ARTICLES NATURE IMMUNOLOGY VOLUME 4 NUMBER 8 AUGUST 2003 765 ICOSL, also known as B7RP-1, B7h and B7-H2, is a member of the immunoglobulin (Ig) superfamily and belongs to the B7 family of co- stimulatory molecules 1,2 , which includes B7H1 (PD-L1) 3,4 , B7H3 (ref. 5) and PD-L2 (B7-DC) 6 . These proteins differ substantially in their expression patterns and regulation, and may control specific aspects of T cell activation, differentiation and function 7 . ICOSL specifically binds to ICOS but not to CD28 or to cytotoxic T lymphocyte antigen 4 (refs. 1,2). ICOSL is constitutively expressed in B cell areas of sec- ondary lymphoid organs 8 and is inducible in nonlymphoid tissues by treatment with bacterial lipopolysaccharide 1 . Mice transgenic for sol- uble ICOSL-Fc develop lymphoid hyperplasia, plasmocytosis and hypergammaglobulinemia 2 . Tumor cells expressing ICOSL are highly susceptible to destruction by CD8 + cytotoxic T cells and are more readily rejected in vivo 9,10 . ICOS is homologous to CD28 but is expressed on T cells only after activation 8,11 . In vitro, ICOS delivers a CD28-independent signal that stimulates production of interleukin 4 (IL-4) and IL-10 but not IL-2 (ref. 11). In vivo, Icos deletion 12–14 results in altered T H 2 differentiation, lack of germinal center formation and impaired isotype switching to IgG1 and IgE. In vivo blockade of ICOS function by monoclonal antibodies substantially prolongs allograft survival 15 and differentially modulates the severity of experimental autoimmune encephalomyelitis 16 and aller- gic airway disease 17 as a function of the phase of the T cell response. To investigate the physiological function of ICOSL, we generated gene-targeted Icosl –/– mice. The phenotype of these mutants was similar to that of Icos –/– mice 12–14 . We used Icosl –/– mice in an adop- tive transfer system to analyze where and when ICOS/ICOSL co- stimulation is required during T cell-dependent (TCD) B cell responses. Our findings indicated that engagement of ICOS-ICOSL was essential for the functional integrity of both T and B cell responses to soluble TCD antigens, and that costimulation through ICOS-ICOSL was pivotal in the primary, but not secondary, phase of helper T cell responses. RESULTS Gene targeting of mouse Icosl We inactivated Icosl by deleting a 1.1-kilobase (kb) genomic region containing base pairs (bp) 153–507 encoding the IgV loop (Fig. 1a). Because this region is essential for binding of B7-2 to the MYPPPY motif of CD28 (ref. 18), we reasoned that this domain might be essen- tial for the interaction of ICOSL with the FDPPPF motif of ICOS 2,11 . Icosl –/– mice were born at the expected mendelian frequency, and proved healthy and fertile. We confirmed the genotype of Icosl –/– mice by Southern blotting (Fig. 1b) and loss of ICOSL protein expression by flow cytometric analysis using the ICOSL-specific monoclonal anti- body 1B7 (Fig. 1c). 1 Advanced Medical Discovery Institute, 620 University Avenue, Toronto, Ontario M5G 2C1, Canada. 2 Ontario Cancer Institute, and Departments of Medical Biophysics and Immunology, University of Toronto, Toronto, Ontario M5G 2C1, Canada. 3 Amgen, One Amgen Center Drive, Thousand Oaks, California 91320-1789, USA. 4 Division of Respiratory Disease and Allergy, Centre for Gene Therapeutics, McMaster University, Hamilton, Ontario L8N 3Z5, Canada. 5 University Hospital Zurich, Department of Pathology, CH-8091, Zurich, Switzerland. 6 Mount Sinai Hospital, Samuel Lunenfeld Research Institute, 600 University Avenue, Toronto, Ontario M5G 1X5, Canada. 7 Present address: Hôpital Necker, INSERM U-580, 161, rue de Sèvres, 75743 Paris Cedex 15, France. Correspondence should be addressed to T.W.M. ([email protected]) or A.T. ([email protected]). Costimulation through the inducible costimulator ligand is essential for both T helper and B cell functions in T cell–dependent B cell responses Tak W Mak 1,2 , Arda Shahinian 1,2 , Steve K Yoshinaga 3 , Andrew Wakeham 1,2 , Louis-Martin Boucher 1,2 , Melania Pintilie 2 , Gordon Duncan 1,2 , Beata U Gajewska 4 , Matthew Gronski 2 , Urs Eriksson 2 , Bernhard Odermatt 5 , Alexandra Ho 1,2 , Denis Bouchard 1,2 , John S Whorisky 3 , Manel Jordana 4 , Pamela S Ohashi 2 , Tony Pawson 6 , Friedhelm Bladt 6 & Anna Tafuri 1,2,7 Costimulation through the inducible costimulator (ICOS) and its ligand (ICOSL) is essential for T cell–dependent B cell responses, but the cellular and temporal dynamics underlying its in vivo effects are poorly defined. Here we have shown that Icosl –/– and Icos –/– mice had similar phenotypes and that ICOS-ICOSL costimulation modulated the early but not late phases of IgG1 affinity maturation. Exploiting the adoptive transfer of T or B cells from primed Icosl –/– mice, we provided genetic evidence that costimulation through ICOSL was essential for primary but not secondary helper T cell responses and for the control of both T and B cell activities, resulting in T cell–dependent IgG1 production. © 2003 Nature Publishing Group http://www.nature.com/natureimmunology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A RT I C L E S
NATURE IMMUNOLOGY VOLUME 4 NUMBER 8 AUGUST 2003 765
ICOSL, also known as B7RP-1, B7h and B7-H2, is a member of theimmunoglobulin (Ig) superfamily and belongs to the B7 family of co-stimulatory molecules1,2, which includes B7H1 (PD-L1)3,4, B7H3 (ref.5) and PD-L2 (B7-DC)6. These proteins differ substantially in theirexpression patterns and regulation, and may control specific aspects ofT cell activation, differentiation and function7. ICOSL specificallybinds to ICOS but not to CD28 or to cytotoxic T lymphocyte antigen 4(refs. 1,2). ICOSL is constitutively expressed in B cell areas of sec-ondary lymphoid organs8 and is inducible in nonlymphoid tissues bytreatment with bacterial lipopolysaccharide1. Mice transgenic for sol-uble ICOSL-Fc develop lymphoid hyperplasia, plasmocytosis andhypergammaglobulinemia2. Tumor cells expressing ICOSL are highlysusceptible to destruction by CD8+ cytotoxic T cells and are morereadily rejected in vivo9,10.
ICOS is homologous to CD28 but is expressed on T cells only afteractivation8,11. In vitro, ICOS delivers a CD28-independent signal thatstimulates production of interleukin 4 (IL-4) and IL-10 but not IL-2 (ref.11). In vivo, Icos deletion12–14 results in altered TH2 differentiation, lackof germinal center formation and impaired isotype switching to IgG1and IgE. In vivo blockade of ICOS function by monoclonal antibodiessubstantially prolongs allograft survival15 and differentially modulatesthe severity of experimental autoimmune encephalomyelitis16 and aller-gic airway disease17 as a function of the phase of the T cell response.
To investigate the physiological function of ICOSL, we generatedgene-targeted Icosl–/– mice. The phenotype of these mutants wassimilar to that of Icos–/– mice12–14. We used Icosl–/– mice in an adop-tive transfer system to analyze where and when ICOS/ICOSL co-stimulation is required during T cell-dependent (TCD) B cellresponses. Our findings indicated that engagement of ICOS-ICOSLwas essential for the functional integrity of both T and B cellresponses to soluble TCD antigens, and that costimulation throughICOS-ICOSL was pivotal in the primary, but not secondary, phaseof helper T cell responses.
