In Vivo Correction of Murine Hereditary Tyrosinemia Type I by FC31 Integrase-Mediated Gene Delivery Patrice K. Held, 1, * Eric C. Olivares, 2 Christina P. Aguilar, 1 Milton Finegold, 3 Michele P. Calos, 2 and Markus Grompe 1 1 Molecular and Medical Genetics, Oregon Health & Science University, Portland, OR 97239, USA 2 Department of Genetics, Stanford University School of Medicine, Stanford, CA 94305, USA 3 Department of Pathology, Baylor College of Medicine, Houston, TX 77030, USA *To whom correspondence and reprint requests should be addressed at Oregon Health & Science University, 3181 SW Sam Jackson Park Road, L103, Portland, OR 97239, USA. Fax: +1 503 494 6886; E-mail: [email protected]. Available online 15 December 2004 Phage fC31 integrase is a site-specific recombinase that mediates efficient integration of circular extrachromosomal DNA into the host genome. Here, the integrase system was used to transfer the fumarylacetoacetate hydrolase (FAH ) gene into the liver of mice affected with hereditary tyrosinemia type 1. Approximately 3.6% of transfected hepatocytes experienced an integration event. The absolute frequency of integration was 1/1374. A higher proportion of integrase- transfected FAH + hepatocytes displayed abnormal morphology (bizarre nuclei, enlarged cells) on day 25 after gene transfer, compared to cells not receiving integrase. The increased frequency of these abnormal cells correlated with the amount of integrase plasmid administered, suggesting some form of integrase toxicity in Fah / livers. The abnormal hepatocyte appearance was transient and livers analyzed after longer selection (90 days) showed 60% repopulation with only normal healthy FAH + hepatocytes. A total of seven different integration sites (accounting for N90% of integration) were identified. Serial transplantation of integrase-corrected hepatocytes to Fah / recipients was successful, suggesting long-term viability of corrected cells and persistent gene expression through many rounds of cell division. The stability of transgene expression, relatively high integration frequency, and significant site specificity that characterize the fC31 integration system suggest that it may have utility in many gene therapy settings. Key Words: nonviral somatic gene therapy, phage integrase, genetic disease INTRODUCTION Currently, most gene therapy applications use viral vectors to transfer therapeutic DNA into cellular targets. While these vectors can be effective in vivo , they also have several disadvantages. First, viral vectors can trigger immune responses at multiple levels [1,2]. Second, large-scale production of clinical-grade vectors is difficult and costly. Finally, some viral vectors can cause inser- tional mutagenesis [3,4]. In contrast, nonviral vectors are less likely to trigger immune responses and are cheaper to produce, but their efficacy in correcting genetic diseases has been hampered by lack of integration and long-term gene expression. Recently, plasmid-based nonviral gene transfer systems capable of mediating stable integration of transgenes have been described. The Sleeping Beauty (SB) transposase/transposon system has been shown to mediate effective transposition of foreign DNA into host cells in vitro and in vivo [5–7]. However, SB transposase- mediated integration occurs at random sites in the genome and therefore could result in insertional muta- genesis, similar to integrating viral vectors. Ideally the integration of foreign DNA into the genome must be site- specific and controlled to prevent activation of onco- genes and disruption of essential or tumor-suppressor genes. Phage fC31 integrase is a site-specific recombinase that mediates efficient integration of plasmid DNA into host cell genomes, including mammalian cells [8]. In nature, it mediates a unidirectional recombination event between the 39-bp attP site of the phage fC31 genome and the related 34-bp attB site in the Streptomyces chromosome [9]. In mammalian cells, fC31 integrase has been shown also to recognize partially identical attP sites in both the murine and the human genome [10,11]. These sites were termed bpseudo attPQ sites. The efficiency of integration at genomic pseudo attP sites was estimated ARTICLE doi:10.1016/j.ymthe.2004.11.001 MOLECULAR THERAPY Vol. 11, No. 3, March 2005 399 Copyright C The American Society of Gene Therapy 1525-0016/$30.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLEdoi101016jymthe200411001
In Vivo Correction of Murine Hereditary TyrosinemiaType I by FC31 Integrase-Mediated Gene Delivery
Patrice K Held1 Eric C Olivares2 Christina P Aguilar1 Milton Finegold3
Michele P Calos2 and Markus Grompe1
1 Molecular and Medical Genetics Oregon Health amp Science University Portland OR 97239 USA2 Department of Genetics Stanford University School of Medicine Stanford CA 94305 USA
3 Department of Pathology Baylor College of Medicine Houston TX 77030 USA
To whom correspondence and reprint requests should be addressed at Oregon Health amp Science University
3181 SW Sam Jackson Park Road L103 Portland OR 97239 USA Fax +1 503 494 6886 E-mail heldpohsuedu
Available online 15 December 2004
MOLECULA
Copyright C
1525-0016$
Phage fC31 integrase is a site-specific recombinase that mediates efficient integration of circularextrachromosomal DNA into the host genome Here the integrase system was used to transfer thefumarylacetoacetate hydrolase (FAH ) gene into the liver of mice affected with hereditarytyrosinemia type 1 Approximately 36 of transfected hepatocytes experienced an integrationevent The absolute frequency of integration was 11374 A higher proportion of integrase-transfected FAH+ hepatocytes displayed abnormal morphology (bizarre nuclei enlarged cells) onday 25 after gene transfer compared to cells not receiving integrase The increased frequency ofthese abnormal cells correlated with the amount of integrase plasmid administered suggestingsome form of integrase toxicity in Fah livers The abnormal hepatocyte appearance was transientand livers analyzed after longer selection (90 days) showed 60 repopulation with only normalhealthy FAH+ hepatocytes A total of seven different integration sites (accounting for N90 ofintegration) were identified Serial transplantation of integrase-corrected hepatocytes to Fah
recipients was successful suggesting long-term viability of corrected cells and persistent geneexpression through many rounds of cell division The stability of transgene expression relativelyhigh integration frequency and significant site specificity that characterize the fC31 integrationsystem suggest that it may have utility in many gene therapy settings
R
Th
30
Key Words nonviral somatic gene therapy phage integrase genetic disease
INTRODUCTION
Currently most gene therapy applications use viralvectors to transfer therapeutic DNA into cellular targetsWhile these vectors can be effective in vivo they also haveseveral disadvantages First viral vectors can triggerimmune responses at multiple levels [12] Secondlarge-scale production of clinical-grade vectors is difficultand costly Finally some viral vectors can cause inser-tional mutagenesis [34] In contrast nonviral vectors areless likely to trigger immune responses and are cheaper toproduce but their efficacy in correcting genetic diseaseshas been hampered by lack of integration and long-termgene expression Recently plasmid-based nonviral genetransfer systems capable of mediating stable integrationof transgenes have been described The Sleeping Beauty(SB) transposasetransposon system has been shown tomediate effective transposition of foreign DNA into hostcells in vitro and in vivo [5ndash7] However SB transposase-
THERAPY Vol 11 No 3 March 2005
e American Society of Gene Therapy
00
mediated integration occurs at random sites in thegenome and therefore could result in insertional muta-genesis similar to integrating viral vectors Ideally theintegration of foreign DNA into the genome must be site-specific and controlled to prevent activation of onco-genes and disruption of essential or tumor-suppressorgenes
Phage fC31 integrase is a site-specific recombinasethat mediates efficient integration of plasmid DNA intohost cell genomes including mammalian cells [8] Innature it mediates a unidirectional recombination eventbetween the 39-bp attP site of the phage fC31 genomeand the related 34-bp attB site in the Streptomyceschromosome [9] In mammalian cells fC31 integrasehas been shown also to recognize partially identical attPsites in both the murine and the human genome [1011]These sites were termed bpseudo attPQ sites The efficiencyof integration at genomic pseudo attP sites was estimated
399
TABLE 1 Absolute and actual integration frequencies of integrase-treated mice
Group 1 2
Plasmid combinations 25 Ag FAH-att 25 Ag integrase (n = 30) 25 Ag FAH-att 25 Ag empty vector (n = 9)
Absolute integration frequency 73 F 38 104 28 F 44 105
Minimum 14880 0
Median 11374 135640Maximum 1465 17165
Corrected integration frequency 36 014
127 1714
FAH + hepatocytes and nodules were counted as single integration events Averages with standard deviations are given In addition the minimum (worst animal) and maximum (best
animal) frequencies in each group are shown
ARTICLE doi101016jymthe200411001
to be approximately 5ndash10 [10] similar to that observedwith SB transposase Pseudo attP sites are rare in both thehuman and the murine genome and therefore thenumber of integration sites is limited In one settingonly two integrations were identified in vivo within themouse genome [10] Therefore fC31 integrase-mediatedintegration is nonrandom and efficient enough to makethe system very attractive for nonviral gene therapy ofgenetic disorders Several papers have been publishedshowing in vivo applications of fC31 integrase Onestudy showed the delivery and site-specific integration ofthe human Factor IX transgene in mouse liver in vivoresulting in production of therapeutic Factor IX levels[10] Further studies reported the ability of fC31 inte-grase to transfect human keratinocytes and to correctinherited skin diseases in a xenotransplantation model[1112]
To characterize further the properties of fC31 inte-grase-mediated gene transfer in vivo we used the murinemodel of hereditary tyrosinemia type 1 (HT1) [13] Thissystem has unique advantages for the characterization ofstable integration events in hepatocytes in vivo HT1 is agenetic liver disease caused by deficiency of fumarylace-toacetate hydrolase (FAH) [13] the enzyme that catalyzesthe last step in the tyrosine degradation pathway In HT1FAH-expressing hepatocytes display a strong proliferativeadvantage and eventually repopulate the entire diseasedliver [1415] Because of this selective advantage hepato-
FIG 1 Frequency of fC31 integrase-medi-
ated stable FAH gene transfer The frequency
of integration in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of sections
400
cytes in which fC31 integrase has mediated integrationof the FAH gene can be identified counted andhistologically evaluated soon after transfection as wellas after several rounds of growth and expansion of FAH+
hepatocyte clonesHere we report the properties of fC31 integrase-
mediated gene transfer in the HT1 mouse model
RESULTS
Frequency of Stable Integration and HistologicalEvaluationThe efficiency of gene transfer into the liver is typicallydetermined by the expression of histochemical markerssuch as h-galactosidase or alkaline phosphatase [2627]However the simple presence or absence of such markersdoes not accurately measure integration frequencybecause gene expression can occur in both the presenceand the absence of chromosomal integration [28] In theFah mouse model single FAH-expressing hepatocytescan selectively expand and form clonal nodules only ifstable integration occurred in the initially transfectedcells [1429] This model is therefore well suited tomeasure the frequency of vector integration and stabletransgene expression
