Correspondences between plants and soil/environmental factors in beech forests of Central Apennines: from homogeneity to complexity Anna Testi Cristina De Nicola Giuseppina Dowgiallo Giuliano Fanelli Received: 28 April 2009 / Accepted: 20 August 2009 Ó Springer-Verlag Italia 2009 Abstract Data have been collected in beech forests of central Apennines through 94 phytosociological releve `s and 37 soil profiles. The main edaphic factors have been ana- lyzed. Environmental factors [light (L), temperature (T), continentality (K), soil moisture (F), reaction (R), nitrogen (N), hemeroby (H)] have been expressed by Ellenberg bioin- dication model and hemeroby index to estimate anthropogenic disturbance. Significant correlations have been found by Pearson correlation test: the distribution of beech forest typical species, such as Cephalanthera damasonium, Aquilegia vulgaris were positively correlated with carbon/nitrogen (C/N) ratio. Five factors (T and L indicator values, hemeroby index, CaCO 3 , C/N ratio) explained almost the whole variability of data set in the canonical correspondence analysis. The relationships found between soil/environ- mental factors and species/communities allow to detect significant differences within an homogeneous habitat providing management indications at fine-scale. Keywords Beech forest Plant distribution Soil Hemeroby Ellenberg indicators Correlation Canonical correspondence analysis 1 Introduction It is well known that forest species are highly correlated with environmental factors (Bo ¨cker et al. 1983; Kaiser and Ka ¨ding 2005; Schaffers and Sykora 2000; Southall et al. 2004; Schmidtlein and Ewald 2003). The challenge in plant ecology is to select a reduced set among the innumerable factors that are implied in the response of species and com- munities. In other words, among the manifold of factors, only a few act as key factors, structuring the ecology of communities (Fanelli et al. 2006a). A. Testi (&) C. De Nicola G. Dowgiallo G. Fanelli Department of Plant Biology, Botanical Garden, University of Rome ‘‘La Sapienza’’, Largo Cristina di Svezia, 24, 00165 Rome, Italy e-mail: [email protected] 123 Rend. Fis. Acc. Lincei DOI 10.1007/s12210-009-0054-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Correspondences between plants and soil/environmentalfactors in beech forests of Central Apennines:from homogeneity to complexity
Anna Testi Æ Cristina De Nicola Æ Giuseppina Dowgiallo ÆGiuliano Fanelli
Received: 28 April 2009 / Accepted: 20 August 2009� Springer-Verlag Italia 2009
Abstract Data have been collected in beech forests of central Apennines through 94
phytosociological releves and 37 soil profiles. The main edaphic factors have been ana-
lyzed. Environmental factors [light (L), temperature (T), continentality (K), soil moisture
(F), reaction (R), nitrogen (N), hemeroby (H)] have been expressed by Ellenberg bioin-
dication model and hemeroby index to estimate anthropogenic disturbance. Significant
correlations have been found by Pearson correlation test: the distribution of beech forest
typical species, such as Cephalanthera damasonium, Aquilegia vulgaris were positively
correlated with carbon/nitrogen (C/N) ratio. Five factors (T and L indicator values,
hemeroby index, CaCO3, C/N ratio) explained almost the whole variability of data set in
the canonical correspondence analysis. The relationships found between soil/environ-
mental factors and species/communities allow to detect significant differences within an
homogeneous habitat providing management indications at fine-scale.
Keywords Beech forest � Plant distribution � Soil � Hemeroby � Ellenberg indicators �Correlation � Canonical correspondence analysis
1 Introduction
It is well known that forest species are highly correlated with environmental factors
(Bocker et al. 1983; Kaiser and Kading 2005; Schaffers and Sykora 2000; Southall et al.
2004; Schmidtlein and Ewald 2003). The challenge in plant ecology is to select a reduced
set among the innumerable factors that are implied in the response of species and com-
munities. In other words, among the manifold of factors, only a few act as key factors,
structuring the ecology of communities (Fanelli et al. 2006a).
A. Testi (&) � C. De Nicola � G. Dowgiallo � G. FanelliDepartment of Plant Biology, Botanical Garden,University of Rome ‘‘La Sapienza’’, Largo Cristina di Svezia, 24,00165 Rome, Italye-mail: [email protected]
123
Rend. Fis. Acc. LinceiDOI 10.1007/s12210-009-0054-8

In ecosystems, two sets of factors are responsible for the distribution of species and
communities: static and dynamic. pH, sand, silt, clay, organic matter, pool of nitrogen in
soil, are examples of the former; mineralization rate, water input and outputs from the
ecosystem, average temperature in the course of the year, are examples of the latter. We
studied in particular static edaphic factors by soil profiles and pedological analyses.
Dynamic factors can be measured only with difficulty (Schimel and Bennet 2004) and with
long-term research effort. These factors are instead easily estimated by use of ecoindica-
tors, such as Ellenberg’s (Ellenberg 1974–1979; Testi et al. 2006a; Diekmann 1995) and
Hemeroby (Kowarik 1990) models.
Studies relating the distribution of species to environmental factors have a long history in
Central Europe; such studies are instead scarce in the Mediterranean area and in particular in
the Apennines, notwithstanding the extensive phytosociological literature. For instance,
Apennine beech woodlands are well known from a phytosociological point of view (e.g.
Feoli and Lagonegro 1982; Pignatti 1998), whereas synecological investigations are lacking.
Aims of this study are:
1. To verify the relationships between single species and soil factors in beech woodlands
of Central Apennines on calcareous rocks;
2. To study the response of the communities to the different soil and environmental
(static and dynamic) factors.
2 Materials and methods
2.1 Study area
The study was carried out within a project Life 04NAT/IT/000190 by the Department of
Plant Biology of the University of Rome (La Sapienza) and the National Forest Service.
The beech forests investigated are located in the Upper Sangro Valley in Central Apen-
nines (Abruzzo region) and include seven sites of community importance (SCI), extended
over an area of 200 km2, with altitudes ranging between 900 and 1,994 m a.s.l. This area
has a long history of silvicultural management, as in most Apennine areas: overgrazing and
fires caused a degradation of this habitat leading to soil thinning as well as to decreasing of
species diversity in the herbaceous layer. For the last 30 years silvicultural management
changed towards an ecosystemic approach: shrubs, herbs, faunistic and microbiological
components are taken into account. The managers stop to consider the forest only as an
high-productivity system, but a naturalistic management promotes the development of
stands with different stages of growth instead of coppiced stands.
The forest is dominated by Fagus sylvatica with a few specimens of Fraxinus excelsior,
Carpinus betulus, Acer campestre and Quercus robur. Occurrences of Taxus baccata and
Ilex aquifolium were recorded in mixed stands with Fagus according to ‘‘European Habitat
Directive 2000’’.
Although the beech stands are rather homogeneous in the composition of the herbaceous
layer, they show differences both in the density and in the species richness of the under-
story: a mosaic-like pattern of species-poor stands with low coverage alternated with richer
stands with clonal populations of Cardamine enneaphyllos, Cardamine bulbifera, Galiumodoratum, Viola reichenbachiana, Corydalis cava.
The main bedrocks in the area are calcareous. Most of the soils found throughout the
study area (De Nicola et al. 2007a) have a moderately deep ABwC profile: they consist of a
Rend. Fis. Acc. Lincei
123

rather thin epipedon with a crumb structure, brownish black colors and very high organic
matter contents (from 10 up to 20%) and a cambic B horizon, subangular blocky, often
containing calcareous gravel and gradually merging into a stony C horizon. Most of these
soils are slightly acid to neutral and free of primary carbonates, but have a rather high base
saturation ([60%), thus they can be classified as Dystric Eutrudepts. Occasionally Typical
Eutrudepts have been found, which are slightly to moderately alkaline throughout the
profile, slightly calcareous and with very high base contents (BS [ 90%). All these soils
are moderately fine textured (silt loam) and have a medium to very high water availability
(AWC = 120–280 mm). Their moisture regime is udic, the temperature regime ranges
from mesic to frigid with altitude.
In strongly eroded areas, the dominant soils are rendzina-types with a xeric moisture
regime (Lithic and Typic Haploxerolls) with a very dark mollic epipedon, very rich in
organic matter (16–26%), 35–50 cm thick, underlain by a stony C horizon, or, more rarely,
by hard limestone. These soils have a loamy texture with abundant calcareous fragments of
various sizes and a rather low water availability (80–100 mm); the matrix is slightly to
moderately calcareous and pH values are neutral or slightly alkaline.
2.2 Vegetation data collecting
In the first step vegetation data were collected in 94 sampling sites chosen to represent:
(1) different vegetation types;
(2) different forest management types.
