Arch Virol (2003) 148: 1969–1986 DOI 10.1007/s00705-003-0149-y Cotton leaf curl disease is associated with multiple monopartite begomoviruses supported by single DNA β S. Mansoor 1 , R. W. Briddon 2 , S. E. Bull 2 , I. D. Bedford 2 , A. Bashir 1 , M. Hussain 1 , M. Saeed 1 , Y. Zafar 1 , K. A. Malik 1 , C. Fauquet 3 , and P. G. Markham 2 1 National Institute of Biotechnology and Genetic Engineering (NIBGE), Faisalabad, Pakistan 2 Department of Disease and Stress Biology, John Innes Centre, Norwich, U.K. 3 ILTAB/Danforth Plant Science Center, St. Louis, Missouri, U.S.A. Received January 13, 2003; accepted May 8, 2003 Published online July 29, 2003 c Springer-Verlag 2003 Summary. For bipartite begomoviruses (family Geminiviridae) trans-replication of the DNA B component by the DNA A-encoded replication-associated protein (Rep) is achieved by virtue of a shared sequence, the “common region”, which contains repeated motifs (iterons) which are sequence-specific Rep binding sites and form part of the origin of replication. Recently cotton leaf curl disease (CLCuD), a major constraint to cotton production on the Indian subcontinent, has been shown to be caused by a monopartite begomovirus (Cotton leaf curl Multan virus [CLCuMV]) and a novel single-stranded DNA satellite molecule termed CLCuD DNA β. The satellite molecule is trans-replicated by CLCuMV but does not possess the iteron sequences of this virus. We have investigated the ability of CLCuD DNA β to interact with three further clones of monopar- tite begomoviruses, isolated from cotton, that have distinct Rep binding speci- ficities. All three cloned viruses were capable of trans-replicating the satellite molecule and inducing CLCuD symptoms in cotton, indicating that the interaction between begomovirus and DNA β is relaxed in comparison to the interaction between DNA A and DNA B components. Field surveys across all the cotton growing regions of Pakistan indicate that dual and multiple infections are the norm for CLCuD with no evidence of synergism. Despite the diversity of be- gomoviruses associated with CLCuD, only a single class of DNA β has been detected, suggesting that this satellite has the capacity to be recruited by unrelated begomoviruses.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Arch Virol (2003) 148: 1969–1986DOI 10.1007/s00705-003-0149-y
Cotton leaf curl disease is associated with multiplemonopartite begomoviruses supported
by single DNA β
S. Mansoor1, R. W. Briddon2, S. E. Bull2, I. D. Bedford2, A. Bashir1,M. Hussain1, M. Saeed1, Y. Zafar1, K. A. Malik1, C. Fauquet3,
and P. G. Markham2
1National Institute of Biotechnology and Genetic Engineering (NIBGE),Faisalabad, Pakistan
2Department of Disease and Stress Biology, John Innes Centre,Norwich, U.K.
3ILTAB/Danforth Plant Science Center, St. Louis, Missouri, U.S.A.
Received January 13, 2003; accepted May 8, 2003Published online July 29, 2003 c© Springer-Verlag 2003
Summary. For bipartite begomoviruses (family Geminiviridae) trans-replicationof the DNA B component by the DNA A-encoded replication-associated protein(Rep) is achieved by virtue of a shared sequence, the “common region”, whichcontains repeated motifs (iterons) which are sequence-specific Rep binding sitesand form part of the origin of replication. Recently cotton leaf curl disease(CLCuD), a major constraint to cotton production on the Indian subcontinent,has been shown to be caused by a monopartite begomovirus (Cotton leaf curlMultan virus [CLCuMV]) and a novel single-stranded DNA satellite moleculetermed CLCuD DNA β. The satellite molecule is trans-replicated by CLCuMVbut does not possess the iteron sequences of this virus. We have investigatedthe ability of CLCuD DNA β to interact with three further clones of monopar-tite begomoviruses, isolated from cotton, that have distinct Rep binding speci-ficities. All three cloned viruses were capable of trans-replicating the satellitemolecule and inducing CLCuD symptoms in cotton, indicating that the interactionbetween begomovirus and DNA β is relaxed in comparison to the interactionbetween DNA A and DNA B components. Field surveys across all the cottongrowing regions of Pakistan indicate that dual and multiple infections are thenorm for CLCuD with no evidence of synergism. Despite the diversity of be-gomoviruses associated with CLCuD, only a single class of DNA β has beendetected, suggesting that this satellite has the capacity to be recruited by unrelatedbegomoviruses.

1970 S. Mansoor et al.
Introduction
The begomoviruses constitute the largest genus of the family Geminiviridae,a group of plant viruses characterized by a twinned, quasi-icosahedral particlemorphology and a single-stranded, circular DNA genome [33]. The genomes ofthe vast majority of begomoviruses consist of two components [33, 27]. The firstof these components (DNA A) encodes all viral functions required for replica-tion, control of gene expression and encapsidation [38]. The second, DNA B,encodes two products involved in movement of the virus between and withinplant cells [26].
A small number of begomoviruses, mostly isolated from tomato, have monopar-tite genomes, appearing to be able to dispense with the functions encoded byDNA B. For these viruses various gene products encoded by their genomes (ahomologue of the DNA A component of bipartite viruses) appear able to take overthe functions of the DNA B products in movement [31]. The group of monopartitebegomoviruses includes Tomato yellow leaf curl virus, a major cause of losses totomato crops globally (reviewed by Czosnek and Laterrot [12]).
Cotton leaf curl disease (CLCuD) is a disorder of cotton that is transmittedby the whitefly Bemisia tabaci [17]. The disease has been shown to be associatedwith several viruses of the genus Begomovirus (family Geminiviridae) which werecollectively referred to as cotton leaf curl virus [6, 17, 18, 23, 25, 41]. The studyof Zhou et al. [41], into the diversity of begomoviruses associated with CLCuD,concluded that there were essentially four variants (represented by clones clc26,clc804a, olc311 and clc72b) of begomoviruses infecting cotton in Pakistan. Morerecent analyses of the full-length sequences of begomoviruses associated withCLCuD (utilising the 89% sequence similarity cut-off for distinguishing speciesfrom strains [14]) has concluded that the viruses analysed by Zhou et al. [41]represent three species. These have been named Cotton leaf curl Alabad virus(CLCuAV; represented by clone clc804a {CLCuAV-[804a]}), Cotton leaf curlMultan virus (CLCuMV; represented by clones clc26 {CLCuMV-[26]} and olc311{CLCuMV-[Ok]}) and Cotton leaf curl Khokhran virus (CLCuKV; representedby clone clc72b {CLCuKV-[72b]}). None of these prior studies, however, showeda causal relationship between the viruses and the disease. An infectious clone ofone of the viruses, CLCuMV, was subsequently produced and shown not to becapable of inducing characteristic leaf curl symptoms in host plants [6]. Althoughclones CLCuMV-[26] and CLCuMV-[Ok] fall into the species CLCuMV, theyhave distinct features (replication associated protein [Rep] binding sites; see laterdiscussion). For the purposes of this paper we shall henceforth refer to thesedistinct strains of CLCuMV as CLCuMVa and CLCuMVb respectively.
Recently Briddon et al. [5] demonstrated that, to induce characteristic leaf curlsymptoms in cotton, CLCuMV requires the presence of a single-stranded DNAsatellite (DNA β). DNA β is reliant on the helper begomovirus for replication,movement in plants and insect transmission, presumably by trans-encapsidationin the begomovirus’ coat protein. The function of DNA β in the infection processremains unclear, although a role in movement (analogous to DNA B), creating

