-dim
ethy
l is
omer
was
the
onl
y co
mpo
nent
ide
ntif
ied.
>
0 c -
o'-
...{3
5.0E6 t
4.0E6
3.0E6
2.0E6
1.0E6
1314
46
37
10 50 5
~ 24 273:3 54 38 43 48 52
31 47~ 51
L 10
57
20 Time (min.)
67
62
64 x 66 i
1 , 4O
72
74
75 78
'3 x
5O
5.0E6
4.0E6
m 3.0E6 O t -
c
.D < 2.0E6-
13 14
101
20
27
37
46 52
67
) 172 74
10 20 3O 40 5O Time (rain.)
FIG. 1. Total ion chromatograms of cuticular hydrocarbons from: (A) Conophthorus conicolens collected from Chihuahua pine from the Chiricahua Mountains in southeast- ern Arizona, and (B) C. ponderosae collected from sugar pine from the Eldomdo National Forest, California. Numbers identify peaks whose compositions are listed in Table 1. Compounds marked with an x were not identified.
6;8 I I 76
19 3033 53 56 60
16 25 = 32 43 58 7
38 5 159 x ~ j ~
i i l L
I1) c-
r
,<
5.0E6
4.0E6'
3.0E6
2.0E6
1.0E6
26
A
40 14
37
71
74
62 / 46 48 E ?
33 35 77
13 28 42 ~2 75
10 54 X]
I
10 20 3O 40 50
Time {min.)
5.0E6
4.0E6
a~ 3.0E6
-O < 2.0E6
1.OE6
3
10
2O
17[ [151i
37 40
67
46 57 62 ) j
48
5~,', ~ 83
72
7~t
20 30 40 Time (min.)
FtG. 2. Total ion chromatograms of cuticular hydrocarbons from: (A) Conophthorus cembroides collected from Mexican pinyon from the Chiricahua Mountains in south- eastern Arizona, and (B) C. edulis collected from pinyon pine from Prescott, Arizona. Numbers identify peaks whose compositions are listed in Table 1. Compounds marked with an x were not identified.
24
3.0E6-
0 c" e~
"0 r
.(3 <
2.0E6 -
1.0E6
13
14 I
i ;
, I
XJ
64
38 57
68
I
10 20 3O 40 Time (rain.)
5.0E6 i
]
4,0E6
72 68
3.0E6 t ,O lie 13 ,t." :::3 .C~ 62 67 < 2.0E6 2 ~
1 9 ~ 63 g'g 1.0E6 37 ~
, ~ ~519 24 273,233 ~ 4B 64
Time (rain.) Fro. 3. Total ion chromatograms of cuticular hydrocarbons from: (A) Conophthortts coniperda collected from eastern white pine from Murphy, North Carolina, and (B) C radiatae collected from Monterey pine from Albany, California. Numbers identify peaks whose compositions are listed in Table 1. Compounds marked with an x were not iden- tified.
5O
0 C;:
"ID CE
,<
0 t - ' -
"10 t -
,<
5.0E6 ~
4 . 0 E 6
3 . 0 E 6
2.0E61
1 . 0 E 8
5.0E6
4.0E6
3 .0E6
2 . 0 E 6
1.0E6-
13
14
26
10
. ~ =
10 20 30
Time (min.)
69 65
24 67 73
20 71
40 50
14 26
24
60
46
37
10 15
. . . . 3'0 . . . .
Time (rain.)
3 . I . I
1'0 . . . . 2'o
65
67
69 71
74
40 50
FIG. 4. Total ion chromatograms of cuticular hydrocarbons from: (A) Conophthorus resinosae collected from red pine from Little Rapids, Ontario, Canada, and (B) C. bank- sianae collected from jack pine twigs from Aubrey Falls, Ontario, Canada. Numbers identify peaks whose compositions are listed in Table 1. Compounds marked with an x were not identified.