RESULTSGene targeting of mouse IcoslWe inactivated Icosl by deleting a 1.1-kilobase (kb) genomic regioncontaining base pairs (bp) 153–507 encoding the IgV loop (Fig. 1a).Because this region is essential for binding of B7-2 to the MYPPPYmotif of CD28 (ref. 18), we reasoned that this domain might be essen-tial for the interaction of ICOSL with the FDPPPF motif of ICOS2,11.Icosl–/– mice were born at the expected mendelian frequency, andproved healthy and fertile. We confirmed the genotype of Icosl–/– miceby Southern blotting (Fig. 1b) and loss of ICOSL protein expression byflow cytometric analysis using the ICOSL-specific monoclonal anti-body 1B7 (Fig. 1c).
1Advanced Medical Discovery Institute, 620 University Avenue, Toronto, Ontario M5G 2C1, Canada. 2Ontario Cancer Institute, and Departments of MedicalBiophysics and Immunology, University of Toronto, Toronto, Ontario M5G 2C1, Canada. 3Amgen, One Amgen Center Drive, Thousand Oaks, California 91320-1789,USA. 4Division of Respiratory Disease and Allergy, Centre for Gene Therapeutics, McMaster University, Hamilton, Ontario L8N 3Z5, Canada. 5University HospitalZurich, Department of Pathology, CH-8091, Zurich, Switzerland. 6Mount Sinai Hospital, Samuel Lunenfeld Research Institute, 600 University Avenue, Toronto,Ontario M5G 1X5, Canada. 7Present address: Hôpital Necker, INSERM U-580, 161, rue de Sèvres, 75743 Paris Cedex 15, France. Correspondence should beaddressed to T.W.M. ([email protected]) or A.T. ([email protected]).
Costimulation through the inducible costimulatorligand is essential for both T helper and B cellfunctions in T cell–dependent B cell responsesTak W Mak1,2, Arda Shahinian1,2, Steve K Yoshinaga3, Andrew Wakeham1,2, Louis-Martin Boucher1,2, Melania Pintilie2, Gordon Duncan1,2, Beata U Gajewska4, Matthew Gronski2, Urs Eriksson2, Bernhard Odermatt5,Alexandra Ho1,2, Denis Bouchard1,2, John S Whorisky3, Manel Jordana4, Pamela S Ohashi2, Tony Pawson6,Friedhelm Bladt6 & Anna Tafuri1,2,7
Costimulation through the inducible costimulator (ICOS) and its ligand (ICOSL) is essential for T cell–dependent B cellresponses, but the cellular and temporal dynamics underlying its in vivo effects are poorly defined. Here we have shown thatIcosl–/– and Icos–/– mice had similar phenotypes and that ICOS-ICOSL costimulation modulated the early but not late phases ofIgG1 affinity maturation. Exploiting the adoptive transfer of T or B cells from primed Icosl–/– mice, we provided genetic evidencethat costimulation through ICOSL was essential for primary but not secondary helper T cell responses and for the control of bothT and B cell activities, resulting in T cell–dependent IgG1 production.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
Reduced IgG1 production in Icosl–/– miceExtensive flow cytometric analyses of cells from 6- to 8-week-oldIcosl–/– and wild-type mice showed that in vivo deletion of Icosl did notaffect T or B cell development or the overall composition of mature Band T cell subsets in the spleen (data not shown).
We assessed B cell function in Icosl–/– mice in vitro and in vivo. Invitro, splenic B cells from Icosl–/– and wild-type mice responded equallyto mitogenic stimulation with lipopolysaccharide and antibody to IgMor CD40 (data not shown). In vivo, measurements of basal immuno-globulin concentrations showed that IgG1 was the only isotype signifi-cantly reduced in Icosl–/– mice (Fig. 2; P = 0.0120). The apparentincrease in serum IgM and decrease in serum IgG2a in the mutantswere not statistically significant. After in vivo challenge with the Tcell–independent antigen trinitrophenyl-Ficoll, Icosl–/– and wild-typemice had similar concentrations of IgM and IgG3 (data not shown).These findings were strongly indicative of a defect in TCD responses inIcosl–/– mice.
Requirement of ICOSL for germinal center formationTo investigate the function of ICOSL in TCD responses, we immu-nized wild-type and mutant mice with keyhole limpet hemocyanin(KLH) in complete Freund’s adjuvant (CFA). This antigenic challengeelicits both TH1 and TH2 responses and promotes B cell production ofIgG2a and IgG1. Serum concentrations of KLH-specific IgG1 were
significantly reduced (P < 0.0001) in Icosl–/– mice, whereas IgG2atiters were not substantially different from those of wild-type mice(Fig. 3a). We assessed germinal center formation, the anatomical hall-mark of ongoing T-B cell cooperation19, in frozen spleen sectionsfrom Icosl–/– and wild-type mice by staining with peanut agglutinin(Fig. 3b) and antibodies to B220 or CD4 (data not shown). The sizesand mean numbers of germinal centers per section (wild-type, 41.2 ±3.9; Icosl–/–, 5.3 ± 2) were notably reduced in Icosl–/– mice (Fig. 3b).Thus, abrogation of Icosl function interfered with B cell isotypeswitching and profoundly impaired cognate interaction between Tand B cells.
Impaired TH2 differentiation in Icosl–/– miceIsotype switching is driven by cytokines released by helper T cells20:IL-4 is required for isotype switching to IgG1 and IgE21, whereas inter-feron-γ (IFN-γ) promotes production of IgG2a22. To investigate theinvolvement of ICOSL in T cell cytokine production, we restimulatedhelper T cells from KLH-primed Icosl–/– and wild-type mice in vitrofor 4 d in the presence of KLH. We then assayed cytokine productionboth in culture supernatants and by intracellular staining. IL-4 and IL-10 concentrations (Fig. 4a) were significantly reduced (P = 0.0095) inIcosl–/– culture supernatants. In contrast, IFN-γwas produced in simi-lar amounts in mutant and wild-type cultures. IL-13 concentrationswere reduced in Icosl–/– culture supernatants compared with that of
766 VOLUME 4 NUMBER 8 AUGUST 2003 NATURE IMMUNOLOGY
Figure 1 In vivo ablation of mouse Icosl. (a) Protein and genomic structure of mouse ICOSL.ICOSL is composed of a signal peptide (SP; aminoacids (aa) 26–46), an IgV loop (aa 62–138), anIgC loop (aa 184–242), a transmembrane region(TM; aa 280–298) and an intracellular domain (IC;aa 299–322). The diagnostic restriction fragments(B, BamHI) and the location of the probe used forSouthern blot analysis are shown, as are thestructure of the Icosl targeting vector and thepredicted structure of the targeted locus. neo,neomycin-resistance gene. (b) Southern blotanalysis of representative tail biopsies of mice withmutated Icosl. Genomic DNA digested with BamHIwas hybridized to the short-arm flanking probe (5′probe) shown in a. Left margin, molecular sizes. (c) Icosl–/– mice lack ICOSL expression. Spleniccell subsets from Icosl–/– mice (thick lines) andwild-type littermates (thin lines) were stained withthe rat antibody to mouse ICOSL 1B7. Stainingwith the isotype control antibody consistentlyoverlapped Icosl–/– staining and has been omitted.Expression of ICOSL is shown on gated CD19+
cells (B), CD11b+CD11c– cells (M, macrophages),CD11c+CD11b– cells (DC), and CD4+ and CD8+
T cells. Results shown are representative of threeindependent experiments.
