Psychopharmacology (1998) 136 : 247255 ' Springer-Verlag 1998 ORIGINAL INVESTIGATION Stephen C. Heinrichs • Alwin Klaassen George F. Koob • Gery Schulteis Serge Ahmed • Errol B. De Souza Corticotropin-releasing factor receptor blockade enhances conditioned aversive properties of cocaine in rats Received: 3 June 1997 / Final version: 28 August 1997 Abstract The behavioral proÞle of corticotropin-releas- ing factor (CRF) in mediating anxiogenic-like and aver- sive responses to stressors may be particularly relevant for dependence and withdrawal in drug-experienced organisms. Moreover, stressful aspects of drug expo- sure in the drug naive organism may also induce CRF system activation. In the present studies, the depen- dence of aversive properties of cocaine on activation of endogenous CRF systems has been evaluated in rats using taste conditioning and runway self-administra- tion paradigms. Systemic cocaine administration (20 mg/kg IP) produced a conditioned saccharin aver- sion which was dose-dependently potentiated by cen- tral administration of the CRF receptor antagonist, D-phe CRF (1241). In addition, IV cocaine adminis- tration (0.75 mg/kg per injection IV) produced runway goal-box avoidance and conditioned place avoidance responses which were signiÞcantly accelerated by CRF antagonist treatment. In contrast, CRF receptor stim- ulation using CRF itself abolished cocaine-induced increases in goal latency in the runway paradigm. This generalized involvement of CRF systems in cocaine- related motivational/associative states is consistent with the comprehensive role of CRF in mediating emo- tional responses to non-drug stressors. Key words Corticotropin-releasing factor • Cocaine • Aversion • Arousal • Learning • Runway • Rat Introduction Psychoactive drug use in both humans and laboratory animals is probably perpetuated by a variety of situa- tional and individual factors, including environmental stressors which interact with the reinforcing properties of the drug itself to determine the likelihood of recur- ring drug use, abuse or addiction (Koob 1996). Psychostimulants such as cocaine manifest potent pos- itive reinforcing actions, as evidenced by cocaine binge- ing during unlimited access self-administration (Markou and Koob 1991) and the high reinforcement eƒcacy of cocaine relative to other drugs of abuse (Deneau et al. 1969). These hedonic properties of cocaine are accompanied by other biological actions of the drug following systemic administration includ- ing conditioned avoidance responses (Goudie 1979), fear-like or aversive states (Fontana and Commissaris 1989; Yang et al. 1992) and mobilization of the behav- ioral and physiological stress axes (Rivier and Vale 1987; Antelman et al. 1992; Yang et al. 1992). For example, Ettenberg and colleagues have modeled the dual reinforcing and anxiogenic behavioral conse- quences of cocaine infusion in rats trained to self- administer the drug by traversing the length of a straight alley (Geist and Ettenberg 1990). Following repeated testing, the latency to enter the goal box where IV cocaine infusions were delivered gradually increased, suggesting a pro-conßict or anxiogenic response which was reversible by pretreatment with diazepam (Ettenberg and Geist 1991). Similarly, rats will run faster down a runway for an injection of mor- phine and once in the goal box eat less food, suggest- ing that morphine treatment unmasks both conditioned rewarding and aversive consequences (White et al. 1977). Furthermore, pre-exposure to environmental stressors enhances the reinforcement value of cocaine in the context of limited access cocaine self-adminis- tration as well as in situations of conditioned cocaine reinforcement (Ramsey and Van Ree 1993; Goeders S.C. Heinrichs (*) • A. Klaassen • G. Schulteis • E.B. De Souza Neurocrine Biosciences Inc., 3050 Science Park Road, San Diego, CA 92121, USA Fax: +1-619-658-7602, e-mail: [email protected] G.F. Koob • S. Ahmed The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92093, USA

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Psychopharmacology (1998) 136 : 247�255 © Springer-Verlag 1998
ORIGINAL INVESTIGATION
Stephen C. Heinrichs · Alwin KlaassenGeorge F. Koob · Gery SchulteisSerge Ahmed · Errol B. De Souza
Corticotropin-releasing factor receptor blockade enhances conditionedaversive properties of cocaine in rats
Received: 3 June 1997/Final version: 28 August 1997
Abstract The behavioral proÞle of corticotropin-releas-ing factor (CRF) in mediating anxiogenic-like and aver-sive responses to stressors may be particularly relevantfor dependence and withdrawal in drug-experiencedorganisms. Moreover, stressful aspects of drug expo-sure in the drug naive organism may also induce CRFsystem activation. In the present studies, the depen-dence of aversive properties of cocaine on activation ofendogenous CRF systems has been evaluated in ratsusing taste conditioning and runway self-administra-tion paradigms. Systemic cocaine administration(20 mg/kg IP) produced a conditioned saccharin aver-sion which was dose-dependently potentiated by cen-tral administration of the CRF receptor antagonist,D-phe CRF (12�41). In addition, IV cocaine adminis-tration (0.75 mg/kg per injection IV) produced runwaygoal-box avoidance and conditioned place avoidanceresponses which were signiÞcantly accelerated by CRFantagonist treatment. In contrast, CRF receptor stim-ulation using CRF itself abolished cocaine-inducedincreases in goal latency in the runway paradigm. Thisgeneralized involvement of CRF systems in cocaine-related motivational /associative states is consistentwith the comprehensive role of CRF in mediating emo-tional responses to non-drug stressors.
Key words Corticotropin-releasing factor ·Cocaine · Aversion · Arousal · Learning ·Runway · Rat
Introduction
Psychoactive drug use in both humans and laboratoryanimals is probably perpetuated by a variety of situa-tional and individual factors, including environmentalstressors which interact with the reinforcing propertiesof the drug itself to determine the likelihood of recur-ring drug use, abuse or addiction (Koob 1996).Psychostimulants such as cocaine manifest potent pos-itive reinforcing actions, as evidenced by cocaine binge-ing during unlimited access self-administration(Markou and Koob 1991) and the high reinforcemente¦cacy of cocaine relative to other drugs of abuse(Deneau et al. 1969). These hedonic properties ofcocaine are accompanied by other biological actionsof the drug following systemic administration includ-ing conditioned avoidance responses (Goudie 1979),fear-like or aversive states (Fontana and Commissaris1989; Yang et al. 1992) and mobilization of the behav-ioral and physiological stress axes (Rivier and Vale1987; Antelman et al. 1992; Yang et al. 1992). Forexample, Ettenberg and colleagues have modeled thedual reinforcing and anxiogenic behavioral conse-quences of cocaine infusion in rats trained to self-administer the drug by traversing the length of astraight alley (Geist and Ettenberg 1990). Followingrepeated testing, the latency to enter the goal box whereIV cocaine infusions were delivered graduallyincreased, suggesting a pro-conßict or anxiogenicresponse which was reversible by pretreatment withdiazepam (Ettenberg and Geist 1991). Similarly, ratswill run faster down a runway for an injection of mor-phine and once in the goal box eat less food, suggest-ing that morphine treatment unmasks both conditionedrewarding and aversive consequences (White et al.1977). Furthermore, pre-exposure to environmentalstressors enhances the reinforcement value of cocainein the context of limited access cocaine self-adminis-tration as well as in situations of conditioned cocainereinforcement (Ramsey and Van Ree 1993; Goeders
S.C. Heinrichs (*) · A. Klaassen · G. Schulteis · E.B. De SouzaNeurocrine Biosciences Inc., 3050 Science Park Road,San Diego, CA 92121, USAFax: +1-619-658-7602, e-mail: [email protected]
G.F. Koob · S. AhmedThe Scripps Research Institute, 10550 North Torrey Pines Road,La Jolla, CA 92093, USA

and Guerin 1994). Since the stressful behavioral andphysiological aspects of cocaine persist in the cocaine-experienced organism, and since anxiogenic-likeresponses are readily apparent following cocaine with-drawal (Antelman et al. 1992; Yang et al. 1992), it seemsreasonable to postulate that biological substratesresponsible for behavioral and physiological responsesto stress may play a role in cocaine seeking behavior(Goeders 1997).