We calculated the integration frequency for experi-mental plasmids and controls We injected group 1 (n =10) with a FAH-expressing construct containing the attB
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therap
y
ARTICLEdoi101016jymthe200411001
site (FAH-att) plus the integrase expression constructacting in trans We treated group 2 (n = 5) with FAH-attplus an empty vector lacking integrase We used apreviously described 11 ratio (25 Ag of each plasmid) ofintegrase (or empty vector) to the FAH expressionconstruct (FAH-att) [10] On day 3 after plasmid injec-tion we discontinued 2-(2-nitro-4-trifluoromethylben-zoyl)-13-cyclohexanedione (NTBC) treatment to permitthe development of liver disease and the selection ofFAH+ hepatocyte nodules We sacrificed all animals 25days postinjection and processed the liver tissue forhistological evaluation We scored several liver sectionsfor each mouse for FAH+ cells and nodules The totalnumber of sections evaluated for each group was 30 forgroup 1 and 9 for group 2 (Table 1 Fig 1) Approximately2 to 3 sections were scored for each individual mousewithin group 1 or 2
In the liver sections from group 1 (mice treated withFAH-att and integrase) the average absolute frequency ofFAH+ nodules was 73 F 38 104 (11374 hepatocytes)The frequency ranged over 10-fold from a maximum of 1465 cells in the best mouse to 14880 in the worst Group2 (FAH-att and an empty vector) displayed minimalintegration at only 28 F 44 105 (135640 cells)and was significantly different from group 1 as evaluatedby a Student t test ( P value b00001) To determine theefficiency of the fC31 integrase-mediated integration ona per-cell basis the DNA transfection frequency must beconsidered To measure this parameter we injected threeFah mice via the tail vein with a h-galactosidaseexpression construct and sacrificed them 3 days laterWe stained liver sections for h-galactosidase and calcu-lated the hepatocyte transfection efficiency at ~2 (datanot shown) This number confirms that Fah mutant
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
mice have lower rates of transfection than typically seenafter hydrodynamic injection [5] The average correctedintegration frequency for group 1 mice was 36 (127)ranging from 1 to 107 Group 2 mice had an averageper-cell integration frequency of 01 (1714) rangingfrom 0 to 06 (Table 1 Fig 1)
Integrase Induced a Higher Frequency of HepatocyteDysplasiaFAH+ hepatocytes observed in histological sections fromgroup 1 mice harvested after 25 days of selectiondisplayed a unique dysplastic phenotype at a higherfrequency than controls Specifically individual FAH+
hepatocytes were often markedly enlarged in compari-son to adjacent FAH-negative hepatocytes (Fig 2A) Inaddition many of the nuclei in these abnormal cellswere massively enlarged giving the hepatocytes abdysplasticQ appearance We devised an index ratingsystem to quantitate the presence of these abnormalcells Nodules in which more than half of the hepato-cytes were clearly abnormal in phenotype were scoredwith a 3+ rating while nodules in which less than half ofthe hepatocytes were abnormal were scored with a 2+rating Nodules that contained only morphologicallynormal hepatocytes were given a 1+ rating (Figs 2D 2Eand 2F) Seventy percent of the nodules in group 1 micecontained abnormal cells (2+ and 3+) (Fig 3) Group 2mice which did not receive integrase did not have any3+ nodules but still had approximately 25 2+ noduleswhich contain a lower fraction of abnormal cells (Figs2C and 3) As an additional control we also injectedFah mice with the Sleeping Beauty transposon andtheir scores paralleled those of the integrase-transfectedmice after sacrifice on day 25 [5] Specifically 30 Ag of
FIG 2 FAH immunohistochemistry in hepa-
tocytes treated with fC31 integrase (A) Sec-
tion from a mouse treated with FAH-att and
integrase after 25 days of selection Arrows
denote hepatocytes with abnormal morpho-
logy (B) Mouse treated with FAH-att and inte-
grase after 90+ days of selection (C) Mouse
treated with FAH-att and empty vector after
25 days of selection (D) Nodule scored as a 1+
in the dysplasia index (no abnormal FAH+
cells) (E) Nodule scored as a 2+ in the
dysplasia index (less than half of the FAH+ cells
are abnormal) (F) Nodule scored as a 3+ in the
dysplasia index (more than half of the FAH+
cells are abnormal)
401
ARTICLE doi101016jymthe200411001
the transposon containing the FAH gene (driven by theSRa promoter) and 1 Ag of the transposase were deliveredtogether by hydrodynamic injection into Fah mice aspreviously described [5] Mice treated with the trans-poson showed hepatocyte dysplasia in a smaller percent-age of the nodules similar to the group 2 controlswhich did not receive any integrase plasmid Approxi-mately 75 of the nodules were healthy (1+) and 25 ofthe nodules were scored as 2+ (Fig 3) The 2+ nodulesfound in group 2 with no integrase and in the SleepingBeauty transposon control suggested that a level ofhepatocyte dysplasia is attributable to the Fah deficiencyitself FAH overexpression andor the hydrodynamicinjection method
In fact prevalent hepatocyte dysplasia as seen in the3+ nodules is a common feature of Fah mutant liverduring NTBC withdrawal without correction [2430]Here we withdrew NTBC 3 days after plasmid injectionand it is therefore possible that this time was too shortto initiate FAH expression To determine whetherremoval of NTBC prior to full FAH expression was thecause of injury and dysplasia in the integrase-trans-fected hepatocytes we gave three additional group 1mice maximal NTBC therapy during and after plasmidinjection They were treated with a higher dose ofNTBC (30 mgL drinking watermdashfour times the normalamount) for a longer period (1 week instead of 3 days)However this did not change the frequency of FAHintegration or the dysplasia index The average integra-tion frequency for these three mice was 74 F 18 104 (11332) and ~35 of the nodules were scored ashighly abnormal (3+)
Importantly the abnormal phenotype describedabove did not persist long term (Fig 2B) Integrase-treated (n = 4) mice sacrificed after more than 90 days of
FIG 3 Nodule dysplasia index in hepatocytes
treated with fC31 integrase The nodule
dysplasia index in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of nodules scored
402
hepatocyte selection had ~70 liver repopulation withonly normal healthy hepatocytes as seen in Fig 2 Thisobservation indicates that the abnormal hepatocytephenotype found 25 days after gene therapy is transientIt is currently unknown whether individual abnormalFAH+ hepatocytes revert to normal cell morphology orwhether normal FAH+ hepatocytes have a selectiveadvantage and simply outgrow the dysplastic cells overtime
Effects of Different Levels of IntegraseTo determine whether the high levels of integraseplasmid administered were the cause of the higherfrequency of abnormal hepatocytes we performedadditional experiments to define the optimal amountsof integrase plasmid for effective gene therapy Weinjected four different groups of Fah mice with thefollowing dilutions of integrase group A (n = 10) wasinjected with 25 Ag of both plasmids integrase andFAH-att (11 ratio) group B (n = 8) received 5 Ag ofintegrase and 25 Ag of FAH-att (15 ratio) group C (n =6) 25 Ag of the integrase and 25 Ag of FAH-att (110ratio) and group D (n = 5) 125 Ag of integrase and25 Ag of FAH-att (120) We evaluated liver sectionsfrom each mouse for integration frequency and mor-phology 25 days after removal from NTBC We saw thehighest percentage of abnormal 3+ nodules in micefrom group A receiving a 11 ratio of Fah-att tointegrase (Fig 3) The proportion of 3+ nodulesdecreased and the frequency of 1+ normal nodulesincreased with decreasing amount of the integraseplasmid Group D mice receiving only 125 Ag of theintegrase plasmid had no abnormal nodules Thus theamount of the integrase plasmid administered corre-lated directly with the frequency of morphologically
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
abnormal hepatocytes The quantity of integrase plas-mid was also strongly correlated with the level of genetransfer The average absolute integration frequencyranged from 11374 with a 11 ratio (group A) toonly 134091 with a 120 ratio (group D) Thesefindings are summarized in Table 2 and Fig 1
Integration Site AnalysisOne major pseudo attP site (mpsL1) favored forintegration into the mouse genome has been describedpreviously [10] Here we determined whether integra-tion into other sites also occurred We stably trans-fected approximately 11000 hepatocytes byhydrodynamic injection of the integrase construct andFAH-att but under selective pressure the FAH+ hep-atocytes expanded to approximately 70 of the livermass (~3 107 hepatocytes) Group 1 mice (n = 3)treated with FAH-att and integrase underwent a periodof selection for ~90 days We then used these long-termsurvivors as hepatocyte donors for serial transplantationinto naRve Fah secondary recipients After therecipient mice survived 90+ days off NTBC we usedthem as hepatocyte donors for a second round of serialtransplantation Tertiary Fah mutant recipients alsounderwent a selection period of 90 days off NTBCLiver tissue harvested and analyzed after the selectionperiod showed high levels of repopulation in thetertiary recipients (N60 by immunohistochemistry)(Fig 2B) All FAH-positive hepatocytes were healthyand showed no signs of dysplasia Additionally weperformed liver function tests on eight tertiary recipientmice The average alanine aminotransferase level was232 unitsL (ranging from 90 to 755 unitsL) which isslightly elevated over normal (100 unitsL) The crea-tinine and conjugated bilirubin levels were 05 mgLand 0 mgdl respectively which are similar to normallevels (creatinine = 0 mgL and bilirubin = 0 mgdl) Onaverage untreated Fah mice have alanine amino-transferase levels of 1000 unitsL creatinine levels of15 mgL and conjugated bilirubin levels of 05 mgdl[25]
TABLE 2 Effect of integrase lev
Group A
Plasmid combinations 25 Ag FAH-att 25 Agintegrase (n = 30)
25 Aginteg
Absolute integration frequency 73 F 38 104 11 FMinimum 14800
Median 11374Maximum 1465
Corrected integration frequency 36
127
Fah mice were treated with varying ratios of integrase to FAH expression construct FAH+ he
(worst animal) and maximum (best animal) frequencies in each group are shown
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
We extracted genomic liver DNA from three repopu-lated (~50ndash70 by immunohistochemistry) serial trans-plant recipients All three recipient mice had differentprimary hepatocyte donors which had been injectedwith the integrase and FAH-att constructs We analyzedintegration sites for the three mice by Southern blot afterdigestion with HindIII or ScaI and probed them using aportion of the human FAH cDNA The previously knownintegration site (mpsL1) was held in common betweenall three mice however several distinct junction frag-ments were observed in addition In mouse 1 DNAdigested with HindIII we identified two bmajor b sites ofintegration (77 and 68 kb) and four or five additionalbminorQ sites based upon band intensity (lane 6 Fig 4)To confirm these integration sites we performed asecond Southern blot on the same mouse 1 DNAdigested with ScaI Again two major sites were identified(60 and 48 kb) (lane 7 Fig 4) We also digested DNAfrom mice 2 and 3 with ScaI (lanes 8 and 9 Fig 4)Mouse 2 had two major sites of integration (60 and68 kb) and mouse 3 had three sites of integration (6068 and 86 kb) The 60-kb band corresponds to thempsL1 and was shared between all mice To identify thenumber of FAH gene copies per cell we performed athird Southern blot using the restriction enzyme PstIwhich cuts twice in FAH leaving an internal fragmentMouse 1 showed greater than 1 copy of FAH per diploidgenome equivalent compared to its matched control(lanes 10 and 11 Fig 4) while mice 2 and 3 bothshowed less than 1 copy of FAH per diploid genomecompared to their matched control (lanes 12 13 and 14Fig 4)