Sampling sites are 100 m2 in size. In each one, vegetation composition was surveyed
with a phytosociological releve. Nomenclature of species follows Pignatti (1982).
A matrix of 175 species 9 94 releves was obtained.
2.3 Sampling design
Among the 94 sampling sites, a subset of 37 plots (Fig. 1) was selected for soil profile
description and pedological analyses. These plots were chosen to represent different plant
communities and different forest management types, based on the following repartitions:
ten plots in coppiced stands, ten in the mature high-growth forest stands, ten in the young
high-growth forest stands and seven in stands under transition to high forest. Soil and
vegetation data have been collected simultaneously.
2.4 Soil factors sampling
In the subset of 37 plots, soil profiles were studied through pits and augering. For field
description of the macromorphological soil characteristics the Soil Survey Division Staff
(1993) was used. The following physical and chemical analyses were carried out on a total
of 111 soil samples, according to USDA methods (1996): pH (in H2O), total calcium
carbonate (CaCO3), particle-size analysis, organic carbon (C) and nitrogen content,
exchangeable bases (Ca??, Na?, K?, Mg??), cation exchange capacity (CEC),
exchangeable acidity (EA), base saturation (BS). Moreover, two other factors were cal-
culated: available water capacity (AWC) and ratio between carbon and nitrogen (C/N).
Available water capacity was estimated using the following equation proposed by Salter
and Williams (1969), based on textural composition and percentage of organic matter:
Rend. Fis. Acc. Lincei
123

AWC ðmm H2O=cm soil depth cm) ¼ 1:475� 0:010 ð% coarse sandÞ þ 0:011 ð% siltÞþ 0:138 ð% organic carbonÞ:
Diagnostic horizons as well as soil profiles were classified to the subgroup level of Soil
Taxonomy (USDA 1999, 2006).
2.5 Environmental factors
Ellenberg indicator values (Ellenberg 1974–1979) represent a synthetic and effective way
to analyze and express ecosystems complexities (Pignatti et al. 2001; Testi et al. 2004);
these ecoindicators express the average realized niche for species along the gradients light
(L), temperature (T), continentality of climate (K), soil moisture (F), soil reaction (R),
nitrogen (N). Ellenberg indicator values are often used in plant ecology research
Fig. 1 Study area is marked with bold lines surrounding the sites of community importance (SCI). Filledcircles correspond to sampling sites following progressive numbers and identified by the acronyms of thecommunities. A Mesophile beech woodlands, B pioneer beech woodlands, C thermophile and ecotonalbeech woodlands, D mixed woodlands
Rend. Fis. Acc. Lincei
123

(Diekmann 1995; Schaffers and Sykora 2000), in comparative studies on plant commu-
nities (Dupre and Diekmann 1998; Testi et al. 2006b), and to relate vegetation patterns to
environmental changes (Pignatti et al. 2001). Since the beginning of phytosociology,
Braun-Blanquet and Jenny (1926) and Guinochet (1938) pointed out that species occur-
rences and plant associations may be defined through an ecological approach based on
factors measured in the field as well as on ecological indicators (Ellenberg 1963); they
foresaw the development of a multidimensional analysis overcoming the approach
exclusively based on floristic analysis.
Closely related to Ellenberg indicator values is the Hemeroby index (H), expressing the
degree of past and present human disturbance on ecosystems according to a ten-point scale
(van der Maarel 1979; Kowarik 1990; Fanelli and De Lillis 2004). Disturbance is a general
ecological factor in nature and it is a primary cause of spatial heterogeneity in ecosystems
(Platt 1975; Loucks et al. 1985; Collins and Glenn 1988; White and Jentsch 2001).
Unfortunately, direct estimation of disturbance and human impact is usually difficult. It is
therefore necessary to evaluate disturbance indirectly by means of changes in the com-
position of communities. So, in practice we do not study disturbance directly, but the
response of vegetation to disturbance (Fanelli and Testi 2008).
The figures for ecoindicators have been taken from a data-bank for the species of the
Mediterranean flora (Fanelli et al. 2006b, c). The values were averaged over the matrix
‘‘species/releves’’, obtaining an ecological matrix where each species had seven indicators
values (L, T, K, F, R, N, H). For each plot, indicator values were weighted on the species
coverages, obtaining an ecological characterization for site.
2.6 Statistical analysis
Pearson correlation test was applied to investigate the relationships between species dis-
tribution and soil factors. Furthermore, since soil and environmental factors are indepen-
dently measured and calculated, also correlations between them were considered. In order
to identify multivariate correspondence between all analyzed soil/environmental factors
expressed by ecoindicators and coverage of all species, canonical correspondence analysis
(CCA) has been applied using SYNTAX 2000 (Podani 2000). The normalized scores for
factors stressing their weight on the CCA axes were utilized to recognize the main gra-
dients along which species and communities were distributed. The CCA results are pre-
sented as ordination triplots with sampling plots, species and soil/environmental factors by
scores for set with axes constrained.
3 Results
3.1 Vegetation
The 94 plots of the complete dataset can be grouped into four types of communities that
have been previously recognized in a separate study (De Nicola et al. 2007a); these types
are represented in a map of the study area (Fig. 1):
• mesophile beech woodlands at higher altitudes; this type is the most widespread in the
study area (Type A).
• pioneer beech woodlands, mainly distributed in the NE slope of the mountain ridges on
stony soils (Type B).
Rend. Fis. Acc. Lincei
123

• thermophile beech woodlands at lower altitudes; this type is quite rare in the study area
(Type C).
• mixed woodlands with turkey oak (Quercus cerris) on marls, at lower altitudes (Type
D).
3.2 Output of correlation analysis
Pearson correlation test performed on the subset of 37 plots where phytosociological
releves and soil profiles were simultaneously carried out, displayed many significant
correlations between species and soil factors. Individual species resulted positively or
negatively correlated with one or more soil factors (Table 1). Furthermore, many signifi-
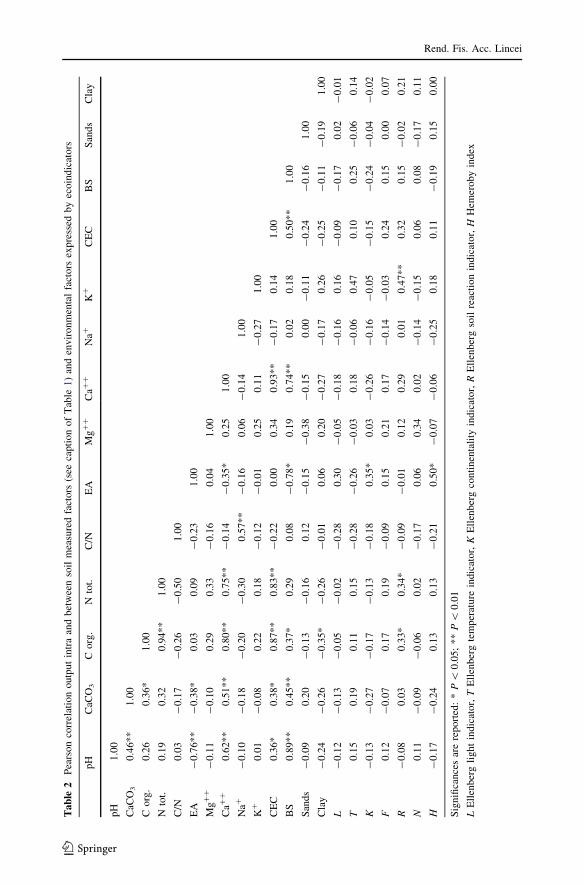
cant correlations emerged also from intra-data set and from measured soil factors versusecoindicators (Table 2).
It is worth mentioning that in many cases correlations between species and soil factors
resulted significant even when correlation coefficients were low (for instance the corre-
lation between Rhamnus alpinus and organic carbon has r2 = 0.34 but is nonetheless
significant).
