930 • Weed Science 51, November–December 2003 Weed Science, 51:930–939. 2003 Cropping system effects on giant foxtail (Setaria faberi ) demography: II. Retrospective perturbation analysis Adam S. Davis Corresponding author. Department of Agronomy, Iowa State University, Ames, IA 50011; present address: Department of Crop and Soil Science, Michigan State University, East Lansing, MI 48824; [email protected] Philip M. Dixon Department of Statistics, Iowa State University, Ames, IA 50011 Matt Liebman Department of Agronomy, Iowa State University, Ames, IA 50011 Cropping system characteristics affect weed management by altering key demograph- ic rates of weeds, including recruitment, seedling survival, fecundity, and seed sur- vival. To facilitate the design and improvement of cropping systems that limit weed population growth, analytical methods are needed to identify weed management ‘‘choke points’’ (weed life stages that vary in response to management and whose variation strongly affects weed population growth rate). The objectives of this study were to (1) determine whether wheat–red clover green manure can limit giant foxtail population growth rate (l) in a wheat–corn–soybean crop sequence and (2) identify choke points in the giant foxtail life cycle with respect to the green manure treatment. Demographic data were used to construct a periodic matrix model of giant foxtail population growth in a wheat–corn–soybean crop sequence, with either a wheat sole crop (W) or a wheat–red clover intercrop (R) in the wheat phase. Identification of choke points was accomplished by adapting the life-table response experiment (LTRE) design for retrospective perturbation analysis of the periodic matrix model. The difference in l (Dl) between the two treatments was decomposed into contri- butions from each parameter in each rotation phase of the periodic model. Each LTRE contribution was equal to the product of the sensitivity of l to changes in a given parameter by the treatment difference in that parameter. Those parameters making large contributions to Dl represented weed management choke points. Giant foxtail population growth rate in the simulation was more than twice as great in the W treatment (l 5 2.54) than in the R treatment (l 5 1.16). Retrospective pertur- bation analysis indicated that the proportion of seeds surviving predation in the wheat phase made the largest LTRE contribution (0.55) to Dl, followed by seedling recruitment in the soybean (0.41) and corn (0.20) phases. By identifying weed management choke points within a given system, retrospective perturbation analysis can target research and management efforts for greater reductions in weed population growth. Nomenclature: Giant foxtail, Setaria faberi Herrm. SETFA; corn, Zea mays L. ‘Pioneer 3512’; red clover, Trifolium pratense L. ‘Cherokee’; soybean, Glycine max L. ‘IA 2039’; spring wheat, Triticum aestivum L. ‘Sharp’. Key words: Life-table response experiment, simulation model, integrated weed management, seed predation, crop rotation, legume green manure. There is an increasing emphasis on weed prevention in agricultural systems in which herbicide use is minimized or eliminated (Bastiaans et al. 2000; Jordan 1996). Weed pre- vention systems seek to limit weed population growth and are predicted to be most successful when they combine sev- eral management tactics (Liebman and Gallandt 1997). Cropping system characteristics that reduce weed population growth rates may provide a weed suppressive foundation upon which to layer complementary management tactics. Because cropping system characteristics can fundamentally alter the abiotic and biotic features of the agroecosystem in which weed populations exist, they may influence demo- graphic rates throughout the entire weed life cycle. This introduces a level of complexity that is difficult to resolve without the analytical tools of demography. These tools make it possible to address complex environmental effects on weed populations with relatively simple models and to understand how changes in individual demographic rates contribute to overall weed population dynamics. Demographic analyses can aid the use of cropping system characteristics in weed management systems in at least two ways. First, population models integrate experimental results across the entire weed life cycle to predict management ef- fects on weed population growth rates (Gonzalez-Andujar and Fernandez-Quintanilla 1991; Jordan et al. 1995). Mod- els are especially useful for systems in which environmental changes affect different weed life stages in contrasting ways (e.g., a management practice that reduces germination but increases fecundity). Second, perturbation analyses (Caswell 2001) weigh the relative importance of management-in- duced variation in individual demographic parameters to overall changes in population growth rate. By identifying the life stage or stages that drive weed population dynamics under a given management scenario, perturbation analyses can focus efforts at improving the weed-suppressive qualities of a given management practice, as well as highlight other life stages to be targeted by complementary weed manage- ment tactics. The demographic analyses described here comprise the second half of an empirical modeling study of cropping sys- tem effects on giant foxtail demography. The empirical work (Davis and Liebman 2003) measured giant foxtail demo-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

930 • Weed Science 51, November–December 2003
Weed Science, 51:930–939. 2003
Cropping system effects on giant foxtail (Setaria faberi )demography: II. Retrospective perturbation analysis
Adam S. DavisCorresponding author. Department of Agronomy,Iowa State University, Ames, IA 50011; presentaddress: Department of Crop and Soil Science,Michigan State University, East Lansing, MI 48824;[email protected]
Philip M. DixonDepartment of Statistics, Iowa State University,Ames, IA 50011
Matt LiebmanDepartment of Agronomy, Iowa State University,Ames, IA 50011
Cropping system characteristics affect weed management by altering key demograph-ic rates of weeds, including recruitment, seedling survival, fecundity, and seed sur-vival. To facilitate the design and improvement of cropping systems that limit weedpopulation growth, analytical methods are needed to identify weed management‘‘choke points’’ (weed life stages that vary in response to management and whosevariation strongly affects weed population growth rate). The objectives of this studywere to (1) determine whether wheat–red clover green manure can limit giant foxtailpopulation growth rate (l) in a wheat–corn–soybean crop sequence and (2) identifychoke points in the giant foxtail life cycle with respect to the green manure treatment.Demographic data were used to construct a periodic matrix model of giant foxtailpopulation growth in a wheat–corn–soybean crop sequence, with either a wheat solecrop (W) or a wheat–red clover intercrop (R) in the wheat phase. Identification ofchoke points was accomplished by adapting the life-table response experiment(LTRE) design for retrospective perturbation analysis of the periodic matrix model.The difference in l (Dl) between the two treatments was decomposed into contri-butions from each parameter in each rotation phase of the periodic model. EachLTRE contribution was equal to the product of the sensitivity of l to changes in agiven parameter by the treatment difference in that parameter. Those parametersmaking large contributions to Dl represented weed management choke points. Giantfoxtail population growth rate in the simulation was more than twice as great in theW treatment (l 5 2.54) than in the R treatment (l 5 1.16). Retrospective pertur-bation analysis indicated that the proportion of seeds surviving predation in thewheat phase made the largest LTRE contribution (0.55) to Dl, followed by seedlingrecruitment in the soybean (0.41) and corn (0.20) phases. By identifying weedmanagement choke points within a given system, retrospective perturbation analysiscan target research and management efforts for greater reductions in weed populationgrowth.
Nomenclature: Giant foxtail, Setaria faberi Herrm. SETFA; corn, Zea mays L.‘Pioneer 3512’; red clover, Trifolium pratense L. ‘Cherokee’; soybean, Glycine max L.‘IA 2039’; spring wheat, Triticum aestivum L. ‘Sharp’.
Key words: Life-table response experiment, simulation model, integrated weedmanagement, seed predation, crop rotation, legume green manure.
