Cornmeal and starch influence the dynamic of fumonisin B, A and C production and masking in Fusarium verticillioides and F. proliferatum Irene Lazzaro a , Claudia Falavigna b , Gianni Galaverna b , Chiara Dall'Asta b , Paola Battilani a, ⁎ a Institute of Entomology and Plant Pathology, Università Cattolica del Sacro Cuore, Via Emilia Parmense 84, 29122 Piacenza, Italy b Department of Food Science, University of Parma, Parco Area delle Scienze 17/A, 43124 Parma, Italy abstract article info Article history: Received 11 March 2013 Received in revised form 5 June 2013 Accepted 11 June 2013 Available online 18 June 2013 Keywords: Mycotoxins Hidden fumonisins Fusarium Maize components Fusarium verticillioides and F. proliferatum can infect maize ears and produce fumonisins. The fumonisin B (FB) series is the most prolifically produced, followed by fumonisin C (FC), A (FA) and P (FP); moreover hidden forms of fumonisins have been detected in maize and derivatives. There is a lack of infor- mation about which maize component may affect fumonisin pattern production. Therefore, in this work we studied the role of cornmeal and corn starch, as the sole source of nutrition, in the production dynamic of all fumonisin series, hidden forms included, in different strains of F. verticillioides and F. proliferatum incu- bated at 25 °C for 7–45 days. Both Fusarium species produced high amounts of FB, following the chemotype FB 1 > FB 2 > FB 3 ; FC and FA were produced in lesser amounts, showing the chemotypes: FA 2 + FA 3 > FA 1 and FC 1 > FC 2 + FC 3 > FC 4 , respectively; while no FP were detected. F. verticillioides was more prolific than F. proliferatum in fumonisin production (ten times more on average) in all the tested conditions. Fumonisin pro- duction was higher in cornmeal than in starch based medium in both Fusarium species; FA and FC were detected only in the former medium. An important role of amylopectin as an inducing factor for fumonisin biosynthesis was suggested, as were acid pH conditions. Fumonisin hidden forms may occur in cornmeal medium, whereas they were never found at significant levels in corn starch medium. © 2013 Elsevier B.V. All rights reserved. 1. Introduction Maize is the second most cultivated crop worldwide (170 million ha) and has the highest average yield per hectare of any crop (~5.2 t/ha, in 2011). It is the most important agricultural commodity in the Americas and third most important, after wheat and barley, in Europe (faostat.fao.org). Several pests and diseases can reduce crop yield but one of the major concerns relates to Fusarium verticillioides and Fusarium proliferatum, the two fungi mainly responsible for pink ear rot on maize that can develop in almost all maize growing areas (Battilani and Logrieco, 2013). These fungi are known to produce fumonisins, myco- toxins that can accumulate in maize kernels during ripening. Fumonisins are considered a risk for human and animal health as fumonisin B 1 (FB 1 ) is known to cause equine leukoencephalomalacia, porcine pulmonary oe- dema, human oesophageal cancer and liver damage in swine (Howard et al., 2001; Marasas et al., 2004; Schroeder et al., 1996); it has been also classified as a possible carcinogen to humans (Group 2B) by the Interna- tional Agency for Research on Cancer (IARC, 1993). Humans and animals are potentially exposed to fumonisin B (FB) worldwide and high levels of FBs are often reported in maize or maize-based food such as corn meal (Battilani et al., 2008; Bittencourt et al., 2005; Garrido et al., 2012; Silva et al., 2007). Fumonisins B, and in particular FB 1 followed by FB 2 , are the most abundant in nature (Rheeder et al., 2002). However these fungi are able to produce three minor series of fumonisin other than FB: fumonisin A (FA), fumonisin C (FC) and fumonisin P (FP). Fumonisin C and FA usu- ally occur at lower concentrations than FB (b 5%) (Bartόk et al., 2006); FP can occur at levels up to 30% of FB when F. verticillioides is grown on cracked maize kernel medium (Musser et al., 1996). Fumonisins C are known to be phyto- and cytotoxic (Abbas et al., 1998) and they can co-occur with FB in naturally contaminated maize (Seo and Lee, 1999). The in vitro production of fumonisin B is well studied (EHC, 2000; Marín et al., 1999; Scott, 2012). The dynamics of production of fumonisin series was studied by Lazzaro et al. (2012) in cultures of F. verticillioides grown on Malt Extract Agar for 45 days. They observed that FB, FA and FC, but no FP were produced. The production of all fumonisin series in F. verticillioides cultures was previously reported by Musser and Plattner (1997), but data on the production versus incubation time were not provided. In addition to major and minor forms the occurrence of hidden fumonisins in maize has been reported (Dall'Asta et al., 2010; Dall'Asta et al., 2012). These forms are not covalently bound derivatives, they are entrapped in food macromolecules such as starch and proteins, thus they are determined with difficulty by common analytical methods. Hid- den fumonisins cannot be fully regarded as masked mycotoxins, since the physical entrapment does not lead to a stable conjugation. In partic- ular, masked mycotoxins are usually stable under gastrointestinal International Journal of Food Microbiology 166 (2013) 21–27 ⁎ Corresponding author. Tel.: +39 0523 599254. E-mail address: [email protected] (P. Battilani). 0168-1605/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.ijfoodmicro.2013.06.011 Contents lists available at SciVerse ScienceDirect International Journal of Food Microbiology journal homepage: www.elsevier.com/locate/ijfoodmicro

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal of Food Microbiology 166 (2013) 21–27
Contents lists available at SciVerse ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r .com/ locate / i j foodmicro
Cornmeal and starch influence the dynamic of fumonisin B, A and Cproduction and masking in Fusarium verticillioides and F. proliferatum
Irene Lazzaro a, Claudia Falavigna b, Gianni Galaverna b, Chiara Dall'Asta b, Paola Battilani a,⁎a Institute of Entomology and Plant Pathology, Università Cattolica del Sacro Cuore, Via Emilia Parmense 84, 29122 Piacenza, Italyb Department of Food Science, University of Parma, Parco Area delle Scienze 17/A, 43124 Parma, Italy
⁎ Corresponding author. Tel.: +39 0523 599254.E-mail address: [email protected] (P. Battilan
0168-1605/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.ijfoodmicro.2013.06.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 11 March 2013Received in revised form 5 June 2013Accepted 11 June 2013Available online 18 June 2013