Stressful aspects of drug exposure in the drug-naiveorganism may produce activation of brain corti-cotropin-releasing factor (CRF) systems (Antelmanet al. 1980; Sarnyai et al. 1993). For example,signiÞcant, regionally selective and dose-dependentincreases in hypothalamic CRF peptide content andCRF mRNA expression are observed following acuteor �binge�-pattern administration of cocaine in rats(Sarnyai et al. 1993; Zhou et al. 1996). This cocaine-induced CRF activation state appears to be critical formobilizing adaptive responses, such as adrenocorti-cotropin secretion, to stressor exposure (Rivier andVale 1987). A generalized involvement of CRF systemsin drug-related, and in particular drug withdrawal-related, negative motivational states is consistent withthe comprehensive role of CRF in mediating the coor-dinated response to environmental stressors (Baldwinet al. 1991; Koob et al. 1993; Rassnick et al. 1993;Heinrichs et al. 1995). However, the physiological roleof brain CRF in mediating biological responses to drugstimuli in drug-inexperienced animals has not beenextensively explored (Sarnyai et al. 1992). Accordingly,the main goal of the present studies was to examinethe impact of central administration of a peptide CRFreceptor antagonist on two measures of the conditionedaversive properties of cocaine: taste conditioning pro-duced by a single, acute injection of cocaine and acqui-sition of cocaine-induced avoidance responding in arepeated access runway paradigm.
Materials and methods
Subjects
Male Wistar rats (n = 202, Charles River) weighing 300�350 g weregroup housed and maintained in a temperature- and light-controlledenvironment on a reversed 12-h light :dark cycle (lights o¤: 0600hours; lights on: 1800 hours) with ad libitum access to rat chow(Teklad). Animals were allowed a 1-week period for acclimation tothe vivarium prior to experimentation. All experimental procedureswere approved by the Animal Care and Use Committee ofNeurocrine Biosciences, Inc.
Intracerebroventricular cannulation and infusion
In order to enable central administration of the CRF peptides, astainless steel guide cannula was implanted, aimed at the lateralventricle. Rats were anesthetized with isoßurane (2�5% in oxygen)and placed in a stereotaxic instrument with the incisor bar set 5 mm
above the interaural line. A 7 mm cannula (23 gauge) was securedto the skull using stainless steel screws and dental cement andoccluded with a 30 gauge obdurator. Implantation coordinates were0.6 mm posterior to bregma, ± 2.0 mm lateral, and 3.2 mm belowthe surface of the skull. After a 7-day surgical recovery period, can-nula patency was conÞrmed by gravity ßow of distilled waterthrough an 8 mm long, 30 gauge injector inserted within the guideto 1 mm beyond its tip. For treatment injections, 5 µl of solutionwas infused over 15 s and the injectors were left in place for 30 sto allow di¤usion of test substance.
Jugular catheterization
Chronic indwelling IV catheters were surgically implanted inisoßurane anesthetized rats (2�5% in oxygen). The left jugular veinwas incised and a 3.5 cm length of PE 10 tubing was inserted andsecured with the tip in proximity to the heart. The enlarged PE 50output end of the catheter was closed with a removable occluderand passed out of an incision in back of the neck so as to facilitatetethering and IV injection. Catheterized rats were single housed fora 2-day post-surgical recovery period, during which time catheterswere ßushed and Þlled with heparinized (1 :5 by volume) physio-logical saline. The injectable antibiotic Timentin (10 mg/animal /day; Smith Kline Beecham) was administered to all animals via theirIV catheter following IV infusions of either saline or cocaine, inorder to ward o¤ infection.
Indwelling IV catheters were checked for patency at the conclu-sion of the experiments in order to exclude animals with non-patentcatheters from data analysis. Brevital Sodium (0.2 ml of 1% metho-hexital sodium, Eli Lilly), a short-acting barbiturate that inducesanesthesia when delivered directly into the bloodstream, was admin-istered to all subjects via the IV catheter. If the animal lost con-sciousness within 5 s of administration, the catheter was judged tobe patent. Three animals with non-patent catheters were droppedaccording to this a priori criterion.
Drugs and peptide
In the taste conditioning studies, cocaine hydrochloride providedby the National Institute on Drug Abuse (NIDA) was dissolved in0.9% saline and administered IP in a volume of 1 ml/kg bodyweight. In the runway studies, cocaine was dissolved in 0.9% salineand administered IV via a jugular catheter in a Þxed volume of0.5 ml/ trial. Rat/human D-phe CRF (12�41) and ovine CRF (1�41)were synthesized by Dr. Nicholas Ling at Neurocrine Biosciences,Inc.. The peptides were solubilized immediately prior to infusion indistilled water adjusted to pHH 6.7 and solutions were kept on ice.
Statistics
Data were analyzed using one-, two- or three-way analysis of vari-ance (ANOVA) with drug or peptide treatment as between-subjects factors and experimental day as a within-subjects factor.In particular, the taste conditioning studies were analyzed usingone-way ANOVA with cocaine and CRF receptor antagonist doseas a between-subjects factor, the 1-week runway studies were ana-lyzed using two-way ANOVA with cocaine, CRF or CRF recep-tor antagonist treatment as a between-subjects factor andexperimental day as a within-subjects factor, and the extended run-way studies were analyzed using three-way ANOVA with cocaineand CRF receptor antagonist treatment as between subjects factorsand experimental day as a within-subjects factor. Post-hoc pairwisecomparisons were performed using Newman-Keuls tests followingidentiÞcation of signiÞcant omnibus e¤ects by ANOVA.
248
Experiment 1: taste conditioning
Apparatus and methods
Colony room water bottles were removed at 1100 hours to beginthe 6-day, single-pairing, two-bottle taste conditioning procedure(Heinrichs et al. 1991). The taste conditioning apparatus consistedof a standard rodent housing cage containing pre-measured aliquotsof water or saccharin solution, or both, in standard cage-top, sip-per-tube bottles. Water deprived (24 h) animals were Þrst placedindividually in the taste conditioning apparatus and allowed a 15-min access to drinking water (Fig. 1). Subjects were returnedsubsequently to their home cages and colony water bottles werereplaced for 24 h. On day 4, following a second 24-h water depri-vation, subjects were again placed individually in the taste condi-tioning apparatus and allowed a 15-min access to a 0.2% saccharinsolution. One group of rats received a single post-saccharin injec-tion of 0, 5, 10, 15, 20 or 40 mg/kg IP doses of cocaine, while aseparate group of rats received two post-saccharin injections of 0,0.1, 1, 10 or 150 µg ICV doses of D-phe CRF (12�41) immediatelyafter saccharin exposure, followed 5 min later by 0, 5 or 20 mg/kgIP doses of cocaine. Rats were returned to their home cagesequipped with water bottles for the following 24 h. On day 6 fol-lowing a third 24-h water deprivation, subjects were placed in thetaste conditioning apparatus and presented with a 15-min choicebetween two bottles, counterbalanced for position upon the cagetop, containing either distilled water or a 0.2% saccharin solution.The measure of taste conditioning was taken to be the di¤erencescore between saccharin solution intake on day 6 following thedrug/peptide pairing and day 4 prior to the pairing.
