This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Crystal structure and molecular dynamics studies of human purine nucleosidephosphorylase complexed with 7-deazaguanine
Rafael Andrade Caceres a,b,1, Luis Fernando Saraiva Macedo Timmers a,1, Ivani Pauli a,Lisandra Marques Gava c, Rodrigo Gay Ducati d, Luiz Augusto Basso b,d, Diógenes Santiago Santos b,d,*,Walter Filgueira de Azevedo Jr. a,b,*
a Faculdade de Biociências, Instituto Nacional de Ciência e Tecnologia em Tuberculose-CNPq, Laboratório de Bioquímica Estrutural,Pontifícia Universidade Católica do Rio Grande do Sul, PUCRS, Porto Alegre, RS, Brazilb Programa de Pós-Graduação em Medicina e Ciências da Saúde, Pontifícia Universidade Católica do Rio Grande do Sul, PUCRS, Porto Alegre, RS, Brazilc Universidade Estadual de Campinas, UNICAMP, Campinas, SP, Brazild Centro de Pesquisas em Biologia Molecular e Funcional, Instituto de Pesquisas Biomédicas, Instituto Nacional de Ciência e Tecnologia em Tuberculose-CNPq,Pontifícia Universidade Católica do Rio Grande do Sul, PUCRS, Porto Alegre, RS, Brazil
a r t i c l e i n f o
Article history:Received 1 September 2009Received in revised form 9 November 2009Accepted 16 November 2009Available online 22 November 2009
Keywords:Purine nucleoside phosphorylase7-DeazaguanineMolecular dynamicsVirtual screeningX-ray diffractionEnzymatic assay
a b s t r a c t
In humans, purine nucleoside phosphorylase (HsPNP) is responsible for degradation of deoxyguanosine,and genetic deficiency of this enzyme leads to profound T-cell mediated immunosuppression. HsPNP is atarget for inhibitor development aiming at T-cell immune response modulation. Here we report the crys-tal structure of HsPNP in complex with 7-deazaguanine (HsPNP:7DG) at 2.75 Å. Molecular dynamics sim-ulations were employed to assess the structural features of HsPNP in both free form and in complex with7DG. Our results show that some regions, responsible for entrance and exit of substrate, present a con-formational variability, which is dissected by dynamics simulation analysis. Enzymatic assays were alsocarried out and revealed that 7-deazaguanine presents a lower inhibitory activity against HsPNP(Ki = 200 lM). The present structure may be employed in both structure-based design of PNP inhibitorsand in development of specific empirical scoring functions.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
Purine nucleoside phosphorylase (PNP) (EC 2.4.2.1) is a key en-zyme of the purine salvage pathway, responsible for the reactionbetween (deoxy)nucleosides and bases, which can in turn be trans-formed to uric acid for excretion or reused in nucleic acid biosyn-thesis (Parks and Agarwal, 1972). This enzyme catalyzes thereversible cleavage, in the presence of inorganic phosphate (Pi),of N-glycosidic bonds of purine (deoxy)nucleosides, except adeno-sine, to produce alpha-ribose 1-phosphate and the equivalent pur-ine base (Kalckar, 1947). The reaction proceeds with inversion ofconfiguration, from beta-nucleosides to alpha-ribose 1-phosphate(Porter, 1992).
Human PNP (HsPNP) participates in the removal of deoxygua-nosine (dGuo), and this process is fundamental for T-cell immunefunctions (Gelfand et al., 1978a,b). The genetic deficiency of HsPNPhas been shown to produce specific T-cell immune deficiency(Giblett et al., 1975). The syndrome results from the increasedblood levels of dGuo and its conversion to dGTP in dividing T-cells.Altered deoxynucleotide pools allosterically block ribonucleotidediphosphate reductase, insert errors into DNA and activate apopto-sis in dividing T-cells (Bzowska et al., 2000; Minkui et al., 2008;Kicska et al., 2001). Since the establishment of the relationship be-tween PNP and T-cell associated autoimmune diseases, an intensesearch for PNP inhibitors has been carried out (Schramm, 2004;Schramm and Grubmeyer, 2004; Ghanem et al., 2008; Knappet al., 2006). Although the mechanism by which HsPNP deficiencycauses impaired T-cell maturation and discrimination is not com-pletely understood, T-cell leukemia and lymphomas can be im-paired by efficient inhibitors that target HsPNP.
The 7-deazaguanine (7DG) (Fig. 1) is an analogue of the HsPNPsubstrate guanine, which may act as a competitive inhibitor andthus form a binary complex with free enzyme.
In the present work we describe the crystallographic structureof HsPNP associated with 7DG, which was solved at 2.75 Å, using
1047-8477/$ - see front matter � 2009 Elsevier Inc. All rights reserved.doi:10.1016/j.jsb.2009.11.010
* Corresponding authors. Address: Centro de Pesquisas em Biologia Molecular eFuncional, PUCRS, Av. Ipiranga, 6681 Prédio 92A, Porto Alegre 90619-900, RS, Brazil.Fax: +55 51 33203629.
E-mail addresses: [email protected] (D.S. Santos), [email protected] (W.F.de Azevedo).
1 These authors contributed equally to this work, the order of authorship beingarbitrary.
Journal of Structural Biology 169 (2010) 379–388
Contents lists available at ScienceDirect
Journal of Structural Biology
journal homepage: www.elsevier .com/locate /y jsbi

Author's personal copy
recombinant HsPNP and synchrotron radiation. Molecular dynam-ics (MD) simulations were applied to verify the stability of thecrystal structure in solution and to give detailed information onthe dynamics properties of the structure in free form and associ-ated with the ligand. We also present a virtual screening (VS) pro-tocol for analogues of 7DG, which were obtained from ZINCdatabase (Irwin and Shoichet, 2005). In addition, steady-statekinetics and equilibrium fluorescence spectroscopy assays fordetermination of the Kd value for 7DG confirmed the affinity of thisligand for HsPNP.
2. Materials and methods
2.1. Crystallization and data collection
Recombinant HsPNP was obtained as previously described (Sil-va et al., 2003). PNP solution was concentrated to 12 mg/mLagainst 20 mM Tris–HCl, pH 8.0 and incubated for 2 h with 7DGin a molar ratio between protein and ligand of 1:3. Hanging dropswere prepared mixing 1 lL of protein solution and 1 lL of reservoirsolution containing 19% ammonium sulfate and 0.5 M sodium cit-rate pH 5.6. The crystals complexed with 7DG appeared 24 h afterthe drops were prepared. The crystals were flash frozen at 100 K
and cryoprotected with 20% of glycerol. X-ray diffraction data werecollected at wavelength 1.431 Å using Synchrotron RadiationSource (Laboratório Nacional de Luz Síncrotron, Campinas, SP, Bra-zil) and CCD detector (MARCCD) with an exposure time of 60 s perimage at a crystal to detector distance of 120 mm. The crystals dif-fracted at 2.75 Å and the data were processed using MOSFLM andscaled using SCALA (Collaborative Computational Project No. 4,1994). Cell parameters for HsPNP complexed with 7DG werea = b = 138.7 and c = 159.4 Å.