Keywords:MycotoxinsHidden fumonisinsFusariumMaize components
Fusarium verticillioides and F. proliferatum can infect maize ears and produce fumonisins.The fumonisin B (FB) series is the most prolifically produced, followed by fumonisin C (FC), A (FA) and P (FP);moreover hidden forms of fumonisins have been detected in maize and derivatives. There is a lack of infor-mation about which maize component may affect fumonisin pattern production. Therefore, in this workwe studied the role of cornmeal and corn starch, as the sole source of nutrition, in the production dynamicof all fumonisin series, hidden forms included, in different strains of F. verticillioides and F. proliferatum incu-bated at 25 °C for 7–45 days. Both Fusarium species produced high amounts of FB, following the chemotypeFB1 > FB2 > FB3; FC and FA were produced in lesser amounts, showing the chemotypes: FA2 + FA3 > FA1
and FC1 > FC2 + FC3 > FC4, respectively; while no FP were detected. F. verticillioides was more prolific thanF. proliferatum in fumonisin production (ten timesmore on average) in all the tested conditions. Fumonisin pro-ductionwas higher in cornmeal than in starch basedmedium in both Fusarium species; FA and FCwere detectedonly in the former medium. An important role of amylopectin as an inducing factor for fumonisin biosynthesiswas suggested, as were acid pH conditions. Fumonisin hidden forms may occur in cornmeal medium, whereasthey were never found at significant levels in corn starch medium.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Maize is the secondmost cultivated cropworldwide (170 million ha)and has the highest average yield per hectare of any crop (~5.2 t/ha, in2011). It is the most important agricultural commodity in the Americasand third most important, after wheat and barley, in Europe(faostat.fao.org). Several pests and diseases can reduce crop yield butone of the major concerns relates to Fusarium verticillioides and Fusariumproliferatum, the two fungi mainly responsible for pink ear rot on maizethat can develop in almost all maize growing areas (Battilani andLogrieco, 2013). These fungi are known to produce fumonisins, myco-toxins that can accumulate in maize kernels during ripening. Fumonisinsare considered a risk for human and animal health as fumonisin B1 (FB1)is known to cause equine leukoencephalomalacia, porcine pulmonary oe-dema, human oesophageal cancer and liver damage in swine (Howard etal., 2001; Marasas et al., 2004; Schroeder et al., 1996); it has been alsoclassified as a possible carcinogen to humans (Group 2B) by the Interna-tional Agency for Research on Cancer (IARC, 1993). Humans and animalsare potentially exposed to fumonisin B (FB)worldwide and high levels ofFBs are often reported in maize or maize-based food such as corn meal(Battilani et al., 2008; Bittencourt et al., 2005; Garrido et al., 2012; Silvaet al., 2007).
i).
rights reserved.
Fumonisins B, and in particular FB1 followed by FB2, are the mostabundant in nature (Rheeder et al., 2002). However these fungi areable to produce threeminor series of fumonisin other than FB: fumonisinA (FA), fumonisin C (FC) and fumonisin P (FP). Fumonisin C and FA usu-ally occur at lower concentrations than FB (b5%) (Bartόk et al., 2006); FPcan occur at levels up to 30% of FB when F. verticillioides is grown oncracked maize kernel medium (Musser et al., 1996). Fumonisins C areknown to be phyto- and cytotoxic (Abbas et al., 1998) and they canco-occur with FB in naturally contaminated maize (Seo and Lee, 1999).
The in vitro production of fumonisin B is well studied (EHC, 2000;Marín et al., 1999; Scott, 2012). The dynamics of production of fumonisinseries was studied by Lazzaro et al. (2012) in cultures of F. verticillioidesgrown on Malt Extract Agar for 45 days. They observed that FB, FA andFC, but no FP were produced. The production of all fumonisin series inF. verticillioides cultures was previously reported by Musser and Plattner(1997), but data on the production versus incubation time were notprovided.
In addition to major and minor forms the occurrence of hiddenfumonisins in maize has been reported (Dall'Asta et al., 2010; Dall'Astaet al., 2012). These forms are not covalently bound derivatives, they areentrapped in food macromolecules such as starch and proteins, thusthey are determinedwith difficulty by common analyticalmethods. Hid-den fumonisins cannot be fully regarded as masked mycotoxins, sincethe physical entrapment does not lead to a stable conjugation. In partic-ular, masked mycotoxins are usually stable under gastrointestinal

22 I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
conditions (Dall'Erta et al., 2013),whereas parent fumonisins are totallyreleased from hidden forms upon digestion (Dall'Asta et al., 2010;Falavigna et al., 2012b).
Nutritional factors are crucial for fumonisin production. Dried rip-ening maize kernels have the following composition (percentage onthe maize seed weight): 65–80% starch, mainly concentrated in theendosperm, 1.4–2% soluble sugars, 8–10% protein and 3.5–4.5% oil,mainly located in the germ, 1.5–2.0% ash and 1.5–2.1% crude fibre, al-most all in the pericarp, and 10–15% water (Dall'Asta et al., 2012;Smith et al., 2004).
Cornmeal is the main product in the diet of many populations livingin South America and Africa, where it is considered a staple food and isoften used to complement the diet of children (www.fao.org). Starch isalso a significant product derived from maize and important in thehuman diet, especially in American countries. In the food industry,starch is used either as a thickening agent or as a precursor of sweet-eners such as corn syrups (www.fao.org). Starch, whose major constit-uent is amylopectin (up to75% compared with 25% amylose, the othercomponent), is the most abundant carbon source for fungi colonizinga ripening maize ear. Amylopectin is known to be an inducing factorfor FB1 production in F. verticillioides (Bluhm and Woloshuk, 2005).
As far as we are aware, there is a lack of information about whichmaize component could affect fumonisin pattern production and themasking phenomenon; only recently, a relation between fatty acids,linoleic acid in particular, and fumonisin production and maskingwas reported Dall'Asta et al. (2012). Contrary to masked mycotoxinssuch as deoxynivalenol-3-glucoside that can be formed exclusivelyby plant enzyme activity (Berthiller et al., 2012) hidden fumonisinsmaypotentially be formed also under in vitro conditions,whenunspecificsupramolecular interactionsmay occur between fumonisins andmediummacroconstituents.
