AIDS RESEARCH AND HUMAN RETROVIRUSES Volume 18, Number 5, 2002, pp. 343–352 © Mary Ann Liebert, Inc. Coreceptor Change Appears after Immune Deficiency Is Established in Children Infected with Different HIV-1 Subtypes C. CASPER, 1,2 L. NAVÉR, 3 P. CLEVESTIG, 1 E. BELFRAGE, 4 T. LEITNER, 5 J. ALBERT, 6 S. LINDGREN, 7 C. OTTENBLAD, 8 A.-B. BOHLIN, 3 E.M. FENYÖ, 1,9 and A. EHRNST 1 ABSTRACT Change of HIV-1 coreceptor use has been connected to progression of disease in children infected with HIV- 1, presumably subtype B. It has not been possible to discern whether the appearance of new viral phenotypes precedes disease development or comes as a consequence of it. We studied the evolution of coreceptor use in HIV-1 isolates from 24 vertically infected children. Their clinical, virological, and immunological status was recorded and the env V3 subtype was determined by DNA sequencing. Coreceptor use was tested on human cell lines, expressing CD4 together with CCR5, CXCR4, and other chemokine receptors. The children car- ried five different env subtypes (nine A, five B, four C, three D, and one G) and one circulating recombinant form, CRF01_AE (n 5 2). Of the 143 isolates, 86 originated from peripheral blood mononuclear cells (PBMCs) and 57 originated from plasma, received at 90 time points. In 52 of 54 paired plasma and PBMC isolates the coreceptor use was concordant. All 74 isolates obtained at 41 time points during the first year of life used CCR5. A change from use of CCR5 to use of CXCR4 occurred in four children infected with subtype A, D, or CRF01_AE after they had reached 1.5 to 5.8 years of age. There was a significant association with de- creased CD4 1 cell levels and severity of disease but, interestingly, the coreceptor change appeared months or even years after the beginning of the immunological deterioration. Thus CXCR4-using virus may emerge as a possible consequence of immune deficiency. The results provide new insights into AIDS development in chil- dren. 343 INTRODUCTION G ENETIC DIVERSITY of human immunodeficiency virus type 1 (HIV-1) is a hallmark of the epidemic of the acquired immunodeficiency syndrome (AIDS). The associated evolu- tion of variants may influence the ability of the immune sys- tem to control HIV-1 infection. Group M represents the main group of HIV-1 strains and has been subdivided into nine dif- ferent subtypes: A, B, C, D, F, G, H, J, and K. 1,2 In addition, several circulating recombinant forms (CRFs) have spread epidemically and may play a role in the global pandemic. The most important intersubtype recombinants are CRF01_AE, CRF02_AG, and CRF03_AB, denoting a recombination of genes from different subtypes as indicated by the name. Oth- 1 Microbiology and Tumor Biology Center, Karolinska Institutet, SE-171 77 Stockholm, Sweden. 2 Neonatal Unit, Department of Women’s and Children’s Health, Karolinska Institutet, SE-171 77 Stockholm, Sweden. 3 Unit of Pediatrics, Department of Clinical Science, Huddinge University Hospital, Karolinska Institutet, SE-171 77 Stockholm, Sweden. 4 Childhood Cancer Research Unit, Department of Women’s and Children’s Health, Karolinska Institutet, SE-171 77 Stockholm, Sweden. 5 Department of Clinical Virology, Swedish Institute for Infectious Disease Control, SE-172 82 Solna, Sweden. 6 Division of Clinical Virology, Department of Immunology, Microbiology, and Pathology, Huddinge University Hospital, Karolinska Insti- tutet, SE-171 77 Stockholm, Sweden. 7 Unit of Obstetrics and Gynecology, Department of Clinical Science, Huddinge University Hospital, Karolinska Institutet, SE-171 77 Stock- holm, Sweden. 8 Unit of Addictive Prevention, Department of Clinical Neuroscience and Internal Medicine, Huddinge University Hospital, Karolinska Insti- tutet, SE-171 77 Stockholm, Sweden. 9 Department of Medical Microbiology, Dermatology, and Infection, Lund University Hospital, SE 221 85, Lund, Sweden.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AIDS RESEARCH AND HUMAN RETROVIRUSESVolume 18, Number 5, 2002, pp. 343–352© Mary Ann Liebert, Inc.
Coreceptor Change Appears after Immune Deficiency IsEstablished in Children Infected with Different
HIV-1 Subtypes
C. CASPER,1,2 L. NAVÉR,3 P. CLEVESTIG,1 E. BELFRAGE,4 T. LEITNER,5 J. ALBERT,6 S. LINDGREN,7
C. OTTENBLAD,8 A.-B. BOHLIN,3 E.M. FENYÖ,1,9 and A. EHRNST1
ABSTRACT
Change of HIV-1 coreceptor use has been connected to progression of disease in children infected with HIV-1, presumably subtype B. It has not been possible to discern whether the appearance of new viral phenotypesprecedes disease development or comes as a consequence of it. We studied the evolution of coreceptor use inHIV-1 isolates from 24 vertically infected children. Their clinical, virological, and immunological status wasrecorded and the env V3 subtype was determined by DNA sequencing. Coreceptor use was tested on humancell lines, expressing CD4 together with CCR5, CXCR4, and other chemokine receptors. The children car-ried five different env subtypes (nine A, five B, four C, three D, and one G) and one circulating recombinantform, CRF01_AE (n 5 2). Of the 143 isolates, 86 originated from peripheral blood mononuclear cells (PBMCs)and 57 originated from plasma, received at 90 time points. In 52 of 54 paired plasma and PBMC isolates thecoreceptor use was concordant. All 74 isolates obtained at 41 time points during the first year of life usedCCR5. A change from use of CCR5 to use of CXCR4 occurred in four children infected with subtype A, D,or CRF01_AE after they had reached 1.5 to 5.8 years of age. There was a significant association with de-creased CD41 cell levels and severity of disease but, interestingly, the coreceptor change appeared months oreven years after the beginning of the immunological deterioration. Thus CXCR4-using virus may emerge asa possible consequence of immune deficiency. The results provide new insights into AIDS development in chil-dren.
343
INTRODUCTION
GENETIC DIVERSITY of human immunodeficiency virus type1 (HIV-1) is a hallmark of the epidemic of the acquired
immunodeficiency syndrome (AIDS). The associated evolu-tion of variants may influence the ability of the immune sys-tem to control HIV-1 infection. Group M represents the main
group of HIV-1 strains and has been subdivided into nine dif-ferent subtypes: A, B, C, D, F, G, H, J, and K.1,2 In addition,several circulating recombinant forms (CRFs) have spreadepidemically and may play a role in the global pandemic. Themost important intersubtype recombinants are CRF01_AE,CRF02_AG, and CRF03_AB, denoting a recombination ofgenes from different subtypes as indicated by the name. Oth-
1Microbiology and Tumor Biology Center, Karolinska Institutet, SE-171 77 Stockholm, Sweden.2Neonatal Unit, Department of Women’s and Children’s Health, Karolinska Institutet, SE-171 77 Stockholm, Sweden.3Unit of Pediatrics, Department of Clinical Science, Huddinge University Hospital, Karolinska Institutet, SE-171 77 Stockholm, Sweden.4Childhood Cancer Research Unit, Department of Women’s and Children’s Health, Karolinska Institutet, SE-171 77 Stockholm, Sweden.5Department of Clinical Virology, Swedish Institute for Infectious Disease Control, SE-172 82 Solna, Sweden.6Division of Clinical Virology, Department of Immunology, Microbiology, and Pathology, Huddinge University Hospital, Karolinska Insti-
tutet, SE-171 77 Stockholm, Sweden.7Unit of Obstetrics and Gynecology, Department of Clinical Science, Huddinge University Hospital, Karolinska Institutet, SE-171 77 Stock-
holm, Sweden.8Unit of Addictive Prevention, Department of Clinical Neuroscience and Internal Medicine, Huddinge University Hospital, Karolinska Insti-
tutet, SE-171 77 Stockholm, Sweden.9Department of Medical Microbiology, Dermatology, and Infection, Lund University Hospital, SE 221 85, Lund, Sweden.