Results
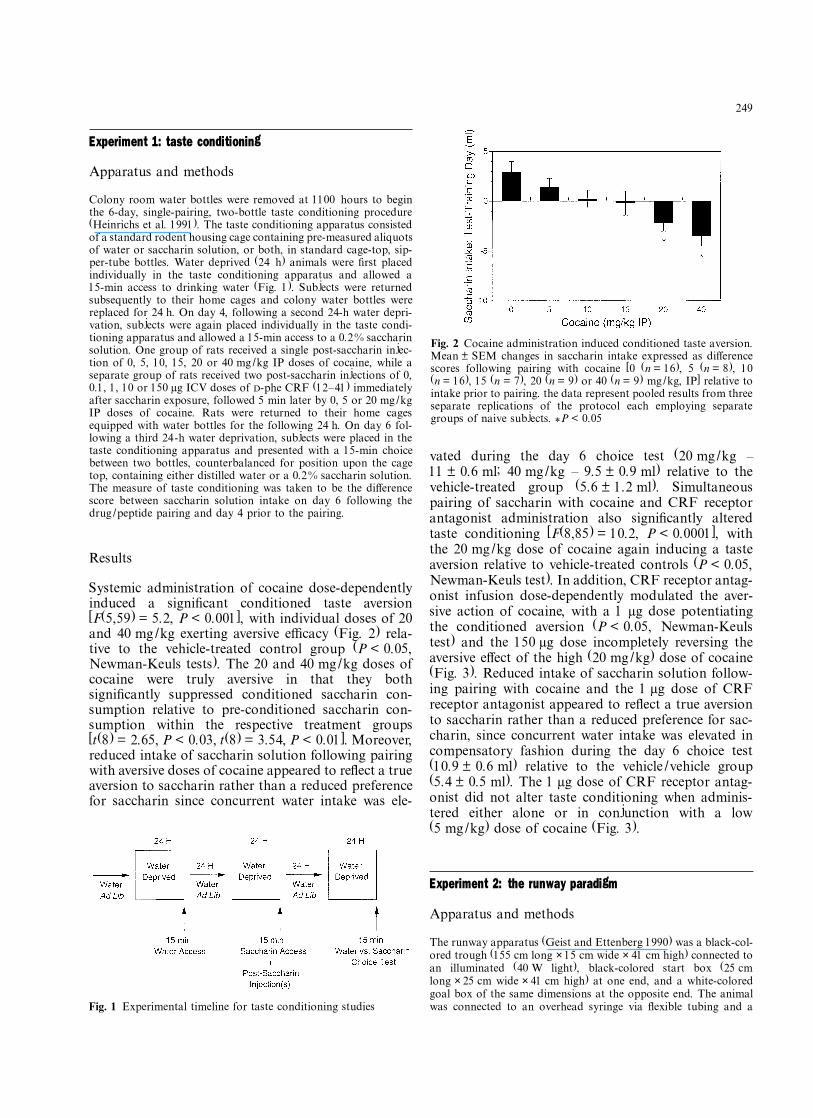
Systemic administration of cocaine dose-dependentlyinduced a signiÞcant conditioned taste aversion[F(5,59) = 5.2, P < 0.001], with individual doses of 20and 40 mg/kg exerting aversive e¦cacy (Fig. 2) rela-tive to the vehicle-treated control group (P < 0.05,Newman-Keuls tests). The 20 and 40 mg/kg doses ofcocaine were truly aversive in that they bothsigniÞcantly suppressed conditioned saccharin con-sumption relative to pre-conditioned saccharin con-sumption within the respective treatment groups[t(8) = 2.65, P < 0.03, t(8) = 3.54, P < 0.01]. Moreover,reduced intake of saccharin solution following pairingwith aversive doses of cocaine appeared to reßect a trueaversion to saccharin rather than a reduced preferencefor saccharin since concurrent water intake was ele-
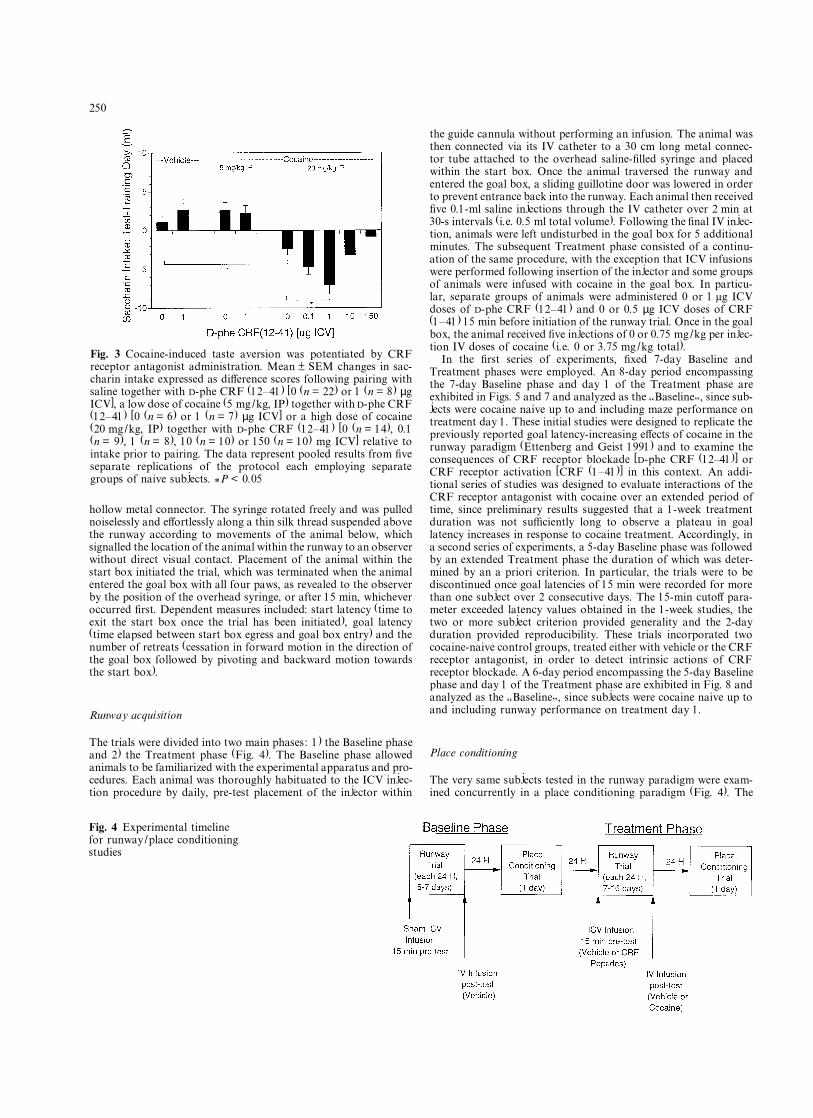
vated during the day 6 choice test (20 mg/kg �11 ± 0.6 ml; 40 mg/kg � 9.5 ± 0.9 ml) relative to thevehicle-treated group (5.6 ± 1.2 ml). Simultaneouspairing of saccharin with cocaine and CRF receptorantagonist administration also signiÞcantly alteredtaste conditioning [F(8,85) = 10.2, P < 0.0001], withthe 20 mg/kg dose of cocaine again inducing a tasteaversion relative to vehicle-treated controls (P < 0.05,Newman-Keuls test). In addition, CRF receptor antag-onist infusion dose-dependently modulated the aver-sive action of cocaine, with a 1 µg dose potentiatingthe conditioned aversion (P < 0.05, Newman-Keulstest) and the 150 µg dose incompletely reversing theaversive e¤ect of the high (20 mg/kg) dose of cocaine(Fig. 3). Reduced intake of saccharin solution follow-ing pairing with cocaine and the 1 µg dose of CRFreceptor antagonist appeared to reßect a true aversionto saccharin rather than a reduced preference for sac-charin, since concurrent water intake was elevated incompensatory fashion during the day 6 choice test(10.9 ± 0.6 ml) relative to the vehicle /vehicle group(5.4 ± 0.5 ml). The 1 µg dose of CRF receptor antag-onist did not alter taste conditioning when adminis-tered either alone or in conjunction with a low(5 mg/kg) dose of cocaine (Fig. 3).