We used a nested inverse PCR approach on genomicDNA from mouse 1 to identify the specific integrationjunctions shown in the HindIII Southern blot (lane 6Fig 4) We identified seven different integration sitesThe vectorndashgenome junctions and their locations withinthe genome are listed in Table 3 In all seven cases theswitch from the attB sequence of the vector to thegenomic sequence occurred at or near a TT coreconfirming the use of fC31-mediated integration The
els on integration frequency
B C D
FAH-att 5 Agrase (n = 8)
25 Ag FAH-att 25 Agintegrase (n = 6)
25 Ag FAH-att 125 Agintegrase (n = 5)
14 104 52 F 66 105 29 F 43 105
0 0 0
19375 119201 13409112558 16101 19784
055 026 015
1181 1385 1690
patocytes and nodules were counted as single integration events In addition the minimum
403
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

TABLE 1 Absolute and actual integration frequencies of integrase-treated mice
Group 1 2
Plasmid combinations 25 Ag FAH-att 25 Ag integrase (n = 30) 25 Ag FAH-att 25 Ag empty vector (n = 9)
Absolute integration frequency 73 F 38 104 28 F 44 105
Minimum 14880 0
Median 11374 135640Maximum 1465 17165
Corrected integration frequency 36 014
127 1714
FAH + hepatocytes and nodules were counted as single integration events Averages with standard deviations are given In addition the minimum (worst animal) and maximum (best
animal) frequencies in each group are shown
ARTICLE doi101016jymthe200411001
to be approximately 5ndash10 [10] similar to that observedwith SB transposase Pseudo attP sites are rare in both thehuman and the murine genome and therefore thenumber of integration sites is limited In one settingonly two integrations were identified in vivo within themouse genome [10] Therefore fC31 integrase-mediatedintegration is nonrandom and efficient enough to makethe system very attractive for nonviral gene therapy ofgenetic disorders Several papers have been publishedshowing in vivo applications of fC31 integrase Onestudy showed the delivery and site-specific integration ofthe human Factor IX transgene in mouse liver in vivoresulting in production of therapeutic Factor IX levels[10] Further studies reported the ability of fC31 inte-grase to transfect human keratinocytes and to correctinherited skin diseases in a xenotransplantation model[1112]
To characterize further the properties of fC31 inte-grase-mediated gene transfer in vivo we used the murinemodel of hereditary tyrosinemia type 1 (HT1) [13] Thissystem has unique advantages for the characterization ofstable integration events in hepatocytes in vivo HT1 is agenetic liver disease caused by deficiency of fumarylace-toacetate hydrolase (FAH) [13] the enzyme that catalyzesthe last step in the tyrosine degradation pathway In HT1FAH-expressing hepatocytes display a strong proliferativeadvantage and eventually repopulate the entire diseasedliver [1415] Because of this selective advantage hepato-
FIG 1 Frequency of fC31 integrase-medi-
ated stable FAH gene transfer The frequency
of integration in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of sections
400
cytes in which fC31 integrase has mediated integrationof the FAH gene can be identified counted andhistologically evaluated soon after transfection as wellas after several rounds of growth and expansion of FAH+
hepatocyte clonesHere we report the properties of fC31 integrase-
mediated gene transfer in the HT1 mouse model
RESULTS
Frequency of Stable Integration and HistologicalEvaluationThe efficiency of gene transfer into the liver is typicallydetermined by the expression of histochemical markerssuch as h-galactosidase or alkaline phosphatase [2627]However the simple presence or absence of such markersdoes not accurately measure integration frequencybecause gene expression can occur in both the presenceand the absence of chromosomal integration [28] In theFah mouse model single FAH-expressing hepatocytescan selectively expand and form clonal nodules only ifstable integration occurred in the initially transfectedcells [1429] This model is therefore well suited tomeasure the frequency of vector integration and stabletransgene expression
We calculated the integration frequency for experi-mental plasmids and controls We injected group 1 (n =10) with a FAH-expressing construct containing the attB
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therap
y
ARTICLEdoi101016jymthe200411001
site (FAH-att) plus the integrase expression constructacting in trans We treated group 2 (n = 5) with FAH-attplus an empty vector lacking integrase We used apreviously described 11 ratio (25 Ag of each plasmid) ofintegrase (or empty vector) to the FAH expressionconstruct (FAH-att) [10] On day 3 after plasmid injec-tion we discontinued 2-(2-nitro-4-trifluoromethylben-zoyl)-13-cyclohexanedione (NTBC) treatment to permitthe development of liver disease and the selection ofFAH+ hepatocyte nodules We sacrificed all animals 25days postinjection and processed the liver tissue forhistological evaluation We scored several liver sectionsfor each mouse for FAH+ cells and nodules The totalnumber of sections evaluated for each group was 30 forgroup 1 and 9 for group 2 (Table 1 Fig 1) Approximately2 to 3 sections were scored for each individual mousewithin group 1 or 2
In the liver sections from group 1 (mice treated withFAH-att and integrase) the average absolute frequency ofFAH+ nodules was 73 F 38 104 (11374 hepatocytes)The frequency ranged over 10-fold from a maximum of 1465 cells in the best mouse to 14880 in the worst Group2 (FAH-att and an empty vector) displayed minimalintegration at only 28 F 44 105 (135640 cells)and was significantly different from group 1 as evaluatedby a Student t test ( P value b00001) To determine theefficiency of the fC31 integrase-mediated integration ona per-cell basis the DNA transfection frequency must beconsidered To measure this parameter we injected threeFah mice via the tail vein with a h-galactosidaseexpression construct and sacrificed them 3 days laterWe stained liver sections for h-galactosidase and calcu-lated the hepatocyte transfection efficiency at ~2 (datanot shown) This number confirms that Fah mutant
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
mice have lower rates of transfection than typically seenafter hydrodynamic injection [5] The average correctedintegration frequency for group 1 mice was 36 (127)ranging from 1 to 107 Group 2 mice had an averageper-cell integration frequency of 01 (1714) rangingfrom 0 to 06 (Table 1 Fig 1)
Integrase Induced a Higher Frequency of HepatocyteDysplasiaFAH+ hepatocytes observed in histological sections fromgroup 1 mice harvested after 25 days of selectiondisplayed a unique dysplastic phenotype at a higherfrequency than controls Specifically individual FAH+
hepatocytes were often markedly enlarged in compari-son to adjacent FAH-negative hepatocytes (Fig 2A) Inaddition many of the nuclei in these abnormal cellswere massively enlarged giving the hepatocytes abdysplasticQ appearance We devised an index ratingsystem to quantitate the presence of these abnormalcells Nodules in which more than half of the hepato-cytes were clearly abnormal in phenotype were scoredwith a 3+ rating while nodules in which less than half ofthe hepatocytes were abnormal were scored with a 2+rating Nodules that contained only morphologicallynormal hepatocytes were given a 1+ rating (Figs 2D 2Eand 2F) Seventy percent of the nodules in group 1 micecontained abnormal cells (2+ and 3+) (Fig 3) Group 2mice which did not receive integrase did not have any3+ nodules but still had approximately 25 2+ noduleswhich contain a lower fraction of abnormal cells (Figs2C and 3) As an additional control we also injectedFah mice with the Sleeping Beauty transposon andtheir scores paralleled those of the integrase-transfectedmice after sacrifice on day 25 [5] Specifically 30 Ag of
FIG 2 FAH immunohistochemistry in hepa-
tocytes treated with fC31 integrase (A) Sec-
tion from a mouse treated with FAH-att and
integrase after 25 days of selection Arrows
denote hepatocytes with abnormal morpho-
logy (B) Mouse treated with FAH-att and inte-
grase after 90+ days of selection (C) Mouse
treated with FAH-att and empty vector after
25 days of selection (D) Nodule scored as a 1+
in the dysplasia index (no abnormal FAH+
cells) (E) Nodule scored as a 2+ in the
dysplasia index (less than half of the FAH+ cells
are abnormal) (F) Nodule scored as a 3+ in the
dysplasia index (more than half of the FAH+
cells are abnormal)
401
ARTICLE doi101016jymthe200411001
the transposon containing the FAH gene (driven by theSRa promoter) and 1 Ag of the transposase were deliveredtogether by hydrodynamic injection into Fah mice aspreviously described [5] Mice treated with the trans-poson showed hepatocyte dysplasia in a smaller percent-age of the nodules similar to the group 2 controlswhich did not receive any integrase plasmid Approxi-mately 75 of the nodules were healthy (1+) and 25 ofthe nodules were scored as 2+ (Fig 3) The 2+ nodulesfound in group 2 with no integrase and in the SleepingBeauty transposon control suggested that a level ofhepatocyte dysplasia is attributable to the Fah deficiencyitself FAH overexpression andor the hydrodynamicinjection method
In fact prevalent hepatocyte dysplasia as seen in the3+ nodules is a common feature of Fah mutant liverduring NTBC withdrawal without correction [2430]Here we withdrew NTBC 3 days after plasmid injectionand it is therefore possible that this time was too shortto initiate FAH expression To determine whetherremoval of NTBC prior to full FAH expression was thecause of injury and dysplasia in the integrase-trans-fected hepatocytes we gave three additional group 1mice maximal NTBC therapy during and after plasmidinjection They were treated with a higher dose ofNTBC (30 mgL drinking watermdashfour times the normalamount) for a longer period (1 week instead of 3 days)However this did not change the frequency of FAHintegration or the dysplasia index The average integra-tion frequency for these three mice was 74 F 18 104 (11332) and ~35 of the nodules were scored ashighly abnormal (3+)
Importantly the abnormal phenotype describedabove did not persist long term (Fig 2B) Integrase-treated (n = 4) mice sacrificed after more than 90 days of
FIG 3 Nodule dysplasia index in hepatocytes
treated with fC31 integrase The nodule
dysplasia index in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of nodules scored
402
hepatocyte selection had ~70 liver repopulation withonly normal healthy hepatocytes as seen in Fig 2 Thisobservation indicates that the abnormal hepatocytephenotype found 25 days after gene therapy is transientIt is currently unknown whether individual abnormalFAH+ hepatocytes revert to normal cell morphology orwhether normal FAH+ hepatocytes have a selectiveadvantage and simply outgrow the dysplastic cells overtime
