Pergarnon Int. 1. Insect Morphol. & Embryol., Vol. 25, No. l/2, pp. 93-114, 1996 Copyright 0 1996 Elsevier Science Ltd Printed in Great Britain. All rights reserved 0020-7322!% $15.00 + 0.00 0020.7322(95)OtKI22-4 CONTRIBUTIONS OF INSECT SYSTEMS TO THE CHROMOSOME THEORY OF INHERITANCE AND TO OUR UNDERSTANDING OF CHROMOSOME BEHAVIOR ON THE SPINDLE Jeffrey G. Ault *Laboratory of Structural Pathology, Wadsworth Center for Laboratories and Research, Albany, NY 12201-0509, U.S.A. and Department of Biomedical Sciences, State University of New York at Albany, Albany, NY 12222, U.S.A. (Accepted 20 November 1995) Abstract-Insect chromosomes were among the first chromosomes to be investigated. Since then, insects have had an important place in cytogenetics. Insect studies were instrumental in proving that genes are on chromosomes and that spindle fibers exist in living cells and are not fixation artifacts. Chromosome micromanipulation studies of insect spermatocytes have provided insight into the physical properties of spindle fibers, the initial orientation (attachment) of a bivalent to the spindle, the reorientation of mal-oriented bivalents, and the cause of orientation stability. Insect spermatocytes have been important material for observing the structure of the meiotic spindle, and for determining the location and force of the chromosome motor during anaphase. In this review, I highlight important contributions of insect systems to the Chromosome Theory of Inheritance and to our understanding of chromosome behavior on the spindle. Copyright 0 1996 Elsevier Science Ltd. Index descriptors (in addition to those in title): Cytogenetics, mitosis, meiosis, spindle fiber, Orthoptera, Diptera, Hemiptera. INTRODUCTION Mitosis and meiosis are essential developmental processes for sexually reproducing eukaryotes. Usually considered variants on the same theme (Wilson, 1928; Schrader, 1953; Nicklas, 1971), both allow the precise handling, without the necessity of a high degree of redundancy, of large amounts of genetic material during cell division (Mazia, 1961). The distribution of chromosomes, the carriers of genetic material, to daughter cells is facilitated by the spindle, a fusiform body that appears prior to, and disassembles after, cell division. Chromosomes (or bivalents, in the case of meiosis I) begin attaching to the spindle shortly after nuclear-envelope breakdown and, through trial and error, form essential bipolar orientations (i.e. attachments to each of the spindle poles; see Nicklas and Ward, 1994). Mistakes can occur during attachment (i.e. mal-orientations), but are soon corrected by the process of reorientation (Hughes-Schrader, 1943a; see also Bauer ef al., 1961). Bipolarly oriented chromosomes (bivalents) move eventually to the equatorial plane of the spindle. Sometime after all of the chromosomes (bivalents) reach the equatorial plane, the sister components of the replicated chromosome *Correspondence address. 93

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pergarnon Int. 1. Insect Morphol. & Embryol., Vol. 25, No. l/2, pp. 93-114, 1996 Copyright 0 1996 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0020-7322!% $15.00 + 0.00
0020.7322(95)OtKI22-4
CONTRIBUTIONS OF INSECT SYSTEMS TO THE CHROMOSOME THEORY OF INHERITANCE AND TO OUR UNDERSTANDING OF CHROMOSOME BEHAVIOR ON THE
SPINDLE
Jeffrey G. Ault
*Laboratory of Structural Pathology, Wadsworth Center for Laboratories and Research, Albany, NY 12201-0509, U.S.A. and Department of Biomedical Sciences, State University of New York at Albany,
Albany, NY 12222, U.S.A.
(Accepted 20 November 1995)
Abstract-Insect chromosomes were among the first chromosomes to be investigated. Since then, insects have had an important place in cytogenetics. Insect studies were instrumental in proving that genes are on chromosomes and that spindle fibers exist in living cells and are not fixation artifacts. Chromosome micromanipulation studies of insect spermatocytes have provided insight into the physical properties of spindle fibers, the initial orientation (attachment) of a bivalent to the spindle, the reorientation of mal-oriented bivalents, and the cause of orientation stability. Insect spermatocytes have been important material for observing the structure of the meiotic spindle, and for determining the location and force of the chromosome motor during anaphase. In this review, I highlight important contributions of insect systems to the Chromosome Theory of Inheritance and to our understanding of chromosome behavior on the spindle. Copyright 0 1996 Elsevier Science Ltd.
Index descriptors (in addition to those in title): Cytogenetics, mitosis, meiosis, spindle fiber, Orthoptera, Diptera, Hemiptera.