Experiment 2: the runway paradigm
Apparatus and methods
The runway apparatus (Geist and Ettenberg 1990) was a black-col-ored trough (155 cm long × 15 cm wide × 41 cm high) connected toan illuminated (40 W light), black-colored start box (25 cmlong × 25 cm wide × 41 cm high) at one end, and a white-coloredgoal box of the same dimensions at the opposite end. The animalwas connected to an overhead syringe via ßexible tubing and a
249
Fig. 1 Experimental timeline for taste conditioning studies
Fig. 2 Cocaine administration induced conditioned taste aversion.Mean ± SEM changes in saccharin intake expressed as di¤erencescores following pairing with cocaine [0 (n = 16), 5 (n = 8), 10(n = 16), 15 (n = 7), 20 (n = 9) or 40 (n = 9) mg/kg, IP] relative tointake prior to pairing. the data represent pooled results from threeseparate replications of the protocol each employing separategroups of naive subjects. *P < 0.05
hollow metal connector. The syringe rotated freely and was pullednoiselessly and e¤ortlessly along a thin silk thread suspended abovethe runway according to movements of the animal below, whichsignalled the location of the animal within the runway to an observerwithout direct visual contact. Placement of the animal within thestart box initiated the trial, which was terminated when the animalentered the goal box with all four paws, as revealed to the observerby the position of the overhead syringe, or after 15 min, whicheveroccurred Þrst. Dependent measures included: start latency (time toexit the start box once the trial has been initiated), goal latency(time elapsed between start box egress and goal box entry) and thenumber of retreats (cessation in forward motion in the direction ofthe goal box followed by pivoting and backward motion towardsthe start box).
Runway acquisition
The trials were divided into two main phases : 1) the Baseline phaseand 2) the Treatment phase (Fig. 4). The Baseline phase allowedanimals to be familiarized with the experimental apparatus and pro-cedures. Each animal was thoroughly habituated to the ICV injec-tion procedure by daily, pre-test placement of the injector within
the guide cannula without performing an infusion. The animal wasthen connected via its IV catheter to a 30 cm long metal connec-tor tube attached to the overhead saline-Þlled syringe and placedwithin the start box. Once the animal traversed the runway andentered the goal box, a sliding guillotine door was lowered in orderto prevent entrance back into the runway. Each animal then receivedÞve 0.1-ml saline injections through the IV catheter over 2 min at30-s intervals (i.e. 0.5 ml total volume). Following the Þnal IV injec-tion, animals were left undisturbed in the goal box for 5 additionalminutes. The subsequent Treatment phase consisted of a continu-ation of the same procedure, with the exception that ICV infusionswere performed following insertion of the injector and some groupsof animals were infused with cocaine in the goal box. In particu-lar, separate groups of animals were administered 0 or 1 µg ICVdoses of D-phe CRF (12�41) and 0 or 0.5 µg ICV doses of CRF(1�41) 15 min before initiation of the runway trial. Once in the goalbox, the animal received Þve injections of 0 or 0.75 mg/kg per injec-tion IV doses of cocaine (i.e. 0 or 3.75 mg/kg total).
In the Þrst series of experiments, Þxed 7-day Baseline andTreatment phases were employed. An 8-day period encompassingthe 7-day Baseline phase and day 1 of the Treatment phase areexhibited in Figs. 5 and 7 and analyzed as the �Baseline�, since sub-jects were cocaine naive up to and including maze performance ontreatment day 1. These initial studies were designed to replicate thepreviously reported goal latency-increasing e¤ects of cocaine in therunway paradigm (Ettenberg and Geist 1991) and to examine theconsequences of CRF receptor blockade [D-phe CRF (12�41)] orCRF receptor activation [CRF (1�41)] in this context. An addi-tional series of studies was designed to evaluate interactions of theCRF receptor antagonist with cocaine over an extended period oftime, since preliminary results suggested that a 1-week treatmentduration was not su¦ciently long to observe a plateau in goallatency increases in response to cocaine treatment. Accordingly, ina second series of experiments, a 5-day Baseline phase was followedby an extended Treatment phase the duration of which was deter-mined by an a priori criterion. In particular, the trials were to bediscontinued once goal latencies of 15 min were recorded for morethan one subject over 2 consecutive days. The 15-min cuto¤ para-meter exceeded latency values obtained in the 1-week studies, thetwo or more subject criterion provided generality and the 2-dayduration provided reproducibility. These trials incorporated twococaine-naive control groups, treated either with vehicle or the CRFreceptor antagonist, in order to detect intrinsic actions of CRFreceptor blockade. A 6-day period encompassing the 5-day Baselinephase and day 1 of the Treatment phase are exhibited in Fig. 8 andanalyzed as the �Baseline�, since subjects were cocaine naive up toand including runway performance on treatment day 1.
Place conditioning
The very same subjects tested in the runway paradigm were exam-ined concurrently in a place conditioning paradigm (Fig. 4). The
250
Fig. 3 Cocaine-induced taste aversion was potentiated by CRFreceptor antagonist administration. Mean ± SEM changes in sac-charin intake expressed as di¤erence scores following pairing withsaline together with D-phe CRF (12�41) [0 (n = 22) or 1 (n = 8) µgICV], a low dose of cocaine (5 mg/kg, IP) together with D-phe CRF(12�41) [0 (n = 6) or 1 (n = 7) µg ICV] or a high dose of cocaine(20 mg/kg, IP) together with D-phe CRF (12�41) [0 (n = 14), 0.1(n = 9), 1 (n = 8), 10 (n = 10) or 150 (n = 10) mg ICV] relative tointake prior to pairing. The data represent pooled results from Þveseparate replications of the protocol each employing separategroups of naive subjects. *P < 0.05
Fig. 4 Experimental timelinefor runway/place conditioningstudies
apparatus consisted of the start and goal boxes employed in therunway studies. The boxes were joined with the openings facingeach other, creating an enclosed environment wherein the animalcould move freely between the compartments. A white light illu-minated the black-colored start box while the white-colored goalbox remained darkened so that testing conditions were consistentwith the runway trial environment. A mirror suspended above theboxes allowed the experimenter to observe the animal�s movementswithout direct visual contact. The test was initiated by placing theanimal in the start box facing the back wall, and the time spent inthe goal box was recorded over 20 min. In both sets of studies, theplace conditioning test was performed 1 day following the end ofthe Baseline phase to determine baseline place preference, and again1 day following the end of the Treatment phase, in order to eval-uate conditioned place preference. Time spent within the goal-boxhalf of the environment following the Treatment phase relative totime spent within the goal-box prior to initiation of experimentaltreatments was taken as the measure of place conditioning. It isnoteworthy that in contrast to the concurrent runway/place con-ditioning design of the present studies, the original study of cocaineactions in the runway paradigm employed separate apparatus andprocedures for determination of cocaine-induced place condition-ing (Ettenberg and Geist 1991). In particular, changes in time spentwithin the drug/peptide-paired goal-box served as a measure ofplace conditioning in the present studies without regard for timespent in the alternative start-box, since subjects were neither admin-istered vehicle nor conÞned within the unpaired start-box, as is stan-dard practice in most stand-alone place conditioning studies(Swerdlow et al. 1989).