Effects of Different Levels of IntegraseTo determine whether the high levels of integraseplasmid administered were the cause of the higherfrequency of abnormal hepatocytes we performedadditional experiments to define the optimal amountsof integrase plasmid for effective gene therapy Weinjected four different groups of Fah mice with thefollowing dilutions of integrase group A (n = 10) wasinjected with 25 Ag of both plasmids integrase andFAH-att (11 ratio) group B (n = 8) received 5 Ag ofintegrase and 25 Ag of FAH-att (15 ratio) group C (n =6) 25 Ag of the integrase and 25 Ag of FAH-att (110ratio) and group D (n = 5) 125 Ag of integrase and25 Ag of FAH-att (120) We evaluated liver sectionsfrom each mouse for integration frequency and mor-phology 25 days after removal from NTBC We saw thehighest percentage of abnormal 3+ nodules in micefrom group A receiving a 11 ratio of Fah-att tointegrase (Fig 3) The proportion of 3+ nodulesdecreased and the frequency of 1+ normal nodulesincreased with decreasing amount of the integraseplasmid Group D mice receiving only 125 Ag of theintegrase plasmid had no abnormal nodules Thus theamount of the integrase plasmid administered corre-lated directly with the frequency of morphologically
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
abnormal hepatocytes The quantity of integrase plas-mid was also strongly correlated with the level of genetransfer The average absolute integration frequencyranged from 11374 with a 11 ratio (group A) toonly 134091 with a 120 ratio (group D) Thesefindings are summarized in Table 2 and Fig 1
Integration Site AnalysisOne major pseudo attP site (mpsL1) favored forintegration into the mouse genome has been describedpreviously [10] Here we determined whether integra-tion into other sites also occurred We stably trans-fected approximately 11000 hepatocytes byhydrodynamic injection of the integrase construct andFAH-att but under selective pressure the FAH+ hep-atocytes expanded to approximately 70 of the livermass (~3 107 hepatocytes) Group 1 mice (n = 3)treated with FAH-att and integrase underwent a periodof selection for ~90 days We then used these long-termsurvivors as hepatocyte donors for serial transplantationinto naRve Fah secondary recipients After therecipient mice survived 90+ days off NTBC we usedthem as hepatocyte donors for a second round of serialtransplantation Tertiary Fah mutant recipients alsounderwent a selection period of 90 days off NTBCLiver tissue harvested and analyzed after the selectionperiod showed high levels of repopulation in thetertiary recipients (N60 by immunohistochemistry)(Fig 2B) All FAH-positive hepatocytes were healthyand showed no signs of dysplasia Additionally weperformed liver function tests on eight tertiary recipientmice The average alanine aminotransferase level was232 unitsL (ranging from 90 to 755 unitsL) which isslightly elevated over normal (100 unitsL) The crea-tinine and conjugated bilirubin levels were 05 mgLand 0 mgdl respectively which are similar to normallevels (creatinine = 0 mgL and bilirubin = 0 mgdl) Onaverage untreated Fah mice have alanine amino-transferase levels of 1000 unitsL creatinine levels of15 mgL and conjugated bilirubin levels of 05 mgdl[25]
TABLE 2 Effect of integrase lev
Group A
Plasmid combinations 25 Ag FAH-att 25 Agintegrase (n = 30)
25 Aginteg
Absolute integration frequency 73 F 38 104 11 FMinimum 14800
Median 11374Maximum 1465
Corrected integration frequency 36
127
Fah mice were treated with varying ratios of integrase to FAH expression construct FAH+ he
(worst animal) and maximum (best animal) frequencies in each group are shown
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
We extracted genomic liver DNA from three repopu-lated (~50ndash70 by immunohistochemistry) serial trans-plant recipients All three recipient mice had differentprimary hepatocyte donors which had been injectedwith the integrase and FAH-att constructs We analyzedintegration sites for the three mice by Southern blot afterdigestion with HindIII or ScaI and probed them using aportion of the human FAH cDNA The previously knownintegration site (mpsL1) was held in common betweenall three mice however several distinct junction frag-ments were observed in addition In mouse 1 DNAdigested with HindIII we identified two bmajor b sites ofintegration (77 and 68 kb) and four or five additionalbminorQ sites based upon band intensity (lane 6 Fig 4)To confirm these integration sites we performed asecond Southern blot on the same mouse 1 DNAdigested with ScaI Again two major sites were identified(60 and 48 kb) (lane 7 Fig 4) We also digested DNAfrom mice 2 and 3 with ScaI (lanes 8 and 9 Fig 4)Mouse 2 had two major sites of integration (60 and68 kb) and mouse 3 had three sites of integration (6068 and 86 kb) The 60-kb band corresponds to thempsL1 and was shared between all mice To identify thenumber of FAH gene copies per cell we performed athird Southern blot using the restriction enzyme PstIwhich cuts twice in FAH leaving an internal fragmentMouse 1 showed greater than 1 copy of FAH per diploidgenome equivalent compared to its matched control(lanes 10 and 11 Fig 4) while mice 2 and 3 bothshowed less than 1 copy of FAH per diploid genomecompared to their matched control (lanes 12 13 and 14Fig 4)
We used a nested inverse PCR approach on genomicDNA from mouse 1 to identify the specific integrationjunctions shown in the HindIII Southern blot (lane 6Fig 4) We identified seven different integration sitesThe vectorndashgenome junctions and their locations withinthe genome are listed in Table 3 In all seven cases theswitch from the attB sequence of the vector to thegenomic sequence occurred at or near a TT coreconfirming the use of fC31-mediated integration The
els on integration frequency
B C D
FAH-att 5 Agrase (n = 8)
25 Ag FAH-att 25 Agintegrase (n = 6)
25 Ag FAH-att 125 Agintegrase (n = 5)
14 104 52 F 66 105 29 F 43 105
0 0 0
19375 119201 13409112558 16101 19784
055 026 015
1181 1385 1690
patocytes and nodules were counted as single integration events In addition the minimum
403
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
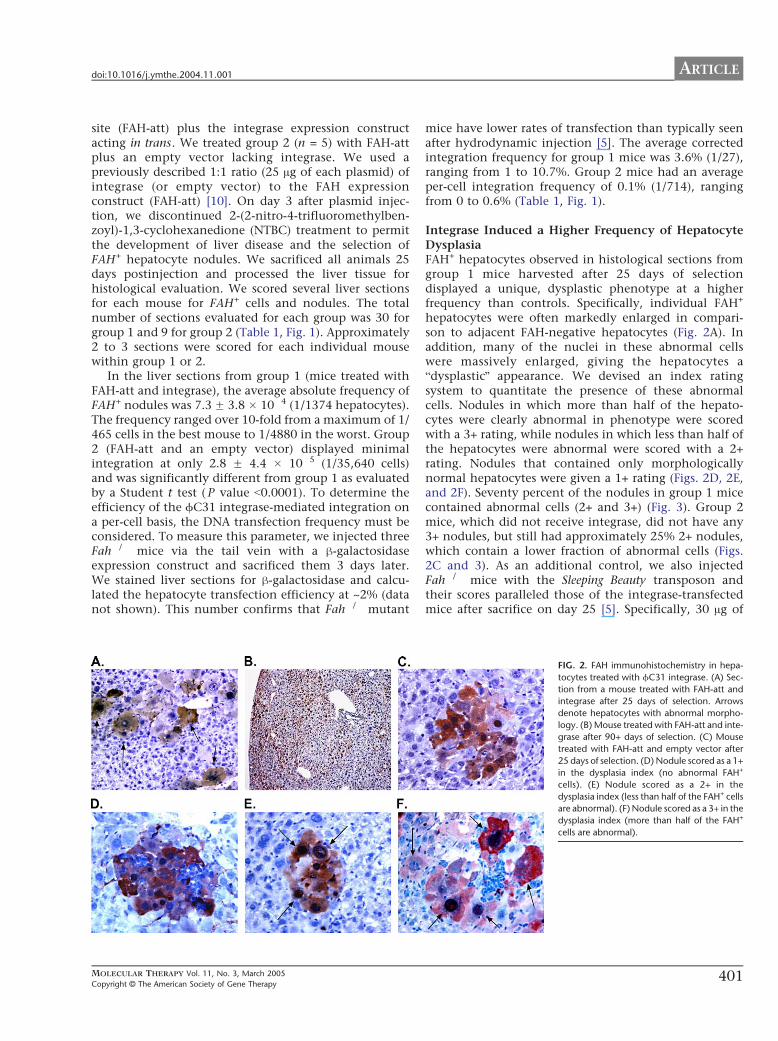
ARTICLEdoi101016jymthe200411001
site (FAH-att) plus the integrase expression constructacting in trans We treated group 2 (n = 5) with FAH-attplus an empty vector lacking integrase We used apreviously described 11 ratio (25 Ag of each plasmid) ofintegrase (or empty vector) to the FAH expressionconstruct (FAH-att) [10] On day 3 after plasmid injec-tion we discontinued 2-(2-nitro-4-trifluoromethylben-zoyl)-13-cyclohexanedione (NTBC) treatment to permitthe development of liver disease and the selection ofFAH+ hepatocyte nodules We sacrificed all animals 25days postinjection and processed the liver tissue forhistological evaluation We scored several liver sectionsfor each mouse for FAH+ cells and nodules The totalnumber of sections evaluated for each group was 30 forgroup 1 and 9 for group 2 (Table 1 Fig 1) Approximately2 to 3 sections were scored for each individual mousewithin group 1 or 2
In the liver sections from group 1 (mice treated withFAH-att and integrase) the average absolute frequency ofFAH+ nodules was 73 F 38 104 (11374 hepatocytes)The frequency ranged over 10-fold from a maximum of 1465 cells in the best mouse to 14880 in the worst Group2 (FAH-att and an empty vector) displayed minimalintegration at only 28 F 44 105 (135640 cells)and was significantly different from group 1 as evaluatedby a Student t test ( P value b00001) To determine theefficiency of the fC31 integrase-mediated integration ona per-cell basis the DNA transfection frequency must beconsidered To measure this parameter we injected threeFah mice via the tail vein with a h-galactosidaseexpression construct and sacrificed them 3 days laterWe stained liver sections for h-galactosidase and calcu-lated the hepatocyte transfection efficiency at ~2 (datanot shown) This number confirms that Fah mutant
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
mice have lower rates of transfection than typically seenafter hydrodynamic injection [5] The average correctedintegration frequency for group 1 mice was 36 (127)ranging from 1 to 107 Group 2 mice had an averageper-cell integration frequency of 01 (1714) rangingfrom 0 to 06 (Table 1 Fig 1)
Integrase Induced a Higher Frequency of HepatocyteDysplasiaFAH+ hepatocytes observed in histological sections fromgroup 1 mice harvested after 25 days of selectiondisplayed a unique dysplastic phenotype at a higherfrequency than controls Specifically individual FAH+
hepatocytes were often markedly enlarged in compari-son to adjacent FAH-negative hepatocytes (Fig 2A) Inaddition many of the nuclei in these abnormal cellswere massively enlarged giving the hepatocytes abdysplasticQ appearance We devised an index ratingsystem to quantitate the presence of these abnormalcells Nodules in which more than half of the hepato-cytes were clearly abnormal in phenotype were scoredwith a 3+ rating while nodules in which less than half ofthe hepatocytes were abnormal were scored with a 2+rating Nodules that contained only morphologicallynormal hepatocytes were given a 1+ rating (Figs 2D 2Eand 2F) Seventy percent of the nodules in group 1 micecontained abnormal cells (2+ and 3+) (Fig 3) Group 2mice which did not receive integrase did not have any3+ nodules but still had approximately 25 2+ noduleswhich contain a lower fraction of abnormal cells (Figs2C and 3) As an additional control we also injectedFah mice with the Sleeping Beauty transposon andtheir scores paralleled those of the integrase-transfectedmice after sacrifice on day 25 [5] Specifically 30 Ag of
FIG 2 FAH immunohistochemistry in hepa-
tocytes treated with fC31 integrase (A) Sec-
tion from a mouse treated with FAH-att and
integrase after 25 days of selection Arrows
denote hepatocytes with abnormal morpho-
logy (B) Mouse treated with FAH-att and inte-
grase after 90+ days of selection (C) Mouse
treated with FAH-att and empty vector after
25 days of selection (D) Nodule scored as a 1+
in the dysplasia index (no abnormal FAH+
cells) (E) Nodule scored as a 2+ in the
dysplasia index (less than half of the FAH+ cells
are abnormal) (F) Nodule scored as a 3+ in the
dysplasia index (more than half of the FAH+
cells are abnormal)
401
ARTICLE doi101016jymthe200411001
the transposon containing the FAH gene (driven by theSRa promoter) and 1 Ag of the transposase were deliveredtogether by hydrodynamic injection into Fah mice aspreviously described [5] Mice treated with the trans-poson showed hepatocyte dysplasia in a smaller percent-age of the nodules similar to the group 2 controlswhich did not receive any integrase plasmid Approxi-mately 75 of the nodules were healthy (1+) and 25 ofthe nodules were scored as 2+ (Fig 3) The 2+ nodulesfound in group 2 with no integrase and in the SleepingBeauty transposon control suggested that a level ofhepatocyte dysplasia is attributable to the Fah deficiencyitself FAH overexpression andor the hydrodynamicinjection method
In fact prevalent hepatocyte dysplasia as seen in the3+ nodules is a common feature of Fah mutant liverduring NTBC withdrawal without correction [2430]Here we withdrew NTBC 3 days after plasmid injectionand it is therefore possible that this time was too shortto initiate FAH expression To determine whetherremoval of NTBC prior to full FAH expression was thecause of injury and dysplasia in the integrase-trans-fected hepatocytes we gave three additional group 1mice maximal NTBC therapy during and after plasmidinjection They were treated with a higher dose ofNTBC (30 mgL drinking watermdashfour times the normalamount) for a longer period (1 week instead of 3 days)However this did not change the frequency of FAHintegration or the dysplasia index The average integra-tion frequency for these three mice was 74 F 18 104 (11332) and ~35 of the nodules were scored ashighly abnormal (3+)