ers can be even more complex, such as CRF04_cpx (complex).3,4
Much of our understanding of the biology and pathogenesis ofHIV-1 comes from analyses of subtype B, which predominatesin Europe and the United States, but which is infrequent inAfrica, where the prevalence of HIV-1 is the highest.
Subtype-specific differences have been observed in vitrowith regard to the use of coreceptors5–8 for virus infection ofcells. The chemokine receptors CCR5 and CXCR4 may be usedas coreceptors,9 whereas CD4 is the major receptor of HIV-1for entry into susceptible cells. HIV-1 isolates can be classifiedinto two major groups based on the ability to use either CCR5or CXCR4 as coreceptor.9 The corresponding phenotypes of therespective viruses are called R5 and X4, or R5X4 when bothcoreceptors are used.10 Earlier, HIV-1 phenotypes were distin-guished by their ability to induce syncytia in MT-2 cells,11
which indicated use of the CXCR4 receptor. Experimentsshowed later that syncytium-inducing virus was synonymouswith virus using (or not using) the CXCR4 receptor.12
The emergence of X4 virus has more recently been associ-ated with disease progression in adults,13 as well as in chil-dren.14 It was not possible, however, to discern whether the appearance of new viral phenotypes preceded disease develop-ment or came as a consequence.13,14 On the other hand, the ap-pearance of syncytium-inducing virus has been observed in chil-dren as an age-related and late phenomenon.15,16 The relationto disease progression is less clear in children but most studiesfind a similar impact on disease progression as in adults,16,17
whereas others failed to find such a correlation.18
The aim of this study was to document the evolution of core-ceptor use of HIV-1 isolates obtained from both plasma and pe-ripheral blood mononuclear cells (PBMCs) in children, infectedwith HIV-1 from their mothers, who originated from many partsof the world. Thus a broad representation of the genetic sub-types of HIV-1 was anticipated. Most children were monitoredfrom birth. Their immunological status and virological statuswere documented prospectively.19–21
MATERIALS AND METHODS
Subjects
The children, later diagnosed to be HIV-1 infected, were bornbetween March 1987 and September 1994. HIV-1 was isolatedfrom 25 of them until February 1996. One infected child wasexcluded, as the virus isolates did not grow on passage. Thus24 children were studied.
Nineteen of the children were born to mothers with a previ-ously known infection or who were detected through a nationalscreening program for HIV-1 in pregnant women.22 They weremonitored prospectively from birth. Five children (nos. 1, 6,13, 17, and 18) were included later, when they were identifiedas HIV-1 infected at the age of 41, 45, 29, 69, and 10 months,respectively. These children were also presumed to have ac-quired HIV-1 from their HIV-1-infected mothers. Their moth-ers had lived outside of Sweden when giving birth, or in onecase the mother became infected close to delivery or soon af-ter and then breast-fed her baby.
No mother or child received zidovudine prophylaxis to re-duce vertical transmission, as the children were born before the
introduction of the ACTG-076 protocol.23 Three mothers wereintravenous drug users and in one case the mode of the mother’sinfection was unknown. The other mothers were classified asheterosexually infected. The mothers gave informed consent toparticipate in the study and were strongly advised not to breast-feed their babies. The five children included at an older agewere breast-fed as well as one child monitored from birth.
Clinical examination and sample collection
Children monitored from birth were examined at birth andat 3-month intervals. Children born in 1991 and later were alsoseen at 3 and 6 weeks and 4.5 months of age.20
Blood samples were collected on all visits, except at 3 weeksand 4.5 months of age. Cord blood was not used. After the firstyear of life, samples for virus isolation were taken at 18 and 24months, and every half or whole year thereafter. Defibrinatedwhole blood was used for virus culture from plasma andPBMCs, as well as for HIV-1 detection by the polymerase chainreaction (PCR). The number of CD41 cells was determined and,retrospectively, plasma RNA levels were determined by use ofa nucleic acid sequence-based amplification (NASBA) HIV-1RNA QT kit (Organon Teknika, The Netherlands). All sampleswere collected before the introduction of highly active anti-retroviral therapy.
Immunological and clinical stages were defined according tothe revised classification system of the Centers for Disease Con-trol and Prevention (CDC, Atlanta, GA) for children less than13 years of age.21 Briefly, there are four clinical stages: N, nosymptoms; A, mild symptoms; B, moderate symptoms; C, se-vere symptoms. The immune deficiency categories are basedon age-specific CD41 cell counts and percentage of total lym-phocytes: In class 1 there is no evidence of suppression, in class2 there is a moderate immune suppression, and in class 3 thereis severe immune suppression.21
HIV-1 diagnosis of the children
The HIV-1 diagnosis was based on virus isolation, and/or HIV-1 DNA PCR, and/or HIV serology,19,20 which in relation to theage of the child was paired with maternal serum. Virus isolationwas carried out as described.24 Whenever the volume of bloodallowed, virus isolation from PBMCs and plasma was performedin parallel. For HIV-1 DNA PCR at least two different sets ofprimers were used from the gag, pol, and/or env region of HIV-1.25 HIV-1 antibodies were detected by an enzyme-linked im-munosorbent assay (ELISA; Abbott, Abbott Park, IL) and con-firmed by Western blot. Children with HIV-specific antibodiesafter 18 months of age were defined as HIV-1 infected and thosewithout HIV-specific antibodies were defined as uninfected.
Passage of viral isolates
Virus was isolated from all blood samples obtained for di-agnosis and from most samples obtained during the first year,but thereafter only from samples taken every half or full year.One hundred and fifty-seven HIV-1 isolates from the initial iso-lation or the first passage were available and used to infect phy-tohemagglutinin-stimulated PBMCs from two healthy blooddonors.24 The passage was successful for 143 isolates, 86 ofwhich originated from PBMCs and 57 from plasma.
CASPER ET AL.344

Assays for coreceptor use by HIV-1
The coreceptor use of the virus isolates was determined us-ing U87.CD4 cells, expressing the chemokine receptor CCR1,CCR2b, CCR3, CCR5, or CXCR4,9 and GHOST(3) cells, ex-pressing CCR5, CXCR4, or the orphan receptor BOB orBonzo.26
Briefly, 24-well plates, each with different U87.CD4 cells,were incubated at 37°C in a humidified 5% CO2 atmosphereuntil the cultures reached half-confluence and were then co-cultured with PBMCs infected with the different primary iso-lates since 10 days. The cell cultures were observed for syn-cytium formation daily for 7 days. An ELISA was used todetermine the amount of p24 antigen in supernatants, collectedon day 1 (before the washes), day 4, and day 7. A culture wellwas considered positive if syncytium formation and increasingamounts of HIV-1 p24 antigen were observed.