Results
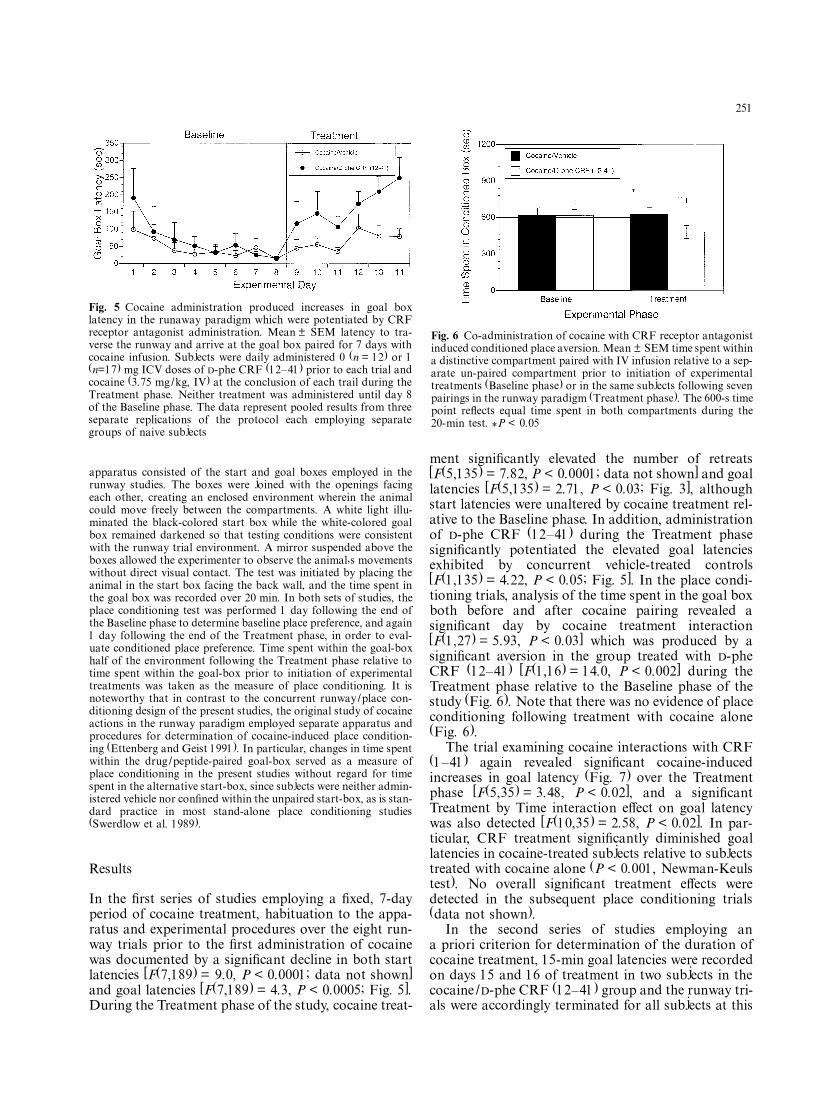
In the Þrst series of studies employing a Þxed, 7-dayperiod of cocaine treatment, habituation to the appa-ratus and experimental procedures over the eight run-way trials prior to the Þrst administration of cocainewas documented by a signiÞcant decline in both startlatencies [F(7,189) = 9.0, P < 0.0001; data not shown]and goal latencies [F(7,189) = 4.3, P < 0.0005; Fig. 5].During the Treatment phase of the study, cocaine treat-
ment signiÞcantly elevated the number of retreats[F(5,135) = 7.82, P < 0.0001; data not shown] and goallatencies [F(5,135) = 2.71, P < 0.03; Fig. 3], althoughstart latencies were unaltered by cocaine treatment rel-ative to the Baseline phase. In addition, administrationof D-phe CRF (12�41) during the Treatment phasesigniÞcantly potentiated the elevated goal latenciesexhibited by concurrent vehicle-treated controls[F(1,135) = 4.22, P < 0.05; Fig. 5]. In the place condi-tioning trials, analysis of the time spent in the goal boxboth before and after cocaine pairing revealed asigniÞcant day by cocaine treatment interaction[F(1,27) = 5.93, P < 0.03] which was produced by asigniÞcant aversion in the group treated with D-pheCRF (12�41) [F(1,16) = 14.0, P < 0.002] during theTreatment phase relative to the Baseline phase of thestudy (Fig. 6). Note that there was no evidence of placeconditioning following treatment with cocaine alone(Fig. 6).
The trial examining cocaine interactions with CRF(1�41) again revealed signiÞcant cocaine-inducedincreases in goal latency (Fig. 7) over the Treatmentphase [F(5,35) = 3.48, P < 0.02], and a signiÞcantTreatment by Time interaction e¤ect on goal latencywas also detected [F(10,35) = 2.58, P < 0.02]. In par-ticular, CRF treatment signiÞcantly diminished goallatencies in cocaine-treated subjects relative to subjectstreated with cocaine alone (P < 0.001, Newman-Keulstest). No overall signiÞcant treatment e¤ects weredetected in the subsequent place conditioning trials(data not shown).