Importantly the abnormal phenotype describedabove did not persist long term (Fig 2B) Integrase-treated (n = 4) mice sacrificed after more than 90 days of
FIG 3 Nodule dysplasia index in hepatocytes
treated with fC31 integrase The nodule
dysplasia index in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of nodules scored
402
hepatocyte selection had ~70 liver repopulation withonly normal healthy hepatocytes as seen in Fig 2 Thisobservation indicates that the abnormal hepatocytephenotype found 25 days after gene therapy is transientIt is currently unknown whether individual abnormalFAH+ hepatocytes revert to normal cell morphology orwhether normal FAH+ hepatocytes have a selectiveadvantage and simply outgrow the dysplastic cells overtime
Effects of Different Levels of IntegraseTo determine whether the high levels of integraseplasmid administered were the cause of the higherfrequency of abnormal hepatocytes we performedadditional experiments to define the optimal amountsof integrase plasmid for effective gene therapy Weinjected four different groups of Fah mice with thefollowing dilutions of integrase group A (n = 10) wasinjected with 25 Ag of both plasmids integrase andFAH-att (11 ratio) group B (n = 8) received 5 Ag ofintegrase and 25 Ag of FAH-att (15 ratio) group C (n =6) 25 Ag of the integrase and 25 Ag of FAH-att (110ratio) and group D (n = 5) 125 Ag of integrase and25 Ag of FAH-att (120) We evaluated liver sectionsfrom each mouse for integration frequency and mor-phology 25 days after removal from NTBC We saw thehighest percentage of abnormal 3+ nodules in micefrom group A receiving a 11 ratio of Fah-att tointegrase (Fig 3) The proportion of 3+ nodulesdecreased and the frequency of 1+ normal nodulesincreased with decreasing amount of the integraseplasmid Group D mice receiving only 125 Ag of theintegrase plasmid had no abnormal nodules Thus theamount of the integrase plasmid administered corre-lated directly with the frequency of morphologically
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
abnormal hepatocytes The quantity of integrase plas-mid was also strongly correlated with the level of genetransfer The average absolute integration frequencyranged from 11374 with a 11 ratio (group A) toonly 134091 with a 120 ratio (group D) Thesefindings are summarized in Table 2 and Fig 1
Integration Site AnalysisOne major pseudo attP site (mpsL1) favored forintegration into the mouse genome has been describedpreviously [10] Here we determined whether integra-tion into other sites also occurred We stably trans-fected approximately 11000 hepatocytes byhydrodynamic injection of the integrase construct andFAH-att but under selective pressure the FAH+ hep-atocytes expanded to approximately 70 of the livermass (~3 107 hepatocytes) Group 1 mice (n = 3)treated with FAH-att and integrase underwent a periodof selection for ~90 days We then used these long-termsurvivors as hepatocyte donors for serial transplantationinto naRve Fah secondary recipients After therecipient mice survived 90+ days off NTBC we usedthem as hepatocyte donors for a second round of serialtransplantation Tertiary Fah mutant recipients alsounderwent a selection period of 90 days off NTBCLiver tissue harvested and analyzed after the selectionperiod showed high levels of repopulation in thetertiary recipients (N60 by immunohistochemistry)(Fig 2B) All FAH-positive hepatocytes were healthyand showed no signs of dysplasia Additionally weperformed liver function tests on eight tertiary recipientmice The average alanine aminotransferase level was232 unitsL (ranging from 90 to 755 unitsL) which isslightly elevated over normal (100 unitsL) The crea-tinine and conjugated bilirubin levels were 05 mgLand 0 mgdl respectively which are similar to normallevels (creatinine = 0 mgL and bilirubin = 0 mgdl) Onaverage untreated Fah mice have alanine amino-transferase levels of 1000 unitsL creatinine levels of15 mgL and conjugated bilirubin levels of 05 mgdl[25]
TABLE 2 Effect of integrase lev
Group A
Plasmid combinations 25 Ag FAH-att 25 Agintegrase (n = 30)
25 Aginteg
Absolute integration frequency 73 F 38 104 11 FMinimum 14800
Median 11374Maximum 1465
Corrected integration frequency 36
127
Fah mice were treated with varying ratios of integrase to FAH expression construct FAH+ he
(worst animal) and maximum (best animal) frequencies in each group are shown
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
We extracted genomic liver DNA from three repopu-lated (~50ndash70 by immunohistochemistry) serial trans-plant recipients All three recipient mice had differentprimary hepatocyte donors which had been injectedwith the integrase and FAH-att constructs We analyzedintegration sites for the three mice by Southern blot afterdigestion with HindIII or ScaI and probed them using aportion of the human FAH cDNA The previously knownintegration site (mpsL1) was held in common betweenall three mice however several distinct junction frag-ments were observed in addition In mouse 1 DNAdigested with HindIII we identified two bmajor b sites ofintegration (77 and 68 kb) and four or five additionalbminorQ sites based upon band intensity (lane 6 Fig 4)To confirm these integration sites we performed asecond Southern blot on the same mouse 1 DNAdigested with ScaI Again two major sites were identified(60 and 48 kb) (lane 7 Fig 4) We also digested DNAfrom mice 2 and 3 with ScaI (lanes 8 and 9 Fig 4)Mouse 2 had two major sites of integration (60 and68 kb) and mouse 3 had three sites of integration (6068 and 86 kb) The 60-kb band corresponds to thempsL1 and was shared between all mice To identify thenumber of FAH gene copies per cell we performed athird Southern blot using the restriction enzyme PstIwhich cuts twice in FAH leaving an internal fragmentMouse 1 showed greater than 1 copy of FAH per diploidgenome equivalent compared to its matched control(lanes 10 and 11 Fig 4) while mice 2 and 3 bothshowed less than 1 copy of FAH per diploid genomecompared to their matched control (lanes 12 13 and 14Fig 4)
We used a nested inverse PCR approach on genomicDNA from mouse 1 to identify the specific integrationjunctions shown in the HindIII Southern blot (lane 6Fig 4) We identified seven different integration sitesThe vectorndashgenome junctions and their locations withinthe genome are listed in Table 3 In all seven cases theswitch from the attB sequence of the vector to thegenomic sequence occurred at or near a TT coreconfirming the use of fC31-mediated integration The
els on integration frequency
B C D
FAH-att 5 Agrase (n = 8)
25 Ag FAH-att 25 Agintegrase (n = 6)
25 Ag FAH-att 125 Agintegrase (n = 5)
14 104 52 F 66 105 29 F 43 105
0 0 0
19375 119201 13409112558 16101 19784
055 026 015
1181 1385 1690
patocytes and nodules were counted as single integration events In addition the minimum
403
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
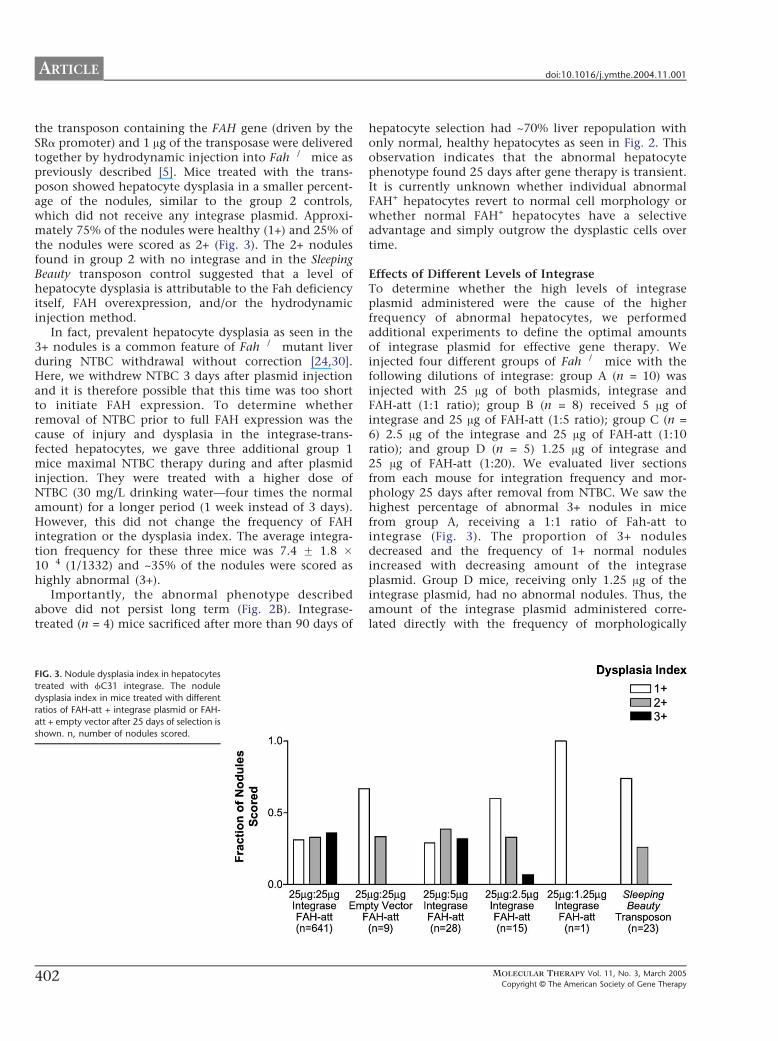
ARTICLE doi101016jymthe200411001
the transposon containing the FAH gene (driven by theSRa promoter) and 1 Ag of the transposase were deliveredtogether by hydrodynamic injection into Fah mice aspreviously described [5] Mice treated with the trans-poson showed hepatocyte dysplasia in a smaller percent-age of the nodules similar to the group 2 controlswhich did not receive any integrase plasmid Approxi-mately 75 of the nodules were healthy (1+) and 25 ofthe nodules were scored as 2+ (Fig 3) The 2+ nodulesfound in group 2 with no integrase and in the SleepingBeauty transposon control suggested that a level ofhepatocyte dysplasia is attributable to the Fah deficiencyitself FAH overexpression andor the hydrodynamicinjection method
In fact prevalent hepatocyte dysplasia as seen in the3+ nodules is a common feature of Fah mutant liverduring NTBC withdrawal without correction [2430]Here we withdrew NTBC 3 days after plasmid injectionand it is therefore possible that this time was too shortto initiate FAH expression To determine whetherremoval of NTBC prior to full FAH expression was thecause of injury and dysplasia in the integrase-trans-fected hepatocytes we gave three additional group 1mice maximal NTBC therapy during and after plasmidinjection They were treated with a higher dose ofNTBC (30 mgL drinking watermdashfour times the normalamount) for a longer period (1 week instead of 3 days)However this did not change the frequency of FAHintegration or the dysplasia index The average integra-tion frequency for these three mice was 74 F 18 104 (11332) and ~35 of the nodules were scored ashighly abnormal (3+)
Importantly the abnormal phenotype describedabove did not persist long term (Fig 2B) Integrase-treated (n = 4) mice sacrificed after more than 90 days of
FIG 3 Nodule dysplasia index in hepatocytes
treated with fC31 integrase The nodule
dysplasia index in mice treated with different
ratios of FAH-att + integrase plasmid or FAH-
att + empty vector after 25 days of selection is
shown n number of nodules scored
402
hepatocyte selection had ~70 liver repopulation withonly normal healthy hepatocytes as seen in Fig 2 Thisobservation indicates that the abnormal hepatocytephenotype found 25 days after gene therapy is transientIt is currently unknown whether individual abnormalFAH+ hepatocytes revert to normal cell morphology orwhether normal FAH+ hepatocytes have a selectiveadvantage and simply outgrow the dysplastic cells overtime
Effects of Different Levels of IntegraseTo determine whether the high levels of integraseplasmid administered were the cause of the higherfrequency of abnormal hepatocytes we performedadditional experiments to define the optimal amountsof integrase plasmid for effective gene therapy Weinjected four different groups of Fah mice with thefollowing dilutions of integrase group A (n = 10) wasinjected with 25 Ag of both plasmids integrase andFAH-att (11 ratio) group B (n = 8) received 5 Ag ofintegrase and 25 Ag of FAH-att (15 ratio) group C (n =6) 25 Ag of the integrase and 25 Ag of FAH-att (110ratio) and group D (n = 5) 125 Ag of integrase and25 Ag of FAH-att (120) We evaluated liver sectionsfrom each mouse for integration frequency and mor-phology 25 days after removal from NTBC We saw thehighest percentage of abnormal 3+ nodules in micefrom group A receiving a 11 ratio of Fah-att tointegrase (Fig 3) The proportion of 3+ nodulesdecreased and the frequency of 1+ normal nodulesincreased with decreasing amount of the integraseplasmid Group D mice receiving only 125 Ag of theintegrase plasmid had no abnormal nodules Thus theamount of the integrase plasmid administered corre-lated directly with the frequency of morphologically
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
abnormal hepatocytes The quantity of integrase plas-mid was also strongly correlated with the level of genetransfer The average absolute integration frequencyranged from 11374 with a 11 ratio (group A) toonly 134091 with a 120 ratio (group D) Thesefindings are summarized in Table 2 and Fig 1
