Exp Brain Res (1997) 117:67–79 Springer-Verlag 1997 RESEARCH ARTICLE Magnus K.O. Burstedt · Benoni B. Edin Roland S. Johansson Coordination of fingertip forces during human manipulation can emerge from independent neural networks controlling each engaged digit Received: 25 October 1996 / Accepted: 3 April 1997 M.K.O. Burstedt ( ) ) · B.B. Edin · R.S. Johansson Department of Physiology, University of Umeå, S-901 87 Umeå, Sweden Tel.: +46-90-7865186, Fax: +46-90-7866683, e-mail: magnus.burstedt @physiol.umu.se Abstract We investigated the coordination of fingertip forces in subjects who lifted an object (i) using the index finger and thumb of their right hand, (ii) using their left and right index fingers, and (iii) cooperatively with anoth- er subject using the right index finger. The forces applied normal and tangential to the two parallel grip surfaces of the test object and the vertical movement of the object were recorded. The friction between the object and the digits was varied independently at each surface between blocks of trials by changing the materials covering the grip surfaces. The objects weight and surface materials were held constant across consecutive trials. The perfor- mance was remarkably similar whether the task was shared by two subjects or carried out unimanually or bi- manually by a single subject. The local friction was the main factor determining the normal:tangential force ratio employed at each digit-object interface. Irrespective of grasp configuration, the subjects adapted the force ratios to the local frictional conditions such that they maintained adequate safety margins against slips at each of the en- gaged digits during the various phases of the lifting task. Importantly, the observed force adjustments were not obligatory mechanical consequences of the task. In all three grasp configurations an incidental slip at one of the digits elicited a normal force increase at both engaged digits such that the normal:tangential force ratio was re- stored at the non-slipping digit and increased at the slip- ping digit. The initial development of the fingertip forces prior to object lift-off revealed that the subjects employed digit-specific anticipatory mechanisms using weight and frictional experiences in the previous trial. Because grasp stability was accomplished in a similar manner whether the task was carried out by one subject or cooperatively by two subjects, it was concluded that anticipatory adjust- ments of the fingertip forces can emerge from the action of anatomically independent neural networks controlling each engaged digit. In contrast, important aspects of the temporal coordination of the digits was organized by a “higher level” sensory – based control that influenced both digits. In lifts by single subjects this control was mast probably based on tactile and visual input and on communication between neural control mechanisms asso- ciated with each digit. In the two-subject grasp configura- tion this synchronization information was based on audi- tory and visual cues. Key words Bimanual motor control · Precision grip · Grip force · Frictional adjustments · Human hand Introduction How humans grasp objects and what sensorimotor mech- anisms subserve this function have been important ques- tions for several research groups during the last decade (for recent overview see Johansson 1996). Many of these studies have focused on the mechanical interaction be- tween the digits and the object while the object is lifted from its support, i.e., from the moment the subject first touches the object until it is replaced and released from the grasp (e.g., Westling and Johansson 1984; Johansson and Westling 1984a, 1988a; Edin et al. 1992; Gordon et al. 1993; Kinoshita et al. 1993). This apparently simple task is a complex sequence of parallel and sequential changes in motor output. Each phase of the lifting task is characterized by a particular goal (e.g., to establish a grasp), the completion of which is verified by specific sensory signals. These signals may be elicited by mechan- ical events associated with object contact, lift-off of the object, touch down, etc. (Westling and Johanssen 1987). An indicator of the complexity of this task is the observa- tion that the robust performance observed in adults is typ- ically not achieved before the age of 8–10 years (Forss- berg et al. 1991, 1992, 1995). Several control mechanisms involved in this task have been identified during the last decade, such as memory-driven feed-forward control and the intermittent use of somatosensory afferent infor-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Exp Brain Res (1997) 117:67±79 � Springer-Verlag 1997
R E S E A R C H A R T I C L E
Magnus K.O. Burstedt ´ Benoni B. EdinRoland S. Johansson
Coordination of fingertip forces during human manipulation can emergefrom independent neural networks controlling each engaged digit
Received: 25 October 1996 / Accepted: 3 April 1997
M.K.O. Burstedt ()) ´ B.B. Edin ´ R.S. JohanssonDepartment of Physiology, University of Umeå,S-901 87 Umeå, SwedenTel.: +46-90-7865186, Fax: +46-90-7866683,e-mail: magnus.burstedt @physiol.umu.se
Abstract We investigated the coordination of fingertipforces in subjects who lifted an object (i) using the indexfinger and thumb of their right hand, (ii) using their leftand right index fingers, and (iii) cooperatively with anoth-er subject using the right index finger. The forces appliednormal and tangential to the two parallel grip surfaces ofthe test object and the vertical movement of the objectwere recorded. The friction between the object and thedigits was varied independently at each surface betweenblocks of trials by changing the materials covering thegrip surfaces. The object�s weight and surface materialswere held constant across consecutive trials. The perfor-mance was remarkably similar whether the task wasshared by two subjects or carried out unimanually or bi-manually by a single subject. The local friction was themain factor determining the normal:tangential force ratioemployed at each digit-object interface. Irrespective ofgrasp configuration, the subjects adapted the force ratiosto the local frictional conditions such that they maintainedadequate safety margins against slips at each of the en-gaged digits during the various phases of the lifting task.Importantly, the observed force adjustments were notobligatory mechanical consequences of the task. In allthree grasp configurations an incidental slip at one ofthe digits elicited a normal force increase at both engageddigits such that the normal:tangential force ratio was re-stored at the non-slipping digit and increased at the slip-ping digit. The initial development of the fingertip forcesprior to object lift-off revealed that the subjects employeddigit-specific anticipatory mechanisms using weight andfrictional experiences in the previous trial. Because graspstability was accomplished in a similar manner whetherthe task was carried out by one subject or cooperativelyby two subjects, it was concluded that anticipatory adjust-ments of the fingertip forces can emerge from the actionof anatomically independent neural networks controlling
each engaged digit. In contrast, important aspects of thetemporal coordination of the digits was organized by aªhigher levelº sensory ± based control that influencedboth digits. In lifts by single subjects this control wasmast probably based on tactile and visual input and oncommunication between neural control mechanisms asso-ciated with each digit. In the two-subject grasp configura-tion this synchronization information was based on audi-tory and visual cues.
Key words Bimanual motor control ´ Precision grip ´Grip force ´ Frictional adjustments ´ Human hand
Introduction
How humans grasp objects and what sensorimotor mech-anisms subserve this function have been important ques-tions for several research groups during the last decade(for recent overview see Johansson 1996). Many of thesestudies have focused on the mechanical interaction be-tween the digits and the object while the object is liftedfrom its support, i.e., from the moment the subject firsttouches the object until it is replaced and released fromthe grasp (e.g., Westling and Johansson 1984; Johanssonand Westling 1984a, 1988a; Edin et al. 1992; Gordonet al. 1993; Kinoshita et al. 1993). This apparently simpletask is a complex sequence of parallel and sequentialchanges in motor output. Each phase of the lifting taskis characterized by a particular goal (e.g., to establish agrasp), the completion of which is verified by specificsensory signals. These signals may be elicited by mechan-ical events associated with object contact, lift-off of theobject, touch down, etc. (Westling and Johanssen 1987).An indicator of the complexity of this task is the observa-tion that the robust performance observed in adults is typ-ically not achieved before the age of 8±10 years (Forss-berg et al. 1991, 1992, 1995). Several control mechanismsinvolved in this task have been identified during the lastdecade, such as memory-driven feed-forward controland the intermittent use of somatosensory afferent infor-

68
mation (cf. Johansson and Cole 1994). Also, partialknowledge has become available about the sensory sig-nals that provide information about various mechanicalevents during a lifting task (Johansson and Westling1987; Westling and Johansson 1987). Furthermore, as-pects of central nervous system (CNS) mechanisms en-gaged in the control of similar tasks have been studiedin man (Müller and Dichgans 1994; Lemon et al. 1995)and in monkey (Espinoza and Smith 1990; Dugas andSmith 1992; Picard and Smith 1992a, b).
When individual subjects lift an object in a precisiongrip between the index finger and the thumb of one hand,the applied fingertip forces are partitioned in a mannerproviding grasp stability at the separate digit-object inter-faces (Edin et al. 1992). That is, subjects adapt the coor-dination of the normal and tangential forces at each digitto the local frictional condition such that they avoid slipsor sliding movements and at the same time excessive fin-gertip forces. The normal:tangential force ratio at eachdigit involved in the task appears to be controlled by dig-it-specific tactile sensory information and an anticipatoryparameter-control policy using sensorimotor memoriesrelated to the local frictional condition in previous liftswith the same object. As such, the ratio employed islargely independent of the frictional condition at the op-posite digit. On the basis of these observations we havehypothesized that the partitioning of forces among thedigits may be controlled by independent neural mecha-nisms operating in a digit-specific manner. Thus, the co-ordinated behavior across the digits would not necessarilyreflect the operation of a global ªcoordinatorº but rathercould emerge through the interactions between the objectand individually controlled digits.