Cotton-infecting begomoviruses 1971
a cellular environment suitable for virus replication and/or overcoming host de-fenses has been proposed [5, 34].
The work described in this paper, taken together with previous findings,demonstrates that four clones of begomoviruses associated with CLCuD, repre-senting three species, are monopartite and require a DNA β component to inducetypical disease symptoms in cotton.A single DNA β component has been identifiedin CLCuD isolates originating from geographically widely dispersed sites and iscapable of interacting with all four cloned viruses to induce leaf curl symptomsin cotton. This is the first report of the association of a single DNA β moleculewith multiple begomoviruses and has implications on our understanding of theevolution of such complexes, as well as to the development of cotton varietiesresistant to CLCuD.
Materials and methods
Sources of virus isolates and propagation
Cotton plants showing typical symptoms of CLCuD were collected from a field in the vicinityof Faisalabad, Pakistan, in 1992. These isolates were maintained by grafting infected scion tohealthy, susceptible cotton (var. S-12) and by whitefly transmission to healthy cotton (var. S-12). A single isolate was transmitted to tobacco (Nicotiana tabacum cv. Samsun) by grafting.All plants were maintained in insect-proof glasshouses at 25 ◦C with supplementary lightingto yield a 16 hour photoperiod.
Cloning of viruses and sequence determination
Clones pCLCUV002 and pCLCUV003 were obtained by cloning from restriction endonu-clease HindIII-digested supercoiled (sc) DNA, as previously described for Beet curly topvirus [8]. The linearised molecules were ligated into pBluescript II SK(+/−) (Stratagene).Partial repeats of pCLCUV002 and pCLCUV003 were produced by cloning approx 550 bpHindIII – EcoRV or 1100 bp HindIII – EcoRI fragments into pBluescript II SK(+/−) toyield clones pCLCUV002[0.2] and pCLCUV003[0.4] respectively. Subsequently the full-length, HindIII fragments of the relevant clones were ligated into the unique HindIII sites ofpCLCUV002[0.2] and pCLCUV003[0.4] to yield clones pCLCUV002[1.2] and pCLCUV003[1.4] respectively.
A further clone (pCLCUV004) was produced by polymerase chain reaction (PCR)-mediated amplification, from N. tabacum graft inoculated with a CLCuD isolate originatingfrom Faisalabad (Pakistan), essentially as described in Briddon et al. [6]. The primers usedwere designed around a unique ClaI restriction endonuclease site identified in the publishedsequence of CLCuAV-[804a] ([41]; EMBL accession no. AJ002452; virion-sense primer [Pak4F] 5′-TTTATCGATCTGAAATGTGCCCCAGTC-3′, complementary-sense primer [Pak 4R]5′-TTTATCGATGGAAGATCTGCAAGAGGA-3′).Amplification products were ligated intothe pGEM-T vector (Promega).
The complete nucleotide sequences of all three full-length clones were determined bydideoxynucleotide chain termination sequencing using the PCR-based “BIG DYE” kit (PerkinElmer Cetus) and specific internal primers (Genosys). Reaction products were resolved on anABI 3700 automated sequencer. Sequence information was stored, assembled and analysedusing Version 10.2 of the program library of the Genetics Computer Group [13].

1972 S. Mansoor et al.
Strategy used to identify potential DNA B components
The techniques used to identify potential DNA B components associated with cotton-infectingbegomoviruses have been described previously [6]. These included PCR using the DNA Bspecific primers reported by Rojas et al. [29], PCR with primers designed to the intergenicregion (equivalent to the common region shared by DNA A and DNA B components) andrestriction endonuclease digestion of purified super-coiled DNA to identify two populationsof molecules.
Detection of cotton leaf curl viruses and DNA β in cotton
Cotton samples showing typical CLCuD symptoms were collected from various locationsin Pakistan (Table 1). Total nucleic acids were extracted from leaf tissue by the CTABmethod, resolved in duplicate 0.5% agarose gels using TNE buffer and blotted to Hybond NX(Amersham) membranes. A full-length PCR product of CLCuKV (produced using primersPak3F and Pak3R), a full length clone of CLCuD DNA β (pCLCβ02; [5]) and a DNA β
molecule (produced by PCR-mediated amplification with the primers described by Briddon
Table 1. PCR-Mediated detection of four begomoviruses in CLCuD-affected cotton plantscollected from cotton-growing areas of Pakistan
No.∗ Location Year Species detected
CLCuMVa CLCuMVb CLCuKV CLCuAV
1 Multan 1992 + + − −2 Faisalabad 1993 + + + −3 Chichawatni 1996 − − + −4 Mianchannu 1996 + − − −5 Rahim Yar Khan 1996 + + − −6 Bahawalpur 1996 + + − −7 Faisalabad 1996 − + − −8 Dera Ghazi Khan 1997 + + + +9 Dera Ghazi Khan 1997 + + + −
10 Rahim Yar Khan 1997 + + − −11 Rahim Yar Khan 1997 + + − −12 Multan 1997 + + + −13 Multan 1997 + − + +14 Dera Ghazi Khan 1997 + + + +15 Multan 1997 + + − +16 Faisalabad 1997 + − − −17 Multan 1997 + + + −18 Multan 1997 + + + −19 Ghotki 1997 + − − +20 Sukkur 1997 + − − +21 Rahim Yar Khan 1998 − + − −22 Faisalabad 1998 + + + −23 Multan 1998 + + + +
∗The sample number corresponds to the lane number in Fig. 4. Samples 21 to 23 wererun on a separate gel (results not shown)