Integration Site AnalysisOne major pseudo attP site (mpsL1) favored forintegration into the mouse genome has been describedpreviously [10] Here we determined whether integra-tion into other sites also occurred We stably trans-fected approximately 11000 hepatocytes byhydrodynamic injection of the integrase construct andFAH-att but under selective pressure the FAH+ hep-atocytes expanded to approximately 70 of the livermass (~3 107 hepatocytes) Group 1 mice (n = 3)treated with FAH-att and integrase underwent a periodof selection for ~90 days We then used these long-termsurvivors as hepatocyte donors for serial transplantationinto naRve Fah secondary recipients After therecipient mice survived 90+ days off NTBC we usedthem as hepatocyte donors for a second round of serialtransplantation Tertiary Fah mutant recipients alsounderwent a selection period of 90 days off NTBCLiver tissue harvested and analyzed after the selectionperiod showed high levels of repopulation in thetertiary recipients (N60 by immunohistochemistry)(Fig 2B) All FAH-positive hepatocytes were healthyand showed no signs of dysplasia Additionally weperformed liver function tests on eight tertiary recipientmice The average alanine aminotransferase level was232 unitsL (ranging from 90 to 755 unitsL) which isslightly elevated over normal (100 unitsL) The crea-tinine and conjugated bilirubin levels were 05 mgLand 0 mgdl respectively which are similar to normallevels (creatinine = 0 mgL and bilirubin = 0 mgdl) Onaverage untreated Fah mice have alanine amino-transferase levels of 1000 unitsL creatinine levels of15 mgL and conjugated bilirubin levels of 05 mgdl[25]
TABLE 2 Effect of integrase lev
Group A
Plasmid combinations 25 Ag FAH-att 25 Agintegrase (n = 30)
25 Aginteg
Absolute integration frequency 73 F 38 104 11 FMinimum 14800
Median 11374Maximum 1465
Corrected integration frequency 36
127
Fah mice were treated with varying ratios of integrase to FAH expression construct FAH+ he
(worst animal) and maximum (best animal) frequencies in each group are shown
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
We extracted genomic liver DNA from three repopu-lated (~50ndash70 by immunohistochemistry) serial trans-plant recipients All three recipient mice had differentprimary hepatocyte donors which had been injectedwith the integrase and FAH-att constructs We analyzedintegration sites for the three mice by Southern blot afterdigestion with HindIII or ScaI and probed them using aportion of the human FAH cDNA The previously knownintegration site (mpsL1) was held in common betweenall three mice however several distinct junction frag-ments were observed in addition In mouse 1 DNAdigested with HindIII we identified two bmajor b sites ofintegration (77 and 68 kb) and four or five additionalbminorQ sites based upon band intensity (lane 6 Fig 4)To confirm these integration sites we performed asecond Southern blot on the same mouse 1 DNAdigested with ScaI Again two major sites were identified(60 and 48 kb) (lane 7 Fig 4) We also digested DNAfrom mice 2 and 3 with ScaI (lanes 8 and 9 Fig 4)Mouse 2 had two major sites of integration (60 and68 kb) and mouse 3 had three sites of integration (6068 and 86 kb) The 60-kb band corresponds to thempsL1 and was shared between all mice To identify thenumber of FAH gene copies per cell we performed athird Southern blot using the restriction enzyme PstIwhich cuts twice in FAH leaving an internal fragmentMouse 1 showed greater than 1 copy of FAH per diploidgenome equivalent compared to its matched control(lanes 10 and 11 Fig 4) while mice 2 and 3 bothshowed less than 1 copy of FAH per diploid genomecompared to their matched control (lanes 12 13 and 14Fig 4)
We used a nested inverse PCR approach on genomicDNA from mouse 1 to identify the specific integrationjunctions shown in the HindIII Southern blot (lane 6Fig 4) We identified seven different integration sitesThe vectorndashgenome junctions and their locations withinthe genome are listed in Table 3 In all seven cases theswitch from the attB sequence of the vector to thegenomic sequence occurred at or near a TT coreconfirming the use of fC31-mediated integration The
els on integration frequency
B C D
FAH-att 5 Agrase (n = 8)
25 Ag FAH-att 25 Agintegrase (n = 6)
25 Ag FAH-att 125 Agintegrase (n = 5)
14 104 52 F 66 105 29 F 43 105
0 0 0
19375 119201 13409112558 16101 19784
055 026 015
1181 1385 1690
patocytes and nodules were counted as single integration events In addition the minimum
403
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

ARTICLEdoi101016jymthe200411001
abnormal hepatocytes The quantity of integrase plas-mid was also strongly correlated with the level of genetransfer The average absolute integration frequencyranged from 11374 with a 11 ratio (group A) toonly 134091 with a 120 ratio (group D) Thesefindings are summarized in Table 2 and Fig 1
Integration Site AnalysisOne major pseudo attP site (mpsL1) favored forintegration into the mouse genome has been describedpreviously [10] Here we determined whether integra-tion into other sites also occurred We stably trans-fected approximately 11000 hepatocytes byhydrodynamic injection of the integrase construct andFAH-att but under selective pressure the FAH+ hep-atocytes expanded to approximately 70 of the livermass (~3 107 hepatocytes) Group 1 mice (n = 3)treated with FAH-att and integrase underwent a periodof selection for ~90 days We then used these long-termsurvivors as hepatocyte donors for serial transplantationinto naRve Fah secondary recipients After therecipient mice survived 90+ days off NTBC we usedthem as hepatocyte donors for a second round of serialtransplantation Tertiary Fah mutant recipients alsounderwent a selection period of 90 days off NTBCLiver tissue harvested and analyzed after the selectionperiod showed high levels of repopulation in thetertiary recipients (N60 by immunohistochemistry)(Fig 2B) All FAH-positive hepatocytes were healthyand showed no signs of dysplasia Additionally weperformed liver function tests on eight tertiary recipientmice The average alanine aminotransferase level was232 unitsL (ranging from 90 to 755 unitsL) which isslightly elevated over normal (100 unitsL) The crea-tinine and conjugated bilirubin levels were 05 mgLand 0 mgdl respectively which are similar to normallevels (creatinine = 0 mgL and bilirubin = 0 mgdl) Onaverage untreated Fah mice have alanine amino-transferase levels of 1000 unitsL creatinine levels of15 mgL and conjugated bilirubin levels of 05 mgdl[25]
TABLE 2 Effect of integrase lev
Group A
Plasmid combinations 25 Ag FAH-att 25 Agintegrase (n = 30)
25 Aginteg
Absolute integration frequency 73 F 38 104 11 FMinimum 14800
Median 11374Maximum 1465
Corrected integration frequency 36
127
Fah mice were treated with varying ratios of integrase to FAH expression construct FAH+ he
(worst animal) and maximum (best animal) frequencies in each group are shown
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
We extracted genomic liver DNA from three repopu-lated (~50ndash70 by immunohistochemistry) serial trans-plant recipients All three recipient mice had differentprimary hepatocyte donors which had been injectedwith the integrase and FAH-att constructs We analyzedintegration sites for the three mice by Southern blot afterdigestion with HindIII or ScaI and probed them using aportion of the human FAH cDNA The previously knownintegration site (mpsL1) was held in common betweenall three mice however several distinct junction frag-ments were observed in addition In mouse 1 DNAdigested with HindIII we identified two bmajor b sites ofintegration (77 and 68 kb) and four or five additionalbminorQ sites based upon band intensity (lane 6 Fig 4)To confirm these integration sites we performed asecond Southern blot on the same mouse 1 DNAdigested with ScaI Again two major sites were identified(60 and 48 kb) (lane 7 Fig 4) We also digested DNAfrom mice 2 and 3 with ScaI (lanes 8 and 9 Fig 4)Mouse 2 had two major sites of integration (60 and68 kb) and mouse 3 had three sites of integration (6068 and 86 kb) The 60-kb band corresponds to thempsL1 and was shared between all mice To identify thenumber of FAH gene copies per cell we performed athird Southern blot using the restriction enzyme PstIwhich cuts twice in FAH leaving an internal fragmentMouse 1 showed greater than 1 copy of FAH per diploidgenome equivalent compared to its matched control(lanes 10 and 11 Fig 4) while mice 2 and 3 bothshowed less than 1 copy of FAH per diploid genomecompared to their matched control (lanes 12 13 and 14Fig 4)
We used a nested inverse PCR approach on genomicDNA from mouse 1 to identify the specific integrationjunctions shown in the HindIII Southern blot (lane 6Fig 4) We identified seven different integration sitesThe vectorndashgenome junctions and their locations withinthe genome are listed in Table 3 In all seven cases theswitch from the attB sequence of the vector to thegenomic sequence occurred at or near a TT coreconfirming the use of fC31-mediated integration The
els on integration frequency
B C D
FAH-att 5 Agrase (n = 8)
25 Ag FAH-att 25 Agintegrase (n = 6)
25 Ag FAH-att 125 Agintegrase (n = 5)
14 104 52 F 66 105 29 F 43 105
0 0 0
19375 119201 13409112558 16101 19784
055 026 015
1181 1385 1690
patocytes and nodules were counted as single integration events In addition the minimum
403
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
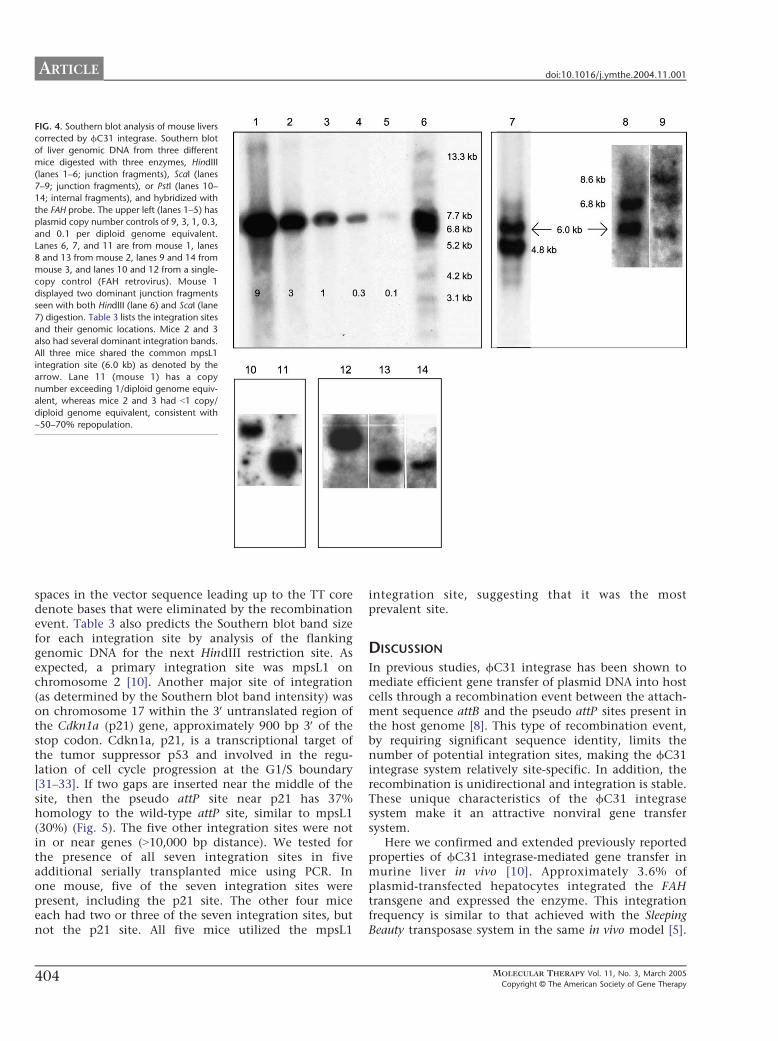
FIG 4 Southern blot analysis of mouse livers
corrected by fC31 integrase Southern blot
of liver genomic DNA from three different
mice digested with three enzymes HindIII
(lanes 1ndash6 junction fragments) ScaI (lanes
7ndash9 junction fragments) or PstI (lanes 10ndash
14 internal fragments) and hybridized with
the FAH probe The upper left (lanes 1ndash5) has
plasmid copy number controls of 9 3 1 03
and 01 per diploid genome equivalent
Lanes 6 7 and 11 are from mouse 1 lanes
8 and 13 from mouse 2 lanes 9 and 14 from
mouse 3 and lanes 10 and 12 from a single-
copy control (FAH retrovirus) Mouse 1
displayed two dominant junction fragments
seen with both HindIII (lane 6) and ScaI (lane
7) digestion Table 3 lists the integration sites
and their genomic locations Mice 2 and 3
also had several dominant integration bands
All three mice shared the common mpsL1
integration site (60 kb) as denoted by the
arrow Lane 11 (mouse 1) has a copy
number exceeding 1diploid genome equiv-
alent whereas mice 2 and 3 had b1 copy
diploid genome equivalent consistent with
~50ndash70 repopulation
ARTICLE doi101016jymthe200411001
spaces in the vector sequence leading up to the TT coredenote bases that were eliminated by the recombinationevent Table 3 also predicts the Southern blot band sizefor each integration site by analysis of the flankinggenomic DNA for the next HindIII restriction site Asexpected a primary integration site was mpsL1 onchromosome 2 [10] Another major site of integration(as determined by the Southern blot band intensity) wason chromosome 17 within the 3Vuntranslated region ofthe Cdkn1a (p21) gene approximately 900 bp 3Vof thestop codon Cdkn1a p21 is a transcriptional target ofthe tumor suppressor p53 and involved in the regu-lation of cell cycle progression at the G1S boundary[31ndash33] If two gaps are inserted near the middle of thesite then the pseudo attP site near p21 has 37homology to the wild-type attP site similar to mpsL1(30) (Fig 5) The five other integration sites were notin or near genes (N10000 bp distance) We tested forthe presence of all seven integration sites in fiveadditional serially transplanted mice using PCR Inone mouse five of the seven integration sites werepresent including the p21 site The other four miceeach had two or three of the seven integration sites butnot the p21 site All five mice utilized the mpsL1
404
integration site suggesting that it was the mostprevalent site
DISCUSSION
In previous studies fC31 integrase has been shown tomediate efficient gene transfer of plasmid DNA into hostcells through a recombination event between the attach-ment sequence attB and the pseudo attP sites present inthe host genome [8] This type of recombination eventby requiring significant sequence identity limits thenumber of potential integration sites making the fC31integrase system relatively site-specific In addition therecombination is unidirectional and integration is stableThese unique characteristics of the fC31 integrasesystem make it an attractive nonviral gene transfersystem