GHOST(3) cells carry the HIV-2 long terminal repeat (LTR)-driven green fluorescent protein (GFP) marker.26 The greenlight is emitted in virus-positive cells and can easily be observedin a fluorescence microscope and/or by flow cytometry. Thesecells express CXCR4 endogenously, which calls for caution ininterpretation of coreceptor use of viruses capable of usingCXCR4. Infection by coculture was performed as withU87.CD4 cells. Three to 4 days after infection, the cultures werevisually inspected in a fluorescence microscope. If fluorescencewas observed for only the cell line expressing CCR5, a p24antigen ELISA was performed on all cell lines. If fluorescencewas observed in other than the CCR5-expressing cells, the wellswere prepared for fluorescence-activated cell sorting (FACS)of p24 antigen after harvest. The proportion of fluorescent cellsand the fluorescence intensity were measured as previously de-scribed.27 Results of flow cytometry were considered positiveif at least 0.5% of the cells emitted green fluorescence at least2-fold greater than that of the negative control. The results ob-tained by fluorescence microscopy and flow cytometry were inagreement.
All isolates were tested for CCR5 and CXCR4 use on bothcell systems and the results showed good agreement. In one in-stance the p24 antigen rose between days 1 and 7, but syncy-tia were apparent only on day 7 in the cell line U87.CD4.CXCR4 and the results were negative in the cell lineGHOST(3)CXCR4. The corresponding HIV-1 phenotypes wereclassified as R5, X4, or R5X4.10
Subtype determination by phylogenetic tree analysis of env DNA sequences
The genetic subtype1,2 of the virus carried by these individ-uals was determined by sequencing viral DNA from thePBMCs28 or after reverse transcription of patient plasma RNA,extracted by binding the nucleic acid to silica, washing it, andremoving it with guanidine thiocyanate (NASBA; OrganonTeknika). In one case PBMCs of the cultured virus were used.The HIV-1 V3 region of the envelope gene was amplified bya nested PCR.28 The PCR product was purified with a QIAquickDNA purification kit (Qiagen, Chatsworth, CA) and sequencedwith a Big Dye terminator cycle sequencing Ready Reactionkit (PE Biosystems, Foster City, CA). The sequences were de-tected with an ABI Prisma 377 DNA sequencer. The genetic
subtype was determined by phylogenetic tree analysis, usingthe programs DNADIST and NEIGHBOR in the PHYLIP pack-age29 and recommended reference sequences from the LosAlamos Database (http://hiv-web.lanl.gov). Seven subtypes hadbeen determined previously.30
Ethical considerations
The study was approved by the Ethics Committee of theKarolinska Institute (Stockholm, Sweden) (no. 98-184).
Statistical analysis
For statistical analyses the Fisher exact test and the Wilcoxontest were used. Probability (p) values ,0.05 were regarded assignificant.
RESULTS
Distribution of coreceptor use among the patients and isolates
The coreceptor use and genetic subtype of the HIV-1 iso-lates obtained from the 24 children, as well as their age andclinical, virological, and immunological status, are as shown inTables 1 and 2. Virus from 20 children (99 isolates) used CCR5only as coreceptor during the entire study period (median, 36months) (Table 1). The appearance of virus with an X4 phe-notype clustered in 4 of 24 (17%) patients (Table 2).
Among all 143 isolates, 121 (85%) used CCR5 and 22 (15%)used CXCR4, alone or in combination with another chemokinereceptor. Thus in one patient CCR5 was used together with theorphan coreceptors Bob and Bonzo (six isolates), followed bythe emergence of R5X4 dual-tropic virus (five isolates) and lossof viruses using Bob and Bonzo. There were altogether eightdual-tropic R5X4 isolates obtained from three children. In onepatient, R5 virus reappeared after X4 virus had been presentalone for 36 months in 10 isolates.
In the four children in whom a change of phenotype was ob-served, R5 virus was present first (14 isolates), followed by thepresence of X4 virus (12 isolates) (Table 2). Thus the presenceof X4 virus not only clustered among the patients, but also overtime among individual children.
Concordance of receptor use in plasma and PBMC isolates
Because we had collected virus isolates from plasma andPBMCs in parallel, it was possible to determine whether virusisolates from plasma and PBMCs displayed a different pheno-type. Results on coreceptor use were available for 54 pairedplasma and PBMC isolates. Two discordant results were ob-tained in two children (nos. 21 and 23) in whom a coreceptorswitch had occurred or was ongoing (Table 2). The coreceptoruse was concordant in the remaining 52 paired plasma andPBMC isolates. This allowed us the inclusion of only one phe-notype per time point in the further analysis. Samples after 1year of age that were received less than 4 months apart wereomitted to comply with the schedule of the prospective study.In this way altogether 90 sample dates were identified (Tables
CORECEPTOR CHANGE OF HIV-1 IN CHILDREN 345
1 and 2), 26 of which belonged to children with a change in vi-ral phenotype to X4 virus.
Distribution of coreceptor use in virus isolates ofdifferent genetic subtypes
The HIV-1 subtypes reflected the variable origin of the moth-ers. Accordingly, the mothers carrying subtype A virus origi-nated from sub-Saharan Africa (n 5 7, one pair of siblings) orfrom Eastern Europe (n 5 1). One of the siblings developed X4virus and altogether two of the nine children, carrying subtypeA. The subtype of one sibling (case 1) was inferred from thatof the other (case 21). Mothers of the children with viruses ofsubtypes C (n 5 4), D (n 5 3), and G (n 5 1) were also fromsub-Saharan Africa. Of the children with subtype D, one car-ried X4 virus. Children born to mothers who were born in Swe-den (n 5 5) had viruses of subtype B, and none developed an
X4 virus. The mothers of those with CRF01_AE were fromAsia (n 5 2) and one child developed X4 virus. Thus 4 chil-dren of 14 with subtype A, D, or CRF01_AE developed X4virus, whereas none of 10 children with subtype B, C, or G de-veloped X4 virus. There was no statistically significant associ-ation between coreceptor use and any of the subtypes or com-binations of them at the level of the individual.
The isolates of subtype A were taken at 34 time points, andthree contained X4 virus. Subtype D isolates came from 12 timepoints, and seven contained X4 virus. CRF01_AE was isolatedat six time points, and two contained X4 virus. Subtype B wascollected at 22 time points without X4 viruses, nor were thereany X4 viruses found during the isolation of subtype C (14 timepoints) or during the isolation of subtype G (2 time points).