Cotton-infecting begomoviruses 1973
et al. [4]) from leaf curl disease affected okra originating from Pakistan [20] were radioactively-labelled with 32P-dCTP (NEN) by oligo-labelling. Duplicate blots were hybridised with theseprobes and were washed at intermediate stringency (1 × SSC, 50 ◦C) for the CLCuKV probeand high stringency (0.1 × SSC, 65 ◦C) for the CLCuD DNA β and okra leaf curl DNA β
probes.
Detection of multiple infections in cotton
Primers specific for four begomovirus species associated with CLCuD were used to assess theprevalence of multiple infections in field isolated cotton. A specific primer pair for the PCR-mediated detection of CLCuMVa was designed to the sequence at the overlap of the Rep andC2 genes (virion sense primer [C1Pak1F]; 5′-GCTTTCCATGGGTTGTAGTTGAACTGGAT-3′) and the 5′ end of the Rep gene (complementary-sense primer [C1Pak1R]; 5′-AAGTACCATGGCCTCCCCCCAAACGTTTTAA-3′). These primers amplify an approximately 1300 bpfragment of the virus.
Primers for the detection of CLCuAV were designed to the intergenic region (Pak-4R,complementary-sense; 5′TTAAGCTTTGAGTCTGCATCGTCGTG CG3′) and the 5′ endof the C3 gene (VBTB, virion-sense; 5′-TGATGAGTTC CCTGTGCGTGAATCCATGGTTGT-3′).
The design of a multiplex PCR procedure for the specific detection of CLCuMVa andCLCuKV has been reported earlier [22]. Briefly, sequences conserved between these twospecies were identified in the Rep gene to which a primer was designed (CLCuV-F, virion-sense; 5′-CTGTCTAGATTTGCATTTAAATTATGAAATTG-3′). Primers (PCL2 andCLCuVPk1-C442) specific for CLCuKV and CLCuMVa were designed to the 5′ end of the repgene (complementary-sense; 5′-CATGCCTCCAAAGCGGAACGGTATTTATT-3′) and the5′ end of the C4 gene (complementary-sense; 5′-CGACCATGGGAGCCCTCATCTCCATGTGC-3′) respectively. These primers amplify fragments of approximately 510 bp and 360 bprespectively.
A CLCuMVb specific primer was designed to the intergenic region 5′ of the hairpin in thecomplementary sense (CRPak2R, complementary-sense; 5′-TGGAGCGTGAGGATTTTGAAATGATTTCTCA-3′) and was used in combination with primer CLCuV-F.
Inoculation of plants with cloned components and insect transmission
Young N. benthamiana and cotton (var. S-12) plants at the 3–4 leaf stage were inocu-lated with cloned viruses and DNA β by a biolistic procedure as described previously [6].Gold particles (0.95 µM) were coated with clones pCLCUV004 (from which the insertwas excised using ClaI) and pCLCβ002 (from which the insert was excised using KpnI).Clones pCLCUV002[1.2] and pCLCUV003[1.4] were linearised with SstI before coatingonto gold particles with KpnI digested pCLCβ02 [5]. Following inoculation, cotton plantswere maintained in a growth cabinet at 35 ◦C with a 16 h photoperiod and N. benthamiana inan insect proof glasshouse at 25 ◦C with supplementary lighting to give a 16 h photoperiod.Plants were checked daily for the appearance of symptoms.
Cotton seedlings, infected with the cloned components of CLCuD by biolistic inoculation,were placed in perspex cages and exposed to approximately 1000 Bemisia tabaci. Followinga 48 h acquisition access period, 4 healthy cotton seedlings were placed in the cage andthe source plant was removed. After approximately 10 days the plants were removed fromthe cage, sprayed with insecticide and maintained in an insect-proof glasshouse at 25 ◦Cwith supplementary lighting to give a 16 h photoperiod. Plants were checked daily for theappearance of symptoms.