There is an increasing emphasis on weed prevention inagricultural systems in which herbicide use is minimized oreliminated (Bastiaans et al. 2000; Jordan 1996). Weed pre-vention systems seek to limit weed population growth andare predicted to be most successful when they combine sev-eral management tactics (Liebman and Gallandt 1997).Cropping system characteristics that reduce weed populationgrowth rates may provide a weed suppressive foundationupon which to layer complementary management tactics.Because cropping system characteristics can fundamentallyalter the abiotic and biotic features of the agroecosystem inwhich weed populations exist, they may influence demo-graphic rates throughout the entire weed life cycle. Thisintroduces a level of complexity that is difficult to resolvewithout the analytical tools of demography. These toolsmake it possible to address complex environmental effectson weed populations with relatively simple models and tounderstand how changes in individual demographic ratescontribute to overall weed population dynamics.
Demographic analyses can aid the use of cropping systemcharacteristics in weed management systems in at least two
ways. First, population models integrate experimental resultsacross the entire weed life cycle to predict management ef-fects on weed population growth rates (Gonzalez-Andujarand Fernandez-Quintanilla 1991; Jordan et al. 1995). Mod-els are especially useful for systems in which environmentalchanges affect different weed life stages in contrasting ways(e.g., a management practice that reduces germination butincreases fecundity). Second, perturbation analyses (Caswell2001) weigh the relative importance of management-in-duced variation in individual demographic parameters tooverall changes in population growth rate. By identifyingthe life stage or stages that drive weed population dynamicsunder a given management scenario, perturbation analysescan focus efforts at improving the weed-suppressive qualitiesof a given management practice, as well as highlight otherlife stages to be targeted by complementary weed manage-ment tactics.
The demographic analyses described here comprise thesecond half of an empirical modeling study of cropping sys-tem effects on giant foxtail demography. The empirical work(Davis and Liebman 2003) measured giant foxtail demo-

Davis et al.: Giant foxtail demography • 931
graphic rates in response to green manure and tillage timingwithin a wheat–corn–soybean crop sequence in the centralUnited States. In the current study, these demographic datawere used to parameterize a matrix population model andconduct perturbation analyses. Our objectives were to (1)predict the effect of wheat–red clover green manure on giantfoxtail population growth rate within a wheat–corn–soybeancrop sequence and (2) identify weed management ‘‘chokepoints’’ in the giant foxtail life cycle (weed life stages thatvary in response to management and whose variation strong-ly affects weed population growth rate). Demographic anal-yses were used to test two hypotheses. First, compared witha wheat sole crop, wheat underseeded with red clover willlimit giant foxtail population growth rate within a wheat–corn–soybean crop sequence. Empirical studies have shownthat wheat–red clover green manure can negatively affectweed recruitment, growth, and seed survival within a sub-sequent corn crop (Conklin et al. 2002; Davis and Liebman2001, 2003). Second, differential survival of newly shed gi-ant foxtail seed will be responsible for most of the differencein population growth rate between management systems(i.e., seed survival would be the primary weed managementchoke point). In a simulation analysis of crop rotation effectsupon weed seedbanks, Jordan et al. (1995) found that over-winter survival of newly dispersed seed in the top 10 cm ofthe soil was of primary importance in determining popula-tion growth rate for velvetleaf (Abutilon theophrasti) andgreen foxtail (Setaria viridis).
Matrix models of weed population dynamics readily ac-commodate many types of life histories observed amongweed taxa and offer a straightforward way of accounting formanagement effects on weed life-stage transitions (for anexcellent overview of matrix population models, see Caswell2001). The basic equation for projecting population growthwith a matrix model is
n 5 Ant11 t [1]
where n is a vector with i rows representing the number ofindividuals in each life stage at time t and t 1 1, and Arepresents the annual projection matrix with i rows and j(5 i) columns containing all life-stage transition probabili-ties (aij) for the weed species being modeled. Such modelscan be linear or nonlinear in their parameters and deter-ministic or stochastic with respect to parameter variability.As a starting point, linear models were used because theysimplified the analysis, but the methods described here canbe applied to nonlinear models as well (Caswell 2001). Forsuch models, population growth rate can be calculated asthe dominant eigenvalue, l1, of A (for an example withcomplete MATLAB code, see Appendix A at http://www.stat.iastate.edu/preprint/AppA.pdf ). Predictions of lunder different management treatments provide a measureof the relative effectiveness of the treatments in limitingweed population growth.
In an agricultural setting, management operations oftenoccur at some subannual time scale or vary across years withthe different phases of a crop rotation. To increase the ca-pacity of matrix-modeling methods to simulate the effectsof management at sub- or supra-annual time scales, it isnecessary to use a periodic model. Periodic models accountexplicitly for variations in life-stage transition probabilitiesin a sequence of environments that repeats itself over time
(Caswell and Trevisan 1994). For a periodic model with mperiods in a complete cycle, the projection equation for aprojection interval starting at period h is
n 5 (B · · · B B · · · B B )nt11 (h21) (1) (m) (h11) (h) t
h 5 1, . . . , m [2]
where n represents a weed population vector at time t andt 1 1, and each B(h) represents a phase-specific projectionmatrix (Caswell 2001).
Perturbation analysis determines what happens to modeloutput when projection matrix parameter values arechanged. Perturbation analyses fall into two general cate-gories, prospective and retrospective, each designed to askdifferent questions (Caswell 2000). Both types of analysesrepresent important tools for ecosystem managers becausethey permit the identification of demographic processes andmanagement interventions that are most likely to havestrong regulatory control over l.
Prospective perturbation analysis, which includes sensitiv-ity and elasticity analysis, is by far the most common typeof perturbation analysis in the applied ecology literature.Sensitivity analysis examines changes in l in response toadditive perturbations to each element aij (i.e., each life-stage transition probability) of A (SA 5 ]l/]aij), whereaselasticity analysis examines changes in l in response to pro-portional perturbations to each aij (EA 5 (aij/l)(]l/]aij))(Caswell 2001). The sensitivities of l to changes in each aijcan be calculated by varying one demographic parameter ata time and recording model output, but this technique islaborious and does not allow the model user to take fulladvantage of computing power. A more direct method is tocalculate eigenvalue sensitivities to perturbations of elementsof the annual projection matrix (Caswell 2001). This pro-cedure has been extended to permit the calculation of ei-genvalue sensitivities of periodic projection matrices (Ca-swell and Trevisan 1994):
TS 5 D SB A(h) (h)
T5 (B · · · B B · · · B ) S [3](h21) (1) (m) (h11) A(h)
where S represents the sensitivity of l to changes in theB(h)elements of periodic projection matrix B(h), DT representsthe transpose of the product of the periodic projection ma-trices excluding B(h), and S represents the sensitivity of lA(h)to changes in the elements of A(h) (the annual projectionmatrix for the interval beginning at time period h). Elastic-ities of l to the elements, bhij (the life-stage transition prob-abilities for each period), of periodic projection matrices arethen computed as
E 5 (b /l)SB hij B(h) (h)[4]
The periodic matrix-modeling approach has been usedpreviously to simulate weed population dynamics in croprotations (Jordan et al. 1995; Mertens et al. 2002; Ullrich2000), using prospective perturbation analysis as a tool todescribe the dynamics of the system. In a sensitivity analysisof crop rotation effects on weed population dynamics, Jor-dan et al. (1995) found that l was most sensitive to changesin overwinter seedbank decline in the upper 10 cm of soil.By identifying overwinter seedbank decline as a parameterwith the potential to greatly influence l, this analysis indi-cated that overwinter seedbank decline represents a weed

932 • Weed Science 51, November–December 2003
management choke point (i.e., a stage in the weed life cyclewhere management interventions may be applied with thegreatest likelihood of reducing l). But what if l is found tobe highly sensitive to a demographic process that also hap-pens to be highly resistant to external manipulation? In sucha case, what appears to be a promising weed managementchoke point with regard to sensitivity analysis may be vir-tually useless in practical terms. Retrospective perturbationanalysis can help assess the utility of putative choke pointsby accounting for both sensitivity and observed variabilityin demographic parameters.