INTRODUCTION
Mitosis and meiosis are essential developmental processes for sexually reproducing eukaryotes. Usually considered variants on the same theme (Wilson, 1928; Schrader, 1953; Nicklas, 1971), both allow the precise handling, without the necessity of a high degree of redundancy, of large amounts of genetic material during cell division (Mazia, 1961). The distribution of chromosomes, the carriers of genetic material, to daughter cells is facilitated by the spindle, a fusiform body that appears prior to, and disassembles after, cell division. Chromosomes (or bivalents, in the case of meiosis I) begin attaching to the spindle shortly after nuclear-envelope breakdown and, through trial and error, form essential bipolar orientations (i.e. attachments to each of the spindle poles; see Nicklas and Ward, 1994). Mistakes can occur during attachment (i.e. mal-orientations), but are soon corrected by the process of reorientation (Hughes-Schrader, 1943a; see also Bauer ef al., 1961). Bipolarly oriented chromosomes (bivalents) move eventually to the equatorial plane of the spindle. Sometime after all of the chromosomes (bivalents) reach the equatorial plane, the sister components of the replicated chromosome
*Correspondence address.
93

94 J. G. Ault
(homologous chromosomes of the bivalent) disjoin and move to opposite poles. Once the chromosomes reach the pole, they decondense and a nuclear envelope is formed around them. This orchestrated movement of chromosomes on the spindle has fascinated scientists for over 100 yr.
The use of insects to investigate chromosomes and their behavior on the spindle has a long and distinguished history. We have learned more about chromosome behavior on the spindle during meiosis from studying insects than from any other class in the animal kingdom. This is due in part to: (1) their extreme suitability for cytological observations, (2) their easy availability through collecting and rearing, (3) the low cost associated with this availability, (4) the large size and diversity of the class, and (5) their historic place as being one of the first classes in which chromosomes were studied. For this review, I will concentrate on those topics of cytogenetics which have benefited the most from studies of chromosome behavior on insect spindles.
It should be noted, in the spirit of this tribute to insects, that even before the importance of insects as model systems for cytogenetics was discovered, insects unwillingly made an important technological contribution to this new science. In 1858, Gerlach described a staining procedure that, for the first time, adequately stained the nuclei, and later the chromosomes, of fixed cells (Moore, 1972). This procedure used a dilute solution of carmine, a dye long used by the Indians of Mexico. It so happens that carmine is made from the crushed and dried bodies of cochineal insects.
CHROMOSOME THEORY OF INHERITANCE
Mitosis was first described in the dividing eggs of the flatworm Mesostomum ehrenbergii (Schneider, 1873) and later more completely in the large clear epithelial cells, and erythroblasts, of the salamander (Flemming, 1879). Meiosis was first described in Ascaris (van Beneden, 1883). However, it was not until the beginning of the 1900s after Mendel’s classic work (Mendel, 1865) on heredity was rediscovered (1901), that the important connection between the behavior of chromosomes on the spindle and heredity became apparent. This led to the Chromosome Theory of Inheritance (Sutton, 1903) and the start of the field of cytogenetics. Studies on insects were instrumental in proving the Chromosome Theory of Inheritance. They linked an inheritable trait (sex) to a single chromosome, demonstrated the individuality of chromosomes, and showed that chromosome behavior on the spindle obeys Mendel’s first and second laws of heredity, the principle of segregation and the principle of independent assortment.
As an interesting side note to this tribute to insect cytogenetics, Mendel, after his successful experiments with plant hybrids, tried to extend his observations to insects by attempting to make hybrid crosses using bees (Voeller, 1968). However, this first attempt to use insects to study heredity failed. It turns out that Hymenoptera are unusual in that the males are haploid, and their chromosomes do not undergo a true reductional division during meiosis (Wilson, 1928). Therefore, genetic markers in hymenopteran males do not segregate or assort independently, as do those in peas. Later, of course, Morgan and his students discovered an ideal genetic model system in the fruit fly Drosophila melanogaster.
The first studies on cell division were done on organisms whose chromosomes were of similar size. Size diversity among chromosomes was first observed in insects, and allowed for a number of important discoveries that led to and supported

Contributions of Insect Systems to Cytogenetics 95
the Chromosome Theory of Inheritance. These important discoveries included the following.
Henking (1891) noted on spindles in spermatocytes of the bug Pyrrhocoris an extra chromosome that he called X. McClung (1902) also observed this extra chromosome, which he called an accessory chromosome, on spindles in various grasshopper species, and recognized that during spermatogenesis half of the sperm produced contained the extra chromosome and half did not. McClung (1902) associated this with sex, and was the first to assign a specific trait (sex) to a specific chromosome. Wilson (1905) later distinguished 2 types of sex determination (X0 and XY) in hemipterans (true bugs).