1974 S. Mansoor et al.
Results
Isolation of begomovirus clones
Full-length clones of three begomoviruses isolated from cotton originating fromPakistan were produced. Clones pCLCUV002 and pCLCUV003 were isolatedfrom purified supercoiled DNA and cloned at unique HindIII restriction endonu-clease sites. Clone pCLCUV004 was produced by PCR-mediated amplificationwith primers designed around a unique ClaI restriction site from nucleic acids iso-lated from Nicotiana tabacum infected with CLCuD by graft inoculation. Sourceplants from which these clones were obtained showed the typical symptoms ofCLCuD (leaf curling, vein darkening, vein swelling and enations on the undersideof leaves which frequently develop into cup-shaped, leaf-like structures) in cotton.The symptoms in graft-inoculated N. tabacum were qualitatively similar to thosein cotton but also showed severe twisting of both the stem and petioles (Fig. 3)and, rather than producing cup-shaped, leaf-like enations, this species producedfrond-like, cup-shaped enations along the main veins.
Attempts to show the presence of a DNA B component for all three of theCLCuD isolates analysed were negative. Although fragments were amplified withthe universal DNA B primers from this material, sequence analysis followingcloning showed no relationship of the amplified molecules to any of the be-gomoviruses (results not shown).
Sequence analysis
The complete nucleotide sequences of clones pCLCUV002, pCLCUV003 andpCLCUV004 were determined. These sequences are available in the EMBL,DDJB and GenBank databases under accession numbers AJ496286, AJ496287and AJ436992 respectively. The clones were 2750 nt, 2750 nt and 2745 nt inlength. All three clones have the typical genome arrangement of the monopartitebegomoviruses (equivalent to the DNAA components of bipartite begomoviruses)consisting of 6 positionally and sequence conserved genes (Fig. 1); two in thevirion-sense (V2 and the coat protein) and four in the complementary-sense(Rep, C2 , C3 and C4). The precise functions of the products of genes V2 andC4 remain unclear. For bipartite begomoviruses the C2 and C3 genes encodethe transcriptional activator protein and the replication enhancer protein respec-tively. The functions of these genes for monopartite begomoviruses has yet to beestablished.
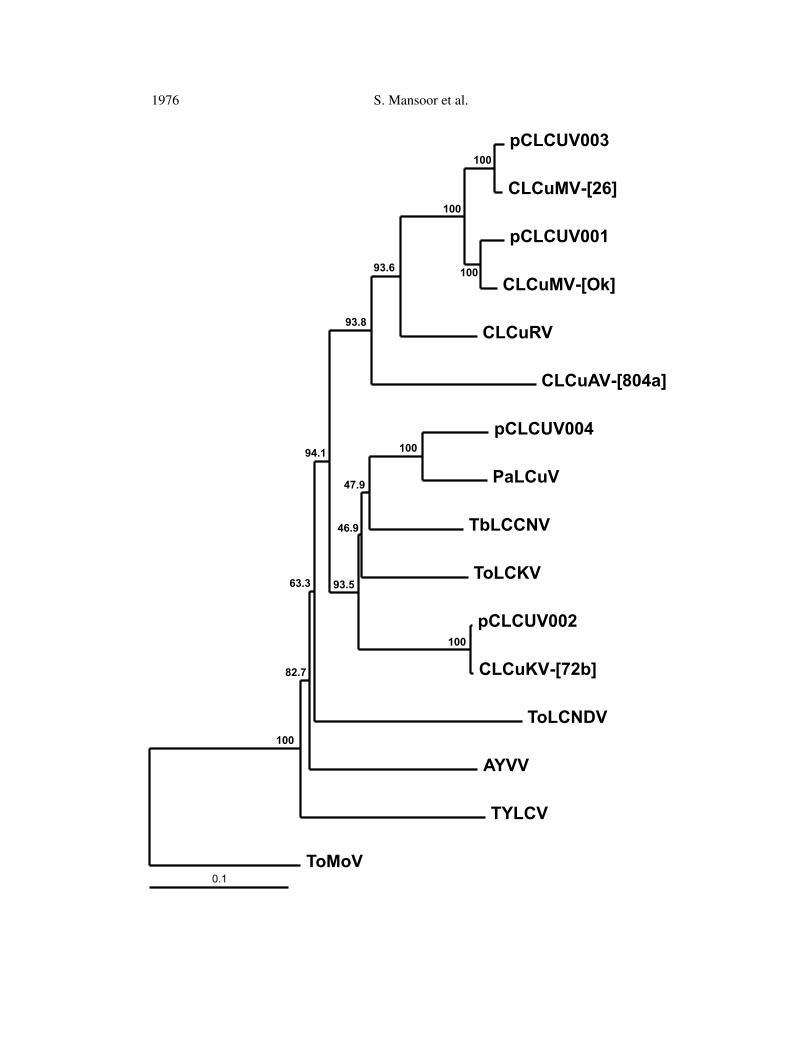
Phylogenetic analyses based upon alignments of the complete nucleotidesequences (Fig. 2) show clones pCLCUV002 and pCLCUV003 to be very similarto and segregate with clones CLCuKV-[72b] (99.6% nucleotide sequence identity)and CLCuMV-[26] (98.8% nucleotide sequence identity) respectively.Alignmentsof the nucleotide sequences of the intergenic regions of these clones with selectedother cotton-isolated begomoviruses originating from the Indian subcontinentindicate that the predicted “iterons” of pCLCUV002 and pCLCUV003 are iden-tical to those of CLCuKV-[72b] and CLCuMV-[26] respectively. These findings

Cotton-infecting begomoviruses 1975
Fig. 1. Typical genome arrangement of the four begomovirus species. The positions andorientation of the 6 conserved (for New World begomoviruses) genes are shown. The positionof the nonanucleotide motif is shown with a black dot. The arrow outside the circle denotesthe position of the sequences of 10 CLCuD associated begomovirus clones shown in thebox. The position of the complementary-sense TATA box is indicated. The predicted iteronsequences, within the intergenic region, for each of the clones is underlined. The numbers in
brackets refer to the clone numbers in Zhou et al. [41]
demonstrate that pCLCUV002 and pCLCUV003 are clones of the begomovirusspecies CLCuKV and CLCuMVa respectively.
In contrast, pCLCUV004 shows the highest levels of nucleotide sequenceidentity (90.6%) to Papaya leaf curl virus (PaLCuV) originating from India [36]and segregates with this virus in the phylogenetic comparisons (Fig. 2). Thepredicted iteron sequences of pCLCUV004 are identical to those of PaLCuV.Clone pCLCUV004 thus represents a distinct strain of PaLCuV for which wepropose the descriptor PaLCuV-[Cot].
Additionally, the alignments of the intergenic sequences and phylogeneticanalyses include the cotton-isolated clone pCLCUV001, previously shown to becapable of inducing typical CLCuD symptoms in cotton in the presence of CLCuDDNA β [5]. This clone shows the highest levels of sequence similarity to CLCuMV(97% nucleotide sequence similarity to CLCuMV-[Ok] of Zhou et al. [41]). Thepredicted iteron sequences of pCLCUV001 are identical to those of CLCuMVb,showing this clone to represent the species CLCuMVb. The alignment also showsthe predicted iteron sequences of CLCuMVa and CLCuMVb differ whereas thoseof PaLCuV and CLCuAV are identical.
1976 S. Mansoor et al.