One approach to retrospective perturbation analysis is thelife-table response experiment (LTRE) (Caswell 2001). TheLTRE approximation for an annual matrix model decom-poses treatment effects on l into the sum, over all aij, ofthe product of treatment differences in aij and the sensitivityof l to changes in each aij. For an LTRE model with a one-way treatment design, the relationship between l for a treat-ment of interest (m) and l for a reference treatment (r) maybe described by
]l(m) (r) (m) (r)l 2 l ø (a 2 a )O ij ij )]aij ij A*
m 5 1, . . . , N [5]
where subscripts i and j refer to matrix rows and columns,respectively, aij refers to individual elements of the A(m) andA(r) projection matrices, and A* 5 (A(m) 1 A(r))/2. The firstterm on the right-hand side of Equation 5 denotes treatmentdifferences in parameter values, and the second term denotesthe sensitivity of l to changes in each aij, evaluated at A*.A parameter that differs only slightly between treatments mand r may still make a substantial contribution to treatmentdifferences in population growth rate if l is very sensitiveto this parameter. Thus, the LTRE provides a weighted mea-sure, under a given set of conditions, of how demographicparameters contribute to treatment effects on l.
As part of the second objective (identifying weed man-agement choke points for giant foxtail), we extended theLTRE approximation for the analysis of individual life-stagetransitions in periodic matrix population models. This ex-tension was essential because it permitted us to use the an-alytical power of the LTRE to search for weed managementchoke points within each of the phases of the crop rotation.Others have analyzed periodic models using the LTRE ap-proach (Rydgren et al. 2001) but did not analyze contri-butions of individual life-stage transitions to treatment dif-ferences in l. We believe this step to be essential for theprecise identification of weed management choke points.
The adaptation of the LTRE approach for use with pe-riodic matrix models offers some unique advantages for an-alyzing integrated weed management systems. First, themodular design and multiple matrix dimensions of a peri-odic model allow the user to simulate complex populationdynamics with a model that is not complicated. Second, theLTRE is specifically designed for the integration of empiricaland modeling efforts; the basic LTRE design is analogousto analysis of variance and may be modified to accommo-date different experimental designs (Caswell 2001). Finally,LTRE analysis of periodic models permits side-by-side com-parisons of numerous system components. For periodic sys-tems where management effects on demographic rates are
known, LTRE analysis is a powerful tool for identifyingweed life stages where management interventions may betargeted for greatest effect.
Materials and Methods
Field Experiment, Parameter Estimation, andModel Structure
Giant foxtail demography was studied in 2000 and 2001within the wheat–corn transition of a wheat–corn–soybeancrop sequence at the Iowa State University Agronomy Farm,Boone, IA. Experimental procedures are detailed in thestudy by Davis and Liebman (2003) and, thus, will be de-scribed only briefly here. In the wheat phase of the cropsequence, spring wheat was grown either as a sole crop (W)or underseeded with red clover (R) as a green manure forthe following corn crop. In fall of the wheat phase, wheatand red clover residues were incorporated to a depth of 20cm with a power takeoff-driven rototiller. Corn was plantedinto tilled residues from the wheat phase in late April in0.76-m-wide rows at 64,500 seeds ha21 and received 90 kgha21 N from urea banded in the corn row on June 5. Soy-bean was sown in 0.76-m-wide rows at 395,000 seeds ha21
in mid-May. The experiment was performed as a random-ized complete block design with four replications.
Giant foxtail seeds were buried in mesh bags in the fieldfrom November through March to permit after-ripening. Inearly April, within the corn phase of the crop sequence, 50giant foxtail seeds m22 were planted in a row parallel to,and offset 4 cm from, the crop row. Germinating seedlingswere tagged with color-coded toothpicks to identify weeklyemergence cohorts. Estimates of percent germination andseedling survival were obtained from a weekly census ofnewly emerged seedlings and dead seedlings. Before seedrain, 30 mature giant foxtail panicles per plot were enclosedin tightly woven, but highly permeable, bags made of wovenvegetable row cover. Panicle length was regressed on seedproduction by the bagged panicles. This regression relation-ship was then used to convert measurements of giant foxtailpanicle length in the rest of the plot into an estimate ofseeds per plant. Two sets of synthetic seedbanks were usedto measure seedbank survival in the top 5 cm of soil, onefor the period of October through March and the other forthe period from March through October. In October, aftertillage, sections of polyvinyl chloride air-duct pipe 7.5 cmin length and 30.5 cm in diameter were driven flush withthe soil surface. Four hundred giant foxtail seeds were thenmixed into the top 5 cm of soil. Seeds from one set ofsynthetic seedbanks were recovered using a seedbank elutri-ator (Wiles et al. 1996) in March, and seeds from the re-maining set of synthetic seedbanks were recovered the fol-lowing October. Point estimates of daily rates of postdis-persal seed predation were made within the wheat, corn, andsoybean phases using the method of Menalled et al. (2000).At several dates after seed rain had begun in August, withineach experimental unit, 200 giant foxtail seeds were placedon a 10- by 10-cm square of Bemist humidifier filter flushwith the soil surface. To determine the effects of wind andrain on seed losses from pads, a second set of seeds wasplaced within the experimental unit during the same timebut was enclosed within a cage made of fine mesh to excludepredators. Seed pads were checked on a daily basis and re-
Davis et al.: Giant foxtail demography • 933
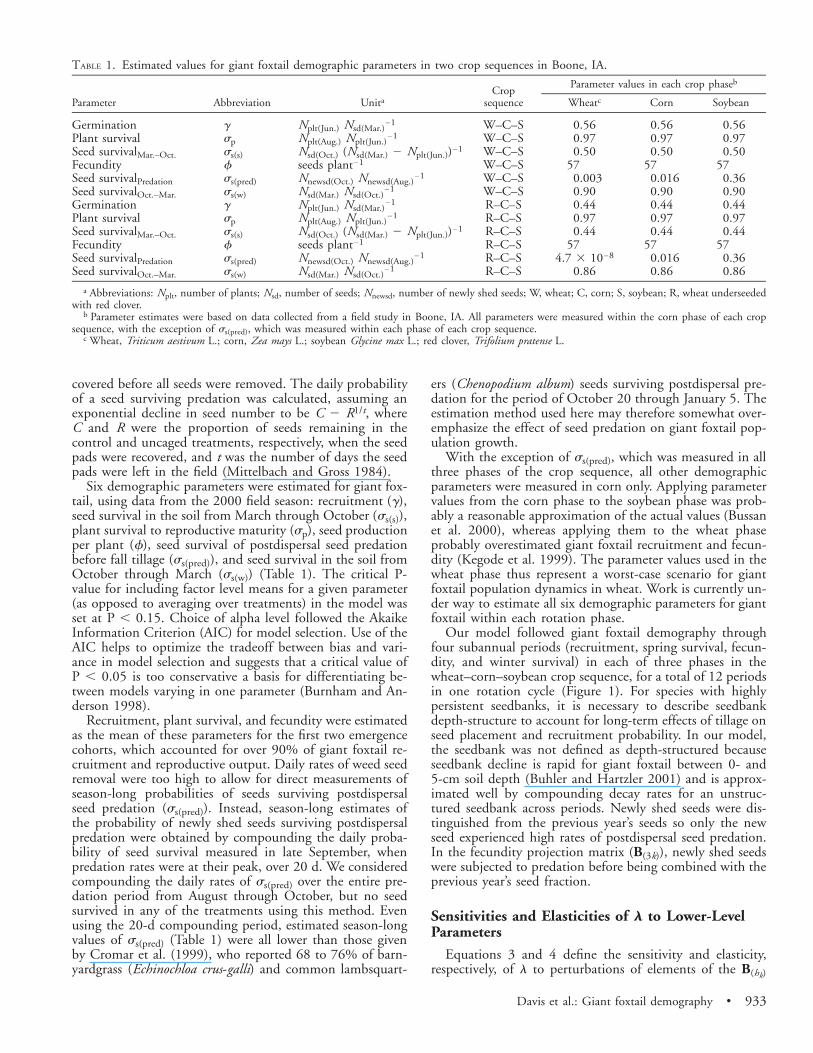
TABLE 1. Estimated values for giant foxtail demographic parameters in two crop sequences in Boone, IA.