For this reason we investigated the influence of corn meal and cornstarch, as sole nutritional sources, on the production dynamic of allfumonisin series and on the masking phenomenon in F. verticillioidesand F. proliferatum cultures.
This could provide more information regarding fungal metabolism,in terms of nutritional needs, and of the mechanism behind fumonisinproduction.Moreover it could benefit the understanding of themaskingphenomenon, which has been observed only inmaize andmaize-basedfood, and whose mechanism has not yet been clarified.
2. Material and methods
2.1. Fungal material
The fungal species used in this study were F. verticillioides and theclosely related species F. proliferatum. Three strains of F. verticillioideswere considered: ITEM1744, ITEM10027 (MPVP 294) and ITEM10026(MPVP 289), the first isolated from maize in South Sardinia, and thelatter two in South Tuscany, Italy. Two strains of Gibberella intermedia(teleomorph of F. proliferatum) were included: ITEM 7595 and ITEM15876 (MPVP328), respectively isolated from maize in Kansas, USA andthe Po valley, Italy. Cultures are stored in 18% glycerol at the fungal collec-tion of the Institute of Entomology and Plant Pathology-UCSC, Piacenza(MPVP) and at the Institute of Sciences of Food Production-CNR, Bari(ITEM; http://server.ispa.cnr.it/ITEM/Collection).
2.2. Inoculum preparation and maize-based substrates
All strainswere grown on Potato Dextrose Agar at 25 °C for 7 d in thedark and at the end of incubation. A portion of the colony (7 mmØ)wasused as inoculum.
All strains were inoculated on two different solid media, cornmeal(CMM), made with cornmeal and sterile distilled water (1:7, volume)and corn starch (CSM), prepared with corn starch and sterile distilledwater (1:1, volume). Commercial maize flour bought at the supermarket
was used for CMMpreparationwhereas a commercial corn starch (CarloErba, Milano, Italy) was used for CSM.
Both substrates were tyndallised by heating for 30 min at 80 °Cthree times, poured into 90 mmØ Petri plates and once cooled, coveredby a cellophane sheet (P400; Cannings, Ltd., Bristol, UK) to facilitate re-moval of the fungal biomass. The fungal inoculumwas added at the cen-tre of the plate; a similar sized portion of uninoculated PDA was addedto control plates. Cultures were incubated for 7, 14, 21, 30 or 45 days at25 °C in the dark and then stored at−20 °C. The trial was performed intriplicate.
Water activity of both media was measured in triplicate using theAqualab LITE (Decagon, Pullman, WA, USA), according to the manu-facturer instructions, before inoculation of plates.
2.3. Sample preparation for free and total fumonisins
Sampleswere prepared according to Lazzaro et al. (2012). Briefly, al-iquots (2 g) of the maize-based mediumwere blended in a high-speedblender (Ultraturrax T18; IKA, Stauffen, Germany) with 8 mL of water/methanol, 30:70 v/v, for 1 min and then centrifuged at 2470 g for15 min. Next, an aliquot (50 μL) of supernatant was diluted with450 μL of water/methanol, 30:70 v/v, before LC–ESI-MS/MS analysis.
Samples (2 g aliquot) for determination of hydrolysed fumonisinswere blended with 20 mL of 2 M KOH, centrifuged for 1 min at 2474 gand then stirred for 60 min. The sample was then combined with20 mL of acetonitrile, stirred for 10 min, and then centrifuged at 2470 gfor 15 min in order to resolve the sample into two layers. A 200 μL por-tion of the upper, acetonitrile-rich layer was evaporated to drynessunder a stream of compressed air, and the residue was re-dissolved in800 μL water/methanol, 30:70 v/v and analysed by LC–ESI-MS/MS.After hydrolysis, samples were examined by LC–ESI-MS/MS for the pres-ence of the hydrolysed forms of FB1, FB2 and FB3.
All the results are expressed as the sum of FB1, FB2, and FB3 equiv-alents, considering a correction factor due to the different molecularweights of parent and hydrolysed compounds and referred to as“total fumonisins after hydrolysis”.
2.4. LC–ESI-MS/MS analysis for the determination of free and totalfumonisins
Free and total fumonisins were determined according to Dall'Astaet al. (2010) without any modification.
Very briefly, LC–ESI-MS/MS analysis was performed by a 2695 Alli-ance separation system (Waters Co., Milford, MA, USA) equipped withaQuattro API triple quadrupolemass spectrometerwith an electrospraysource (Micromass; Waters, Manchester, UK). Chromatographic condi-tions were set as follows: the column was a 250 mm × 2.1 mm i.d.,5 μm, XTerra C18; the flow rate was 0.2 mL/min; the column tempera-ture was set at 30 °C; the injection volume was 10 μL; gradient elutionwas performedusingbidistilledwater (eluentA) andmethanol (eluent B)both acidified with 0.2% formic acid: initial condition at 70% A, 0–2 minisocratic step, 2–5 min linear gradient to 45% B, 5–25 min linear gradientto 90% B, 25–35 min isocratic step at 90% B, 35–36 min linear gradient to70% A, and reequilibration step at 70% A for 15 min (total analysis time:50 min). MS parameters: ESI + (positive ionisation mode); capillaryvoltage, 4.0 kV; cone voltage, 50 V; extractor voltage, 2 V; source blocktemperature, 120 °C; desolvation temperature, 350 °C; cone gas flowand desolvation gas flow (nitrogen), 50 L/h and 700 L/h, respectively.
Detection was performed using a multiple reaction monitoring(MRM) mode by monitoring two transitions for each analyte, asfollows: 722.4 → 334.4, 722.4 → 352.3 for FB1, 706.4 → 336.4 and706.4 → 318.4 for FB2 and FB3, 406.5 → 370.5 and 406.5 → 388.5for HFB1, 390.5 → 336.4 and 390.5 → 372.5 for HFB2 and HFB3,564.1 → 334.4 and 564.1 → 352.2 for PHFB1, 548.2 → 336.4and 548.2 → 354.20 for PHFB2 and PHFB3, 532.2 → 338.2 and532.2 → 320.2 for PHFB4.

Table 1ANOVA of the effects of the species (Fusarium verticillioides and F. proliferatum) and theincubation time (7–45 days) on the production of fumonisin B and hidden forms infungal cultures grown on cornmeal medium.