Cotton-infecting begomoviruses 1977
Infectivity of begomoviruses with CLCuD DNA β
The results of biolistic inoculation of the three cotton isolated clones to both cottonand N. benthamiana, in the presence of CLCuD DNA β (clone pCLCβ02; [5]),is shown in Table 2. All three clones were infectious to both the experimentalhost N. benthamiana and to cotton. Symptoms of infection for N. benthamianatypically appeared within 12 days of inoculation. The symptoms induced inall cases were downward leaf curling, swelling of the veins on the undersidesof leaves and vein yellowing with some minor swelling of the veins on theupper leaf surface (Fig. 3). The yellowing of veins on the upper leaf surface wasmore pronounced for infections with pCLCUV003 than the other clones. Initiallyinfected N. benthamiana plants showed little chlorosis with a mild yellow mottlingdeveloping within a week of the first symptoms. These symptoms are very similarto the symptoms reported for infection of N. benthamiana with AYVV in thepresence of AYVD DNA β [34] with the exception that, for the cotton clones, thevein yellowing symptoms were more pronounced.
Symptoms of infection in cotton were initially downward cupping of theyoungest leaf approximately 18 days post inoculation. All subsequently
Table 2. Infectivity of clones of CLCuD-associated begomoviruses by biolistic inoculationwith CLCuD DNA β
Clone Infectivity (number of plants infected/number of plants inoculated)
Nicotiana benthamiana Cotton
Exp. I Exp. II Exp. III Exp. I Exp. II Exp. III
pCLCUV002 3/10 3/10 2/10 7/13 5/16 3/15pCLCUV003 5/14 6/10 4/10 1/15 3/14 5/18pCLCUV004 3/10 4/10 3/10 3/27 4/16 3/15
�Fig. 2. Neighbor-joining phylogenetic dendrogram based upon alignments of the completenucleotide sequences of the genomes (or DNAA genomic components) of CLCuD-associatedand selected other begomoviruses. Vertical distances are arbitrary, horizontal distances areproportional to calculated mutation distances. Numbers at nodes indicate percentage bootstrapscores (1000 replicates). The viruses included in the analysis were Tomato leaf curl Karnatakavirus (ToLCKV; accession no. U38239), Tomato leaf curl New Delhi virus (ToLCNDV; acc.no. U15016), Ageratum yellow vein virus (AYVV; AF314144), Tomato yellow leaf curl virus(TYLCV; X15656) Tobacco curly shoot virus (TbCSV; acc. no. AJ420318), Cotton leafcurl Rajasthan virus (CLCuRV; acc. no. AF363011), Cotton leaf curl Alabad virus-[804a](CLCuAV-[804a]; acc. no. AJ002455), Cotton leaf curl Multan virus-[26] (CLCuMV-[26];acc. no. AJ002458) Cotton leaf curl Multan virus-[Ok] (CLCuMV-[Ok]; acc. no. AJ002459),Cotton leaf curl Kokhran virus-[72b] (CLCuKV-[72b]; acc. no. AJ0024480 and Papaya leafcurl virus (PaLCuV; Y15934). The tree was rooted on the sequence of Tomato mottle virus(ToMoV) DNA A component (accession no. M90494), a distantly related (to begomovirusesof the Old World) bipartite begomovirus

1978 S. Mansoor et al.

Cotton-infecting begomoviruses 1979
developing leaves then showed either upward or downward leaf curl, vein swelling(on the underside of leaves), vein darkening and the formation of enations on veinson the undersides of leaves (Fig. 3). In a small number of cases enations developedinto cup-shaped, leaf-like structures on the main veins (Fig. 3). These symptomsare typical of CLCuD [7] and have been reported previously for pCLCUV001with CLCuD DNA β in cotton [5].
The three clones of CLCuD-associated begomoviruses were also shown to beable to infect N. benthamiana in the absence of CLCuD DNA β. Typically 2 out of10 plants inoculated showed symptoms of infection approximately 12 days post-inoculation. The symptoms induced by all three clones were qualitatively similarto those of infection with CLCuD DNA β, although the leaf curling, rugosityand vein yellowing were less pronounced (Fig. 3). No infections of cotton weredetected, following biolistic inoculation with any of the three clones, in the absenceof DNA β.
Of the three clones analysed only pCLCUV002 was whitefly transmitted fromcotton plants, infected by biolistic inoculation, to healthy test cotton seedlings.Two of four test cotton seedlings showed typical symptoms of CLCuD approxi-mately three weeks after exposure to viruliferous insects. The symptoms exhibitedby these plants were mild leaf curling and vein darkening.
Field survey
Samples of cotton showing the typical symptoms of CLCuD were collected fromselected sites across all cotton growing regions of Pakistan and subjected toPCR analysis using primers specific for four begomoviruses. The results of thisanalysis are presented in Table 1. Of 23 cotton samples analysed, 18 showed thepresence of more than one begomovirus species, of which three plants showed thepresence of all four viruses. The most commonly detected species was CLCuMVa
(detected in 20 samples) while CLCuAV was detected in the fewest samples (7).Sequence analyses suggest that the primers used to detect CLCuAV would alsoamplify PaLCuV. Prior to this analysis PaLCuV had not previously been detectedin cotton and this cross-reactivity was not considered. However, in light of thedemonstration that PaLCuV can cause CLCuD (in the presence of CLCuD DNAβ), a positive reaction with the CLCuAV primers could indicate the presence ofeither CLCuAV or PaLCuV in this PCR-based analysis.
As well as by PCR, nucleic acids extracted from field collected cotton sampleswere also examined by Southern blot analysis (Fig. 4). Duplicate blots were
�Fig. 3. Symptoms induced by infection of Nicotiana tabacum with a wild type isolateof CLCuD by graft inoculation (A). Nicotiana benthamiana plants infected by biolisticinoculation with clones pCLCUV003 and CLCuD DNA β (B) and clone pCLCUV003 only(D) compared to a healthy plant (C). Foliar symptoms of cotton induced by infection withclones pCLCUV003 and CLCuD DNA β (E) compared to a leaf from a healthy plant (F)photographed under transmitted light

1980 S. Mansoor et al.
Fig
.4.
Dup
licat
eSo
uthe
rnbl
ots
prob
edw
ithra
dioa
ctiv
ely-
labe
led
prob
esof
CL
CuK
V(A
)an
dC
LC
uDD
NA
β(B
).N
ucle
icac
idsa
mpl
esel
ectr
opho
rese
din
lane
s1
to20
wer
eex
trac
ted
from
field
colle
cted
cotto
nsa
mpl
es,
orig
inat
ing
from
acro
ssal
lco
tton
grow
ing
regi
ons
ofPa
kist
an,s
how
ing
typi
cals
ympt
oms
ofC
LC
uD