a b
c
Figure 2 Reduced basal concentrations of IgG1.Serum concentrations of IgM, IgG1, IgG2a,IgG2b, IgG3 and IgA in 8-week-old wild-type (�; n = 6) and Icosl–/– (�; n = 9) mice were analyzedby ELISA. Titers for wild-type control mice wereaveraged in each litter and considered to be100%, and the antibody titer for each mouse wascalculated as a percentage of the wild-type (WT)mean. Horizontal lines, mean antibody titers. Datarepresent the combined results of threeindependent experiments. *, P < 0.0001.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
wild-type supernatants, but this difference did not reach statistical sig-nificance (P > 0.1; data not shown). To rule out the possibility ofdecreased proliferation of Icosl–/– T cells, we measured incorporationof [3H]thymidine and did intracellular staining with bromodeoxyuri-dine and 7-amino-actinomycin D in cultures of enriched CD4+ T cellsfrom KLH-primed Icosl–/– and wild-type mice. Mutant and wild-typein vitro cultures had similar rates of [3H]thymidine incorporation atboth 72 and 96 h of culture (data not shown). In both mutant andwild-type cultures, comparable percentages of CD4+CD25+ T cellsincorporated bromodeoxyuridine and 7-amino-actinomycin D tosimilar extent at both 72 and 96 h (Fig. 4b). CD4+CD25+ T cells frommutant and wild-type cultures also showed similar staining withannexin V and 7-amino-actinomycin D (data not shown), indicatingthat ICOS-ICOSL costimulation was not essential for proliferation orapoptosis of CD4+ T cells. To investigate whether ICOSL costimula-tion enhanced IL-4 and IL-10 production or influenced the process ofTH2 differentiation itself, we used intracellular staining for IL-4, IL-10
and IFN-γ. Although production of IL-4, IL-10 and IFN-γon a per-cellbasis (fluorescence intensity) was similar in Icosl–/– and wild-typeCD4+CD25+ T cells, the percentage of T cells producing IL-4- and IL-10 was substantially decreased in the mutants. In contrast, we foundno difference in the percentage of mutant and wild-type T cells pro-ducing IFN-γ (Fig. 4c). Thus, our data indicated that ICOSL costimu-lation is pivotal in the control of TH2 T cell differentiation.
Impaired IgG1 affinity maturation in Icosl–/– miceThe observations described above prompted us to focus on TH2-mediated responses, particularly IgG1 affinity maturation23,24 andallergic reactions25. Immunization of wild-type mice with (4-hydroxy-3-nitrophenyl)–acetyl-ovalbumin (NP-OVA) adsorbed to alum isoften used to study affinity maturation23,24. This protocol induceshighly skewed TH2 responses associated with the production of IgG1and IgE antibodies. Icosl–/– mice immunized with NP-OVA-alumshowed a significant reduction in NP-specific IgG1 during the primary
NATURE IMMUNOLOGY VOLUME 4 NUMBER 8 AUGUST 2003 767
Figure 3 Impaired germinal center formation. (a) Reduced IgG1 in Icosl–/– mice after immunization with a TCD antigen. Wild-type (�; n = 10) and Icosl–/–
(�; n = 11) mice were immunized with KLH-CFA, and titers of KLH-specific antibodies were measured at day 14 after immunization. Data are presented aspercentage of wild-type (WT) mean. *, P < 0.05. (b) Impaired germinal center formation in response to KLH-CFA. Spleens were ‘snap-frozen’ on day 10 afterimmunization with KLH-CFA and germinal center formation was analyzed by staining with peanut agglutinin as described48. Original magnification, ×25.
a b
Figure 4 Selective defect in TH2 differentiation.(a) Reduced production of TH2 cytokines byIcosl–/– T cells. Enriched T cells from Icosl–/– andwild-type littermates immunized in vivo with KLH-CFA were restimulated in vitro with KLH. Icosl–/–
T cells (filled bars) produced significantly lowerconcentrations of IL-4 (P = 0.0095) and IL-10 (P = 0.0095) than did wild-type T cells (openbars). IFN-γ production was unaltered in theabsence of ICOSL. *, P < 0.01. (b) Loss of ICOSLfunction affects neither CD4+ T cell proliferationnor apoptosis in response to antigenic challenge.Intracellular incorporation of bromodeoxyuridine(BrdU) and 7-amino-actinomycin D (7-AAD) byCD4+ (at 72 h) and CD4+CD25+ T cells (at 72 and96 h) was similar in enriched CD4-APC culturesfrom KLH-CFA-primed Icosl–/– (right) and wild-type(left) littermates. Upper right corners, percentagesof cells in G1, S and G2/M phase (key, far upperright). (c) Defective TH2 polarization in Icosl–/–
mice. Flow cytometric analysis of intracellularproduction of IFN-γ, IL-4 and IL-10 by wild-typeand Icosl–/– T cells restimulated in vitro with KLH.Staining for IFN-γ and IL-4 is shown on gatedFSChiCD4+CD25+ T cells, whereas staining for IL-10 is shown on gated FSChiCD25+ T cells. Data are representative of three independentexperiments. Numbers by boxes, percentage ofcells enclosed in gate.