In the present study we specifically tested the hypoth-esis of digit-specific control of the fingertip force vectorsin lifting tasks favorable for exploiting anticipatory con-trol mechanisms. We compared the behavior when thelifting task was performed by two subjects, each of whomcontributed one finger, with the behavior when it was car-ried out by a single subject using one hand or two hands.If the performance by two people is similar to that by asingle person we must conclude that grasp stability canbe mediated by neural controllers that are independentin terms of neural connections. A partial account of theresults in this study has been presented in a preliminaryform (Burstedt et al. 1992).
Subjects and methods
Subjects, apparatus and general procedure
Ten healthy, adult subjects (18±30 years old; 5 men and 5 women)participated after giving their informed consent. They did not knowthe specific purpose of the experiments.
Subjects sat in an ordinary chair with their upper arms parallel tothe trunk and the unsupported forearms extended anteriorly.Throughout the trials, the subjects could see the instrumented test ob-ject that was initially located on a small table in front of them. Uponinstruction the subjects grasped the test object, lifted it up to a heightof about 5 cm and held it steady in the air for 2±4 s before replacing
it. They were told to lift the object at their preferred speed but did notreceive instructions about the forces to apply. Subjects were asked towash their hands with soap and water before each experiment.
The test object was similar to the one used in previous studies(Edin et al. 1992) (Fig. 1A). The mass of the object was 665 g.The parallel and vertical grip surfaces were 30 mm apart and the sur-face area of each surface measured 40�60 mm. The center points ofthe surfaces were located 125 mm above the object�s center of grav-ity. Different frictional conditions were achieved by exchanging thegrip surfaces (see below).
The fingertip forces were measured at both grip surfaces as or-thogonal components using a multiple-element strain gauge trans-ducer system (DC, 120 Hz): the normal force was directed perpen-dicular to the grip surface and the vertical tangential force parallelto the surface. The error in normal and tangential force measure-ments was <5% over the entire grip surfaces.
Vertical movements of the test object were measured by an ultra-sonic position transducer with a transmitter located at the top of theobject and a receiver on a support above the test object (DC, 40 Hz).Contact between the object and the supporting table was detectedgalvanically. The test object carried an accelerometer to facilitatedetection of friction slips between the finger and the manipulandum.The accelerometer signal (10±600 Hz) was root-mean-square(r.m.s.) processed with rise and decay times of 3 and 6 ms, respec-tively (Johansson and Westling 1984a).
Experimental procedures
The lifting tasks were analyzed for three grasp configurations:(i) During unimanual lifts, the subjects lifted the test object with a
precision grasp between the tips of the index finger and thrumbof the right hand. The three ulnar fingers were kept flexed dur-ing the task.
(ii) During bimanual lifts by one subject, the subjects used the tipsof their right and left index fingers to lift the test object. Theremaining ulnar fingers were flexed and the thumbs were ex-tended and abducted. No contact between the hands was al-lowed.
(iii) During bimanual lifts by two subjects, the subjects faced eachother while lifting the test object cooperatively using the indexfinger of their right hands, one fingertip on each of the two gripsurfaces of the object. The remaining ulnar fingers were flexedand the thumbs were extended and abducted. The subjects werenot allowed to talk to each other during the trials.
To initiate the trials involving a single subject, the verbal instructionªliftº was given. The subject then reached forward to grip the object.The start of lifting trials involving two subjects was synchronized bya verbal count down (ªready±steady±golº) upon which both subjectsmoved the right index finger to contact the grip surface located some3 cm in the left direction. Under all grasp configurations the instruc-tion ªreplaceº was used to terminate the trial. During both bimanualgrasp configurations, the object was oriented with the grip surfacesin the subjects� sagittal plane. In the unimanual condition, the objectwas rotated counter-clockwise some 45� in the horizontal plane for amore natural grasp. Thus, in this condition the subjects could not seethe final approach of the index finger towards the object.
It is possible to adjust normal:tangential force ratios by applyingthe digits asymmetrically to the object and thereby creating an inter-nal torque that allows the load to be shifted predominantly to thelower digit. The focus in the present study was not, however, to ex-amine the mechanisms involved in grasp selection. Therefore, toavoid asymmetrical points of force attack, markers were put at thevertical midpoints of the grip surfaces and the subjects were askedto place each digit at the level of these markers. Moreover, trialsin which the normal forces were notably different at the two digitswere excluded from the analyses.
All subjects were familiarized with the task prior to the actualexperimental series. The grip surfaces were always covered by sand-paper during these trials. When two subjects lifted the test object ittended to move horizontally or tilt before lift-off during the first

69
practice trials. The experiment did not begin until at least two suc-cessful trials without appreciable tilt or horizontal movement hadbeen completed. This always occurred within five trials.
Surface materials and frictional measurements
Three different surface structures that represented low, intermediateand high surface friction were used (Fig. 4A): rayon, suede and fine-grain sandpaper (no. 320) (Westling and Johansson 1984). The co-efficient of static friction was determined for each digit-object inter-face in separate trials at the beginning of each experimental series.
Subjects held the object in air and the normal and tangential forceswere measured while frictional slips were provoked alternately atone digit at a time (Edin et al. 1992). The subjects were asked togradually decrease the normal force and simultaneously tilt the ob-ject laterally until slips occurred. Slips were detected using both theaccelerometer signal and the slip-induced reciprocal changes in thetangential forces at the two digits. The ratio between the normal andtangential force was recorded when a slip was initiated. This ratio,called the slip ratio, represented the inverse of the coefficient of stat-ic friction. Slip ratios were also determined from trials during whichslips occurred spontaneously. For each subject there was one slip ra-tio value registered for each combination of digit, surface materialand grasp configuration. This value was the mean of the slip ratiomeasurements (5£n£10) for each surface material and digit obtainedin each series of lifts. The difference between the normal:tangentialforce ratio employed and the slip ratio will be referred to as the safe-ty margin in line with previous studies on human precision grip (Jo-hansson and Westling 1984a).
Series of lift trials
With three surface materials and two grip surfaces there are ninepossible combinations of surface materials. The lifting series con-sisted of blocks of eight consecutive trials of each surface combina-tion out of which six were attended to; the first two trials in eachcombination were excluded from the analysis because initial adjust-ments to new frictional conditions were not the focus of the study(cf. Edin et al. 1992). Five subjects (2 men and 3 women) carriedout the lifting series for all three grasp configurations. In the seriescarried out bimanually by two subjects, five more subjects were re-cruited as counterparts. Thus, a total of 810 trials were recorded(9�6�5�3). Fifteen trials were excluded from the analysis becauseof sampling errors. The excluded trials were performed by differentsubjects and with different surface combinations and thus their ex-clusion would not be expected to bias the results.
Fig. 1A±C Instrumented test object, sample trial and measure-ments. A Schematic illustration of the apparatus. B Single trial inwhich the thumb contacted sandpaper and the index finger contactedthe more slippery suede. From top to bottom, superimposed nor-mal:tangential force ratios, normal forces, tangential forces, verticalposition and a signal that indicated when the object lost its contactwith the supporting table. The vertical distance between the appliedforce ratio and the slip ratio (horizontal lines) represents the safetymargin against slips. The vertical lines indicate points in time usedfor measurements: a moment of initial contact between the fingerand the object, b onset of tangential force increase, c moment oflift-off; d®e a 0.5-s period starting 0.7 s after lift-off representingthe static hold phase. C Initial part of the trial in B illustrates howa and b were identified for one of the digits (thumb). Normal andtangential forces are shown together with their first and second timedifferentials. Starting 100 ms before the normal force reached 0.2 N(i.e., 100 ms before a1), the moment was identified when the firsttime differential of the normal force reached 5 N/s (a2). Within atime window of �35 ms before and 15 ms after this, the local max-imum of the second time derivative was located (a). The onset of thetangential force increase (b) was defined analogously using the timederivatives of the tangential force record
70
Data collection and analysis
Using a custom-built data acquisition and analysis system (SC/ZOOM, Department of Physiology, Umeå University) on a DOS-op-erated 486 system, the transducer signals were sampled at 400/s with12-bit resolution and stored on a computer disk. The normal:tangen-tial force ratio was computed for each digit as a function of time.The first time derivatives of the various signals were obtained bya symmetrical numerical differentiation using a moving time win-dow of �5 samples.
Each lifting trial was divided into a series of phases (Johanssonand Westling 1984a). For each trial, these phases were initially iden-tified by means of computer algorithms and then interactively veri-fied by the experimenters by visual inspection of force records fromeach trial.
