Cortinarius section Sanguinei in North America Tuula Niskanen 1 Kare Liimatainen Department of Biosciences, Plant Biology, P.O. Box 65, FI-00014 University of Helsinki, Finland Joseph F. Ammirati Department of Biology, Box 351330, University of Washington, Seattle, Washington 98195-1800 Karen Hughes Ecology and Evolutionary Biology, 437 Hesler Biology Building, University of Tennessee, Knoxville, Tennessee 37996-1100 Abstract: The North American species of Cortinarius section Sanguinei were studied using morphological characters and ITS and RPB2 sequence data. Several type collections also were examined. Four species were identified: C. harrisonii sp. nov, C. neosanguineus sp. nov., C. sanguineus and C. sierraensis comb. nov. Of these, C. sanguineus also occurs in Europe together with C. puniceus, a fifth member of the section. Typical features of these species include 6 red, fairly small basidiomata, stipe basal mycelium often with yellow to reddish yellow tints, amygdaloid to ellipsoid spores, and aniline-red lamellar trama and pileipellis hyphae when mounted in KOH. Two other species with red lamellae C. marylandensis comb. nov. and C. smithii stat. nov. & nom. nov. also are discussed. Key words: Cortinariaceae, Dermocybe, molecular systematics, multigene phylogeny, phylogeography INTRODUCTION Cortinarius is the largest genus of Agaricales and has worldwide distribution. It is a taxonomically difficult group, and there remain many undescribed species. Little is known about the distribution of the species within larger geographical areas or the differences in the species composition between continents. The most extensive taxonomic studies have concentrated on European species (e.g. Frøslev et al. 2007; Ortega et al. 2008; Niskanen et al. 2009, 2011; Sua ´rez-Santiago et al. 2009). Fewer studies have been completed for North American species and these have focused primarily on subgenus Phlegmacium (Moser and Peintner 2002; Garnica et al. 2009, 2011). A large study on Cortinarius species diversity from British Columbia, however, was published by Harrower et al. (2011). The species of sect. Sanguinei M.M. Moser (subge- nus Dermocybe) in North America have been studied by Ammirati (1972), Keller and Ammirati (1983), Ammirati and Smith (1984), Ammirati (1989) and Liu et al. (1995). Based on morphology all the species with red lamellae were included in the section, but in the first study to use molecular data Liu et al. (1995) indicated that section Sanguinei was polyphyletic. This result later was supported by the larger molec- ular study by Liu et al. (1997) of subgenus Dermocybe and the study by Niskanen et al. (2012) of the red species of Dermocybe in Europe. Based on these studies the section was delimited to include three validly described species, C. sanguineus (Wulfen : Fr.) Fr., C. puniceus P.D. Orton and D. sierraensis Ammirati 1989. Typical for the species are red basidiomata and pigments in the emodin series in addition to the dermorubin and dermocybin series (Keller and Ammirati 1983, Høiland 1984,). Dermocybe marylandensis Ammirati and C. phoeniceus var. occiden- talis A.H. Sm. were not included in sect. Sanguinei but were placed in/D. semisanguinea by Liu et al. (1997). Furthermore, Cortinarius californicus A.H. Sm. was shown to belong to subgenus Telamonia. The study by Niskanen et al. (2012) included only European species of sect. Sanguinei and no extensive molecular study of North American species has been done to date. Therefore we present here a treatment of the species in sect. Sanguinei from North America based on morphology and molecular data. MATERIALS AND METHODS Fungal specimens.—The specimens included in this study are provided (TABLE I). Material includes the species of subge- nus Dermocybe studied in Niskanen et al. (2012), except C. mirandus for which only the ITS1 region was succesfully amplified and some of the species included in sect. Sanguinei by Ammirati and Smith (1984). To improve the study of phylogenetic relationships, type specimens of some Dermo- cybe species described from North America were included. Herbarium acronyms follow Thiers (http://sciweb.nybg. org/science2/IndexHerbariorum.asp). Collectors abbreviat- ed by the initials JFA, KL and TN refer to the authors. Molecular analysis.— DNA was extracted from a few milligrams of dried material (a piece of lamella) with the NucleoSpin Plant kit (Macherey-Nagel). Primers ITS 1F and ITS 4 (White et al. 1990, Gardes and Bruns 1993) were used to amplify ITS regions. The primer combinations ITS1F/ ITS2 and ITS3/ITS4 were used on problematic material. Specific primers cort6F and b7.1R (Frøslev et al. 2005) were used to amplify the RPB2 6–7 region. The same primer pairs Submitted 20 Mar 2012; accepted for publication 24 Jul 2012. 1 Corresponding author. E-mail: [email protected] Mycologia, 105(2), 2013, pp. 344–356. DOI: 10.3852/12-086 # 2013 by The Mycological Society of America, Lawrence, KS 66044-8897 344

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cortinarius section Sanguinei in North America
Tuula Niskanen1
Kare LiimatainenDepartment of Biosciences, Plant Biology, P.O. Box65, FI-00014 University of Helsinki, Finland
Joseph F. AmmiratiDepartment of Biology, Box 351330, University ofWashington, Seattle, Washington 98195-1800
Karen HughesEcology and Evolutionary Biology, 437 Hesler BiologyBuilding, University of Tennessee, Knoxville, Tennessee37996-1100
Abstract: The North American species of Cortinariussection Sanguinei were studied using morphologicalcharacters and ITS and RPB2 sequence data. Severaltype collections also were examined. Four species wereidentified: C. harrisonii sp. nov, C. neosanguineus sp.nov., C. sanguineus and C. sierraensis comb. nov. Ofthese, C. sanguineus also occurs in Europe togetherwith C. puniceus, a fifth member of the section. Typicalfeatures of these species include 6 red, fairly smallbasidiomata, stipe basal mycelium often with yellow toreddish yellow tints, amygdaloid to ellipsoid spores,and aniline-red lamellar trama and pileipellis hyphaewhen mounted in KOH. Two other species with redlamellae C. marylandensis comb. nov. and C. smithiistat. nov. & nom. nov. also are discussed.
Key words: Cortinariaceae, Dermocybe, molecularsystematics, multigene phylogeny, phylogeography
INTRODUCTION
Cortinarius is the largest genus of Agaricales and hasworldwide distribution. It is a taxonomically difficultgroup, and there remain many undescribed species.Little is known about the distribution of the specieswithin larger geographical areas or the differences inthe species composition between continents. The mostextensive taxonomic studies have concentrated onEuropean species (e.g. Frøslev et al. 2007; Ortega et al.2008; Niskanen et al. 2009, 2011; Suarez-Santiago et al.2009). Fewer studies have been completed for NorthAmerican species and these have focused primarily onsubgenus Phlegmacium (Moser and Peintner 2002;Garnica et al. 2009, 2011). A large study on Cortinariusspecies diversity from British Columbia, however, waspublished by Harrower et al. (2011).
The species of sect. Sanguinei M.M. Moser (subge-nus Dermocybe) in North America have been studiedby Ammirati (1972), Keller and Ammirati (1983),Ammirati and Smith (1984), Ammirati (1989) andLiu et al. (1995). Based on morphology all the specieswith red lamellae were included in the section, but inthe first study to use molecular data Liu et al. (1995)indicated that section Sanguinei was polyphyletic.This result later was supported by the larger molec-ular study by Liu et al. (1997) of subgenus Dermocybeand the study by Niskanen et al. (2012) of the redspecies of Dermocybe in Europe.
Based on these studies the section was delimited toinclude three validly described species, C. sanguineus(Wulfen : Fr.) Fr., C. puniceus P.D. Orton and D.sierraensis Ammirati 1989. Typical for the species arered basidiomata and pigments in the emodin series inaddition to the dermorubin and dermocybin series(Keller and Ammirati 1983, Høiland 1984,). Dermocybemarylandensis Ammirati and C. phoeniceus var. occiden-talis A.H. Sm. were not included in sect. Sanguinei butwere placed in/D. semisanguinea by Liu et al. (1997).Furthermore, Cortinarius californicus A.H. Sm. wasshown to belong to subgenus Telamonia.