a b
c
γ
γ
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y
A RT I C L E S
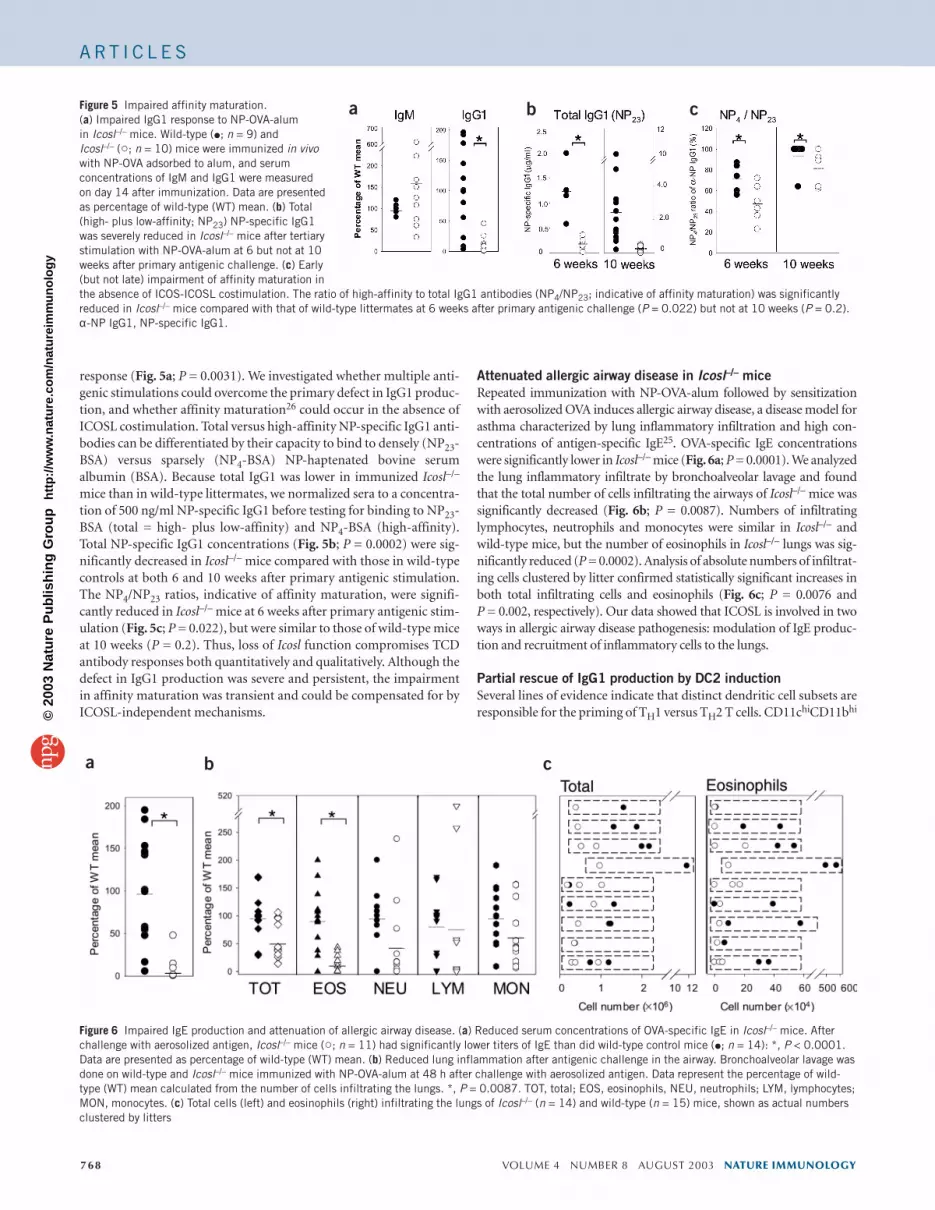
response (Fig. 5a; P = 0.0031). We investigated whether multiple anti-genic stimulations could overcome the primary defect in IgG1 produc-tion, and whether affinity maturation26 could occur in the absence ofICOSL costimulation. Total versus high-affinity NP-specific IgG1 anti-bodies can be differentiated by their capacity to bind to densely (NP23-BSA) versus sparsely (NP4-BSA) NP-haptenated bovine serumalbumin (BSA). Because total IgG1 was lower in immunized Icosl–/–
mice than in wild-type littermates, we normalized sera to a concentra-tion of 500 ng/ml NP-specific IgG1 before testing for binding to NP23-BSA (total = high- plus low-affinity) and NP4-BSA (high-affinity).Total NP-specific IgG1 concentrations (Fig. 5b; P = 0.0002) were sig-nificantly decreased in Icosl–/– mice compared with those in wild-typecontrols at both 6 and 10 weeks after primary antigenic stimulation.The NP4/NP23 ratios, indicative of affinity maturation, were signifi-cantly reduced in Icosl–/– mice at 6 weeks after primary antigenic stim-ulation (Fig. 5c; P = 0.022), but were similar to those of wild-type miceat 10 weeks (P = 0.2). Thus, loss of Icosl function compromises TCDantibody responses both quantitatively and qualitatively. Although thedefect in IgG1 production was severe and persistent, the impairmentin affinity maturation was transient and could be compensated for byICOSL-independent mechanisms.
Attenuated allergic airway disease in Icosl–/– miceRepeated immunization with NP-OVA-alum followed by sensitizationwith aerosolized OVA induces allergic airway disease, a disease model forasthma characterized by lung inflammatory infiltration and high con-centrations of antigen-specific IgE25. OVA-specific IgE concentrationswere significantly lower in Icosl–/– mice (Fig. 6a; P = 0.0001). We analyzedthe lung inflammatory infiltrate by bronchoalveolar lavage and foundthat the total number of cells infiltrating the airways of Icosl–/– mice wassignificantly decreased (Fig. 6b; P = 0.0087). Numbers of infiltratinglymphocytes, neutrophils and monocytes were similar in Icosl–/– andwild-type mice, but the number of eosinophils in Icosl–/– lungs was sig-nificantly reduced (P = 0.0002). Analysis of absolute numbers of infiltrat-ing cells clustered by litter confirmed statistically significant increases inboth total infiltrating cells and eosinophils (Fig. 6c; P = 0.0076 and P = 0.002, respectively). Our data showed that ICOSL is involved in twoways in allergic airway disease pathogenesis: modulation of IgE produc-tion and recruitment of inflammatory cells to the lungs.
Partial rescue of IgG1 production by DC2 inductionSeveral lines of evidence indicate that distinct dendritic cell subsets areresponsible for the priming of TH1 versus TH2 T cells. CD11chiCD11bhi
768 VOLUME 4 NUMBER 8 AUGUST 2003 NATURE IMMUNOLOGY
Figure 5 Impaired affinity maturation.(a) Impaired IgG1 response to NP-OVA-alum in Icosl–/– mice. Wild-type (�; n = 9) and Icosl–/– (�; n = 10) mice were immunized in vivowith NP-OVA adsorbed to alum, and serumconcentrations of IgM and IgG1 were measuredon day 14 after immunization. Data are presentedas percentage of wild-type (WT) mean. (b) Total(high- plus low-affinity; NP23) NP-specific IgG1was severely reduced in Icosl–/– mice after tertiarystimulation with NP-OVA-alum at 6 but not at 10weeks after primary antigenic challenge. (c) Early(but not late) impairment of affinity maturation inthe absence of ICOS-ICOSL costimulation. The ratio of high-affinity to total IgG1 antibodies (NP4/NP23; indicative of affinity maturation) was significantlyreduced in Icosl–/– mice compared with that of wild-type littermates at 6 weeks after primary antigenic challenge (P = 0.022) but not at 10 weeks (P = 0.2).α-NP IgG1, NP-specific IgG1.
a b c
Figure 6 Impaired IgE production and attenuation of allergic airway disease. (a) Reduced serum concentrations of OVA-specific IgE in Icosl–/– mice. Afterchallenge with aerosolized antigen, Icosl–/– mice (�; n = 11) had significantly lower titers of IgE than did wild-type control mice (�; n = 14): *, P < 0.0001.Data are presented as percentage of wild-type (WT) mean. (b) Reduced lung inflammation after antigenic challenge in the airway. Bronchoalveolar lavage wasdone on wild-type and Icosl–/– mice immunized with NP-OVA-alum at 48 h after challenge with aerosolized antigen. Data represent the percentage of wild-type (WT) mean calculated from the number of cells infiltrating the lungs. *, P = 0.0087. TOT, total; EOS, eosinophils, NEU, neutrophils; LYM, lymphocytes;MON, monocytes. (c) Total cells (left) and eosinophils (right) infiltrating the lungs of Icosl–/– (n = 14) and wild-type (n = 15) mice, shown as actual numbersclustered by litters
a b c
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
‘myeloid’ dendritic cells (DC2) preferentially induce T cell productionof IL-4 and IL-10, whereas CD11chiCD11b– ‘lymphoid’ dendritic cells(DC1) promote production of IFN-γ27,28. In vivo injection of wild-typemice with polyethylene glycol–modified granulocyte-macrophagecolony-stimulating factor (PEG-GM-CSF) has a TH2-type adjuvanteffect that results in DC2 expansion29,30. After mice were injected withPEG-GM-CSF, the expansion of ‘myeloid’ dendritic cells (R2), assessedby flow cytometry, was similar in wild-type and Icosl–/– littermates (Fig. 7a). We detected ICOSL expression on ‘lymphoid’ dendritic cells(R1), ‘myeloid’ dendritic cells (R2) and macrophages (R3) in wild-typemice (Fig. 7b). In Icosl–/– mice, immunization with NP-OVA after treat-ment with PEG-GM-CSF restored NP-specific IgG1 production towild-type amounts in only 50% (three of six) of the litters examined(Fig. 7c). The IgG1 concentrations of Icosl–/– mice treated with PEG-GM-CSF (calculated as a percentage of the wild-type mean) increasedfrom 8.7 ± 12.9% for the primary response to NP-OVA-alum (Fig. 5a)and 21.2 ± 21.7% for the tertiary response (Fig. 5b, NP23) to 40.9 ±47.2% (Fig. 7c). However, the incomplete rescue of IgG1 productionled us to look for ICOS-ICOSL-dependent events that could not be res-cued by enhanced DC2 activity.