Factors FB1a FBtotb TOT Hidden
Species **c ** **F. verticillioides10026 250.1 a 379.7 a 366.2 a 2.010027 288.7 a 408.8 a 469.7 a 37.31744 160.2 a 249.2 a 230.0 a 0.7
F. proliferatum7595 67.8 b 115.1 b 115.1 c 0.7328 17.3 b 33.0 b 47.1 b 15.3
Time ** ** **7 6.8 c 11.7 c 11.3 c 0.814 79.9 b 129.4 b 123.7 b 3.821 229.7 a 374.5 a 407.3 a 13.230 200.7 a 318.7 a 311.3 a 9.745 277.7 a 368.9 a 394.1 a 29.2
Strain ∗ time ** ** **
a Fumonisins are measured in mg/kg.b FBtot = FB1 + FB2 + FB3 + FB4 + FB5 (fumonisin before alkaline hydrolysis);
TOT = fumonisin after alkaline hydrolysis; MASK = hidden fumonisins (TOT–FBtot).c ** = significance p ≤ 0.01; * = significance p ≤ 0.05; ns = not significant.
23I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
The first transition reported was used for quantification, while thesecond transition was chosen as qualifier. Calibration curves (calibrationrange 10–1000 μg/kg) were used for free fumonisin and hydrolysedfumonisin quantification. For hydrolysed fumonisin quantification, stan-dards were obtained by hydrolysing FB1, FB2 and FB3 as reported byDall'Asta et al. (2010).
Limits of detection (LOD) and quantification (LOQ)were respective-ly 5 μg/kg and 10 μg/kg for fumonisins and hydrolysed fumonisins.
2.5. LC–ESI-MS/MS analysis for the determination of FB, FA and FCanalogues
Major andminor fumonisins were determined according to Falavignaet al. (2012a). The instrument setup described above was used also forthis analysis.
The separation was obtained on an XTerra C18 (250 mm × 2.1 mmi.d., 5 μm), using a flow rate of 0.2 mL/min. The column temperaturewas set at 30 °C; the injection volume was 10 μL. Gradient elution wasachieved using bidistilled water (eluent A) and acetonitrile (eluent B)both acidified with 0.2% formic acid: initial condition at 100% A,0–5 min isocratic step, 5–30 min linear gradient to 100% B, 30–35 minisocratic step, 35–36 min linear gradient to 100% A and reequilibrationstep at 100% A for 14 min (total analysis time: 50 min). MS parameterswere the following: ESI + (positive ionisation mode); capillary voltage,4.0 kV; cone voltage, 30 V; extractor voltage, 2 V; source block temper-ature, 120 °C; desolvation temperature, 350 °C; and cone gas flow anddesolvation gas flow (nitrogen), 50 L/h and 700 L/h, respectively.
Minor fumonisins were detected using a MRM mode, by moni-toring two transitions for each analytes, as follows: 690.1 → 320.1and 690.1 → 338.2 for FB4; 738.4 → 368.1 and 738.4 → 350.1 forFB5; 764.1 → 334.1 and 764.1 → 394.1 for FA1; 748.1 → 336.1 and748.1 → 378.1 for FA2 and FA3; 707.8 → 337.8 and 707.8 → 319.1for FC1; and 692.2 → 322.1 and 692.2 → 340.2 FC2 and FC3. Thefirst transition reported was used for quantification, while the secondtransition was chosen as qualifier.
As already reported by Lazzaro et al. (2012), A- and C-seriesfumonisins as well as FB4 and FB5 were quantified using FB1-calibrationcurve (calibration range 250–5000 μg/L), since analytical standards arenot commercially available for minor fumonisins.
2.6. pH measurements
pH measurements were performed in the medium using a pH 212Microprocessor pH meter (Hanna Instruments, Modena, Italy). Beforemeasurements, the pH meter was calibrated using an appropriatebuffer solution. Each sample underwent to pH measurements beforeanalysis.
2.7. Statistical analysis
Statistical analyses were performed using SPSS v.19.0 (SPSS Italia,Bologna, Italy). Data were statistically analysed by using a OneWay-ANOVA Test transforming all fumonisin concentration valuesby y = ln(x + 1) before analysis; mean data were statistically com-pared by a post-hoc Tukey test (α = 0.05).
3. Results
3.1. Production of fumonisins on cornmeal medium
Fumonisins B, FA and FC were produced by all F. verticillioides andF. proliferatum strains, in particular F. verticillioides strains were signif-icantly (p ≤ 0.01) the strongest producers of all fumonisin series; nosignificant differences were observed between strains belonging tothe same species (Table 1; Fig. 1).
Fumonisins A were produced after 21 days of incubation byITEM10026 and ITEM10027; ITEM1744 started FA production after7 days of incubation and was the highest producer (6400 μg/kg inaverage of total FA produced). For F. proliferatum, ITEM7595 pro-duced FA only at 45 days of incubation and ITEM15876 did not syn-thesize FA at all (Fig. 1).
All the three F. verticillioides strains produced FC from 7 days, withITEM 10027 the highest producer (57600 μg/kg in average of total FCproduced), whereas both F. proliferatum strains showed FC produc-tion only at 21 and 45 days (Fig. 1). Fumonisin P were analysed, butnever detected in any of the conditions considered in this study. Nodata analysis was performed for FA and FC because the productionwas observed only in a limited part of scheduled observations. Theproduction of FB, FA and FC by each F. verticillioides and F. proliferatumstrain is detailed in Table 3.
Fumonisin production followed a similar trend for all considered se-ries during the incubation time: FB significantly (p ≤ 0.01) increasedup to 21 days of incubation, FA followed the same trend while thepeak of production for FCwas registered at 30 days. The two-way inter-action strain ∗ timewas always highly significant (p ≤ 0.01) for FB pro-duction, thus all data were represented based on this interaction(Fig. 1).
For FB1–5, production at 7 dayswas significantly lower than the otherincubation time points in all strains tested. ITEM10026, ITEM1744 andITEM15876 produced FB1–5 maximally at 21 days, whereas ITEM10027produced maximally at 30 days, then decreased up to 45 days; on thecontrary, in ITEM10026, production at 45 days was significantly thehighest (p ≤ 0.01). For FA, the peak of production was observed at21 days (ITEM1744, which is the minor FB1–5 producer) or 30 days(ITEM10026 and ITEM10027); again ITEM7595 produced higheramounts of FA at 45 days. Fumonisin C production in F. verticillioideswas lower at 7 days and also at 45 dayswhere it decreased significantly(p ≤ 0.01). Peaks were observed at 21 days (ITEM1744, ITEM10026and ITEM15876), 30 days (ITEM10027) and again at 45 days inITEM7595 (Fig. 1).