2.2. Structure resolution and refinement
The crystal structure of HsPNP complexed with 7DG was deter-mined by standard molecular replacement methods using the pro-gram AMoRe (Navaza, 1994), incorporated in the CCP4 programpackage (Collaborative Computational Project No. 4, 1994), usingas search model the structure of HsPNP complexed with 8-aza-guanine (PDB access code: 1V41). Structure refinement was per-formed using the program REFMAC (Collaborative ComputationalProject No. 4, 1994). The atomic positions acquired from molecularreplacement were used to start the crystallographic refinement.The overall stereochemical quality of the final model was assessedby the program PROCHECK (Laskowski et al., 1993). The trimericstructure and atomic models were superposed using the programLSQKAB from CCP4 (Collaborative Computational Project No. 4,1994).
2.3. Molecular dynamics simulations
The MD simulations were performed with the trimeric structureof the HsPNP to provide a more realistic analysis, since the HsPNPis biologically active as a trimer. The MD simulations were carriedout in periodic boundary conditions using the GROMACS 3.3.1 (vander Spoel et al., 2005) package using the Gromos 96.1 (53A6) forcefield (Oostenbrink et al., 2005). The molecular topology file andforce field parameters, except for the charges for the inhibitor7DG, were generated by the program PRODRG (van Aalten et al.,
Fig. 1. 7-Deazaguanine chemical structure.
Fig. 2. (A) (top right) Classical binding site of HsPNP; the 7DG is anchored by the two main residues of the binding pocket (Glu201 and Asn243). (B) (bottom right) Electrondensity (2Fobs � Fcalc) map of 7DG in binding pocket.
380 R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388

Author's personal copy
1996). The partial atomic charges of 7DG were determined by GA-MESS program (Schmidt et al., 1993), which were submitted to sin-gle-point ab initio calculations at RHF 6-31G* level in order toobtain Löwdin derived charges. Before MD simulations, the uncom-plexed and complexed models were solvated with the explicit SPC/E water model (Berendsen et al., 1981) embedded in 90.20 Å �85.86 Å � 48.95 Å or 90.99 Å � 86.13 Å � 49.62 Å boxes, respec-tively, in which the minimum distance between the protein surfaceand the box face was 10 Å in both cases. To neutralize the negativecharge density of the systems, sodium ions were added. The simu-lation systems were composed of 42,246 (HsPNP) and 42,306(HsPNP:7DG) water molecules in total. The systems were then sub-jected to a steepest descent followed by conjugate gradient energyminimization until a tolerance of 1000 kJ/mol, step by step. The en-tire systems were minimized. The temperature of the system wasthen increased from 50 to 300 K in five steps (50–100, 100–150,150–200, 200–250 and 250–300 K), and the velocities at each stepwere reassigned according to the Maxwell–Boltzmann distributionat that temperature and equilibrated for 10 ps, except the last partof thermalization phase, that was for 50 ps. The systems were thensubmitted to a short MD with position restrains for a period of30 ps and, afterwards, performed a full MD without restrains. Fi-nally, the 4 ns MD simulations were performed under normal tem-perature (300 K) and pressure (1 bar), using a temperaturecoupling time constant of 0.1 ps and a pressure coupling time con-stant of 1.0 ps. The value of the isothermal compressibility was setto 4.5 � 10�5 bar�1 for water simulations. All bond lengths includ-ing hydrogen atoms were constrained by the LINCS algorithm(Hess et al., 1997). The electrostatic interactions were calculatedby using the Particle-mesh Ewald (PME) algorithm (Darden et al.,1993), with interpolation order of 4 and a grid spacing of0.12 nm. The cutoff for van der Waals interactions was 1.0 Å. Sim-ulations were performed with a time step of 2 fs, and coordinateswere saved every 1 ps. Examinations of the molecular structureswere achieved using the Pymol (Delano and Lam, 2005) andVMD (Humphrey et al., 1996) programs. All simulations were car-ried out using the GROMACS program on a DuoQuad Core 550Xeon – 3.00 GHz server. Analysis were performed using the fea-tures within the GROMACS package.
2.4. Essential dynamics analysis
Essential dynamics (ED), also known as principal componentanalysis (PCA), is a method commonly used for dissecting thedynamics of proteins and their importance in biological processes,like protein folding or substrate binding. The ED analysis is a tech-nique that reduces the complexity of the data and extracts the con-certed motion in simulations that are essentially correlated andpresumably meaningful for biological function (Amadei et al.,1993). In the ED analysis, a variance/covariance matrix was con-structed from the trajectories after removal of the rotational andtranslational movements. A set of eigenvectors and eigenvalueswas identified by diagonalizing the matrix. The eigenvalues repre-sented the amplitude of the eigenvectors along the multidimen-sional space, and the displacements of atoms along eacheigenvector showed the concerted motions of protein along eachdirection. An assumption of ED analysis is that the correlated mo-tions for the function of the protein are described by eigenvectorswith large eigenvalues. The movements of protein in the essentialsubspace were identified by projecting the Cartesian trajectory coor-dinates along the most important eigenvectors from the analysis.
2.5. Molecular docking protocol
The flexible docking simulations (FDS) were performed usingMolDock 3.2.0 (Thomsen and Christensen, 2006), which is used
to predict protein–ligand complexes. The docking scoring functionused for MolDock is based on piecewise linear potential, as de-scribed elsewhere (Thomsen and Christensen, 2006). Docking sim-ulations is important for the development of new drugs becauseFDS is based on three basic tasks, which are: (1) characterizationof the binding site; (2) positioning of the ligand into the bindingsite; and (3) evaluating the strength of interaction for a specific li-gand–receptor complex (Bourne and Weissig, 2003).
The first step of the docking simulation was performed as avalidation of the docking protocol (redocking), focused on HsPNPcrystallographic structure associated with 7DG. We used the fol-lowing parameters as a protocol for this enzyme: MolDockScore[GRID], grid resolution (0.30 Å), radius of the sphere (12 Å), thealgorithm of MolDock SE, number of runs (10), maximum itera-tions (1500), maximum population size (100, and the otherparameters were used as default. The validation results wereanalyzed based on the Re-rank Score (Thomsen and Christensen,2006).
2.6. Virtual screening protocol
One attractive attribute of ZINC database is a compilation ofpredefined subsets of small molecules (Irwin and Shoichet,2005). These subsets were defined based on physicochemical crite-ria in order to assist VS initiatives. Another valuable option is the
Table 1Data collection and refinement statistics. Values in parentheses refer to the highestresolution shell.
X-ray wavelength (Å) 1.431Temperature (K) 100Resolution range (Å) 47.972–2.75Number of measurements with I > 2r(I) 15,364Number of independent reflections 14,793Space group R32Unit-cell parametersa = b (Å) 138.7c (Å) 159.4a = b (�) 90c (�) 120Highest resolution shell (Å) 2.9–2.75Data completeness (%) 99.1 (99.1)Rmerge (%)a 8.1 (33.1)Resolution range used in refinement (Å) 43.68–2.75Rfactor (%)b 20.9Rfree (%)c 26.9Observed RMSD from the ideal geometryBond lengths (Å) 0.04Bond angles (�) 3.5B values (Å)2d
Main chain 43.82Side chains 44.20Ligand 25.83Waters 39.96Sulfate groups 33.14Residues in most favored regions of the
Ramachandran plot (%)80.3
Residues in additionally allowed regionsof the Ramachandran plot (%)
16
Residues in generously allowed regionsof the Ramachandran plot (%)
3.3
Residues in disallowed regions of theRamachandran plot (%)
0.4
Number of water molecules 24Number of sulfate groups 3
a Rmerge =P
|Ii � hIi|/P
|Ii|, where Ii is the scaled intensity of the ith measurementand hIii is the mean intensity for that reflection.
b Rfactor = 100 �P
|Fobs � Fcalc|/P
(Fobs), the sums being taken over all reflectionswith F/r(F) > 2 cutoff.
c Rfree = Rfactor for 10% of the data, which were not included during crystallo-graphic refinement.
d B values = average B values for all non-hydrogen atoms.