A possible difference between subtypes could be related tothe circulation of infectious virus in plasma. From all the timepoints there were 56 (40%) plasma isolates versus 84 (60%)
CASPER ET AL.346
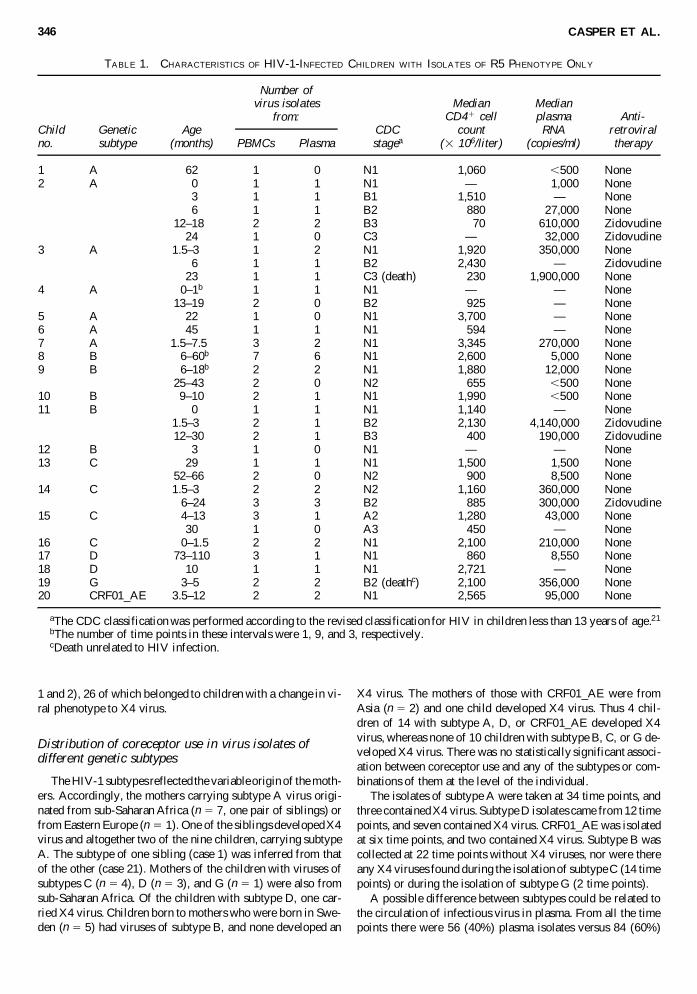
TABLE 1. CHARACTERISTICS OF HIV-1-INFECTED CHILDREN WITH ISOLATES OF R5 PHENOTYPE ONLY
Median MedianCD41 cell plasma Anti-
Child Genetic Age CDC count RNA retroviralno. subtype (months) PBMCs Plasma stagea (3 106/liter) (copies/ml) therapy
1 A 62 1 0 N1 1,060 ,500 None2 A 0 1 1 N1 — 1,000 None
3 1 1 B1 1,510 —00 None6 1 1 B2 880 27,000 None
12–180 2 2 B3 70 610,000 Zidovudine24 1 0 C3 — 32,000 Zidovudine
3 A 1.5–300 1 2 N1 1,920 350,000 None6 1 1 B2 2,430 —00 Zidovudine
23 1 1 C3 (death) 230 1,900,000 None4 A 0–1b 1 1 N1 — —00 None
13–190 2 0 B2 925 —00 None5 A 22 1 0 N1 3,700 —00 None6 A 45 1 1 N1 594 —00 None7 A 1.5–7.50 3 2 N1 3,345 270,000 None8 B 06–60b 7 6 N1 2,600 5,000 None9 B 06–18b 2 2 N1 1,880 12,000 None
25–430 2 0 N2 655 ,500 None10 B 9–10 2 1 N1 1,990 ,500 None11 B 0 1 1 N1 1,140 —00 None
1.5–300 2 1 B2 2,130 4,140,000 Zidovudine12–300 2 1 B3 400 190,000 Zidovudine
12 B 3 1 0 N1 — —00 None13 C 29 1 1 N1 1,500 1,500 None
52–660 2 0 N2 900 8,500 None14 C 1.5–300 2 2 N2 1,160 360,000 None
.6–24 3 3 B2 885 300,000 Zidovudine15 C .4–13 3 1 A2 1,280 43,000 None
30 1 0 A3 450 —00 None16 C 00–1.5 2 2 N1 2,100 210,000 None17 D 73–110 3 1 N1 860 8,550 None18 D 10 1 1 N1 2,721 —00 None19 G 3–5. 2 2 B2 (deathc) 2,100 356,000 None20 CRF01_AE 3.5–120 2 2 N1 2,565 95,000 None
aThe CDC classification was performed according to the revised classification for HIV in children less than 13 years of age.21
bThe number of time points in these intervals were 1, 9, and 3, respectively.cDeath unrelated to HIV infection.
Number of virus isolates
from:
from PBMCs. The proportion of plasma isolates was distrib-uted among the subtypes as follows. In subtype A there were20 of 52 (38.46%) plasma isolates, in subtype B there were12 of 31 (38.7%), in subtype C there were 9 of 21 (39.1%),in subtype D there were 10 of 21 (47.6%), in CRF01_AEthere were 3 of 9 (33.3%), and in subtype G there were 2 of4 (50%). None of the differences was statistically significant,p . 0.1.
Relationship between RNA load and coreceptor use
There was no obvious association between RNA load andthe presence of X4 virus. The X4 virus appeared along with be-tween 45,000 and 230,000 HIV-1 RNA copies/ml. The highestpeaks were observed in four children with the R5 phenotypeonly. An analysis of the RNA peak copy numbers in the twogroups of children showed no significant difference, using theWilcoxon test, nor did age at the peak. There was no tendencyof RNA copy numbers per milliliter to increase along with theappearance of the X4 phenotype.
Change of coreceptor use in relation to immune deficiency
As both coreceptor use and disease progression toward AIDSshow dynamic evolution over time, it was important to analyzethese components in relation to one another. Were the childrenwith X4 viruses simply more ill than those carrying only R5viruses?
At the last time of observation there were 11 children whowere in immunological class 1 and 5 children in class 2, all ofwhom harbored R5 virus. There were eight children who hadreached class 3, of whom four had an X4 virus. The differencewas statistically significant when comparing children in classes1 and 2 with those in class 3, p 5 0.007, as evidence for theappearance of X4 virus late in the evolution of the immune de-ficiency.
