Correlations of Regional Cerebral Metabolism With Memory Performance and Executive Function in Patients With Herpes Encephalitis or Frontal Lobe Lesions Laurence J. Reed, Daniel Lasserson, Paul Marsden, Peter Bright, Nicola Stanhope, and Michael D. Kopelman King’s College London, Institute of Psychiatry Cerebral [ 18 F]fluorodeoxyglucose positron emission tomography ( 18 FDG-PET) data from patients suf- fering amnesia following herpes encephalitis (n 7) or frontal lobe pathology (n 14) were compared with data from age-matched nonamnesic subjects (n 10). All subjects received structural MRI, resting 18 FDG-PET scans, and neuropsychological evaluation. PET data were analyzed using complementary statistical parametric mapping and region-of-interest methods. Differential patterns of hypometabolism were found in patients relative to healthy controls. Factor analysis of the neuropsychological data revealed that memory performance was associated with retrosplenial and medial temporal metabolism, and executive function was associated with dorsolateral frontal metabolism. The association between memory performance and retrosplenial metabolism remained statistically significant after accounting for measures of cerebral atrophy using MRI. The significance of the retrosplenium as a major relay station between the thalamus and the medial temporal and frontal lobes—sensitive to changes in either—is discussed in the light of the findings. In both the neuropsychological (lesion) and neuroimaging (functional activation) literatures, there is considerable debate re- garding the respective contributions of the medial temporal struc- tures and the frontal lobes in memory formation (Kopelman, 2002). Whereas lesion studies had pointed to a direct role of the medial temporal lobes in memory “consolidation” and of the frontal lobes in the more “executive” aspects of memory, early functional imaging studies highlighted the frequency with which the frontal lobes were activated during memory encoding and retrieval (Grasby et al., 1993; Kapur et al., 1994; Shallice et al., 1994). Moreover, subsequent lesion studies have drawn attention to the direct effects of frontal lesions on various aspects of memory processing (Kopelman & Stanhope, 1997, 1998; Wheeler, Stuss, Tulving, 1995). One question that follows from the overlap in the effects of frontal and medial temporal lesions on memory pro- cesses concerns whether these result from distal metabolic effects on the medial temporal lobes (in the case of large frontal lesions) or on the frontal lobes (in the case of medial temporal lesions). The present article addresses this issue using [ 18 F]fluorodeoxyglucose positron emission tomography ( 18 FDG-PET) measures of metab- olism, and it examines the degree of association between medial temporal and frontal metabolism and memory performance and executive function, respectively. Studies of “resting” 18 FDG-PET in patients are potentially im- portant in a number of different ways: First, they detect the overall location and extent of any abnormalities in glucose metabolism arising directly from cerebral pathology. Second, they may reveal sites of altered function that are distant from the primary structural pathology (Fazio et al., 1992; Kapur, Thompson, Kartsounis, & Abbot, 1999; Kuhl et al., 1980). Third, the current interest in the neuroimaging of memory activation in healthy subjects (Kopel- man, Stevens, Foli, & Grasby, 1998; Maguire, 2001a; Montaldi et al., 1998; Rugg, 2002; Schacter, Alpert, Savage, Rauch, & Albert, 1996) suggests that abnormalities in baseline or “resting” metab- olism should ideally be specified before such techniques are ap- plied to patient groups, especially where group-mapping methods are applied. In particular, 18 FDG-PET is cerebral-blood-flow in- sensitive (Phelps et al., 1979) and, hence, not susceptible to the confounding factors of cerebrovascular disease and drug exposure, unlike methods used in activation studies. In previous 18 FDG-PET studies, differing techniques of analysis (assumptions, statistics, inference) have been used, and any inconsistencies and discrepan- cies between these different methods need to be clearly identified (Reed et al., 1999). Patients with severe memory disorder resulting from acute hy- poxic episodes have been investigated in a series of studies. Kuwert et al. (1993) reported thalamic hypometabolism in 3 pa- tients, medial temporal hypometabolism in 2 patients, and involve- ment of the caudate nucleus and cerebellum in 1 case each. Markowitsch, Weber-Luxenberger, Ewald, Kessler, and Heiss (1997) found altered metabolism in the thalami, in the medial and lateral temporal lobes, and in the occipitoparietal regions bilater- ally in a case of cardiac arrest in which MRI revealed atrophy without “specific brain damage.” Reed et al.’s (1999) study of 4 Laurence J. Reed, Division of Psychological Medicine, Institute of Psychiatry, King’s College London, United Kingdom; Clinical PET Cen- tre, Guy’s, King’s and St Thomas’s School of Medicine, St Thomas’s Hospital, King’s College London; Brain Image Analysis Unit, Department of Psychological Medicine, Institute of Psychiatry, King’s College London. Daniel Lasserson, Peter Bright, Nicola Stanhope, and Michael D. Kopel- man, Division of Psychiatry and Psychology, St Thomas’s Hospital, King’s College London. Paul Marsden, Clinical PET Centre, Guy’s, King’s and St Thomas’s School of Medicine, St Thomas’s Hospital, King’s College London. This research was funded by a Wellcome project grant to Michael D. Kopelman. Correspondence concerning this article should be addressed to Laurence J. Reed, Division of Psychological Medicine, Maudsley Hospital, Denmark Hill London SE5 8AF, United Kingdom, or Michael D. Kopelman, Uni- versity Department of Psychiatry and Psychology, 3rd Floor, Block 8 South Wing, St Thomas’s Hospital, London SE1 7EH, United Kingdom. E-mail: [email protected] or [email protected] Neuropsychology Copyright 2005 by the American Psychological Association 2005, Vol. 19, No. 5, 555–565 0894-4105/05/$12.00 DOI: 10.1037/0894-4105.19.5.555 555

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Correlations of Regional Cerebral Metabolism With Memory Performanceand Executive Function in Patients With Herpes Encephalitis or Frontal
Lobe Lesions
Laurence J. Reed, Daniel Lasserson, Paul Marsden, Peter Bright, Nicola Stanhope, and Michael D. KopelmanKing’s College London, Institute of Psychiatry
Cerebral [18F]fluorodeoxyglucose positron emission tomography (18FDG-PET) data from patients suf-fering amnesia following herpes encephalitis (n � 7) or frontal lobe pathology (n � 14) were comparedwith data from age-matched nonamnesic subjects (n � 10). All subjects received structural MRI, resting18FDG-PET scans, and neuropsychological evaluation. PET data were analyzed using complementarystatistical parametric mapping and region-of-interest methods. Differential patterns of hypometabolismwere found in patients relative to healthy controls. Factor analysis of the neuropsychological datarevealed that memory performance was associated with retrosplenial and medial temporal metabolism,and executive function was associated with dorsolateral frontal metabolism. The association betweenmemory performance and retrosplenial metabolism remained statistically significant after accounting formeasures of cerebral atrophy using MRI. The significance of the retrosplenium as a major relay stationbetween the thalamus and the medial temporal and frontal lobes—sensitive to changes in either—isdiscussed in the light of the findings.
In both the neuropsychological (lesion) and neuroimaging(functional activation) literatures, there is considerable debate re-garding the respective contributions of the medial temporal struc-tures and the frontal lobes in memory formation (Kopelman,2002). Whereas lesion studies had pointed to a direct role of themedial temporal lobes in memory “consolidation” and of thefrontal lobes in the more “executive” aspects of memory, earlyfunctional imaging studies highlighted the frequency with whichthe frontal lobes were activated during memory encoding andretrieval (Grasby et al., 1993; Kapur et al., 1994; Shallice et al.,1994). Moreover, subsequent lesion studies have drawn attentionto the direct effects of frontal lesions on various aspects of memoryprocessing (Kopelman & Stanhope, 1997, 1998; Wheeler, Stuss,Tulving, 1995). One question that follows from the overlap in theeffects of frontal and medial temporal lesions on memory pro-cesses concerns whether these result from distal metabolic effectson the medial temporal lobes (in the case of large frontal lesions)or on the frontal lobes (in the case of medial temporal lesions). The
present article addresses this issue using [18F]fluorodeoxyglucosepositron emission tomography (18FDG-PET) measures of metab-olism, and it examines the degree of association between medialtemporal and frontal metabolism and memory performance andexecutive function, respectively.
Studies of “resting” 18FDG-PET in patients are potentially im-portant in a number of different ways: First, they detect the overalllocation and extent of any abnormalities in glucose metabolismarising directly from cerebral pathology. Second, they may revealsites of altered function that are distant from the primary structuralpathology (Fazio et al., 1992; Kapur, Thompson, Kartsounis, &Abbot, 1999; Kuhl et al., 1980). Third, the current interest in theneuroimaging of memory activation in healthy subjects (Kopel-man, Stevens, Foli, & Grasby, 1998; Maguire, 2001a; Montaldi etal., 1998; Rugg, 2002; Schacter, Alpert, Savage, Rauch, & Albert,1996) suggests that abnormalities in baseline or “resting” metab-olism should ideally be specified before such techniques are ap-plied to patient groups, especially where group-mapping methodsare applied. In particular, 18FDG-PET is cerebral-blood-flow in-sensitive (Phelps et al., 1979) and, hence, not susceptible to theconfounding factors of cerebrovascular disease and drug exposure,unlike methods used in activation studies. In previous 18FDG-PETstudies, differing techniques of analysis (assumptions, statistics,inference) have been used, and any inconsistencies and discrepan-cies between these different methods need to be clearly identified(Reed et al., 1999).