R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388 381

Author's personal copy
possibility to generate your own subset of potential inhibitors. Thisoption permits an effective subset to be employed for other groupsinterested in the same problem. With that in mind, we employedthe 7DG core to carry out a search in the ZINC database (Irwinand Shoichet, 2005; Timmers et al., 2008) to build a small-mole-cule database, using the pharmacophoric fingerprint of 7DG witha Tanimoto coefficient cutoff of 60%. This set was constituted by79 compounds, for which 8 molecules satisfy the Lipinski’s ruleof five (Lipinski et al., 1997).
2.7. Enzymatic assay
All chemicals were of analytical-reagent grade and were usedwithout further purification. The reagent 7DG was taken from Sigmaand 2-amino-6-mercapto-7-methylpurine ribonucleoside (MESG)is commercially available in the Enzchek phosphate assay kit fromMolecular Probes. 7DG was solubilized in H2O pH 4.0. HsPNP expres-sion and purification were conducted as described elsewhere (Silvaet al., 2003). All enzyme activity assays were performed at 25 �C and50 mM Tris at pH 7.6. The HsPNP conversion of 2-amino-6-mer-capto-7-methylpurine from MESG (e = 11,000 M�1 cm�1 at 360 nm(Webb, 1992)) was monitored spectrophotometrically by thechange in absorbance in the presence of Pi. The concentration of7DG that reduces HsPNP enzyme activity by half (IC50) was deter-mined at MESG (358 lM) and Pi (94 lM) concentrations close totheir KM values in either absence or presence of 7DG in the reactionmixture. The pH of the cuvette (reaction mixture) was monitored toensure that it was kept constant, since the pH of the 7DG stock solu-tion was 4.0.
2.8. Fluorescence spectroscopy
Fluorescence titration of HsPNP by 7DG was carried out at 25 �Cby making microliter additions of 2 or 4 mM 7DG stock solutions
(final concentration ranging from 1.0 to 104.13 lM) to 2 mL of3 lM HsPNP in 50 mM Tris at pH 7.6, keeping the dilution to amaximum of 2.75%. Measurements of intrinsic protein fluores-cence of HsPNP employed excitation wavelength at 300 nm andemission wavelength ranging from 310 to 420 nm (maximumkEM = 335 nm). The slits for excitation and emission wavelengthswere, respectively, 3.0 and 20 nm. To account for the inner filter ef-fect due to 7DG absorption of excitation light, two cuvettes wereplaced in series so that the contents of the first cuvette (either buf-fer or 7DG) acted as a filter of the excitation light and the lightemitted from the second cuvette detected.
2.9. Experimental data analysis
Data from equilibrium fluorescence spectroscopy were fitted toEq. (1), the Hill equation (Hill, 1913), where F is the observed fluo-rescence signal, Fmax is the maximal fluorescence, F/Fmax ratio is thedegree of saturation, n represents the total number of binding sites,and K0 is the mean dissociation constant for HsPNP:7DG binarycomplex formation, which is comprised of interaction factors andthe intrinsic dissociation constant.
F=Fmax ¼ An=ðK 0 þ AnÞ ð1Þ
3. Results and discussion
3.1. Molecular replacement and crystallographic refinement
The standard procedure of molecular replacement using AMoRe(Navaza, 1994) was used to solve the structure. The best modelgenerated a correlation of 79.3% and Rfactor of 28.8%. The highestmagnitude of the correlation coefficient function obtained for theEuler angles were a = 117.45�, b = 59.77�, and c = 153.53�. The frac-tional coordinates are Tx = 0.1645, Ty = 0.3749, and Tz = 0.0322. At
Fig. 3. Schematic drawing of trimeric HsPNP:7DG complex. The protein is presented as cartoon and 7DG and SO4 as sticks.
382 R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388

Author's personal copy
this stage Fobs � Fcalc omit maps were calculated. These mapsshowed clear electron density for the 7DG in the complex. Fig. 2Bshows the electron density (Fobs � Fcalc) map where we clearlysee the bound ligand. Atomic coordinates for 7DG have been in-cluded in the model and all crystallographic refinement using REF-MAC (Collaborative Computational Project No. 4, 1994) continuedwith maximum likelihood protocol, followed by alternate cyclesof positional refinement and manual rebuilding using MIFit 3.1.0(McRee, 2004). Finally, the positions of 7DG, water, and sulfatemolecules were checked and corrected in Fobs � Fcalc maps. The fi-nal model has an Rfactor of 20.9% and Rfree of 26.9%, with 24 watermolecules, three sulfate ions, and 7DG. The X-ray and refinementstatistics are shown in Table 1.
3.2. Crystallographic structure of HsPNP:7DG
The structure of HsPNP:7DG follows the same fold as otherHsPNPs, which are described elsewhere (Timmers et al., 2008).The data obtained of the crystallography reveals that the structureis a trimer (Fig. 3) in solution and each monomer displays an alpha/beta fold consisting of a mixed beta-sheet surrounded by alpha-helices. The analysis of the electron density maps at final stagesof the crystallographic refinement shows that the ligand (7DG) isfound into the classical binding site of the HsPNP (Fig. 2A) and an-chored by the two main residues of the binding pocket (Glu201and Asn243). When analyzed the differences among the proteinin free form and associated with the ligand, we observed confor-mational changes, as expected, mainly in the regions close to thebinding pocket, and the loop region which includes the residuesLeu240–Val260.
The interactions of the ligand and the HsPNP were evaluatedwith the program Ligplot (Wallace et al., 1995), which revealstwo intermolecular hydrogen bonds between the ligand and theresidues Glu201 and Asn243, as commented above, and hydropho-bic contact with other residues of the binding pocket (Ala116,Ala117, Gly118, Val217, and Phe200) (Table 2).