The study by Niskanen et al. (2012) included onlyEuropean species of sect. Sanguinei and no extensivemolecular study of North American species has beendone to date. Therefore we present here a treatmentof the species in sect. Sanguinei from North Americabased on morphology and molecular data.
MATERIALS AND METHODS
Fungal specimens.—The specimens included in this study areprovided (TABLE I). Material includes the species of subge-nus Dermocybe studied in Niskanen et al. (2012), except C.mirandus for which only the ITS1 region was succesfullyamplified and some of the species included in sect. Sanguineiby Ammirati and Smith (1984). To improve the study ofphylogenetic relationships, type specimens of some Dermo-cybe species described from North America were included.Herbarium acronyms follow Thiers (http://sciweb.nybg.org/science2/IndexHerbariorum.asp). Collectors abbreviat-ed by the initials JFA, KL and TN refer to the authors.
Molecular analysis.—DNA was extracted from a fewmilligrams of dried material (a piece of lamella) with theNucleoSpin Plant kit (Macherey-Nagel). Primers ITS 1F andITS 4 (White et al. 1990, Gardes and Bruns 1993) were usedto amplify ITS regions. The primer combinations ITS1F/ITS2 and ITS3/ITS4 were used on problematic material.Specific primers cort6F and b7.1R (Frøslev et al. 2005) wereused to amplify the RPB2 6–7 region. The same primer pairs
Submitted 20 Mar 2012; accepted for publication 24 Jul 2012.1 Corresponding author. E-mail: [email protected]
Mycologia, 105(2), 2013, pp. 344–356. DOI: 10.3852/12-086# 2013 by The Mycological Society of America, Lawrence, KS 66044-8897
344

TABLE I. Specimens included in this study
Species Voucher Herb. Locality
GenBank
GenBank nameaITS RPB2
Section Sanguinei
C. harrisoniiAmmirati,Niskanen & Liimat.
RH2331 NYBG USA, Massachusetts U56060 D. sanguinea
C. harrisonii(holotype)
ASM3128 EIU USA, North-Carolina JX045675
C. harrisonii KH11308 DAOM USA, North-Carolina JX045676C. harrisonii AHS14870 MICH USA, Tennessee U56069 D. subcinnabarinaC. harrisonii TENN61657 TENN USA, Tennessee JX045677C. neosanguineus
Ammirati,Liimat. & Niskanen
F16299 UBC Canada, BritishColumbia
EU486445 C. sanguineus
C. neosanguineus SMI171 UBC Canada, BritishColumbia
FJ157116 C. sanguineus
C. neosanguineus F14888 UBC Canada, BritishColumbia
FJ157089 C. sanguineus
C. neosanguineus JFA11655 WTU USA, Oregon JX045680C. neosanguineus AHS79922 MICH USA, Oregon U56059 D. sanguineaC. neosanguineus JFA8533 WTU USA, Washington U56056 D. sanguineaC. neosanguineus JFA11548 WTU USA, Washington JX045679 JX045702C. neosanguineus
(holotype)TN09-130 H (WTU) USA, Washington JX045678
C. puniceus P.D.Orton (5C.rubrosanguineus(holotype))
G00110215 G France JN114091 JX045703
C. puniceus 7371 United Kingdom FJ876159 C. sp.C. sanguineus
(Wulfen: Fr.) Fr.TN07-329 H Canada Newfoundland JX045684
C. sanguineus TN10-071 H Canada Newfoundland JX045682C. sanguineus KH14.10.1974 DAOM Canada Nova Scotia JX045683C. sanguineus DM9.9.73/27 TRTC Canada Ontario U56046 D. mallochiiC. sanguineus TN10-023 H Canada Quebec JX045681C. sanguineus AHS74847 MICH USA, Michigan JX045685C. sanguineus JFA9748 WTU Austria U56057 D. sanguineaC. sanguineus TN04-561 H Finland JN114102C. sanguineus var.
aurantiovaginatusFillion & Moenne-Locc. (holotype)
G00110216 G France JN114100
C. sanguineus TUB011822 TUB Germany AY669582C. sanguineus 1726 MCVE Italy JF907874C. sanguineus TN06-014 H Norway JN114106C. sanguineus IK03-003 H Slovakia JN114108C. sanguineus
(neotype)SL22091940 UPS Sweden JN114099
C. sanguineus CFP737 S Sweden JN114101 JX045704C. sierraensis
(Ammirati)Ammirati,Niskanen & Liimat.(holotype)
HDT32671 SFSU USA, California U56068,JX045687
D. sierraensis
C. sierraensis HDT21106 SFSU USA, California JX045686C. sierraensis DBB03415 DBb USA, California JX045688
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 345

TABLE I. Continued
Species Voucher Herb. Locality
GenBank
GenBank nameaITS RPB2
Other Dermocybe species
C. aurantiobasisAmmirati & A.H.Sm. (holotype)
AHS24213 MICH USA, Oregon JX045670
C. bataillei (J. Favreex Mos.) Høil.
CFP1078 S Switzerland JX045666 JX045697
C. cascadensisAmmirati & A.H.Sm. (holotype)
JFA6156 MICH USA, Oregon JX045671
C. chrysolitusKauffman(holotype)
19 Sept. 1914,Barcode10332
MICH USA, New York JX045672
C. cinnamomeus(L.) Fr.
CFP623 S Sweden JN114080 JX045698
C. cistoadelphus(G. Moreno,Poder, Kirchm.,Esteve-Rav. &Heykoop) G.Moreno(holotype)
AH18359 WTU Spain JN114078 JX045699
C. croceoconus Fr. CFP429 S Sweden JX045674 JX045706C. croceus (Schaeff.)
GrayCFP651 S Sweden JN114079 JX045700
C. fervidus P.D.Orton
CFP334 S Sweden JN114087 JX045705
C. humboldtensisAmmirati & A.H.Sm. (holotype)
AHS56392 MICH USA, California JX045673
C. idahoensisAmmirati & A.H.Sm. (holotype)
AHS71073 MICH USA, Idaho JX045669
C. malicorius Fr. TN04-406 H Finland JX045668 JX045701C. marylandensis
(Ammirati) Ammirati,Niskanen & Liimat.
JFA12010 WTU Costa Rica JX045694
C. marylandensis DMS1772 WTU USA, Delaware JX045692C. marylandensis JFA7935 WTU USA, Florida U56047C. marylandensis
(holotype)JFA7936 WTU USA, Florida U56048,
JX045690C. marylandensis SM053189 WTU USA, Texas JX045693C. marylandensis JFA6481 WTU USA, Virginia JX045691C. marylandensis JFA6482 WTU USA, Virginia U56070 C. sp.C. phoeniceus
(epitype)CFP742 S Sweden JN114084
C. semisanguineus(Fr.) Gillet
CFP333 S Sweden JN114090
C. smithii Ammirati,Niskanen & Liimat.
VMS7 UBC Canada, BritishColumbia
FJ717526 C. sp.
C. smithii (holotype) AHS3485 MICH USA, Oregon JX045689C. smithii JFA9886 WTU USA, Washington U56054 D. phoeniceaC. sommerfeltii Høil. CFP594 S Sweden JN114083 JX045695C. uliginosus Berk. CFP1049 S Sweden JN114081
346 MYCOLOGIA

were used in direct sequencing. PCR amplification andsequencing followed Niskanen et al. (2009). Sequences wereassembled and edited with Sequencher 4.1 (Gene CodesCorp., Ann Arbor, Michigan). A total of 29 new ITS sequencesand 12 RPB2 sequences were produced for this study.