CUTICULAR HYDROCARBONS OF CONE BEETLES 1 191
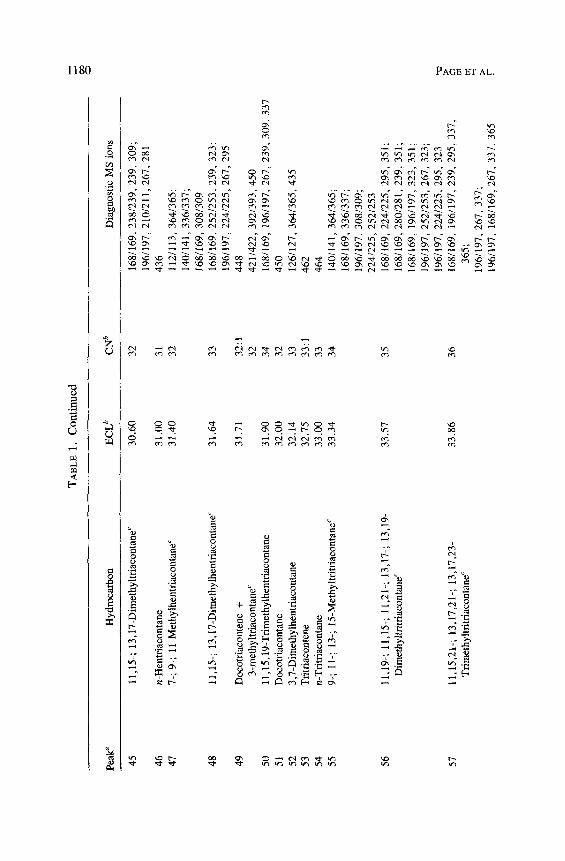
Significant (>0.5% total hydrocarbon) internally branched monometh- ylalkanes occur in these Conophthorus species as complex isomeric mixtures. The mass spectra of peaks with ECL of approximately 0.7 before the corre- sponding Cn have been identified as having methyl branches on odd-numbered carbons (9 through 15). They have strong ions at m/z 140: 141, 168:169, 196 : 197 and 224 : 225, indicating alpha cleavage internal to the branches at 9, 11, 13, and 15 carbons. C. conicolens, C. cembroides, and C. edulis produce the only homologous series of isomeric mixtures that includes all 11- and 13- (Me) Cn compounds having odd-numbered parent chains from 11-; 13-(Me) C25 to 11-; 13(Me) C34. The 9-(Me) Cn isomers are present from 9-(Me) C25 to 9-(Me) C35. The only exception is C. conicolens, which produces only 9-(Me) C29 and adds the 15-(Me) Cn isomers at 15-(Me) C33, 15-(Me) C39, 15-(Me) C4~, and 15-(Me) C43.
There were only five individual or isomeric mixtures of internally branched monomethylalkanes identified with even-numbered parent chains (Tables 1 and 2). C. ponderosae and C. coniperda both produce 12-; 13-(Me) C26. 15-(Me) C28 is found in C. coniperda and 11-; 12-; 13-(Me) C38 in C. conicolens. Traces of ll-; 12-(Me) and 13-(Me) C34 are found in C. ponderosae and C. cem- broides. Small amounts of single-component 5- and 7-monomethyalkanes are present in C. ponderosae and C. coniperda. Trace amounts of 5-(Me) C25, 5-(Me) C27, and 5-(Me) C29 were identified in all of the C. ponderosae popu- lations. C. coniperda is the only species with a single-component 7-(Me) C,, identified as 7-(Me) C27.