3.3. Virtual screening
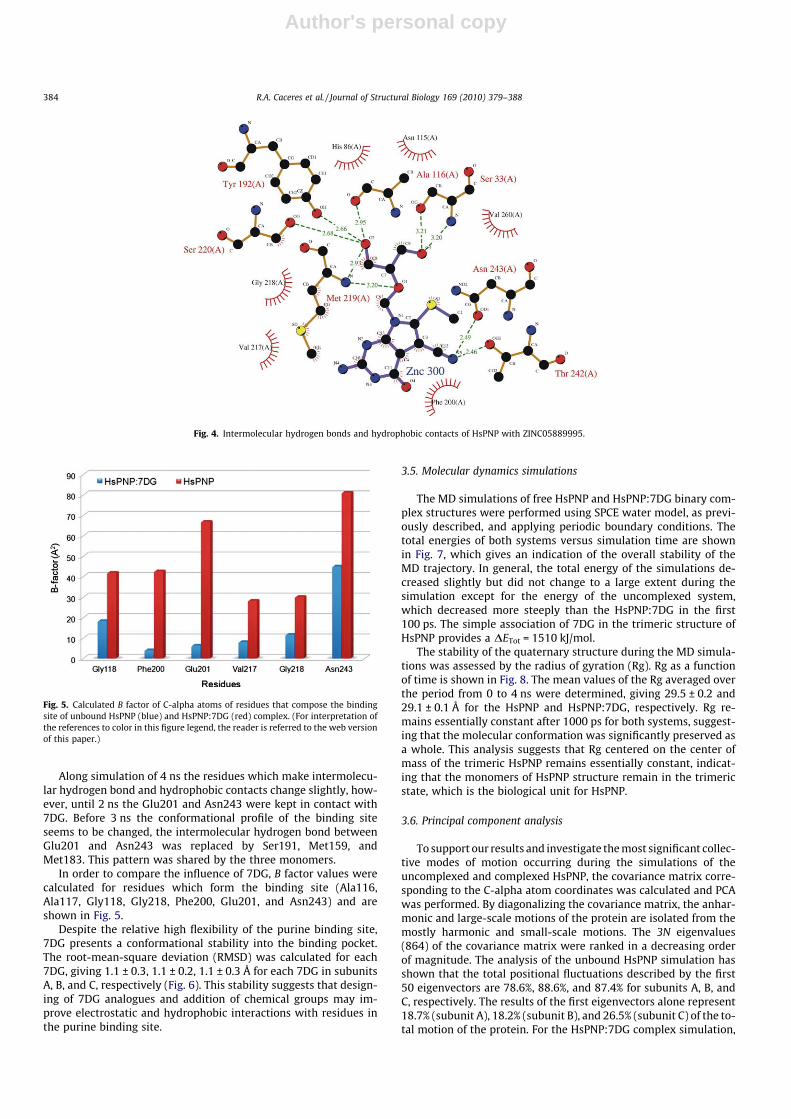
We have applied VS for a set of 79 molecules from the ZINCdatabase in order to identify new potential inhibitors for HsPNPbased on the analogues of 7DG. The FDS results were analyzed tak-ing into account the piecewise linear potential (PLP), which is ob-tained from the MolDock program. The lowest local energies wereachieved for five molecules of a set composed by 79 ligands fromthe ZINC database, which are ZINC05889995, ZINC05890010,ZINC05117045, ZINC05890018, and ZINC05425329, suggestingthat these molecules could be potential inhibitors. The interactionsof the ZINC05889995 ligand with HsPNP were assessed (Fig. 4)with the program Ligplot (Wallace et al., 1995). It is obvious thatexperimental analysis should be carried out aiming to verify theinhibition activity. However, none of the five molecules are com-mercially available becoming necessary to synthesize it to furtherassays.
3.4. Interactions of HsPNP with 7-deazaguanine
The specificity and affinity between enzyme and a ligand de-pend on directional hydrogen bonds and ionic interactions, as wellas on shape complementarity of the contact surfaces of both part-ners (Canduri et al., 2001). In order to better describe and under-stand the 7DG interactions in the binding site under solution wemonitored each subunit binding pocket. Analysis of the hydrogenbonds between 7DG and HsPNP revealed two intermolecularhydrogen bonds involving the residues Glu201 at 2.7 Å andAsn243 at 2.6 Å, in all monomers (Table 2). Ta
ble
2H
ydro
gen
bond
san
dhy
drop
hobi
cco
ntac
tsof
HsP
NP
wit
h7D
Gof
mon
omer
sdu
ring
MD
sim
ulat
ions
.
Subu
nit
ASu
bun
itB
Subu
nit
C
Hid
roge
nbo
nd
Hid
roph
obic
con
tact
Hid
roge
nbo
nd
Hid
roph
obic
con
tact
Hid
roge
nbo
nd
Hid
roph
obic
con
tact
Tim
eR
es(C
hai
n)
Ato
ms
Dis
t(Å
)R
es(C
hai
n)
Tim
eR
es(C
hai
n)
Ato
ms
Dis
t(Å
)R
es(C
hai
n)
Tim
eR
es(C
hai
n)
Ato
ms
Dis
t(Å
)R
es(C
hai
n)
Init
ial
stru
ctu
reG
lu20
1(A
)O
E2?
N2
2.7
Ala
116(
A)
Init
ial
stru
ctu
reG
lu20
1(B
)O
E2?
N2
2.7
Ala
116(
B)
Init
ial
stru
ctu
reG
lu20
1(C
)O
E2?
N2
2.7
Ala
116(
C)
Asn
243(
A)
ND
2?
O7
2.6
Ala
117(
A)
Asn
243(
B)
ND
2?
O7
2.6
Ala
117(
B)
Asn
243(
C)
ND
2?
O7
2.6
Ala
117(
C)
Gly
118(
A)
Gly
118(
B)
Gly
118(
C)
Phe2
00(A
)Ph
e200
(B)
Phe2
00(C
)V
al21
7(A
)V
al21
7(B
)V
al21
7(C
)G
ly21
8(A
)G
ly21
8(B
)G
ly21
8(C
)1
ns
Glu
201(
A)
OE1
?N
23.
0Ph
e200
(A)
1n
sG
lu20
1(B
)O
E1?
N2
3.0
Phe1
59(A
)1
ns
Gly
197(
C)
O?
N11
3.0
Val
195(
C)
Val
217(
A)
OE1
?N
83.
3V
al21
7(B
)A
sn24
3(C
)N
D2?
O7
2.9
Ser1
99(C
)M
et21
9(A
)A
sn24
3(B
)N
D2?
O7
2.9
Gly
218(
B)
Met
219(
C)
2n
sG
lu20
1(A
)O
E2?
N2
3.2
Phe2
00(A
)2
ns
Glu
201(
B)
OE1
?N
23.
1Ph
e159
(A)
2n
sG
lu20
1(C
)O
E2?
N2
2.8
Val
195(
C)
Asn
243(
A)
ND
2?
O7
3.5
Val
217(
A)
Asn
243(
B)
ND
2?
O7
2.7
Phe2
00(B
)O
E2?
N8
3.2
Phe2
00(C
)G
ly21
8(A
)V
al21
7(B
)A
sn24
3(C
)N
D2?
O7
2.9
Val
217(
C)
Gly
218(
B)
Gly
218(
C)
3n
sG
lu20
1(A
)O
E1?
N2
2.9
Met
219(
A)
3n
sG
lu20
1(B
)O
E1?
N2
3.0
Gly
218(
B)
3n
sG
lu20
1(B
)O
E1?
N2
3.1
Gly
218(
C)
OE1
?N
83.
1A
sn24
3(B
)N
D2?
O7
2.8
Asn
243(
B)
ND
2?
O7
2.7
Phe2
00(C
)V
al21
7(C
)4
ns
Met
218(
A)
SD?
N11
3.2
Phe2
00(A
)4
ns
Glu
201(
B)
OE1
?N
23.
0A
la11
7(B
)4
ns
Glu
201(
C)
OE1
?N
23.
2Ph
e159
(B)
Glu
201(
A)
OE1
?N
22.
8V
al21
7(A
)A
sn24
3(B
)N
D2?
O7
2.8
Gly
118(
B)
Asn
243(
C)
ND
2?
O7
3.0
Phe2
00(C
)A
sn24
3(A
)N
D2?