Parameter Abbreviation UnitaCrop
sequence
Parameter values in each crop phaseb
Wheatc Corn Soybean
GerminationPlant survivalSeed survivalMar.–Oct.FecunditySeed survivalPredationSeed survivalOct.–Mar.GerminationPlant survivalSeed survivalMar.–Oct.FecunditySeed survivalPredationSeed survivalOct.–Mar.
gspss(s)fss(pred)ss(w)gspss(s)fss(pred)ss(w)
Nplt(Jun.) Nsd(Mar.)21
Nplt(Aug.) Nplt(Jun.)21
Nsd(Oct.) (Nsd(Mar.) 2 Nplt(Jun.))21
seeds plant21
Nnewsd(Oct.) Nnewsd(Aug.)21
Nsd(Mar.) Nsd(Oct.)21
Nplt(Jun.) Nsd(Mar.)21
Nplt(Aug.) Nplt(Jun.)21
Nsd(Oct.) (Nsd(Mar.) 2 Nplt(Jun.))21
seeds plant21
Nnewsd(Oct.) Nnewsd(Aug.)21
Nsd(Mar.) Nsd(Oct.)21
W–C–SW–C–SW–C–SW–C–SW–C–SW–C–SR–C–SR–C–SR–C–SR–C–SR–C–SR–C–S
0.560.970.50
570.0030.900.440.970.44
574.7 3 1028
0.86
0.560.970.50
570.0160.900.440.970.44
570.0160.86
0.560.970.50
570.360.900.440.970.44
570.360.86
a Abbreviations: Nplt, number of plants; Nsd, number of seeds; Nnewsd, number of newly shed seeds; W, wheat; C, corn; S, soybean; R, wheat underseededwith red clover.
b Parameter estimates were based on data collected from a field study in Boone, IA. All parameters were measured within the corn phase of each cropsequence, with the exception of ss(pred), which was measured within each phase of each crop sequence.
c Wheat, Triticum aestivum L.; corn, Zea mays L.; soybean Glycine max L.; red clover, Trifolium pratense L.
covered before all seeds were removed. The daily probabilityof a seed surviving predation was calculated, assuming anexponential decline in seed number to be C 2 R1/t, whereC and R were the proportion of seeds remaining in thecontrol and uncaged treatments, respectively, when the seedpads were recovered, and t was the number of days the seedpads were left in the field (Mittelbach and Gross 1984).
Six demographic parameters were estimated for giant fox-tail, using data from the 2000 field season: recruitment (g),seed survival in the soil from March through October (ss(s)),plant survival to reproductive maturity (sp), seed productionper plant (f), seed survival of postdispersal seed predationbefore fall tillage (ss(pred)), and seed survival in the soil fromOctober through March (ss(w)) (Table 1). The critical P-value for including factor level means for a given parameter(as opposed to averaging over treatments) in the model wasset at P , 0.15. Choice of alpha level followed the AkaikeInformation Criterion (AIC) for model selection. Use of theAIC helps to optimize the tradeoff between bias and vari-ance in model selection and suggests that a critical value ofP , 0.05 is too conservative a basis for differentiating be-tween models varying in one parameter (Burnham and An-derson 1998).
Recruitment, plant survival, and fecundity were estimatedas the mean of these parameters for the first two emergencecohorts, which accounted for over 90% of giant foxtail re-cruitment and reproductive output. Daily rates of weed seedremoval were too high to allow for direct measurements ofseason-long probabilities of seeds surviving postdispersalseed predation (ss(pred)). Instead, season-long estimates ofthe probability of newly shed seeds surviving postdispersalpredation were obtained by compounding the daily proba-bility of seed survival measured in late September, whenpredation rates were at their peak, over 20 d. We consideredcompounding the daily rates of ss(pred) over the entire pre-dation period from August through October, but no seedsurvived in any of the treatments using this method. Evenusing the 20-d compounding period, estimated season-longvalues of ss(pred) (Table 1) were all lower than those givenby Cromar et al. (1999), who reported 68 to 76% of barn-yardgrass (Echinochloa crus-galli) and common lambsquart-
ers (Chenopodium album) seeds surviving postdispersal pre-dation for the period of October 20 through January 5. Theestimation method used here may therefore somewhat over-emphasize the effect of seed predation on giant foxtail pop-ulation growth.
With the exception of ss(pred), which was measured in allthree phases of the crop sequence, all other demographicparameters were measured in corn only. Applying parametervalues from the corn phase to the soybean phase was prob-ably a reasonable approximation of the actual values (Bussanet al. 2000), whereas applying them to the wheat phaseprobably overestimated giant foxtail recruitment and fecun-dity (Kegode et al. 1999). The parameter values used in thewheat phase thus represent a worst-case scenario for giantfoxtail population dynamics in wheat. Work is currently un-der way to estimate all six demographic parameters for giantfoxtail within each rotation phase.