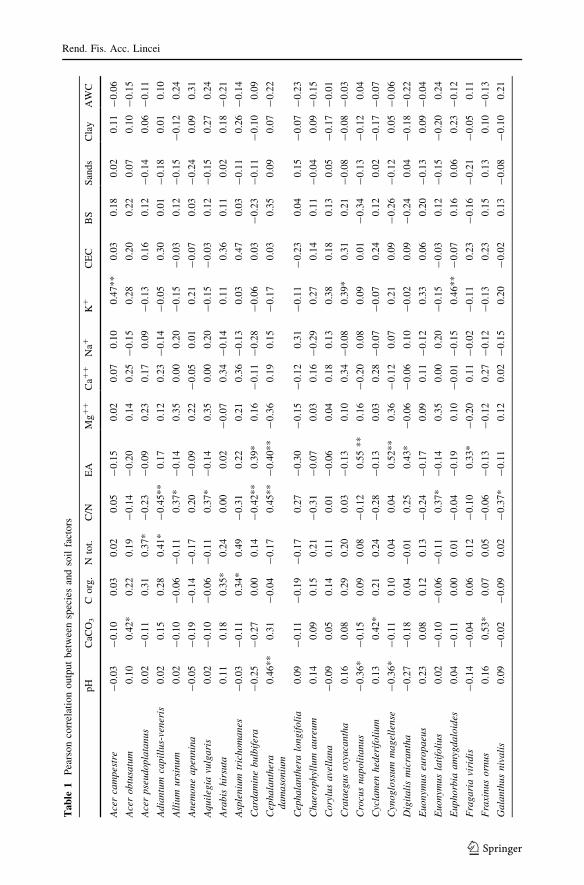
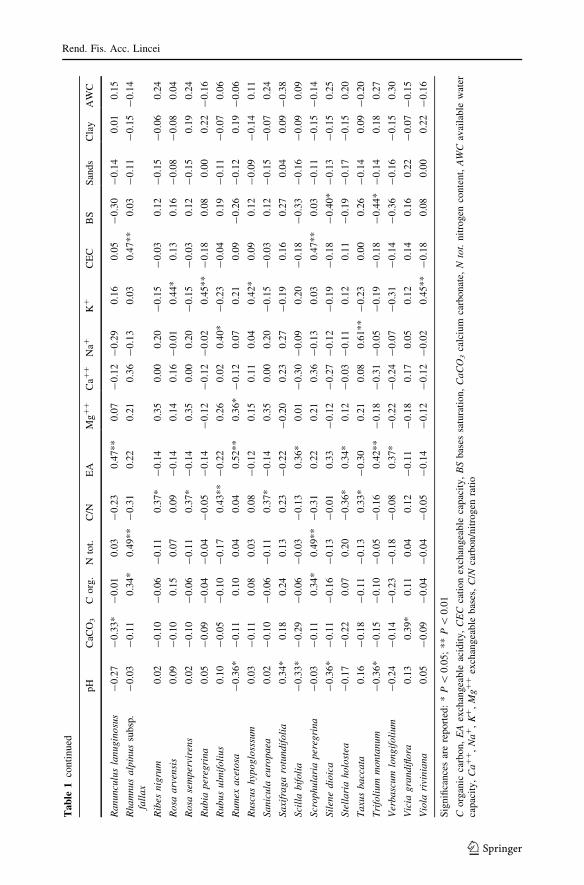
The most important correlations reported in Tables 1 and 2 are described below:
CaCO3 Soils with high content of calcium carbonate are positively correlated with tree
species such as Laburnum anagyroides, Fraxinus ornus, Pinus nigra subsp. nigra and Acerobtusatum: these species colonize sites where competition with Fagus is low (Pignatti
1998). CaCO3 is also positively correlated with pH, CEC, organic carbon, BS, Ca?? and
negatively with EA.
C Organic carbon is correlated with species occurring at the edge of the forest, such as
Rhamnus alpinus subsp. fallax, Arabis hirsuta, Asplenium trichomanes, Milium effusum,
Scrophularia peregrina, exhibiting high requirement for soil nutrients (Wilson et al. 2001),
as shown by high values of nitrogen Ellenberg indicator (N), ranged between 7 and 8.
Organic carbon is also correlated with soil reaction indicator value (R) and with several soil
factors: CEC, BS, CaCO3, nitrogen content, Ca??.
Nitrogen content Several species distributed in the different communities are positively
correlated with nitrogen content. This factor is also correlated with R indicator and with
other soil factors such as organic carbon, Ca??, and CEC. Correlation values for organic
carbon, Ca?? and CEC are very high (0.75–0.83) (Table 2).
C/N ratio Carbon/nitrogen (C/N) ratio is the factor that is correlated with the largest
number of different species; these are mainly mesophile species (more common in Type A
woodlands): Taxus baccata, Cephalanthera damasonium, Ilex aquifolium, Lathyrus vernus,
Euonymus latifolius, Ribes nigrum, Sanicula europaea, Allium ursinum, Aquilegiavulgaris, Hieracium sylvaticum.
pH Correlations with pH allow to discriminate subacidophile species (Poa alpina,
Cynoglossum magellense, Crocus napolitanus, Polystichum lonchitis, Rumex acetosa,
Scilla bifolia), with negative coefficient values and basophile ones (Cephalantheradamasonium, Saxifraga rotundifolia) with positive correlation: the first are dominant in
open spaces represented by wood-clearings and pasturelands; the second occur in beech
forest.
Rend. Fis. Acc. Lincei
123
Tab
le1
Pea
rson
corr
elat
ion
outp
ut
bet
wee
nsp
ecie
san
dso
ilfa
ctors
pH
CaC
O3
Co
rg.
Nto
t.C
/NE
AM
g?
?C
a??
Na?
K?
CE
CB
SS
and
sC
lay
AW
C
Ace
rca
mp
estr
e-
0.0
3-
0.1
00
.03
0.0
20
.05
-0
.15
0.0
20
.07
0.1
00
.47
**
0.0
30
.18
0.0
20
.11
-0
.06
Ace
ro
btu
satu
m0
.10
0.4
2*
0.2
20
.19
-0
.14
-0
.20
0.1
40
.25
-0
.15
0.2
80
.20
0.2
20
.07
0.1
0-
0.1
5
Ace
rp
seu
do
pla
tan
us
0.0
2-
0.1
10
.31
0.3
7*
-0
.23
-0
.09
0.2
30
.17
0.0
9-
0.1
30
.16
0.1
2-
0.1
40
.06
-0
.11
Ad
ian
tum
cap
illu
s-ve
ner
is0
.02
0.1
50
.28
0.4
1*
-0
.45
**
0.1
70
.12
0.2
3-
0.1
4-
0.0
50
.30
0.0
1-
0.1
80
.01
0.1
0
All
ium
urs
inu
m0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
-0
.12
0.2
4
An
emo
ne
ap
enn
ina
-0
.05
-0
.19
-0
.14
-0
.17
0.2
0-
0.0
90
.22
-0
.05
0.0
10
.21
-0
.07
0.0
3-
0.2
40
.09
0.3
1
Aq
uil
egia
vulg
ari
s0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
0.2
70
.24
Ara
bis
hir
suta
0.1
10
.18
0.3
5*
0.2
40
.00
0.0
2-
0.0
70
.34
-0
.14
0.1
10
.36
0.1
10
.02
0.1
8-
0.2
1
Asp
len
ium
tric
ho
ma
nes
-0
.03
-0
.11
0.3
4*
0.4
9-
0.3
10
.22
0.2
10
.36
-0
.13
0.0
30
.47
0.0
3-
0.1
10
.26
-0
.14
Ca
rda
min
eb
ulb
ifer
a-
0.2
5-
0.2
70
.00
0.1
4-
0.4
2*
*0
.39
*0
.16
-0
.11
-0
.28
-0
.06
0.0
3-
0.2
3-
0.1
1-
0.1
00
.09
Cep
ha
lan
ther
ad
am
aso
niu
m0
.46
**
0.3
1-
0.0
4-
0.1
70
.45
**
-0
.40
**
-0
.36
0.1
90
.15
-0
.17
0.0
30
.35
0.0
90
.07
-0
.22
Cep
ha
lan
ther
alo
ng
ifo
lia
0.0
9-
0.1
1-
0.1
9-
0.1
70
.27
-0
.30
-0
.15
-0
.12
0.3
1-
0.1
1-
0.2
30
.04
0.1
5-
0.0
7-
0.2
3
Ch
aer
op
hyl
lum
au
reu
m0
.14
0.0
90
.15
0.2
1-
0.3
1-
0.0
70
.03
0.1
6-
0.2
90
.27
0.1
40
.11
-0
.04
0.0
9-
0.1
5
Co
rylu
sa
vell
an
a-
0.0
90
.05
0.1
40
.11
0.0
1-
0.0
60
.04
0.1
80
.13
0.3
80
.18
0.1
30
.05
-0
.17
-0
.01
Cra
taeg
us
oxy
aca
nth
a0
.16
0.0
80
.29
0.2
00
.03
-0
.13
0.1
00
.34
-0
.08
0.3
9*
0.3
10
.21
-0
.08
-0
.08
-0
.03
Cro
cus
na
po
lita
nu
s-
0.3
6*
-0
.15
0.0
90
.08
-0
.12
0.5
5*
*0
.16
-0
.20
0.0
80
.09
0.0
1-
0.3
4-
0.1
3-
0.1
20
.04
Cyc
lam
enh
eder
ifo
liu
m0
.13
0.4
2*
0.2
10
.24
-0
.28
-0
.13
0.0
30
.28
-0
.07
-0
.07
0.2
40
.12
0.0
2-
0.1
7-
0.0
7
Cyn
og
loss
um
ma
gel
len
se-
0.3
6*
-0
.11
0.1
00
.04
0.0
40
.52
**
0.3
6-
0.1
20
.07
0.2
10
.09
-0
.26
-0
.12
0.0
5-
0.0
6
Dig
ita
lis
mic
ran
tha
-0
.27
-0
.18
0.0
4-
0.0
10
.25
0.4
3*
-0
.06
-0
.06
0.1
0-
0.0
20
.09
-0
.24
0.0
4-
0.1
8-
0.2
2
Eu
on
ymu
seu
rop
aeu
s0
.23
0.0
80
.12
0.1
3-
0.2
4-
0.1
70
.09
0.1
1-
0.1
20
.33
0.0
60
.20
-0
.13
0.0
9-
0.0
4
Eu
on
ymu
sla
tifo
liu
s0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
-0
.20
0.2
4
Eu
ph
orb
iaa
myg
da
loid
es0
.04
-0
.11
0.0
00
.01
-0
.04
-0
.19
0.1
0-
0.0
1-
0.1
50
.46
**
-0
.07
0.1
60
.06
0.2
3-
0.1
2
Fra
ga
ria
viri
dis
-0
.14
-0
.04
0.0
60
.12
-0
.10
0.3
3*
-0
.20
0.1
1-
0.0
2-
0.1
10
.23
-0
.16
-0
.21
-0
.05
0.1
1
Fra
xin
us
orn
us
0.1
60
.53
*0
.07
0.0
5-
0.0
6-
0.1
3-
0.1
20
.27
-0
.12
-0
.13
0.2
30
.15
0.1
30
.10
-0
.13
Ga
lan
thu
sn
iva
lis
0.0
9-
0.0
2-
0.0
90
.02
-0
.37
*-
0.1
10
.12
0.0
2-
0.1
50
.20
-0
.02
0.1
3-
0.0
8-
0.1
00
.21
Rend. Fis. Acc. Lincei
123

Tab
le1
con
tin
ued
pH
CaC
O3
Co
rg.