O7
3.1
Phe2
00(B
)V
al21
7(C
)V
al21
7(B
)G
ly21
8(C
)
R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388 383
Author's personal copy
Along simulation of 4 ns the residues which make intermolecu-lar hydrogen bond and hydrophobic contacts change slightly, how-ever, until 2 ns the Glu201 and Asn243 were kept in contact with7DG. Before 3 ns the conformational profile of the binding siteseems to be changed, the intermolecular hydrogen bond betweenGlu201 and Asn243 was replaced by Ser191, Met159, andMet183. This pattern was shared by the three monomers.
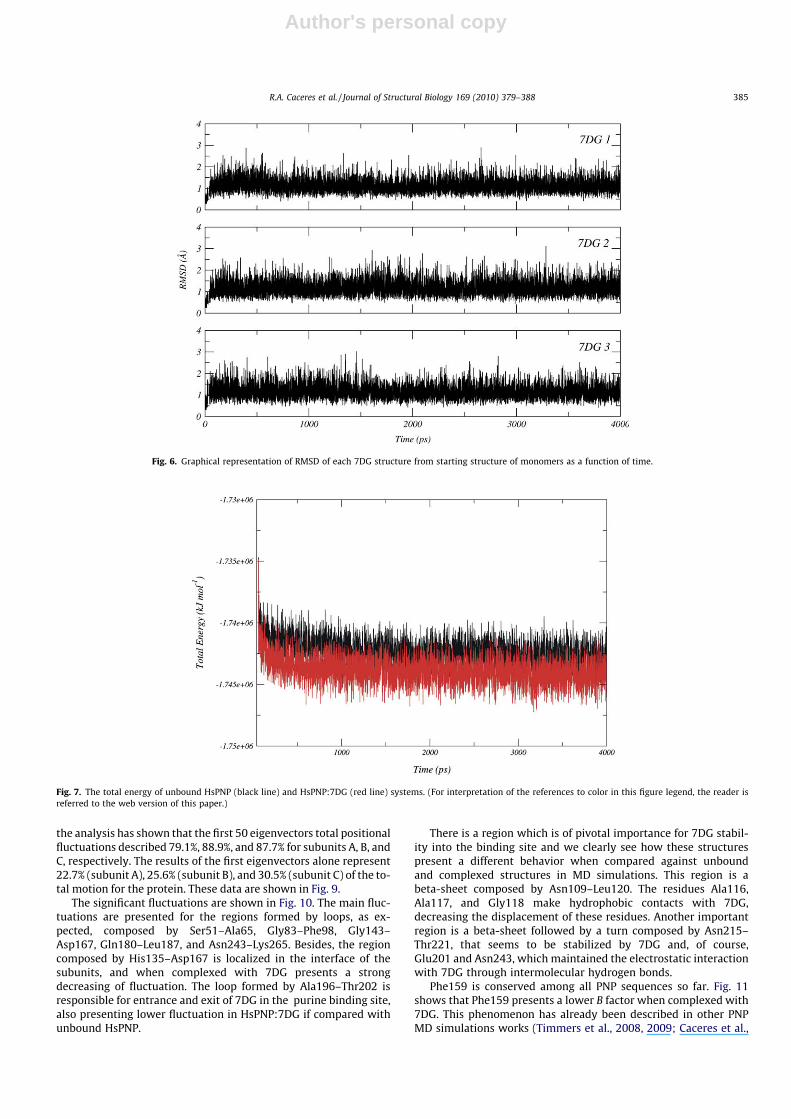
In order to compare the influence of 7DG, B factor values werecalculated for residues which form the binding site (Ala116,Ala117, Gly118, Gly218, Phe200, Glu201, and Asn243) and areshown in Fig. 5.
Despite the relative high flexibility of the purine binding site,7DG presents a conformational stability into the binding pocket.The root-mean-square deviation (RMSD) was calculated for each7DG, giving 1.1 ± 0.3, 1.1 ± 0.2, 1.1 ± 0.3 Å for each 7DG in subunitsA, B, and C, respectively (Fig. 6). This stability suggests that design-ing of 7DG analogues and addition of chemical groups may im-prove electrostatic and hydrophobic interactions with residues inthe purine binding site.
3.5. Molecular dynamics simulations
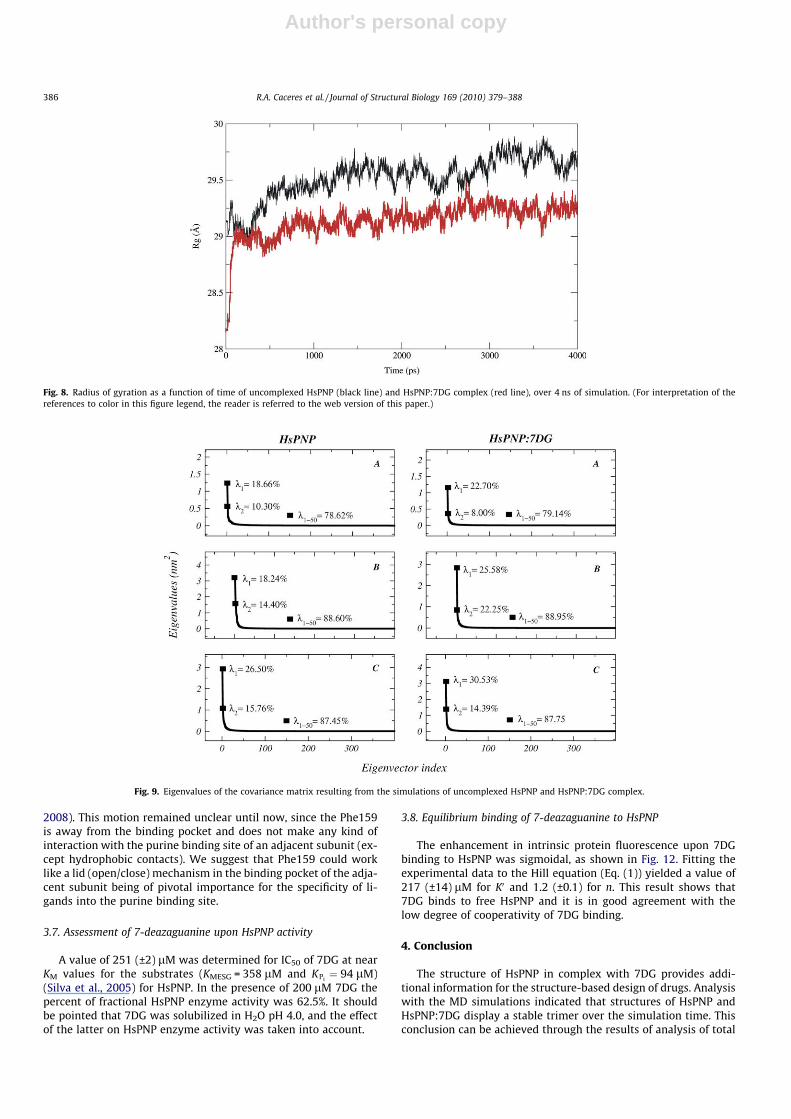
The MD simulations of free HsPNP and HsPNP:7DG binary com-plex structures were performed using SPCE water model, as previ-ously described, and applying periodic boundary conditions. Thetotal energies of both systems versus simulation time are shownin Fig. 7, which gives an indication of the overall stability of theMD trajectory. In general, the total energy of the simulations de-creased slightly but did not change to a large extent during thesimulation except for the energy of the uncomplexed system,which decreased more steeply than the HsPNP:7DG in the first100 ps. The simple association of 7DG in the trimeric structure ofHsPNP provides a DETot = 1510 kJ/mol.