Patients with severe memory disorder resulting from acute hy-poxic episodes have been investigated in a series of studies.Kuwert et al. (1993) reported thalamic hypometabolism in 3 pa-tients, medial temporal hypometabolism in 2 patients, and involve-ment of the caudate nucleus and cerebellum in 1 case each.Markowitsch, Weber-Luxenberger, Ewald, Kessler, and Heiss(1997) found altered metabolism in the thalami, in the medial andlateral temporal lobes, and in the occipitoparietal regions bilater-ally in a case of cardiac arrest in which MRI revealed atrophywithout “specific brain damage.” Reed et al.’s (1999) study of 4
Laurence J. Reed, Division of Psychological Medicine, Institute ofPsychiatry, King’s College London, United Kingdom; Clinical PET Cen-tre, Guy’s, King’s and St Thomas’s School of Medicine, St Thomas’sHospital, King’s College London; Brain Image Analysis Unit, Departmentof Psychological Medicine, Institute of Psychiatry, King’s College London.Daniel Lasserson, Peter Bright, Nicola Stanhope, and Michael D. Kopel-man, Division of Psychiatry and Psychology, St Thomas’s Hospital, King’sCollege London. Paul Marsden, Clinical PET Centre, Guy’s, King’s and StThomas’s School of Medicine, St Thomas’s Hospital, King’s CollegeLondon.
This research was funded by a Wellcome project grant to Michael D.Kopelman.
Correspondence concerning this article should be addressed to LaurenceJ. Reed, Division of Psychological Medicine, Maudsley Hospital, DenmarkHill London SE5 8AF, United Kingdom, or Michael D. Kopelman, Uni-versity Department of Psychiatry and Psychology, 3rd Floor, Block 8South Wing, St Thomas’s Hospital, London SE1 7EH, United Kingdom.E-mail: [email protected] or [email protected]
Neuropsychology Copyright 2005 by the American Psychological Association2005, Vol. 19, No. 5, 555–565 0894-4105/05/$12.00 DOI: 10.1037/0894-4105.19.5.555
555

patients who had suffered severe amnesia following episodes ofacute hypoxia revealed bilateral thalamic and retrosplenial hypo-metabolism, associated with evidence of medial temporal lobe(hippocampal and parahippocampal) atrophy on MRI brain scans.These findings were interpreted as indicating that structural lesionscan indeed have distal metabolic effects on brain structures else-where. Somewhat similar but more severe abnormalities werefound in a profoundly amnesic patient following an overdose ofMDMA (ecstasy): In this case, the thalami and retrosplenium wereinvolved bilaterally as well as the left medial temporal region(Kopelman, Reed et al., 2001).
There have also been 18FDG-PET studies of patients whosechronic memory disorder resulted from the alcoholic Korsakoffsyndrome. Paller et al. (1997) found widely distributed cerebralhypometabolism in the absence of thalamic or medial temporalchange. Martin et al. (1992) found little difference between Kor-sakoff patients and controls, and Joyce et al. (1994) reportedhypometabolism in the anterior cingulate and medial temporallobes. In a recent study by Reed et al. (2003), Korsakoff patientswere shown to have white matter hypermetabolism and relativehypometabolism in diencephalic and retrosplenial gray matter,with a variable degree of frontotemporal hypometabolism acrossindividual participants.
Fazio et al. (1992) proposed that in organic amnesia, there is asignificant reduction of metabolism throughout limbic– dien-cephalic circuits on FDG-PET, whatever the site of the primarystructural pathology. This proposal was based on findings from 11amnesic patients with a variety of underlying pathologies. How-ever, Fazio et al. did not report the individual findings from theirdifferent patient groups, raising concerns as to the generalizabilityof their findings (but see Signorini et al., 1999). Aupee et al. (2001)used voxel-based mapping methods to compare FDG-PET mea-sures of metabolism in 3 Korsakoff patients and 2 hypoxic pa-tients. In this group as a whole, the authors found bilateral hypo-metabolism in the thalamus, the posterior cingulate, and the mesialprefrontal cortex (near the anterior cingulate gyrus) as well as inthe left supramarginal and middle temporal gyri. This overallpattern was also seen in individual patients, although there weresome minor differences.
The contribution of the retrosplenial cortex to memory forma-tion, seldom recognized in the lesion literature (e.g., Rudge &Warrington, 1991; Valenstein et al., 1987), has much more fre-quently been identified within recent functional-imaging/neuro-anatomical-mapping investigations. Studies of the trajectories ofcholinergic pathways arising from the nucleus basalis show adense innervation of the retrosplenial cortex by the medial fiberbundle, strongly implicated in memory processing (Selden, Gitel-man, Salamon-Murayama, Parrish, & Mesulam, 1998). More gen-erally, Morris, Petrides, and Pandya (1999) used anterograde andretrograde tracers to demonstrate close connectivity between theretrosplenial cortex and the medial temporal lobes and dorsolateralprefrontal cortex. In functional-imaging studies, Kennedy et al.(1995) reported impairments in retrosplenial cortical metabolismin familial Alzheimer dementia, and more recently, Desgranges etal. (2002) reported retrosplenial cortical hypometabolism that cor-related with recall test scores in early Alzheimer dementia. Workin our own group has shown retrosplenial hypometabolism inamnesia arising from anoxia (Reed et al., 1999), MDMA toxicity(Kopelman, Reed, et al., 2001), and the Korsakoff syndrome (Reedet al., 2003). Taken together, these reports suggest that hypome-
tabolism in this region is commonly associated with memoryimpairments, whatever the underlying etiology.
In the present article, we report findings in two more groups ofmemory-disordered patients: those with herpes encephalitis (withstructural pathology affecting the temporal lobes, as demonstratedon MRI) and those with large frontal lobe lesions (as delineated onstructural MRI). In particular, we examine the pattern of associa-tion between our FDG-PET findings and measures of memory andexecutive test function, using factor analyses from a series ofcognitive tests. Both region-of-interest (ROI) and voxel-basedstatistical parametric mapping (SPM) analyses were used becausetheir differing assumptions make it important to identify conver-gent or discrepant findings (Desgranges et al., 1998; Kennedy etal., 1995; Reed et al., 1999, 2003). In particular, the large lesionspresent in this patient group lead to significant distortions incerebral anatomy that may interfere with transformation into stan-dard reference space used in SPM: Hence, ROI-based analysisprovides a means to address this potential artifact.
The main purposes of this study were
1. to examine the pattern of metabolic impairment in thecase of the large lesions seen in the herpes encephalitisand frontal lesion groups and to investigate whether thereare indeed common patterns of abnormal metabolism,possibly involving brain regions distal to the principalsites of lesion within these groups.
2. More fundamentally, we examined the regional meta-bolic correlates of anterograde memory impairment andexecutive dysfunction in these memory-disordered pa-tients. Our a priori hypotheses were (a) that anterogradememory impairment would be reflected in impaired met-abolic activity in the memory circuits—specifically in-cluding the medial temporal lobes, retrosplenium, andthalamus—and that (b) executive dysfunction would beassociated with impaired metabolic activity in frontal,particularly dorsolateral, cortical structures.
3. Lastly, we examined whether the relationships observedcould be accounted for by cerebral atrophy using mea-sures derived from quantitative MRI.
Method
Subject Groups
The samples from which these patients were recruited have been de-scribed in greater detail elsewhere (Kopleman, Lasserson, et al., 2001;Kopelman, Stanhope, & Kingsley, 1999). There were 8 patients who wereassessed 2 weeks following a bilateral frontal tractotomy. In this operation,yttrium rods were implanted in the frontal lobes for treatment of chronicaffective disorders. On MRI scans, the yttrium rods could be seen in themedial and midfrontal white matter, surrounded by a small area of radiationnecrosis and a much larger area of edema. These patients were assessedafter any confusion had subsided, usually in the 2nd or 3rd postoperativeweek, on the basis of the observations of Kartsounis, Poynton, Bridges, andBartlett (1991) that these patients behaved most like patients with largefrontal lesions at this time. Figures 1A and 1B show the PET changesobserved in comparison with control subjects; these changes were highlyconsistent across individual subjects in this subgroup. Kartsounis et al.demonstrated statistically significant impairments on executive–frontaltests 2 weeks postoperatively, although those deficits were not presenteither preoperatively (despite severe depression) or 6 months later.
There were 6 patients with focal frontal lesions. One patient had oldleucotomy scars from a bilateral operation 18 years before testing, and the
556 REED ET AL.
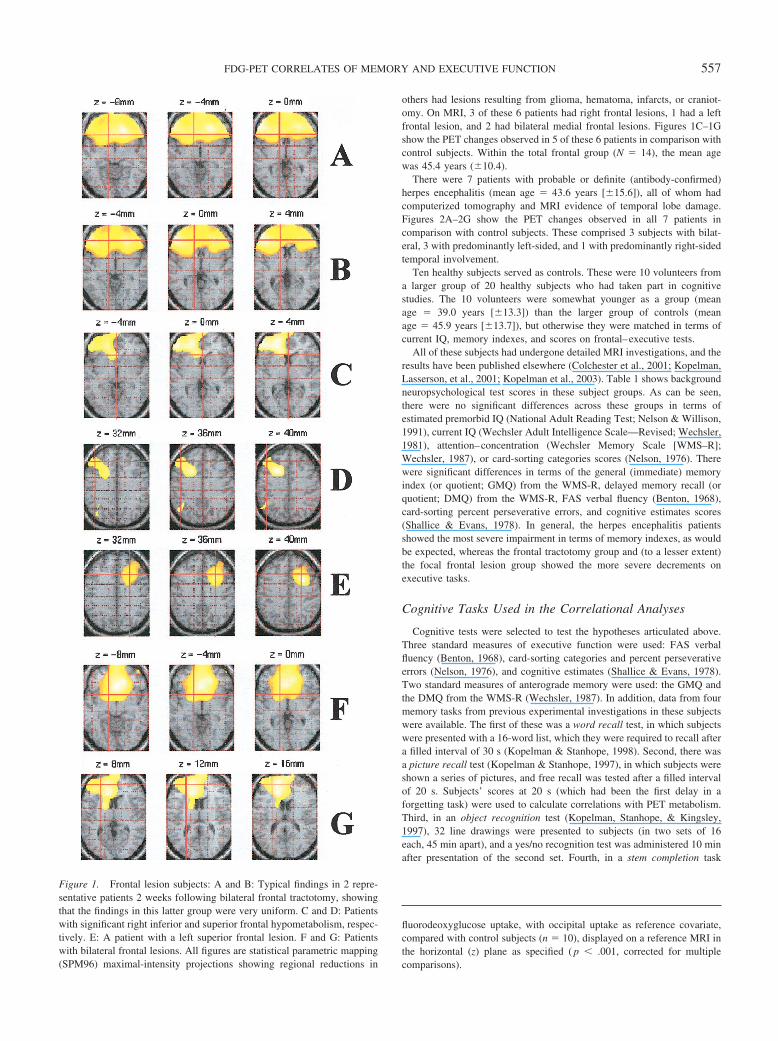
others had lesions resulting from glioma, hematoma, infarcts, or craniot-omy. On MRI, 3 of these 6 patients had right frontal lesions, 1 had a leftfrontal lesion, and 2 had bilateral medial frontal lesions. Figures 1C–1Gshow the PET changes observed in 5 of these 6 patients in comparison withcontrol subjects. Within the total frontal group (N � 14), the mean agewas 45.4 years (�10.4).