Nto
t.C
/NE
AM
g?
?C
a??
Na?
K?
CE
CB
SS
and
sC
lay
AW
C
Ga
liu
ma
pa
rin
e-
0.1
6-
0.1
2-
0.0
90
.02
-0
.25
0.2
7-
0.1
0-
0.0
2-
0.2
00
.09
0.0
7-
0.1
3-
0.0
4-
0.0
40
.11
Gen
zia
na
vern
a-
0.1
4-
0.1
10
.02
0.0
7-
0.2
10
.26
-0
.13
-0
.16
0.0
4-
0.0
7-
0.0
8-
0.2
2-
0.0
60
.19
0.1
3
Ger
an
ium
ma
cro
rrh
izu
m0
.24
0.2
6-
0.2
3-
0.1
8-
0.2
5-
0.2
40
.09
-0
.11
0.0
0-
0.0
9-
0.2
00
.17
-0
.06
-0
.15
0.2
0
Ger
an
ium
rob
erti
an
um
-0
.06
-0
.27
0.0
80
.20
-0
.33
*0
.14
-0
.02
0.0
10
.15
-0
.08
0.0
7-
0.0
5-
0.1
9-
0.2
1-
0.0
1
Gle
cho
ma
hed
era
cea
0.0
3-
0.1
10
.08
0.0
30
.08
-0
.12
0.1
50
.11
0.0
40
.42
**
0.0
90
.12
-0
.09
-0
.14
0.1
1
Hie
raci
um
sylv
ati
cum
0.1
1-
0.1
6-
0.0
3-
0.1
50
.36
*-
0.0
3-
0.1
80
.08
0.3
8-
0.1
70
.08
0.1
10
.06
-0
.31
-0
.27
Ilex
aq
uif
oli
um
-0
.30
-0
.15
-0
.28
-0
.28
0.3
5*
-0
.10
0.1
7-
0.2
30
.45
**
-0
.24
-0
.26
-0
.21
0.1
80
.11
0.4
1*
Jun
iper
us
com
mu
nis
0.1
20
.06
-0
.07
-0
.18
0.4
4*
*-
0.1
9-
0.2
5-
0.0
30
.13
0.2
9-
0.1
10
.16
0.0
9-
0.0
6-
0.2
9
La
bu
rnu
ma
na
gyr
oid
es0
.18
0.4
4*
0.0
4-
0.0
60
.24
-0
.21
-0
.20
0.2
60
.19
0.0
20
.19
0.2
60
.20
0.0
5-
0.3
0
La
miu
mg
arg
an
icu
m-
0.2
1-
0.2
7-
0.0
8-
0.0
7-
0.1
10
.50
**
-0
.11
-0
.28
-0
.11
-0
.02
-0
.12
-0
.35
0.0
50
.08
0.0
9
La
thyr
us
vern
us
0.2
5-
0.0
30
.07
-0
.06
0.5
3*
*-
0.4
1*
0.0
50
.19
0.4
4-
0.2
10
.06
0.3
00
.04
0.0
5-
0.2
1
Lig
ust
rum
vulg
are
0.1
4-
0.0
5-
0.0
6-
0.0
7-
0.0
7-
0.2
60
.05
-0
.04
0.0
10
.56
**
-0
.13
0.1
9-
0.0
80
.03
0.0
3
Lo
nic
era
alp
igen
a0
.02
-0
.11
0.3
10
.37
*-
0.2
3-
0.0
90
.23
0.1
70
.09
-0
.13
0.1
60
.12
-0
.14
-0
.16
-0
.11
Lu
zula
pil
osa
-0
.25
0.0
0-
0.1
4-
0.1
60
.15
-0
.05
-0
.27
-0
.21
0.1
2-
0.1
8-
0.2
5-
0.3
70
.48
**
0.0
50
.06
Ma
lus
sylv
estr
is0
.03
-0
.11
0.0
80
.03
0.0
8-
0.1
20
.15
0.1
10
.04
0.4
2*
0.0
90
.12
-0
.09
-0
.14
0.1
1
Mil
ium
efu
sum
0.3
1-
0.0
70
.33
*0
.28
0.0
6-
0.1
40
.05
0.2
80
.12
-0
.20
0.2
40
.23
-0
.07
0.1
9-
0.3
9
Myc
elis
mu
rali
s-
0.0
20
.01
0.2
30
.30
-0
.37
*0
.21
-0
.04
0.1
3-
0.5
8-
0.0
70
.19
-0
.08
0.0
40
.21
0.0
1
Myo
soti
ssy
lva
tica
0.0
0-
0.1
10
.20
0.3
2-
0.3
5*
0.0
80
.48
0.2
6-
0.1
70
.11
0.3
30
.06
-0
.20
-0
.07
-0
.07
Neo
ttia
nid
us
avi
s0
.25
0.3
10
.31
0.3
9*
-0
.29
-0
.14
-0
.06
0.4
1-
0.1
50
.06
0.3
70
.26
0.0
1-
0.1
1-
0.1
9
Ph
ylli
tis
sco
lop
end
riu
m0
.02
-0
.11
0.3
10
.37
*-
0.2
3-
0.0
90
.23
0.1
70
.09
-0
.13
0.1
60
.12
-0
.14
0.0
2-
0.1
1
Pin
us
nig
rasu
bsp
.n
igra
0.1
60
.53
**
0.0
70
.05
-0
.06
-0
.13
-0
.12
0.2
7-
0.1
2-
0.1
30
.23
0.1
50
.13
0.0
9-
0.1
3
Po
aa
lpin
a-
0.3
6*
-0
.15
-0
.10
-0
.05
-0
.16
0.4
2*
*-
0.1
8-
0.3
1-
0.0
5-
0.1
9-
0.1
8-
0.4
4*
-0
.14
0.1
90
.27
Po
lyst
ich
um
lon
chit
is-
0.4
1*
-0
.15
-0
.07
-0
.08
-0
.04
0.3
4*
0.3
1-
0.2
00
.03
0.1
8-
0.0
6-
0.2
00
.08
0.0
2-
0.0
2
Pri
mu
lavu
lga
ris
-0
.11
-0
.15
-0
.18
-0
.14
-0
.10
-0
.13
-0
.04
-0
.19
-0
.04
0.3
6*
-0
.25
0.0
40
.16
0.3
1-
0.0
9
Qu
ercu
sce
rris
-0
.03
-0
.10
0.0
10
.02
-0
.07
-0
.15
-0
.15
-0
.10
0.0
40
.55
**
-0
.16
-0
.01
0.1
00
.14
-0
.14
Ra
nu
ncu
lus
fica
ria
0.2
50
.18
-0
.24
-0
.18
-0
.25
-0
.28
0.1
1-
0.1
30
.01
0.0
4-
0.2
30
.20
-0
.10
-0
.10
0.1
9
Rend. Fis. Acc. Lincei
123
Tab
le1
con
tin
ued
pH
CaC
O3
Co
rg.