Our model followed giant foxtail demography throughfour subannual periods (recruitment, spring survival, fecun-dity, and winter survival) in each of three phases in thewheat–corn–soybean crop sequence, for a total of 12 periodsin one rotation cycle (Figure 1). For species with highlypersistent seedbanks, it is necessary to describe seedbankdepth-structure to account for long-term effects of tillage onseed placement and recruitment probability. In our model,the seedbank was not defined as depth-structured becauseseedbank decline is rapid for giant foxtail between 0- and5-cm soil depth (Buhler and Hartzler 2001) and is approx-imated well by compounding decay rates for an unstruc-tured seedbank across periods. Newly shed seeds were dis-tinguished from the previous year’s seeds so only the newseed experienced high rates of postdispersal seed predation.In the fecundity projection matrix (B(3k)), newly shed seedswere subjected to predation before being combined with theprevious year’s seed fraction.
Sensitivities and Elasticities of l to Lower-LevelParameters
Equations 3 and 4 define the sensitivity and elasticity,respectively, of l to perturbations of elements of the B(h )k

934 • Weed Science 51, November–December 2003
FIGURE 1. Periodic matrix model used to simulate giant foxtail populationgrowth under contrasting crop sequences. Abbreviations: nt, a vector of thenumber of seeds (s) and plants (p) in the population at a given point intime (t); each B represents a subannual projection matrix accounting for(h )ka single life-stage transition (h) taking place in a given phase (k) of the cropsequence; lower-level demographic parameters in projection matrices arerepresented by Greek symbols, explained in Table 1.
FIGURE 2. Projected growth of giant foxtail seedbank over six cycles of awheat–corn–soybean (W) or wheat–red clover–corn–soybean (R) crop se-quence. The symbol l represents the population growth rate over the entirecrop sequence rather than the annual population growth rate.
making up the periodic model, where h refers to the periodand k refers to the rotation phase. At the core of these pe-riodic sensitivity and elasticity calculations is a sensitivitymatrix of l with respect to A . The elements of A are(h ) (h )k kdetermined by the product of the periodic matrices over allperiods and rotation phases, starting with period h in rota-tion phase k (at the right-hand side of the product matrix),such that A 5 (B(h21)· · · B(1)B(m)· · · B(h11)B(h))k. For the(h )ksimple system described in this study, all A have only one(h )knonzero element, a11, which contains the product of all thedemographic parameters in each of the rotation phases:[(ss(w))(ss(s))(1 2 g) 1 (ss(w))(f)(ss(pred))(sp)(g)]k. The left-hand side term describes the fate of dormant seeds, and theright-hand side term describes new inputs to the seedbank.Perturbations to a11 involve this entire string of parameters,yet for management purposes, it is important to know howl responds to the perturbation of lower-level demographicparameters (i.e., individual life-stage transitions), such as gor f. This problem is solved by applying the chain rule fordifferentiation (Caswell 2001), so that for a given demo-graphic parameter x
(h )k]l ]l ]bij5 and [6]O(h ) (h ) (h )k k k]x ]b ]xi,j ij
(h ) (h ) (h )k k kx ]l x ]l ]bij5 [7]O(h ) (h ) (h )k k kl ]x l ]b ]xi,j ij
where Equations 6 and 7 describe the sensitivity and elas-ticity, respectively, of l with respect to x.
LTRE Analysis of a Periodic Matrix PopulationModel
The LTRE equation was rewritten to decompose treat-ment effects on l into the sum, over all periods and phases,of the product of treatment differences in each periodic ma-trix element bhijk and the sensitivity of l to changes in eachbhijk. The equation used to describe the LTRE approxima-tion for a periodic matrix model is
(m) (r) (m) (r) T (h ) (h )*l 2 l ø (b 2 b )D S zk kO hijk hijk A Ahijk
m 5 1, . . . , N [8]
where m is a treatment of interest, r is a reference treatment,h and k refer to period and rotation phase, respectively, iand j refer to matrix rows and columns, respectively, bhijkrefers to individual elements of the periodic projection ma-trices, DT represents the transpose of the product of thephase-specific projection matrices excluding B (see Equa-(h )ktion 3), S represents the sensitivity of l to changes inA(h )kthe elements of A *, and A * 5 (A m 1 A )r/2. The(h ) (h ) (h ) (h )k k k kmatrices A m and A r were calculated as the products of(h ) (h )k ktreatment-specific B and then averaged over A m and(h ) (h )k kA r to obtain A *.(h ) (h )k k
To simplify the presentation of the LTRE results by mak-ing LTRE contributions positive, the R treatment was des-ignated as the reference treatment (r), and the W treatmentwas designated as the treatment of interest (m). After thecontribution of each bhijk to treatment differences in l wascalculated, contributions were summed over all bhijk. Thissum was compared with l(m) 2 l(r) from the simulationmodel to obtain percentage error in the LTRE approxima-tion, calculated as (zDlLTRE 2 DlModelz)/DlModel.
Computational MethodsMatrix calculations for the example included in this study
were performed using MATLAB Version 5.0 (MathWorks1997). MATLAB was used to predict l and calculate sen-sitivity of l to changes in each bhijk within each treatment(see Appendix B, at http://www.stat.iastate.edu/preprint/AppB.pdf, for MATLAB code). Parameter contributions totreatment differences in l were calculated by multiplyingtreatment differences in bhij by the sensitivity of l to changesin each bhij.
Results and Discussion
Population GrowthProjecting the initial giant foxtail seedbank through six
rotation cycles showed population increases for the W(wheat sole crop in the wheat phase) and R (wheat under-seeded with red clover in the wheat phase) treatments (Fig-ure 2). Population growth rate, however, was more than twotimes greater in the W treatment (l 5 2.54) than in the Rtreatment (l 5 1.16), supporting Hypothesis 1 (the R treat-

Davis et al.: Giant foxtail demography • 935
TABLE 2. Elasticity of population growth rate (l) to demographic parameters of giant foxtail grown under contrasting crop sequences.
Cropsequencea
Cropphaseb
Demographic parameter
g ss(s) sp f ss(pred) ss(w)
elasticity of l
W–C–S WheatCornSoybean
2 0.600.480.96
0.700.230.02
0.300.770.98
0.300.770.98
0.300.770.98
1.01.01.0
R–C–S WheatCornSoybean
2 0.780.470.95
1.00.300.03
4.6 3 1026
0.700.97
4.6 3 1026
0.700.97
4.6 3 1026
0.700.97
1.01.01.0
a Crop sequence and demographic parameter abbreviations are described in Table 1.b Wheat, Triticum aestivum L.; corn, Zea mays L.; soybean Glycine max L.; red clover, Trifolium pratense L.
ment should limit giant foxtail population growth in awheat–corn–soybean crop sequence). These values of l werecalculated over the entire rotation cycle for use in the pe-riodic LTRE and thus were greater than the annual popu-lation growth rates in the W and R treatments (l1/3 5 1.36and l1/3 5 1.05, respectively). Figure 2 shows that in bothcrop sequences, population size declined sharply in thewheat phase, declined moderately in the corn phase, andincreased rapidly in the soybean phase. As noted earlier, re-cruitment and fecundity of giant foxtail were probably over-estimated in the wheat phase. The sharp population declinein the wheat phase is therefore primarily due to the lowprobability of seeds surviving predation in this phase in ei-ther the W or the R treatment.