The initial contact between a digit and the object and the onsetof tangential force increase were defined at each grip surface fromthe normal and tangential forces, respectively, using a computer al-gorithm described in Fig. 1C. The definition of tangential force on-set was, however, associated with a larger uncertainty than that ofthe initial contact because the subjects might induce fluctuationsin the tangential force when they contacted the object prior to onsetof the lifting drive (cf. tangential force record in Fig. 1C). This couldhave biased the measurement towards an early tangential force on-set. The time between the moments of initial contact of the first digitto touch the object (leading digit) and the second (lagging digit), andthe time between the onset of tangential force increase at the twosurfaces were determined.
For each digit, the pre-load phase was defined as the period fromthe moment of contact to the onset of the tangential force increase(a±b in Fig. 1B, C) and the duration of the load phase as the periodfrom the onset of tangential force increase until lift-off (b±c in Fig.1B), i.e., when the galvanic contact between the object and the sup-port table was initially broken. The static hold phase was defined asa 500-ms period (d±e) starting 0.7 s after lift-off, while subjects heldthe object stationary in the air. For each digit, the static normal andtangential forces were measured as the average forces, respectively,during this period.
Statistical procedures
Numerical values of normal:tangential force ratios, normal force andtiming information were transferred to a statistical program (STA-TISTICA, Statsoft). All variables were logarithmically transformedto obtain approximately normal distributions. Population statisticsare therefore presented as geometric means ´/� standard deviationsand refer to data pooled across subjects unless stated otherwise [no-tably, log(mean)�log(SD) correspond to means ´/�SD]. Two be-tween-groups multivariate analysis of variance (MANOVA) designswere performed. The first design was a 5�3 MANOVA with sub-jects included as a random factor (5 subjects �3 grasp configura-tions). Two dependent variables were analyzed: (i) the absolute timebetween the moments of initial contact and (ii) the absolute time be-tween the onset of tangential force increase at the two grip surfaces.The second design was a 3�3�3�2 MANOVA (3 grasp configura-
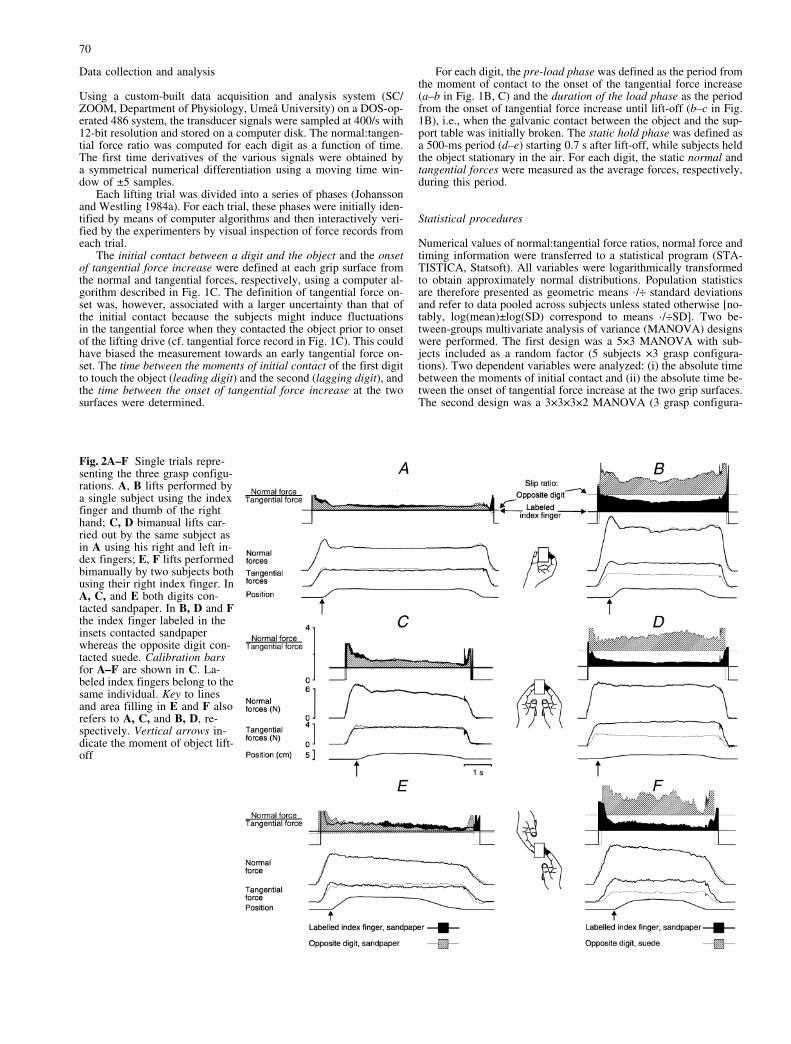
Fig. 2A±F Single trials repre-senting the three grasp configu-rations. A, B lifts performed bya single subject using the indexfinger and thumb of the righthand; C, D bimanual lifts car-ried out by the same subject asin A using his right and left in-dex fingers; E, F lifts performedbimanually by two subjects bothusing their right index finger. InA, C, and E both digits con-tacted sandpaper. In B, D and Fthe index finger labeled in theinsets contacted sandpaperwhereas the opposite digit con-tacted suede. Calibration barsfor A±F are shown in C. La-beled index fingers belong to thesame individual. Key to linesand area filling in E and F alsorefers to A, C, and B, D, re-spectively. Vertical arrows in-dicate the moment of object lift-off

71
tions�3 surface materials at the digit-object interface �3 surface ma-terials at the opposite digit-object interface �2 leading or laggingdigit). With this design six dependent variables were analyzed: (i)the duration of the pre-load phase, (ii) the duration of the load phase,(iii) the normal force applied at the onset of tangential force in-crease, and (iv) the normal force, (v) normal:tangential force ratioand (vi) safety margin during the static hold phase. In the later de-sign, data were aggregated from records involving the right indexfinger when the task was performed by a single subject and, duringthe series with two subjects, from the same subjects that participatedin the single-subject condition. Planned comparisons were used totest specific hypotheses in both designs. Before applying the MAN-OVA analyses, the assumption of homogeneity of variances wastested according to Sen and Puri (1968).
A Pearson product-moment correlation matrix was used to iden-tify correlated variables and linear regression was used to test thedependence of correlated variables. Test outcomes associated withP values <0.01 were considered ªsignificantº. In particular, all re-ported correlation coefficients were significant with P<0.01.
Results
General performance
The behavior during bimanual lifts by two subjects wasremarkably similar to that in bimanual and unimanual liftsby one subject (Fig. 2). Irrespective of grasp configura-tion, for each of the engaged digits the subjects adaptedthe ratio between the normal and tangential force to thelocal frictional conditions such that they maintained ade-quate safety margins against frictional slips. With thesame surface material at both digit-object interfaces theforces applied at the two surfaces were similar (Fig. 2A,C, E). In contrast, when the two involved digits were incontact with surfaces of different friction, the total tan-gential force was partitioned asymmetrically with thelarger tangential force applied by the digit in contact withthe less slippery surface. Although influenced by the fric-tional condition at both digits as described below, the nor-mal forces were similar (Figs. 2B, D, F, 3). As a conse-
quence, the normal:tangential ratio employed was higherat the digit in contact with the more slippery surface thanat the digit with higher friction and an adequate safetymargin to prevent slips was maintained at both digit-ob-ject interfaces.
Because the subjects employed different tangentialforces at the two digits already during the load phase,the object must have tilted after lift-off. However, the ac-tual tilt was barely noticeable due to the object�s low cen-ter of gravity. Calculation using the tangential force re-cords indicated that the maximum tilting during the holdphase was 6.1� and that in 95% of the trials the tilting didnot exceed 3.5�. This calculation was justified because thenormal forces did not contribute to a tilting torque, i.e.,they were approximately symmetrically applied (see Sub-jects and methods).
Because the object�s weight and surface textures wereheld constant across consecutive trials, the subjects wereexpected to employ anticipatory control strategies (Joh-annson and Westling 1984a, 1988a). Indeed, the tangen-tial force at the two digits diverged from the onset of forceincrease in anticipation of the frictional conditions expe-rienced in the previous trial. This occurred in bimanualand unimanual lifts by one subject (Figs. 2B, D; cf. Edinet al. 1992) but also in bimanual lifts by two subjects (Fig.2F; see also Fig. 3). Moreover, under all three grasp con-figurations the weight of the object clearly was anticipat-ed since the rate of increase in the applied fingertip forcesdeclined during the load phase prior to object lift-off (Jo-hansson and Westling 1988a).
Figure 3 illustrates the inter-trial variability in the liftingbehavior typically observed in single subjects under thethree grasp configurations. The variability in the appliedforces was expected to be higher during bimanual lifts bytwo subjects than in bimanual and unimanual lifts by onesubject. This was, however, evident neither from visual in-spection of the records (cf. Fig. 3A±C) nor from the popu-lation statistics (standard deviations in Figs. 4A and 5).