Negative controls, corresponding to CMM plates without fungalinoculum, did not contain any fumonisin. The water activity of themedium without inoculum was 0.99.
3.2. Production of fumonisins on corn starch medium
Fumonisin B was the only series detected on CSM. F. verticillioidesstrains produced significantly less FB1–5 than on CMM (18.3 vs

Fig. 1. Fumonisin B, FA and FC production profiles by F. verticillioides (ITEM10026–ITEM10027–ITEM1744) and F. proliferatum strains (ITEM7595–ITEM15876) cultured on cornmealmedium in a 45 day-long incubation period.
Table 2ANOVA of the effects of the fungal species (Fusarium verticillioides and F. proliferatum)and the incubation time (7–45 days) on the production of fumonisin B and hiddenforms in fungal cultures grown on corn starch medium.
Factors FB1a FBtotb TOT Hidden
Species **c ** **F. verticillioides10026 32.1 b 44.2 b 44.4 b 0.210027 5.5 b 8.4 b 12.0 b 3.01744 1.1 b 2.2 b 2.0 b 0.0
F. proliferatum7595 0.6 c 1.5 c 1.5 c 0.015876 116.6 a 147.9 a 147.9 a 1.0
Time ** ** **7 0.9 c 1.9 c 2.9 c 1.114 4.8 bc 7.7 bc 8.7 bc 0.721 30.7 ab 37.6 ab 41.8 ab 0.530 37.9 ab 47.7 ab 54.2 ab 0.245 81.6 a 109.2 a 123.3 a 1.5
Strain ∗ time ** ** **
a Fumonisins are measured in mg/kg.b FBtot = FB1 + FB2 + FB3 + FB4 + FB5; TOT = fumonisin after alkaline hydrolysis;
MASK = hidden fumonisins (TOT–FBtot).c Significance p ≤ 0.01.
24 I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
346 mg/kg in average of total FB1–5 produced), as did F. proliferatumITEM7595 (1.5 vs 115.1 mg/kg) (Table 2). F. proliferatum ITEM15876produced the highest amount of FB1–5, including comparison withF. verticillioides strains, with a production 5 times higher, in average,than on CMM (Table 2); therefore, significant differences between the2 strains of F. proliferatum were observed.
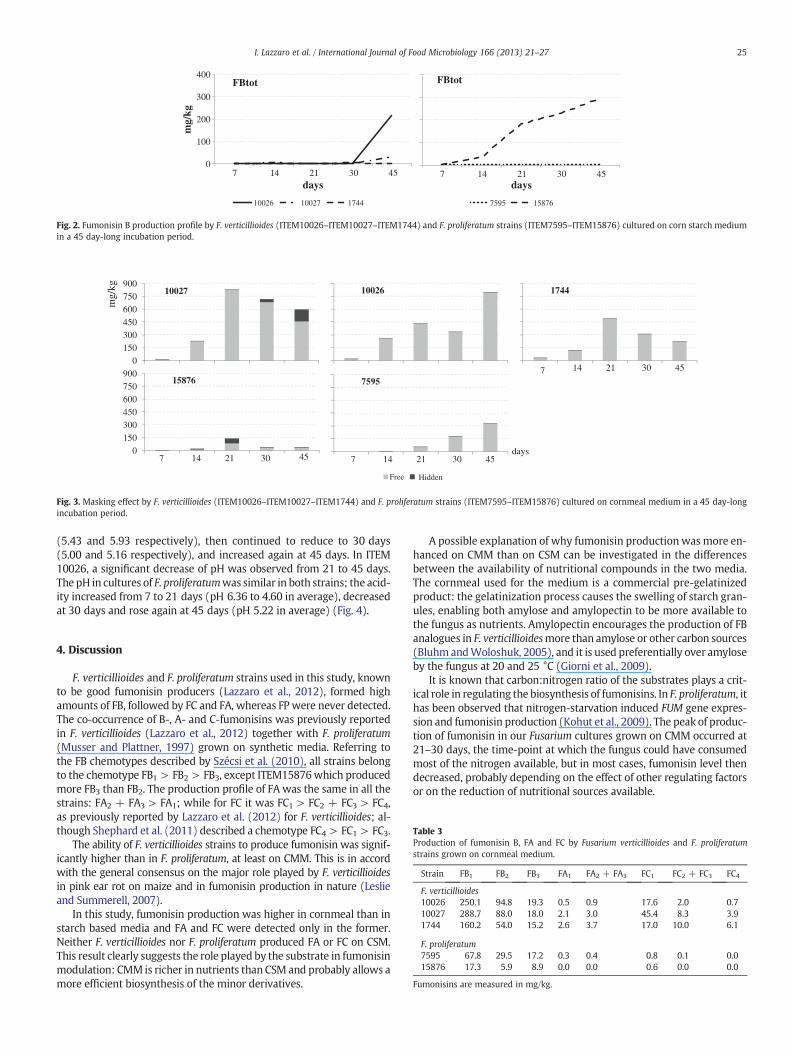
Fumonisin B production over time significantly increased up to45 days of incubation. The two-way interaction strain ∗ time was al-ways highly significant (p ≤ 0.01) for fumonisin production, so alldata were presented based on this interaction (Fig. 2).
Fumonisin B production followed a similar trend in all F. verticillioidesand F. proliferatum strains, with a peak at 30 days for ITEM1744, and at45 days for all the other strains (Fig. 2).
Negative controls, corresponding to CSMplateswithout fungal inoc-ulum did not contain any fumonisin. The water activity of the mediumwithout inoculum was 0.99.
3.3. Hidden fumonisin on cornmeal medium
Hidden fumonisins were measured on CMM as the difference be-tween total FB1–3 found after hydrolysis and free FB1–3, quantified withthe standard analytical approach. F. verticillioides strains seemed to pro-duce more hidden forms than F. proliferatum, the highest amount beingfound for ITEM10027 (37.3 mg/kg; Fig. 3). For all the strains, the hiddenFB1–3 amount was very limited and the highest mean amount was pro-duced after 45 days of incubation (Table 2).