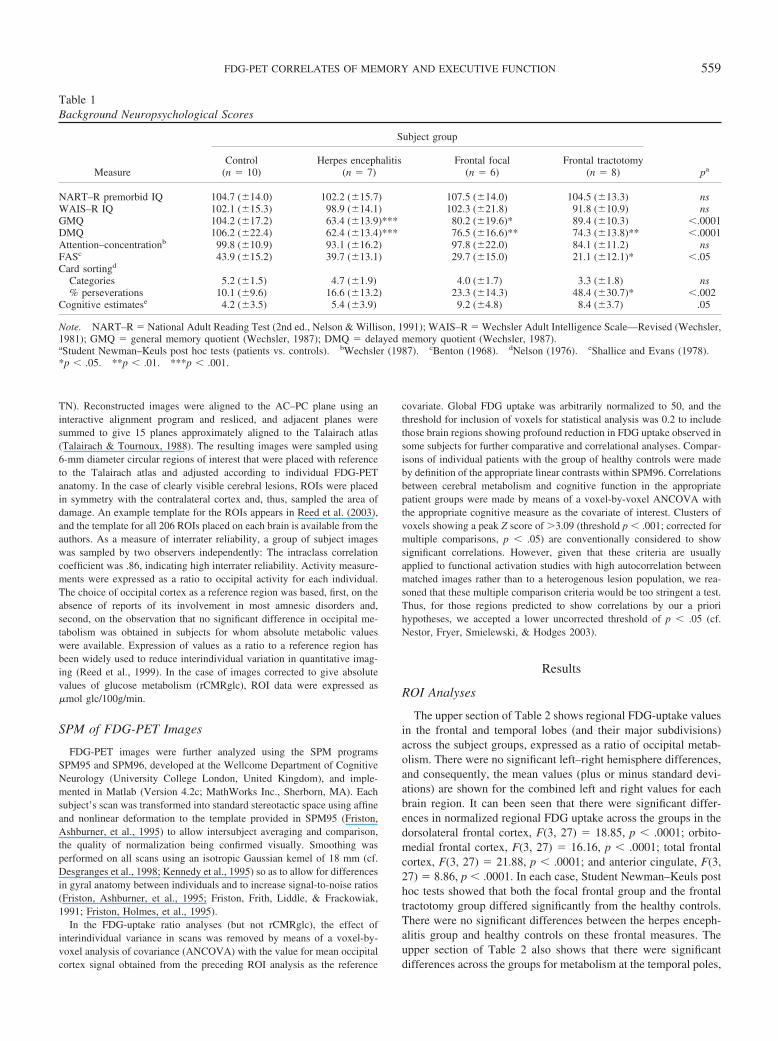
There were 7 patients with probable or definite (antibody-confirmed)herpes encephalitis (mean age � 43.6 years [�15.6]), all of whom hadcomputerized tomography and MRI evidence of temporal lobe damage.Figures 2A–2G show the PET changes observed in all 7 patients incomparison with control subjects. These comprised 3 subjects with bilat-eral, 3 with predominantly left-sided, and 1 with predominantly right-sidedtemporal involvement.
Ten healthy subjects served as controls. These were 10 volunteers froma larger group of 20 healthy subjects who had taken part in cognitivestudies. The 10 volunteers were somewhat younger as a group (meanage � 39.0 years [�13.3]) than the larger group of controls (meanage � 45.9 years [�13.7]), but otherwise they were matched in terms ofcurrent IQ, memory indexes, and scores on frontal–executive tests.
All of these subjects had undergone detailed MRI investigations, and theresults have been published elsewhere (Colchester et al., 2001; Kopelman,Lasserson, et al., 2001; Kopelman et al., 2003). Table 1 shows backgroundneuropsychological test scores in these subject groups. As can be seen,there were no significant differences across these groups in terms ofestimated premorbid IQ (National Adult Reading Test; Nelson & Willison,1991), current IQ (Wechsler Adult Intelligence Scale—Revised; Wechsler,1981), attention–concentration (Wechsler Memory Scale [WMS–R];Wechsler, 1987), or card-sorting categories scores (Nelson, 1976). Therewere significant differences in terms of the general (immediate) memoryindex (or quotient; GMQ) from the WMS-R, delayed memory recall (orquotient; DMQ) from the WMS-R, FAS verbal fluency (Benton, 1968),card-sorting percent perseverative errors, and cognitive estimates scores(Shallice & Evans, 1978). In general, the herpes encephalitis patientsshowed the most severe impairment in terms of memory indexes, as wouldbe expected, whereas the frontal tractotomy group and (to a lesser extent)the focal frontal lesion group showed the more severe decrements onexecutive tasks.
Cognitive Tasks Used in the Correlational Analyses
Cognitive tests were selected to test the hypotheses articulated above.Three standard measures of executive function were used: FAS verbalfluency (Benton, 1968), card-sorting categories and percent perseverativeerrors (Nelson, 1976), and cognitive estimates (Shallice & Evans, 1978).Two standard measures of anterograde memory were used: the GMQ andthe DMQ from the WMS-R (Wechsler, 1987). In addition, data from fourmemory tasks from previous experimental investigations in these subjectswere available. The first of these was a word recall test, in which subjectswere presented with a 16-word list, which they were required to recall aftera filled interval of 30 s (Kopelman & Stanhope, 1998). Second, there wasa picture recall test (Kopelman & Stanhope, 1997), in which subjects wereshown a series of pictures, and free recall was tested after a filled intervalof 20 s. Subjects’ scores at 20 s (which had been the first delay in aforgetting task) were used to calculate correlations with PET metabolism.Third, in an object recognition test (Kopelman, Stanhope, & Kingsley,1997), 32 line drawings were presented to subjects (in two sets of 16each, 45 min apart), and a yes/no recognition test was administered 10 minafter presentation of the second set. Fourth, in a stem completion task
Figure 1. Frontal lesion subjects: A and B: Typical findings in 2 repre-sentative patients 2 weeks following bilateral frontal tractotomy, showingthat the findings in this latter group were very uniform. C and D: Patientswith significant right inferior and superior frontal hypometabolism, respec-tively. E: A patient with a left superior frontal lesion. F and G: Patientswith bilateral frontal lesions. All figures are statistical parametric mapping(SPM96) maximal-intensity projections showing regional reductions in
fluorodeoxyglucose uptake, with occipital uptake as reference covariate,compared with control subjects (n � 10), displayed on a reference MRI inthe horizontal (z) plane as specified ( p � .001, corrected for multiplecomparisons).
557FDG-PET CORRELATES OF MEMORY AND EXECUTIVE FUNCTION

(Kopelman & Stanhope, 1997), subjects were presented with lists of 17words (12 target words and 5 buffer items). At a 1-min delay, subjects werepresented with 12 three-letter word stems and given, first, an inclusioninstruction (to use the stems to recall the words presented) and then anexclusion instruction (to complete the stems with words other than thosepreviously presented). From these results, a recollection score (explicitmemory) and an automaticity score (implicit memory) were calculatedusing Jacoby and Kelley’s (1992) formula. All cognitive measures weresubject to z transformation before principal components analysis withvarimax rotation, using the Statistical Package for Social Sciences forWindows (Version 10).
Acquisition of FDG-PET Data
Measurements of cerebral metabolic rate for glucose (rCMRglc) wereperformed using the tracer 18FDG (Huang et al., 1980; Phelps et al., 1979)using a CTI ECAT 951R PET camera (CTI/Siemens, Knoxville, TN). Thefield of view was 10.8 cm and centered on the cerebral areas of interestcomprising frontal and temporal lobes and the diencephalon. In mostsubjects, this resulted in the loss of superior parietal and inferior cerebellarcortex. Each reconstructed image was displayed in a matrix of 128 �128 � 31 voxel format, each voxel measuring 2.09 � 2.09 � 3.43 mm;image resolution was 8.5 � 8.5 � 5.5 mm full-width at half-maximum(FWHM).