It was not surprising that model projections indicatedthat the giant foxtail population grew larger (i.e., l . 1) inthe W and R treatments. No external forms of weed controlwere applied; therefore, the only constraints on giant foxtailpopulation growth were those due to cropping system ef-fects. What was surprising was that the rate of giant foxtailpopulation growth was twice as great in the W treatment asin the R treatment. Analysis of the primary experimentalresults (Davis and Liebman 2003) provided a detailed de-scription of treatment effects on individual demographicprocesses but did not integrate such effects into the overalllife cycle. This latter step is necessary for identifying weedmanagement choke points (i.e., determining the relative im-portance of treatment effects on individual life-stage tran-sitions to changes in l). Fortunately, perturbation analysisoffers a method for recovering more information from thedata. To highlight the relative merits of prospective and ret-rospective perturbation analyses for identifying weed man-agement choke points, results for elasticity analysis and themodified LTRE analysis were compared. Elasticity analysishas been the primary method of perturbation analysis usedin the earlier studies of weed population dynamics (Gon-zalez-Andujar and Fernandez-Quintanilla 1991; Jordan et al.1995; Mertens et al. 2002).
Elasticity Analysis
Relative rankings of the elasticity of l to giant foxtaildemographic parameters were the same for the corn andsoybean phases in the W and R treatments (Table 2). Elas-ticity of l to ss(w) was ranked first, sp, f, and ss(pred) wereranked second, g was ranked third, and ss(s) was rankedfourth.
Elasticity rankings for the wheat phase differed from those
in the corn and soybean phases. Within the wheat phase,elasticity rankings differed between the W and R treatments.In the W treatment, elasticity of l to ss(w) was ranked first;ss(s) was ranked second; g was ranked third (with a negativeelasticity value); and sp, f, and ss(pred) were ranked fourth.In the R treatment, elasticity of l to ss(w) and ss(s) wasranked first; g was ranked second (with a negative elasticityvalue); and sp, f, and ss(pred) were ranked third. The neg-ative elasticity of l to g in the wheat phase described asituation where seeds that gave rise to new plants actuallyreduced the population growth rate because these seeds werelost from the seedbank, but almost all seeds produced bythe new plants were consumed by predators.
The high elasticity of l to ss(w) was to be expected foran annual plant, for which seed survival during the seed-only phase represents a bottleneck that the entire populationmust pass through each year. The pattern of elasticity rank-ings for the remaining parameters yielded more informationabout how the W and R treatments differed. Elasticity rank-ings were reversed in the wheat phase compared with thecorn and soybean phases for the parameters other than ss(w).These parameters could be divided into groups: (1) param-eters affecting inputs to the seedbank (g, sp, f, and ss(pred))and (2) parameters important to the preservation of existingseeds (ss(s) and g). Recruitment (g) appears in both groupsbecause the recruitment of seedlings is necessary for seedproduction, yet 1 2 g, or dormancy, promotes the survivalof existing seeds.
In the corn and soybean phases, elasticities for seed inputparameters were ranked higher than those for seed preser-vation parameters, whereas in the wheat phase, elasticitiesfor seed preservation parameters were ranked higher thanthose for seed input parameters. It makes sense that the seedinput parameters should be important to the populationgrowth of an annual seed-bearing plant; this is the only way,besides immigration, in which the population can grow.Why then should the ranking have changed so that within-season seed preservation became more important than seedinputs in the wheat phase (especially within the R treat-ment)? The explanation can be found in the exact value ofthe elasticities of l to each of the seed input–related param-eters in the wheat phase: 4.6 3 1026. This is relatively sim-ilar to the season-long probability (4.6 3 1028) of newlyshed seeds in the wheat–red clover intercrop surviving post-dispersal seed predation. The extremely low value of ss(pred)reflected the nearly complete elimination of all inputs to theseedbank during the wheat phase in the R treatment, mak-ing the survival of those seeds that did not germinate more

936 • Weed Science 51, November–December 2003
FIGURE 3. Difference in values for giant foxtail demographic parameters inthe W and R treatments (see Figure 2 for explanation of abbreviations),where R was designated as the reference treatment (from which values ofdemographic parameters in the W treatment were subtracted).
FIGURE 4. Sensitivity of l to changes in lower-level giant foxtail demograph-ic parameters in corn, wheat, and soybean. The sensitivities were evaluatedfor a projection matrix taken as the average of the W and R treatments.
FIGURE 5. The contributions of lower-level giant foxtail demographic pa-rameters to the difference in l between the W and R treatments. Eachcontribution represents the product of the difference in parameter valuesbetween the two treatments (Figure 3) and the sensitivity of l to changesin that parameter (Figure 4).
important to long-term giant foxtail population growth (asseen in the large elasticity of l to ss(w) and in the negativeelasticity of l to g).
LTRE AnalysisUse of Akaike’s Information Criterion (Burnham and An-
derson 1998) in our parameter estimation process resultedin four of the six demographic parameters (g, ss(s), ss(w),and ss(pred)) differing between the W and R treatments (Fig-ure 3). Treatment differences in parameter values were great-est for g (0.12), followed by ss(s) (0.06), ss(w) (0.04), andss(pred) (0.003). Survival of postdispersal seed predation,ss(pred), varied between the two crop sequences only in thewheat phase.
Sensitivity of l to giant foxtail demographic parameters(Figure 4) could be divided into three groups on the basisof magnitude: (1) sensitivity of l to ss(pred) in the corn(S 5 57) and wheat (S 5 184) phases was at least anB B(h ) (h )k korder of magnitude greater than all other sensitivities, (2)sensitivity of l to g, sp, ss(s), ss(w), and ss(pred) (in soybean)was in an intermediate range (S varied from 0.66 to 4.9),B(h )kand (3) sensitivity of l to f was at least an order of mag-nitude lower than the intermediate range (S varied fromB(h )k0.005 to 0.03). Although the sensitivity of l to g in thewheat phase was intermediate in magnitude, it was distinctfrom the other sensitivity values in that its sign was negative.
The sum of contributions to treatment differences in lover all bhijk was 1.38, equal to the difference in l betweenthe two crop sequences predicted by the simulation model(Figure 2). Contributions of the bhijk to treatment differ-ences in l (Figure 5) were divided into two groups basedon sign: a negative contribution from g in the wheat phase(2 0.28) and positive contributions from the rest of theparameters, summing to 1.66. Among the positive contri-butions to l, there were two distinct groups based on mag-nitude of the contribution. Contributions of ss(pred) in thewheat phase (0.55), g in the corn (0.20) and soybean (0.41)phases, and ss(s) in the wheat phase (0.18) were at least twiceas large as all other contributions, which ranged from 0.01to 0.09. Plant survival and fecundity did not contribute totreatment differences in l. Three parameters, ss(pred) in thewheat phase and g in the corn and soybean phases, account-ed for 84% of the total contribution to l. The primaryimportance of ss(pred) as a weed management choke point
in wheat offers some support for Hypothesis 2 (that differ-ential survival of newly shed seed would account for mostof the treatment difference in l). A strict interpretation ofelasticity values implies that overwinter seed survival (withan elasticity of 1 in all rotation phases) should always be animportant weed management choke point. However, whenactual parameter variability in response to management wasfactored in, using the LTRE, it became apparent that Hy-pothesis 2 oversimplified matters because g was an impor-tant choke point in both the corn and the soybean phases.