As already reported for free fumonisins, the two-way interactionstrain ∗ time was always highly significant (p ≤ 0.01) also for totalFB1–3 production.
3.4. Hidden fumonisin on corn starch medium
Hidden fumonisins were measured on CSM as the difference be-tween total FB1–3 found after hydrolysis and free FB1–3. Although inthis case the hidden fumonisin amount was generally lower thanon CMM, F. verticillioides strains produced more hidden forms thanF. proliferatum, with the highest amount in this case also found in
ITEM10027 (3.0 mg/kg). Similarly to data obtained on CMM, the hiddenFB1–3 amount was highest after 45 days of incubation (Table 3).
As already reported for free fumonisins, the two-way interactionstrain ∗ time was always highly significant (p ≤ 0.01) also for totalFB production.
3.5. pH measurement in CMM
pH was measured only in CMM, because starch gelatinization inwater did not allow a reliable measurement in CSM.
The pH of CMM before inoculation with fungal strain was 6.12.Changes in pH were observed during the incubation of fungal strains,and these were different between the two Fusarium species. ForF. verticillioides, in ITEM10027 and ITEM1744 pH changed at 14 days
7 14 21 30 45
FBtot
7595
0
100
200
300
400
7 14 21 30 45m
g/kg
days days
FBtot
10026 10027 1744 15876
Fig. 2. Fumonisin B production profile by F. verticillioides (ITEM10026–ITEM10027–ITEM1744) and F. proliferatum strains (ITEM7595–ITEM15876) cultured on corn starch mediumin a 45 day-long incubation period.
mg/
kg 174410026
7595
Free
days
0150300450600750900
10027
0150300450600750900
15876
Hidden
7 14 21 30 45
7 14 21 30 457 14 21 30 45
Fig. 3. Masking effect by F. verticillioides (ITEM10026–ITEM10027–ITEM1744) and F. proliferatum strains (ITEM7595–ITEM15876) cultured on cornmeal medium in a 45 day-longincubation period.
25I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
(5.43 and 5.93 respectively), then continued to reduce to 30 days(5.00 and 5.16 respectively), and increased again at 45 days. In ITEM10026, a significant decrease of pH was observed from 21 to 45 days.The pH in cultures of F. proliferatumwas similar in both strains; the acid-ity increased from 7 to 21 days (pH 6.36 to 4.60 in average), decreasedat 30 days and rose again at 45 days (pH 5.22 in average) (Fig. 4).
Table 3Production of fumonisin B, FA and FC by Fusarium verticillioides and F. proliferatumstrains grown on cornmeal medium.
Strain FB1 FB2 FB3 FA1 FA2 + FA3 FC1 FC2 + FC3 FC4
F. verticillioides10026 250.1 94.8 19.3 0.5 0.9 17.6 2.0 0.710027 288.7 88.0 18.0 2.1 3.0 45.4 8.3 3.91744 160.2 54.0 15.2 2.6 3.7 17.0 10.0 6.1
F. proliferatum7595 67.8 29.5 17.2 0.3 0.4 0.8 0.1 0.015876 17.3 5.9 8.9 0.0 0.0 0.6 0.0 0.0
Fumonisins are measured in mg/kg.
4. Discussion
F. verticillioides and F. proliferatum strains used in this study, knownto be good fumonisin producers (Lazzaro et al., 2012), formed highamounts of FB, followed by FC and FA, whereas FP were never detected.The co-occurrence of B-, A- and C-fumonisins was previously reportedin F. verticillioides (Lazzaro et al., 2012) together with F. proliferatum(Musser and Plattner, 1997) grown on synthetic media. Referring tothe FB chemotypes described by Szécsi et al. (2010), all strains belongto the chemotype FB1 > FB2 > FB3, except ITEM15876which producedmore FB3 than FB2. The production profile of FA was the same in all thestrains: FA2 + FA3 > FA1; while for FC it was FC1 > FC2 + FC3 > FC4,as previously reported by Lazzaro et al. (2012) for F. verticillioides; al-though Shephard et al. (2011) described a chemotype FC4 > FC1 > FC3.
The ability of F. verticillioides strains to produce fumonisin was signif-icantly higher than in F. proliferatum, at least on CMM. This is in accordwith the general consensus on the major role played by F. verticillioidesin pink ear rot on maize and in fumonisin production in nature (Leslieand Summerell, 2007).
In this study, fumonisin production was higher in cornmeal than instarch based media and FA and FC were detected only in the former.Neither F. verticillioides nor F. proliferatum produced FA or FC on CSM.This result clearly suggests the role played by the substrate in fumonisinmodulation: CMM is richer in nutrients than CSM and probably allows amore efficient biosynthesis of the minor derivatives.
A possible explanation of why fumonisin production was more en-hanced on CMM than on CSM can be investigated in the differencesbetween the availability of nutritional compounds in the two media.The cornmeal used for the medium is a commercial pre-gelatinizedproduct: the gelatinization process causes the swelling of starch gran-ules, enabling both amylose and amylopectin to be more available tothe fungus as nutrients. Amylopectin encourages the production of FBanalogues in F. verticillioidesmore than amylose or other carbon sources(BluhmandWoloshuk, 2005), and it is used preferentially over amyloseby the fungus at 20 and 25 °C (Giorni et al., 2009).
It is known that carbon:nitrogen ratio of the substrates plays a crit-ical role in regulating the biosynthesis of fumonisins. In F. proliferatum, ithas been observed that nitrogen-starvation induced FUM gene expres-sion and fumonisin production (Kohut et al., 2009). The peak of produc-tion of fumonisin in our Fusarium cultures grown on CMM occurred at21–30 days, the time-point at which the fungus could have consumedmost of the nitrogen available, but in most cases, fumonisin level thendecreased, probably depending on the effect of other regulating factorsor on the reduction of nutritional sources available.

0
2
4
6
8
7 14 21 30 45 7 14 21 30 45days
ph
1002610027 1587675951744
Fig. 4. Measurements of pH in cornmeal medium in F. verticillioides (ITEM10026–ITEM10027–ITEM1744) and F. proliferatum (ITEM7595–ITEM15876) strains in a 45-day period ofincubation.