The stability of the quaternary structure during the MD simula-tions was assessed by the radius of gyration (Rg). Rg as a functionof time is shown in Fig. 8. The mean values of the Rg averaged overthe period from 0 to 4 ns were determined, giving 29.5 ± 0.2 and29.1 ± 0.1 Å for the HsPNP and HsPNP:7DG, respectively. Rg re-mains essentially constant after 1000 ps for both systems, suggest-ing that the molecular conformation was significantly preserved asa whole. This analysis suggests that Rg centered on the center ofmass of the trimeric HsPNP remains essentially constant, indicat-ing that the monomers of HsPNP structure remain in the trimericstate, which is the biological unit for HsPNP.
3.6. Principal component analysis
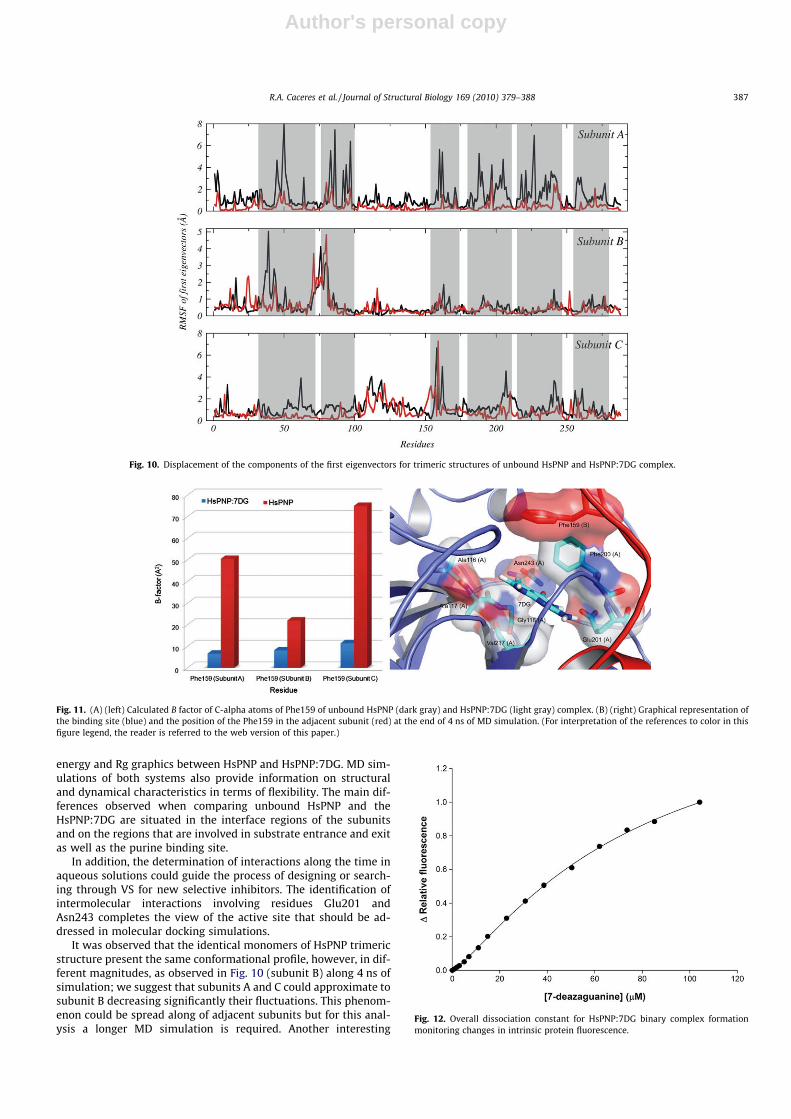
To support our results and investigate the most significant collec-tive modes of motion occurring during the simulations of theuncomplexed and complexed HsPNP, the covariance matrix corre-sponding to the C-alpha atom coordinates was calculated and PCAwas performed. By diagonalizing the covariance matrix, the anhar-monic and large-scale motions of the protein are isolated from themostly harmonic and small-scale motions. The 3N eigenvalues(864) of the covariance matrix were ranked in a decreasing orderof magnitude. The analysis of the unbound HsPNP simulation hasshown that the total positional fluctuations described by the first50 eigenvectors are 78.6%, 88.6%, and 87.4% for subunits A, B, andC, respectively. The results of the first eigenvectors alone represent18.7% (subunit A), 18.2% (subunit B), and 26.5% (subunit C) of the to-tal motion of the protein. For the HsPNP:7DG complex simulation,
Fig. 4. Intermolecular hydrogen bonds and hydrophobic contacts of HsPNP with ZINC05889995.
Fig. 5. Calculated B factor of C-alpha atoms of residues that compose the bindingsite of unbound HsPNP (blue) and HsPNP:7DG (red) complex. (For interpretation ofthe references to color in this figure legend, the reader is referred to the web versionof this paper.)
384 R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388
Author's personal copy
the analysis has shown that the first 50 eigenvectors total positionalfluctuations described 79.1%, 88.9%, and 87.7% for subunits A, B, andC, respectively. The results of the first eigenvectors alone represent22.7% (subunit A), 25.6% (subunit B), and 30.5% (subunit C) of the to-tal motion for the protein. These data are shown in Fig. 9.
The significant fluctuations are shown in Fig. 10. The main fluc-tuations are presented for the regions formed by loops, as ex-pected, composed by Ser51–Ala65, Gly83–Phe98, Gly143–Asp167, Gln180–Leu187, and Asn243–Lys265. Besides, the regioncomposed by His135–Asp167 is localized in the interface of thesubunits, and when complexed with 7DG presents a strongdecreasing of fluctuation. The loop formed by Ala196–Thr202 isresponsible for entrance and exit of 7DG in the purine binding site,also presenting lower fluctuation in HsPNP:7DG if compared withunbound HsPNP.
There is a region which is of pivotal importance for 7DG stabil-ity into the binding site and we clearly see how these structurespresent a different behavior when compared against unboundand complexed structures in MD simulations. This region is abeta-sheet composed by Asn109–Leu120. The residues Ala116,Ala117, and Gly118 make hydrophobic contacts with 7DG,decreasing the displacement of these residues. Another importantregion is a beta-sheet followed by a turn composed by Asn215–Thr221, that seems to be stabilized by 7DG and, of course,Glu201 and Asn243, which maintained the electrostatic interactionwith 7DG through intermolecular hydrogen bonds.
Phe159 is conserved among all PNP sequences so far. Fig. 11shows that Phe159 presents a lower B factor when complexed with7DG. This phenomenon has already been described in other PNPMD simulations works (Timmers et al., 2008, 2009; Caceres et al.,
Fig. 6. Graphical representation of RMSD of each 7DG structure from starting structure of monomers as a function of time.
Fig. 7. The total energy of unbound HsPNP (black line) and HsPNP:7DG (red line) systems. (For interpretation of the references to color in this figure legend, the reader isreferred to the web version of this paper.)
R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388 385
Author's personal copy
2008). This motion remained unclear until now, since the Phe159is away from the binding pocket and does not make any kind ofinteraction with the purine binding site of an adjacent subunit (ex-cept hydrophobic contacts). We suggest that Phe159 could worklike a lid (open/close) mechanism in the binding pocket of the adja-cent subunit being of pivotal importance for the specificity of li-gands into the purine binding site.