Dimethylalkanes in insects generally have methyl branches on odd-num- bered carbons separated by an odd number of carbons (Blomquist et al., 1987). All of the dimethylalkanes produced by these species of Conophthorus fit this pattern. The majority of the methyl branches are separated by three carbons. They ~nclude the 3,7-(Me)2 Cn (3,7-dimethylalkanes) (C. ponderosae, sugar pine, and C. conicolens), the 11,15- and 13,17-(Me)2 Cn (C. conicolens, C. ponderosae, C. cembroides, C. edulis, C. radiatae), 15,19-(Me)2 C35 (C. con- icolens, C. ponderosae, C. coniperda) and 9,13-(Me)2 C27 in C. conicolens. The remainder of the dimethylalkanes have methyl branches separated by 5, 7, 9, and 11 carbons. Dimethylalkanes with methyl branches separated by five carbons include 9,15-(Me)2 C27 (C. cembroides, C. edulis), 13,19-(Me) 2 C n (C. conicolens, C. ponderosae, C. cembroides, C. edulis, and C. radiatae), and 15,21-(Me)2 C37 in all species except C. cembroides and C. edulis. All species include representatives of dimethylalkanes separated by 7 carbons: ll,19-(Me)2 C33; 13,21-(Me)2 C35; 13,21-(Me)2 C37 , and 13,21-(Me)2 C41. The only component with methyl branches separated by nine carbons was 11,21- (Me)2 C33 in an isomeric mixture in C. ponderosae. The one dimethylalkane with 11 carbons separating methyl branches was 11,23-(Me)2 C35 in C. coni- colens and C. ponderosae.
We identified isomeric mixtures of species specific dimethylalkanes in C. ponderosae, C. conicolens, C. cembroides, and C. edulis (Table 2). Dimeth- ylalkanes that eluted about 0.9 carbon units in front of the corresponding n-

1192 PAGE ET AL.
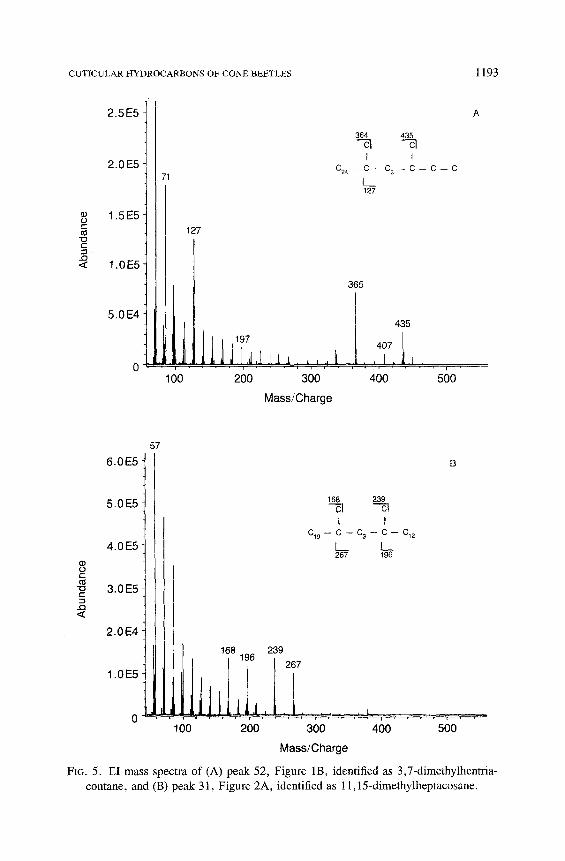
alkane were identified as 3,7-(Me) 2 C n isomers. An example of the mass spec- trum of this type of component is interpreted as 3,7-(Me)2 C31 (Figure 5A). The (M-29) + and (M-57) + ions at m/z 435 and 407 localize one methyl branch on the 3-position. The ion fragments at m/z 364/365 and 127 show that the other methyl branch is on carbon 7. The absence of other diagnostic ions indi- cates that this isomer is the only prominent one in this peak. In C. ponderosae three 3,7-(Me)2 C, (peaks 44, 52, and 58) were unique to beetles infesting sugar pine cones. Thus far we have not found these components in adults infesting ponderosa pine or western white pine cones.
Dimethylalkanes with both methyl branches positioned internally elute ca. 1.4 ECL in front of the corresponding n-alkane. Unique species-specific di- methylalkanes with such ECLs are present in C. conicolens, C. cembroides, and C. edulis (Table 2). The hydrocarbon mixtures of C. cembroides and C. edulis contain significant amounts of 9,15- and 11,15-(Me)2 C27 (peak 31); 11,15- and 13,17-(Me) 2 C29 (peak 40), and 11,15- and 13,17-(Me) 2 C31 (peak 48). An example of the mass spectrum of this type of dimethylalkane was inter- preted as 11,15-(Me) 2 C27 (Figure 5B). The fragments at m/z 168/169 and 196/ 197, where the even-to-odd ratio is greater than one, arise from cleavage inter- nal to each of the carbons bearing methyl branches. The ion fragment at m/z 239 and 267, in which the odd-numbered ions predominate, arise from cleavage external to the two branching methyl groups. The mass spectra of the other internally branched dimethylalkanes were interpreted in a similar manner (Tables 1 and 2).