Fig. 3A±C Five superimposedconsecutive trials in each of thethree grasp configurations. Tri-als were performed unimanually(A) or bimanually (B) by thesame subject. C Trials carriedout cooperatively by two sub-jects; the subject in A and B isrepresented by the labeled indexfinger. A±C Labeled index fin-ger contacted sandpaper and theopposite digit suede. Horizontallines represent slip ratios foreach digit separately
72
Thus, the initial analysis indicated that the grasp con-figuration had little influence on the general performanceand on the applied fingertip forces.
Normal:tangential force ratios during the hold phase
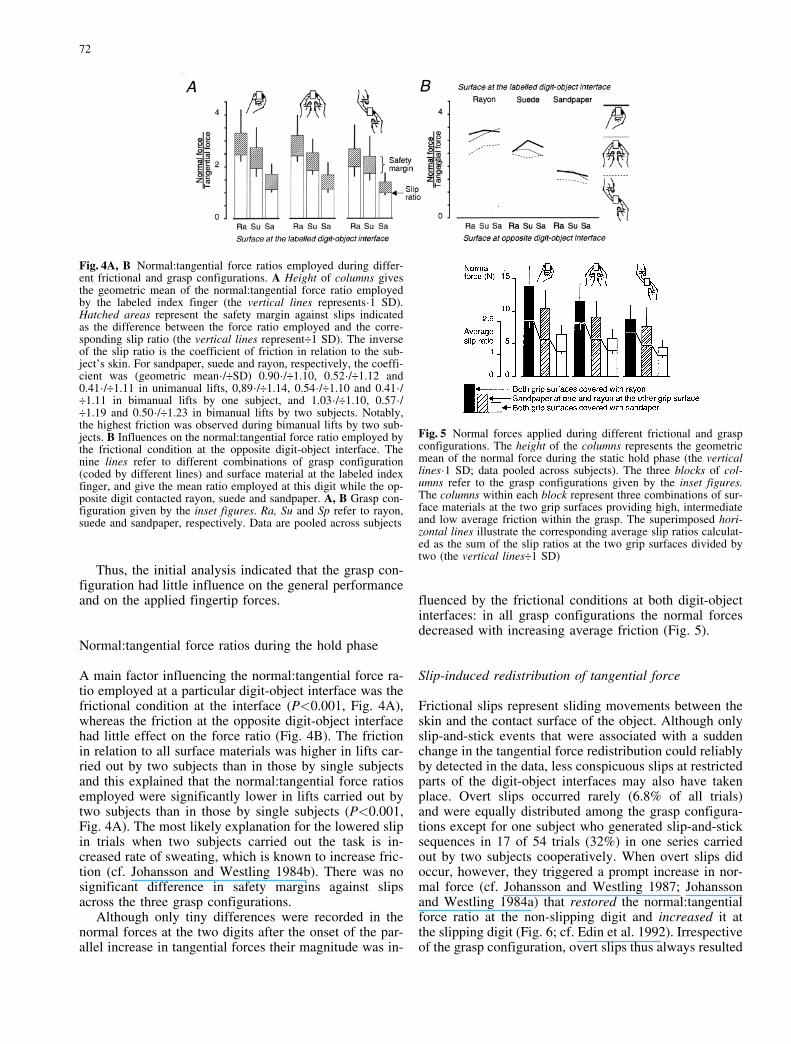
A main factor influencing the normal:tangential force ra-tio employed at a particular digit-object interface was thefrictional condition at the interface (P<0.001, Fig. 4A),whereas the friction at the opposite digit-object interfacehad little effect on the force ratio (Fig. 4B). The frictionin relation to all surface materials was higher in lifts car-ried out by two subjects than in those by single subjectsand this explained that the normal:tangential force ratiosemployed were significantly lower in lifts carried out bytwo subjects than in those by single subjects (P<0.001,Fig. 4A). The most likely explanation for the lowered slipin trials when two subjects carried out the task is in-creased rate of sweating, which is known to increase fric-tion (cf. Johansson and Westling 1984b). There was nosignificant difference in safety margins against slipsacross the three grasp configurations.
Although only tiny differences were recorded in thenormal forces at the two digits after the onset of the par-allel increase in tangential forces their magnitude was in-
fluenced by the frictional conditions at both digit-objectinterfaces: in all grasp configurations the normal forcesdecreased with increasing average friction (Fig. 5).
Slip-induced redistribution of tangential force
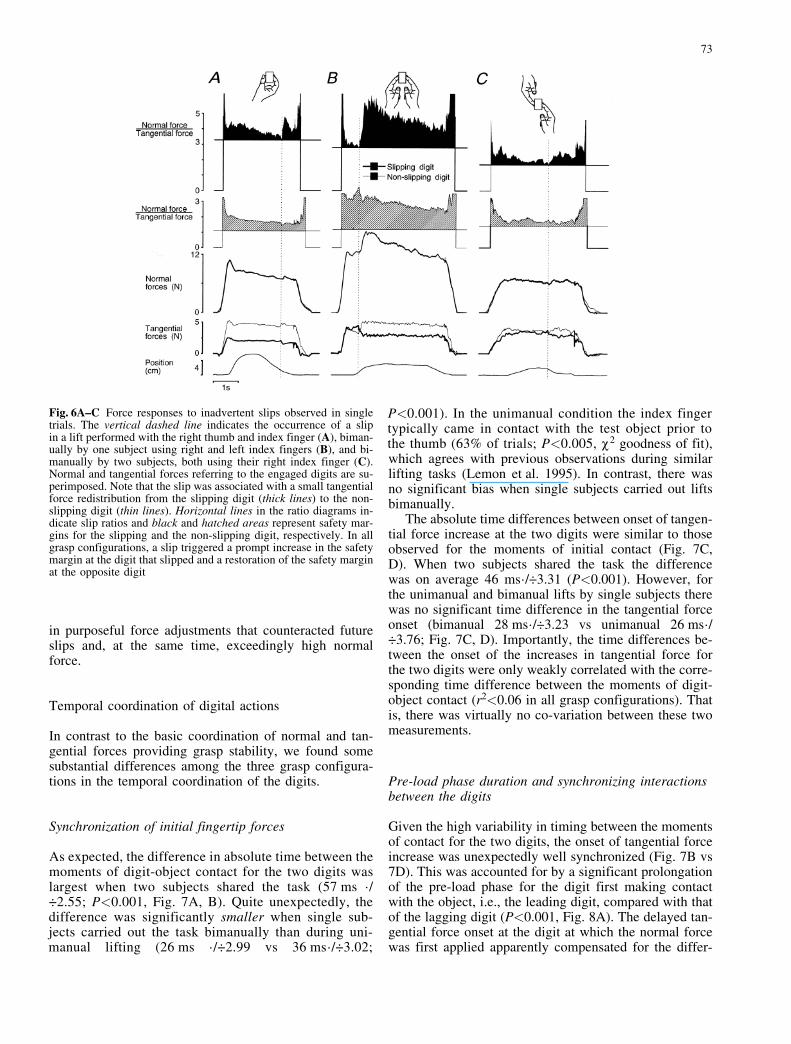
Frictional slips represent sliding movements between theskin and the contact surface of the object. Although onlyslip-and-stick events that were associated with a suddenchange in the tangential force redistribution could reliablyby detected in the data, less conspicuous slips at restrictedparts of the digit-object interfaces may also have takenplace. Overt slips occurred rarely (6.8% of all trials)and were equally distributed among the grasp configura-tions except for one subject who generated slip-and-sticksequences in 17 of 54 trials (32%) in one series carriedout by two subjects cooperatively. When overt slips didoccur, however, they triggered a prompt increase in nor-mal force (cf. Johansson and Westling 1987; Johanssonand Westling 1984a) that restored the normal:tangentialforce ratio at the non-slipping digit and increased it atthe slipping digit (Fig. 6; cf. Edin et al. 1992). Irrespectiveof the grasp configuration, overt slips thus always resulted
Fig. 4A, B Normal:tangential force ratios employed during differ-ent frictional and grasp configurations. A Height of columns givesthe geometric mean of the normal:tangential force ratio employedby the labeled index finger (the vertical lines represents´1 SD).Hatched areas represent the safety margin against slips indicatedas the difference between the force ratio employed and the corre-sponding slip ratio (the vertical lines represent�1 SD). The inverseof the slip ratio is the coefficient of friction in relation to the sub-ject�s skin. For sandpaper, suede and rayon, respectively, the coeffi-cient was (geometric mean´/�SD) 0.90´/�1.10, 0.52´/�1.12 and0.41´/�1.11 in unimanual lifts, 0,89´/�1.14, 0.54´/�1.10 and 0.41´/�1.11 in bimanual lifts by one subject, and 1.03´/�1.10, 0.57´/�1.19 and 0.50´/�1.23 in bimanual lifts by two subjects. Notably,the highest friction was observed during bimanual lifts by two sub-jects. B Influences on the normal:tangential force ratio employed bythe frictional condition at the opposite digit-object interface. Thenine lines refer to different combinations of grasp configuration(coded by different lines) and surface material at the labeled indexfinger, and give the mean ratio employed at this digit while the op-posite digit contacted rayon, suede and sandpaper. A, B Grasp con-figuration given by the inset figures. Ra, Su and Sp refer to rayon,suede and sandpaper, respectively. Data are pooled across subjects
Fig. 5 Normal forces applied during different frictional and graspconfigurations. The height of the columns represents the geometricmean of the normal force during the static hold phase (the verticallines´1 SD; data pooled across subjects). The three blocks of col-umns refer to the grasp configurations given by the inset figures.The columns within each block represent three combinations of sur-face materials at the two grip surfaces providing high, intermediateand low average friction within the grasp. The superimposed hori-zontal lines illustrate the corresponding average slip ratios calculat-ed as the sum of the slip ratios at the two grip surfaces divided bytwo (the vertical lines�1 SD)
73
Fig. 6A±C Force responses to inadvertent slips observed in singletrials. The vertical dashed line indicates the occurrence of a slipin a lift performed with the right thumb and index finger (A), biman-ually by one subject using right and left index fingers (B), and bi-manually by two subjects, both using their right index finger (C).Normal and tangential forces referring to the engaged digits are su-perimposed. Note that the slip was associated with a small tangentialforce redistribution from the slipping digit (thick lines) to the non-slipping digit (thin lines). Horizontal lines in the ratio diagrams in-dicate slip ratios and black and hatched areas represent safety mar-gins for the slipping and the non-slipping digit, respectively. In allgrasp configurations, a slip triggered a prompt increase in the safetymargin at the digit that slipped and a restoration of the safety marginat the opposite digit
in purposeful force adjustments that counteracted futureslips and, at the same time, exceedingly high normalforce.