3.7. Assessment of 7-deazaguanine upon HsPNP activity
A value of 251 (±2) lM was determined for IC50 of 7DG at nearKM values for the substrates (KMESG = 358 lM and KPi
¼ 94 lM)(Silva et al., 2005) for HsPNP. In the presence of 200 lM 7DG thepercent of fractional HsPNP enzyme activity was 62.5%. It shouldbe pointed that 7DG was solubilized in H2O pH 4.0, and the effectof the latter on HsPNP enzyme activity was taken into account.
3.8. Equilibrium binding of 7-deazaguanine to HsPNP
The enhancement in intrinsic protein fluorescence upon 7DGbinding to HsPNP was sigmoidal, as shown in Fig. 12. Fitting theexperimental data to the Hill equation (Eq. (1)) yielded a value of217 (±14) lM for K0 and 1.2 (±0.1) for n. This result shows that7DG binds to free HsPNP and it is in good agreement with thelow degree of cooperativity of 7DG binding.
4. Conclusion
The structure of HsPNP in complex with 7DG provides addi-tional information for the structure-based design of drugs. Analysiswith the MD simulations indicated that structures of HsPNP andHsPNP:7DG display a stable trimer over the simulation time. Thisconclusion can be achieved through the results of analysis of total
Fig. 8. Radius of gyration as a function of time of uncomplexed HsPNP (black line) and HsPNP:7DG complex (red line), over 4 ns of simulation. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web version of this paper.)
Fig. 9. Eigenvalues of the covariance matrix resulting from the simulations of uncomplexed HsPNP and HsPNP:7DG complex.
386 R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388
Author's personal copy
energy and Rg graphics between HsPNP and HsPNP:7DG. MD sim-ulations of both systems also provide information on structuraland dynamical characteristics in terms of flexibility. The main dif-ferences observed when comparing unbound HsPNP and theHsPNP:7DG are situated in the interface regions of the subunitsand on the regions that are involved in substrate entrance and exitas well as the purine binding site.
In addition, the determination of interactions along the time inaqueous solutions could guide the process of designing or search-ing through VS for new selective inhibitors. The identification ofintermolecular interactions involving residues Glu201 andAsn243 completes the view of the active site that should be ad-dressed in molecular docking simulations.
It was observed that the identical monomers of HsPNP trimericstructure present the same conformational profile, however, in dif-ferent magnitudes, as observed in Fig. 10 (subunit B) along 4 ns ofsimulation; we suggest that subunits A and C could approximate tosubunit B decreasing significantly their fluctuations. This phenom-enon could be spread along of adjacent subunits but for this anal-ysis a longer MD simulation is required. Another interesting
Fig. 10. Displacement of the components of the first eigenvectors for trimeric structures of unbound HsPNP and HsPNP:7DG complex.
Fig. 11. (A) (left) Calculated B factor of C-alpha atoms of Phe159 of unbound HsPNP (dark gray) and HsPNP:7DG (light gray) complex. (B) (right) Graphical representation ofthe binding site (blue) and the position of the Phe159 in the adjacent subunit (red) at the end of 4 ns of MD simulation. (For interpretation of the references to color in thisfigure legend, the reader is referred to the web version of this paper.)
Fig. 12. Overall dissociation constant for HsPNP:7DG binary complex formationmonitoring changes in intrinsic protein fluorescence.
R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388 387

Author's personal copy
behavior was observed in Phe159, which seems to be involved inligand specificity. When the purine binding site is occupied by li-gand, the Phe159 of the adjacent subunit moves closer, comparedwith unbound HsPNP, which reveals a lid mechanism.
In the VS simulations, we identified a possible inhibitor pre-sumably more efficient than 7DG; since this ligand is not commer-cially available, chemical synthesis is required for enzymaticassays. However, the knowledge-based structure could supplysome clues. In conformity with Fig. 4, the presence of a highernumber of intermolecular hydrogen bonds (Ser33, Ala116,Tyr192, Met219, Ser220, Thr242, and Asn243) and hydrophobiccontacts (His86, Asn115, Phe200, Val217, Gly218, and Val260) al-lows us to suggest that ZINC05889995 could be a more effectiveinhibitor than 7DG.
Equilibrium binding and enzymatic assays revealed that 7DGcan bind to free HsPNP and that it has an inhibitory effect onHsPNP enzyme activity. Nevertheless, these data are very impor-tant although considered not so valuable by many structural biol-ogists. The structures with weakly binding ligands are moredifficult to obtain and could be very useful in the development ofempirical scoring functions by taking part of the training sets ofthese functions (Böhm and Schneider, 2003).
Deposit
The atomic coordinates and structure factors for theHsPNP:7DG structure have been deposited at the Protein DataBank, access code: 3INY.
Acknowledgments
This work was supported by the National Institute of Scienceand Technology on Tuberculosis (Decit/SCTIE/MS-MCT-CNPq-FNDCT-CAPES) and Millennium Initiative Program (CNPq), Brazil.W.F.A. (300851/98-7), D.S.S. (304051/1975-06), and L.A.B.(520182/99-5) are research career awardees of the National Coun-cil for Scientific and Technological Development of Brazil (CNPq).R.A.C., L.F.S.M.T., and I.P. would like to thank CNPq for thefellowship.
References
Amadei, A., Linssen, A.B.M., Berendsen, H.J.C., 1993. Essential dynamics of proteins.Proteins 17, 412–425.
Berendsen, H.J.C., Postma, J.P.M., van Gunsteren, W.F., Hermans, J., 1981. Interactionmodels for water in relation to protein hydration. In: Pullman, B. (Ed.),Intermolecular Forces. D. Reidel Publishing Company, Netherlands.
Böhm, H.J., Schneider, G., 2003. Protein–Ligand Interactions – From MolecularRecognition to Drug Design. Willey-VCH Verlag GmbH & Co. KGaA, Weinheim,Germany.
Bourne, P.E., Weissig, H., 2003. Structural and methods of docking and liganddesign. In: Bourne, P.E., Weissig, H. (Eds.), Structural Bioinformatics. Wiley-Liss,California.
Bzowska, A., Kulikowska, E., Shugar, D., 2000. Purine nucleoside phosphorylases:properties, functions, and clinical aspects. Pharmacol. Ther. 88, 349–425.
Caceres, R.A., Timmers, L.F.S., Dias, R., Basso, L.A., Santos, D.S., de Azevedo Jr., W.F.,2008. Molecular modeling and dynamics simulations of PNP from Streptococcusagalactiae. Bioorgan. Med. Chem. 16, 4984–4993.
Canduri, F., Teodoro, L.G., Fadel, V., Lorenzi, C.C., Hial, V., Gomes, R.A., Neto, J.R., deAzevedo Jr., W.F., 2001. Structure of human uropepsin at 2.45 Å resolution. ActaCrystallogr. D 57, 1560–1570.
Collaborative Computational Project No. 4, 1994. The CCP4 suite: programs forprotein crystallography. Acta Crystallogr. D 50, 760–763.
Darden, T., York, D., Pedersen, L., 1993. Particle mesh Ewald: an N log(N) method forEwald sums in large systems. J. Chem. Phys. 98, 10089–10092.
Delano, W.L., Lam, J.W., 2005. PyMOL: a communications tool for computationalmodels. Abstr. Pap. Am. Chem. Soc. 203, U1371–U1372.