Nto
t.C
/NE
AM
g?
?C
a??
Na?
K?
CE
CB
SS
and
sC
lay
AW
C
Ra
nu
ncu
lus
lan
ug
ino
sus
-0
.27
-0
.33
*-
0.0
10
.03
-0
.23
0.4
7*
*0
.07
-0
.12
-0
.29
0.1
60
.05
-0
.30
-0
.14
0.0
10
.15
Rh
am
nu
sa
lpin
us
sub
sp.
fall
ax
-0
.03
-0
.11
0.3
4*
0.4
9*
*-
0.3
10
.22
0.2
10
.36
-0
.13
0.0
30
.47
**
0.0
3-
0.1
1-
0.1
5-
0.1
4
Rib
esn
igru
m0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
-0
.06
0.2
4
Ro
saa
rven
sis
0.0
9-
0.1
00
.15
0.0
70
.09
-0
.14
0.1
40
.16
-0
.01
0.4
4*
0.1
30
.16
-0
.08
-0
.08
0.0
4
Ro
sase
mp
ervi
ren
s0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
0.1
90
.24
Ru
bia
per
egri
na
0.0
5-
0.0
9-
0.0
4-
0.0
4-
0.0
5-
0.1
4-
0.1
2-
0.1
2-
0.0
20
.45
**
-0
.18
0.0
80
.00
0.2
2-
0.1
6
Ru
bu
su
lmif
oli
us
0.1
0-
0.0
5-
0.1
0-
0.1
70
.43
**
-0
.22
0.2
60
.02
0.4
0*
-0
.23
-0
.04
0.1
9-
0.1
1-
0.0
70
.06
Ru
mex
ace
tosa
-0
.36
*-
0.1
10
.10
0.0
40
.04
0.5
2*
*0
.36
*-
0.1
20
.07
0.2
10
.09
-0
.26
-0
.12
0.1
9-
0.0
6
Ru
scu
sh
ypo
glo
sssu
m0
.03
-0
.11
0.0
80
.03
0.0
8-
0.1
20
.15
0.1
10
.04
0.4
2*
0.0
90
.12
-0
.09
-0
.14
0.1
1
Sa
nic
ula
euro
pa
ea0
.02
-0
.10
-0
.06
-0
.11
0.3
7*
-0
.14
0.3
50
.00
0.2
0-
0.1
5-
0.0
30
.12
-0
.15
-0
.07
0.2
4
Sa
xifr
ag
aro
tun
dif
oli
a0
.34
*0
.18
0.2
40
.13
0.2
3-
0.2
2-
0.2
00
.23
0.2
7-
0.1
90
.16
0.2
70
.04
0.0
9-
0.3
8
Sci
lla
bif
oli
a-
0.3
3*
-0
.29
-0
.06
-0
.03
-0
.13
0.3
6*
0.0
1-
0.3
0-
0.0
90
.20
-0
.18
-0
.33
-0
.16
-0
.09
0.0
9
Scr
op
hu
lari
ap
ereg
rin
a-
0.0
3-
0.1
10
.34
*0
.49
**
-0
.31
0.2
20
.21
0.3
6-
0.1
30
.03
0.4
7*
*0
.03
-0
.11
-0
.15
-0
.14
Sil
ene
dio
ica
-0
.36
*-
0.1
1-
0.1
6-
0.1
3-
0.0
10
.33
-0
.12
-0
.27
-0
.12
-0
.19
-0
.18
-0
.40
*-
0.1
3-
0.1
50
.25
Ste
lla
ria
ho
lost
ea-
0.1
7-
0.2
20
.07
0.2
0-
0.3
6*
0.3
4*
0.1
2-
0.0
3-
0.1
10
.12
0.1
1-
0.1
9-
0.1
7-
0.1
50
.20
Ta
xus
ba
cca
ta0
.16
-0
.18
-0
.11
-0
.13
0.3
3*
-0
.30
0.2
10
.08
0.6
1*
*-
0.2
30
.00
0.2
6-
0.1
40
.09
-0
.20
Tri
foli
um
mo
nta
nu
m-
0.3
6*
-0
.15
-0
.10
-0
.05
-0
.16
0.4
2*
*-
0.1
8-
0.3
1-
0.0
5-
0.1
9-
0.1
8-
0.4
4*
-0
.14
0.1
80
.27
Ver
ba
scu
mlo
ng
ifo
liu
m-
0.2
4-
0.1
4-
0.2
3-
0.1
8-
0.0
80
.37
*-
0.2
2-
0.2
4-
0.0
7-
0.3
1-
0.1
4-
0.3
6-
0.1
6-
0.1
50
.30
Vic
iag
ran
difl
ora
0.1
30
.39
*0
.11
0.0
40
.12
-0
.11
-0
.18
0.1
70
.05
0.1
20
.14
0.1
60
.22
-0
.07
-0
.15
Vio
lari
vin
ian
a0
.05
-0
.09
-0
.04
-0
.04
-0
.05
-0
.14
-0
.12
-0
.12
-0
.02
0.4
5*
*-
0.1
80
.08
0.0
00
.22
-0
.16
Sig
nifi
can
ces
are
rep
ort
ed:
*P
\0
.05
;*
*P
\0
.01
Co
rgan
icca
rbo
n,
EA
exch
ang
eab
leac
idit
y,
CE
Cca
tio
nex
chan
gea
ble
cap
acit
y,
BS
bas
essa
tura
tio
n,
Ca
CO
3ca
lciu
mca
rbo
nat
e,N
tot.
nit
rog
enco
nte
nt,
AW
Cav
aila
ble
wat
erca
pac
ity
,C
a?
?,
Na
?,
K?
,M
g?
?ex
chan
gea
ble
bas
es,
C/N
carb
on
/nit
rog
enra
tio
Rend. Fis. Acc. Lincei
123
Ta
ble
2P
ears
on
corr
elat
ion
ou
tpu
tin
tra
and
bet
wee
nso
ilm
easu
red
fact
ors
(see
capti
on
of
Tab
le1)
and
env
iro
nm
enta
lfa
cto
rsex
pre
ssed
by
eco
ind
icat
ors
pH
CaC
O3
Co
rg.
Nto
t.C
/NE
AM
g?
?C
a??
Na?
K?
CE
CB
SS
and
sC
lay
pH
1.0
0
CaC
O3
0.4
6*
*1
.00
Co
rg.
0.2
60
.36*
1.0
0
Nto
t.0
.19
0.3
20
.94*
*1
.00
C/N
0.0
3-
0.1
7-
0.2
6-
0.5
01
.00
EA
-0
.76*
*-
0.3
8*
0.0
30
.09
-0
.23
1.0
0
Mg
??
-0
.11
-0
.10
0.2
90
.33
-0
.16
0.0
41
.00
Ca?
?0
.62*
*0
.51*
*0
.80*
*0
.75
**
-0
.14
-0
.35*
0.2
51
.00
Na?
-0
.10
-0
.18
-0
.20
-0
.30
0.5
7*
*-
0.1
60
.06
-0
.14
1.0
0
K?