Intragenomic polymorphisms were observed as mixedpeaks in chromatographic data. The ITS sequences werecompared with the material in the public databases(GenBank: http://www.ncbi.nlm.nih.gov/ and UNITE:http://unite.ut.ee/) with BLAST queries. A sequence ofevery species was compared with all other sequences withBLAST to estimate genetic distances. These sequences andthe sequences of the closest ones were aligned withMUSCLE (Edgar 2004) on the European BioinformaticsInstitute server (http://www.ebi.ac.uk/Tools/muscle/index.html). These differences were counted by eye fromthe alignments: (i) the observed number of variable sitesshowing how many sites contain intraspecific and/orintragenomic polymorphisms; and (ii) the differencesbetween the closest species as minimum evolutionary eventsincluding indels (multiple base indels treated as onechange), transitions and transversions. Only differencesshared by all specimens of the same species were counted.
Based on Garnica et al. (2005) we chose Phlegmacium asan outgroup for our phylogenetic analysis. The combinedITS and RPB2 alignment from sequences of 63 specimenswas produced with MUSCLE (Edgar 2004) under defaultsettings and followed by manual adjustments in BioEdit(www.mbio.ncsu.edu/BioEdit/bioedit.html). The align-ment is 1416 nucleotides long (including gaps) andavailable at TreeBASE under S12726 (http://www.treebase.org/treebase-web/home.html).
Bayesian inference (BI) was performed with the programMrBayes 3.1.1 (Huelsenbeck and Ronquist 2003). Weanalyzed the entire dataset with the GTR model includinga gamma-shape parameter and estimating the proportion ofinvariable sites. Two independent runs with four chains ineach were performed for 3 000 000 generations withsampling at every 100th generation. All trees sampledbefore stationarity were discarded using a 25% safetymargin (burn-in of 7500 trees, 750 000 generations). The
sampled trees were combined in a 50% majority ruleconsensus phylogram with posterior probabilities (PP). Theanalysis was run on computer clusters of the CSC, IT Centerfor Science, Espoo, Finland.
Morphological studies.—Morphological descriptions arebased on material collected by the authors and/or othermycologists. Capitalized color notations in descriptions offresh characteristics are from Ridgway (1912), uncapitilizedcolor terms are regarded as useful approximations. Thenames used in the descriptions of exsiccata colors followMunsell (2009) soil color charts. UV observations weremade on dried specimens at 366 nm. Microscopiccharacteristics were observed from dried material mountedin 5% KOH, except the dextrinoidity of the spores wasobserved in Melzer’s reagent. Measurements were madewith an ocular micrometer with 100 3 oil-immersion lens.Spore measurements are from those deposited on the theveil or apex of the stipe; 20 basidiospores were measuredfrom one basidiome from each collection. The length andwidth were measured from each spore, and their length/width ratios (Q value) were calculated. Average values formeasurements from one basidiome were counted, andtheir range is indicated in point averages/Q averagesunder each species. The lamellar trama and basidia alsowere examined. The pileipellis structure was studied fromboth radial freehand sections and scalps from the pileuscenter or midway between the center and margin. Themeasurements of the elements of pileipellis were madefrom radial freehand sections.
RESULTS
Molecular analysis.—Based on our phylogeneticanalysis (FIG. 1) sect. Sanguinei is strongly supported(PP 1.00) and includes five well supported species (PP1.00). The section is further split into two groups,although with low support. The first group (PP 0.54)comprises western North American species C. neo-sanguineus and C. sierraensis and the second (PP
TABLE I. Continued
Species Voucher Herb. Locality
GenBank
GenBank nameaITS RPB2
C. vitiosus (M.M.Moser) Niskanen,Kytov., Liimat. &S. Laine
TN 04-221 H Finland JN114095 JX045696
C. vitiosus EO44 OULU Russia, SakhaRepublic, Yakutia
JX045667
C. vitiosus (holotype) MM1974/0117 IB Sweden JN114098
Outgroup
C. barbarorum TSJ2000-069 C Italy DQ083773 DQ083881C. sodagnitus TF2001-094 C Denmark DQ083812 DQ083920
a GenBank name 5 Name in the GenBank database if different from the current name.b DB 5 D. Bojantchev pers herb.
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 347
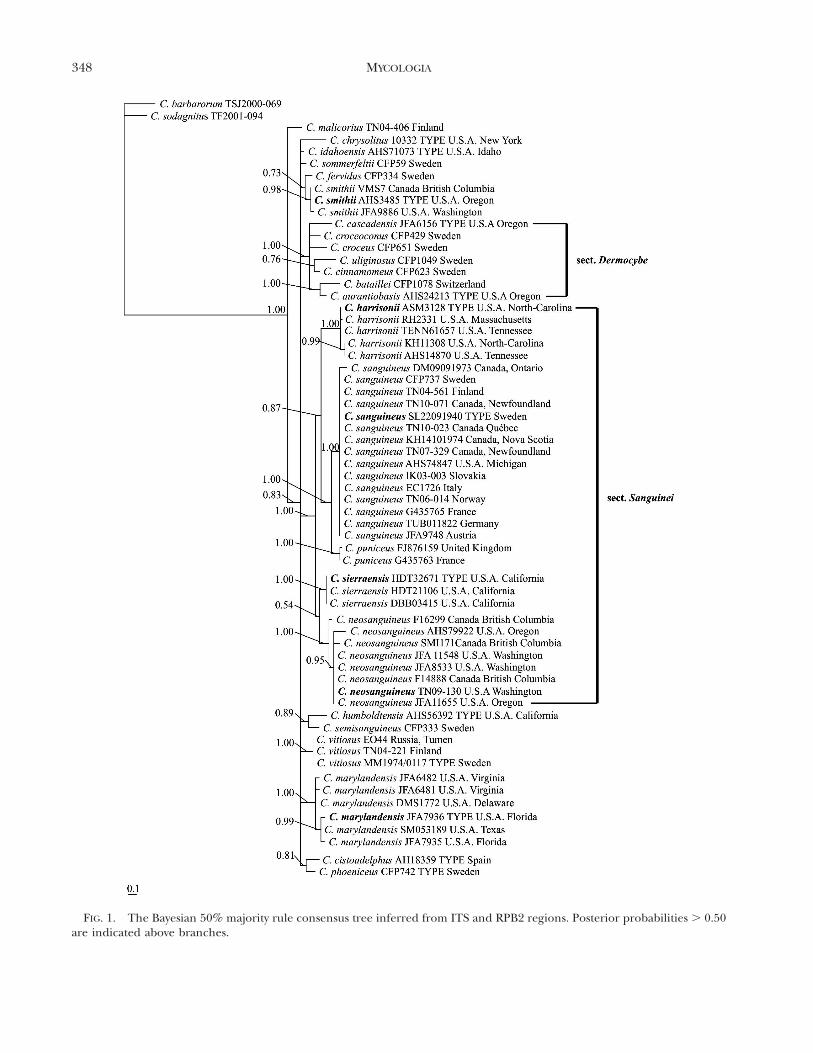
FIG. 1. The Bayesian 50% majority rule consensus tree inferred from ITS and RPB2 regions. Posterior probabilities . 0.50are indicated above branches.
348 MYCOLOGIA

0.87) eastern North American and European C.harrisonii, C. puniceus and C. sanguineus. As in Liuet al. (1997) C. marylandensis and C. smithii (syn. C.phoeniceus var. occidentalis) did not group togetherwith species in sect. Sanguinei. Furthermore, thelatter is not related to C. phoeniceus but instead to C.fervidus P.D. Orton (PP 0.73). The species arepresented in TAXONOMY.
From other groups of subgenus Dermocybe the sect.Dermocybe is also well supported (PP 1.00) in ouranalysis. In addition C. humboldtensis Ammirati &A.H. Sm./C. semisanguineus (Fr.)Gillet (PP 0.89) andC. cistoadelphus (G. Moreno, Poder, Kirchm., Esteve-Rav. & Heykoop) G. Moreno/C. phoeniceus (Vent.)Maire (PP 0.81) receive some support.