The subject’s head was placed in the scanner with a supportive headrestto minimize head movement, aligned axially and to the orbitomeatal line(approximating to the AC–PC line), which was checked at intervals duringimage acquisition and corrected if drift was identified. Subjects werescanned lying supine in a darkened room with their eyes closed. Subjectswere fitted with an intravenous cannula into the right antecubital vein,allowing injection of approximately 185 MBq FDG as a slow bolus over30 s. Thirty min after injection, images were acquired over a 30-minperiod. Nine out of 10 controls, 5 out of 7 herpes patients, 5 out 6 focalfrontal patients, and 5 out of 8 frontal tractotomy patients were agreeableto and tolerated arterialized-venous sampling, allowing correction of datato true metabolic rate or rCMRglc using the autoradiographic method(Huang et al., 1980; Phelps et al., 1979).
Strategy for Analysis of FDG-PET Data
Two complementary approaches (ROI, SPM) were adopted for analysisof the FDG-PET images. ROI analysis was performed on the anatomicalFDG-PET images both to derive measures necessary for occipital corticalnormalization and to serve as a check on the validity of the spatialnormalization routines used in voxel-based parametric methods (SPM).Although such spatial normalization routines are customarily accepted asvalid in healthy subject and some patient groups, the asymmetrical natureand large volume of the lesions present in our patient groups made itrelevant to cross-validate the SPM findings using ROI, as has been donepreviously in a minority of studies (Desgranges et al., 1998; Kennedy et al.,1995; Reed et al., 1999, 2003).
ROI Analysis of FDG-PET Data
All image transformations and analyses were performed on a SunSPARC workstation (Sun Computers Europe Inc., Surrey, United King-dom) using proprietary image-analysis software (CTI/Siemens, Knoxville,
Figure 2. Temporal lesion subjects: The findings in patients with herpesencephalitis, illustrating the highly variable pattern of temporal lobe in-volvement, with bilateral temporal lobe hypometabolism in some andunilateral left or right hypometabolism in others. All figures are statistical
parametric mapping (SPM96) maximal-intensity projections showing re-gional reductions in fluorodeoxyglucose uptake, with occipital uptake asreference covariate, compared with control subjects (n � 10), displayed ona reference MRI in the horizontal (z) plane as specified ( p � .001,corrected for multiple comparisons).
558 REED ET AL.
TN). Reconstructed images were aligned to the AC–PC plane using aninteractive alignment program and resliced, and adjacent planes weresummed to give 15 planes approximately aligned to the Talairach atlas(Talairach & Tournoux, 1988). The resulting images were sampled using6-mm diameter circular regions of interest that were placed with referenceto the Talairach atlas and adjusted according to individual FDG-PETanatomy. In the case of clearly visible cerebral lesions, ROIs were placedin symmetry with the contralateral cortex and, thus, sampled the area ofdamage. An example template for the ROIs appears in Reed et al. (2003),and the template for all 206 ROIs placed on each brain is available from theauthors. As a measure of interrater reliability, a group of subject imageswas sampled by two observers independently: The intraclass correlationcoefficient was .86, indicating high interrater reliability. Activity measure-ments were expressed as a ratio to occipital activity for each individual.The choice of occipital cortex as a reference region was based, first, on theabsence of reports of its involvement in most amnesic disorders and,second, on the observation that no significant difference in occipital me-tabolism was obtained in subjects for whom absolute metabolic valueswere available. Expression of values as a ratio to a reference region hasbeen widely used to reduce interindividual variation in quantitative imag-ing (Reed et al., 1999). In the case of images corrected to give absolutevalues of glucose metabolism (rCMRglc), ROI data were expressed as�mol glc/100g/min.
SPM of FDG-PET Images
FDG-PET images were further analyzed using the SPM programsSPM95 and SPM96, developed at the Wellcome Department of CognitiveNeurology (University College London, United Kingdom), and imple-mented in Matlab (Version 4.2c; MathWorks Inc., Sherborn, MA). Eachsubject’s scan was transformed into standard stereotactic space using affineand nonlinear deformation to the template provided in SPM95 (Friston,Ashburner, et al., 1995) to allow intersubject averaging and comparison,the quality of normalization being confirmed visually. Smoothing wasperformed on all scans using an isotropic Gaussian kemel of 18 mm (cf.Desgranges et al., 1998; Kennedy et al., 1995) so as to allow for differencesin gyral anatomy between individuals and to increase signal-to-noise ratios(Friston, Ashburner, et al., 1995; Friston, Frith, Liddle, & Frackowiak,1991; Friston, Holmes, et al., 1995).
In the FDG-uptake ratio analyses (but not rCMRglc), the effect ofinterindividual variance in scans was removed by means of a voxel-by-voxel analysis of covariance (ANCOVA) with the value for mean occipitalcortex signal obtained from the preceding ROI analysis as the reference
covariate. Global FDG uptake was arbitrarily normalized to 50, and thethreshold for inclusion of voxels for statistical analysis was 0.2 to includethose brain regions showing profound reduction in FDG uptake observed insome subjects for further comparative and correlational analyses. Compar-isons of individual patients with the group of healthy controls were madeby definition of the appropriate linear contrasts within SPM96. Correlationsbetween cerebral metabolism and cognitive function in the appropriatepatient groups were made by means of a voxel-by-voxel ANCOVA withthe appropriate cognitive measure as the covariate of interest. Clusters ofvoxels showing a peak Z score of �3.09 (threshold p � .001; corrected formultiple comparisons, p � .05) are conventionally considered to showsignificant correlations. However, given that these criteria are usuallyapplied to functional activation studies with high autocorrelation betweenmatched images rather than to a heterogenous lesion population, we rea-soned that these multiple comparison criteria would be too stringent a test.Thus, for those regions predicted to show correlations by our a priorihypotheses, we accepted a lower uncorrected threshold of p � .05 (cf.Nestor, Fryer, Smielewski, & Hodges 2003).
Results
ROI Analyses
The upper section of Table 2 shows regional FDG-uptake valuesin the frontal and temporal lobes (and their major subdivisions)across the subject groups, expressed as a ratio of occipital metab-olism. There were no significant left–right hemisphere differences,and consequently, the mean values (plus or minus standard devi-ations) are shown for the combined left and right values for eachbrain region. It can been seen that there were significant differ-ences in normalized regional FDG uptake across the groups in thedorsolateral frontal cortex, F(3, 27) � 18.85, p � .0001; orbito-medial frontal cortex, F(3, 27) � 16.16, p � .0001; total frontalcortex, F(3, 27) � 21.88, p � .0001; and anterior cingulate, F(3,27) � 8.86, p � .0001. In each case, Student Newman–Keuls posthoc tests showed that both the focal frontal group and the frontaltractotomy group differed significantly from the healthy controls.There were no significant differences between the herpes enceph-alitis group and healthy controls on these frontal measures. Theupper section of Table 2 also shows that there were significantdifferences across the groups for metabolism at the temporal poles,
Table 1Background Neuropsychological Scores
Measure
Subject group
paControl
(n � 10)Herpes encephalitis
(n � 7)Frontal focal
(n � 6)Frontal tractotomy
(n � 8)
NART–R premorbid IQ 104.7 (�14.0) 102.2 (�15.7) 107.5 (�14.0) 104.5 (�13.3) nsWAIS–R IQ 102.1 (�15.3) 98.9 (�14.1) 102.3 (�21.8) 91.8 (�10.9) nsGMQ 104.2 (�17.2) 63.4 (�13.9)*** 80.2 (�19.6)* 89.4 (�10.3) �.0001DMQ 106.2 (�22.4) 62.4 (�13.4)*** 76.5 (�16.6)** 74.3 (�13.8)** �.0001Attention–concentrationb 99.8 (�10.9) 93.1 (�16.2) 97.8 (�22.0) 84.1 (�11.2) nsFASc 43.9 (�15.2) 39.7 (�13.1) 29.7 (�15.0) 21.1 (�12.1)* �.05Card sortingd
Categories 5.2 (�1.5) 4.7 (�1.9) 4.0 (�1.7) 3.3 (�1.8) ns% perseverations 10.1 (�9.6) 16.6 (�13.2) 23.3 (�14.3) 48.4 (�30.7)* �.002
Cognitive estimatese 4.2 (�3.5) 5.4 (�3.9) 9.2 (�4.8) 8.4 (�3.7) .05
Note. NART–R � National Adult Reading Test (2nd ed., Nelson & Willison, 1991); WAIS–R � Wechsler Adult Intelligence Scale—Revised (Wechsler,1981); GMQ � general memory quotient (Wechsler, 1987); DMQ � delayed memory quotient (Wechsler, 1987).aStudent Newman–Keuls post hoc tests (patients vs. controls). bWechsler (1987). cBenton (1968). dNelson (1976). eShallice and Evans (1978).*p � .05. **p � .01. ***p � .001.
559FDG-PET CORRELATES OF MEMORY AND EXECUTIVE FUNCTION

F(3, 27) � 9.15, p � .0001; the lateral temporal cortex, F(3,27) � 3.54, p � .05; the medial temporal lobes, F(3, 27) � 14.96,p � .0001; and total temporal lobe metabolism, F(3, 27) � 7.21,p � .001. In each of these cases, the post hoc test indicated that theherpes encephalitis group showed significant hypometabolism rel-ative to the healthy controls, whereas the two frontal groups didnot differ significantly from controls. In contrast, there were nosignificant differences between the groups in the inferior temporallobe or in the anterior, posterior, inferior, or total thalamus; retro-splenium; parietal cortex; caudate; putamen; or cerebellum.
The lower section of Table 2 shows the findings from thosepatients in whom measures of absolute glucose metabolism(rCMRglc) values were obtained. In broad terms, the findings wereconsistent with the findings for FDG-uptake ratios in the largergroup, except that the findings in the dorsolateral frontal and lateraltemporal regions were no longer statistically significant, whereasthe herpes encephalitis group now showed significantly reducedmetabolism in the inferior temporal region. There were no signif-icant differences across the groups in other brain regions on thisanalysis. It should be noted that the coefficient of variance wasapproximately 20% in measures of rCMRglc compared with co-efficients of about 5% when expressed relative to occipital metab-olism, emphasizing the sensitivity of the latter measure.