Montgomery (1901) used hemipterans to show for the first time that chromosomes were not alike. He saw that chromosomes on the spindle could differ in size. After studying the size diversity and the haploid number of the spermatogonial chromosomes of 42 hemipteran species, he concluded that maternal chromosomes paired only with paternal chromosomes during meiosis. In some species, 2 chromosomes were larger than the others. These 2 chromosomes, one of maternal origin and the other of paternal origin, always paired together. Maternal-paternal chromosome pairing was also evident in those species where the haploid number of chromosomes was uneven; if pairing of chromosomes with the same origin occurred, univalents would have resulted. Therefore, Montgomery (1901) demonstrated that pairing of chromosomes, like that of Mendelian factors (genes), brought maternal and paternal components together.
Sutton (1902) demonstrated the individuality of chromosomes in the grasshopper Bruchystolu magna. He found that its 23 chromosomes consisted of 11 chromosome pairs and a sex chromosome. Each pair was morphologically distinct from the other pairs and the sex chromosome. The same chromosome complement was observed on spindles in different cell generations and in cells from different parts of the grasshopper. He concluded that chromosomes retain their individuality between cell divisions and “that the constant morphological differences between the ordinary chromosomes are the visible expression of physiological or qualitative differences”. Sutton (1902) also correctly suggested that “the association of paternal and maternal chromosomes in pairs and their subsequent separation during the reducing division constitutes the physical basis of the Mendelian law of heredity”, i.e. the principle of segregation. A year later, Sutton (1903) proposed the Chromosome Theory of Inheritance and elegantly described the relationship between Mendel’s observations and the behavior of chromosomes. He showed that the behavior of pairs of chromosomes on the spindle during meiosis parallels that of pairs of Mendelian factors. During meiosis, the chromosomes of a pair, one maternal and one paternal, segregate to different gametes as predicted by Mendel’s principle of segregation. Sutton (1903) also concluded that the fate of one pair of homologous chromosomes was independent of any other, which was the cytological basis of Mendel’s principle of independent assortment.
It was Carothers (1913), however, who provided the actual cytological demonstration of Mendel’s principle of independent assortment. While studying chromosome behavior on the spindles of 3 grasshopper species, Bruchystolu magna, Arphiu simplex, and Dissosteiru curolina, she found that chromosomes of an homologous pair could be heteromorphic, i.e. one being larger than the other. She observed that the segregation of this pair of chromosomes to opposite poles of the spindle was independent of that of the accessory (sex) chromosome. Of 300 primary spermatocytes in B. magna scored, 154 (51.3%) showed the larger chromosome segregating with the accessory chromosome,

96 J. G. Ault
Pl WHITE-EYED FEMALE X RED-EYED MALE
Normal Gametes
Fl RED-EYED DAUGHTERS WHITE-EYED SONS
W
Fig. 1. Criss-cross inheritance. This diagram shows the relationship of sex-chromosome transmission and criss-cross expression of maternal and paternal phenotypes in sons and daughters, respectively. The X chromosome is marked with an eye-color gene, the Y chromosome is not. The red-eye allele
(w’) of that gene is dominant to the white-eye allele (w).
while the remaining 146 had the smaller chromosome segregating with it. Indeed, chromosomes assorted independently of each other, as did Mendelian factors.
Around the time when cytologists were providing cytological proof for the Chromosome Theory of Inheritance, Morgan and his students were observing and collecting heritable traits found in their Drosophila melunoguster stocks. They, like Mendel, made true-breeding strains with these traits. Morgan (1910, 1911) found that certain traits (e.g. eye color) appeared not to behave in simple Mendelian fashion. When comparing the progeny from reciprocal crosses, the phenotypic ratios were different. In one of the 2 reciprocal crosses, a criss-cross pattern, in which the female progeny had the paternal phenotype and the male progeny the maternal phenotype, occurred. Morgan (1910) suggested that the factors (genes) associated with these traits were linked to the sex factor X (X chromosome). Indeed, Stevens (1908) had earlier showed that the D. melunoguster female had 2 copies of the X chromosome, while the male had only one along with a Y chromosome. Recessive alleles of genes on maternal X chromosomes would be masked by dominant alleles on the paternal X in female progeny, but expressed in male progeny, resulting in the criss-cross phenotypic pattern (Fig. 1). Though Morgan was not the first to observe sex-linked traits (Doncaster and Raynor (1906) first observed sex linkage in moths), his experiments were the start of Drosophila genetics. They also set the stage for the definitive cytogenetic proof of the Chromosome Theory of Inheritance.