ICOSL is required for both T and B cell functionsThe ICOS-ICOSL costimulatory axis underlying TCD B cell responsescomprises two functionally distinct interfaces: the interaction of T cellswith professional or semi-professional antigen-presenting cells (APC-T) to induce T cell priming, differentiation and acquisition of helperfunction, and the antigen-driven interaction of primed T cells with Bcells (T-B) that results in antibody production and isotype switching. Todetermine the function of ICOSL in each arm of this axis, CD4+ T cellswere primed in the presence or absence of ICOSL and adoptively trans-ferred into naive recipients together with Icosl–/– or wild-type B cellsfrom primed donors (Fig. 8a). To test the efficiency of T-B interaction in
the absence of ICOS-ICOSL costimulation, wild-type T cells and Icosl–/–
B cells were transferred together into wild-type recipients. Similarly, totest whether ICOS-ICOSL engagement was required for effective helperT cell differentiation, T cells from Icosl–/– mice and wild-type B cells weretransferred together into Icosl–/– recipients. First, we investigatedwhether our adoptive transfer system could measure NP-specificresponses of donor origin. We assessed the response to NP-OVA inincomplete Freund’s adjuvant (IFA) in the following control situations:naive wild-type recipients irradiated (4 Gy) and immunized with NP-OVA-IFA (no adoptive transfer); irradiated naive recipients adoptivelytransferred with CD4+ T cells and B cells from virgin wild-type mice(TVBV→WT); irradiated naive recipients adoptively transferred withCD4+ T cells and B cells from wild-type mice immunized with PBS-CFA(TWTBWT + PBS-CFA→WT). All control recipient mice produced lowor undetectable amounts of NP-specific IgG1. In contrast, recipients ofCD4+ T cells and B cells from wild-type mice that had been immunizedwith NP-OVA-CFA mounted a vigorous NP-specific IgG1 response(TWTBWT + NP-OVA-CFA→WT). However, the abrogation of ICOS-ICOSL engagement during either APC-T interaction (TMutBWT→Icosl–/–; APC-T) or secondary T-B interaction (TWTBMut→WT; T-B)resulted in substantial defects in IgG1 production (Fig. 8b).
We then assessed the temporal requirements for ICOS-ICOSL co-stimulation in primary and secondary TCD responses using the sameadoptive transfer system. When the ICOS-ICOSL pathway wasactively engaged during priming and/or differentiation of CD4+ Tcells, the absence of functional ICOS-ICOSL costimulation duringthe recall phase (TWTBWT→Icosl–/–) did not substantially reduceserum NP-specific IgG1 (Fig. 8c). In contrast, when costimulationthrough ICOSL was not provided during the primary response,engagement of this costimulatory pathway in the recall phase couldnot rescue the deficient IgG1 production (TMutBWT→WT). Thus,costimulation through ICOS-ICOSL is required for both T and B cell
NATURE IMMUNOLOGY VOLUME 4 NUMBER 8 AUGUST 2003 769
Figure 7 Partial rescue of defective IgG1production by PEG-modified GM-CSF. (a) DC2expansion. Repetitive in vivo injections of PEG-modified GM-CSF induced expansion ofCD11c+CD11b+ myeloid dendritic cells in wild-type and Icosl–/– mice. Percentages of ‘lymphoid’CD11c+CD11b– dendritic cells (R1), myeloidCD11c+CD11b+ dendritic cells (R2) andCD11c–CD11b+ macrophages (R3) were similarin wild-type and Icosl–/– mice (R1, 2.33% versus2.55%; R2, 6.65% versus 6.93%; R3, 6.27%versus 6.28%, respectively). Results shown arefrom one of three independent experiments.Percentages of cells in R1 and R3 did not changemuch from baseline in either wild-type or Icosl–/–
mice. CD11chiCD11bhi dendritic cellsconsistently represented less than 1% of totalspleen cells in both wild-type and Icosl–/– mice.(b) ICOSL expression. After 5 d of in vivotreatment with PEG-modified GM-CSF, ICOSLexpression was enhanced over basal levels on allsubsets in wild-type mice (thin lines), but wasabsent on cells derived from Icosl–/– mice (thicklines). Staining with the isotype control antibodyconsistently overlapped that of Icosl–/– and hasbeen omitted. R1, R2 and R3, defined in a. (c) NP-specific IgG1 production. Wild-type andIcosl–/– mice treated with PEG-modified GM-CSF were immunized with NP-OVA. Concentrations of NP-specific IgG1 were measured 7 days after secondaryimmunization. Data are shown as absolute values found within litters (left; �, wild-type, n = 12; �, Icosl–/–, n = 8) and as the percentage of the wild-type(WT) mean (right). Controls (n = 4) injected with PBS instead of PEG-modified GM-CSF and immunized with NP-OVA had low or undetectable concentrationsof NP-specific IgG1 (gray diamond).
a
b
c
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
functions and in the primary, but not secondary, phase of TCD anti-body production.
DISCUSSIONEffective collaboration between cognate lymphocytes, which is modu-lated by costimulation, is essential for the generation of immuneresponses to TCD antigens. In this study, we used a genetic approachto define the physiological function of the ICOS ligand and to dissectthe cellular and temporal requirements for ICOS-ICOSL costimula-tion in TCD B cell responses.
The phenotype of Icosl–/– mice was similar to that of Icos–/– mice,thus providing genetic evidence for the widely held hypothesis thatICOSL is the only functional counterpart of ICOS. The basal serumconcentrations of IgG1 were reduced in Icosl–/– mice, indicating adefect in T-B collaboration31. Consistent with this serumimmunoglobulin profile, in vivo challenge with KLH-CFA resulted inselective impairment of TH2 polarization but intact IFN-γ productionand TH1 polarization. This defect was not accounted for by alteredCD4+ T cell proliferation or apoptosis, indicating a crucial function
for ICOS-ICOSL costimulation in the processof TH2 differentiation.
The impaired germinal center formation inIcosl–/– mice indicated that loss of ICOSLinterferes with the generation of the siteswhere T-B cross-talk physically occurs. A sim-ilar defect is induced by in vivo blockade ofICOSL32. Reduced germinal center formationcan also result from impaired engagement ofCD28 by B7.1 or B7.2 (refs. 33,34), or ofCD40 by CD40L35,36. The interplay betweenthese costimulatory pathways is only partiallyunderstood; it is known they are intercon-nected through IL-4. Indeed, germinal centerformation is reduced in Il4–/– mice and isalmost abolished in Cd28–/–Il4–/– mice37.Furthermore, costimulation through B7.1and/or B7.2 enhances ICOS expression38 andstabilizes CD40L expression on activated Tcells39. In vitro, CD40 signaling promotesICOSL expression on B cells, counteracting Bcell receptor and IL-4R-mediated downregu-lation of ICOSL32. In vivo, CD40 costimula-tion rescues the defective IgG1 production ofIcos–/– mice13.