SPM Analysis
Figure 1 shows frontal hypometabolism in typical patients withfrontal lobe lesions. Figures 1A and 1B show the typical findings
in 2 patients 2 weeks following bilateral frontal tractotomy: Thefindings in this group were very uniform and involved extensiveregions of the orbitomedial, anterior cingulate, and dorsolateralprefrontal cortices. Figures 1C–1G show findings from 5 focalfrontal patients, indicating the variety of regional frontal corticalinvolvement. Figure 2 shows the findings in all 7 patients withherpes encephalitis, showing bilateral temporal lobe hypometabo-lism in 3 cases, unilateral left hypometabolism in 3 others, andunilateral right medial temporal involvement in the last.
FDG-PET Cognitive Correlations
To avoid the problems of multiple comparisons across a largenumber of individual cognitive tests and regional FDG-PET mea-surements, we used findings from a factor analysis of our cognitivetests, which had been carried out in the total sample of patients inthe context of our earlier neuropsychological investigations (Ko-pelman & Stanhope, 1997, 1998; Kopelman et al., 1997). Table 3shows the findings from this factor analysis, in which three mainfactors emerged. The first factor, accounting for 37.2% of the totalvariance, appeared to be a memory factor, with high loadings ofGMQ, DMQ, word recall, picture recall, and picture recognitionand recollection and a negative loading of the automaticity score.The second factor, accounting for 27.1% of the variance, appearedto be a marker of executive function, with negative loadings ofFAS verbal fluency and card-sorting categories and positive load-ings of card-sorting perseverations and cognitive estimates scores.
Table 2Regional FDG-PET Findings in Herpes Encephalitis and Frontal Lesion Patients
Region
Subject group
pControl
(n � 10)Herpes encephalitis
(n � 7)Frontal focal
(n � 6)Frontal tractotomy
(n � 8)
Regional FDG-uptake values relative to occipital metabolismFrontal
Dorsolateral frontal 1.02 (�0.06) 0.99 (�0.08) 0.87 (�0.07)** 0.76 (�0.12)*** �.0001Orbitomedial frontal 0.91 (�0.06) 0.82 (�0.13) 0.74 (�0.19)* 0.51 (�0.11)*** �.0001Anterior cingulate 1.01 (�0.07) 0.93 (�0.09) 0.72 (�0.21)** 0.76 (�0.13)** �.0001Total 1.00 (�0.05) 0.95 (�0.09) 0.83 (�0.09)** 0.69 (�0.12)*** �.0001
TemporalTemporal pole 0.84 (�0.04) 0.62 (�0.12)*** 0.82 (�0.08) 0.81 (�0.11) �.0001Lateral temporal 0.92 (�0.03) 0.81 (�0.10)* 0.90 (�0.07) 0.91 (�0.08) �.05Inferior temporal 0.86 (�0.05) 0.77 (�0.29) 0.81 (�0.09) 0.78 (�0.11) nsMedial temporal 0.76 (�0.04) 0.55 (�0.10)*** 0.76 (�0.07) 0.76 (�0.08) �.0001Total 0.85 (�0.04) 0.69 (�0.11)** 0.83 (�0.06) 0.84 (�0.09) �.001
Regional glucose metabolism (rCMRglc): �mol/100g/min
FrontalDorsolateral frontal 38.8 (�6.4) 30.9 (�8.6) 32.3 (�9.5) 29.9 (�9.5) nsOrbitomedial frontal 38.6 (�7.0) 34.1 (�13.2) 30.3 (�5.8)† 15.1 (�5.2)*** �.001Anterior cingulate 41.3 (�7.7) 34.2 (�9.2) 22.2 (�12.5)** 19.6 (�4.5)** �.001Total 39.1 (�6.7) 32.3 (�9.9) 30.3 (�6.9)† 24.2 (�7.5)* �.02
TemporalTemporal pole 25.7 (�5.0) 15.4 (�5.5)* 21.5 (�4.8) 24.3 (�5.7) �.02Lateral temporal 36.9 (�5.9) 24.2 (�7.7) 34.0 (�10.8) 34.7 (�9.8) nsInferior temporal 33.4 (�5.1) 21.3 (�6.7)‡ 31.3 (�10.2) 32.9 (�8.1) �.05Medial temporal 30.0 (�4.4) 19.5 (�6.7)* 29.2 (�7.7) 32.5 (�7.5)* �.02Total 33.7 (�5.3) 21.7 (�6.7)‡ 31.3 (�9.7) 32.9 (�8.5) �.05
Note. FDG-PET � fluorodeoxyglucose positron emission tomography.Student Newman–Keuls post hoc tests (patients vs. controls): *p � .05. **p � .01. ***p � .001.t tests (patients vs. controls, where analysis of variance was significant, and post hoc test was nonsignificant): †p � .05. ‡p � .01.
560 REED ET AL.
A third factor identified was harder to interpret with implicitautomaticity memory scores and explicit picture recall scoresloading in opposite directions.
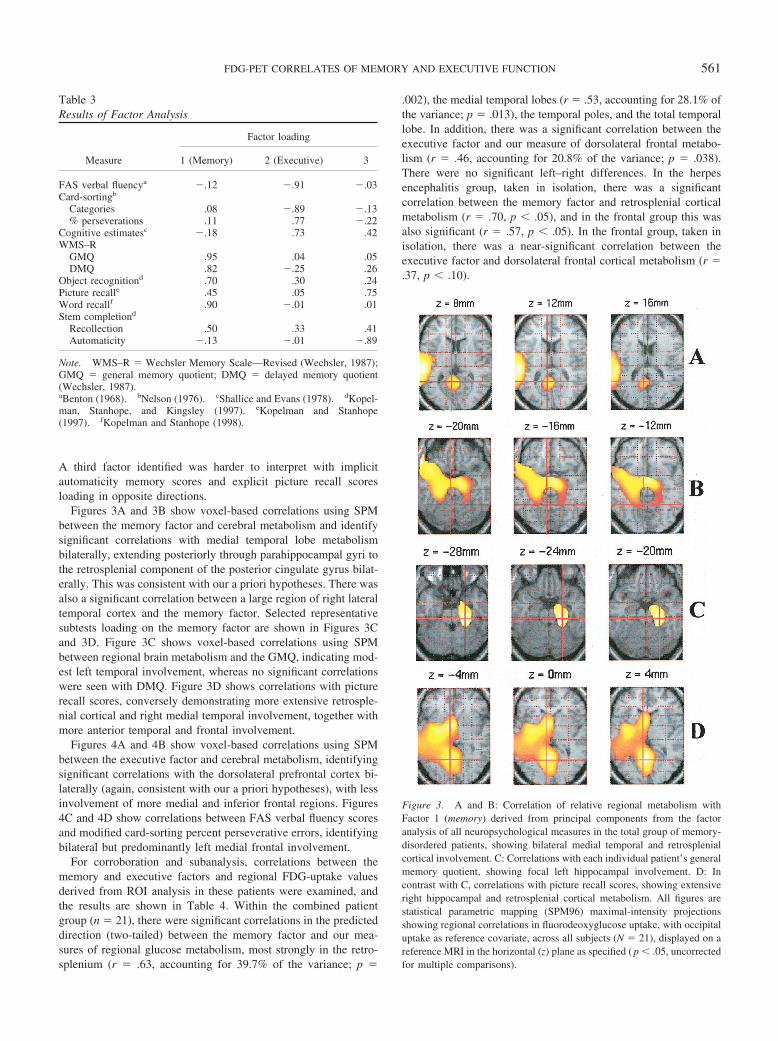
Figures 3A and 3B show voxel-based correlations using SPMbetween the memory factor and cerebral metabolism and identifysignificant correlations with medial temporal lobe metabolismbilaterally, extending posteriorly through parahippocampal gyri tothe retrosplenial component of the posterior cingulate gyrus bilat-erally. This was consistent with our a priori hypotheses. There wasalso a significant correlation between a large region of right lateraltemporal cortex and the memory factor. Selected representativesubtests loading on the memory factor are shown in Figures 3Cand 3D. Figure 3C shows voxel-based correlations using SPMbetween regional brain metabolism and the GMQ, indicating mod-est left temporal involvement, whereas no significant correlationswere seen with DMQ. Figure 3D shows correlations with picturerecall scores, conversely demonstrating more extensive retrosple-nial cortical and right medial temporal involvement, together withmore anterior temporal and frontal involvement.
Figures 4A and 4B show voxel-based correlations using SPMbetween the executive factor and cerebral metabolism, identifyingsignificant correlations with the dorsolateral prefrontal cortex bi-laterally (again, consistent with our a priori hypotheses), with lessinvolvement of more medial and inferior frontal regions. Figures4C and 4D show correlations between FAS verbal fluency scoresand modified card-sorting percent perseverative errors, identifyingbilateral but predominantly left medial frontal involvement.
For corroboration and subanalysis, correlations between thememory and executive factors and regional FDG-uptake valuesderived from ROI analysis in these patients were examined, andthe results are shown in Table 4. Within the combined patientgroup (n � 21), there were significant correlations in the predicteddirection (two-tailed) between the memory factor and our mea-sures of regional glucose metabolism, most strongly in the retro-splenium (r � .63, accounting for 39.7% of the variance; p �
.002), the medial temporal lobes (r � .53, accounting for 28.1% ofthe variance; p � .013), the temporal poles, and the total temporallobe. In addition, there was a significant correlation between theexecutive factor and our measure of dorsolateral frontal metabo-lism (r � .46, accounting for 20.8% of the variance; p � .038).There were no significant left–right differences. In the herpesencephalitis group, taken in isolation, there was a significantcorrelation between the memory factor and retrosplenial corticalmetabolism (r � .70, p � .05), and in the frontal group this wasalso significant (r � .57, p � .05). In the frontal group, taken inisolation, there was a near-significant correlation between theexecutive factor and dorsolateral frontal cortical metabolism (r �.37, p � .10).