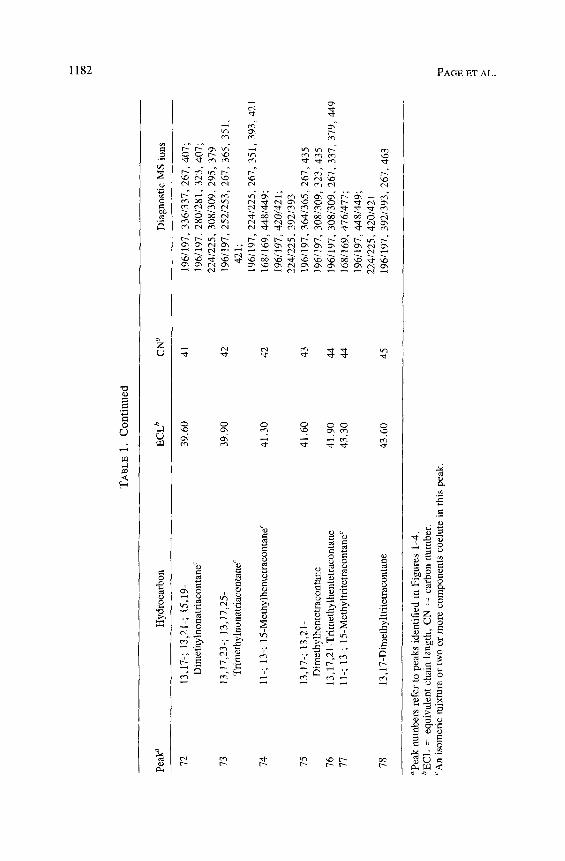
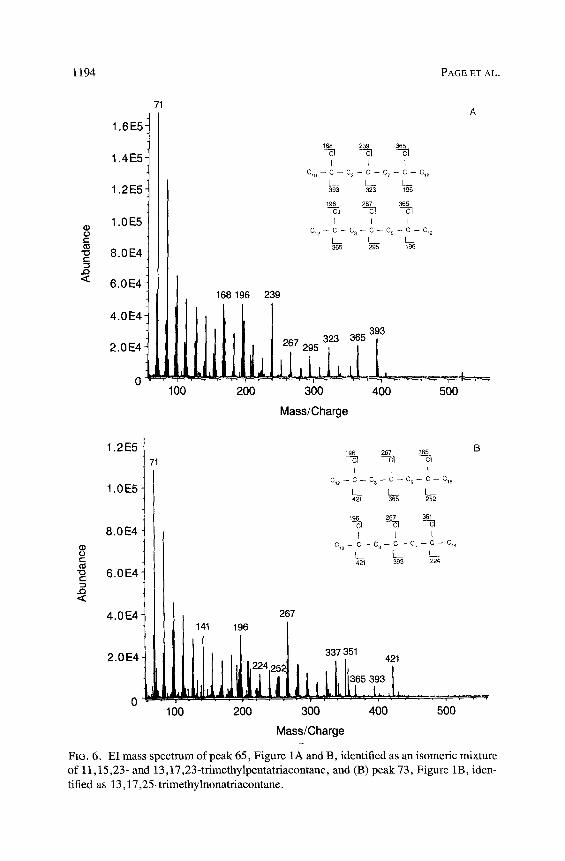
Homologous series of high-molecular-weight trimethylalkanes are found in all eight species of Conophthorus. Each component has an ECL of approx- imately 2.1 less than the corresponding n-alkane. C. cembroides and C. edulis contain fewer trimethylalkanes than the other six species. C. radiatae, C. con- iperda, C. resinosae, and C. banksianae appear to only produce trimethylal- kanes with parent carbon chains of 35, 37, and 39. C. conicolens and C. ponderosae produce isomeric mixtures of trimethylalkanes from parent carbon chains 33-41. We identified a homologous series of 13,17,23-(Me)3 Cn (13,17,23-trimethylalkanes) in C. conicolens and C. ponderosae starting with 13,17,23-(Me) 3 C33 (Figure 6A). The component 13,17,23-(Me) 3 C35 (peak 64) is found in all Conophthorus species characterized in this paper. The significant even-mass ions m/z 196 : 197, with an even-to-odd number ratio of about 1, are interpreted as arising from a cleavage internal to the methyl branches at either end of the molecule. The ions at m/z 267, 295, and 365 arise from cleavage external to the methyl branch on positions 13 and 23 and on both sides of the methyl branch on position 17. The odd-to-even ratio for these ions is greater than 1, as secondary carbonium ions containing two of three methyl branches supress formation of even-mass ions (Blomquist et al. 1987). C. conicolens and C. ponderosae clearly have the most complex series of trimethylalkanes. In these species, we have identified a unique trimethylalkane (peak 73) that occurs