All species formed well supported, monophyleticgroups in our analysis. They also are distinct asdefined by their morphological differences and havesmaller intraspecific genetic variation compared withthe interspecific variation. The two closest Sanguineispecies, C. sanguineus and C. puniceus, differ by sixevolutionary events in their ITS regions while thedifference of C. sierraensis and C. neosanguineus is 11evolutionary events and that of C. sanguineus and C.harrisonii 16 evolutionary events. Intragenomic poly-morphisms were observed in 9–50% of the sampleswithin species. Polymorphisms were not observed intwo species, C. sierraensis and C. neosanguineus.
TAXONOMY
Cortinarius sect. Sanguinei Kuhner & Romagn. exM.M. Moser, Beih Nova Hedwig 52:480 (1975) em.Niskanen, Liimat. & AmmiratiType species: Cortinarius sanguineus (Wulfen : Fr.) Fr.
Basidiomata 6 red, fairly small. Universal veil sparse,often indistinct. Basal mycelium at least in exsiccataoften reddish yellow to yellow, in C. puniceus and C.sierraensis also weak red to pale red. Spores amygdaloidto ellipsoid, moderately verrucose, nondextrinoid toweakly dextrinoid, rarely moderately dextrinoid. La-mellar trama hyphae not encrusted, aniline red inKOH. Scalp sections of the pileipellis aniline red inKOH. Major pigments include the emodin seriesin addition to the dermorubin and dermocybin series,but not confirmed for C. harrisonii (Keller andAmmirati 1983, Høiland 1984). With conifers ordeciduous trees. Distribution, as presently known, inthe northern hemisphere.
From other described North American species, onlythe red C. marylandensis (Ammirati) Ammirati,Niskanen & Liimat. is reminiscent of the species ofsect. Sanguinei. It occurs from Costa Rica throughoutthe southern and eastern United States north to
Delaware and is associated with hardwoods. Thecoloration of the basidiomata is closest to C.sierraensis and C. harrisonii, but C. harrisonii hassomewhat smaller spores and C. sierraensis has largerspores than C. marylandensis. Based on pigmentchemistry C. marylandensis differs from the speciesof section Sanguinei by having endocrocin but notemodin (Keller and Ammirati 1983, Liu et al. 1995).
1. Cortinarius sanguineus (Wulfen : Fr.) Fr., Epicr.Syst. mycol.: 288 (1838). FIG. 2A
Basionym: Agaricus sanguineus Wulfen in Jacquin, Miscell.austriac. 2:107 (1781): sanctioned in Fr., Syst. mycol.1:229 (1821). Type: SWEDEN. Smaland: Femsjo, 22 Sep1940, S. Lundell (UPS, NEOTYPE, designated by Høiland1984). GenBank No. JN114099.
Cortinarius sanguineus var. aurantiovaginatus Fillion &Moenne-Locc. in Bidaud et al., Atlas des Cortinaires6:192 (1994). Type: FRANCE. Hte-Savoie: Plateau desGlieres, on an old stump of Picea abies, 1500 m, 3 Aug1993, P. Moenne-Loccoz 3475, G00110216 (G, HOLO-TYPE). GenBank No. JN114100.
Illustrations: Bidaud et al. (1994: pl. 129), Brandrudet al. (1989: pl. A57).
Pileus 2–5 cm broad, hemispherical, later lowconvex to almost plane, sometimes slightly umbo-nate, fibrillose-tomentose, often with fibrillosescales, dry; rich red (Garnet Brown) to deep red(Maroon to deep Garnet Brown) or tinted with themargin, margin rich red (Garnet Brown to Ox-Blood Red) or duller red (Coral Red), sometimesstreaked deep red to fuscous red (Maroon toVictoria Lake). Lamellae medium-spaced, emargi-nated to adnexed, at first rich red (Ox-Blood Red,dull Carmine or Garnet Brown). Stipe 4–10 cmlong, 0.3–0.8 cm thick, cylindrical or slightly clavate,rich red (Garnet Brown, Ox-Blood Red) or dullerred (Coral Red), paler than the pileus. Cortina darkred, ochraceous or golden brown. Universal veilcarmine red, fibrillose, sparse and difficult toobserve. Basal mycelium dull ochraceous (Ochra-ceous-Buff). Context in pileus watery deep red(Maroon to Garnet Brown), faded to lighter red(near Coral Red). In stipe rich red (Garnet Brown,Ox-Blood Red) or deeper red (Maroon) in thelower portion. Odor in lamellae cedar-like, especial-ly when slightly dried. Exsiccata: Pileus dusky red(7.5R 3/3–3/4, 10R 3/3), reddish brown (2.5YR 4/4) to dark reddish brown (2.5YR 3/3–3/4), stipeweak red (10R 4/3–4/4) with reddish yellow (5YR6/8–7/8, 7.5YR 7/8) mycelium. UV observationsnegative, whole basidiome.
Spores 7–8(–8.5) 3 4.5–5.2 mm, av. 5 7.4–7.7 3 4.6–4.7 mm, Q 5 1.45–1.7(–1.75), Qav. 5 1.59–1.62 (fiveNorth American collections), amygdaloid to ellipsoid,
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 349

moderately verrucose, weakly dextrinoid. Lamellartrama hyphae not encrusted, with aniline red pigmentand granules in KOH. Lamellar edge partly fertile,especially young basidiomata with clavate to broadlyclavate marginal cells 10–24 3 6–11 mm. Basidia four-spored, 22.5–30.5 3 6.0–8.0 mm, with granulosecontent, in KOH hyaline to aniline red. Pileipellis acutis with ascending terminal elements. Scalp prepa-ration aniline red in KOH. Uppermost hyphae (about4–6 layers of hyphae) 5–15(–20) mm wide, hyaline topinkish red or grayish pink, not or finely encrusted.Lower hyphae 5–15 mm wide, often aniline red withaniline red granules, not encrusted. Hypoderm notdifferentiated. Clamp connections present.
Ecology and distribution: In hemiboreal to borealmossy coniferous forests, also in mountainous areas,at least with Picea perhaps also with Abies and Tsuga,often on damp and somewhat richer ground. Mid-Julyto October. Known in eastern North America, fromMichigan (USA.) to Newfoundland (Canada). Occursalso in northern Europe and mountainous areas ofcentral and southern Europe.
Differential diagnosis: Typical for C. sanguineus isfibrillose-tomentose to fibrillose scaly cap and occur-rence in hemiboreal to boreal coniferous forests.Cortinarius neosanguineus is most similar to C.sanguineus but differs from the latter by somewhatdarker, purplish red pileus and generally a moreaniline-red lamellar trama hyphae in KOH. Inaddition, the pileus in exsiccata is a bit more brownishyellow and the basal mycelium more yellow. Cortinar-ius neosanguineus occurs in the Pacific Northwest ofNorth America, while C. sanguineus is currentlyknown from eastern North America and Europe.Cortinarius harrisonii occurs in the eastern UnitedStates and C. sierraensis in the Sierra Nevada; bothhave more red orange to brown orange pileus than C.sanguineus. These differences are clear also in theexsiccata. The spores of C. harrisonii are the smallestof section Sanguinei while the spores of C. sierraensisare the largest of the group.
In our phylogenetic analysis C. sanguineus speci-mens from North America and Europe formed aclade with strong support. However, one intraspecificvariation site in the ITS regions exists where NorthAmerican collections have a T and European collec-tions have a C or Y (intragenomic base polymor-phisms). Morphologically they do not differ, exceptthe spores of the European collections were onaverage slightly broader (Qav. 5 1.53–1.58). Inaddition, DM09091973/U56046 (Canada, Ontario)has a couple of unique differences in ITS region, butbased on morphology it also fits in C. sanguineus.