When the analysis was extended to the number of samplingtime points, only one per immunological class (the last time pointfor each class) for each individual was included. The associationbetween the development of the immune deficiency and the X4
CORECEPTOR CHANGE OF HIV-1 IN CHILDREN 347
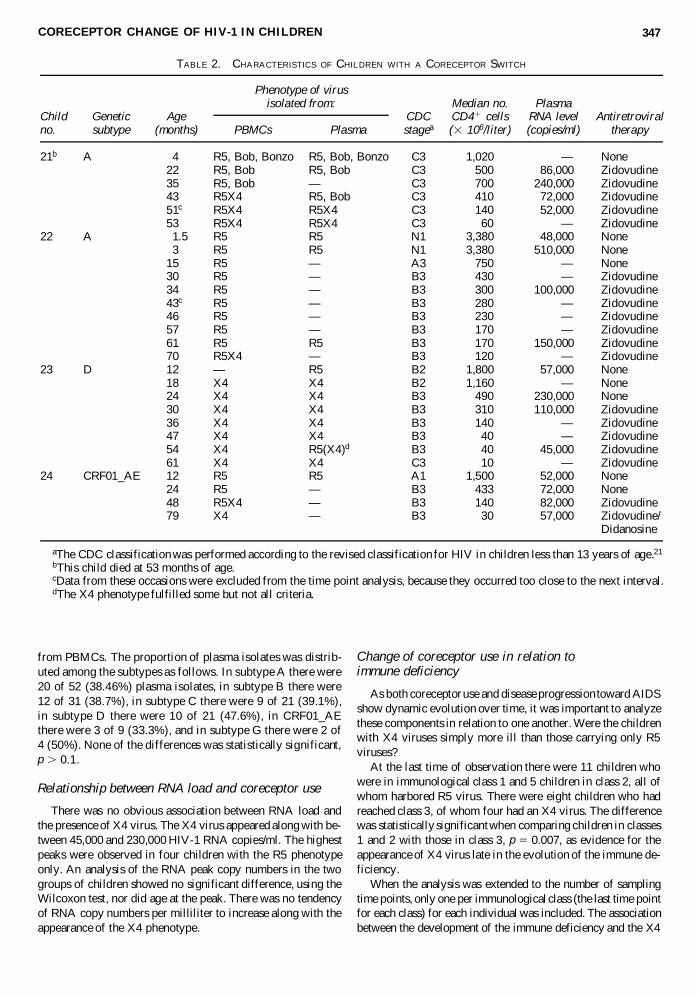
TABLE 2. CHARACTERISTICS OF CHILDREN WITH A CORECEPTOR SWITCH
Median no. PlasmaChild Genetic Age CDC CD41 cells RNA level Antiretroviralno. subtype (months) PBMCs Plasma stagea (3 106/liter) (copies/ml) therapy
21b A 4 R5, Bob, Bonzo R5, Bob, Bonzo C3 1,020 — None22 R5, Bob R5, Bob C3 500 86,000 Zidovudine35 R5, Bob — C3 700 240,000 Zidovudine43 R5X4 R5, Bob C3 410 72,000 Zidovudine51c R5X4 R5X4 C3 140 52,000 Zidovudine53 R5X4 R5X4 C3 60 — Zidovudine
22 A 51.5 R5 R5 N1 3,380 48,000 None3 R5 R5 N1 3,380 510,000 None
15 R5 — A3 750 — None30 R5 — B3 430 — Zidovudine34 R5 — B3 300 100,000 Zidovudine43c R5 — B3 280 — Zidovudine46 R5 — B3 230 — Zidovudine57 R5 — B3 170 — Zidovudine61 R5 R5 B3 170 150,000 Zidovudine70 R5X4 — B3 120 — Zidovudine
23 D 12 — R5 B2 1,800 57,000 None18 X4 X4 B2 1,160 — None24 X4 X4 B3 490 230,000 None30 X4 X4 B3 310 110,000 Zidovudine36 X4 X4 B3 140 — Zidovudine47 X4 X4 B3 40 — Zidovudine54 X4 R5(X4)d B3 40 45,000 Zidovudine61 X4 X4 C3 10 — Zidovudine
24 CRF01_AE 12 R5 R5 A1 1,500 52,000 None24 R5 — B3 433 72,000 None48 R5X4 — B3 140 82,000 Zidovudine79 X4 — B3 30 57,000 Zidovudine/
Didanosine
aThe CDC classification was performed according to the revised classification for HIV in children less than 13 years of age.21
bThis child died at 53 months of age.cData from these occasions were excluded from the time point analysis, because they occurred too close to the next interval.dThe X4 phenotype fulfilled some but not all criteria.
Phenotype of virus isolated from:

phenotype became even clearer (Table 3A). These comparisonswere statistically significant not only for classes 1 and 2 versusclass 3, but also for class 1 versus classes 2 and 3.
However, during the 26 time points of sampling in the fourchildren in whom HIV-1 displayed a change of phenotype, thedistribution of samples over immunological class was similarfor R5 and X4 phenotypes (Table 2). This was even more ob-vious in the whole material, where there were more time pointswhen only R5 virus was present as compared with X4 in class2 and an equal number during class 3 (Table 3A). Thus, whereasX4 virus appeared when signs of severe immune deficiencywere present, the reverse was not true.
Change of coreceptor use in relation to clinical stage
The analysis was extended to the relationship between thepresence of X4 virus and clinical stage at the individual level.Most of the children were still in clinical stage N (n 5 13) orA (n 5 1) at the end of the study, all of whom had R5 virusesonly. There were six children in stage B2 with an X4 virus, andfour in stage C, of whom two had an X4 virus. A comparisonbetween children in stage N versus those in other stages wasstatistically significant, p 5 0.045, as was a comparison be-tween those in stages N and A, versus those in stages B and C,p 5 0.02, but not between those in stages N, A, and B versusthose in stage C, p . 0.1.
When the outcome was analyzed in relation to time points,including only one per clinical stage (the last one) for each pa-tient, it was obvious that X4 virus was not present in the asymp-tomatic phase of infection (Table 3B). This association was alsostatistically significant, even more so than at the individuallevel. Still, there were as many or more sample dates for chil-dren in stages B and C with only R5 viruses versus those with
X4 viruses. Thus, also with regard to clinical stage, X4 virusappeared only when signs of disease were present, but the re-verse was not true.
Change of coreceptor use and disease developmentover time
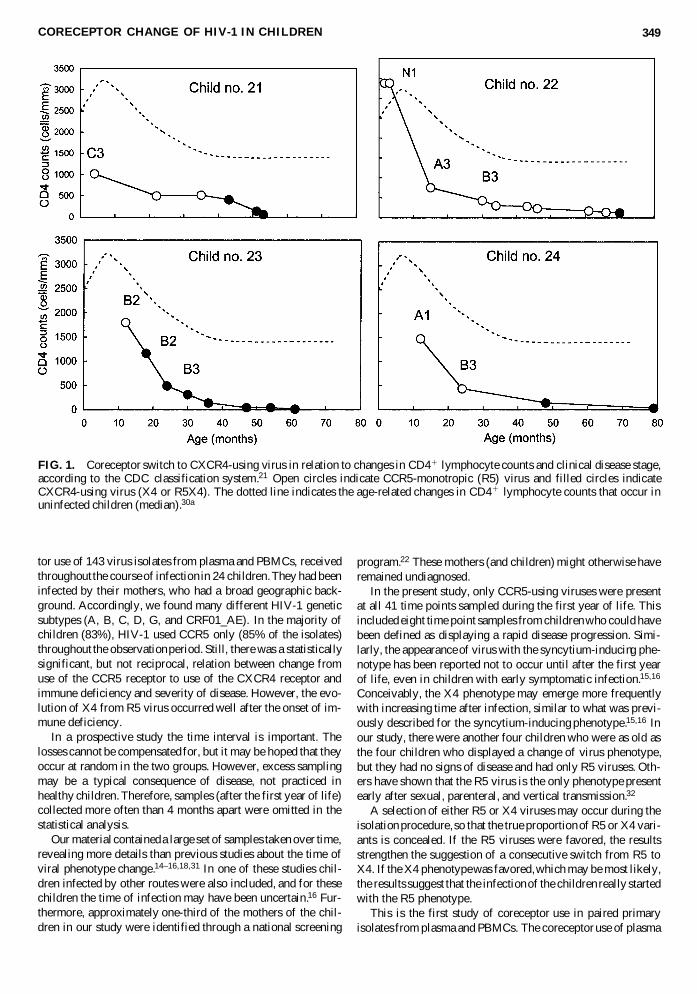
An important question in disease development toward AIDSis whether the observed development of viral phenotype is pre-disposing for, or occurs as a result of, disease development. Inthe present material there were enough time points of observa-tion to allow a resolution of the temporal relationship betweenphenotype switch, CD41 cell decline, and clinical disease progression in the four patients who developed X4 viruses (Fig. 121,30a). In all four patients, the X4 virus appeared wellafter establishment of the immune deficiency.