0.0
1-
0.0
80
.22
0.1
8-
0.1
2-
0.0
10
.25
0.1
1-
0.2
71
.00
CE
C0
.36*
0.3
8*
0.8
7*
*0
.83
**
-0
.22
0.0
00
.34
0.9
3*
*-
0.1
70
.14
1.0
0
BS
0.8
9*
*0
.45*
*0
.37*
0.2
90
.08
-0
.78*
0.1
90
.74*
*0
.02
0.1
80
.50*
*1
.00
San
ds
-0
.09
0.2
0-
0.1
3-
0.1
60
.12
-0
.15
-0
.38
-0
.15
0.0
0-
0.1
1-
0.2
4-
0.1
61
.00
Cla
y-
0.2
4-
0.2
6-
0.3
5*
-0
.26
-0
.01
0.0
60
.20
-0
.27
-0
.17
0.2
6-
0.2
5-
0.1
1-
0.1
91
.00
L-
0.1
2-
0.1
3-
0.0
5-
0.0
2-
0.2
80
.30
-0
.05
-0
.18
-0
.16
0.1
6-
0.0
9-
0.1
70
.02
-0
.01
T0
.15
0.1
90
.11
0.1
5-
0.2
8-
0.2
6-
0.0
30
.18
-0
.06
0.4
70
.10
0.2
5-
0.0
60
.14
K-
0.1
3-
0.2
7-
0.1
7-
0.1
3-
0.1
80
.35*
0.0
3-
0.2
6-
0.1
6-
0.0
5-
0.1
5-
0.2
4-
0.0
4-
0.0
2
F0
.12
-0
.07
0.1
70
.19
-0
.09
0.1
50
.21
0.1
7-
0.1
4-
0.0
30
.24
0.1
50
.00
0.0
7
R-
0.0
80
.03
0.3
3*
0.3
4*
-0
.09
-0
.01
0.1
20
.29
0.0
10
.47
**
0.3
20
.15
-0
.02
0.2
1
N0
.11
-0
.09
-0
.06
0.0
2-
0.1
70
.06
0.3
40
.02
-0
.14
-0
.15
0.0
60
.08
-0
.17
0.1
1
H-
0.1
7-
0.2
40
.13
0.1
3-
0.2
10
.50*
-0
.07
-0
.06
-0
.25
0.1
80
.11
-0
.19
0.1
50
.00
Sig
nifi
cance
sar
ere
port
ed:
*P
\0
.05;
**
P\
0.0
1
LE
llen
ber
gli
gh
tin
dic
ato
r,T
Ell
enber
gte
mp
erat
ure
indic
ato
r,K
Ell
enb
erg
con
tin
enta
lity
ind
icat
or,
RE
llen
ber
gso
ilre
acti
on
indic
ator,
HH
emer
oby
ind
ex
Rend. Fis. Acc. Lincei
123

K? Potassium shows positive correlations only with species such as Quercus cerris, Acercampestre, Ligustrum vulgare, Euphorbia amygdaloides, Rubia peregrina, species typical
of mixed woodlands.
Soil exchangeable acidity EA is positively correlated with species prevailing on the
forest fringes and wood clearings (Digitalis micrantha, Cynoglossum magellense, Fragariaviridis, Lamium garganicum, Ranunculus lanuginosus, Stellaria holostea, Trifoliummontanum, Verbascum longifolium). This soil factor is correlated positively with hemeroby
index (H), continentality indicator (K), and negatively with BS and Ca??.
3.3 CCA output
Variances explained by the first three axes are 37% (I), 33% (II) and 26% (III), respec-
tively. First axis was positively correlated with temperature indicator (T); second axis was
negatively correlated with hemeroby (H) and positively with carbon/nitrogen ratio (C/N);
third axis was positively correlated with soil nitrogen indicator (N) and negatively with
CaCO3 and Ca?? in the soil (Table 3).
The dataset is ordered along two main axes (Fig. 2): axis I separates thermophile
woodlands (both thermophile beech and mixed oak woodlands) on the positive side and
mesophile beech woodlands on the negative side; axis II separates plots with high C/Nratio, low Hemeroby index (types A and C beech woodlands), from plots with low C/Nratio, high Hemeroby index (type B beech woodland and D mixed oak woodlands).
Species are distributed on the plane of CCA in four groups:
1� A large group of typical species of mesophile beech woodlands under low light
conditions is distributed in the upper left quadrant of CCA: Taxus baccata, Geraniumversicolor, Paris quadrifolia, Prenanthes purpurea, Lathyrus vernus, Polypodiumvulgare, Actaea spicata, Cephalanthera damasonium, Cephalanthera longifolia,
Polystichum setiferum, Corydalis cava, Adenostyles australis, Hieracium sylvaticum,
Galium odoratum. A few of these species such as Taxus baccata, Lathyrus vernus,
Cephalanthera damasonium, Hieracium sylvaticum, resulted directly correlated with
C/N ratio by Pearson test (Table 1) and by CCA (Fig. 2). According to Ellenberg
indicator values (1979), all the species of this group exhibit a low requirement for
light (values ranged from 1 to 3), and in fact, are distributed in the negative sector of
light indicator. These species occur mainly in the community identified as Type A.
2� A smaller group of species such as: Arabis alpina, Galium album, Lamiumgarganicum, Digitalis micrantha, Doronicum columnae, Campanula glomerata,
Laburnum anagyroides is distributed in the lower left sector of CCA. Tree species,
Table 3 Significant (P \ 0.05)correlations of soil factors andecoindicators versus CCA axes
T temperature indicator, N soilnutrients indicator, H hemerobyindex, CaCO3 calcium carbonate,C/N carbon/nitrogen ratio, Ca??
exchangeable base
Factors CCA axes
I axis II axis III axis
T 0:56
N 0:40
H -0.49
CaCO3 -0.40
C/N 0:52
Ca?? -0.41
Rend. Fis. Acc. Lincei
123
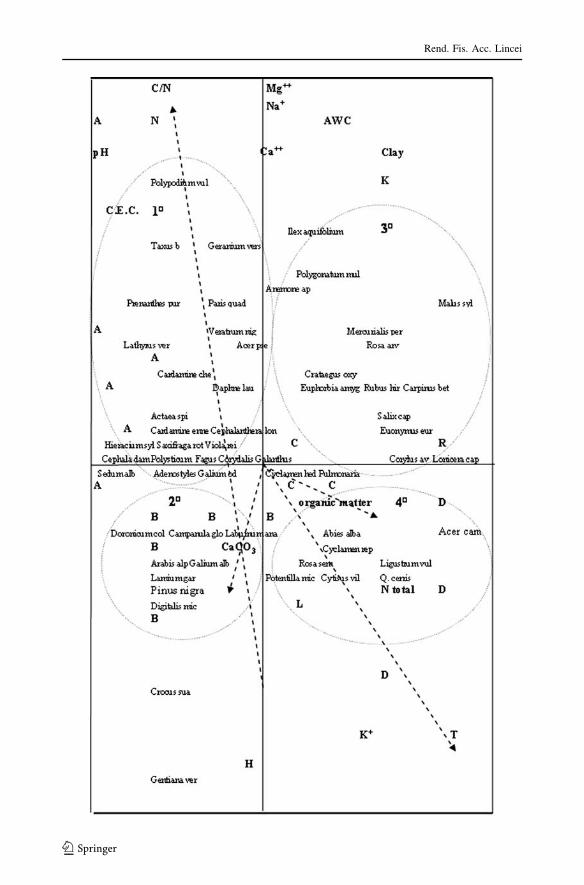
Rend. Fis. Acc. Lincei
123

such as Laburnum anagyroides and Pinus nigra subsp. nigra, resulted correlated with
CaCO3 both in CCA and in Pearson test. Some herbaceous species of this group—
Digitalis micrantha and Lamium garganicum—are fringe species distributed in the
sector of CCA related to high H and are positively correlated with EA (Table 1). These
species are abundant in pioneer beech woodlands (type B community) on soils rich in
CaCO3.
3� Ilex aquifolium, Polygonatum multiflorum, Anemone apennina, Mercurialis perennis,
Carpinus betulus, Euphorbia amygdaloides, Euonymus europaeus, Corylus avellana,
Rosa arvensis, Malus sylvestris, Crataegus oxyacantha are distributed in the upper
right quadrant of CCA, correlated with AWC, clay, Na?. Ilex aquifolium is correlated
with AWC and Na? also in the Pearson test, Anemone apennina with AWC (Table 1).
The species of this group range from thermophile beech woodlands of lower altitude
with Ilex aquifolium to ecotonal woodlands with Carpinus betulus, Malus sylvestris(type C community).
4� The fourth group of species is distributed in the lower right sector of CCA, that is
correlated with total nitrogen and characterized by high light indicator (L) and
hemeroby (H). Some of these species (Ligustrum vulgare, Quercus cerris, Acercampestre) are correlated with K? both in CCA and in Pearson test (Table 1) and are
characteristic of mixed oak woodlands (type D community).
In summary, species of group 1� are distributed along the direction of C/N ratio, species
of group 2� along the direction of CaCO3, species of groups 3� and 4� along the direction
of T and L indicators and K?.
4 Discussion
Among the many interrelationships detected in this study, between species and soil/
environmental factors, only a few emerge as the main structuring factors of this vegetation.
The other factors are not unimportant, but explain only a secondary variability, as shown in
the ordination diagram (Fig. 2).