In the second series of studies employing ana priori criterion for determination of the duration ofcocaine treatment, 15-min goal latencies were recordedon days 15 and 16 of treatment in two subjects in thecocaine/D-phe CRF (12�41) group and the runway tri-als were accordingly terminated for all subjects at this
251
Fig. 5 Cocaine administration produced increases in goal boxlatency in the runaway paradigm which were potentiated by CRFreceptor antagonist administration. Mean ± SEM latency to tra-verse the runway and arrive at the goal box paired for 7 days withcocaine infusion. Subjects were daily administered 0 (n = 12) or 1(n=17) mg ICV doses of D-phe CRF (12�41) prior to each trial andcocaine (3.75 mg/kg, IV) at the conclusion of each trail during theTreatment phase. Neither treatment was administered until day 8of the Baseline phase. The data represent pooled results from threeseparate replications of the protocol each employing separategroups of naive subjects
Fig. 6 Co-administration of cocaine with CRF receptor antagonistinduced conditioned place aversion. Mean ± SEM time spent withina distinctive compartment paired with IV infusion relative to a sep-arate un-paired compartment prior to initiation of experimentaltreatments (Baseline phase) or in the same subjects following sevenpairings in the runway paradigm (Treatment phase). The 600-s timepoint reßects equal time spent in both compartments during the20-min test. *P < 0.05
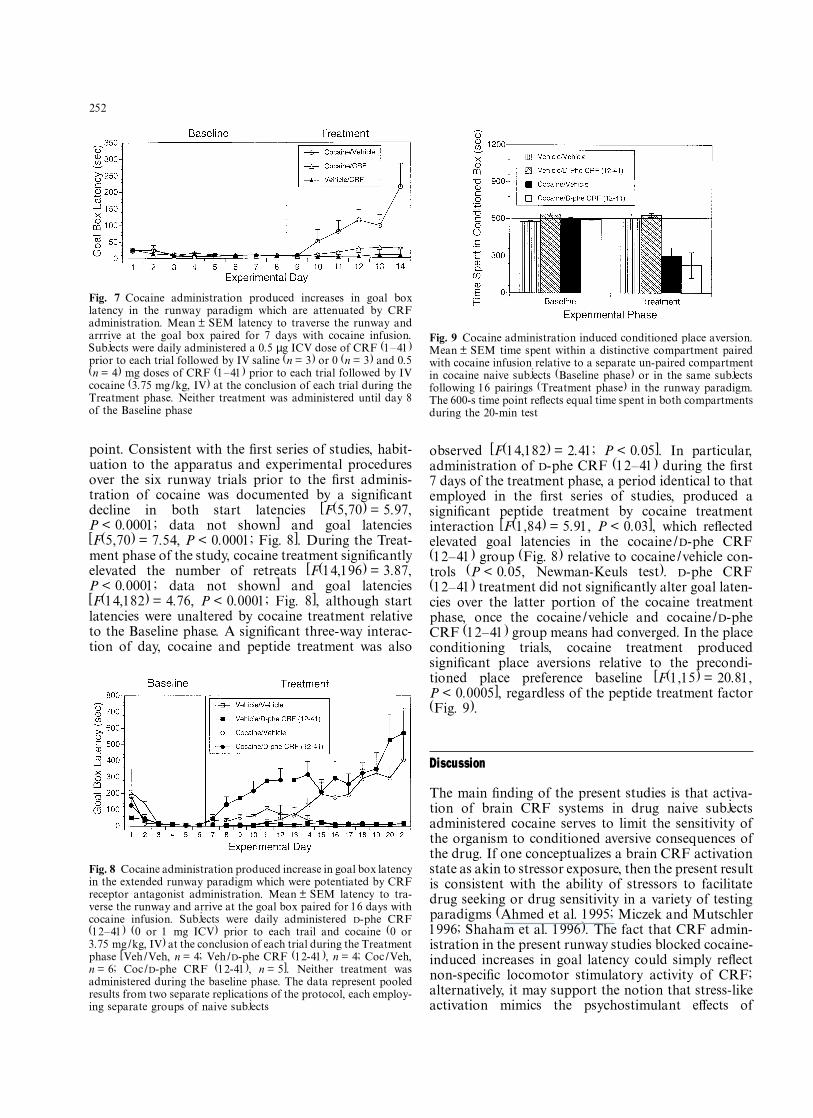
point. Consistent with the Þrst series of studies, habit-uation to the apparatus and experimental proceduresover the six runway trials prior to the Þrst adminis-tration of cocaine was documented by a signiÞcantdecline in both start latencies [F(5,70) = 5.97,P < 0.0001; data not shown] and goal latencies[F(5,70) = 7.54, P < 0.0001; Fig. 8]. During the Treat-ment phase of the study, cocaine treatment signiÞcantlyelevated the number of retreats [F(14,196) = 3.87,P < 0.0001; data not shown] and goal latencies[F(14,182) = 4.76, P < 0.0001; Fig. 8], although startlatencies were unaltered by cocaine treatment relativeto the Baseline phase. A signiÞcant three-way interac-tion of day, cocaine and peptide treatment was also
observed [F(14,182) = 2.41; P < 0.05]. In particular,administration of D-phe CRF (12�41) during the Þrst7 days of the treatment phase, a period identical to thatemployed in the Þrst series of studies, produced asigniÞcant peptide treatment by cocaine treatmentinteraction [F(1,84) = 5.91, P < 0.03], which reßectedelevated goal latencies in the cocaine/ D-phe CRF(12�41) group (Fig. 8) relative to cocaine/vehicle con-trols (P < 0.05, Newman-Keuls test). D-phe CRF(12�41) treatment did not signiÞcantly alter goal laten-cies over the latter portion of the cocaine treatmentphase, once the cocaine/vehicle and cocaine/D-pheCRF (12�41) group means had converged. In the placeconditioning trials, cocaine treatment producedsigniÞcant place aversions relative to the precondi-tioned place preference baseline [F(1,15) = 20.81,P < 0.0005], regardless of the peptide treatment factor(Fig. 9).
Discussion
The main Þnding of the present studies is that activa-tion of brain CRF systems in drug naive subjectsadministered cocaine serves to limit the sensitivity ofthe organism to conditioned aversive consequences ofthe drug. If one conceptualizes a brain CRF activationstate as akin to stressor exposure, then the present resultis consistent with the ability of stressors to facilitatedrug seeking or drug sensitivity in a variety of testingparadigms (Ahmed et al. 1995; Miczek and Mutschler1996; Shaham et al. 1996). The fact that CRF admin-istration in the present runway studies blocked cocaine-induced increases in goal latency could simply reßectnon-speciÞc locomotor stimulatory activity of CRF;alternatively, it may support the notion that stress-likeactivation mimics the psychostimulant e¤ects of
252
Fig. 7 Cocaine administration produced increases in goal boxlatency in the runway paradigm which are attenuated by CRFadministration. Mean ± SEM latency to traverse the runway andarrrive at the goal box paired for 7 days with cocaine infusion.Subjects were daily administered a 0.5 µg ICV dose of CRF (1�41)prior to each trial followed by IV saline (n = 3) or 0 (n = 3) and 0.5(n = 4) mg doses of CRF (1�41) prior to each trial followed by IVcocaine (3.75 mg/kg, IV) at the conclusion of each trial during theTreatment phase. Neither treatment was administered until day 8of the Baseline phase
Fig. 8 Cocaine administration produced increase in goal box latencyin the extended runway paradigm which were potentiated by CRFreceptor antagonist administration. Mean ± SEM latency to tra-verse the runway and arrive at the goal box paired for 16 days withcocaine infusion. Subjects were daily administered D-phe CRF(12�41) (0 or 1 mg ICV) prior to each trail and cocaine (0 or3.75 mg/kg, IV) at the conclusion of each trial during the Treatmentphase [Veh/Veh, n = 4; Veh/D-phe CRF (12-41), n = 4; Coc/Veh,n = 6; Coc/D-phe CRF (12-41), n = 5]. Neither treatment wasadministered during the baseline phase. The data represent pooledresults from two separate replications of the protocol, each employ-ing separate groups of naive subjects
Fig. 9 Cocaine administration induced conditioned place aversion.Mean ± SEM time spent within a distinctive compartment pairedwith cocaine infusion relative to a separate un-paired compartmentin cocaine naive subjects (Baseline phase) or in the same subjectsfollowing 16 pairings (Treatment phase) in the runway paradigm.The 600-s time point reßects equal time spent in both compartmentsduring the 20-min test

cocaine (Ahmed and Koob 1997), thereby limiting theconditioned aversive properties of cocaine. This latterhypothesis is supported by a recent report that CRFadministration facilitates, and CRF receptor antago-nist administration impairs, reinstatement of extin-guished responding for heroin (Shaham et al. 1997).