Cotton-infecting begomoviruses 1981
probed with a full-length PCR product of the begomovirus CLCuKV, CLCuDDNA β and an as yet uncharacterised DNA β molecule originating from okra. TheCLCuKV probe hybridised to all samples examined. At the washing stringencyemployed (an intermediate stringency) this probe would be expected to detectall begomoviruses and a positive reaction can thus only be taken to signify thepresence of a begomovirus. In contrast, the CLCuD DNA β probed blots werewashed at high stringency and would thus be expected to only detect closelyrelated molecules. Again all cotton samples gave a positive reaction to this probe.DNA β molecules contain an approximately 80 bp region of near sequence identity(the “satellite conserved region” [3]) which might be expected to cross-hybridisebetween distinct DNA β molecules. To investigate this possibility the cottonsamples were hybridised to a DNA β molecule isolated from okra and againwashed at high stringency. No hybridisation was detected (results not shown),indicating that, under the conditions employed, the CLCuD DNA β probe wasonly detecting the DNA β isolated from cotton.
Discussion
DNA β is a single-stranded DNA satellite which is required by some be-gomoviruses to symptomatically infect their natural hosts. Two DNA β moleculeshave thus far been characterised, that associated with CLCuD [5] and a furthersatellite associated with Ageratum yellow vein disease (AYVD; [34]). Like thesatellite molecules associated with a number of RNA viruses (reviewed by Collmerand Howell [11]), DNA β relies upon its helper begomovirus for replication, move-ment within plants and transmission between plants [5, 34]. We have shown thatthe DNA β component associated with CLCuD is able to productively interact withfour distinct monopartite, cotton-infecting begomoviruses to induce characteristicleaf curl symptoms in cotton [5; this study].
The replication of geminiviruses occurs by a rolling circle mechanism withthe origin of replication consisting of a conserved stem loop structure, with theloop sequence TAATATTAC, which is nicked by the virus-encoded replication-associated protein (Rep) during the initiation of replication, and a number ofupstream repeat sequences (“iterons”) which are the sequence-specific bindingsites for Rep [1, 10, 16]. For bipartite begomoviruses, trans-replication of theDNA B component is achieved by virtue of a shared sequence, the “commonregion”, which is present on both components and contains the stem loop structureand the iterons. The Rep-iteron interaction is highly sequence specific [9, 16]such that interspecific replication can occur only if iterons are conserved or verysimilar [35]. In contrast to DNA B components, the DNA β components do notcontain the iterons of their respective helper begomoviruses suggesting that someother motif(s), cryptic Rep binding sites, act to facility trans-replication of thesemolecules [5]. Our demonstration that four cotton-infecting begomoviruses, eachwith distinct Rep binding sites, are able to productively interact with CLCuD DNAβ indicates that there is a relaxed relationship between DNA β molecules andbegomoviruses, rather than the specific interaction characteristic of begomovirus

1982 S. Mansoor et al.
DNAA and DNA B components. Furthermore, these results provide circumstantialevidence to suggest that the symptoms characteristic of CLCuD are a feature ofCLCuD DNA β rather than of the cotton-infecting begomoviruses with which itis associated.
CLCuMVb is the only monopartite begomovirus for which a causal relation-ship with CLCuD has previously been shown and this virus requires the satelliteDNA β to induce symptoms in cotton. Here we demonstrate that two furtherbegomovirus species (CLCuKV and PaLCuV), isolated from field collected cottonsamples, are involved in the CLCuD epidemic. CLCuMV and CLCuKV havepreviously been shown to be associated with diseased cotton but no infectiousclones were produced to prove a causal relationship [41]. This is also the first reportof PaLCuV being associated with CLCuD and able to induce typical symptoms incotton experimentally, in the presence of CLCuD DNA β. PaLCuV occurs widelyacross the Indian subcontinent in papaya (Carica papaya, Caricaecae; [36]).
As well as the species already mentioned (CLCuKV, CLCuMV and PaLCuV),several other begomovirus species have been implicated in CLCuD, including athird species resulting from the report by Zhou et al. [41], represented by cloneCLCuAV-[804a] and a species originating from Rajasthan, India (Cotton leaf curlRajasthan virus [CLCuRV]).
These findings make it clear that the epidemic of CLCuD occurring acrossPakistan and western India has a complex genetic structure, involving numerousdistinct begomovirus species. The involvement of more than two begomovirusesin a disease epidemic is unique for geminiviruses. Although a number of symp-tomatically indistinguishable diseases are known to be caused by geneticallydistinct begomoviruses, these viruses usually have different geographical hostranges. For example, cassava mosaic disease (CMD) in Africa is associated withsix distinct begomovirus species, African cassava mosaic virus (ACMV; [39]),East African cassava mosaic virus (EACMV; [19]), East African cassava mosaicCameroon virus [15], East African cassava mosaic Malawi virus [15], SouthAfrican cassava mosaic virus [2] and East African cassava mosaic Zanzibarvirus [24] which occur in different geographic areas with little overlap. However,the ongoing epidemic in eastern Africa has been shown to be associated withtwo begomovirus species, ACMV and EACMV (specifically EACMV-Uganda, anatural recombinant between ACMV and EACMV) [40, 28]. The severity of theepidemic is attributed to a synergistic interaction between the two species [15, 28].For CLCuD, despite the frequency of dual and multiple infection of cotton plantsin the field, no apparent synergism is evident. The severity of symptoms appearsmore related to the cultivar of cotton infected, the level of whitefly infestation andthe age at which plants were infected than to the species/strain of virus associatedwith the infection [7, 29, 37].
The common denominator of CLCuD across the affected areas of Pakistanand western India appears to be CLCuD DNA β. Although our understanding ofthe function of DNA β in the disease process and the diversity of this moleculeis still far from complete, only a single class of this satellite has been identifiedin cotton [3]. It is therefore likely that CLCuD DNA β is the cornerstone that