CUTICULAR HYDROCARBONS OF CONE BEETLES 1193
2.5E5
2.0E5
1.5E5
< 1.0E5
5 . 0 E 4
0 -'-
A
364 435
71 C24-- C-- C3-- C-- C -- C L_ 127
127
365
. . . . , [ L . . LJ . . . . . l I . . T . I . J _ ,
100 200 300 400 500
Mass/Charge
6.0E5
5.0E5
4.0E5
O E
-o 3.0E5 E $3 ,<
2.0E4
57
168 239
1 I Clo-- C - - C 3 - C - - C~2
L._ L_ 267 196
16 1i 2391 267 1.0E5
100 200 300 400 500
Mass/Charge
FIG. 5. E1 mass spectra of (A) peak 52, Figure 1B, identified as 3,7-dimethylhentria- contane, and (B) peak 3 l, Figure 2A, identified as l 1,15-dimethylheptacosane.
1194 PAGE ET AL.
0 t -
" 0 r
. . 0 <
1.6E5"
1.4E5
1.2E5
1.0E5
8.0 E4
6.0E4
71
168 2 3 9 3 6 5
I L
C l o - - C - - C 3 - - C - - C 7 - - C - - C12
L__ I__ L._ 3 9 3 3 2 3 196
196 2 6 7 3 6 5
--dl -~ I I
C12 - - c - c 3 - C - C 5 - c - c12
i_._ [__ [_ 3 6 5 2 9 5 196
168 196 239
2.0E4 2 5 393
i i t I Ii f
0 ~ ' - ' - ~ - " ~ - ' - ' - " ~ ; " ' ~ ' ' " ~ -;~ ' " " ' - * " " :
1 O0 200 300 400 500
Mass/Charge
1.2E5
1.0E5
71
8.0E4
0 r
"~ 6.0E4 t--
..(3 <
4.0E4
2.0 E4 .,t
0 ' ~ "
196 2 6 7 3 6 5
b I I
C~2 - - C - - C3 - - C - - C 5 - - C - - C l a
L L._ L._ 421 3 6 5 2 5 2
1 9 6 2 6 7 351
--dl -gl -51 t L I C~2 - - C - - C 3 - - C - - C 7 - - C - - C14
L_ L_ L 421 3 9 3 2 2 4
1 267 141 196 I
, _ r . , , . ; , ; �9 , . . . . I , �9 , �9 i .
100 200 300 400 500
Mass/Charge
FIG. 6. E1 mass spectrum of peak 65, Figure 1A and B, identified as an isomeric mixture of 11,15,23- and 13,17,23-trimethylpentatriacontane, and (B) peak 73, Figure 1B, iden- tified as 13,17,25-trimethylnonatriacontane.