The present experiments were designed speciÞcallyto examine the interaction of CRF systems with aver-sive doses of cocaine in testing contexts selected forsensitivity to conditioned aversion/avoidance. Thus, asigniÞcantly aversive dose of cocaine (20 mg/kg IP) wasselected from the cocaine dose-response taste condi-tioning study (Fig. 2) in order to evaluate actions ofthe CRF receptor antagonist on cocaine-induced tasteaversion. Similarly, a 1 µg dose of D-phe CRF (12�41)was selected from the CRF receptor antagonist dose-response taste conditioning study (Fig. 3) as the onlydose of CRF receptor antagonist which signiÞcantlymodiÞed the aversive properties of cocaine.Accordingly, this single e¦cacious dose of CRF recep-tor antagonist was carried forward into another modelsensitive to cocaine-conditioned avoidance responses,the runway paradigm, again employing a single doseof cocaine (3.75 mg/kg IV) with documented aversiveproperties in this model (Ettenberg and Geist 1991).Thus, the a priori experimental aim of studying the roleof CRF in cocaine-induced learned aversion wasachieved by a rational selection of dosage and testingparadigm. The observed e¦cacy of CRF receptorblockade in consistently facilitating learned aversion tococaine in two separate taste and place conditioningparadigms thought to reßect the aversive properties ofpsychostimulant drugs (Turenne et al. 1996) begs thequestion of potential consequences of CRF receptorblockade in testing contexts sensitive to the positivelyreinforcing properties of psychostimulants.
A variety of possible alternative explanations for thepostulated aversion potentiating actions of D-phe CRF(12�41) related to intrinsic or unrelated e¤ects of theCRF receptor antagonist are not supported by the pre-sent data. In particular, CRF receptor antagonistadministration did not alter the goal latency or placeconditioning measures in the absence of cocaine treat-ment. Similarly, the runway paradigm embodies sev-eral similarities to a classical conßict situation involvingapproach towards a goal box and simultaneous avoid-ance of the goal box, as evidenced by the appearanceof retreat behavior which is reversible by benzodi-azepine administration (Ettenberg and Geist 1991).However, the anxiolytic-like and anti-stress proÞle ofCRF receptor antagonists, as revealed in a variety ofanimal models of anxiety employing spontaneousexploration, noxious stimuli and fear-conditioning,does not transfer readily, if at all, to the classical conßictparadigm (Dunn and Berridge 1990). Thus, the lack ofe¤ect of CRF receptor antagonists on conßict behav-iors in general, and on runway retreat behavior in particular, argues against the exertion of pro- or
anti-conßict actions of D-phe CRF (12�41) in the present cocaine/CRF receptor antagonist interactionstudies.
Another non-speciÞc mechanism by which reactiv-ity to cocaine in the runway paradigm could be alteredwould be sedative or response-inhibitory properties ofCRF receptor antagonist treatment. However, treat-ment with D-phe CRF (12�41) altered neither latencyto exit the start box nor the frequency of retreat behav-iors while transiting the runway, which together helpto reveal the relative degree of positive and negativeattributes of the cocaine-conditioned goal box (Geistand Ettenberg 1997). Moreover, the peptide exerted neither changes in these indices nor alterations in thegoal latency measure in cocaine-naive subjects testedin the runway over 21 days (Fig. 8). In addition,in the extended treatment series of runway studies(Fig. 8), CRF receptor antagonist treatment did notsigniÞcantly alter any of the runway performance mea-sures once vehicle-treated controls had achievedincreased goal latencies. Finally, central administrationof D-phe CRF (12�41) in e¤ective anti-stress doses doesnot alter cocaine responding in animals well trainedchronically to self-administer IV cocaine (0.25 mg/injection) on a limited access schedule (Ahmedet al. 1996). Taken together, these observations argueagainst sedative consequences of CRF receptor block-ade and suggest that CRF receptor antagonists aremore e¤ective in behavioral situations requiring achange in behavior, as opposed to a highly trainedresponse which has already been learned or a habitualresponse which does not mobilize the aversive /anxio-genic-like proÞle of CRF activation.
Place conditioning tests examining the relative pref-erence for cocaine-paired and unpaired compartmentsperformed immediately prior to and following cocaineexposure in the runway paradigm (Figs 6 and 9) pro-vide further evidence for conditioned aversive proper-ties of cocaine administration which were potentiatedby central administration of CRF receptor antagonist.In particular, the combination of cocaine and D-pheCRF (12�41) treatment in the Þrst series of runwaystudies produced a coordinated increase in runway goallatencies in combination with a conditioned place aver-sion (Figs. 5 and 6). The more robust and prolongedincrease in runway goal latencies recorded for bothcocaine-treated groups in the extended runway trialsresulted in robust place aversions which also did notdistinguish the two groups based on the peptide treat-ment factor (Figs 8 and 9). Thus, the place condition-ing results appear to validate the detection of aversiveproperties of cocaine using the runway goal latencymeasure and provide further support for the notionthat CRF receptor antagonist treatment acceleratesand/or enhances the conditioned aversive properties ofcocaine. This view is supported by the fact thattwo of Þve cocaine/D-phe CRF (12�41)-treated, butnone of six cocaine/vehicle-treated, subjects met the
253

a priori criterion of su¦ciently long goal latencies toterminate the extended runway trials. This robust avoid-ance of the goal box environment may be indicative ofan extinction event in which the negative conditionede¤ects of cocaine came to outweigh positive condi-tioned e¤ects of the drug.
Given the role of stress and stress-relief in drugintake, the characteristic proÞle of CRF receptor antag-onists may be of interest (Sarnyai et al. 1992; Cadoret al. 1993). The CRF receptor antagonists are partic-ularly e¤ective in reversing the behavioral changes pro-duced by drug withdrawal stressors without anymarked behavioral actions in the unperturbed animal(Baldwin et al. 1991). Increases in brain CRF (e.g. fol-lowing stressor exposure) seem to be an important pre-requisite for the anxiolytic-like e¤ectiveness of CRFreceptor antagonists (Koob et al. 1993). This anti-stressproÞle distinguishes the CRF receptor antagonists fromanxiolytics of the benzodiazepine class which act moreindependently of the arousal state of the animal(Goeders et al. 1993). The present results, together withpublished reports (Rodrîguez de Fonseca et al. 1997),suggest that brain CRF activation may accompanyboth initial exposure to a drug and withdrawal of thedrug following a period of chronic administration. Inparticular, brain CRF system activation, by its arousal-enhancing actions, appears both to limit aversiveactions of cocaine when drug naive and to promotethose same aversive actions during drug withdrawalfollowing chronic drug administration.
Acknowledgements We thank Michael Arends for help with man-uscript preparation. This research was supported by NIDA grantsDA09503 to S.C.H. and E.B.D.S., DA04398 and DA08467 toG.F.K. and S.A. and NIDDK grant DK26741 to G.F.K.