Table 3Results of Factor Analysis
Measure
Factor loading
1 (Memory) 2 (Executive) 3
FAS verbal fluencya �.12 �.91 �.03Card-sortingb
Categories .08 �.89 �.13% perseverations .11 .77 �.22
Cognitive estimatesc �.18 .73 .42WMS–R
GMQ .95 .04 .05DMQ .82 �.25 .26
Object recognitiond .70 .30 .24Picture recalle .45 .05 .75Word recallf .90 �.01 .01Stem completiond
Recollection .50 .33 .41Automaticity �.13 �.01 �.89
Note. WMS–R � Wechsler Memory Scale—Revised (Wechsler, 1987);GMQ � general memory quotient; DMQ � delayed memory quotient(Wechsler, 1987).aBenton (1968). bNelson (1976). cShallice and Evans (1978). dKopel-man, Stanhope, and Kingsley (1997). eKopelman and Stanhope(1997). fKopelman and Stanhope (1998).
Figure 3. A and B: Correlation of relative regional metabolism withFactor 1 (memory) derived from principal components from the factoranalysis of all neuropsychological measures in the total group of memory-disordered patients, showing bilateral medial temporal and retrosplenialcortical involvement. C: Correlations with each individual patient’s generalmemory quotient, showing focal left hippocampal involvement. D: Incontrast with C, correlations with picture recall scores, showing extensiveright hippocampal and retrosplenial cortical metabolism. All figures arestatistical parametric mapping (SPM96) maximal-intensity projectionsshowing regional correlations in fluorodeoxyglucose uptake, with occipitaluptake as reference covariate, across all subjects (N � 21), displayed on areference MRI in the horizontal (z) plane as specified ( p � .05, uncorrectedfor multiple comparisons).
561FDG-PET CORRELATES OF MEMORY AND EXECUTIVE FUNCTION

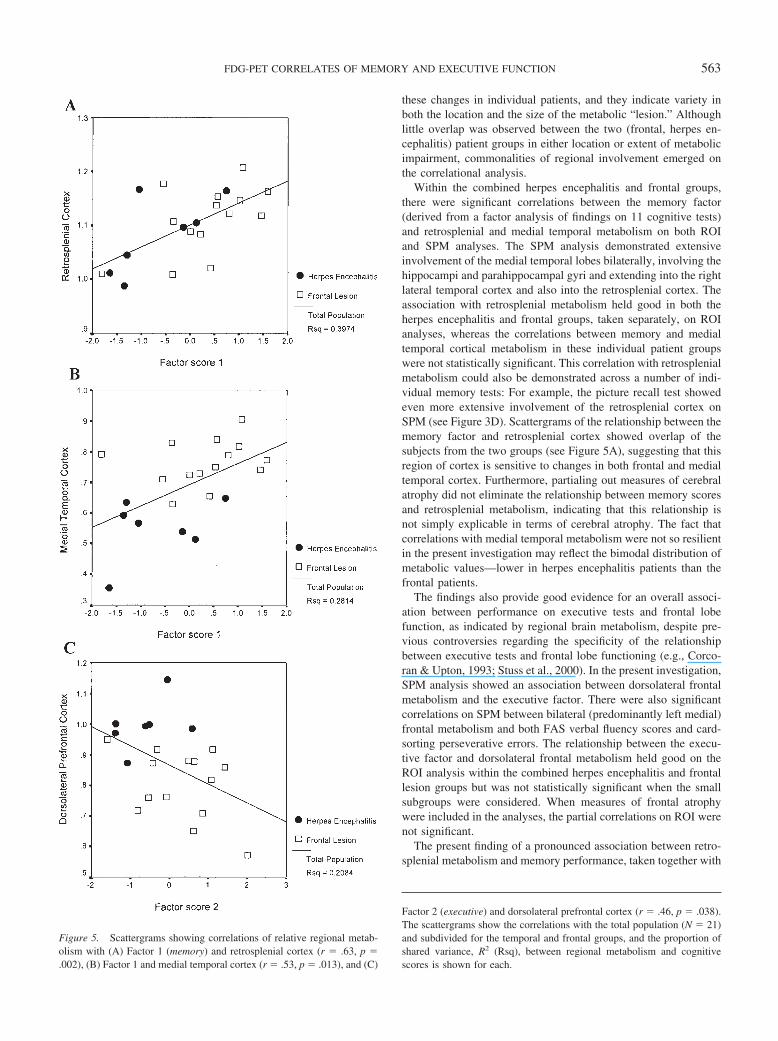
Selected scattergrams displaying these relationships are shownin Figure 5. As expected, herpes encephalitis patients tended toshow the lowest medial temporal metabolism, and frontal patientsshowed the lowest dorsolateral prefrontal metabolism, but therewas a more even distribution of the subgroups for retrosplenialmetabolism, and common regression curves could be drawn for thetwo subgroups across all three scatters, accounting for 20.8% ( p �.04) to 39.7% ( p � .002) of the variance.
To examine the effect of cerebral atrophy on the regionalcorrelations observed, we entered MRI measures of regional brainvolumes into a partial correlation analysis. These were derivedfrom prior volumetric MRI segmentation analyses reported inColchester et al. (2001), comprising whole-brain volume, frontallobe volume, temporal and medial temporal lobe volumes, andremaining posterior cortical volumes. Retrosplenial cortical vol-
umes themselves were unavailable given the lack of unequivocalneuroanatomical boundaries and, thus, posterior brain volume wasused as a marker of general cortical atrophy in this group ofpatients, in whom pathology was primarily in the frontal andtemporal lobes. In this partial correlation analysis, the correlationbetween the memory factor and medial temporal metabolism wasno longer statistically significant. The correlation between execu-tive and dorsolateral prefrontal cortex was reduced from �.46 to�.37 (ns) when total frontal volume was used as a partial correlate.However, although the correlation between the memory factor andretrosplenial cortical metabolism was reduced from .63 to .39, afterpartialing out of the remaining brain volume, this remained sig-nificant on a one-tailed test with a p value of .03.
Discussion
Distinct patterns of metabolic change were observed in theherpes encephalitis and frontal lesion groups. On ROI analysis, thetwo frontal groups showed hypometabolism in the dorsolateral andorbitomedial frontal cortex as well as in the total frontal cortex andthe anterior cingulate. This was more pronounced in the frontaltractotomy group—in whom there were widespread and bilateralfrontal metabolic changes—than in the focal frontal group, in 4 ofwhom the lesions were unilateral. In contrast, the herpes enceph-alitis group showed severe hypometabolism throughout the tem-poral lobes on ROI analysis but did not show statistically signif-icant frontal hypometabolism. As such, these findings corroboratethe use of these groups to exemplify frontal and temporal lobeamnesia in earlier cognitive studies (e.g., Kopelman & Stanhope,1997; Kopelman et al., 1999; Stanhope, Guinan, & Kopelman,1998). The SPM analyses have been used in this article to illustrate
Figure 4. A and B: Correlation of relative regional metabolism withFactor 2 (executive) derived from principal components from the factoranalysis of all neuropsychological measures in the total group of memory-disordered patients, showing bilateral frontal cortical involvement. C:Correlations with each individual patient’s FAS score (Benton, 1968). D:Correlations with percent perseverative errors of the modified card-sortingtest (Nelson, 1976). All figures are statistical parametric mapping (SPM96)maximal-intensity projections showing regional correlations in fluorode-oxyglucose uptake, with occipital uptake as reference covariate, across allsubjects (N � 21), displayed on a reference MRI in the horizontal (z) planeas specified ( p � .05, uncorrected for multiple comparisons).
Table 4Correlations in Herpes Encephalitis and Frontal Lesion GroupsBetween FDG Regional Uptake and Memoryand Executive Factors
Region
Factor
1 (Memory) 2 (Executive)
FrontalDorsolateral frontal �.31 �.46*Orbitomedial frontal �.17 �.29Anterior cingulate .27 �.26Total �.28 �.43
TemporalTemporal pole .49* .21Lateral temporal .39 �.17Inferior temporal .21 .05Medial temporal .53* .38Total .45* .23
ThalamusAnterior thalamus .00 �.14Posterior thalamus .16 �.11Inferior thalamus .25 �.18Total .10 �.14
Retrosplenial .63** .05Parietal .09 .04Caudate �.04 �.03Putamen .17 .08Cerebellum �.17 �.09
Note. FDG � fluorodeoxyglucose.*p � .05 (two-tailed). **p � .01 (two-tailed).
562 REED ET AL.
these changes in individual patients, and they indicate variety inboth the location and the size of the metabolic “lesion.” Althoughlittle overlap was observed between the two (frontal, herpes en-cephalitis) patient groups in either location or extent of metabolicimpairment, commonalities of regional involvement emerged onthe correlational analysis.