Here we confirmed and extended previously reportedproperties of fC31 integrase-mediated gene transfer inmurine liver in vivo [10] Approximately 36 ofplasmid-transfected hepatocytes integrated the FAHtransgene and expressed the enzyme This integrationfrequency is similar to that achieved with the SleepingBeauty transposase system in the same in vivo model [5]
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

TABLE 3 Junctions of FAH expression vector to genomic DNA
Chromosome
location Sequence vector-genome junction
Neighboring
gene(s)
Predicted
band
2 (mpsL1) tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcc
AGGGTACTACTCAGATGGTGACAGAGATGACAGGCTGCCCACATGAA
77 kb
7 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtAGGACATATTCCCAGTGACCCACCAGGGTCCACA
GACAACAGCATGGGTATGGCGCCCAG
42 kb
10 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccc
AGGSTCCTTATCTGCTAAGAATAGCTGTGTTTGGTCTGGCTCGGGAGACCCAT
69 kb
12 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgt
TACTCATGGTCTCAAGGTTATGACTCACAACCTGGACCAATTTTGGGAGGAAGTTGCT
52 kb
14 tcgacgatgtaggtcacggtctcgaagc
GCGGTCCTTTCAGGAGTGTGGTGACTCAAAGTCC
TCAGAGTCACCAAATC
Tkt (10 kb 5V)Prkcd (1 kb 3V)
133 kb
17 tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgccct
GCATATACATTCCCTTCCAGTCCACTGAGCTGTG
GGGCAAGTGCCTAGATATG
Cdkn1a
(in 3VUTR)
68 kb
X tcgacgatgtaggtcacggtctcgaagccgcggtgcgggtgccagggcgtgcccACACAATGGCATCTTAGAATTCTTTTCAACACATA
TAATTTTGCGGTGGACTCAAGAGAGTTACCC
31 kb
The vector sequence is in lowercase and the genomic DNA sequence is in uppercase For each intagration site the corresponding chromosome number and location in relation neighboring
genes are given Southern blot (Fig 4) band size (HindIII digest) for each integration is shown
ARTICLEdoi101016jymthe200411001
Furthermore the enhanced site specificity of integrationwas confirmed The mpsL1 site which is distant fromany genes was by far the most utilized site and wasfound in all treated mice However six additional sitesof integration were identified It is likely that all sevensites represent areas of high FAH expression andorintegration that allow for selective expansion It is alsoplausible that other sites of integration did occurinitially but due to minimal expression of FAH andorplacement within the genome the clone was not able toexpand Interestingly one pseudo attP site was in the 3Vuntranslated region of the p21 gene This site wasdetected in two mice but went undetected in otherintegrase-treated mice Importantly however there isvery little sequence conservation between the 3VUTRs ofthe human and the murine p21 genes Therefore theobservation made here in mouse does not predict thatthe human p21 gene will also target for fC31 integrase-mediated integration
The FAH copy number was determined to be approxi-mately 1 copy per diploid genome equivalent Howeverin one mouse (mouse 1 Fig 4) which had the p21 locus
FIG 5 Sequence alignment of pseudo attP sites and the wild-type attP Wild-type
the wild-type attP have 30 identity Chromosome 17 and wild-type attP have
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
as a dominant integration site the copy number wassignificantly higher than 1 It is therefore possible thatthe higher copy number was caused by the disruption ofthe p21 gene In other samples in which integration intop21 did not occur there was V1 copy of FAH per diploidgenome equivalent
We also found that fC31 integrase-transfected hepa-tocytes displayed morphological abnormalities at ahigher frequency than controls This phenomenonwas dependent on the amount of integrase expressionplasmid present High levels of hepatocyte dysplasiahave not been previously seen with other methods of invivo gene transfer into the liver of Fah miceincluding oncoretroviral vectors AAV2 and SB trans-posase [534ndash36] The abnormal cell morphology couldto some extent be ascribed to Fah deficiency itselfbecause dysplastic hepatocytes were seen at low fre-quencies in controls and are seen at high frequencyupon withdrawal of NTBC [2430] However it isprobable that integrase itself is also responsible forthe dysplastic appearance of hepatocytes The precisecause of these morphological changes is currently
attP and pseudo attP sites mpsL1 and chromosome 17 are shown MpsL1 and
37 similarity if a 2-bp gap is inserted into the pseudo site near the core
405
ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

ARTICLE doi101016jymthe200411001
unclear and must be further investigated A theoreticalpossibility is that fC31 integrase causes some level ofchromosomal instability fC31 integrase is a site-specific recombinase as is the unrelated enzyme CreIt has recently been suggested that Cre-recombinasemay mediate billegitimate recombinationQ in vivo iecreate genomic rearrangements even in the absence of aloxP site in the target genome [37] Since the humanand murine genomes contain multiple pseudo attP sites(at least seven in mouse hepatocytes) it is conceivablethat fC31 integrase expressed at high levels could alsomediate recombination between pseudo attP sites ondifferent chromosomes or even between more distantlyrelated DNA sequences It is possible that some aspectof integrase behavior such as putative nicking atpseudo att sites is tolerated in normal cells but ismore detrimental in the perturbed hepatocyte environ-ment of the Fah disease model The microenviron-ment of the Fah-mutant liver is abnormal even in thepresence of NTBC as suggested by the approximate 10-fold lower than normal transfection efficiency of thehydrodynamic liver delivery method observed here andin previous studies [5] the measurable levels ofspontaneous hepatocyte dysplasia (Fig 3) and thechanges in liver gene expression described for thisdisease setting [30]
Abnormal cellular morphology has not been reportedin previous studies of in vivo or in vitro fC31 integrasegene transfer suggesting that the phenomenon isspecific to the Fah model [810ndash12] For examplefC31 integrase-treated human keratinocytes were biop-sied from grafts at 4 8 12 and 14 weeks after graftingand analyzed with the result that the integrase-treatedcells were indistinguishable from normal cells [11] ThefC31 integrase has been used extensively in mouse EScells to perform genomic manipulations and to generatetransgenic mice with no adverse consequences and EScells expressing fC31 integrase under the PGK promoterhave been used to generate lines of mice that containthe fC31 integrase gene in every cell [3839] These micedisplay normal development and fertility suggestingthat modest levels fC31 integrase may have littletoxicity Transgenic Drosophila have also been generatedby using fC31 integrase with no ill effects [40] In thepresent study FAH-expressing hepatocytes were ofnormal appearance when histology was performed 90days after injection This finding indicates that normalhepatocytes are growth selected or that abnormalhepatocytes revert to a normal morphology with timeIn either case the transient presence of cells withabnormal morphology does not appear to have patho-logical consequences
Overall our findings suggest that fC31 integrase iseffective for integrating the FAH gene into hepatocytesafter codelivery of a plasmid encoding FAH and bearingan attB site and a plasmid expressing the fC31
406
integrase Integration was site-specific with seven inte-gration sites documented one being predominantHowever the insertion site into the mouse p21 geneserves as a reminder that the human genome maycontain bunwantedQ pseudo sites that may be used at alesser frequency Successful serial transfer of correctedhepatocytes to secondary and tertiary recipients indi-cated that the integration events were stable and thatgene expression was persistent over time and throughmultiple rounds of cell division The elevated frequencyof transient hepatocyte dysplasia seen here suggestedthat there is some level of toxicity associated withintegrase expression in the Fah model These typesof transient morphological abnormalities have beenobserved to date only in the Fah setting Normalhealthy livers without dysplastic cells were observed by90 days With careful monitoring for toxicity and forsafety of integration sites the fC31 integrase maybe acandidate gene transfer system for clinical use
MATERIALS AND METHODS
Vector construction Plasmid constructs of pCMVInt (CMV promoter
driving integrase expression) and pCS (empty cassette) were previously
described [910] The full-length human FAH cDNA expression cassette
contained in phFAFFA2 (14 kb) [16] was cloned into a blunted EcoRI site
of the expression plasmid pCD-SRa [17] The resulting plasmid
pCDSRahFAH expresses the FAH enzyme under the transcriptional
control of the SRa promoter and contains the SV40 polyadenylation
sequence (entire expression cassette is 346 kb) This cassette was removed
by digestion with SalI and cloned into the XhoI sites of the construct pBC-
hAAT-B replacing the hAAT cassette [10] resulting in a 677-kb plasmid
pBCB-FAH which contains an attB site
Animal husbandry integrase injection hepatocyte selection and
integration frequency FahDexon5 [13] strain mice were handled according
to NIH guidelines for animal care with the approval of the institutional
animal care and utilization committee of the Oregon Health amp Science
University All FahDexon5 mice were treated with NTBC-containing water
at a concentration of 75 mgL (provided by S Lindstedt Gothenborg
Sweden) [18] Fah mice (ranging in weight from 18 to 22 g) underwent
hydrodynamic tail vein injection over 5ndash8 s administering plasmid
constructs diluted in 09 saline solution (approximately 2 ml) as
previously described [1920] After an injection recovery period of 3 days
the mice were removed from the protective drug NTBC to allow for
expansion of FAH+ hepatocytes NTBC therapy was reinstated for 1 week
whenever animals lost more than 25 of their pretreatment weight
NTBC was removed again after weight gain equaled or exceeded the
starting weight
To measure the integration frequency Fah mice hydrodynamically
injected with different constructs were removed from NTBC 3 days after
injection for a selection period of 25 days [1920] The mice were then
sacrificed and sections of at least 07 cm2 were analyzed by FAH
immunohistology A blinded observer scored the presence of FAH+
hepatocytes using a Leica microscope DM RX (Leica Microsystems
Bannockburn IL USA) The surface area of the liver section was determined
by scanning the glass slides along with a size standard using a Microtec 2
scanner at a resolution of 254 dpi Adobe PhotoShop 502 software was used
to select and count the pixels corresponding to the liver sections A typical
mouse liver contains 186000 hepatocytescm2 [21]
The absolute frequency of integration was determined by dividing
the number of FAH+ clusters and individual FAH+ cells by the total
number of hepatocytes scored for a given section In the Fah mouse
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

ARTICLEdoi101016jymthe200411001
hydrodynamic tail vein injection transfects only 2 of all hepatocytes
(data not shown and Ref [5]) Therefore the absolute frequency must be
divided by 2 to calculate the actual transfection efficiency The actual
transfection efficiency reflects the number of hepatocytes that integrated
the integrase constructs assuming all hepatocytes were exposed to the
construct
Histology and immunohistology Liver tissues fixed in 10 phosphate-
buffered formalin pH 74 were dehydrated in 100 ethanol and
embedded in paraffin wax at 588C Four-micrometer sections were
rehydrated and stained with hematoxylin and eosin and with a
polyclonal rabbit antibody against rat Fah (kindly provided by Robert