The LTRE analysis of the periodic matrix model sharedgeneral features with the elasticity analysis. Factors that in-fluenced seed production were more important during thecorn and soybean phases of the crop sequence, and factorsthat influenced seed preservation were more important dur-ing the wheat phase. Low values for ss(pred) in the wheatphase meant that seeds would contribute more to popula-tion growth by remaining in the soil than by being recruitedinto the aboveground population, where their offspring werelikely to be consumed. What distinguished the LTRE ap-proach was the level of detail it provided. For each bhijk, theproduct of multiplying treatment differences in parametervalue by the sensitivity of l to that particular parameter gavethe contribution of that parameter to treatment differencesin l. Thus, it was possible to understand the exact degree

Davis et al.: Giant foxtail demography • 937
of importance of each of the demographic parameters tooverall population growth rate. This only holds for systemswhere detailed demographic data exist.
In the elasticity analysis, factors affecting seed inputs wereimportant in corn and soybean phases of the crop sequence,with g being slightly less important than the other param-eters. In the LTRE analysis, however, it became apparentthat of the four parameters influencing inputs to the seed-bank in the corn and soybean phases, g, sp, f, and ss(pred),only g showed any variation due to crop sequence in thesystem being studied, and, therefore, only g contributed todifferences in growth rate between the two crop sequences.Within the wheat phase, elasticity analysis revealed that var-iation in parameters affecting inputs to the seedbank, in-cluding ss(pred), would be less important than variation inparameters affecting preservation of seeds already in theseedbank.
When the experimental data were taken into account,however, the extremely low value of ss(pred) in the wheatrotation, combined with the strong regulatory role of seedsurvival in the annual plant habit, meant that additive per-turbations to ss(pred) would have a large impact on l. Thehigh sensitivity of l to ss(pred) (at S 5 184, the largestB(h )ksensitivity value in the model) in the wheat phase, multi-plied by modest differences in ss(pred) between the R andW treatments, gave rise to the largest single LTRE contri-bution (0.55) to treatment differences in l.
The large contribution made by ss(pred) in the wheatphase is in contrast to the prediction made by elasticity anal-ysis and also helps in explaining the reversal in elasticityrankings for the wheat phase compared with the corn andsoybean phases. The acute seed predation bottleneck in thewheat phase made seed preservation parameters more im-portant than those in the corn and soybean phases, whereseed production proceeded relatively unimpeded. Both gand ss(s) contributed to treatment differences in l withinthe wheat phase, although their contributions were made inopposite directions: variation in g tended to increase l inthe R treatment compared with the W treatment (thus re-ducing Dl), whereas variation in ss(s) tended to decrease lin the R treatment compared with the W treatment (thusincreasing Dl). The negative contribution made by g wasdue to the seed preservation value of the increase in dor-mancy being associated with a lower value of g in the Rtreatment than in the W treatment (Figure 3). Finally, al-though overwinter seed survival was identified by elasticityanalysis as the single parameter whose variation would mostaffect l, variation in ss(w) between treatments was small,and it made only a small contribution to treatment differ-ences in l.
The periodic LTRE performed well for the experimentalsystem being studied. It is likely that the close agreementbetween the LTRE approximation and Dl predicted by thesimulation model reflects the lack of age or stage structurein our model. In such a case, the linear approximation ofthe LTRE modeled the data very precisely. For plants withmore complex life histories, as for annuals with structuredseedbanks or perennials, greater error should be expected inthe LTRE approximation.
Elasticity and LTRE analyses were useful in understand-ing the results of the experiment, but LTRE analysis gavemuch more specific information pertaining to the experi-
mental system. The LTRE identified ss(pred) and g as weedmanagement choke points, whereas elasticity analysis onlyindicated whether factors affecting seed inputs or seed pres-ervation were important in a given rotation phase. This isnot to question the importance of elasticity analysis or pro-spective perturbation analysis in general but rather to callattention to the suitability of retrospective perturbationanalysis for analyzing experimental results in the hope thatit will be used more frequently in the future. Through theidentification of weed management choke points for a givensystem, LTRE analysis can focus research and weed man-agement efforts on weed life stages that not only vary inresponse to management but whose variation also results insubstantial changes in population growth rate. Because theresults obtained from detailed demographic investigationsare very system-specific, choke points identified for onecropping system should be treated as hypothetical when ex-trapolating to other systems.
The modeling exercise presented here was intended toelucidate how cropping system design itself could affectweed population growth. The parameter values used in thismodel were therefore obtained in the absence of externalweed control measures. Giant foxtail seedling survival ratein the absence of weed control was very high (97%), whereasunder production conditions, seedling survival rate wouldbe likely to vary between 0 and 20%, depending on thecontrol tactics used (Buhler et al. 1992; Mulder and Doll1993). A second set of simulations (results not shown) wasperformed to examine crop sequence effects on giant foxtaildemography under a production scenario in which weedcontrol with rotary hoeing and interrow cultivation resultedin only 10% of giant foxtail seedlings surviving to repro-ductive maturity. Under the low–seedling survival (LSS) sce-nario, the model projected rapid giant foxtail populationdeclines under both treatments, with a slightly more rapiddecline in the R treatment (l 5 0.055) than in the Wtreatment (l 5 0.068). Sensitivity analysis indicated that lin the LSS scenario was most sensitive to variation in ss(pred)in the wheat and corn phases, as was observed for the high–seedling survival (HSS) scenario.
The consistently high sensitivity of l to ss(pred) underboth the HSS and the LSS scenarios indicates that furtherresearch into cropping system effects on postdispersal weedseed predation should be a high priority. It is especially im-portant, for modeling purposes, to develop an improved un-derstanding of (1) the proportion of weed seeds consumedby seed predators across the entire growing season and (2)the effects of predator density and weed seed density on seedpredation rate. Simple compounding of daily seed predationrate over the period of seed rain, the method used here toestimate season-long seed predation rates, probably resultsin unrealistically high values of seed predation. To under-stand what levels of seed predation would be necessary tomaintain or reduce the giant foxtail population under theHSS and LSS scenarios, l was plotted against the mean ofss(pred) over the three phases of the crop rotation (Figure 6).The threshold values for the percentage of seeds survivingseed predation, below which the giant foxtail populationwould decrease, were approximately 3 and 4% for the Wand R treatments, respectively, in the HSS scenario, and 27and 37% for the W and R treatments, respectively, in theLSS scenario. It remains to be seen how closely these values

938 • Weed Science 51, November–December 2003
FIGURE 6. The sensitivity of giant foxtail population growth rate (l) to the mean percentage, over all rotation phases, of seeds surviving postdispersal seedpredation (ss(pred)). Results are shown for contrasting weed management scenarios, in which either (a) 97% (HSS, high seed survival) or (b) 10% (LSS,low seed survival) of giant foxtail seedlings survived to reproductive maturity. The dotted horizontal line represents constant population size over time (l5 1). Note different x-axis scales in (a) and (b).
correspond to season-long empirical measurements of giantfoxtail seed predation.