Temporal coordination of digital actions
In contrast to the basic coordination of normal and tan-gential forces providing grasp stability, we found somesubstantial differences among the three grasp configura-tions in the temporal coordination of the digits.
Synchronization of initial fingertip forces
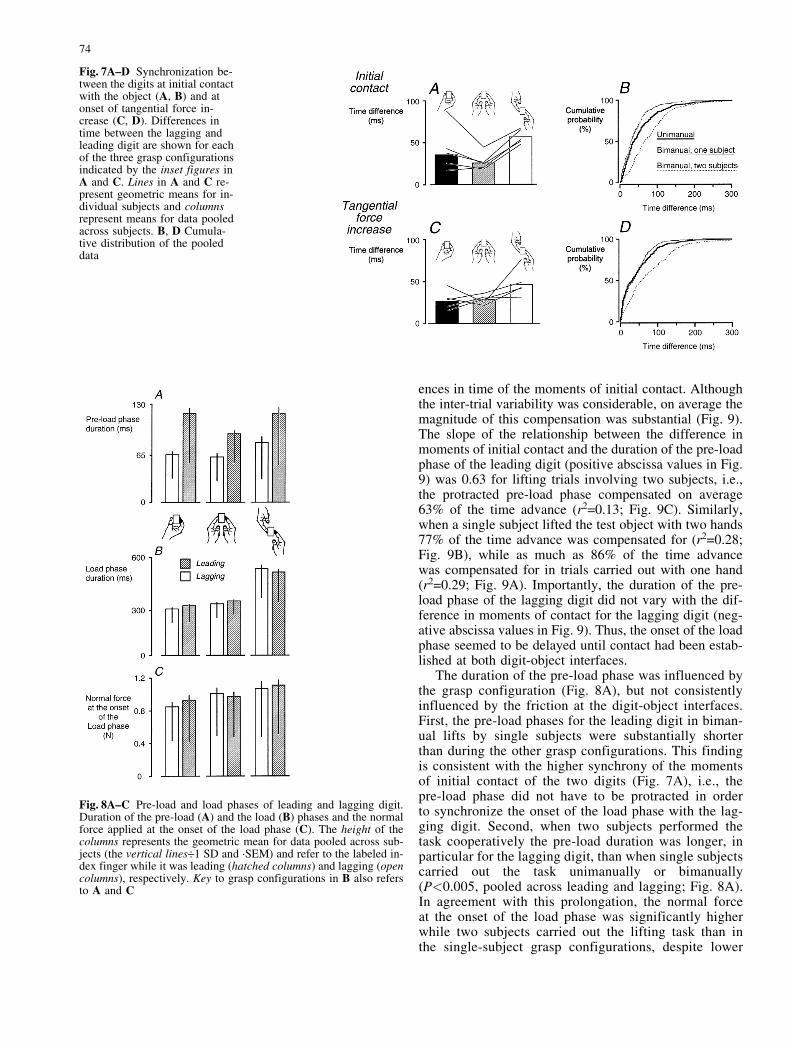
As expected, the difference in absolute time between themoments of digit-object contact for the two digits waslargest when two subjects shared the task (57 ms ´/�2.55; P<0.001, Fig. 7A, B). Quite unexpectedly, thedifference was significantly smaller when single sub-jects carried out the task bimanually than during uni-manual lifting (26 ms ´/�2.99 vs 36 ms´/�3.02;
P<0.001). In the unimanual condition the index fingertypically came in contact with the test object prior tothe thumb (63% of trials; P<0.005, c2 goodness of fit),which agrees with previous observations during similarlifting tasks (Lemon et al. 1995). In contrast, there wasno significant bias when single subjects carried out liftsbimanually.
The absolute time differences between onset of tangen-tial force increase at the two digits were similar to thoseobserved for the moments of initial contact (Fig. 7C,D). When two subjects shared the task the differencewas on average 46 ms´/�3.31 (P<0.001). However, forthe unimanual and bimanual lifts by single subjects therewas no significant time difference in the tangential forceonset (bimanual 28 ms´/�3.23 vs unimanual 26 ms´/�3.76; Fig. 7C, D). Importantly, the time differences be-tween the onset of the increases in tangential force forthe two digits were only weakly correlated with the corre-sponding time difference between the moments of digit-object contact (r2<0.06 in all grasp configurations). Thatis, there was virtually no co-variation between these twomeasurements.
Pre-load phase duration and synchronizing interactionsbetween the digits
Given the high variability in timing between the momentsof contact for the two digits, the onset of tangential forceincrease was unexpectedly well synchronized (Fig. 7B vs7D). This was accounted for by a significant prolongationof the pre-load phase for the digit first making contactwith the object, i.e., the leading digit, compared with thatof the lagging digit (P<0.001, Fig. 8A). The delayed tan-gential force onset at the digit at which the normal forcewas first applied apparently compensated for the differ-
74
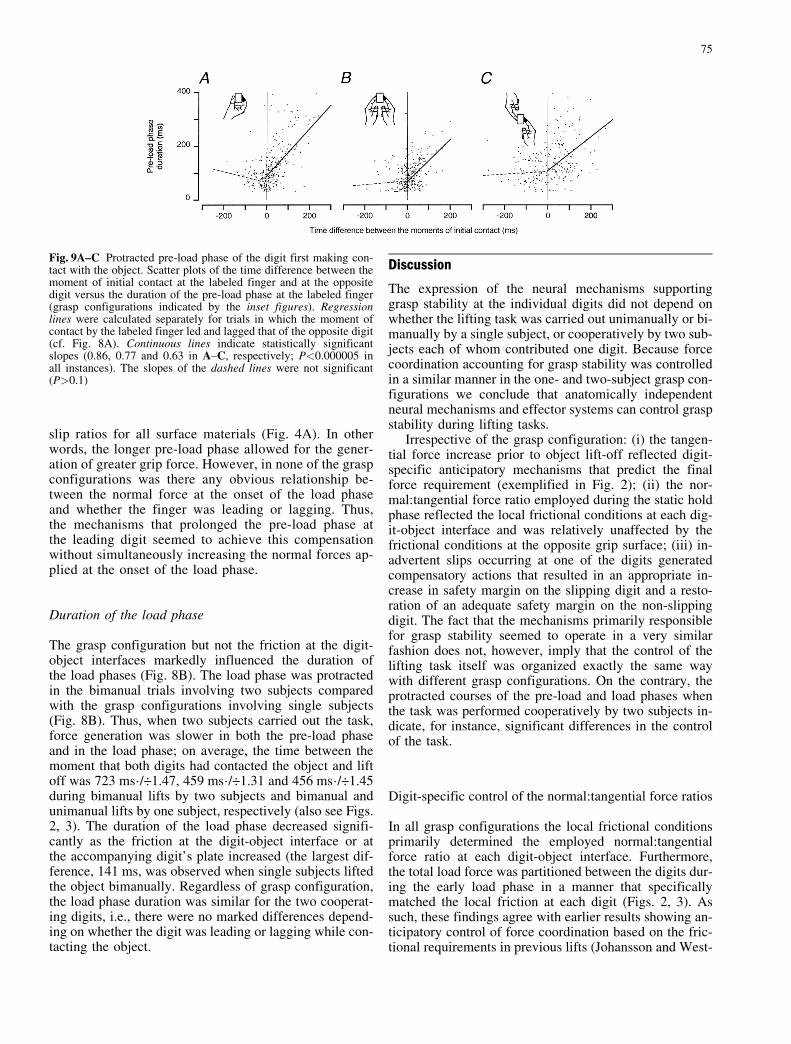
ences in time of the moments of initial contact. Althoughthe inter-trial variability was considerable, on average themagnitude of this compensation was substantial (Fig. 9).The slope of the relationship between the difference inmoments of initial contact and the duration of the pre-loadphase of the leading digit (positive abscissa values in Fig.9) was 0.63 for lifting trials involving two subjects, i.e.,the protracted pre-load phase compensated on average63% of the time advance (r2=0.13; Fig. 9C). Similarly,when a single subject lifted the test object with two hands77% of the time advance was compensated for (r2=0.28;Fig. 9B), while as much as 86% of the time advancewas compensated for in trials carried out with one hand(r2=0.29; Fig. 9A). Importantly, the duration of the pre-load phase of the lagging digit did not vary with the dif-ference in moments of contact for the lagging digit (neg-ative abscissa values in Fig. 9). Thus, the onset of the loadphase seemed to be delayed until contact had been estab-lished at both digit-object interfaces.