Appearance of the X4 phenotype in relation tozidovudine treatment
All four children carrying the X4 phenotype were treatedwith zidovudine, while the X4 phenotype was present. In twoof them the zidovudine treatment had begun before the ap-pearance of the X4 phenotype. In one child the R5 phenotypereappeared during treatment. Four of the children carrying theR5 phenotype only were also treated with zidovudine. The zi-dovudine treatment did not seem to influence the character ofthe phenotype expressed.
DISCUSSION
To further understand the role of coreceptor use by HIV-1in AIDS development in children, we determined the corecep-
CASPER ET AL.348
TABLE 3. PHENOTYPE DISTRIBUTION AT SAMPLING OF DIFFERENT IMMUNOLOGICAL
CLASSES AND DURING DIFFERENT CLINICAL STAGES
A. Phenotype Distribution at Sampling during Different Immunological Classesa
Immunological class
1 2 3 S
Presence of the X4 phenotype 0 1 4 5Absence of the X4 phenotype 20 9 4 33
S: 20 10 8 38
B. Phenotype Distribution at Sampling during Different Clinical Stagesb
Clinical stage
N A B C S
Presence of the X4 phenotype 0 0 3 2 5Absence of the X4 phenotype 19 3 6 2 30
S: 19 3 9 4 35
aFisher’s exact test, one sided, yielded for class 1 versus classes 2 1 3, p 5 0.017; classes 1 1 2 versus class 3, p 5 0.004.
bFisher’s exact test, one sided, yielded for N versus A 1 B 1 C, p 5 0.013; N 1 A ver-sus B 1 C, p 5 0.004; N 1 A 1 B versus C, p 5 0.083.
tor use of 143 virus isolates from plasma and PBMCs, receivedthroughout the course of infection in 24 children. They had beeninfected by their mothers, who had a broad geographic back-ground. Accordingly, we found many different HIV-1 geneticsubtypes (A, B, C, D, G, and CRF01_AE). In the majority ofchildren (83%), HIV-1 used CCR5 only (85% of the isolates)throughout the observation period. Still, there was a statisticallysignificant, but not reciprocal, relation between change fromuse of the CCR5 receptor to use of the CXCR4 receptor andimmune deficiency and severity of disease. However, the evo-lution of X4 from R5 virus occurred well after the onset of im-mune deficiency.
In a prospective study the time interval is important. Thelosses cannot be compensated for, but it may be hoped that theyoccur at random in the two groups. However, excess samplingmay be a typical consequence of disease, not practiced inhealthy children. Therefore, samples (after the first year of life)collected more often than 4 months apart were omitted in thestatistical analysis.
Our material contained a large set of samples taken over time,revealing more details than previous studies about the time ofviral phenotype change.14–16,18,31 In one of these studies chil-dren infected by other routes were also included, and for thesechildren the time of infection may have been uncertain.16 Fur-thermore, approximately one-third of the mothers of the chil-dren in our study were identified through a national screening
program.22 These mothers (and children) might otherwise haveremained undiagnosed.
In the present study, only CCR5-using viruses were presentat all 41 time points sampled during the first year of life. Thisincluded eight time point samples from children who could havebeen defined as displaying a rapid disease progression. Simi-larly, the appearance of virus with the syncytium-inducing phe-notype has been reported not to occur until after the first yearof life, even in children with early symptomatic infection.15,16
Conceivably, the X4 phenotype may emerge more frequentlywith increasing time after infection, similar to what was previ-ously described for the syncytium-inducing phenotype.15,16 Inour study, there were another four children who were as old asthe four children who displayed a change of virus phenotype,but they had no signs of disease and had only R5 viruses. Oth-ers have shown that the R5 virus is the only phenotype presentearly after sexual, parenteral, and vertical transmission.32
A selection of either R5 or X4 viruses may occur during theisolation procedure, so that the true proportion of R5 or X4 vari-ants is concealed. If the R5 viruses were favored, the resultsstrengthen the suggestion of a consecutive switch from R5 toX4. If the X4 phenotype was favored, which may be most likely,the results suggest that the infection of the children really startedwith the R5 phenotype.
This is the first study of coreceptor use in paired primaryisolates from plasma and PBMCs. The coreceptor use of plasma
CORECEPTOR CHANGE OF HIV-1 IN CHILDREN 349
FIG. 1. Coreceptor switch to CXCR4-using virus in relation to changes in CD41 lymphocyte counts and clinical disease stage,according to the CDC classification system.21 Open circles indicate CCR5-monotropic (R5) virus and filled circles indicateCXCR4-using virus (X4 or R5X4). The dotted line indicates the age-related changes in CD41 lymphocyte counts that occur inuninfected children (median).30a

and PBMC isolates was highly concordant. All except 2 of the54 pairs displayed identical coreceptor use. This finding wasnot entirely expected, because it has been reported that plasmavirus may mirror the actively replicating virus population,whereas PBMC virus may represent an archive of older virusvariants.33 However, genetic studies of viruses in both PBMCsand plasma will include all HIV-1 quasispecies, whether theyare infectious or not, whereas the phenotypes of coreceptor usereflect infectious virus only. Also, others have found a closecorrelation between the amounts of infectious virus in PBMCsand the amounts of HIV-1 RNA in plasma, suggesting that thesetwo populations are similar.34
Subtype-specific in vitro differences between phenotypes ofHIV-1 have been reported.5–8 The role of HIV-1 subtypes in invivo differences is ambiguous.35–39 Our material was too smallto allow a statistical analysis with regard to evaluation of therole of the genetic subtype in the evolution of coreceptor use.Also, the differences in clinical stage of the HIV-1 infectionmay have influenced any relation between coreceptor use andphenotype. Nevertheless, the finding that virus with the X4 phe-notype could be isolated from children carrying subtypes A, D,or CRF01_AE, but not subtype C, is in agreement with earlierstudies. Thus syncytium-inducing virus is rare among isolatesof subtype C,5,6,8 even among AIDS patients.5,8,40 In addition,CXCR4-monotropic use is more common among subtype D iso-lates.5 In the present study the child with the greatest numberof X4 viruses, over the greatest time span, carried HIV-1 ofsubtype D.
We also observed the use of the orphan receptors Bob andBonzo among subtype A isolates, together with the R5 pheno-type, to occur as previously observed.27 It is uncertain what rolethe coreceptors other than CCR5 and CXCR4 may play in vivo.
In a study of adults, it was suggested that the disease coursein patients with the R5 phenotype may be more related to highRNA values than in patients with the X4 virus, in whom thedisease course depended more on a rapid CD41 cell decline.41
This finding could not be confirmed here, but the highest peaksof RNA copy numbers were found in children carrying the R5phenotype only. In a previous study, the X4 phenotype, if itwas present already during the first months of life, was associ-ated with children displaying rapid disease progression.14 Sim-ilarly, it has been observed in one adult case that the syncytium-inducing virus evolved early in the asymptomatic phase ofHIV-1 infection.42 This study suggested that the syncytium-in-ducing virus may result in faster disease progression. We couldnot compare differences in disease progression after the ap-pearance of the X4 phenotype, as these children were alreadygreatly immunosuppressed.