Our data have shown ICOS-ICOSL isessential in costimulation in modulating,both quantitatively and qualitatively, theearly phases of IgG1 production and affinitymaturation. We have also demonstrated thatICOS-ICOSL-independent mechanisms canrescue affinity maturation but not antibodyproduction at later stages of the immuneresponse.
We found that Icosl deletion leads todecreased serum IgE and inflammatory air-way infiltration. Eosinophilic infiltration ismodulated by local production of IL-4, IL-5and IL-13 (ref. 40). Although IL-5 and IL-13production was not altered in Icosl–/– T cellcultures, IL-4 was significantly reduced. Ourdata were consistent with previous findings
obtained for Icos–/– mice12 and in vivo ICOS blockade17,41,42.Discrepancies in eosinophilic bronchoalveolar infiltration notedbetween Icos–/– mice12, in vivo ICOS blockade experiments17,41,42 andIcosl–/– mice are probably due to differences in immunization proto-cols. The function of ICOS-ICOSL costimulation in enhancing allergicreactions must be taken into consideration if this costimulatory path-way is to be exploited in vaccine design.
Several lines of evidence indicate that the signals provided by differ-ent dendritic cell subsets dictate TH1-TH2 differentiation27,28. In ourstudy, ICOSL was not differentially expressed on wild-type DC1 versusDC2 cells, and in vivo treatment of Icosl–/– mice with PEG-GM-CSF29,30 led to only a partial rescue of IgG1 production. The equiva-lent expression of ICOSL on DC1 and DC2 subsets and the normalexpansion of Icosl–/– DC2 cells argue against the restricted involvementof ICOSL in DC2 function.
ICOS-expressing T cells interact with ICOSL during two differentphases of a response to a TCD antigen: first, during the T cell primingand differentiation phase, in which professional or semi-professionalAPCs promote T cell acquisition of helper function; and second, during
770 VOLUME 4 NUMBER 8 AUGUST 2003 NATURE IMMUNOLOGY
Figure 8 Cellular and temporal requirements for ICOS/B7RP-1 costimulation in TCD responses. (a) Adoptive transfer system. Donor wild-type or Icosl–/– mice were immunized with NP-OVA-CFA. Then, 10 d later, combinations of splenic CD4+T cells and B cells derived from different donors wereadoptively transferred into irradiated naive recipients, which were immunized with NP-OVA-IFA at thetime of the transfer. Serum concentrations of NP-specific IgG1 were measured 1 week after transfer andimmunization. (b) Costimulation through ICOSL is required for both T and B cell functions in response toTCD antigens. NP-specific IgG1 detected in the serum of recipient mice was exclusively produced by theinteraction of CD4+T and B cells of donor origin, as shown by the low/undetectable titers of IgG1 in micereceiving no cell transfer (�; n = 4), virgin CD4+T and B cells (�; n = 3), or CD4+T and B cells from wild-type mice immunized with PBSCFA (∆; n = 8). Abrogation of ICOS-ICOSL costimulation at the level ofeither T cell interaction with APCs (�; n = 10; TMutBWT→Icosl–/–) or of B cell interaction with primed Tcells (�; n = 10; TWTBMut→WT) resulted in much less IgG1 production than that of mice with an intactICOS-ICOSL pathway (�; n = 7). (c) Costimulation through ICOSL is required in the priming phase ofTCD responses. Priming of naive T cells in the presence of a functional ICOS-ICOSL pathway resulted in the production of normal levels of IgG1, whether costimulation through ICOSL was provided in the secondary phase of the response (�, controls, n = 7; TWTBWT→WT) or not (gray circles; n = 7;TWTBWT→Icosl–/–). In contrast, selective abrogation of costimulation through ICOSL during primingresulted in a reduction of IgG1 levels that could not be overcome by active engagement of ICOS-ICOSLduring the recall phase of the response (gray upwards triangles; n = 7; TMutBWT→WT). wild-typerecipients of TMutBmut had low or undetectable levels of IgG1 (gray downwards triangles; n = 3). V, virgin;WT, wild-type; Mut, mutant; 1° or 2° immun., primary or secondary immunization.
a
b c
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
the delivery of T cell help to B cells. Our adoptive transfer systemenabled us to differentially analyze these two phases and showed thatICOSL costimulation is required for the integrity of both T and B cellfunctions during TCD antibody responses. Thus, the defect in IgG1production in Icosl–/– mice is not merely due to altered TH2 differentia-tion and insufficient production of IL-4 and IL-10 but also resides inthe inability of Icosl–/– B cells to receive T cell help after antigenic boost-ing. The mechanisms ensuring ICOSL expression on activated B cells,such as CD40-CD40L signaling32, are therefore of critical importancefor optimal TCD antibody responses.
Our results confirm previous reports15 that ICOS-ICOSL costimu-lation is essential during antigen priming. However, unlike CD8+
cytotoxic T cells9,10 and CD4+ ‘inflammatory’ cells2,43, CD4+ helper Tcells do not require ICOS-ICOSL costimulation during the recallphase of TCD responses. Although ICOS expression is similar on allthese T cell subsets2,44, ICOS-ICOSL costimulation may be differen-tially required for their very distinct effector functions. Alternatively,these discrepancies may arise from differences in technical approaches,such as the use of blocking reagents in vivo2 or during in vitro prim-ing43. The high frequency of antigen-specific T cells in TCR transgenicanimals may magnify effects otherwise undetectable in bulk T cellpopulations9,10.
In conclusion, our studies shed light on the cellular and temporaldynamics underlying ICOS-ICOSL costimulation in TCD responses,and emphasized the functional complexity of this costimulatory path-way. These factors must be borne in mind if the ICOS-ICOSL pathwayis to become a target for therapeutic intervention in transplantationand autoimmune disorders45,46.
METHODSGeneration of Icosl–/– mice. Mouse Icosl was isolated from a 129/J library usinga full-length (1,190-bp) cDNA probe. The targeting vector replaced a 1.1-kbgenomic fragment with a neomycin-resistance cassette in sense orientation rel-ative to Icosl transcription. Most of the putative exon 2 and the entire exon 3,which encode the leader sequence and the first IgV domain containing theputative ICOS-binding site, were deleted. The targeting vector was electropo-rated into E14 embryonic stem cells (129/Ola) at day 14 of gestation. AfterG418 selection, homologous recombinants were identified by PCR and verifiedby Southern blotting. Chimeric mice generated from Icosl+/– embryonic stemcell clones were crossed with C57BL/6 females to produce Icosl+/– mice.Germline transmission of the Icosl mutation was assessed in tail biopsy DNA bySouthern blot analysis and was obtained from two independently targetedembryonic stem cell lines. The wild-type and mutant alleles yielded bands of5.8 kb and 4.5 kb, respectively. The Icosl–/– mice used in this study were F2-F3
backcrosses onto the C57BL/6 background. All animals were housed and main-tained in accordance with the protocols of the Ontario Cancer Institute AnimalCare Committee.
Flow cytometric analyses. To assess ICOSL expression, spleen cells were stainedwith an irrelevant isotype control antibody or fluorescein isothiocyanate–labeled 1B7, a rat monoclonal antibody to mouse ICOSL of the IgG2b isotype gen-erated as described before47. 1B7 specifically binds to mouse ICOSL and blocksbinding between mouse ICOS-Fc and ICOSL. B cell development in the bonemarrow was assessed by staining for IgM, IgD, B220, CD43, BP-1 and CD24. T celldevelopment in the thymus was assessed by staining for CD4, CD8, CD3, TCRαβ,CD25 and CD44. In the spleen, the numbers and relative percentages of mature Bcells were assessed by staining for B220, CD21 and CD23, whereas those of T cellswere assessed with CD25, CD69, CD44, CD45RB and CD62L. All antibodies andreagents for surface and intracellular staining were from PharMingen. Cell stain-ing was detected by flow cytometry using a FACSCalibur (Becton Dickinson) anddata were analyzed using CellQuest software.