The duration of the pre-load phase was influenced bythe grasp configuration (Fig. 8A), but not consistentlyinfluenced by the friction at the digit-object interfaces.First, the pre-load phases for the leading digit in biman-ual lifts by single subjects were substantially shorterthan during the other grasp configurations. This findingis consistent with the higher synchrony of the momentsof initial contact of the two digits (Fig. 7A), i.e., thepre-load phase did not have to be protracted in orderto synchronize the onset of the load phase with the lag-ging digit. Second, when two subjects performed thetask cooperatively the pre-load duration was longer, inparticular for the lagging digit, than when single subjectscarried out the task unimanually or bimanually(P<0.005, pooled across leading and lagging; Fig. 8A).In agreement with this prolongation, the normal forceat the onset of the load phase was significantly higherwhile two subjects carried out the lifting task than inthe single-subject grasp configurations, despite lower
Fig. 7A±D Synchronization be-tween the digits at initial contactwith the object (A, B) and atonset of tangential force in-crease (C, D). Differences intime between the lagging andleading digit are shown for eachof the three grasp configurationsindicated by the inset figures inA and C. Lines in A and C re-present geometric means for in-dividual subjects and columnsrepresent means for data pooledacross subjects. B, D Cumula-tive distribution of the pooleddata
Fig. 8A±C Pre-load and load phases of leading and lagging digit.Duration of the pre-load (A) and the load (B) phases and the normalforce applied at the onset of the load phase (C). The height of thecolumns represents the geometric mean for data pooled across sub-jects (the vertical lines�1 SD and ´SEM) and refer to the labeled in-dex finger while it was leading (hatched columns) and lagging (opencolumns), respectively. Key to grasp configurations in B also refersto A and C
75
slip ratios for all surface materials (Fig. 4A). In otherwords, the longer pre-load phase allowed for the gener-ation of greater grip force. However, in none of the graspconfigurations was there any obvious relationship be-tween the normal force at the onset of the load phaseand whether the finger was leading or lagging. Thus,the mechanisms that prolonged the pre-load phase atthe leading digit seemed to achieve this compensationwithout simultaneously increasing the normal forces ap-plied at the onset of the load phase.
Duration of the load phase
The grasp configuration but not the friction at the digit-object interfaces markedly influenced the duration ofthe load phases (Fig. 8B). The load phase was protractedin the bimanual trials involving two subjects comparedwith the grasp configurations involving single subjects(Fig. 8B). Thus, when two subjects carried out the task,force generation was slower in both the pre-load phaseand in the load phase; on average, the time between themoment that both digits had contacted the object and liftoff was 723 ms´/�1.47, 459 ms´/�1.31 and 456 ms´/�1.45during bimanual lifts by two subjects and bimanual andunimanual lifts by one subject, respectively (also see Figs.2, 3). The duration of the load phase decreased signifi-cantly as the friction at the digit-object interface or atthe accompanying digit�s plate increased (the largest dif-ference, 141 ms, was observed when single subjects liftedthe object bimanually. Regardless of grasp configuration,the load phase duration was similar for the two cooperat-ing digits, i.e., there were no marked differences depend-ing on whether the digit was leading or lagging while con-tacting the object.
Discussion
The expression of the neural mechanisms supportinggrasp stability at the individual digits did not depend onwhether the lifting task was carried out unimanually or bi-manually by a single subject, or cooperatively by two sub-jects each of whom contributed one digit. Because forcecoordination accounting for grasp stability was controlledin a similar manner in the one- and two-subject grasp con-figurations we conclude that anatomically independentneural mechanisms and effector systems can control graspstability during lifting tasks.
Irrespective of the grasp configuration: (i) the tangen-tial force increase prior to object lift-off reflected digit-specific anticipatory mechanisms that predict the finalforce requirement (exemplified in Fig. 2); (ii) the nor-mal:tangential force ratio employed during the static holdphase reflected the local frictional conditions at each dig-it-object interface and was relatively unaffected by thefrictional conditions at the opposite grip surface; (iii) in-advertent slips occurring at one of the digits generatedcompensatory actions that resulted in an appropriate in-crease in safety margin on the slipping digit and a resto-ration of an adequate safety margin on the non-slippingdigit. The fact that the mechanisms primarily responsiblefor grasp stability seemed to operate in a very similarfashion does not, however, imply that the control of thelifting task itself was organized exactly the same waywith different grasp configurations. On the contrary, theprotracted courses of the pre-load and load phases whenthe task was performed cooperatively by two subjects in-dicate, for instance, significant differences in the controlof the task.
Digit-specific control of the normal:tangential force ratios
In all grasp configurations the local frictional conditionsprimarily determined the employed normal:tangentialforce ratio at each digit-object interface. Furthermore,the total load force was partitioned between the digits dur-ing the early load phase in a manner that specificallymatched the local friction at each digit (Figs. 2, 3). Assuch, these findings agree with earlier results showing an-ticipatory control of force coordination based on the fric-tional requirements in previous lifts (Johansson and West-
Fig. 9A±C Protracted pre-load phase of the digit first making con-tact with the object. Scatter plots of the time difference between themoment of initial contact at the labeled finger and at the oppositedigit versus the duration of the pre-load phase at the labeled finger(grasp configurations indicated by the inset figures). Regressionlines were calculated separately for trials in which the moment ofcontact by the labeled finger led and lagged that of the opposite digit(cf. Fig. 8A). Continuous lines indicate statistically significantslopes (0.86, 0.77 and 0.63 in A±C, respectively; P<0.000005 inall instances). The slopes of the dashed lines were not significant(P>0.1)

76
ling 1984a) and that the underlying mechanisms can beexpressed on a digit-specific basis (Edin et al. 1992).The rate of lifting was dependent on the friction, but thisdoes not explain the different rates of tangential force in-crease at the digits when they contacted different materi-als. The rate of increase in the tangential forces decreasedbefore the moment of lift-off and this indicates that theload development and the partitioning of the load wascontrolled by anticipatory mechanisms. Thus, subjects ac-tively coordinated the development of the fingertip forcesat the digits to obtain force ratios adjusted to the localfrictional condition. Moreover, the mechanisms that pro-vide this anticipatory control can operate through inde-pendent neural networks separately controlling the twodigits because they were expressed also when two sub-jects cooperatively performed the lifting task.