Within the combined herpes encephalitis and frontal groups,there were significant correlations between the memory factor(derived from a factor analysis of findings on 11 cognitive tests)and retrosplenial and medial temporal metabolism on both ROIand SPM analyses. The SPM analysis demonstrated extensiveinvolvement of the medial temporal lobes bilaterally, involving thehippocampi and parahippocampal gyri and extending into the rightlateral temporal cortex and also into the retrosplenial cortex. Theassociation with retrosplenial metabolism held good in both theherpes encephalitis and frontal groups, taken separately, on ROIanalyses, whereas the correlations between memory and medialtemporal cortical metabolism in these individual patient groupswere not statistically significant. This correlation with retrosplenialmetabolism could also be demonstrated across a number of indi-vidual memory tests: For example, the picture recall test showedeven more extensive involvement of the retrosplenial cortex onSPM (see Figure 3D). Scattergrams of the relationship between thememory factor and retrosplenial cortex showed overlap of thesubjects from the two groups (see Figure 5A), suggesting that thisregion of cortex is sensitive to changes in both frontal and medialtemporal cortex. Furthermore, partialing out measures of cerebralatrophy did not eliminate the relationship between memory scoresand retrosplenial metabolism, indicating that this relationship isnot simply explicable in terms of cerebral atrophy. The fact thatcorrelations with medial temporal metabolism were not so resilientin the present investigation may reflect the bimodal distribution ofmetabolic values—lower in herpes encephalitis patients than thefrontal patients.
The findings also provide good evidence for an overall associ-ation between performance on executive tests and frontal lobefunction, as indicated by regional brain metabolism, despite pre-vious controversies regarding the specificity of the relationshipbetween executive tests and frontal lobe functioning (e.g., Corco-ran & Upton, 1993; Stuss et al., 2000). In the present investigation,SPM analysis showed an association between dorsolateral frontalmetabolism and the executive factor. There were also significantcorrelations on SPM between bilateral (predominantly left medial)frontal metabolism and both FAS verbal fluency scores and card-sorting perseverative errors. The relationship between the execu-tive factor and dorsolateral frontal metabolism held good on theROI analysis within the combined herpes encephalitis and frontallesion groups but was not statistically significant when the smallsubgroups were considered. When measures of frontal atrophywere included in the analyses, the partial correlations on ROI werenot significant.
The present finding of a pronounced association between retro-splenial metabolism and memory performance, taken together with
Figure 5. Scattergrams showing correlations of relative regional metab-olism with (A) Factor 1 (memory) and retrosplenial cortex (r � .63, p �.002), (B) Factor 1 and medial temporal cortex (r � .53, p � .013), and (C)
Factor 2 (executive) and dorsolateral prefrontal cortex (r � .46, p � .038).The scattergrams show the correlations with the total population (N � 21)and subdivided for the temporal and frontal groups, and the proportion ofshared variance, R2 (Rsq), between regional metabolism and cognitivescores is shown for each.
563FDG-PET CORRELATES OF MEMORY AND EXECUTIVE FUNCTION

previous findings of medial temporal hypometabolism in herpespatients, thalamic hypometabolism in hypoxic patients (Reed et al.,1999), and relative diencephalic hypometabolism in Korsakoffpatients (Reed et al., 2003), is broadly consistent with previousindications of hypometabolism within these limbic–diencephalicstructures in much smaller groups of memory-disordered patients(Aupee et al., 2001; Fazio et al., 1992) and also in Alzheimerpatients (Desgranges et al., 1998, 2002). This accords with thefindings of lesion studies indicating the critical role of limbic–diencephalic circuitry in memory formation.
Retrosplenial activation has been obtained in a number of func-tional imaging investigations of healthy subjects, particularly withrespect to topographical memory, as well as having been impli-cated in previous neuropsychological studies of memory-disor-dered patients (e.g., Maguire, 2001b; Valenstein et al., 1987). Theretrosplenial cortex is interconnected with both the hippocampusand the anterior thalamus (Valenstein et al., 1987), which arecritical in memory formation (Harding, Halliday, Caine, & Kril,2000). Anatomical studies using anterograde and retrograde trac-ing of pathways subtending the retrosplenial cortex in the rhesusmonkey (Morris et al., 1999) have shown that this area has recip-rocal connections with the middorsolateral part of the prefrontalcortex and with the posterior parahippocampal cortex and theentorhinal cortex. Morris et al. also showed reciprocal connectionswith the lateroposterior thalamic nucleus as well as with limbicthalamic nuclei, thus clearly implicating retrosplenial cortex in anetwork linking the frontal, temporal, and diencephalic regionsinvolved in memory processing. An FDG-PET study in a patientwith retrosplenial pathology and severe amnesia revealed thalamicas well as retrosplenial hypometabolism, which was interpreted interms of a disconnection syndrome (Heilman et al., 1990). More-over, in less severe cases of Alzheimer’s dementia, correlationshave previously been reported between verbal memory and retro-splenial and medial temporal metabolism (Desgranges et al.,2002). Taken together, these findings suggest that the retrosple-nium may be particularly sensitive to changes either “upstream” or“downstream” in the memory circuits.
In conclusion, the present study clearly indicates that differingpatterns of altered metabolism can be obtained in patients whosememory impairments result from sites of pathology within thefrontal and temporal lobes. Furthermore, it has also demonstratedthat meaningful correlations can be obtained between patterns of“resting” regional brain metabolism and cognitive tasks performedwithin a few days of the PET scan. Factor analysis of neuropsy-chological data revealed that memory performance was associatedwith retrosplenial and medial temporal metabolism, and executivefunction was associated with dorsolateral frontal metabolism. Thestrongest and most consistent correlation within the present patientgroups was between anterograde memory function and retrosple-nial metabolism, and we have suggested that the retrosplenium isparticularly sensitive to functional change both “upstream” and“downstream.” These observations add to the growing evidence ofthe importance of the retrosplenial cortex in memory performanceand, thus, corroborate the links between cerebral metabolism andbehavioral performance (Keri & Gulyas, 2003).
References
Aupee, A. M., Desgranges, B., Eustache, F., Lalevee, C., de la Sayette, V.,Viader, F., & Baron, J. C. (2001). Voxel-based mapping of brain
hypometabolism in permanent amnesia with PET. NeuroImage, 13,1164–1173.
Benton, A. L. (1968). Differential behavioral effects of frontal lobe disease.Neuropsychologia, 6, 53–60.
Colchester, A., Kingsley, D., Lasserson, D., Kendall, B., Bello, F., Rush,C., et al. (2001). Structural MRI volumetric analysis in patients withorganic amnesia, 1: Methods and comparative findings across diagnosticgroups. Journal of Neurology, Neurosurgery and Psychiatry, 71, 13–22.
Corcoran, R., & Upton, D. (1993). A role for the hippocampus in cardsorting? Cortex, 29, 293–304.
Desgranges, B., Baron, J.-C., de la Sayette, V., Petit-Taboue, M.-C.,Benali, K., Landeau, B., et al. (1998). The neural substrates of memorysystems impairment in Alzheimer’s disease: A PET study of restingbrain glucose utilization. Brain, 121, 611–631.
Desgranges, B., Baron, J.-C., Lalevee, C., Giffard, B., Viader, F., de laSayette, V., & Eustache, F. (2002). The neural substrates of episodicmemory impairment in Alzheimer’s disease as revealed by FDG–PET:Relationship to degree of deterioration. Brain, 125, 1116–1124.
Fazio, F., Perani, D., Gilardi, M. C., Colombo, F., Cappa, S. F., Vallar, G.,et al. (1992). Metabolic impairment in human amnesia: A PET study ofmemory networks. Journal of Cerebral Blood Flow & Metabolism, 12,353–358.
Friston, K. J., Ashburner, J., Frith, C. D., Poline, J.-B., Heather, J. D., &Frackowiak, R. S. J. (1995). Spatial registration and normalization ofimages. Human Brain Mapping, 3, 165–189.
Friston, K. J., Frith, C. D., Liddle, P. F., & Frackowiak, R. S. (1991).Comparing functional (PET) images: The assessment of significantchange. Journal of Cerebral Blood Flow & Metabolism, 11, 690–699.
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J.-B., Frith, C. D., &Frackowiak, R. S. J. (1995). Statistical parametric maps in functionalimaging: A general linear approach. Human Brain Mapping, 2, 189–210.
Grasby, P. M., Frith, C. D., Friston, K. J., Bench, C., Frackowiak, R. S., &Dolan, R. (1993). Functional mapping of brain areas implicated inauditory–verbal memory function. Brain, 116, 1–20.
Harding, A., Halliday, G., Caine, D., & Kril, J. (2000). Degeneration ofanterior thalamic nuclei differentiates alcoholics with amnesia. Brain,123, 141–154.
Heilman, K. M., Bowers, D., Watson, T., Day, A., Valenstein, E., Ham-mond, E., & Duara, R. (1990). Frontal hypermetabolism and thalamichypometabolism in a patient with abnormal orienting and retrosplenialamnesia. Neuropsychologia, 28, 161–169.
Huang, S. C., Phelps, M. E., Hoffman, E. J., Sideris, K., Selin, C. J., &Kuhl, D. E. (1980). Noninvasive determination of local cerebral meta-bolic rate of glucose in man. American Journal of Physiology, 238,E69–E82.
Jacoby, L. L., & Kelley, C. (1992). Unconscious influences of memory:Dissociations and automaticity. In D. Milner & M. D. Rugg (Eds.), Theneuropsychology of consciousness (pp. 201–233). London: AcademicPress.
Joyce, E. M., Rio, D. E., Ruttimann, U. E., Rohrbaugh, J. W., Martin, P. R.,Rawlings, R. R., & Eckhardt, M. J. (1994). Decreased cingulate andprecuneate glucose utilization in alcoholic Korsakoff’s syndrome. Psy-chiatry Research, 54, 225–239.
Kapur, N., Thompson, P., Kartsounis, L. D., & Abbot, P. (1999). Retro-grade amnesia: Clinical and methodological caveats. Neuropsycholo-gia, 37, 27–30.
Kapur, S., Craik, F. I., Tulving, E., Wilson, A. A., Houle, S., Brown, G. M.(1994). Neuroanatomical correlates of encoding in episodic memory:Levels of processing effect. Proceedings of National Academy of Sci-ence, USA, 91, 2008–2011.