In crosses displaying the criss-cross pattern of sex-linked inheritance, Bridges (1916) observed exceptions among the progeny where males showed the paternal phenotype and females the maternal phenotype. Bridges (1916) postulated that, if the Chromosome Theory of Inheritance were correct, exceptional females must have maternal X chromosomes and exceptional males must have the paternal X chromosome. He hypothesized that this could occur if the maternal X chromosomes failed to disjoin

Contributions of Insect Systems to Cytogenetics 97
Pl WHITE-EYED FEMALE X RED-EYED MALE
Nondisjunctional Gametes
Normal Gametes
Fl EXCEPTIONAL EXCEPTIONAL WHITE-EYED DAUGHTERS RED-EYED MALES
TT r ww W’ 1 Fig. 2. The predicted maternal sex-chromosome non-disjunctional gametes and the confirming sex-chromosome content of exceptions to criss-cross inheritance. The X chromosome is marked with an eye-color gene, the Y chromosome is not. The red-eye allele (w’) of that gene is dominant
to the white-eye allele (w).
during the reductional division of oogenesis, resulting in an egg with either both maternal X chromosomes or none at all. The egg with both maternal X chromosomes would produce a female with the sex-linked recessive traits of the mother, if fertilized by a Y-containing sperm. The egg without maternal X chromosomes would produce a male with the sex-linked traits of the father, if fertilized by an X-containing sperm. Bridges (1916) tested his hypothesis cytologically and found, indeed, that exceptional females had 2 X chromosomes and a Y, while exceptional males had only an X chromosome (Fig. 2). The fact that Bridges (1916) could predict chromosome behavior from the behavior of genes left no doubt that genes are on chromosomes.
THE EXISTENCE OF SPINDLE FIBERS
When chromosome behavior on the spindle was first observed in living dividing cells, the chromosomes were the most prominent structures. Spindles were difficult to see and were usually only evident as fusiform clear areas devoid of mitochondria and large inclusions. Fibers within the spindle were observed in fixed and stained material; however, the reality of spindle fibers in living cells was of major debate (Schrader, 1953). The general belief at the time when Wilson (1928) reprinted the third edition of his classic book The Cell in Development and Heredity was that spindle fibers were fixation artifacts and that the spindle consisted of a weak non-fibrous gel. Insect studies helped, in part, to correct this belief and to formulate our current view of spindle structure.
Bela? (1929) provided some of the first indirect experimental evidence of the existence of spindle fibers within the living dividing cell. He studied the effects of hypertonicity on spermatocyte spindles of the grasshopper Chorthippus Zineatus. He found that spindles shrank in hypertonic media and that shrinkage was greater transversely than

98 J. G. Ault
longitudinally. The forces caused by cell shrinkage sometimes caused the spindle to fracture or bend. Splits occurred only in planes parallel to the longitudinal axis. Sharply bent spindles displayed bent fibers after the cells were fixed. The fibers came closer together in the bent region as if they existed as pliable rods in the living cell. In addition, Belai (1929) noted that Brownian movement of particles usually occurred longitudinally rather than transversely within the spindle. Together, these observations suggested that the spindle has longitudinal structure. Similar support for longitudinal structure within the spindle also came from centrifugation studies of organisms other than insects (reviewed in Schrader, 1953).
Not all spindles in living cells appeared structureless. In certain optically favorable living material, spindle fibers were observed using conventional light microscopy. These exceptions included the spindles of the cleavage cells in the mite Pediculopsis gruminum (Cooper, 1941) and of spermatocytes in the coccid Steatococcus tuberculutus (Hughes- Schrader and Ris, 1941). The spindle fibers observed in these living cells resembled those found in fixed cells.
Schmidt (1939) observed optical anisotropy in spindles of living dividing cells using polarized light and concluded that longitudinal differentiation occurred in the living spindle. However, the definitive study showing that spindle fibers exist in living cells came with technical improvements to the polarization microscope. An analysis of the limiting factors in polarization microscopy (InouC, 1952a, b) allowed for the develop- ment of an improved microscope with increased resolution and sensitivity. With this new microscope, InouC (1953) was able to see, for the first time, spindle fibers in living cells in a variety of organisms including the spermatocytes of the grasshoppers Chortophaga, Melanoplus, and Dissosteira and of the fruit fly Drosophila melanogaster. The spindle fibers in living cells corresponded to those found in fixed cells. To quote InouC (1953), “the demonstration of spindle fibers in living ceils by means of a polarization microscope, and the very close resemblance of the structure of living spindles to figures obtained from good fixed and stained preparations, give final proof for the reality of spindle fibers in living cells”.