This study did not address possible differences be-tween the three grasp configurations regarding the mech-anisms responsible for the initial adjustments to the newfrictional condition, i.e., how the normal:tangential forceratios were in fact updated after a change in surface com-bination (but for a unimanual configuration see Edin et al.1992). We cannot, for instance, exclude the possibilitythat passive mechanisms such as frictional creep or slipscontributed to a tangential force distribution accordingto the local frictional conditions in the very first trial(s)with a new surface combination. If so, subjects may havelearned the normal:tangential force ratios that were ade-quate for grasp stability and applied those in the subse-quent trials. Notably, during all grasp configurations sub-jects occasionally failed to partition the load according tothe frictional condition at the digit-object interfaces, i.e.,the coordination of the fingertip forces was not a mereconsequence of the mechanics of the task. Moreover, inall grasp configurations we observed purposeful adjust-ments of the normal:tangential force ratios at both digitsin response to inadvertent frictional slips. Such slips,which typically occur at one digit only, may appear atany time during object manipulation but are most easily,observed when they occur during the static hold phase(Johansson and Westling 1984a): the tangential force sud-denly drops at the slipping digit and simultaneously in-creases at the opposite digit (cf. Fig. 6). Slip-induced mo-tor responses during the hold phase appear after a shortlatency (ca. 70 ms) and consist primarily of an increasein normal force that raises the safety margin against slipsat the digit that slipped and restores it at the non-slippingdigit. That a similar increase in the normal force was trig-gered also when two subjects shared the task indicatesthat the slip-induced responses can be mediated by sepa-rate neural networks. It must be concluded that the slip-evoked afferent activity at the slipping digit is sufficientto trigger digit-specific adjustments of the normal:tangen-tial force ratios. The increased normal force at the slip-ping digit per se may cause a passive normal force in-crease at the other digit. In addition, it seems likely thatthe slip-induced tangential force increase at the non-slip-ping digit elicited a normal force increase also at this digitand thereby contributed by restoring its normal:tangential
force ratio. Indeed, a slip-triggered increase in normalforce appears after a latency that is similar to increasesin normal force in response to a small, transiently im-posed increase in tangential force while holding an object(cf. Johansson and Westling 1987; Johansson et al. 1992;Edin et al. 1993).
Both fast adapting mechanoreceptors (FAI and FAII)and dynamically sensitive slowly adapting (SAI) afferentsare activated at the slipping digit (Johansson and Westling1987; see also Srinivasan et al. 1990). Both FAI and SAIafferents are likely to be activated at the non-slipping dig-it when the safety margin falls precipitously as a result ofthe sudden increase in tangential force (cf. ªlocalized slipresponsesº in Johansson and Westling 1987). Further-more, small increases in the tangential force have beenshown to elicit responses in FAI, SAI and SAII afferents(Westling and Johansson 1987; Macefield et al. 1996). Insum, there are known afferent mechanisms that might ac-count for the concurrent adjustments of the fingertip forc-es observed at the digits following inadvertent slips.
Temporal coordination
Even if independent neural mechanisms can achieve graspstability during a lifting task, various aspects of inter-digitcoordination necessarily require the operation of ªhigher-levelº controls systems that influence both digits. Onesuch aspect is the synchronizing of digital actions in thetime domain. It is crucial, of course, that the sequentialphases of the tasks develop in parallel at the two digit-ob-ject interfaces. In particular, an object may fall over ormove sideways before lift-off if the fingertip forces arenot applied more or less simultaneously. Indeed, this hap-pened when two subjects lifted the test object during thefirst practice trials. However, even after practice the syn-chronization of initial contact observed when two subjectsshared the task was poor in comparison with when it wasperformed by a single subject. Apparently, the improvedsynchronization in the latter case was accounted for bycommunication between neural control mechanisms asso-ciated with each digit. In the two-subject grasp configura-tion synchronization information must have been con-fined to auditory and visual cues. Visual informationcan be used not only to guide reaching movements(Jeannerod 1994), but also to extrapolate end-points oftrajectories (Jakobson and Goodale 1991, Servos andGoodale 1994), and thereby support spatiotemporal coor-dination.
In single subjects bimanual lifting was better synchro-nized in terms of the initial digit-object contacts than uni-manual lifting. In contrast to the unimanual task, in thebimanual task both grip surfaces were visible to the sub-ject; in the unimanual task only the thumb surface wasvisible. The fact that the final approach of the index fin-ger towards the object could not be under direct visualguidance may have resulted in greater temporal scatter.Evidence from monkeys and humans, however, seemsto indicate that visual information may not be crucial

77
for synchronizing bimanual actions (Johansson andWestling 1988b; Wiesendanger et al. 1992, 1994; Ka-zennikov et al. 1994).
The time between the moments of initial contact withthe object by the two digits varied from one trial to thenext and so did the duration of the pre-load phases ofthe separate digits. Given the near independence of thesevariables one would expect that the difference in onset ofthe tangential forces at the digits involved would varymore than that of the initial digit-object contacts. Thiswas, however, not the case. Instead, the digit that firstmade contact with the object showed a protracted pre-load phase which to a significant degree compensatedfor the lag at the opposite digit-object interface. For uni-manual lifting trials this synchronization could be ex-plained by the conjoint activation of the arm muscles thatare primarily responsible for the lifting action, but thisprovides a poor explanation for synchronization in liftsperformed bimanually. Thus there are mechanisms thatactively tend to synchronize the application of tangentialforces at different digits involved in a lifting task. In liftsperformed by single subjects this synchronization couldhave been implemented by neural connections betweenrelevant networks. These mechanisms may not, however,exclusively depend on transfer of information betweenprocesses that reside within a single nervous system be-cause a significant compensation was observed also whenthe test object was lifted cooperatively by two subjects.Again, visual information may have been used. By visu-ally assessing the approach of both hands involved inthe task subjects may have compensated for temporal dif-ferences in lifting trials. Indeed, in collateral experimentswith pairs of blindfolded subjects both the timing of theinitial digit-object contacts and the onsets of the tangen-tial force increases were considerably less synchronized(M.K.O. Burstedt, unpublished observations). Thus, themechanisms providing synchronization of load force ap-plication may use somatosensory signals related to objectcontact, and visual and auditory information. However,the compensation was clearly weaker in lifts by two sub-jects than in lifts by a single subject, suggesting that co-ordination based on touch information processed withinthe subject�s nervous system played a primary role duringunimanual as well as bimanual lifts.
Independent control versus global coordination
The present results indicate that the coordinated applica-tion of fingertip forces supporting grasp stability canemerge from structurally independent neural mechanisms.It appears that the primary function of the proposed digit-specific mechanisms is to secure appropriate normal:tan-gential force ratio with respect to the prevailing frictionalconditions, i.e., both exceedingly high fingertip forces andslips are avoided at the separate digit-object interfaces.Although this hypothesis accounts only for some aspectsof the control of the lifting task, it has several attractivefeatures as discussed previously (Edin et al. 1992). In par-
ticular, important aspects of the distribution of forcesamong digits in manipulative tasks would not require neu-ral mechanisms that explicitly coordinate the engageddigits but emerge from digit-specific controllers duringobject interaction. A good example is the grip adjustmentsthat arose after a frictional slip. In all three grasp config-urations the normal force increased not only at the slip-ping digit but also at the opposite digit. Also other aspectsof inter-digital coordination may have emerged by digit-specific controllers in phases of lift trials while substantialforce were applied to the object, allowing a good couplingof mechanical events between the digits through the ob-ject. While the object was in air, for instance, actionson one side of the object would have easily influencedthe opposite side because the object was supported onlyby the two digits. Moreover, the slow position changesthat characterized these phases would allow ample timefor sensory-based coordination including coordinationbased on visual cues. Indeed, although the replacementphase was carried out more slowly in trials involvingtwo subjects (Figs. 2, 3), it was always successfully per-formed when initiated by a simple ªreplaceº command.Finally, the mechanical transient related to the table con-tact is effectively encoded by cutaneous mechanorecep-tors (Johansson and Westling 1987, 1988b) and, as such,may have triggered the release phase in all three graspconfigurations.
The proposed scheme of digit-specific control of theforce ratio for grasp stability is particularly attractivewhen one considers that grasp stability applies to a varietyof grips, e.g., one- and two-handed grips as shown in thepresent results, inverted grips and multi-digit grips (Fla-nagan and Tresilian 1994; Kinoshita et al. 1995). Howev-er, this low-level control must be subordinated to a higherlevel of control related to other aspects of manipulativetasks. As noted above, this pertains to the temporal syn-chronization of digital actions before substantial manipu-lative forces are applied to the object. In addition, a higherlevel of control is required for selecting grasp configura-tions and for coping with various constraints imposed bythe physical properties of the object such as its weight andsize (Johansson and Westling 1988a; Gordon et al. 1991,1993) and by the task (Johansson and Westling 1984a,1988b; Flanagan and Wing 1993; Kinoshita et al. 1993,1996; Flanagan and Tresilian 1994).
An organization similar to the one suggested in thisstudy for the control of the fingertip forces for grasp sta-bility may account for motor behaviors such as lip move-ments during speech (Gracco and Abbs 1985). Mechani-cal depression of the lower lip during lip closure just priorthe generation of a ªpº sound results in phase-dependentreflex responses in the orbicularis oris muscle. During theend of the closing phase, the largest responses occur in theupper lip and this has been interpreted as reflecting ªnon-autogenicº reflexes originating in mechanoreceptors ofthe lower lip (Gracco and Abbs 1985). It is, however, pos-sible to interpret these findings as reflecting independentand parallel control of each lip operating until mechano-receptors in the vermilion signify goal completion, i.e.,

78
lip closure. Hence, mechanical depression of the lower lipwould delay the adequate mechanical stimulation of theupper lip resulting in a more vigorous motor response inthe upper lip (Gracco and Abbs 1985).