26 I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
Indeed it has been shown that other mechanisms, such as pH varia-tion, seem to be involved in fumonisin activation (Picot et al., 2010). Liq-uid cultures of F. proliferatum produced fumonisins at acid pH (2 to 4.5)(Keller et al., 1997) and it was also observed that fumonisin biosynthesisis repressed at alkaline condition, due to the activity of gene PAC1 inF. verticillioides (Flaherty et al., 2003). In our study, pH levels in Fusariumcultures were less acid than in Keller et al. (1997), and did not go below4.50; however, in agreement with that study, we observed a high pro-duction of fumonisin corresponding to the lowest pH values both forF. verticillioides and F. proliferatum strains. The influence of both amylo-pectin and pH on fumonisin accumulation in maize kernels was con-firmed in field experiments: the highest fumonisin production occurredduring the dent stage of maize, which corresponded to an acidifica-tion in kernel pH and the maximum amylopectin content (Picot etal., 2011).
F. verticillioides strains used in this studywere analysed in a previousstudy for their fumonisin production profile onmalt extractmedium andthe fumonisin productionwas lower but with a comparable profile, withFA and FC trend similar to that of FB (Lazzaro et al., 2012). Fumonisin Bproduction was 200 times lower (average of total FB produced) than incultures grownonCMM, and FA and FC 100 and 400 times lower, respec-tively. This huge difference in fumonisin biosynthesis can be ascribed tothedifferent growthmedia used in the two studies:malt extractmediumis composed of simple sugars and peptides, whereas CMM containsstarch (72.1%), proteins (8.7%), fibre (3.1%), lipids (2.7%) and solublesugars (1.5%). This substrate is thus a good source of amylose and, in par-ticular, of amylopectinwhich is considered, as just stressed, an importantinducing factor in fumonisin production. In addition, although specificdata are still lacking, the other components may also play an importantrole in fumonisin biosynthesis modulation. Further studies should beplanned to better clarify this effect, including the expression of FUMgenes in different nutritional regimens.
In this study, the occurrence of hidden fumonisins has also beenaddressed. Although the amount of hidden fumonisins found underthe studied conditions was quite low compared to that reported inprevious studies using field samples of maize kernels (Dall'Asta et al.,2012), the data suggest thatmasking effectsmay occur in CMM,whereashidden fumonisins have never been found at significant levels in CSM.Since the accumulation of fumonisins is generally lower on CSM, this dif-ference can be ascribed both to the different production induced by theconsidered substrates or to the media composition. On the other hand,the occurrence of low amounts of hidden fumonisins in CMM seems tosuggest that the masking phenomenon is mainly matrix-driven andcould be described as a physical interaction between mycotoxins andmatrix constituents. Further studies should be performed in order to bet-ter understand the role played by the substrate in accumulation of hid-den fumonisins, i.e. testing other minor components of maize kernel(e.g. sucrose, fatty acids, proteins), trying to identify their degree of in-volvement in fumonisin production and in themasking effect processes.In addition, in planta studies should be crucial to clarify the eventualrole in the masking actions of the living plant in the host–pathogeninteraction.
Acknowledgements
This workwas supported by theMinistry of University and Research(project PRIN 2009).
References
Abbas, H.K., Shier, W.T., Seo, J.A., Lee, Y.W., Musser, S.M., 1998. Phytotoxicity and cytotox-icity of the fumonisin C and P series of mycotoxins from Fusarium spp. fungi. Toxicon12, 2033–2037.
Bartόk, T., Szécsi, Á., Szekeres, A., Mesterházy, Á., Bartόk, M., 2006. Detection of newfumonisin mycotoxins and fumonisin-like compounds by reversed-phase high-performance liquid chromatography/electrospray ionization ion trap mass spec-trometry. Rapid Communications in Mass Spectrometry 20, 2447–2462.
Battilani, P., Logrieco, A., 2013. Global risk maps for mycotoxins in wheat and maize. In:Leslie, J., Logrieco, A. (Eds.), Mycotoxin Reduction in Grain Chains: A Practical Guide.Wiley-Blackwell.
Battilani, P., Pietri, A., Barbano, C., Scandolara, A., Bertuzzi, T., Marocco, A., 2008. Logisticregression modelling of cropping systems to predict fumonisin contamination inmaize. Journal of Agricultural and Food Chemistry 56, 10433–10438.
Berthiller, Crews, C., Dall'Asta, C., De Saeger, S., Haesaert, G., Karlovsky, P., Oswald, P.,Seefelder, W., Speijers, G., Stroka, J., 2012. Masked mycotoxins: a review. MolecularNutrition & Food Research 57, 165–186.
Bittencourt, A.B.F., Oliveira, C.A.F., Dilkin, P., Corrêa, B., 2005. Mycotoxin occurrence incorn meal and flour traded in São Paulo, Brazil. Food Control 16, 117–120.
Bluhm, B.H., Woloshuk, C.P., 2005. Amylopectin induces fumonisin B1 production byFusarium verticillioides during colonization of maize kernels. Molecular Plant-Microbe Interactions 18, 1333–1339.
Dall'Asta, C., Falavigna, C., Galaverna, G., Dossena, A., Marchelli, R., 2010. In vitro diges-tion assay for determination of hidden fumonisins in maize. Journal of Agriculturaland Food Chemistry 58, 12042–12047.
Dall'Asta, C., Falavigna, C., Galaverna, G., Battilani, P., 2012. Role of maize hybrids andtheir chemical composition in Fusarium infection, fumonisin production andmasking.Journal of Agricultural and Food Chemistry 60, 3800–3808.
Dall'Erta, A., Cirlini, M., Dall'Asta, M., Del Rio, D., Galaverna, G., Dall'Asta, C., 2013.Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota re-leasing their aglycones. Chemical Research in Toxicology 26, 305–312.
EHC, 2000. Environmental Health Criteria 219: Fumonisin B1, International Programmeon Chemical Safety (IPCS; UNEP, ILO and WHO). In: Marasas, W.H.O., Miller, J.D.,Riley, R.T., Visconti, A. (Eds.), WHO, Geneva.
Falavigna, C., Cirlini, M., Galaverna, G., Sforza, S., Dossena, A., Dall'Asta, C., 2012a. LC/ESI-MS/MS analysis outlines the different fumonisin patterns produced byF. verticillioides in culture media and inmaize kernels. Journal of Mass Spectrometry47, 1170–1176.
Falavigna, C., Cirlini, M., Galaverna, G., Dall'Asta, C., 2012b. Masked fumonisins inprocessed food: co-occurrence of hidden and bound forms and their stability underdigestiveconditions. World Mycotoxin Journal 5, 325–334.