The main axis of variation is related to temperature. It distinguishes woodlands at lower
(900–1,100 m a.s.l.) and higher altitudes (about 1,400–1,700 m a.s.l.), indifferently from
the dominant tree species (either beech or oak). These two ranges correspond to the
samnitic (supramediterranean) and subatlantic (montane) vegetation belts respectively
(Pignatti 1979). This thermal distinction is also influenced by silvicultural management
interacting with temperature. The intense tree cutting caused over time a shifting of the
mesophile beech woodlands with low light requirement towards ecotonal conditions,
favoring the ingression of species of the 3� and 4� groups, typical of mixed beech
woodlands with Carpinus betulus at intermediate altitudes and with Quercus cerris at the
lowest altitudes. Interestingly, Ilex aquifolium, occupies a marginal position in the 3�
Fig. 2 CCA triplot. Four species groups are highlighted by circles. Horizontal axis is positively correlatedwith temperature indicator (T); vertical axis is negatively correlated with Hemeroby index (H) and positivelywith carbon/nitrogen ratio (C/N). Mesophile beech woodlands (A) with species of group 1� on the upper leftside; pioneer beech woodlands (B) with species of group 2� in the low left sector; thermophile and ecotonalbeech woodlands (C) with species of group 3� on the right upper side; mixed woodlands (D) with species ofgroup 4� on the low right sector of the triplot. A, B, C, D Communities; L, T, K, R, N, H environmentalindicators; soil factors pH, organic matter, CEC, N total, C/N, AWC, Clay, Ca??, Na?, K?, Mg??, CaCO3;species groups = 1�, 2�, 3�, 4�
b
Rend. Fis. Acc. Lincei
123

group, in the upper sector of CCA, and it is correlated with AWC and high C/N ratio
(Table 1; Fig. 2). Beech woodlands with Ilex, a woodland type very characteristic of
Southern Apennines and protected under the Habitat Directive, in Abruzzo reach their
northern distribution limit. This vegetation type is therefore quite fragmentary in the study
area, and occurs only on ‘‘good’’ soils rich in carbon and with high water availability.
Axis II discriminates disturbed woodlands (2� and 4� groups of species) with low values of
C/N ratio from more mature woodlands with high values of C/N ratio (group 1�). C/N ratio is
currently used as an index of soil fertility in woodlands (Vesterdal et al. 2008). Therefore, it
seems that the woodlands here considered are divided between woodlands on more fertile and
less fertile soils (values range from 8 to 19). It is interesting that this axis is also correlated
with hemeroby indicator (Table 3). Disturbance in these woodlands is mainly represented by
anthropogenic activities such as cutting and grazing by sheep and natural impact such as
permanence of snow and slope; these factors lead to soil impoverishment and erosion, and
therefore, probably, to a reduction in C/N ratio and soil fertility (Hedde et al. 2008). In many
regions of the world, anthropogenic activities alter C and N cycling process and C seques-
tration (Huygens et al. 2007). In the study area, several species of more mature beech
woodlands are correlated with high total nitrogen content (Table 1), whose availability
increases with the level of anthropogenic influence (Schimel and Bennet 2004). Probably, in
these forests, the historical impact has enhanced mineralization activity (Noirfalise 1956;
Ellenberg 1996; Gonnert 1989; Schmidt 1970; Leschner et al. 2006), generating unbalanced
cycles in respect to undisturbed ecosystems characterized by closed biogeochemical cycles
and a high capacity of nutrients retention: temperate Nothofagus forest of South Chile is one
of the last remote areas in the world exhibiting tight N cycling (Perakis and Hedin 2001).
A third axis of CCA, explaining a minor but important fraction of variability, is cor-
related with N indicator and CaCO3 (Table 3). This factor emphasizes a rare but very
distinctive pioneer beech woodland type. Species exclusive of this community are
Laburnum anagyroides, Pinus nigra subsp. nigra, Fraxinus ornus, Vicia grandiflora. These
species occur where the dominant soils are rendzina-types overlying a stony C horizon,
often on steep slopes. These soils have a loamy texture with abundant calcareous fragments
of various sizes and a rather low water availability (values range between 80 and 100 mm).
In these more eroded soils, the tree roots exploit the horizons rich in CaCO3, near the
parent rock (De Nicola et al. 2007b). These species, although correlated with CaCO3, are
not correlated with pH. In the soils investigated CaCO3 is related to the high stoniness, and
not to the reaction of the fine fraction of soil. For this reason, we can assess that these
species are not basophile, but are rather pioneer species.
Species such as Arabis hirsuta, Asplenium trichomanes, Rhamnus alpinus subsp. fallaxare correlated with organic carbon, another secondary factor that emerges only in Pearson
correlation (Table 1) and not in CCA (Fig. 2). These species are more abundant in dis-
turbed and pioneer beech woodlands, on rendzina-type soils with high organic carbon
content (16–26%). Organic carbon resulted also correlated with several soil and envi-
ronmental factors: soil reaction (R) indicator, nitrogen content, Ca??, CaCO3 (Table 2).
These relationships show that soil nutrient regime is a composite gradient of several soil
chemical variables (Wilson et al. 2001).
5 Conclusions
Notwithstanding the apparent uniformity of the floristic composition of the forest vege-
tation in the study area, the distribution of the different woodland types cannot be
Rend. Fis. Acc. Lincei
123

explained by a single set of environmental factors. From the whole set of 26 soil/envi-
ronmental factors utilized, CCA results allowed to detect 2 groups of soil/environmental
factors explaining the distribution of vegetation types and plant species in this area:
• soil factors: C/N, CaCO3;
• environmental indicators: temperature, light and hemeroby.
Each group of environmental factors explains the distribution in the territory of one of
the four communities present: mesophile beech woodlands, pioneer beech woodlands,
thermophile and ecotonal beech woodlands, and mixed woodlands.
In our analyses, the second group of species emerges as diagnostic of a very specialized
pioneer community (type B), because a stressing soil factor such as CaCO3 exerts a
selective pressure that filters a few specialized species from the large and variable floristic
pool of beech woodlands. In mesophile and mature beech woodlands (type A) with high
C/N ratio, more favorable conditions allow the growth of less specialized species and the
community is less distinctive.
The results of the two different statistical analyses provided responses at two levels:
linear correlation analysis (Table 1) allowed to detect the relationships between single
species and soil factors, while CCA ordination (Fig. 2) demonstrated that the response of
the communities is not always correspondent to species responses: a community is not the
sum of the species, but a network of relationships that cannot be detected always through
direct and linear correspondences between biotic and abiotic components. This result
supports the fact that vegetation exhibits a capacity of self-organization (Pignatti et al.
2002); for this reason the requirements of the communities for the different soil factors are
not always overlapping or coincident with the requirements of the individual species
(Table 1) of the community (Fig. 2).
The model found for the calcareous Apennine beech forests can be represented by a
multidimensional scheme emphasizing the network of relationships between species,
communities and soil/environmental factors. Even if abiotic factors classified as key fac-
tors showed to be more important in driving biotic elements of the ecosystem, nevertheless
the entire net of linkages contributes to stability and diversity of Fagus forest habitat,
determining spatial heterogeneity at species and community levels.
The model found has to be tested in other areas with different lithology, geomorphology
and soil features to enlarge the pattern of environmental conditions. Future researches
promise to be very rewarding.
Acknowledgments This study was funded by National Forest Service (Castel di Sangro, Abruzzo, Italy).We also acknowledge Dr. Sammarone and Dr. Posillico for graphic support and field data collecting.