CUT1CULAR HYDROCARBONS OF CONE BEETLES 1195
in an isomeric mixture whose mass spectrum is interpreted as 13,17,25-(Me)3 C39 (Figure 6B). This component has even-mass ions at m/z 196/197 and 224/ 225 and significant odd ions at m/z 267, 351,393 and 421.
D I S C U S S I O N
The cuticular hydrocarbons of Conophthorus, as is the case for many insect species (Blomquist et al., 1987), consist of a complex mixture of unsaturated and straight-chain and methyl-branched, saturated components. The relatively large number of components, ease of analysis, and species-specific composi- tions for many insects make them attractive characters for use in chemotaxon- omy (Carlson and Bolten, 1984; Gastner and Nation, 1986; Haverty et al., 1988; Howard et al., 1988; Lockey, 1982; Vander Meer, 1986).
The cuticular hydrocarbons, particularly the unsaturated and methyl- branched components, are synthesized by the insect (Blomquist et al., 1987). A small portion of the n-alkanes in several species can arise from the diet (Blomquist and Jackson, 1973; Nelson et al., 1971), but unsaturated and methyl- branched hydrocarbons are rare in plants. Thus, the prevalence of unsaturated and methyl-branched components in Conophthorus, while expected, is partic- ularly useful.
Conophthorus is a genus in which many of the species are difficult to sep- arate on the basis of morphological characters alone. In this report we have identified all the major cuticular hydrocarbons of eight of the 15 described spe- cies of Conophthorus (Flores and Bright, 1987; Wood, 1982). We hope to develop a new suite of characters for the entire genus, similar to our studies with the dampwood termites, Zootermopsis (Haverty et al., 1988; Haverty and Thorne, 1989; Thome and Haverty, 1989). The cuticular waxes of these species of Conophthorus contain many diagnostic hydrocarbons, which are useful for separation of these species.
Reports of trimethylalkanes in cuticular lipids of Coleoptera are rare, although they have been reported from other groups of insects (Blomquist et al., 1987). So far, trimethylalkanes have been positively identified only in Cylindrinotus laevioctostriatus (Goeze) and Phylan gibbus (Fabricius) in the Tenebrionidae (Lockey, 1981). Trimethyalkanes may be ubiquitous in all spe- cies of Conophthorus. Each of the eight species we examined in this study produces significant amounts of trimethylalkanes after C3o. The spacing between methyl groups in other Coleoptera has been a [3,3] sequence. We have identi- fied trimethylalkanes with [3,5] and [3,7] spacing. The [3,5] spacing of 11,15,21- and 13,17,23-(Me)3 Cn isomers is the most common in all of these species of Conophthorus. This spacing has been identified so far only in the components 11,15,21- and 13,17,23-(Me)3 C35 and 11,15,21- and 13,17,23- (Me)3 C37 in the female tsetse fly, Glossina pallidipes (Austen) (Nelson and

1196 PAGE ET AL.
Carlson, 1986). We have identified the components 11,15,21- and 13,17,23- (Me)3 C33 and 13,17,25-(Me)3 C39 in several species of Conophthorus, in addi- tion to those trimethylalkanes reported in G. pallidipes. This appears to be the first report of a trimethylalkane with a [3,7] spacing sequence between methyl branches.
The composition of the cuticular wax components in three populations of the polyphagous species Conophthorus ponderosae from three different hosts is one of the most complex. With one possible exception, the hydrocarbon blends are qualitatively the same in all three of the populations we examined; the exceptions are the 3,7-(Me) 2 Cn in the populations from sugar pine. It may be possible that C. ponderosae in ponderosae pine and western white pine produce these dimethylalkanes in significantly lower quantities. Future analyses with more adults from additional populatations may resolve their presence or absence. If one agrees that hydrocarbon profiles are species specific (Haverty et al., 1988; Howard and Blomquist, 1982; Howard et al., 1982; Vander Meer, 1986), we would infer that two populations evaluated here (from ponderosa pine and west- ern white pine) are of the same species or at least very closely related.