Flaherty, J.E., Pirttilä, A.M., Bluhm, B.H., Woloshuk, C.P., 2003. PAC1, a pH-regulatorygene from Fusarium verticillioides. Applied and Environmental Microbiology 69,5222–5227.
Garrido, C.E., Hernandez Pezzani, C., Pacin, A., 2012. Mycotoxins occurrence inArgentina's maize (Zea mays L.), from 1999 to 2010. Food Control 25, 660–665.
Giorni, P., Magan, N., Battilani, P., 2009. Environmental factors modify carbon nutritionalpatterns and niche overlap between Aspergillus flavus and Fusarium verticillioidesstrains from maize. International Journal of Food Microbiology 130, 213–218.
Howard, P.C., Eppley, R.M., Stack, M.E., Warbritton, A., Voss, K.A., Lorentzen, R.J.,Kovach, R.M., Bucci, T.J., 2001. Fumonisin B-1 carcinogenicity in a two-year feedingstudy using F344 rats and B6C3F(1) mice. Environmental Health Perspectives 109(Suppl. 2), 277–282.
IARC, 1993. IARC monographs on the evaluation of carcinogenic risks to humans. SomeNaturally Occurring Substances: Food Items and Constituents, Heterocyclic Aro-matic Amines and Mycotoxins. IARC Press, Lyon, pp. 445–466.
Keller, S.E., Sullivan, T.M., Chirtel, S., 1997. Factors affecting the growth of Fusariumproliferatum and the production of fumonisin B1: oxygen and pH. Journal of Indus-trial Microbiology and Biotechnology 19, 305–309.

27I. Lazzaro et al. / International Journal of Food Microbiology 166 (2013) 21–27
Kohut, G., Adám, A.L., Fazekas, B., Hornok, L., 2009. N-starvation stress induced FUMgene expression and fumonisin production is mediated via the HOG-type MAPKpathway in Fusarium proliferatum. International Journal of Food Microbiology 130,65–69.
Lazzaro, I., Falavigna, C., Dall'Asta, C., Proctor, R.H., Galaverna, G., Battilani, P., 2012.Fumonisins B, A and C production and masking in Fusarium verticillioides andF. proliferatum grown with different water activity regimes. International Journalof Food Microbiology 159, 93–100.
Leslie, J.F., Summerell, B.A., 2007. The Fusarium Laboratory Manual. Blackwell Publishing,Ames, Iowa, USA.
Marasas, W.F., Riley, R.T., Hendricks, K.A., Stevens, V.L., Sadler, T.W., Gelineau-Van Waes, J.,Missmer, S.A., Cabrera, J., Torres, O., Gelderblom, W.C., Allegood, J., Martinez, C.,Maddox, J., Miller, J.D., Starr, L., Sullards, M.C., Roman, A.V., Voss, K.A., Wang, E., MerrillJr., A.H., 2004. Fumonisins disrupt sphingolipid metabolism, folate transport, and neuraltube development in embryo culture and in vivo: a potential risk factor for human neu-ral tube defects among populations consuming fumonisin-contaminated maize. Journalof Nutrition 134, 711–716.
Marín, S., Magan, N., Belli, N., Ramos, A.J., Canela, R., Sanchis, V., 1999. Two-dimensionalprofiles of fumonisin B1 production by Fusarium moniliforme and Fusarium proliferatumin relation to environmental factors and potential for modelling toxin formation inmaize grain. International Journal of Food Microbiology 51, 159–167.
Musser, S.M., Plattner, R.D., 1997. Fumonisin composition in cultures of Fusariummoniliforme, Fusarium proliferatum, and Fusarium nygami. Journal of Agriculturaland Food Chemistry 45, 1169–1173.
Musser, S.M., Gay, M.L., Mazzola, E.P., Plattner, R.D., 1996. Identification of a new se-ries of fumonisin containing 3-hydroxypiridine. Journal of Natural Products 59,970–972.
Picot, A., Barreau, C., Pinson-Gadais, L., Caron, D., Lannou, C., Richard-Forget, F., 2010.Factors of the Fusarium verticillioides-maize environment modulating fumonisinproduction. Critical Reviews in Microbiology 1–11.
Picot, A., Barreau, C., Pinson-Gadais, L., Piraux, F., Caron, D., Lannou, C., Richard-Forget,F., 2011. The dent stage of maize kernels is the most conducive for fumonisin bio-synthesis under field conditions. Applied and Environmental Microbiology 77,8382–8390.
Rheeder, J.P., Marasas, W.O., Vismer, H.F., 2002. Production of fumonisin analogs byFusarium species. Applied and Environmental Microbiology 68, 2101–2105.
Schroeder, J.J., Smith, E.R., Plattner, R.D., Abbas, H., Yoo, H.S., Merrill Jr., A.H., 1996. Evi-dence for disruption of sphingolipid metabolism as a contributing factor in the toxic-ity and carcinogenicity of fumonisins. Natural Toxins 4, 3–15.
Scott, P.M., 2012. Recent research on fumonisin: a review. Food Additives and Contaminants:Part A 29, 242–248.
Seo, J.A., Lee, Y.W., 1999. Natural occurrence of the C series of fumonisins in moldycorn. Applied and Environmental Microbiology 65, 1331–1334.
Shephard, G.S., van der Westhuizen, L., Sewram, V., van Zyl, J., Rheeder, J.P., 2011. Oc-currence of the C-series fumonisins in maize from the former Transkei region ofSouth Africa. Food Additives and Contaminants: Part A 28, 1712–1716.
Silva, L.J.G., Lino, C.M., Pena, A., Moltó, J.C., 2007. Occurrence of fumonisins B1 and B2 inPortuguese maize and maize-based foods intended for human consumption. FoodAdditives and Contaminants: Part A 24, 381–390.
Smith, W.C., Betrán, J., Runge, C.A., 2004. Corn: Origin, History, Technology and Produc-tion. John Wiley & Sons Inc., Hoboken, NJ, USA (976 pp).
Szécsi, Á., Szekeres, A., Bartόk, T., Oros, G., Bartόk, M., Mesterhazy, Á., 2010. FumonisinB1–4-producing capacity of Hungarian Fusarium verticillioides isolates. World MycotoxinJournal 3, 67–76.
Related Documents