Cotton-infecting begomoviruses 1983
is driving the epidemic. This satellite molecule is well adapted to the “exotic”cotton varieties (particularly the variety S-12 in which the epidemic is believedto have started [7]) under cultivation on the Indian subcontinent and is able to berecruited by diverse begomoviruses, resulting in the present situation of multiplevirus species supporting, apparently, a single satellite. The recruitment by otherbegomoviruses is likely to occur through co-infection of alternate hosts, mostprobably within the Malvaceae, with other begomovirus/DNA β complexes andsubsequent transmission back to cotton by the vector whitefly Bemisia tabaci.Several malvaceous plant species harbour begomovirus/DNA β complexes, in-cluding hollyhock, okra and hibiscus [3], that could have acted as a source ofviruses for the diversity that we now encounter in cotton.
Our inability to insect transmit all but clone pCLCUV002 using B. tabaciremains something of a conundrum. Rather than being due to a defect in theclones, we attribute this to the problem of maintaining this insect species atthe high temperatures (35 ◦C and above) required for the reliable induction ofCLCuD symptoms in cotton. CLCuD symptomatic cotton plants rapidly losesymptoms when maintained at 25 ◦C, whereas B. tabaci suffers an unacceptablemortality rate when maintained at 35 ◦C. In the field, B. tabaci congregate onthe undersides of leaves near soil level, providing relief from the high daytimetemperatures (frequently 40 ◦C and above) during the cotton growing season inPakistan and India. Our attempts to conduct the insect transmissions at 25 ◦Cand then transfer the plants to cabinets maintained at 35 ◦C have thus far provenunreliable.
Extensive efforts have been made to develop resistance to CLCuD in elite cot-ton varieties by both traditional breeding/selection and utilising pathogen-derivedstrategies. However, with any resistance strategy, a highly diverse pathogen is anegative indicator for the durability of resistance, suggesting that conventionalresistance against CLCuD may not be long-lasting. In fact, recent reports fromPakistan suggest that, at least in some areas, resistance against CLCuD has alreadybroken down [21]. Our findings suggest that a pathogen-derived strategy based onCLCuD DNA β (a molecule with little apparent variation) might provide a moredurable resistance to this disease.
We are presently examining the diversity of both the begomoviruses and theDNA β satellite associated with CLCuD to establish the relationship between, aswell as the evolution of, the components of this begomovirus/DNA β complex.
Acknowledgements
This work was supported by grants from the Common Fund for Commodities (overseen by theInternational Cotton Advisory Committee under project no. ICAC/07), the Federal Ministryof FoodAgriculture and Livestock (Pakistan), theAsian Development Bank and the Institut deRecherche pour le Developpement. The CFC/ICAC project was a collaboration between thethe University ofArizona (Tucson, USA), the National Institute of Biotechnology and GeneticEngineering (Faisalabad, Pakistan), the Cotton Research Institute (Faisalabad, Pakistan) andthe JIC. Their inputs to this work are gratefully acknowledged. The authors also acknowledgethe support of the Biotechnology and Biological Sciences Research Council.

1984 S. Mansoor et al.
Viruses were held and manipulated (in the UK) with the authority of the Ministry ofAgriculture Fisheries and Food under the Plant Health (Great Britain) Order 1993 (SI no.1993/1320); license numbers PHF1419C/1907(6/96) and PHL 11/2557(2/1998).
References1. Arguello-Astorga G, Herrera-Estrella L, Rivera-Bustamante R (1994) Experimental and
theoretical definition of geminivirus origin of replication. Plant Mol Biol 26: 553–5562. Berrie LC, Rybicki EP, Rey MEC (2001) Complete nucleotide sequence and host range
of South African cassava mosaic virus: further evidence for recombination amongstbegomoviruses. J Gen Virol 82: 53–58
3. Briddon RW, Bull SE, Amin I, Idris AM, Mansoor S, Bedford ID, Dhawan P, Rishi N,Siwatch SS, Abdel-Salam AM, Brown JK, Zafar Y, Markham PG (2003) Diversity ofDNA β: a satellite molecule associated with some monopartite begomoviruses. Virology,in press
4. Briddon RW, Bull SE, Mansoor S, Amin I, Markham PG (2002) Universal primers for thePCR-mediated amplification of DNA β; a molecule associated with some monopartitebegomoviruses. Mol Biotechnology 20: 315–318
5. Briddon R, Mansoor S, Bedford I, Pinner M, Saunders K, Stanley J, Zafar Y, Malik K,Markham P (2001) Identification of DNA components required for induction of cottonleaf curl disease. Virology 285: 234–243
6. Briddon RW, Mansoor S, Bedford ID, Pinner MS, Markham PG (2000) Clones of cottonleaf curl geminivirus induce symptoms atypical of cotton leaf curl disease. Virus Genes20: 17–24
7. Briddon RW, Markham PG (2000) Cotton leaf curl virus disease. Virus Res 71: 151–1598. Briddon RW, Stenger DC, Bedford ID, Stanley J, Izadpanah K, Markham PG (1998)
Comparison of a beet curly top virus isolate from the old world with those from the newworld. Eur J Plant Path 104: 77–84
9. Chatterji A, Chatterji U, Beachy RN, Fauquet C (2000) Sequence parameters thatdetermine specificity of binding of the replication-associated protein to its cognate sitein two strains of Tomato leaf curl virus-New Delhi. Virology 273: 341–350
10. Chatterji A, Padidam M, Beachy RN, Fauquet CM (1999) Identification of replicationspecificity determinants in two strains of tomato leaf curl virus from New Delhi. J Virol73: 5481–5489
11. Collmer CW, Howell SH (1992) Role of satellite RNA in the expression of symptomscaused by plant viruses. Ann Rev Phytopathol 30: 419–442
12. Czosnek H, Laterrot H (1997) A worldwide survey of tomato yellow leaf curl viruses.Arch Virol 142: 1391–1406
13. Devereaux J, Haeberli P, Smithies O (1984) A comprehensive set of sequence analysisprograms for the VAX. Nucleic Acids Res 12: 387–395
14. Fauquet CM, Bisaro DM, Briddon RW, Brown JK, Harrison BD, Rybicki EP, StengerDC, Stanley J (2003) Revision of taxonomic criteria for species demarcation in the familyGeminiviridae, and an updated list of begomovirus species. Arch Virol 148: 405–421
15. FondongVN, Pita JS, Rey MEC, de KochkoA, Beachy RN, Fauquet CM (2000) Evidencefor synergism between African cassava mosaic virus and a new double-recombinantgeminivirus infecting cassava in Cameroon. J Gen Virol 81: 287–297
16. Fontes EPB, Gladfelter HJ, Schaffer RL, Petty ITD, Hanley-Bowdoin L (1994)Geminivirus replication origins have a modular organization. Plant Cell 6: 405–416
17. Hameed S, Khalid S, Ehsan-ul-Haq, Hashrni AA (1993) Cotton leaf curl disease inPakistan caused by a whitefly-transmitted geminivirus. Plant Dis 78: 529