References
Bocker P, Kowarik I, Bornkamm R (1983) Untersuchungen zur Anwendung der Zeigerwerte nach Ellen-berg. Verh Ges Okol 11:35–56
Braun-Blanquet J, Jenny H (1926) Vegetations-Entwicklung und Bodenbildung in der alpinen Stufe derZentralalpen. Schweiz. Naturforsch. Gesell. Bd. LXIII- Abh. 2
Collins SL, Glenn SM (1988) Disturbance and community structure in North American Prairies. In: DuringHJ, Werger MJA, Willems JH (eds) Diversity and pattern in plant communities. SPB AcademicPublishing, The Hague, pp 131–143
De Nicola C, Fanelli G, Dowgiallo G, Potena G, Sammarone L, Posillico M, Testi A (2007a) Soilparameters as indicators of succession in beech forest (central-Apennines). In: 16th workshop ofEuropean vegetation survey, March 2007, Rome
Rend. Fis. Acc. Lincei
123

De Nicola C, Fanelli G, Potena G, Sammarone L, Posillico M, Testi A, Dowgiallo G (2007b) Soil andhumus parameters as indicators of different species distribution in central Apennine beech forests. In:5th international congress of the European society for soil conservation: changing soils in a changingworld: the soils of tomorrow, June 25–30, Palermo, Italy
Diekmann M (1995) Use and improvement of Ellenberg’s indicator values in deciduous forests of theBoreo-nemoral zone in Sweden. Ecography 18:178–189
Dupre C, Diekmann M (1998) Prediction of occurrence of vascular plants in deciduous forests of SouthSweden by means of Ellenberg indicator values. Appl Veg Sci 1:139–150
Ellenberg H (1963) Vegetation Mitteleuropas mit den Alpen. Einfuhrung in die Phytologie IV, 2, StuttgartEllenberg H (1974–1979) Zeigerwerte der Gefaßpflanzen Mitteleuropas (Indicator values of vascular plants
in Central Europe). Scripta Geobot., 9 (2nd edn, 1979), GottingenEllenberg H (1996) Vegetation Mitteleuropas mit den Alpen, 5th edn. Ulmer, StuttgartFanelli G, De Lillis M (2004) Relative growth rate and hemerobiotic state in the assessment of disturbance
gradients. Appl Veg Sci 7:133–140Fanelli G, Testi A (2008) Detecting large and fine scale patterns of disturbance in towns by means of plant
species inventories: maps of hemeroby in the town of Rome. In: Wagner LN (ed) Urbanization 21stcentury issues and challenges. Nova Publisher, New York, pp 197–211
Fanelli G, Tescarollo P, Testi A (2006a) Ecological indicators applied to urban and suburban flora. EcolIndic 6:444–457
Fanelli G, Pignatti S, Testi A (2006b) An application case of ecological indicator values (Zeigerwerte)calculated with a simple algorithmic approach. Plant Biosyst 141(1):15–21
Fanelli G, Testi A, Pignatti S (2006c) Ecological indicator values for species in Central and Southern Italyflora. In: Il Sistema Ambientale della Tenuta Presidenziale di Castelporziano. Ricerche sulla com-plessita di un ecosistema forestale costiero mediterraneo. Accademia delle Scienze, Scritti e Doc-umenti XXXVII, Seconda Serie vol II, pp 505–564
Feoli E, Lagonegro M (1982) Syntaxonomical analysis of beech woods in the Apennines (Italy) using theprogram package IAHOPA. Plant Ecol 50(3):129–190
Gonnert T (1989) Okologische Bedingungen verschiedener Laubwaldgesellschaften des Nordwestdeusch-landen Tieflandes. Diss Bot 136:1–124
Guinochet M (1938) Etudes sur la vegetation de l’etage Alpin dans le bassin superieur de la Tinee (AM).These Doctorat d’Etat, Lyon
Hedde M, Aubert M, Thibaud D, Bureau F (2008) Dynamics of soil carbon in a beech wood chronosequenceforest. Forest Ecol Manage 255:193–202
Huygens D, R}utting T, Boeckx P, Van Cleemput O, Godoy R, M}uller C (2007) Soil nitrogen conservationmechanisms in a pristine south Chilean Nothofagus forest ecosystem. J Soil Biol Biochem.doi: 10.1016/j.soilbio.2007.04.013
Kaiser TH, Kading H (2005) Proposal for a transformation scale between bioindicatively determined watersupply levels of grassland sites and mean moisture indicator values according to Ellenberg. Arc AgricSoil Sci 51:241–246
Kowarik I (1990) Some responses of flora and vegetation to urbanization in central Europe: plant and plantcommunities in urban environments. In: Sukopp H, Heiny S, Kowarik I (eds) Urban ecology. SPBAcademic Publishing, The Hague, pp 45–74
Leschner C, Meier IC, Hertel D (2006) On the niche of Fagus sylvatica: soil nutrient status in 50 CentralEuropean beech stands on a broad range of bedrock types. Ann For Sci 63:355–368
Loucks OL, Plumb-Mentjes ML, Rogers D (1985) Gap processes and large-scale disturbances in sandprairies. In: Pickett STA, White PS (eds) The ecology of natural disturbance and patch dynamics.Academic Press, San Diego, pp 71–83
Noirfalise A (1956) La hetraie ardennaise. Bull Inst Agron et Stat Rech Gembloux 24:203–240Perakis SS, Hedin LO (2001) Fluxes and rates of nitrogen in soils of an unpolluted old-growth temperate
forest, southern. Chile Ecol 82:2245–2260Pignatti S (1979) I piani di vegetazione in Italia. Giorn Bot Ital 13:411–428Pignatti S (1982) Flora d’Italia. Edagricole, BolognaPignatti S (1998) Boschi d’Italia. Ed. UTET, TorinoPignatti S, Bianco PM, Fanelli G, Guarino R, Petersen J, Tescarollo P (2001) Reliability and effectiveness of
Ellenberg indices in checking flora and vegetation changes induced by climatic variation. In: Korner C,Walter JR, Burga CA, Edwards PJ (eds) Fingerprints of climate changes: adapted behaviour andshifting species ranges. Kluwer Accademy/Plenum Publishers, New York/London, pp 281–304
Pignatti S, Box EO, Fujiwara K (2002) A new paradigm for the XXI century. Ann Bot 2:31–58Platt WJ (1975) The colonization and formation of equilibrium plant species associations on badger dis-
turbances in a tall-grass prairie. Ecol Monogr 45:285–305
Rend. Fis. Acc. Lincei
123

Podani J (2000) SYN-TAX-PC computer programs for multivariate data analysis in ecology and system-atics—user’s guide. Scientia Publishing, Budapest
Salter PJ, Williams JB (1969) The influence of texture on the moisture of soils: relationships betweenparticle size composition and moisture content at the upper and lower limits of available water. J SoilSci 20:126–131
Schaffers A, Sykora K (2000) Reliability of Ellenberg indicator values for moisture, nitrogen and soilreaction: a comparison with field measurements. J Veg Sci 11:225–244
Schimel JP, Bennet J (2004) Nitrogen mineralization: challenges of a changing paradigm. Ecology85(3):591–602
Schmidt W (1970) Untersuchungen uber die Phosphorversorgung niersachsischer Nuchenwaldgesellschaf-ten. Scripta Geobot 1:1–205
Schmidtlein S, Ewald J (2003) Landscape patterns of indicator plants for soil acidity in the Bavarian Alps.J Biogeogr 30:1493–1503
Soil Survey Division Staff (1993) Soil survey manual. USDA Handbook 18Southall EJ, Dale MP, Kent M (2004) Spatial and temporal analysis of vegetation mosaics for conservation:
poor fen communities in a Cornish valley mire. J Biogeogr 30:1427–1443Testi A, Crosti R, Dowgiallo G, Tescarollo P, De Nicola C, Guidotti S, Bianco PM, Serafini Sauli A (2004)
Soil water availability as a discriminant factor in forest vegetation: preliminary results on sub-coastalmixed oak woodlands in central-southern Latium (Central Italy). Ann Bot IV:49–64
Testi A, Cara E, Fanelli G (2006a) An example of realization of Gis ecological maps derived from Ellenbergindicator values in the Biological Reserve of Donana National Park (Spain). Rend Fis Acc Lincei9(18):1–17
Testi A, De Nicola C, Guidotti S, Serafini-Sauli A, Fanelli G, Pignatti S (2006b) Vegetation ecology ofCastelporziano woodlands. In: Il Sistema Ambientale della Tenuta Presidenziale di Castelporziano.Ricerche sulla complessita di un ecosistema forestale costiero mediterraneo. Accademia delle Scienze,Scritti e Documenti XXXVII, Seconda Serie, vol II, pp 565–605
USDA Natural Resources Conservation Service (1996) Soil taxonomy. A basic system of soil classificationfor making and interpreting soil surveys. Agriculture handbook, no 436, 2nd edn, p 87
USDA Natural Resources Conservation Service (1999) Soil taxonomy. A basic system of soil classificationfor making and interpreting soil surveys. Agriculture handbook, no 436, 2nd edn, p 87
USDA Natural Resources Conservation Service (2006) Keys to soil taxonomy, 10th edn, p 341Van der Maarel E (1979) Transformation of cover-abundance values in phytosociology and its effects on
community similarity. Vegetation 39:97–114Vesterdal L, Schmidt IK, Callesen I, Nilsson LO, Gundersen P (2008) Carbon and nitrogen in forest floor
and mineral soil under six common European tree species. For Ecol Manage 255:35–48White PS, Jentsch A (2001) The search for generality in studies of disturbance and ecosystem dynamics.
Prog Bot 62:399–449Wilson McG, Pyatt DG, Malcolm DC, Connolly T (2001) The use of ground vegetation and humus type as
indicators of soil nutrient regime for an ecological site classification of British forests. For EcolManage 140:101–116
Rend. Fis. Acc. Lincei
123
Related Documents