Tanguay University of Laval Laval QC Canada) The antibody was
diluted in PBS pH 74 and applied at concentrations of 1300000 at 378Cfor 30 min Endogenous peroxidase activity was blocked with 3 H2O2
and methanol Avidin and biotin pretreatment was used to prevent
endogenous staining The secondary antibody was biotinylated goat anti-
rabbit IgG used at 1250 dilution (BA-1000 Vector Laboratories Burlin-
game CA USA) Color development was performed with the AEC
detection kit (Cat No 250-020 Ventana Medical Systems Tucson AZ
USA)
Dysplasia index To evaluate quantitatively the dysplastic-appearing
hepatocytes a dysplasia index scoring system was devised FAH+ nodules
in which more than half of the individual hepatocytes displayed an
abnormal phenotype were scored as 3+ FAH+ nodules in which less than
half of the individual hepatocytes displayed an abnormal phenotype were
scored as 2+ and nodules in which all hepatocytes looked normal were
scored as 1+ The observer was blinded to the transferred plasmid DNA
administered for each mouse
Hepatocyte transplantation Integrase-injected Fah mice that sur-
vived for more than 3 months off of NTBC were sacrificed and
hepatocytes were isolated for serial transplantation according to a
previously described protocol [22] For each round of serial transplanta-
tion 500000 hepatocytes were intrasplenically injected into Fah
recipient mice One week postinjection the recipient mice were removed
from NTBC to allow for selection and expansion of FAH+ hepatocytes
Liver function tests Animals were sacrificed by decapitation and blood
was collected in Microtainer plasma separator tubes with lithium heparin
(BectonndashDickinson Vacutainer Systems Franklin Lakes NJ USA) After a
brief centrifugation the plasma was frozen at 808C Twenty microliters
of plasma was mixed with 80 Al of a 7 (wv) bovine serum albumin
solution and assayed for aspartate serine aminotransferase bilirubin and
creatinine levels using a Kodak Ektachem 700 chemistry analyzer (East-
man Kodak Rochester NY USA)
Southern blot analysis Southern blot analysis was performed on DNA
from freshly obtained or frozen (808C) liver tissue extracted from treated
Fah mice Genomic liver DNA was isolated from ~350 mg of tissue
using the Qiagen genomic DNA isolation kit (Qiagen Valencia CA USA)
Ten micrograms of genomic DNA was digested with the restriction
enzyme HindIII or ScaI or PstI (Roche Indianapolis IN) Capillary transfer
and hybridizations were performed according to standard protocols [23]
A 478-bp fragment from nucleotide 373 to 850 of the human FAH cDNA
was isolated by PCR and radioactively labeled with dCTP to probe the
digested DNA Hybridization was detected by the radioactive signal [24]
The integration sites and copy number were evaluated
Pseudo site rescue by nested inverse pcr Mice that received FAH-att and
integrase were sacrificed Ten micrograms of liver genomic DNA was
digested with a group of restriction enzymes (NheI SpeI and XbaI) that
have incompatible cohesive ends and cut at least once in FAH-att The
digests were extracted with phenolchloroform precipitated with
ethanol and resuspended in 500 Al of 1 ligation buffer to which
was added 1000 units of T4 DNA ligase (New England Biolabs Beverly
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
MA USA) Low-concentration ligations were incubated at 178C over-
night extracted with phenolchloroform precipitated with ethanol and
resuspended in 20 Al TE Primary amplification with primers attB-F2 and
attL-PCR-1 was carried out as previously described [10] The PCR
products were purified using the QiaQuick PCR purification kit (Qiagen)
and a portion of the eluate was reamplified with nested primers attB-F3
and attL-iPCR-2 using the same protocol cited above All amplification
products were excised from the gel purified and cloned into pCR21-
TOPO using the TOPO Cloning Kit (Invitrogen Carlsbad CA USA)
Insert-containing colonies were sequenced using standard primers
previously described [25] Using specific primers designed for each
integration site in combination with vector-specific primers attB-F3
and hAATB-R the recombination junctions were then amplified from
the genomic liver DNA
To validate putative integration sites further additional primers
corresponding to sequences on either side of the vectorndashgenome junction
were designed and seven integration sites were confirmed by this
method The BlastN mouse genome database (wwwncbinlmmnihgov
genomeseqMnBlasthtml) was then used to identify the precise genomic
location and scan for the presence of nearby genes
ACKNOWLEDGMENT
This work was supported by National Institutes of Health Grants DK048252
(MG) and HL68112 (MPC)
RECEIVED FOR PUBLICATION JULY 22 2004 ACCEPTED NOVEMBER 1 2004
REFERENCES1 Hernandez Y J et al (1999) Latent adeno-associated virus infection elicits humoral
but not cell-mediated immune responses in a nonhuman primate model J Virol 73
8549 ndash 8558
2 Yang Y Li Q Ertl H C and Wilson J M (1995) Cellular and humoral immune
responses to viral antigens create barriers to lung-directed gene therapy with
recombinant adenoviruses J Virol 69 2004 ndash 2015
3 Nakai H Iwaki Y Kay M A and Couto L B (1999) Isolation of recombinant
adeno-associated virus vectorndashcellular DNA junctions from mouse liver J Virol 73
5438 ndash 5447
4 Hacein-Bey-Abina S et al (2003) LMO2-associated clonal T cell proliferation in two
patients after gene therapy for SCID-X1 Science 302 415 ndash 419
5 Montini E et al (2002) In vivo correction of murine tyrosinemia type I by DNA-
mediated transposition Mol Ther 6 759 ndash 769
6 Yant S R et al (2000) Somatic integration and long-term transgene expression
in normal and haemophilic mice using a DNA transposon system Nat Genet 25
35 ndash 41
7 Ivics Z Hackett P B Plasterk R H and Izsvak Z (1997) Molecular reconstruction
of Sleeping Beauty a Tc1-like transposon from fish and its transposition in human cells
Cell 91 501 ndash 510
8 Thyagarajan B Olivares E C Hollis R P Ginsburg D S and Calos M P (2001)
Site-specific genomic integration in mammalian cells mediated by phage phiC31
integrase Mol Cell Biol 21 3926 ndash 3934
9 Groth A C Olivares E C Thyagarajan B and Calos M P (2000) A phage
integrase directs efficient site-specific integration in human cells Proc Natl Acad Sci
USA 97 5995 ndash 6000
10 Olivares E C et al (2002) Site-specific genomic integration produces therapeutic
Factor IX levels in mice Nat Biotechnol 20 1124 ndash 1128
11 Ortiz-Urda S et al (2002) Stable nonviral genetic correction of inherited human skin
disease Nat Med 8 1166 ndash 1170
12 Ortiz-Urda S et al (2003) PhiC31 integrase-mediated nonviral genetic correction of
junctional epidermolysis bullosa Hum Gene Ther 14 923 ndash 928
13 Grompe M et al (1993) Loss of fumarylacetoacetate hydrolase is responsible for
the neonatal hepatic dysfunction phenotype of lethal albino mice Genes Dev 7
2298 ndash 2307
14 Overturf K et al (1998) Ex vivo hepatic gene therapy of a mouse model of hereditary
tyrosinemia type I Hum Gene Ther 9 295 ndash 304
15 Manning K Al-Dhalimy M Finegold M and Grompe M (1999) In vivo suppressor
mutations correct a murine model of hereditary tyrosinemia type I Proc Natl Acad Sci
USA 96 11928 ndash 11933
16 Phaneuf D et al (1991) Cloning and expression of the cDNA encoding human
fumarylacetoacetate hydrolase the enzyme deficient in hereditary tyrosinemia
assignment of the gene to chromosome 15 Am J Hum Genet 48 525 ndash 535
17 Takebe Y et al (1988) SR alpha promoter an efficient and versatile mammalian
cDNA expression system composed of the simian virus 40 early promoter and the R-U5
407
ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy

ARTICLE doi101016jymthe200411001
segment of human T-cell leukemia virus type 1 long terminal repeat Mol Cell Biol 8
466 ndash 472
18 Grompe M et al (1995) Pharmacological correction of neonatal lethal hepatic
dysfunction in a murine model of hereditary tyrosinaemia type I Nat Genet 10
453 ndash 460
19 Song Y K Liu F Zhang G and Liu D (2002) Hydrodynamics-based transfection
simple and efficient method for introducing and expressing transgenes in animals by
intravenous injection of DNA Methods Enzymol 346 92 ndash 105
20 Liu F Song Y and Liu D (1999) Hydrodynamics-based transfection in animals by
systemic administration of plasmid DNA Gene Ther 6 1258 ndash 1266
21 Wang X et al (2002) Kinetics of liver repopulation after bone marrow trans-
plantation Am J Pathol 161 565 ndash 574
22 Overturf K al-Dhalimy M Ou C N Finegold M and Grompe M (1997) Serial
transplantation reveals the stem-cell-like regenerative potential of adult mouse
hepatocytes Am J Pathol 151 1273 ndash 1280
23 Sambrook J and Russell D W (2001) Molecular Cloning A Laboratory Manual 3rd
ed Cold Spring Harbor Laboratory Press Cold Spring Harbor NY 2001
24 Al-Dhalimy M Overturf K Finegold M and Grompe M (2002) Long-term
therapy with NTBC and tyrosine-restricted diet in a murine model of hereditary
tyrosinemia type I Mol Genet Metab 75 38 ndash 45
25 Akerley B J et al (1998) Systematic identification of essential genes by in vitro
mariner mutagenesis Proc Natl Acad Sci USA 95 8927 ndash 8932
26 Gusella G L et al (2002) Lentiviral gene transduction of kidney Hum Gene Ther 13
407 ndash 414
27 Mujtaba T Han S S Fischer I Sandgren E P and Rao M S (2002) Stable
expression of the alkaline phosphatase marker gene by neural cells in culture and after
transplantation into the CNS using cells derived from a transgenic rat Exp Neurol 174
48 ndash 57
28 Chen Z Y et al (2001) Linear DNAs concatemerize in vivo and result in sustained
transgene expression in mouse liver Mol Ther 3 403 ndash 410
408
29 Chen S J Tazelaar J Moscioni A D and Wilson J M (2000) In vivo
selection of hepatocytes transduced with adeno-associated viral vectors Mol Ther
1 414 ndash 422
30 Luijerink M C et al (2003) Extensive changes in liver gene expression induced by
hereditary tyrosinemia type I are not normalized by treatment with 2-(2-nitro-4-
trifluoromethylbenzoyl)-13-cyclohexanedione (NTBC) J Hepatol 39 901 ndash 909
31 el-Deiry W S et al (1993) WAF1 a potential mediator of p53 tumor suppression Cell
75 817 ndash 825
32 Harper J W Adami G R Wei N Keyomarsi K and Elledge S J (1993) The p21
Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases Cell
75 805 ndash 816
33 Xiong Y et al (1993) p21 is a universal inhibitor of cyclin kinases Nature 366
701 ndash 704
34 Nakai H et al (2003) Helper-independent and AAV-ITR-independent chromosomal
integration of double-stranded linear DNA vectors in mice Mol Ther 7 101 ndash 111
35 Overturf K et al (1996) Hepatocytes corrected by gene therapy are selected in vivo
in a murine model of hereditary tyrosinaemia type I Nat Genet 12 266 ndash 273
36 Overturf K et al (1997) Adenovirus-mediated gene therapy in a mouse model of
hereditary tyrosinemia type I Hum Gene Ther 8 513 ndash 521
37 Schmidt E E Taylor D S Prigge J R Barnett S and Capecchi M R (2000)
Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse sper-
matids Proc Natl Acad Sci USA 97 13702 ndash 13707
38 Belteki G Gertsenstein M Ow D W and Nagy A (2003) Site-specific cassette
exchange and germline transmission with mouse ES cells expressing phiC31 integrase
Nat Biotechnol 21 321 ndash 324
39 Hollis R P et al (2003) Phage integrases for the construction and manipulation of
transgenic mammals Reprod Biol Endocrinol 1 79
40 Groth A C Fish M Nusse R and Calos M P (2004) Construction of transgenic
Drosophila by using the site-specific integrase from phage phiC31 Genetics 166
1775 ndash 1782
MOLECULAR THERAPY Vol 11 No 3 March 2005
Copyright C The American Society of Gene Therapy
Related Documents