Specimens examined: CANADA. NEWFOUNDLAND ANDLABRADOR: Avalon Peninsula, Brother Brennan Environmental
Education Center, conifer-dominated forest, 29 Sep 2007,M. Voitk (TN 07-329), H7001143 (H). Northern Peninsula,Pistolet Bay Provincial Park, Picea-dominated forest, 7 Sep2010, M. Burzynski (TN 10-071) (H). NOVA SCOTIA:Richmond, Isle Madame, 14 Oct 1974, K. Harrison(DAOM). ONTARIO: Black Sturgeon Lake, conifer dom-inated forest, 9 Sep 1973, D. Malloch (WTU). QUEBEC:Hervey-Junction, near the railroad, coniferous forest, 27Aug 2010, KL & TN 10-023 (H). USA. MICHIGAN:Cheboygan County, under conifers, 8 Aug 1964, A.H.Smith 74847 (MICH).
2. Cortinarius harrisonii Ammirati, Niskanen &Liimat. sp. nov. FIG. 2B
MycoBank MB800333Cortinarius sanguineus var. subcinnabarinus Ammirati &
A.H. Sm., McIlvainea 6:62 (1984), nom. inval., Art. 36.1.
Basidiomata red-orange to red. Pileus innatelyfibrillose or with small, recurved fibrillose scales onthe margin. Exsiccata reddish brown to red. Sporesellipsoid to amygdaloid, 7–8(–8.5) 3 4–5 mm.
Typus: USA. North-Carolina: Macon County, High-land, Sunset Road Trail, 21 Aug 1984, gregarious, soiland duff of road cut, hardwoods and Tsuga, A. S.Methven 3128 (EIU, HOLOTYPE). GenBankNo. JX045675.
Etymology: The species is named in honor ofKenneth A. Harrison Sr., a mycologist from Wolfville,Nova Scotia.
Pileus 3–6(–9) cm broad, convex to obtusely conic,becoming conic-umbonate or broadly convex toplane in age, innately fibrillose or with small, recurvedfibrillose scales on the margin, disk often smooth andnearly glabrous, dry; deep red-orange (Brazil Red) tobrownish orange (Sanford’s Brown) or brownish red(Burnt Sienna) overall or disk deeper red (almostGarnet Brown) at times. Lamellae almost crowded,adnate with a slight tooth or adnexed, deep orange-red (English Red) to darker red, becoming brownishorange (Xanthine Orange) to orangish brown (Am-ber Brown). Stipe 4–8 cm long, 0.4–0.8 cm thick,equal, evenly deep rusty orange (Orange Rufous),becoming sordid reddish brown in places with age.Cortina no observations. Universal veil indistinct.Basal mycelium yellow. Context in pileus thick, firm,dull pinkish vinaceous (Old Rose). In stipe solid, darkreddish except for narrow cortex which is 6
concolorous with the stipe surface. Odor indistinctive.Exsiccata pileus reddish brown (2.5YR 5/4–4/4), red(2.5YR 5/6) to yellowish red (5YR 5/6), stipe red (2.5YR 5/6–4/6), reddish brown (2.5YR 4/4) to yellowishred (5YR 5/6) with reddish yellow (7.5YR 6/8–7/8) toalmost yellow (10YR 8/8–7/8) mycelium. UV obser-vations negative, whole basidiome.
Spores 7–8(–8.5) 3 4–5 mm, av. 5 7.2–7.5 3 4.3–4.5 mm, Q 5 1.55–1.85(–1.90), Qav. 5 1.60–1.76 (five
350 MYCOLOGIA

collections), ellipsoid to slightly amygdaloid, moder-ately verrucose, nondextrinoid to weakly dextrinoid.Lamellar trama hyphae not encrusted, aniline red inKOH. Lamellar edge partly fertile, especially youngbasidiomata with clavate- to balloon-shaped marginalcells, 10.5–17 3 7–10 mm. Basidia four-spored, 23–303 5–7.5 mm, in KOH hyaline or aniline red. Pileipellisa cutis with few ascending terminal elements. Scalppreparation aniline red in KOH. Uppermost hyphae(about 4–6 layers of hyphae) 5–15 mm wide, hyaline,not encrusted. Lower hyphae 5–20 mm wide, oftenaniline red, some with aniline red granules, notencrusted. Hypoderm not differentiated. Clampconnections present.
Ecology and distribution: In deciduous hardwood andmixed forests, often with Tsuga. August to September.
Known only from eastern North America, fromMassachusetts into the Appalachian mountain range.
Differential diagnosis: Cortinarius harrisonii is a red-orange to red species with innately fibrillose, tofibrillose scaly pileus and small spores. The pileus inrelation to stipe is often wide, as it also can be in C.sanguineus. Cortinarius sanguineus differs from C.harrisonii by deeper red basidiomata, also seen inexsiccata, and the larger spores. Cortinarius sierraensishas more brownish pileus and larger spores. Also thedistribution is very different; to date C. sierraensis isknown only from Sierra Nevada, California.
Specimens studied: USA. MASSACHUSETTS: HampshireCounty, vicinity of Skinner State Park, Route 47, Hadley, 24Sep 1977, R. Halling 2331 (NYBG). NORTH CAROLINA:Macon County, Highland, Sunset Road Trail, gregarious,
FIG. 2. A. Cortinarius sanguineus TN04-56. B. C. harrisonii TENN61657. C. C. neosanguineus TN09-130. D. C. sierraensisDBB03415. A, C by K. Liimatainen, B by Karen Hughes, D by Dimitar Bojantchev.
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 351

soil and duff of road cut, hardwoods and Tsuga, 21 Aug1984, A.S. Methven 3128 (EIU–HOLOTYPE). Swain Coun-ty, Deep Creek, under Rhododendron in deciduous woods,wet area, 25 Sep 1971, K. Harrison 11308 (DAOM).TENNESSEE: Elkmont, 17 Aug 1939, A.H. Smith 14870(MICH). Sevier County, Great Smoky Mountains NationalPark, Greenbrier, Old Settlers Trail, under mixed hardwoodand Tsuga, 25 Sep 2006, R. Tulloss 13394, TENN61657(TENN).
3. Cortinarius neosanguineus Ammirati, Liimat. &Niskanen sp. nov. FIG. 2C
MycoBank MB800334Cortinarius purpureosanguineus Ammirati, 1972:76–80, in-
valid, Latin description published in dissertation only.Form of Cortinarius sanguineus, McIlvanea 6(2): 61–62
(1984).
Basidiomata vivid red to purplish red. Pileusminutely to distinctly fibrillose-squamulose. Exsiccatareddish brown with reddish yellow to yellow myceli-um. Spores slightly amygdaloid to ellipsoid, 7–8.5(–9)3 4.5–5.2(–5.5) mm. In coniferous forests of PacificNorthwest of North America.
Type: USA. Washington: Olympic Peninsula, SolDuc Trail, 25 Oct 2009, old, mossy coniferous forestdominated by Pseudotsuga menziesii and Tsugaheterophylla with some Picea sitchensis, Liimatainen& Niskanen 09-130 (H–holotype; WTU isotype).GenBank No. JX045678.
Etymology: The epithet refers to the similarity withC. sanguineus with which it has been confused.
Pileus 1–4 cm diam, broadly conic to convex orbroadly convex, margin straight at first, minutely todistinctly fibrillose-squamulose, some becoming ri-mose, dry; vivid red (Rose Red) to slightly pinkeroverall or with dark purplish red (Bordeaux) disk,often more purple red when mature, sometimes witha discolored (blackish) zone near the margin.Lamellae: medium spaced, adnate to adnexed, vividred (Rose Red) to dark purplish red (Bordeaux,Pansy Purple), in age duller red, edges even usuallyretaining red to purple red tones. Stipe 3–6 cm long,0.2–0.4 cm thick, equal, appressed fibrillose to silky,concolorous with lamellae or slightly paler, stainingdarker (purple red) on base when handled. Cortinaconcolorous with lamellae. Universal veil indistinct.Basal mycelium vinaceous ochraceous to reddishyellow, sometimes coating the lower part of the stipe.Context in pileus thin, concolorous with surface. Instipe solid, concolorous with surface throughout.Odor in lamellae cedar-like or not distinctive.Exsiccata pileus reddish brown (2.5YR 4/4) to darkreddish brown (2.5YR 3/3–3/4), sometimes moredusky red (10R 3/3), stipe weak red (10R 4/4) toreddish brown (2.5YR 4/4) with reddish yellow(7.5YR 6/8–7/8) to almost yellow (10YR 8/8–7/8)
mycelium. UV observations negative, whole basi-diome.