Future Directions
Our results indicate that improved knowledge of agro-nomic management effects on postdispersal seed predationwould be useful to farmers. They also indicate that, wher-ever possible, farmers should consider taking steps to in-crease rates of postdispersal seed predation, such as (1) post-poning fall tillage to allow seed predators foraging on thesoil surface sufficient time to find and consume newly dis-persed seed, (2) diversifying crop rotations with legume–small grain intercrops to provide within-field habitat for seedpredators (Davis and Liebman 2003), and (3) increasinglandscape complexity of the farming operation to improvehabitat for seed predators migrating from areas surroundingfields (Menalled et al. 2000).
Much weed management and research effort has been di-rected at killing weed seedlings. In many respects this hasbeen a successful strategy, but its very success has had theunintended consequence of obscuring other opportunitiesfor management interventions within the weed life cycle.Demographic analyses can open up new research avenues inthe development of integrated weed management systemsby (1) assessing the impact of cropping system characteristicsupon weed population growth, (2) identifying weed man-agement choke points with respect to specific managementpractices, and (3) predicting the weed management out-comes of combining complementary tactics that target dif-ferent points in the weed life cycle. By focusing attentionon combinations of practices most likely to lead to improve-ments in weed management, demographic analyses will helpset the research agenda for the next generation of integratedweed management systems.
AcknowledgmentsThe authors gratefully acknowledge Doug Buhler, Cynthia Cam-
bardella, Bob Hartzler, Tom Kaspar, Fabian Menalled, David Sund-
berg, and members of the field crew for their insight, critical feed-back, and hard work during all phases of this project. Partial fund-ing for this research was provided generously by the USDA NRICompetitive Grants Program and by the Leopold Center at IowaState University.
Literature Cited
Bastiaans, L., M. J. Kropff, J. Goudriaan, and H. H. Van Laar. 2000.Design of weed management systems with a reduced reliance on her-bicides poses new challenges and prerequisites for modeling crop-weedinteractions. Field Crop Res. 67:161–179.
Buhler, D. D., J. L. Gunsolus, and D. F. Ralston. 1992. Integrated weedmanagement techniques to reduce herbicide inputs in soybean. WeedSci. 84:973–978.
Buhler, D. D. and R. G. Hartzler. 2001. Emergence and persistence of seedof velvetleaf, common waterhemp, woolly cupgrass, and giant foxtail.Weed Sci. 49:230–235.
Burnham, K. P. and D. R. Anderson. 1998. Model Selection and Inference:A Practical Information-Theoretic Approach. New York: Springer Ver-lag. pp. 23–74.
Bussan, A. J., C. M. Boerboom, and D. E. Stoltenberg. 2000. Response ofSetaria faberi demographic processes to herbicide rates. Weed Sci. 48:445–453.
Caswell, H. 2000. Prospective and retrospective perturbation analyses: theirroles in conservation biology. Ecology 81:619–627.
Caswell, H. 2001. Matrix Population Models: Construction, Analysis andInterpretation. 2nd ed. Sunderland, MA: Sinauer. pp. 206–262.
Caswell, H. and M. C. Trevisan. 1994. Sensitivity analysis of periodic ma-trix models. Ecology 75:1299–1303.
Conklin, A. E., M. S. Erich, M. Liebman, D. Lambert, E. R. Gallandt,and W. A. Halteman. 2002. Effects of red clover (Trifolium pratense)green manure and compost soil amendments on wild mustard (Brassicakaber) growth and incidence of disease. Plant Soil 238:245–256.
Cromar, H. E., S. D. Murphy, and C. J. Swanton. 1999. Influence of tillageand crop residue on postdispersal predation of weed seeds. Weed Sci.47:184–194.
Davis, A. S. and M. Liebman. 2001. Nitrogen source influences wild mus-tard growth and competitive effect on sweet corn. Weed Sci. 49:558–566.
Davis, A. S. and M. Liebman. 2003. Cropping system effects on giantfoxtail demography: I. Green manure and tillage timing. Weed Sci.51:919–929.
Gonzalez-Andujar, J. L. and C. Fernandez-Quintanilla. 1991. Modeling thepopulation dynamics of Avena sterilis under dry-land cereal croppingsystems. J. Appl. Ecol. 28:16–27.

Davis et al.: Giant foxtail demography • 939
Jordan, N. 1996. Weed prevention: priority research for alternative weedmanagement. J. Prod. Agric. 9:485–490.
Jordan, N., D. A. Mortensen, D. M. Prenzlow, and K. C. Cox. 1995.Simulation analysis of crop rotation effects on weed seedbanks. Am.J. Bot. 82:390–398.
Kegode, G. O., F. Forcella, and B. R. Durgan. 1999. Limiting green andyellow foxtail (Setaria viridis and S. glauca) seed production followingspring wheat (Triticum aestivum) harvest. Weed Technol. 13:43–47.
Liebman, M. and E. R. Gallandt. 1997. Many little hammers: ecologicalapproaches for management of crop-weed interactions. Pages 291–343in L. E. Jackson, ed. Ecology in Agriculture. San Diego, CA: Academ-ic.
MathWorks. 1997. MATLAB. Version 5.0. Natick, MA: The MathWorks.Menalled, F. D., P. C. Marino, K. A. Renner, and D. A. Landis. 2000.
Post-dispersal weed seed predation in Michigan crop fields as a func-tion of agricultural landscape structure. Agric. Ecosyst. Environ. 77:193–202.
Mertens, S. K., F. Van Den Bosch, and J. A.P. Heesterbeek. 2002. Weed
populations and crop rotations: exploring dynamics of a structuredsystem. Ecol. Appl. 12:1125–1141.
Mittelbach, G. G. and K. L. Gross. 1984. Experimental studies of seedpredation in old fields. Oecologia 65:7–13.
Mulder, T. A. and J. D. Doll. 1993. Integrating reduced herbicide use withmechanical weeding in corn (Zea mays). Weed Technol. 7:382–389.
Rydgren, K., H. DeKroon, H. Økland, and J. Van Groenendael. 2001.Effects of fine-scale disturbances on the demography and populationdynamics of the clonal moss Hylocomium splendens. J. Ecol. 89:395–405.
Ullrich, S. D. 2000. Weed Population Dynamics in Potato Cropping Sys-tems as Affected by Crop, Cultivation and Primary Tillage. Ph.D.dissertation. University of Maine, Orono, ME. 171 p.
Wiles, L. J., D. H. Barlin, E. E. Schweizer, H. R. Duke, and D. E. Whitt.1996. A new soil sampler and elutriator for collecting and extractingweed seeds from soil. Weed Technol. 20:35–41.
Received September 30, 2002, and approved April 29, 2003.
Related Documents











![Weed control and response of yellow and white popcorn ... al-2020AgronomyJ… · species consisted of green foxtail (Setaria viridis [L.] P. 460 BARNES ET AL. TABLE 1 Common and chemical](https://static.cupdf.com/doc/110x72/6060f79536fa65272f303ced/weed-control-and-response-of-yellow-and-white-popcorn-al-2020agronomyj-species.jpg)