Determination of serum isotype-specific immunoglobulin and affinity mat-uration. Concentrations of serum polyclonal IgM, IgG1, IgG2a, IgG2b, IgG3
and IgA antibodies were determined using isotype-specific enzyme-linkedimmunosorbent assay (ELISA; Southern Biotechnology). Serum immunoglo-bulin titers were determined by comparing serial dilutions of sera with isotypecontrol standard curves using the SOFTmaxPRO (Molecular Devices) ELISAanalysis program.
To assess affinity maturation, we used two consecutive ELISAs. In the firstELISA, we determined the amounts of total (high- plus low-affinity) NP-specific IgG1 present in the sera of Icosl–/– and wild-type mice (NP23-BSA). Inthe second ELISA, we normalized samples to a concentration of 500 ng/ml NP-specific IgG1 and subjected them to serial dilution. We assessed the bindingactivities of wild-type or Icosl–/– sera to NP23-BSA or to NP4-BSA by comparingthe binding curve of the serial serum dilutions to the control curve of theH33Lγ1 antibody of known affinity for NP (Ka= 2.0 × 107 M–1; provided by G.Kelsoe, Duke University). NP4/NP23 ratios were calculated from OD valuesobtained at concentrations of 125 or 63 ng/ml total NP-specific IgG1.
In vivo immunizations. For TCD antigens, mice were immunized intraperi-toneally with 50 µg KLH (Calbiochem) in CFA (Sigma) or 100 µg NP18-OVA(Bioresearch Technologies) adsorbed to alum. Blood was collected from mousetails on days 7 and 14 after immunization, and serum antigen- and isotype-spe-cific antibodies were quantified by ELISA as described before14. For affinity mat-uration, mice were immunized three times with 100 µg NP18-OVA and alum, andblood was obtained at 6 and 10 weeks after primary antigenic stimulation. For Tcell–independent antigens, mice were injected intraperitoneally with 25 µg trini-trophenyl-Ficoll (Bioresearch Technologies) and blood was collected on day 7after immunization. To test the effect of PEG-GM-CSF on NP-specific IgG1 pro-duction, mice were treated for 5 consecutive days with 5 µg PEG-GM-CSF inPBS, injected intraperitoneally, as described before29,30. On day 5, in addition toPEG-GM-CSF, the mice received 400 µg NP18-OVA in PBS subcutaneously.
Adoptive transfer experiments. Donor mice were immunized intraperi-toneally with 50 µg NP18-OVA in CFA and were killed on days 9–11 afterimmunization. CD4+ T cells and B220+ B cells were isolated using theMiniMacs magnetic cell separation system, according the manufacturer’sinstructions. CD4+ T cells (2 × 106 to 3 × 106) and B220+ B cells (10 × 106 to 12× 106) were injected intravenously into irradiated (4 Gy) recipients.Immediately after cell transfer, the recipients were immunized intraperitoneallywith 50 µg NP18-OVA in IFA. Serum was collected by cardiac puncture on day 7after transfer and NP-specific IgG1 concentrations were assessed by ELISA.
Cytokine production. Mice were injected subcutaneously with 50 µg KLH-CFA and draining lymph nodes were isolated on day 10 after immunization.Lymph node cells depleted of B220+ and CD8+ cells by treatment with mag-netic beads coated with antibodies to B220 and CD8 (Dynal) were cultured in12-well plates in the presence of 100 µg/ml soluble KLH. Culture supernatantswere collected after 90 h, and concentrations of IL-4, IL-10, IL-13 and IFN-γwere determined by immunoassay (R&D Systems). To determine intracellularcytokine expression, stimulated cells were cultured for an additional 4 h in thepresence of phorbol-ester myristate acetate (10 ng/ml) plus ionomycin (100 ng/ml) plus Golgi STOP (PharMingen), and were subsequently stainedand analyzed by flow cytometry.
Allergic airway disease induction and analysis. About 2–3 weeks after tertiaryimmunization with 100 µg NP-OVA adsorbed to alum, mice were exposed to1% OVA aerosol for two 1-h periods spaced 4 h apart. At 2 d after aerosol sensi-tization, mice were killed to obtain serum for OVA-specific IgE detection andfor analysis of lung inflammatory cells by bronchoalveolar lavage25. Differentialcounts of bronchoalveolar lavage cells were determined from samples of at least500 leukocytes, using standard hemocytological criteria to classify cells as neu-trophils, eosinophils, lymphocytes or monocytes.
Statistical analyses. Results were analyzed as percentage of wild-type mean andas raw data stratified by mouse litters. The Wilcoxon-Mann-Whitney testimplemented on StatXact 4.0.1 was used without strata and with strata.Because of multiple comparisons in the basal immunoglobulin and bron-choalveolar lavage experiments, P values between 0.01 and 0.05 were consid-ered marginally significant. For the rest of the experiments, results wereconsidered statistically significant at P < 0.05.
NATURE IMMUNOLOGY VOLUME 4 NUMBER 8 AUGUST 2003 771
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y

A RT I C L E S
ACKNOWLEDGMENTSWe thank T. Horan, M. Zhang, K. Gaida and D. Yanagihara for preparation of the1B7 monoclonal antibody; G. Kelsoe, M.J. Schlomchik, K. Pfeffer, J. Dewey and A. Hebermann for help with the affinity maturation experiments; M. Bachmannfor discussions; M. Saunders for scientific editing; and I. Ng for administrativeassistance. This work was supported by the Canadian Institutes of Health Research,Amgen and the Canadian Network for Vaccines and Immunotherapeutics ofCancer and Chronic Viral Diseases.
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Received 17 October 2002; accepted 28 May 2003Published online 29 June 2003; doi:10.1038/ni947
1. Swallow, M.M., Wallin, J.J. & Sha, W.C. B7h, a novel costimulatory homolog of B7.1and B7.2, is induced by TNF-α. Immunity 11, 423–432 (1999).
2. Yoshinaga, S.K. et al. T-cell co-stimulation through B7RP-1 and ICOS. Nature 402,827–832 (1999).
3. Dong, H., Zhu, G., Tamada, K. & Chen, L. B7-H1, a third member of the B7 family,co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 5,1365–1369 (1999).
4. Freeman, G.J. et al. Engagement of the PD-1 immunoinhibitory receptor by a novelB7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med.192, 1027–1034 (2000).
5. Chapoval, A.I. et al. B7-H3: a costimulatory molecule for T cell activation and IFN-γproduction. Nat. Immunol. 2, 269–274 (2001).
6. Latchman, Y. et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation.Nat. Immunol. 2, 261–268 (2001).
7. Carreno, B.M. & Collins, M. The B7 family of ligands and its receptors: new pathwaysfor costimulation and inhibition of immune responses. Annu. Rev. Immunol. 20,29–53 (2002).
8. Yoshinaga, S.K. et al. Characterization of a new human B7-related protein: B7RP-1 isthe ligand to the co-stimulatory protein ICOS. Int. Immunol. 12, 1439–1447 (2000).
9. Liu, X. et al. B7H costimulates clonal expansion of, and cognate destruction of tumorcells by, CD8+ T lymphocytes in vivo. J. Exp. Med. 194, 1339–1348 (2001).