Gelfand, E.W., Dosch, H.M., Biggar, W.D., Fox, I.H., 1978a. Partial purine nucleosidephosphorylase deficiency. Studies of lymphocyte function. J. Clin. Invest. 61,1071–1080.
Gelfand, E.W., Fox, I.H., Stuckey, M., Dosch, H.M., 1978b. Normal B-lymphocytefunction in patients with Lesch–Nyhan syndrome and HGPRT deficiency. Clin.Exp. Immunol. 31, 205–208.
Giblett, E.R., Ammann, A.J., Wara, D.W., Sandman, R., Diamond, L.K., 1975.Nucleoside-phosphorylase deficiency in a child with severely defective T-cellimmunity and normal B-cell immunity. Lancet 1, 1010–1013.
Hess, B., Bekker, H., Berendsen, H.J.C., Fraaije, J.G.E.M., 1997. LINCS: a linearconstraint solver for molecular simulations. J. Comput. Chem. 18, 1463–1472.
Hill, A.V., 1913. The combinations of haemoglobin with oxygen and with carbonmonoxide. I. Biochem. J. 7, 471–480.
Humphrey, W., Dalke, A., Schulten, K., 1996. VMD: visual molecular dynamics. J.Mol. Graph. 14, 33–38. 27–28.
Irwin, J.J., Shoichet, B.K., 2005. ZINC – a free database of commercially availablecompounds for virtual screening. J. Chem. Inf. Model. 45, 177–182.
Kalckar, H.M., 1947. Differential spectrophotometry of purine compounds by meansof specific enzymes. 1. Determination of hydroxypurine compounds. J. Biol.Chem. 167, 429–443.
Kicska, G.A., Long, L., Hörig, H., Fairchild, C., Tyler, P.C., Furneaux, R.H., Schramm,V.L., Kaufman, H.L., 2001. Immucillin H, a powerful transition-state analoginhibitor of purine nucleoside phosphorylase, selectively inhibits human Tlymphocytes. Proc. Natl. Acad. Sci. USA 98, 4593–4598.
Knapp, M., Bellamacina, C., Murray, J.M., Bussiere, D.E., 2006. Targeting cancer: thechallenges and successes of structure-based drug design against the humanpurinome. Curr. Top. Med. Chem. 6, 1129–1159.
Laskowski, R.A., MaCarthur, M.W., Moss, D.S., Thornton, J.M., 1993. PROCHECK: aprogram to check the stereochemical quality of protein structures. J. Appl.Crystallogr. 26, 283–291.
Lipinski, C.A., Lombardo, F., Dominy, B.W., Feeney, P.J., 1997. Experimental andcomputational approaches to estimate solubility and permeability in drugdiscovery and development settings. Adv. Drug. Deliv. Rev. 23, 3–25.
Ghanem, M., Saen-Oon, S., Zhadin, N., Wing, C., Cahill, S.M., Schwartz, S.D.,Callender, R., Schramm, V.L., 2008. Tryptophan-free human PNP revealscatalytic site interactions. Biochemistry 47, 3202–3215.
McRee, D.E., 2004. Differential evolution for protein crystallographic optimizations.Acta Crystallogr. D 60, 2276–2279.
Minkui, L., Lei, L., Schramm, V.L., 2008. Remote mutations alter transition-statestructure of human purine nucleoside phosphorylase. Biochemistry 47, 2565–2576.
Navaza, J., 1994. AMoRe: an automated package for molecular replacement. ActaCrystallogr. A 50, 157–163.
Oostenbrink, C., Soares, T.A., van der Vegt, N.F., van Gunsteren, W.F., 2005.Validation of the 53A6 GROMOS force field. Eur. Biophys J. 34, 273–284.
Parks Jr., R.E., Agarwal, R.P., 1972. In: Boyer, P.D. (Ed.), The Enzymes. AcademicPress, New York.
Porter, D.J., 1992. Purine nucleoside phosphorylase. Kinetic mechanism of theenzyme from calf spleen. J. Biol. Chem. 267, 7342–7351.
Schmidt, M.W., Baldridge, K.K., Boatz, J.A., Elbert, S.T., Gordon, M.S., Jensen, J.H.,Koseki, S., Matsunaga, N., Nguyen, K.A., Su, S., Windus, T.L., Dupuis, M.,Montgomery Jr., J.A., 1993. General atomic and molecular electronicstructuresystem. J. Comput. Chem. 14, 1347–1363.
Schramm, V.L., 2004. Immucillins as antibiotics for T-cell proliferation and malaria.Nucleos. Nucleot. Nucl. 23, 1305–1311.
Schramm, V.L., Grubmeyer, C., 2004. Phosphoribosyltransferose mechanisms androles in nucleic acid metabolism. Prog. Nucleic Acid Res. Mol. Biol. 78, 261–304.
Silva, R.G., Carvalho, L.P., Oliveira, J.S., Pinto, C.A., Mendes, M.A., Palma, M.S., Basso,L.A., Santos, D.S., 2003. Cloning, overexpression, and purification of functionalhuman purine nucleoside phosphorylase. Protein Expr. Purif. 27, 158–164.
Silva, R.G., Pereira, J.H., Canduri, F., de Azevedo Jr., W.F., Basso, L.A., Santos, D.S.,2005. Kinetics and crystal structure of human purine nucleoside phosphorylasein complex with 7-methyl-6-thio-guanosine. Arch. Biochem. Biophys. 442, 49–58.
Thomsen, R., Christensen, M.H., 2006. MolDock: a new technique for high-accuracymolecular docking. J. Med. Chem. 49, 3315–3321.
Timmers, L.F.S.M., Pauli, I., Caceres, R.A., de Azevedo Jr., W.F., 2008. Drug-bindingdatabases. Curr. Drug Targets 9, 1092–1099.
Timmers, L.F., Caceres, R.A., Vivan, A.L., Gava, L.M., Dias, R., Ducati, R.G., Basso, L.A.,Santos, D.S., de Azevedo Jr., W.F., 2008. Structural studies of human purinenucleoside phosphorylase: towards a new specific empirical scoring function.Arch. Biochem. Biophys. 479, 28–38.
Timmers, L.F., Caceres, R.A., Dias, R., Basso, L.A., Santos, D.S., de Azevedo Jr., W.F.,2009. Molecular modeling, dynamics and docking studies of purine nucleosidephosphorylase from Streptococcus pyogenes. Biophys. Chem. 142, 7–16.
van Aalten, D.M.F., Bywater, B., Findlay, J.B.C., Hendlich, M., Hooft, R.W.W., Vriend,G.J., 1996. PRODRG, a program for generating molecular topologies and uniquemolecular descriptors from coordinates of small molecules. J. Comput. AidedMol. Des. 10, 255–262.
van der Spoel, D., Lindahl, E., Hess, B., Groenhof, G., Mark, A.E., Berendsen, H.J., 2005.GROMACS: fast, flexible, and free. J. Comput. Chem. 26, 1701–1718.
Wallace, A.C., Laskowski, R.A., Thornton, J.M., 1995. LIGPLOT: a program to generateschematic diagrams of protein–ligand interactions. Protein Eng. 8, 127–134.
Webb, M.R., 1992. A continuous spectrophotometric assay for inorganic phosphateand for measuring phosphate release kinetics in biological systems. Proc. Natl.Acad. Sci. USA 89, 4884–4887.
388 R.A. Caceres et al. / Journal of Structural Biology 169 (2010) 379–388
Related Documents