Spores 7–8.5(–9) 3 4.5–5.2(–5.5) mm, av. 5 7.5–8.13 4.8–5.0 mm, Q 5 1.5–1.8, Qav. 5 1.54–1.67 (eightcollections), slightly amygdaloid to ellipsoid, moder-ately verrucose, weakly to moderately dextrinoid.Lamellar trama hyphae not encrusted, stronglyaniline red in KOH, in places with aniline red,granulose content. Lamellar edge partly fertile,especially young basidiomata with clavate to bal-loon-shaped marginal cells, 10.5–22 3 8–12 mm.Basidia four-spored, 24–32 3 6–9 mm, often withgranulose content, in KOH hyaline or aniline-red.Pileipellis a cutis with some ascending terminalelements. Scalp preparation aniline red in KOH.Uppermost hyphae (about 3–5 layers hyphae) 5–20 mm wide, pinkish red to grayish pink, most hyphaenot encrusted but often some of the uppermosthyphae finely or zebra-striped encrusted. Lowerhyphae 5–20(–25) mm wide, often aniline red withaniline red and bluish purple granules, not encrust-ed. Hypoderm not differentiated. Clamp connec-tions present.
Ecology and distribution: In coniferous forests(Pseudotsuga, Tsuga, Abies, Picea). September toNovember. Known from western North America, inthe Pacific Northwest and south into California.
Differential diagnosis: Cortinarius neosanguineus isa vivid red to dark purplish red species with afibrillose-squamulose pileus. Typically the width ofthe pileus is small compared to the length of thestipe. Cortinarius sanguineus can be similar, butoften the pileus is wider in relation to stipe.Cortinarius sanguineus also has a bit lighter pileus,and less aniline-red pigment in lamellae prepara-tions in KOH. The exsiccata are somewhat redderand the basal mycelium is reddish yellow. To datethese two species have not been observed to co-occurin any region of North America. Cortinariussierraensis and C. harrisonii differ from C. neosan-guineus by red-orange to brown-orange basidiomata,difference also observed in exsiccata. In addition,the spores of C. sierraensis are larger and the sporesof C. harrisonii smaller than the ones of C.neosanguineus.
Specimens examined: CANADA. BRITISH COLUMBIA:Capilano River Regional Park, 3735 Capilano Road, NorthVancouver, 100 m, on the ground at the base of conifertrees, usually surrounded by a bed of moss, 27 Oct 2007, V.Woloshen, A. Macdonald, M. Ignaszewski vw271007, F16299(UBC). Oona River (Porcher Island), along river, 22 Oct2003, M. Kranabetter SMI171, F18509 (UBC). SaturnaIsland, Ecological Reserve 15, Trail 5/polygon, 22, 30 m,coniferous forest, 22 Oct 2000, P. Janszen 318 ER, F14888(UBC). USA. OREGON: Polk County, van Duzer Corridor,Tsuga heterophylla, Pseudotsuga menziesii, Picea sitchensis, 14
352 MYCOLOGIA

Nov 1970, D. Goetz (AHS 79922) (MICH). WASHINGTON:King County, Watermain Woods, damp conifer woods, 24Oct 1981, JFA 8533 (WTU). Near Mount Hood, PioneerWoman’s Grave, on rotten conifer wood, forest ofPseudotsuga, Tsuga and Abies, E. Horak (JFA 11655)(WTU). Olympic Peninsula, Big Quilcene River Trail,BHWA, Abies, Tsuga, Pseudotsuga, in deep humus, 11 Oct1995, JFA 11548 (WTU). Olympic Peninsula, Sol Duc Trail,25 Oct 2009, old, mossy coniferous forest dominated byPseudotsuga menziesii and Tsuga heterophylla with somePicea sitchensis, KL & TN 09-130 (H, HOLOTYPE; NY,ISOTYPE).
4. Cortinarius sierraensis (Ammirati) Ammirati, Nis-kanen & Liimat. comb. nov. FIG. 2D
MycoBank MB800335Basionym: Dermocybe sierraensis Ammirati, Mycotaxon 34:33
(1989). Type: U.S.A. California: Alpine County, BlueLakes, in soil under lodgepole pines (Pinus contorta), 17Aug 1974, H.D. Thiers 32671 (SFSU, holotype). GenBankNo. U56068, JX045687.
Cortinarius sanguineus var. sierraensis Gerw. Keller &Ammirati, Mycotaxon 18:364 (1983), nom.inval., Art. 36.1.
Cortinarius sanguineus var. sierraensis Ammirati & A.H.Sm., McIlvanea 6:62–63 (1984), nom.inval., Art. 36.1.
Pileus 2–4 cm diam, convex when young becom-ing plano-convex to plane, margin incurved todecurved to plane, no veil fibrils on edge or surface,glabrous to finely silky appressed fibrillose, usuallyshiny with age, dry; brown orange (CinnamonRufous) to brownish orange (Tawny) during allstages of development. Lamellae close, adnate toadnexed, narrow, reddish brown (Hays Russet) toorange red (Grenadine Red) during all stages ofdevelopment, edges concolorous with faces. Stipe 2–4 cm long, 0.2–0.4 cm thick, cylindrical, deepvinaceous to almost concolorous with pileus surface.Cortina no observations. Universal veil fibrils con-colorous with stipe surface. Basal mycelium vina-ceous to yellowish orange. Context in pileus pink,unchanging when exposed or bruised, 1–2 mmthick, in stipe solid, concolorous with surface. Odorindistinct or slightly raphanoid. Exsiccata pileusreddish brown (2.5YR 5/4–4/4), stipe reddishbrown (2.5YR 5/4–4/4) to weak red (10R 4/4) withlight reddish (2.5YR 6/4) mycelium with somereddish yellow patches (7.5YR 7/8–6/8). UV obser-vations negative, whole basidiome.
Spores 7.5–9 3 5–5.5(–5.8) mm, av. 5 8.0–8.4 3
5.2–5.3 mm, Q 5 1.45–1.7, Qav. 5 1.53–1.58 (threecollections), ellipsoid, less commonly somewhatamygdaloid, moderately verrucose, indextrinoid. La-mellar trama hyphae not encrusted, aniline red, withsome aniline red granules. Lamellar edge partlyfertile, especially young basidiomata with clavate tosometimes balloon-shaped marginal cells, 13–20 3
8–10 mm. Basidia four-spored, 23–30 3 6.5–8.5 mm,hyaline to aniline red. Pileipellis a cutis with someascending terminal elements. Scalp preparationaniline red in KOH, sometimes with orange tints.Uppermost hyphae (about 2–4 layers of hyphae) 5–15 mm wide, not or finely encrusted. Lower hyphae 5–20 mm wide, often aniline red with aniline redgranules, not encrusted. Hypoderm not differentiat-ed. Clamp connections present.
Ecology and distribution: Under lodgepole pine(Pinus contorta). August and September. To dateknown only from the higher elevations of the SierraNevada, California.
Differential diagnosis: Cortinarius sierraensis is rec-ognized by the brownish red-orange, silky fibrillosepileus and large spores. Cortinarius harrisonii has themost similar basidiomata coloration but has muchsmaller spores and occurs in eastern North America.Cortinarius sanguineus and C. neosanguineus aredarker red, have smaller spores and has a westerndistribution.