Kartsounis, L. D., Poynton, A., Bridges, P. K., & Bartlett, J. R. (1991).Neuropsychological correlates of stereotactic subcaudate surgery. Brain,114, 2657–2673.
564 REED ET AL.

Kennedy, A. M., Newman, S. K., Frackowiak, R. S., Cunningham, V. J.,Roques, P., Stevens, J., et al. (1995). Chromosome 14 linked familialAlzheimer’s disease. A clinico-pathological study of a single pedigree.Brain, 118, 185–205.
Keri, S., & Gulyas, B. (2003). Four facets of a single brain: Behaviour,cerebral blood flow/metabolism, neuronal activity and neurotransmitterdynamics. NeuroReport, 14, 1097–1106.
Kopelman, M. D. (2002). Disorders of memory. Brain, 125, 2152–2190.Kopelman, M. D., Lasserson, D., Kingsley, D., Bello, F., Rush, C., Stan-
hope, N., et al. (2001). Structural MRI volumetric analysis in patientswith organic amnesia, 2: Correlations with anterograde memory andexecutive tests in 40 patients. Journal of Neurology, Neurosurgery andPsychiatry, 71, 23–28.
Kopelman, M. D., Lasserson, D., Kingsley, D. R., Bello, F., Rush, C.,Stanhope, N., et al. (2003). Retrograde amnesia and the volume ofcritical brain structures. Hippocampus, 13, 879–891.
Kopelman, M. D., Reed, L. J., Marsden, P., Mayes, A. R., Jaldow, E.,Laing, H., & Isaac, C. (2001). Amnesic syndrome and severe ataxiafollowing the recreational use of 3,4-methylene-dioxymethamphetamine(MDMA, “Ecstasy”) and other substances. Neurocase, 7, 423–432.
Kopelman, M. D., & Stanhope, N. (1997). Rates of forgetting in organicamnesia following temporal lobe, diencephalic, or frontal lobe lesions.Neuropsychology, 11, 343–356.
Kopelman, M. D., & Stanhope, N. (1998). Recall and recognition memoryin patients with focal frontal, temporal lobe and diencephalic lesions.Neuropsychologia, 36, 785–796.
Kopelman, M. D., Stanhope, N., & Kingsley, D. (1997). Temporal andspatial memory in patients with focal frontal, temporal lobe, and dien-cephalic lesions. Neuropsychologia, 35, 1533–1545.
Kopelman, M. D., Stanhope, N., & Kingsley, D. (1999). Retrogradeamnesia in patients with diencephalic, temporal lobe or frontal lesions.Neuropsychologia, 37, 939–958.
Kopelman, M. D., Stevens, T. G., Foli, S., & Grasby, P. (1998). PETactivation of the medial temporal lobe in learning. Brain, 121, 875–887.
Kuhl, D. E., Phelps, M. E., Kowell, A. P., Metter, E. J., Selin, C., & Winter,J. (1980). Effects of stroke on local cerebral metabolism and perfusion:Mapping by emission computed tomography of 18FDG 13NH3. Annals ofNeurology, 8, 47–60.
Kuwert, T., Homberg, V., Steinmetz, H., Unverhau, S., Langen, K.-J.,Herzog, H., & Feinendegen, L. E. (1993). Posthypoxic amnesia: Re-gional cerebral glucose consumption measured by positron emissiontomography. Journal of the Neurological Sciences, 118, 10–16.
Maguire, E. A. (2001a). Neuroimaging studies of autobiographical eventmemory. Philosophical Transactions of the Royal Society of London,Series B, 356, 1441–1451.
Maguire, E. A. (2001b). The retrosplenial contribution to human naviga-tion: A review of lesion and neuroimaging findings. Scandinavian Jour-nal of Psychology, 42, 225–238.
Markowitsch, H. J., Weber-Luxenberger, G., Ewald, K., Kessler, J., &Heiss, W.-D. (1997). Patients with heart attacks are not valid models formedial temporal lobe amnesia: A neuropsychological and FDG-PETstudy with consequences for memory research. European Journal ofNeurology, 4, 178–184.
Martin, P. R., Rio, D., Adinoff, B., Johnson, J. L., Bisserbe, J. C.,Rawlings, R. R., et al. (1992). Regional cerebral glucose utilization inchronic organic mental disorders associated with alcoholism. Journal ofNeuropsychiatry and Clinical Neurosciences, 4, 159–167.
Montaldi, D., Mayes, A. R., Barnes, A., Pirie, H., Hadley, D. M., Patterson,J., & Wyper, D. J. (1998). Associative encoding of pictures activates themedial temporal lobes. Human Brain Mapping, 6, 85–104.
Morris, R., Petrides, M., & Pandya, D. N. (1999). Architecture and con-nections of retrosplenial area 30 in the rhesus monkey (macaca mulatta).European Journal of Neuroscience, 11, 2506–2518.
Nelson, H. E. (1976). A modified card sorting test sensitive to frontal lobedeficits. Cortex, 12, 313–324.
Nelson, H. E., & Willison, J. R. (1991). National Adult Reading Test(NART): Test manual (2nd ed.). Windsor, England: NFER-Nelson.
Nestor, P. J., Fryer, T. D., Smielewski, P., & Hodges, J. R. (2003). Annalsof Neurology, 54, 343–351.
Paller, K. A., Acharya, A., Richardson, B. C., Plaisant, O., Shimamura,A. P., Reed, B. R., & Jagust, W. J. (1997). Functional neuroimaging ofcortical dysfunction in alcoholic Korsakoff’s syndrome. Journal of Cog-nitive Neuroscience, 9, 277–293.
Phelps, M. E., Huang, S. C., Hoffman, E. J., Selin, C., Sokoloff, L., &Kuhl, D. E. (1979). Tomographic measurement of local cerebral glucosemetabolic rate in humans with (F-18)2-fluoro-2-deoxy-D-glucose: Val-idation of method. Annals of Neurology, 6, 371–388.
Reed, L. J., Lasserson, D., Marsden, P., Stanhope, N., Stevens, T., Bello,F., et al. (2003). FDG-PET findings in the Wernicke–Korsakoff syn-drome. Cortex, 39, 1027–1045.
Reed, L. J., Marsden, P., Lasserson, D., Sheldon, N., Lewis, P., Stanhope,N., et al. (1999). FDG-PET analysis and findings in amnesia resultingfrom hypoxia. Memory, 7, 599–614.
Rudge, P., & Warrington, E. K. (1991). Selective impairment of memoryand visual perception in splenial tumours. Brain, 114, 349–360.
Rugg, M. D. (2002). Functional neuroimaging of memory. In A. D.Baddeley, M. D. Kopelman, & B. A. Wilson (Eds.), Handbook ofmemory disorders (2nd ed., pp. 57–80). Chichester, England: Wiley.
Schacter, D. L., Alpert, N. M., Savage, C. R., Rauch, S. L., & Albert, M. S.(1996). Conscious recollection and the human hippocampal formation.Proceedings of the National Academy of Sciences, USA, 93, 321–325.
Selden, N. R., Gitelman, D. R., Salamon-Murayama, N., Parrish, T. B., &Mesulam, M.-M. (1998). Trajectories of cholinergic pathways within thecerebral hemispheres of the human brain. Brain, 121, 2249–2257.
Shallice, T., & Evans, M. E. (1978). The involvement of the frontal lobesin cognitive estimation. Cortex, 14, 294–303.
Shallice, T., Fletcher, P., Frith, C. D., Grasby, P., Frackowiak, R. S., &Dolan, R. J. (1994, April 14). Brain regions associated with acquisitionand retrieval of verbal episodic memory. Nature, 368, 633–635.
Signorini, M., Paulesu, E., Friston, K., Perani, D., Colleluori, A., Lucig-nani, et al. (1999). Rapid assessment of regional cerebral metabolicabnormalities in single subjects with quantitative and nonquantitative[18F]FDG PET: A clinical validation of statistical parametric mapping.NeuroImage, 9, 63–80.
Stanhope, N., Guinan, E., & Kopelman, M. D. (1998). Frequency judge-ments of abstract designs by patients with diencephalic, temporal lobe orfrontal lobe lesions. Neuropsychologia, 36, 1387–1396.
Stuss, D. T., Levine, B., Alexander, M. P., Hong, J., Palumbo, C., Hamer,L., et al. (2000). Wisconsin Card Sorting Test performance in patientswith focal frontal and posterior brain damage: Effects of lesion locationand test structure on separable cognitive processes. Neuropsycholo-gia, 38, 388–402.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotactic atlas of thehuman brain. Stuttgart, Germany: Thieme.
Valenstein, E., Bowers, D., Verfaellie, M., Heilman, K. M., Day, A., &Watson, R. T. (1987). Retrosplenial amnesia. Brain, 110, 1631–1646.
Wechsler, D. (1981). Wechsler Adult Intelligence Scale—Revised. SanAntonio, TX: Psychological Corporation.
Wechsler, D. (1987). Wechsler Memory Scale—Revised. San Antonio, TX:Psychological Corporation.
Wheeler, M. A., Stuss, D. T., & Tulving, E. (1995). Frontal lobe damageproduces episodic memory impairment. Journal of the InternationalNeuropsychological Society, 1, 525–536.
Received July 18, 2003Revision received April 13, 2004
Accepted April 30, 2004 �
565FDG-PET CORRELATES OF MEMORY AND EXECUTIVE FUNCTION
Related Documents






![Clinical characteristics and long-term prognosis of ... · other possible causes, such as viral encephalitis and herpes simplex encephalitis. Relapsing anti-NMDAR encephalitis [5]](https://static.cupdf.com/doc/110x72/5f5cb978af3eab35a02f3630/clinical-characteristics-and-long-term-prognosis-of-other-possible-causes-such.jpg)