References
Ahmed SH, Koob GF (1997) Cocaine- but not food-seeking behav-ior is reinstated by stress after extinction. Psychopharmacology132:289�295
Ahmed SH, Stinus L, Le Moal M, Cador M (1995) Social depri-vation enhances the vulnerability of male Wistar rats to stres-sor- and amphetamine-induced behavioral sensitization.Psychopharmacology 117:116�124
Ahmed SH, Epping-Jordan MP, Gri¦n PS, Markou A, HeinrichsSC, De Souza EB, Rivier J, Koob GF (1996) Maintenance ofcocaine self-administration is not a¤ected by pharmacologicalblockade of brain CRF receptors. Soc Neurosci Abstr22:923
Antelman SM, Eichler AJ, Black CA, Kocan D (1980)Interchangeability between stress and amphetamine in sensiti-zation. Science 207[329]:329�331
Antelman SM, Caggiula AR, Knopf S, Kocan DJ, Edwards DJ(1992) Amphetamine or haloperidol 2 weeks earlier antagonizedthe plasma corticosterone response to amphetamine; evidencefor the stressful / foreign nature of drugs. Psychopharmacology107:331�336
Baldwin HA, Rassnick S, Rivier J, Koob GF, Britton KT (1991)CRF antagonist reverses the �anxiogenic� response to ethanolwithdrawal in the rat. Psychopharmacology 103:227�232
Cador M, Cole BJ, Koob GF, Stinus L, Le Moal M (1993) Centraladministration of corticotropin releasing factor induces long-term sensitization to d-amphetamine. Brain Res 606:181�186
Deneau GA, Yanagita T, Seevers MH (1969) Self-administrationof psychoactive substances by the monkey. Psychopharmaco-logia 16:30�48
Dunn AJ, Berridge CW (1990) Physiological and behavioralresponses to corticotropin-releasing factor administration: isCRF a mediator of anxiety of stress responses? Brain Res Rev15:71�100
Ettenberg A, Geist TD (1991) Animal model for investigating theanxiogenic e¤ects of self-administered cocaine. Psychopharma-cology 103:455�461
Fontana DJ, Commissaris RL (1989) E¤ects of cocaine on conßictbehavior in the rat. Life Sci 45:819�827
Geist TD, Ettenberg A (1990) A simple method for studying intra-venous drug reinforcement in a runway. Pharmacol BiochemBehav 36:703�706
Geist TD, Ettenberg A (1997) Concurrent positive and negativegoalbox events produce runway behaviors comparable to thoseof cocaine-reinforced rats. Pharmacol Biochem Behav57:145�150
Goeders NE (1997) A neuroendocrine role in cocaine reinforce-ment. Psychoneuroendocrinology 4 :237�259
Goeders NE, Guerin GF (1994) Non-contingent electric footshockfacilitates the acquisition of intravenous cocaine self-adminis-tration in rats. Psychopharmacology 114:63�70
Goeders NE, McNulty MA, Guerin GF (1993) E¤ects of alprazo-lam on intravenous cocaine self-administration in rats. Pharma-col Biochem Behav 44:471�474
Goudie AJ (1979) Aversive stimulus properties of drugs. Neuro-pharmacology 18:971�979
Heinrichs SC, Britton KT, Koob GF (1991) Both conditioned tastepreference and aversion induced by corticotropin-releasing fac-tor. Pharmacol Biochem Behav 40:717�721
Heinrichs SC, Menzaghi F, Schulteis G, Koob GF, Stinus L (1995)Suppression of corticotropin-releasing factor in the amygdalaattenuates aversive consequences of morphine withdrawal.Behav Pharmacol 6 :74�80
Koob GF (1996) Drug addiction: the yin and yang of hedonichomeostasis. Neuron 16:893�896
Koob GF, Heinrichs SC, Merlo-Pich E, Menzaghi F, Baldwin H,Miczek K, Britton KT (1993) The role of corticotropin-releas-ing factor in behavioural responses to stress. In: Chadwick DJ,Marsh J, Ackrill K (eds) CIBA Foundation symposium on cor-ticotropin-releasing factor, 172. Wiley, Chichester, pp 277�295
Markou A, Koob GF (1991) Post-cocaine anhedonia: an animalmodel of cocaine withdrawal. Neuropsychopharmacology 4 :17�26
Miczek KA, Mutschler NH (1996) Activational e¤ects of socialstress on IV cocaine self-administration in rats. Psychopharma-cology 128:256�264
Ramsey NF, Van Ree JM (1993) Emotional but not physical stressenhances intravenous cocaine self-administration in drug-naiverats. Brain Res 608:216�222
Rassnick S, Heinrichs SC, Britton KT, Koob GF (1993)Microinjection of a corticotropin-releasing factor antagonistinto the central nucleus of the amygdala reverses anxiogenic-like e¤ects of ethanol withdrawal. Brain Res 605:25�32
Rivier C, Vale W (1987) Cocaine stimulates adrenocorticotropin(ACTH) secretion through a corticotropin-releasing factor(CRF)-mediated mechanism. Brain Res 422:403�406
Rodrîguez de Fonseca F, Carrera MR, Navarro M, Koob GF,Weiss F (1997) Activation of corticotropin-releasing factor inthe limbic system during cannabinoid withdrawal. Science 276:2050�2054
Sarnyai Z, Höhn J, Szabó G, Penke B (1992) Critical role of endoge-nous corticotropin-releasing factor (CRF) in the mediationof the behavioral action of cocaine in rats. Life Sci 51:2019�2024
254

Sarnyai Z, Bíró É, Gardi J, Vecsernyés M, Julesz J, Telegdy G(1993) Alterations of corticotropin-releasing factor-likeimmunoreactivity in di¤erent brain regions after acute cocaineadministration in rats. Brain Res 616:315�319
Shaham Y, Rajabi H, Stewart J (1996) Relapse to heroin-seekingin rats under opioid maintenance: the e¤ects of stress, heroinpriming, and withdrawal. J Neurosci 16:1957�1963
Shaham Y, Funk D, Erb S, Brown TJ, Walker C-D, Stewart J(1997) Corticotropin-releasing factor, but not corticosterone, isinvolved in stress-induced relapse to heroin-seeking in rats. JNeurosci 17:2605�2614
Swerdlow NR, Gilbert D, Koob GF (1989) Conditioned drug e¤ectson spatial preference. In: Boulton AA, Baker BB, GreenshawAJ (eds) Neuromethods. Psychopharmacology, vol 13. TheHumana Press, Clifton, N.J., pp 399�446
Turenne SD, Miles C, Parker LA, Siegel S (1996) Individualdi¤erences in reactivity to the rewarding/aversive properties ofdrugs: assessment by taste and place conditioning. PharmacolBiochem Behav 53:511�516
White N, Sklar L, Amit Z (1977) The reinforcing action of mor-phine and its paradoxical side e¤ect. Psychopharmacology52:63�66
Yang X-M, Gorman AL, Dunn AJ, Goeders NE (1992) Anxiogenice¤ects of acute and chronic cocaine administration: neuro-chemical and behavioral studies. Pharmacol Biochem Behav41:643�650
Zhou Y, Spangler R, LaForge S, Maggos CE, Ho A, Kreek MJ(1996) Corticotropin-releasing factor and type 1 corticotropin-releasing factor receptor messenger RNAs in rat brain and pitu-itary during �binge�-pattern cocaine administration andchronic withdrawal. J Pharmacol Exp Ther 279:351�358
255
Related Documents