Specimens examined: USA. CALIFORNIA: Alpine County,Blue Lakes, in soil under lodgepole pines, 17 Aug 1974,H.D. Thiers 32671 (SFSU–HOLOTYPE). Mariposa County,Yosemite National Park, Hwy 120 between Porcupine Flatand Crane’s Flat, in humus under conifers, 14 Sep 1967,H.D. Thiers 21106 (SFSU). Yosemite National Park, off Hwy120, Tioga Pass, in an area of small lakes and lodgepolepine, 17 Aug 2008, D. Bojantchev DBB03415 (D. Bojantchevpers herb)
COMMENTS ON SOME OTHER SPECIES WITH RED LAMELLAE
5. Cortinarius marylandensis (Ammirati) Ammirati,Niskanen & Liimat. comb. nov.
MycoBank MB800336Basionym: Dermocybe marylandensis Ammirati, Beih. Sydowia
10:144 (1995). Type: USA. FLORIDA: Alachua, Devil’sMill Hopper, mixed hardwoods and Pinus, 26 Aug 1977,D. Malloch, W. Sundberg & J.F. Ammirati 7936 (WTU,HOLOTYPE). GenBank No. U56048, JX045690.
Cortinarius marylandensis Gerw. Keller & Ammirati,Mycotaxon 18:364 (1983). (Nom. inval., Art. 36.1, publishedas ‘‘nom. prov.’’).
Cortinarius marylandensis Ammirati & A.H. Sm., McIl-vainea 6:58 (1984). (Nom. inval., Art. 36.1, published as‘‘nom. prov.’’).
Cortinarius marylandensis Ammirati & A.H. Sm., Fieldguide to southern mushrooms: 213 (1985). (Nom. inval., Art.36.1, published as ‘‘nom. prov.’’).
Note: Recent molecular phylogenetic studies indi-cate that Dermocybe is a monophyletic group includedin the genus Cortinarius (Liu et al. 1997, Peintneret al. 2004, Garnica et al. 2005). Therefore, we herecombine Cortinarius marylandensis in genus Cortinar-ius. The species previously was known only from the
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 353

eastern United States (Liu et al. 1995) but here is alsoreported from Costa Rica. Due to the two intrage-nomic length polymorphism sites in the ITS region,we were not able to get a good quality 5.8S regionsequence from this specimen and therefore it wasexcluded from the analysis. Based on ITS1 and ITS2regions, however, the specimen is identified as C.marylandensis. For morphological species descriptionof C. marylandensis the reader is referred to Ammiratiand Smith (1984) and Liu et al. (1995).
Specimens examined: COSTA RICA. Savegre Sedero LosRobles, 11 Jun 1996, under Quercus copeyensis and Quercussemannii, JFA12010 (WTU). USA. DELAWARE: New CastleCounty, Alaposa Park, mixed hardwoods woods, 18 Jul 1975(WTU). FLORIDA: Alachua County, Devil’s Mill Hopper,mixed hardwoods and Pinus, 26 Aug 1977, D. Malloch, W.Sundberg & JFA 7936 (WTU, HOLOTYPE); University ofFlorida Horticulture Station, mixed hardwoods and Pinus,26 Aug 1977, JFA7935 (WTU). TEXAS: Liberty County, BigCreek Scenic Area, 31 May 1989, coll. Susan Metzler(WTU). VIRGINIA: King George County, King George,mixed woods, 27 Aug 1973, coll. Joe Salvino, JFA6481,JFA6482 (WTU).
6. Cortinarius smithii Ammirati, Niskanen & Liimat.stat. nov. & nom. nov.
MycoBank MB800337Basionym Cortinarius phoeniceus var. occidentalis A.H. Sm.,
Contr. Univ. Mich. Herb. 2:30 (1939). Type: USA.OREGON: Florence, mixed conifers (Picea, Pinus,Pseudotsuga), 14 Nov 1935, A.H. Smith 3485 (MICH,HOLOTYPE). GenBank No. JX045689.
Etymology: The species is named in honor of AlexanderH. Smith who first described this species as a variety of C.phoeniceus.
Dermocybe phoenicea var. occidentalis (A.H. Sm.) Ammir-ati, Mycotaxon 34:26 (1989). (Nom. inval., Art. 43.1).
Note: Cortinarius smithii formed a well supportedclade in our phylogenetic analysis. The analysis alsoshows that C. smithii is not related to C. phoeniceus butto C. fervidus (PP 0.73) from which is differs by sixevolutionary events in the ITS regions. The differenceis about the same as in the species pair C. sanguineus/C. puniceus in the same subgenus. Morphologically C.smithii and C. fervidus differ in coloration of thebasidiomata. Cortinarius smithii has rich red (GarnetBrown to Ox-Blood Red) pileus and purplish red tobright red (Bordeaux) lamellae, while C. fervidustypically has red-brown pileus and rusty red lamellae.The spores of the two species are similar (6.5–7.5 3
4–4.7 mm). Cortinarius smithii occurs in the PacificNorthwest of North America, while C. fervidus is so faronly known from Europe but may also occur ineastern North America. Based on morphology andmolecular data we conclude that C. smithii should betreated as a species. Description of the species is
presented for example in Smith (1939) and Ammiratiand Smith (1984).
Specimens examined: USA. OREGON: Florence, mixedconifers (Picea, Pinus, Pseudotsuga), 14 Nov 1935, A.H.Smith 3485 (MICH, HOLOTYPE). WASHINGTON: SkagitCounty, Easy Pass Trailhead, mixed conifers (Pinus, Picea,Abies, Pseudotsuga, Tsuga), 13 Oct 1988, JFA9886 (WTU).
Additional specimens: CANADA. BRITISH COLUMBIA:Manning Provincial Park, conifer forest, 6 Oct 2007, B.Didier, J. Holkko, E. Harrower, J. Pendray VMS7, F17110(UBC)
KEY TO RED SPECIES OF SUBGENUS DERMOCYBE IN
NORTH AMERICA
1. In hardwood or mixed forests . . . . . . . . . . . . . . . 219. In conifer forests . . . . . . . . . . . . . . . . . . . . . . . . 3
2. Basal mycelium yellow . . . . . . . . . C. harrisonii29. Basal mycelium pale vinaceous pink to pinkish
buff . . . . . . . . . . . . . . . . . . . C. marylandensis3. Pileus deep red-orange to brownish red-orange;
exsiccata reddish brown to yellowish red; inmountains of eastern United States or SierraNevada . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
39. Pileus rich red, vivid red to dark purplish red;exsiccata dusky red, reddish brown to dark reddishbrown; from California into British Columbia or inhemiboreal and boreal coniferous forests . . . . . . 54. Spores , 5 mm wide; in mountains of eastern
United States . . . . . . . . . . . . . . . C. harrisonii49. Spores . 5 mm wide; in Sierra Nevada. . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . C. sierraensis5. From California into the Pacific Northwest of
North America; vivid red to dark purplish, basalmycelium yellowish red to yellow. . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . C. neosanguineus59. At least in eastern North America, in hemiboreal
and boreal forests; pileus rich red to deep red,basal mycelium yellowish red . . . . . . C. sanguineus
DISCUSSION
Sanguinei.—The section includes at least five speciesfrom North America and Europe. Typical for thespecies are 6 red, fairly small basidiomata, basalmycelium often with yellow to reddish yellow tintsand indistinct universal veil. The spores are amygda-loid to ellipsoid, lamellar trama hyphae not encrust-ed and aniline red in KOH as are the pileipellishyphae. The pigments include the emodin series inaddition to the dermorubin and dermocybin series,but not studied from C. harrisonii (Keller andAmmirati 1983, Høiland 1984). The species areassociated with conifers or deciduous trees. To datethe section is known only from the northernhemisphere.
354 MYCOLOGIA

The best characteristics for identification of thespecies within the section are the color of thebasidiomata, also in the exsiccata, the structure ofthe pileus surface (smooth/scaly) and the size ofspores. Currently the known distributions of thespecies are not overlapping and they have differencesin their ecology as well.
Phylogenetic relationships.—In our analysis two largeinfrageneric groups were strongly supported, sect.Dermocybe and sect. Sanguinei correlating well withthe results by Liu et al. (1995) and Niskanen et al.(2012). Several smaller groups also were formed. Thenumber of studied Dermocybe species was hitherto thelargest indicating that when more species are sampledin the future a more natural classification will beachieved and previously single species in reality will berepresentatives of larger clades.