Cotton-infecting begomoviruses 1985
18. Harrison BD, Liu YL, Khalid S, Hameed S, Otim-Nape GW, Robinson DJ (1997)Detection and relationships of cotton leaf curl virus and allied whitefly-transmittedgeminiviruses occurring in Pakistan. Ann Appl Biol 130: 61–75
19. Hong YG, Robinson DJ, Harrison BD (1993) Nucleotide sequence evidence for theoccurrence of three distinct whitefly-transmitted geminiviruses in cassava. J Gen Virol74: 2437–2443
20. Mansoor S, Amin I, Hussain M, Zafar Y, Bull S, Briddon RW, Markham PG (2001)Association of a disease complex involving a begomovirus, DNA 1 and a distinct DNAβ with leaf curl disease of okra in Paksitan. Plant Dis 85: 922
21. Mansoor S, Amin I, Iram S, Hussain M, Zafar Y, Malik KA, Briddon RW (2003) Thebreakdown of resistance in cotton to cotton leaf curl disease in Pakistan. New DiseaseReports vol 7 [http://www.bspp.org.uk/ndr/]
22. Mansoor S, Bashir A, Khan SH, Hussain M, Saeed M, Zafar Y, Markham PG, MalikKA (1999) Rapid multiplex PCR for the specific detection of two whitefly-transmittedgeminivirus species associated with cotton leaf curl disease in Pakistan. Pak J Bot 31:115–123
23. Mansoor S, Bedford I, Pinner MS, Stanley J, Markham PG (1993) A whitefly-transmittedgeminivirus associated with cotton leaf curl disease in Pakistan. Pak J Bot 25: 105–107
24. Maruthi MN, Colvin J, Seal S, Thresh JM (2001) First report of a distinct begomovirusinfecting cassava from Zanzibar. Plant Dis 86: 187
25. Nadeem A, Weng Z, Nelson MR, Xiong Z (1997) Cotton leaf crumple virus andcotton leaf curl virus are distantly related geminiviruses. Mol Plant Pathol on-line:http://www.bspp.org.uk/mppol.1997/0612nadeem
26. Noueiry AO, Lucas WJ, Gilbertson RL (1994) Two proteins of a plant DNA viruscoordinate nuclear and plasmodesmal transport. Cell 76: 925–932
27. Padidam M, Beachy RN, Fauquet CM (1995) Classification and identification ofgeminiviruses using sequence comparisons. J Gen Virol 76: 249–263
28. Pita JS, Fondong VN, Sangre A, Otim-Nape GW, Ogwal S, Fauquet CM (2001)Recombination, pseudorecombination and synergism of geminiviruses are determinantkeys to the epidemic of severe cassava mosaic disease in Uganda. J Gen Virol 82:655–665
29. Rahman H-ur-, Khan WS, Khan M-ud-D, Shah MKN (2001) Stability of cotton cultivarsunder leaf curl virus epidemic in Pakistan. Field Crops Res 69: 251–257
30. Rojas MR, Gilbertson RL, Russell DR, Maxwell DP (1993) Use of degenerate primersin the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis77: 340–347
31. Rojas MR, Jiang H, Salati R, Xoconostle-Cazares B, Sudarshana MR, Lucas WJ,Gilbertson RL (2001) Functional analysis of proteins involved in movement of themonopartite begomovirus, Tomato yellow leaf curl virus. Virology 291: 110–125
32. Rybicki EP (1994) A phylogenetic and evolutionary justification for three genera ofGeminiviridae. Arch Virol 139: 49–77
33. Rybicki EP, Briddon RW, Brown JE, Fauquet CM, Maxwell DP, Harrison BD, MarkhamPG, Bisaro DM, Robinson D, Stanley J (2000) Geminiviridae. In: van RegenmortelMHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J,Mayo MA, McGeoch DJ, Pringle CR, Wicker RB (eds) Virus Taxonomy. Seventh Reportof the International Committee on Taxonomy of Viruses. Academic Press, San Diego,pp 285–297
34. Saunders K, Bedford ID, Briddon RW, Markham PG, Wong SM, Stanley J (2000) Anovel virus complex causes Ageratum yellow vein disease. Proc Natl Acad Sci (USA)97: 6890–6895

1986 S. Mansoor et al.: Cotton-infecting begomoviruses
35. Saunders K, Salim N, MaliVR, MalathiVG, Briddon RW, Markham PG, Stanley J (2002)Characterisation of Sri Lankan cassava mosaic virus and Indian cassava mosaic virus:evidence for acquisition of a DNA B component by a monopartite begomovirus. Virology293: 63–74
36. Saxena S, Hallan V, Singh BP, Sane PV (1998) Nucleotide sequence and intergeminiviralhomologies of the DNA-A of papaya leaf curl geminivirus from India. Biochem MolBiol Int 45: 101–113
37. Singh J, Sohi AS, Brar DS, Denholm I, Russell D, Briddon R (1998) Management ofcotton leaf curl virus disease in India. In: Proc World Cotton Research Conference 2,Athens, Greece, pp 941–944
38. Stanley J (1983) Infectivity of the cloned geminivirus genome requires sequences fromboth DNAs. Nature 305: 643–645
39. Stanley J, Gay MG (1983) Nucleotide sequence of cassava latent virus DNA. Nature 301:260–262
40. Zhou X, Liu Y, Calvert L, Munoz C, Otim-Nape GW, Robinson DJ, Harrison BD (1997)Evidence that DNA A of a geminivirus associated with severe cassava mosaic disease inUganda has arisen by interspecific recombination. J Gen Virol 78: 2101–2111
41. Zhou X, LiuY, Robinson DJ, Harrison BD (1998) Four variants among Pakistani isolatesof cotton leaf curl virus and their affinities to DNA-A of geminivirus isolates from okra.J Gen Virol 79: 915–923
Author’s address: Rob W. Briddon, Department of Disease and Stress Biology, JohnInnes Centre, Norwich Research Park, Colney Lane, Norwich NR4 7UH, U.K.; e-mail:[email protected]
Related Documents

![[з:] girl, curl [з:] girl, curl small, Paulа small, Paulа [⊃:][⊃:]](https://static.cupdf.com/doc/110x72/56649f3f5503460f94c5fbe5/-girl-curl-girl-curl-small-paul-small-paul-.jpg)