Whereas there was a clear association between the appear-ance of the X4 phenotype and HIV-1 disease, the opposite wasnot true, as in studies of HIV-1 syncytium-inducing viruses inchildren15,16 and adults.43 This motivates a conservative viewof the possible role of X4 virus in disease development in chil-dren. The CXCR4-using virus may, however, be present in or-gans but not in the blood; it would therefore be inaccessible fortesting and thus go undetected.44
The results showed that change in the use of coreceptor fromCCR6 to CXCR4 was clearly associated with HIV-1 diseaseand severe immune deficiency. More importantly, the changeoccurred after establishment of the immune deficiency, emerg-
ing as a possible consequence. The results shed new light onthe role of the X4 phenotype in AIDS development in children.
ACKNOWLEDGMENTS
We thank Kajsa Aperia, Elisabeth Carlenor, Yu Shi, EllenSölver, Kerstin Andreasson, and Robert Fredriksson for tech-nical assistance. This work was supported by grants from theSwedish Medical Research Council, the Swedish Physiciansagainst AIDS Research Foundation, the General MaternityFoundation, the Sven Jerring Foundation, and the SamaritenFoundation.
REFERENCES
1. Carr JK, Foley BT, Leitner T, Korber B, and McCutchan F: Ref-erence sequences representing the principal genetic diversity ofHIV-1 in the pandemic. In: Human Retroviruses and AIDS: A Com-pilation and Analysis of Nucleic Acid and Amino Acid Sequences(Korber B, Kiuken C, Foley B, Hahn B, McCutchan F, Mellors J,and Sodroski J, eds.). Theoretical Biology and Biophysics GroupT-10, Los Alamos Laboratories, Los Alamos, New Mexico, 1998,pp. III10–III19.
2. Robertson DL, Anderson JP, Bradac JA, Carr JK, Foley B,Funkhouser RK, Gao F, Hahn BH, Kalish ML, Kuiken C, LearnGH, Leitner T, McCutchan F, Osmanov S, Peeters M, PieniazekD, Salminen M, Sharp PM, Wolinsky S, and Korber B: HIV-1nomenclature proposal. Science 2000;288:55–56.
3. Gao F, Robertson DL, Carruthers CD, Li Y, Bailes E, KostrikisLG, Salminen MO, Bibollet-Ruche F, Peeters M, Ho DD, ShawGM, Sharp PM, and Hahn BH: An isolate of human immunodefi-ciency virus type 1 originally classified as subtype I represents acomplex mosaic comprising three different group M subtypes (A,G, and I). J Virol 1998;72:10234–10241.
4. Nasioulas G, Paraskevis D, Magiorkinis E, Theodoridou M, andHatzakis A: Molecular analysis of the full-length genome of HIVtype 1 subtype I: Evidence of A/G/I recombination. AIDS Res HumRetroviruses 1999;15:745–758.
5. Tscherning C, Alaeus A, Fredriksson R, Björndal Å, Deng HK,Littman DR, Fenyö EM, and Albert J: Differences in chemokinecoreceptor usage between genetic subtypes of HIV-1. Virology1998;241:181–188.
6. Peeters M, Vincent R, Perret JL, Lasky M, Patrel D, Liegeois F,Courgnaud V, Seng R, Matton T, Molinier S, and Delaporte E: Ev-idence for differences in MT2 cell tropism according to geneticsubtypes of HVI-1: Syncytium-inducing variants seem rare amongsubtype C HIV-1 viruses. J AIDS Hum Retrovirol 1999;20:115–121.
7. Abebe A, Demissie D, Goudsmit J, Brouwer M, Kuiken CL, Pol-lakis G, Schuitemaker H, Fontanet AL, and Rinke de Wit TF: HIV-1 subtype C syncytium- and non-syncytium-inducing phenotypesand coreceptor usage among Ethiopian patients with AIDS. AIDS1999;13:1305–1311.
8. Björndal Å, Sönnerborg A, Tscherning C, Albert J, and Fenyö EM:Phenotypic characteristics of human immunodeficiency virus type1 subtype C isolate of Ethiopian AIDS patients. AIDS Res HumRetroviruses 1999;15:647–653.
9. Deng HK, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M,Dimarzio P, Marmon S, Sutton RE, Hill CM, Davis CB, PeiperSC, Schall TJ, Littman DR, and Landau NR: Identification of amajor co-receptor for primary isolates of HIV-1. Nature 1996;381:661–666.
CASPER ET AL.350

10. Berger EA, Doms RW, Fenyö EM, Korber BTM, Littman DR,Moore JP, Sattentau QJ, Schuitemaker H, Sodroski J, and WeissRA: A new classification for HIV-1. Nature 1998;391:240.
11. Koot M, Vos AH, Keet RP, de Goede RE, Dercksen MW, Terp-stra FG, Coutinho RA, Miedema F, and Tersmette M: HIV-1 bio-logical phenotype in long-term infected individuals evaluated withan MT-2 cocultivation assay. AIDS 1992;6:49–54.
12. Björndal Å, Deng H, Jansson M, Fiore JR, Colognesi C, KarlssonA, Albert J, Scarlatti G, Littman DR, and Fenyö EM: Coreceptorusage of primary human immunodeficiency virus type 1 isolatesvaries according to biological phenotype. J Virol 1997;71:7478–7487.
13. Connor RI, Sheridan KE, Ceradini D, Choe S, and Landau NR:Change in coreceptor use correlates with disease progression inHIV-1-infected individuals. J Exp Med 1997;185:621–628.
14. Scarlatti G, Tresoldi E, Björndal Å, Fredriksson R, Colognesi C,Deng HK, Malnati MS, Plebani A, Siccardi AG, Littman DR,Fenyö EM, and Lusso P: In vivo evolution of HIV-1 co-receptorusage and sensitivity to chemokine-mediated suppression. Nat Med1997;3:1259–1265.
15. Spencer LT, Ogino MT, Dankner WM, and Spector SA: Clinicalsignificance of human immunodeficiency virus type 1 phenotypesin infected children. J Infect Dis 1994;169:491–495.
16. Fitzgibbon JE, Gaur S, Gavai M, Gregory P, Frenkel LD, and JohnJF Jr: Effect of the HIV-1 syncytium-inducing phenotype on dis-ease stage in vertically-infected children. J Med Virol 1998;55:56–63.
17. De Rossi A, Masiero S, Giaquinto C, Ruga E, Comar M, GiaccaM, and Chieco-Bianchi L: Dynamics of viral replication in infantswith vertically acquired human immunodeficiency virus type 1 in-fection. J Clin Invest 1996;97:323–330.
18. Gupta P, Urbach A, Cosentino L, Thampatty P, and Brady M: HIV-1 isolates from children with or without AIDS have similar in vitrobiologic properties. AIDS 1993;7:1561–1564.
19. Navér L, Ehrnst A, Belfrage E, Sönnerborg A, Lidin-Janson G,Christensson B, Ljung R, and Bohlin A-B: Long-term pattern ofHIV-1 RNA load in perinatally infected children. Scand J InfectDis 1999;31:337–343.
20. Navér L, Ehrnst A, Belfrage E, Blomberg J, Christensson B, Fors-gren M, Lidin-Janson G, Lindgren S, Ljung R, Sönnerborg A, andBohlin AB: Broad spectrum of time of detection, primary symp-toms and disease progression in infants with HIV-1 infection. EurJ Clin Microbiol Infect Dis 2001;20:159–166.
21. Centers for Disease Control and Prevention: Revised classificationsystem for HIV infection in children less than 13 years of age.MMWR 1994;43:1–10.