The evolutionary history of section Sanguinei wasnot fully resolved but our results indicate that mostlikely early in the evolution two lineages were formedgiving rise to western North American C. sierraensisand C. neosanguineus and eastern North American C.harrisonii. Later C. sanguineus evolved and mighthave spread with Picea to the Old World through aBeringian land bridge or directly from eastern NorthAmerica to Europe. In the Europe the last step ofevolution happened when C. puniceus arose. Also, inthe study by Garnica et al. (2011) North America wasproposed as the center of the origin for twoPhlegmacium species, C. arcuatorum Rob. Henry andC. elegantior (Fr.) Fr.
A new report of C. marylandensis from Costa Ricashows an interesting connection between the moun-tainous Quercus forests of Central America and thedeciduous forests of southern and eastern NorthAmerica. In addition, C. vitiosus (M.M. Moser)Niskanen, Kytov., Liimat. & S. Laine previously knownonly from northern Europe (Niskanen et al. 2012) ishere reported from Russia, Sakha Republic, Yakutia,eastern Siberia. The collection is from a Larix gmeliniiforest, although in Europe the species is assumed tobe associated with Picea. These examples show thatthere is still much more to be learned about this largemycorrhizal genus and the importance of continuingto sample material from as wide geographical area aspossible.
ACKNOWLEDGMENTS
We thank Andrus and Maria Voitk, Michael Burzynski,Renee Lebeuf and Andre Paul for support during fieldcollecting in Canada. We also thank Emma Harrower forproviding information on C. neosanguineus collections,Dimitar Bojantchev for providing material and photo of C.sierraensis and Esteri Ohenoja for providing material of C.
vitiosus. Curators of the herbaria of DAOM, EIU, G, H, IB,MICH, NYBG, S, TENN, TRTC, UBC and WTU are thankedfor kindly sending material. Financial support for this studywas received from the Emil Aaltonen Foundation, theAcademy of Finland (project 129052), Daniel E. StuntzMemorial Foundation and from NSF DEB 1144974 to R.Petersen and K. Hughes.
LITERATURE CITED
Ammirati JF. 1972. The section Dermocybe of Cortinarius inNorth America [doctoral disseration]. An Arbor: Univ.Michigan Press. 282 p.
———. 1989. Dermocybe, subgenus Dermocybe, section San-guineae in northern California. Mycotaxon 34:21–36.
———, Smith AH. 1984. Cortinarius II. A preliminarytreatment of species in the subgenus Dermocybe, sectionSanguinei, in North America north of Mexico. McIlvai-nea 6:54–64.
Edgar RC. 2004. MUSCLE: multiple sequence alignmentwith high accuracy and high throughput. Nucleic AcidsRes 32:1792–1797, doi:10.1093/nar/gkh340
Frøslev TG, Jeppesen TS, Læssoe T, Kjøller R. 2007.Molecular phylogenetics and delimitation of speciesin Cortinarius section Calochroi (Basidiomycota, Agar-icales) in Europe. Mol Phylogen Evol 44:217–227,doi:10.1016/j.ympev.2006.11.013
———, Matheny PB, Hibbett DS. 2005. Lower levelrelationships in the mushroom genus Cortinarius(Basidiomycota, Agaricales): a comparison of RPB1,RPB2, and ITS phylogenies. Mol Phylogen Evol 37:602–618, doi:10.1016/j.ympev.2005.06.016
Gardes M, Bruns TD. 1993. ITS primers with enhancedspecifity for basidiomycetes. Application to the identi-fication of mycorrhizae and rusts. Mol Ecol 2:113–118,doi:10.1111/j.1365-294X.1993.tb00005.x
Garnica S, Spahn P, Oertel B, Ammirati J, Oberwinkler F.2011. Tracking the evolutionary history of Cortinariusspecies in section Calochroi, with transoceanic disjunctdistributions. BMC Evol Biol 11:213, doi:10.1186/1471-2148-11-213
———, Weiß M, Oertel B, Ammirati J, Oberwinkler F. 2009.Phylogenetic relationships in Cortinarius, sectionCalochroi, inferred from nuclear DNA sequences.BMC Evol Biol 9:1, doi:10.1186/1471-2148-9-1
———, ———, ———, Oberwinkler F. 2005. A frameworkfor a phylogenetic classification in the genus Cortinar-ius (Basidiomycota, Agaricales) derived from morpho-logical and molecular data. Can J Bot 83:1457–1477,doi:10.1139/b05-107
Harrower E, Ammirati JF, Cappuccino AA, Ceska O,Kranabetter JM, Kroeger P, Lim S, Taylor T, BerbeeML. 2011. Cortinarius species diversity in BritishColumbia and molecular phylogenetic comparisonwith European specimen sequences. Botany 89:799–810, doi:10.1139/b11-065
Høiland K. 1984. Cortinarius subgenus Dermocybe. OperaBot 71:1–113 (1983).
NISKANEN ET AL.: CORTINARIUS SECTION SANGUINEI 355

Huelsenbeck JP, Ronquist F. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinfor-matics 19:1572–1574, doi:10.1093/bioinformatics/btg180
Keller G, Ammirati JF. 1983. Chemotaxonomic significanceof anthraquinone derivatives in North Americanspecies of Dermocybe, section Sanguineae. Mycotaxon18:357–377.
Liu Y, Rogers SO, Ammirati JF, Keller G. 1995. Dermocybe,section Sanguineae: a look at species relationships withinthe sanguineus complex. Beih. Sydowia 10:142–154.
———, ———, ———. 1997. Phylogenetic relationships inDermocybe and related Cortinarius taxa based onnuclear ribosomal DNA internal transcribed spacers.Can J Bot 75:519–532, doi:10.1139/b97-058
Moser M, Peintner U. 2002. The species complexCortinarius scaurus-Cortinarius herpeticus based onmorphological and molecular data. Micol Veg Medi-terr 17:3–17.
Munsell. 2009. Munsell soil color charts. Grand Rapids,Michigan: Munsell Color. 13 p.
Niskanen T, Kytovuori I, Liimatainen K. 2009. Cortinarius sect.Brunnei (Basidiomycota, Agaricales) in North Europe.Mycol Res 113:182–206, doi:10.1016/j.mycres.2008.10.006
———, ———, ———. 2011. Cortinarius sect. Armillati in NorthEurope. Mycologia 103:1080–1101, doi:10.3852/10-350
———, Laine S, Liimatainen K, Kytovuori I. 2012.Cortinarius sanguineus and equally red species in
Europe with an emphasis on northern Europeanmaterial. Mycologia 104:242–253, doi:10.3852/11-137
Ortega A, Suarez-Santiago VN, Reyes JD. 2008. Morpholog-ical and ITS identification of Cortinarius species(section Calochroi) collected in Mediterranean Quercuswoodlands. Fungal Divers 29:73–88.
Peintner U, Moncalvo J-M, Vilgalys R. 2004. Toward a betterunderstanding of the infrageneric relationships inCortinarius (Agaricales, Basidiomycota). Mycologia 96:1042–1058, doi:10.2307/3762088
Ridgway R. 1912. Color standards and color nomencla-ture. Washington DC: Published by the author. 43 p.,53 p.
Smith A. 1939. Studies in the genus Cortinarius I.Contributions from the University of Michigan Her-barium, vol 2. 1–42.
Suarez-Santiago VN, Ortega A, Peintner U, Lopez-Flores I.2009. Study on Cortinarius subgenus Telamonia sectionHydrocybe in Europe, with special emphasis on Medi-terranean taxa. Mycol Res 113:1070–1090, doi:10.1016/j.mycres.2009.07.006
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes forphylogenetics. In: Michael AJ, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols: a guide to the methodsand applications. New York: Academic Press. p 315–322.
356 MYCOLOGIA
Related Documents