It is thus possible that various sensorimotor behaviorsmay be generated by control elements operating in paral-lel and largely independently. Indeed, functional posturaland locomotive behaviors can be generated by uncoupledneural networks (Kato 1994). Cats with one half of theirlumbar cord isolated from both supralumbar and contra-lateral influences recover to stand and walk. The symp-toms that dominate are related to timing of leg move-ments and ªtransferº of placing information from oneleg to another, i.e., parameters of behavioral relevancethat are not locally available. Similarly, a high degreeof independence regarding the control of the legs has beenobserved in experiments in which cats and humans walkon a treadmill with split belts (Forssberg et al. 1980; Dietzet al. 1994). It is quite clear that the ipsilateral leg is large-ly controlled by its own sensorimotor apparatus. Accord-ingly, interlimb coordination in locomotion is highly flex-ible and may be achieved in a variety of ways constrainedby the overall demands of ªhigher-levelº equilibrium con-trol.
Acknowledgements This study was supported by the SwedishMedical Research Council (project 08667), Department of NavalResearch, Arlington, Va., USA (grant no. N00014-92-J-1919), theGöran Gustafssons Foundation for Research in Natural Sciencesand Medicine, and Umeå University.
References
Burstedt KOM, Westling G, Johansson RS, Edin BB (1992) Co-or-dinated manipulative behaviour can emerge by independent neu-ral systems. ENA-satellit symposium on neural control of eye,head, and limb movement, Ohlstadt/Munich, Germany, Sept10±13
Dietz V, Zijlstra W, Duysens J (1994) Human neuronal interlimb co-ordination during split-belt locomotion. Exp Brain Res101:513±520
Dugas C, Smith AM (1992) Responses of cerebellar Purkinje cells toslip of a hand-held object. J Neurophysiol 67:483±495
Edin BB, Westling G, Johansson RS (1992) Independent control ofhuman finger-tip forces at individual digits during precision lift-ing. J Physiol (Lond) 450:547±564
Edin BB, Howe R, Westling G, Cutkosky M (1993) A physiologicalmethod for relaying frictional information to a human teleoper-ator. IEEE Trans Syst Man Cybern 23:427±432
Espinoza E, Smith AM (1990) Purkinje cell simple spike activityduring grasping and lifting objects of different textures andweights. J Neurophysiol 64:698±714
Flanagan JR, Tresilian JR (1994) Grip load force coupling: a generalcontrol strategy for transporting objects. J Exp Psychol HumPercept Perform 20:944±957
Flanagan JR, Wing AM (1993) Modulation of grip force with loadforce during point-to-point arm movements. Exp Brain Res95:131±143
Forssberg H, Grillner S, Halbertsma J, Rossignol S (1980) The loco-motion of the low spinal cat. II. Interlimb coordination. ActaPhysiol Scand 108:283±295
Forssberg H, Eliasson AC, Kinoshita H, Johansson RS, Westling G(1991) Development of human precision grip. 1. Basic coordina-tion of force. Exp Brain Res 85:451±457
Forssberg H, Kinoshita H, Eliasson AC, Johansson RS, Westling G,Gordon AM (1992) Development of human precision grip. II.Anticipatory control of isometric forces targeted for object�sweight. Exp Brain Res 90:393±398
Forssberg H, Eliasson AC, Kinoshita H, Westling G, Johansson RS(1995) Development of human precision grip. IV. Tactile adap-tation of isometric finger forces to the frictional condition. ExpBrain Res 104:323±330
Gordon AM, Forssberg H, Johansson RS, Westling G (1991) Inte-gration of sensory information during the programming of pre-cision grip: comments on the contributions of size cues. ExpBrain Res 85:226±229
Gordon AM, Westling G, Cole KJ, Johansson RS (1993) Memory rep-resentations underlying motor commands used during manipula-tion of common and novel objects. J Neurophysiol 69:1789±1796
Gracco VL, Abbs JH (1985) Dynamic control of the perioral systemduring speech: kinematic analyses of autogenic and nonautogen-ic sensorimotor processes. J Neurophysiol 54:418±432
Jakobson LS, Goodale MA (1991) Factors affecting higher-ordermovement planning: a kinematic analysis of human prehension.Exp Brain Res 86:199±208
Jeannerod M (1994) Object orientated action. In: Bennett KMB,Castiello U (eds) Insights in the reach to grasp movement. Else-vier, North-Holland, Amsterdam, pp 3±15
Johansson RS (1996) Sensory control of dextrous manipulation inhumans. In: Wing AM, Haggard P, Flanagan JR (eds) Handand brain: the neurophysiology and psychology of hand move-ments. Academic Press, San Diego, pp 381±414
Johansson RS, Cole KJ (1994) Grasp stability during manipulativeactions. Can J Physiol Pharmacol 72:511±524
Johansson RS, Westling G (1984a) Roles of glabrous skin receptorsand sensorimotor memory in automatic control of precision gripwhen lifting rougher or more slippery objects. Exp Brain Res56:550±564
Johansson RS, Westling G (1984b) Influences of cutaneous sensoryinput on the motor coordination during precision manipulation.In: von Euler C, FranzØn O, Lindblom U, Ottoson D (eds) So-matosensory mechanisms. Macmillan, London, pp 249±260
Johansson RS, Westling G (1987) Signals in tactile afferents fromthe fingers eliciting adaptive motor responses during precisiongrip. Exp Brain Res 66:141±154
Johansson RS, Westling G (1988a) Coordinated isometric musclecommands adequately and erroneously programmed for theweight during lifting task with precision grip. Exp Brain Res270:1±13
Johansson RS, Westling G (1988b) Programmed and triggered ac-tions to rapid load changes during precision grip. Exp BrainRes 271:1±15
Johansson RS, Häger C, Riso R (1992) Somatosensory control ofprecision grip during unpredictable pulling loads. II. Changesin load force rate. Exp Brain Res 89:192±203
Kato M (1994) Interlimb coordination during locomotor activities.Spinal-intact cats and chronic cats with horizontal and longitudi-nal separation of the spinal cord. In: Swinnen S, Heuer H, Mass-ion J, Casaer P (eds) Interlimb coordination: neural, dynamic, andcognitive constraints. Academic Press, San Diego, pp 75±96
Kazennikov O, Wicki U, Corboz M, Hyland B, Palmeri A, RouillerEM, Wiesendanger M (1994) Temporal structure of bimanualgoal-directed movement sequence in monkeys. Eur J Neurosci6:203±210
Kinoshita H, Ikuta K, Kawai S, Udo M (1993) Effects of liftingspeed and height on the regulation of forces during lifting tasksusing a precision grip. J Hum Mov Stud 25:151±175
Kinoshita H, Kawai S, Ikuta K (1995) Contributions and coordina-tion of individual fingers in multiple finger prehension. Ergo-nomics 38:1212±1230
Kinoshita H, Kawai S, Ikuta K, Teraoka T (1996) Individual fingerforces acting on a grasped object during shaking actions. Ergo-nomics 39:243±256
Lemon RN, Johansson RS, Westling G (1995) Corticospinal controlduring reach, grasp and precision lift in man. J Neurosci15:6145±6156

79
Macefield VG, Häger-Ross C, Johansson RS (1996) Control of gripforce during restraint of an object held between finger andthumb: responses of cutaneous afferents from the digits. ExpBrain Res 108:155±171
Müller F, Dichgans J (1994) Dyscoordination of pinch and lift forcesduring grasp in patients with cerebellar lesions. Exp Brain Res101:485±492
Picard N, Smith A (1992a) Primary motor cortical responses topertubations of prehension in monkey. J Neurophysiol68:1882±1894
Picard N, Smith A (1992b) Primary motor cortical activity related tothe weight and texture of grasped objects in monkey. J Neuro-physiol 68:1867±1881
Sen PK, Puri ML (1968) On a class of multivariate multisample rankorder tests. II. Test for homogeneity of dispersion matrices.Sankhya 30:1±22
Servos P, Goodale MA (1994) Binocular vision and the online con-trol of human prehension. Exp Brain Res 98:119±127
Srinivasan MA, Whitehouse JM, LaMotte RH (1990) Tactile detec-tion of slip: surface microgeometry and peripheral neural codes.J Neurophysiol 63:1323±1332
Westling G, Johansson RS (1984) Factors influencing the force con-trol during precision grip. Exp Brain Res 53:277±284
Westling G, Johansson RS (1987) Responses in glabrous skin mech-anoreceptors during precision grip in humans. Exp Brain Res66:128±140
Wiesendanger M, Corboz M, Hyland B, Palmeri A, Maier V, Rouil-ler E (1992) Bimanual synergies in primates. Exp Brain Res22:45±64
Wiesendanger M, Kaluzny P, Kazennikov O, Palmeri A, Perrig S(1994) Temporal coordination in bimanual actions. Can J Phy-siol Pharmacol 72:591±594
Related Documents