10. Wallin, J.J., Liang, L., Bakardjiev, A. & Sha, W.C. Enhancement of CD8+ T cellresponses by ICOS/B7h costimulation. J. Immunol. 167, 132–139 (2001).
11. Hutloff, A. et al. ICOS is an inducible T-cell co-stimulator structurally and function-ally related to CD28. Nature 397, 263–266 (1999).
12. Dong, C. et al. ICOS co-stimulatory receptor is essential for T-cell activation and func-tion. Nature 409, 97–101 (2001).
13. McAdam, A.J. et al. ICOS is critical for CD40-mediated antibody class switching.Nature 409, 102–105 (2001).
14. Tafuri, A. et al. ICOS is essential for effective T-helper-cell responses. Nature 409,105–109 (2001).
15. Ozkaynak, E. et al. Importance of ICOS-B7RP-1 costimulation in acute and chronicallograft rejection. Nat. Immunol. 2, 591–596 (2001).
16. Rottman, J.B. et al. The costimulatory molecule ICOS plays an important role in theimmunopathogenesis of EAE. Nat. Immunol. 2, 605–611 (2001).
17. Gonzalo, J.A. et al. ICOS is critical for T helper cell-mediated lung mucosal inflam-matory responses. Nat. Immunol. 2, 597–604 (2001).
18. Rennert, P. et al. The IgV domain of human B7-2 (CD86) is sufficient to co-stimulateT lymphocytes and induce cytokine secretion. Int. Immunol. 9, 805–813 (1997).
19. MacLennan, I.C. Germinal centers. Annu. Rev. Immunol. 12, 117–139 (1994).20. Finkelman, F.D. et al. Lymphokine control of in vivo immunoglobulin isotype selec-
tion. Annu. Rev. Immunol. 8, 303–333 (1990).21. Kuhn, R., Rajewsky, K. & Muller, W. Generation and analysis of interleukin-4 defi-
cient mice. Science 254, 707–710 (1991).
22. Snapper, C.M. & Paul, W.E. Interferon-γ and B cell stimulatory factor-1 reciprocallyregulate Ig isotype production. Science 236, 944–947 (1987).
23. Matsumoto, M. et al. Affinity maturation without germinal centres in lymphotoxin-α-deficient mice. Nature 382, 462–466 (1996).
24. Futterer, A., Mink, K., Luz, A., Kosco-Vilbois, M.H. & Pfeffer, K. The lymphotoxin βreceptor controls organogenesis and affinity maturation in peripheral lymphoid tis-sues. Immunity 9, 59–70 (1998).
25. Ohkawara, Y. et al. Cytokine and eosinophil responses in the lung, peripheral blood,and bone marrow compartments in a murine model of allergen-induced airwaysinflammation. Am. J. Respir. Cell. Mol. Biol. 16, 510–520 (1997).
26. Tarlinton, D.M., Light, A., Nossal, G.J. & Smith, K.G. Affinity maturation of the primaryresponse by V gene diversification. Curr. Top. Microbiol. Immunol. 229, 71–83 (1998).
27. Moser, M. & Murphy, K.M. Dendritic cell regulation of TH1-TH2 development. Nat.Immunol. 1, 199–205 (2000).
28. Liu, Y.J., Kanzler, H., Soumelis, V. & Gilliet, M. Dendritic cell lineage, plasticity andcross-regulation. Nat. Immunol. 2, 585–589 (2001).
29. Pulendran, B. et al. Distinct dendritic cell subsets differentially regulate the class ofimmune response in vivo. Proc. Natl. Acad. Sci. USA 96, 1036–1041 (1999).
30. Daro, E. et al. Polyethylene glycol-modified GM-CSF expands CD11bhighCD11chigh
but notCD11blowCD11chigh murine dendritic cells in vivo: a comparative analysis withFlt3 ligand. J. Immunol. 165, 49–58 (2000).
31. Grusby, M.J. & Glimcher, L.H. Immune responses in MHC class II-deficient mice.Annu. Rev. Immunol. 13, 417–435 (1995).
32. Liang, L., Porter, E.M. & Sha, W.C. Constitutive expression of the B7h ligand forinducible costimulator on naive B cells is extinguished after activation by distinct Bcell receptor and interleukin 4 receptor-mediated pathways and can be rescued byCD40 signaling. J. Exp. Med. 196, 97–108 (2002).
33. Ferguson, S.E., Han, S., Kelsoe, G. & Thompson, C.B. CD28 is required for germinalcenter formation. J. Immunol. 156, 4576–4581 (1996).
34. Borriello, F. et al. B7-1 and B7-2 have overlapping, critical roles in immunoglobulinclass switching and germinal center formation. Immunity 6, 303–313 (1997).
35. Kawabe, T. et al. The immune responses in CD40-deficient mice: impaired immunoglob-ulin class switching and germinal center formation. Immunity 1, 167–178 (1994).
36. Xu, J. et al. Mice deficient for the CD40 ligand. Immunity 1, 423–431 (1994).37. Reiter, R. & Pfeffer, K. Impaired germinal centre formation and humoral immune
response in the absence of CD28 and interleukin-4. Immunology 106, 222–228 (2002).38. McAdam, A.J. et al. Mouse inducible costimulatory molecule (ICOS) expression is
enhanced by CD28 costimulation and regulates differentiation of CD4+ T cells. J. Immunol. 165, 5035–5040 (2000).
39. Jaiswal, A.I. & Croft, M. CD40 ligand induction on T cell subsets by peptide-present-ing B cells: implications for development of the primary T and B cell response. J. Immunol. 159, 2282–2291 (1997).
40. Coyle, A.J., Lloyd, C.M. & Gutierrez-Ramos, J.C. Biotherapeutic targets for the treat-ment of allergic airway disease. Am. J. Respir. Crit. Care Med. 162, S179–184 (2000).
41. Coyle, A.J. et al. The CD28-related molecule ICOS is required for effective T cell-dependent immune responses. Immunity 13, 95–105 (2000).
42. Tesciuba, A.G. et al. Inducible costimulator regulates TH2-mediated inflammation,but not TH2 differentiation, in a model of allergic airway disease. J. Immunol. 167,1996–2003 (2001).
43. Sporici, R.A. et al. ICOS ligand costimulation is required for T-cell encephalitogenic-ity. Clin. Immunol. 100, 277–288 (2001).
44. Campbell, D.J., Kim, C.H. & Butcher, E.C. Separable effector T cell populations spe-cialized for B cell help or tissue inflammation. Nat. Immunol. 2, 876–881 (2001).
45. Sporici, R.A. & Perrin, P.J. Costimulation of memory T-cells by ICOS: a potential ther-apeutic target for autoimmunity? Clin. Immunol. 100, 263–269 (2001).
46. Sperling, A.I. ICOS costimulation: is it the key to selective immunotherapy? Clin.Immunol. 100, 261–262 (2001).
47. Hudson, L. & Hay, F.C. in Practical Immunology 2nd edn. (Blackwell ScientificPublications, St. Louis, MO, 1980).
48. Nishina, H. et al. Impaired CD28-mediated interleukin 2 production and proliferationin stress kinase SAPK/ERK1 kinase (SEK1)/mitogen-activated protein kinase kinase4 (MKK4)-deficient T lymphocytes. J. Exp. Med. 186, 941–953 (1997).
772 VOLUME 4 NUMBER 8 AUGUST 2003 NATURE IMMUNOLOGY
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reim
mu
no
log
y
Related Documents