The status of C. cembroides and C. edulis as distinct, separate species is in question (S.L. Wood, personal communication). Our analyses of their hydro- carbon components suggest that they are closely related. The only difference in hydrocarbon patterns is the replacement of a dimethylalkane at ECL = 26.6 with an alkene (ECL = 26.7) in C. cemroides. Extensive sampling of Conophthorus from the nut pines in the southwestern United States and northern Mexico should clarify the relationship of these two similar species.
The hydrocarbon patterns of C. resinosae and C. banksianae are qualita- tively identical; no unique hydrocarbon components were found between these two species from many populations in Canada and the United States. They do possess several components that easily separate them from the other six species discussed in this paper. This evidence supports the suspicion that C. banksianae is not a valid species (S.L. Wood and P. de Groot, personal communications). Further elucidation with biological and genetic studies will be necessary to resolve this problem.
Future studies will involve further characterization of the cuticular hydro- carbons of additional species of Conophthorus and quantitative evaluation of intra- and interspecific variation. It is hoped these studies will clarify and/or validate the taxonomy of this genus, especially the polyphagous species, C. ponderosae and the C. edulis complex. Once we have characterized the cuti- cular hydrocarbons of all of the extant species of Conophthorus, we will develop a dichotomous key to all species. To be useful for dichotomous keys, hydro- carbons should be abundant, not minor, components (at least 1%, but preferably > 5 % of the total hydrocarbon mixture). They should be unique or present in only a few of the species, or conversely, they should be common in most of

CUTICULAR HYDROCARBONS OF CONE BEETLES 1 197
the species yet completely absent, rare, or present in insignificant quantities in one or a few. Furthermore, they should have a unique elution time so that they do not coelute with another hydrocarbon in the same species, nor should they elute at a time similar to a different hydrocarbon in a different species.
Acknowledgments--We thank Paul Tilden, Gary DeBarr, Peter deGroot, and Patrick Shea for their help in collecting Conophthorus specimens; and Gary DeBarr, Ralph Howard, and Peter de Groot for providing helpful reviews of early versions of the manuscript. This work was sup- ported, in part, by a grant to M.I.H., M.P., and G.J.B. from the Internal Competitive Grant Program, Pacific Southwest Forest and Range Experiment Station.
REFERENCES
BLAILOCK, T.T., BLOMQUIST, G.J., and JACKSON, L.L. 1976. Biosynthesis of 2-methylalkanes in the crickets Nemobius fasciatus and Gryllus pennsylvanicus. Biochem. Biophys. Res. Com- mun. 68:841-849.
BLOMQUIST, G.J., and JACKSON, L.L. 1973. Incorporation of labelled dietary n-alkanes into the cuticular lipids of the grasshopper Melanoplus sanquinipes. J. Insect Physiol. 19:1639-1647.
BLOMQU1ST, G.J., NELSON, D.R., and DE RENOBALES, M. 1987. Chemistry, biochemistry, and physiology of insect cuticular lipids. Arch. Insect Biochem. Physiol. 6:227-265.
CARLSON, D.A., and BOLTEN, A.B. 1984. Identification of africanized and european honey bees, using extracted hydrocarbons. Bull. Entomol. Soc. Am. 30:32-35.
FLORES, J.L., and BRIGHT, D.E. 1987. A new species of Conophthorus from Mexico: Descriptions and biological notes (Coleoptera: Scolytidae). Coleopterists Bull. 41 : 181-184.
FURNISS, R.L., and CAROLIN, V.M. 1977. Western Forest Insects. U.S. Dept. Agric. Misc. Publ. No. 1339. 654 pp.
GASTNER, J.L., and NATION, J.L. 1986. Cuticular lipids for species recognition of mole crickets (Orthoptera: Gryllotalpidae): II. Scapteriscus abbreviatus, S. acletus, S. vicinus, S. sp. and Neocurtilla hexadactyla. Arch. Insect Biochem. Physiol. 3:126-134.
HAVERTY, M.I., and THORNE, B.L. 1989. Agonistic behavior correlated with hydrocarbon phen- otypes in dampwood termites, Zootermopsis (Isoptera: Termopsidae). J. Insect Behav. 2:223- 243.