22. Lindgren S, Bohlin A-B, Forsgren M, Arneborn M, Ottenblad C,Lidman K, Anzén B, von Sydow M, and Böttiger M: Screening forHIV-1 antibodies in pregnancy: Results from the Swedish nationalprogramme. Br Med J 1993;307:1447–1451.
23. Connor EM, Sperling RS, Gelber R, Kiselev P, Scott G, O’Sulli-van MJ, Van Dyke R, Bey M, Shearer W, Jacobson RL, JimenezE, O’Neill E, Bazin B, Delfraissy J-F, Culnane M, Coombs R,Elkins M, Moye J, Stratton P, and Balsley J: Reduction of mater-nal–infant transmission of human immunodeficiency virus type 1with zidovudine treatment. Pediatric AIDS Clinical Trials GroupProtocol 076 Study Group. N Engl J Med 1994;331:1173–1180.
24. Ehrnst A, Sönnerborg A, Bergdahl S, and Strannegård Ö: Efficientisolation of HIV from plasma during different stages of HIV in-fection. J Med Virol 1988;26:23–32.
25. Sönnerborg A, Abens J, Johansson B, and Strannegård Ö: Detec-tion of human immunodeficiency virus-1 by polymerase chain re-action and virus cultivation. J Med Virol 1990;31:234–240.
26. Deng HK, Unutmaz D, Kewal Ramani VN, and Littman DR: Ex-pression cloning of new receptors used by simian and human im-munodeficiency viruses. Nature 1997;388:296–300.
27. Tscherning-Casper C, Vödrös D, Menu E, Aperia K, FredrikssonR, Dolcini G, Chaouat G, Barré-Sinoussi F, Albert J, Fenyö EM,and the European Network for in Utero Transmission of HIV-1:Coreceptor usage of HIV-1 isolates representing different geneticsubtypes obtained from pregnant Cameroonian women. J AIDS2000;24:1–9.
28. Leitner T, Korovina G, Marquina S, Smolskaya T, and Albert J:Molecular epidemiology and MT-2 cell tropism of HIV type 1 vari-ant. AIDS Res Hum Retroviruses 1996;12:1595–1603.
29. Felsenstein J: PHYLIP: Phylogenetic Inference Package, version3.52c. University of Washington, Seattle, Washington, 1993.
30. Contag CH, Ehrnst A, Duda J, Bohlin A-B, Lindgren S, Learn GH,and Mullins JI: Mother-to-infant transmission of human immuno-deficiency virus type 1 involving five envelope sequence subtypes.J Virol 1997;71:1292–1300.
30a. European Collaborative Study: Age-related standards for T lym-phocyte subsets based on uninfected children born to human im-munodeficiency virus 1-infected women. Pediatr Infect Dis J1992;11:1018–1026.
31. Resino S, Gurbindo D, Bellón Cano JM, Sanchez-Ramón S, andMuñoz-Fernández MA: Predictive markers of clinical outcome invertically HIV-1-infected infants. A prospective longitudinal study.Pediatr Res 2000;47:509–515.
32. van’t Wout AB, Kootstra NA, Mulder-Kampinga GA, Albrecht-van Lent N, Scherpbier HJ, Veenstra J, Boer K, Coutinho RA,Miedema F, and Schuitemaker H: Macrophage-tropic variants ini-tiate human immunodeficiency virus type 1 infection after sexual,parenteral, and vertical transmission. J Clin Invest 1994;94:2060–2067.
33. Simmonds P, Zhang LQ, McOmish F, Balfe P, Ludlam CA, andBrown AJ: Discontinuous sequence change of human immunode-ficiency virus (HIV) type 1 env sequences in plasma viral and lym-phocyte-associated proviral populations in vivo: Implications formodels of HIV pathogenesis. J Virol 1991;65:6266–6276.
34. Blaak H, de Wolf F, van’t Wout AB, Pakker NG, Bakker M,Goudsmit J, and Schuitemaker H: Temporal relationship betweenhuman immunodeficiency virus type 1 RNA levels in serum andcellular infectious load in peripheral blood. J Infect Dis 1997;176:1383–1387.
35. Hu DJ, Buvé A, Baggs J, van der Groen G, and Dondero TJ: Whatrole does HIV-1 subtype play in transmission and pathogenesis?An epidemiological perspective. AIDS 1999;13:873–881.
36. Kanki PJ, Hamel DJ, Sankalé JL, Hsieh C-c, Thior I, Barin F,Woodcock SA, Guèye-Ndiaye A, Zhang E, Montano M, Siby T,Marlink R, NDoye I, Essex ME, and Mboup S: Human immuno-deficiency virus type 1 subtypes differ in disease progression. J In-fect Dis 1999;179:68–73.
37. Alaeus A, Lidman K, Björkman A, Giesecke J, and Albert J: Sim-ilar rate of disease progression among individuals infected withHIV-1 genetic subtypes A–D. AIDS 1999;13:901–907.
38. Neilson JR, John GC, Carr JK, Lewis P, Kreiss JK, Jackson S, Nduati RW, Mbori-Ngacha D, Panteleeff DD, Bodrug S, GiachettiC, Bott MA, Richardson BA, Bwayo J, Ndinya-Achola J, and Over-baugh J: Subtypes of human immunodeficiency virus type 1 anddisease stage among women in Nairobi, Kenya. J Virol1999;73:4393–4403.
39. Hu DJ, Vanichseni S, Mastro TD, Raktham S, Young NL, MockPA, Subbarao S, Parekh BS, Srisuwanvilai L, Sutthent R, Wasi C,Heneine W, and Choopanya K: Viral load differences in early in-fection with two HIV-1 subtypes. AIDS 2001;15:683–691.
40. Morris L, Cilliers T, Bredell H, Phoswa M, and Martin DJ: CCR5is the major coreceptor used by HIV-1 subtype C isolates from pa-tients with tuberculosis. AIDS Res Hum Retroviruses 2001;17:697–701.
41. Jurriaans S, Van Gemen B, Weverling GJ, Van Strijp D, Nara P,Coutinho R, Koot M, Schuitemaker H, and Goudsmit J: The nat-
CORECEPTOR CHANGE OF HIV-1 IN CHILDREN 351

ural history of HIV-1 infection: Virus load and virus phenotype in-dependent determinants of clinical course? Virology 1994;204:223–233.
42. Connor RI and Ho DD: Human immunodeficiency virus type 1variants with increased replicative capacity develop during theasymptomatic stage before disease progression. J Virol 1994;68:4400–4408.
43. Koot M, Keet IP, Vos AH, de Goede RE, Roos MT, Coutinho RA,Miedema F, Schellekens PT, and Tersmette M: Prognostic valueof HIV-1 syncytium-inducing phenotype for rate of CD41 cell de-pletion and progression to AIDS. Ann Intern Med 1993;118:742–743.
44. Chun T-W, Davey RT Jr, Ostrowski M, Shawn Justement J, En-
gel D, Mullins JI, and Fauci AS: Relationship between pre-exist-ing viral reservoirs and the re-emergence of plasma viremia afterdiscontinuation of highly active anti-retroviral therapy. Nat Med2000;6:757–761.
Address reprint requests to:Anneka Ehrnst
Microbiology and Tumor Biology Center, Box 280Karolinska Institutet
SE-171 77 Stockholm, Sweden
E-mail: [email protected]
CASPER ET AL.352
Related Documents