HAVERTY, M.I., PAGE, M., NELSON, L.J., and BLOMQUIST, G.J. 1988. Cuticular hydrocarbons of dampwood termites, Zootermopsis: Intra- and intercolony variation and potential as taxonomic characters. J. Chem. Ecol. 14:1035-1058.
HEDLIN, A.F., YATES, H.O., Ill, TOVAR, D.C., EBEL, B.H., KOERBER, T.K., and MERKEL, E.P. 1980. Cone and seed insects of North American conifers. Canadian Forestry Service. 122 pp.
HOWARD, R.W., and BLOMQUtST, G.J. 1982. Chemical ecology and biochemistry of insect hydro- carbons. Annu. Rev. Entomol. 27:149-172.
HOWARD, R.W., MCDANIEL, C.A., NELSON, D.R., BLOMQUIST, G.J., GELBAUM, L.T., and ZAL- KOW, L.H. 1982. Cuticular hydrocarbons as possible species and caste-recognition cues in Reticulitermes sp. J. Chem. Ecol. 8:1227-1239.
HOWARD, R.W., THORNE, B.L., LEWNGS, S.C., and MCDANmL, C.A. 1988. Cuticular hydrocar- bons as chemotaxonomic characters for Nasutitermes corniger (Motschulsky) and N. ephratae (Holmgren) (Isoptera: Termitidae). Ann. Entomol. Soc. Am. 81:395-399.
JACKSON, L.L., and BLOMQUIST, G.J. 1976. Insect waxes, pp. 201-233, in P.E. Kolattududy (ed.). Chemistry and Biochemistry of Natural Waxes. Elsevier, Amsterdam.

1198 PAGE ET AL.
LOCKEY, K.H. 1981. Cuticular hydrocarbons of adult Cylindrinotus laevioctostriatus (Goeze) and Phylan gibbus (Fabricius) (Coleoptera: Tenebrionidae). Insect Biochem. 11:549-561.
LOCKEr, K.H. 1982. Hydrocarbons of adult Onymacris plana (Peringuey) and Onymacris rugati- pennis (Haag) (Coleoptem: Tenebrionidae). Insect Biochem. 12:69-81.
NELSON, D.R. 1978. Long-chained methyl-branched hydrocarbons: Occurrence, biosynthesis, and function. Adv. Insect Physiol. 13:1-33.
NELSON, D.R., and CARLSON, D.A. 1986. Cuticular hydrocarbons of the tsetse flies Glossina mor- sitans morsitans, G. austeni and G. pallidipes. Insect Biochem. 16:403-416.
NELSON, D.R., and SUKKESTAD, D.R. 1970. Normal and branched aliphatic hydrocarbons from the eggs of the tobacco homworm. Biochemistry 9:4601-4611.
NELSON, D.R., SUKKESTAD, D.R., and TERRANOVA, A.C. 1971. Hydrocarbon composition of the integument, fat body, hemolymph and diet of the tobacco homworm. Life Sci. 10:411-419.
NELSON, D.R., SUKKESTAD, D.R., and ZAYLSKIE, R.G. 1972. Mass spectra of methyl-branched hydrocarbons from eggs of the tobacco homworm. J. Lipid Res. 13:413-421.
POMON1S, J.G., FATLAND, C.L., NELSON, D.R, and ZAYLSKIE, R.G. 1978. Insect hydrocarbons. Corroboration of structure by synthesis and mass spectrometry of mono- and dimethylalkanes. J. Chem. Ecol. 4:27-39.
THORNE, B.L., and HAVERTY, M.I. 1989. Accurate identification of Zootermposis species (Isop- tera: Termposidae) based on a mandibular character of non-soldier castes. Ann. Entomol, Soc. Am. 82:262-266.
VANDER MEER, R.K. 1986. Chemical taxonomy as a tool for separating Solenopsis sp., pp. 316- 326, in C.F. Lofgren, R.K. Vander Meet (eds.). Fire Ants and Leaf Cutting Ants: Biology and Management. Westview Press, Boulder, Colorado.
WOOD, S.L. 1982. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Nat. Mere. 6:1359 pp.
Related Documents