THE MECHANICS OF CHROMOSOME BEHAVIOR ON THE SPINDLE
One area of functional cytogenetics in which insect studies have dominated is that of understanding the mechanics of chromosome behavior on the spindle. A powerful technique in these studies has been the manipulation of chromosomes on the spindle using a microneedle. Chromosome micromanipulation can be followed with live-cell observations and recordings using various light-microscope technologies, such as phase-contrast and polarization microscopy, or the cell can be fixed and processed for electron microscopy. Insect primary spermatocytes, particularly those of grasshoppers, have been the cell type of choice in these types of studies.
Kite and Chambers (1912) probably performed the first chromosome micromanipula- tion study when they dissected spermatocytes of squash bugs, grasshoppers and crickets using a fine glass needle in a Barber’s pipette holder. They showed that spindles retained their shape when freed from cytoplasm. While dissecting out single chromosomes from prophase, metaphase and telophase cells, a chromosome with a spindle fiber still attached was removed from a metaphase cell. The fiber was described as “a slightly refractive gel”. Chambers (1914) made similar dissections of the spermatocytes of the

Contributions of Insect Systems to Cytogenetics 99
grasshopper Dissosteira carolina and of the cockroach Periplaneta americana and came to the conclusion that the spindle was a hyaline gel of higher viscosity than the surrounding cytoplasm. He was able to easily remove chromosomes from the spindle, and therefore could not confirm the existence of spindle fibers.
Carlson (1952) performed less-destructive micromanipulations on chromosomes and spindles of dividing neuroblasts of the grasshopper Chortophaga viridifasciata. The microneedle was pushed into the cell without penetrating through the plasma membrane; i.e. the microneedle remained outside the cell and the cell remained alive. From his probing of the spindle with the microneedle, Carlson found that the spindle at metaphase was a semisolid body, consisting largely of longitudinally oriented components, in a fluid cytoplasm, and that the spindle was more “liquid” during prometaphase and anaphase. Most importantly, contrary to Chambers (1914), Carlson (1952) observed that chromosomes were attached to the spindle at their centromere (the term “kinetochore” is now usually used to describe the functional structure found at the chromosomal attachment site) region from prometaphase through early anaphase. Carlson’s observations verified and extended earlier evidence from microsurgical punctures of Tradescantia stamen hair cells (Wada, 1935) and from centrifugation studies (Schrader, 1935; Beams and King, 1936; Shimamura, 1940), that chromosomes were mechanically attached to the spindle (reviewed in Schrader, 1953).
Chromosome micromanipulation technology improved with the introduction of the piezoelectric micromanipulator (Ellis, 1962), and experiments became more sophisti- cated. Using this superior micromanipulator and following Carlson’s lead of performing less-destructive micromanipulations on chromosomes, from outside the cell, Nicklas and others performed a series of elegant chromosome micromanipulation experiments on grasshopper spermatocytes which contributed significantly to our knowledge about the mechanics of chromosome behavior on the spindle. These experiments provided insight into the physical properties of the spindle fiber, the initial orientation (attachment) of a bivalent to the spindle, the reorientation of mal-oriented bivalents, and the cause of orientation stability (summarized in Nicklas, 1974, 1985).
Nicklas and Staehly (1967) began this series of experiments by confirming the existence of chromosome spindle attachment in spermatocytes of the grasshopper Melanoplus differentialis and going on to characterize its physical properties. By tugging laterally on individual bivalents, they found that bivalents were mechanically attached individually to the spindle via their spindle fibers (seen with a polarization microscope) and that spindle fibers were anchored to the rest of the spindle near the pole. During these tugs, as the manipulated bivalent displayed extreme elasticity, the kinetochore-to- pole distances remained relatively unchanged, demonstrating the inelasticity of spindle fibers (Nicklas and Staehly, 1967). Only after prolonged or severe manipulation did kinetochore-to-pole distance increase slightly. During early to mid-anaphase, disjoined half-bivalents remained firmly attached to the pole. Besides lateral displacements, an individual half-bivalent could now be easily pushed close to the attached pole, by the microneedle, without affecting the movements of other half-bivalents, including its homolog. At late anaphase, the strength of chromosome spindle attachments lessened and individual half-bivalents could be displaced to the opposite pole, inducing artificial chromosome non-disjunction (Nicklas and Staehly, 1967).
Begg and Ellis (1979a, b) performed similar micromanipulations on spermatocytes of the grasshopper Trimeratropis maritima and of the crane fly Nephrotoma ferruginea.

100 J. G. Ault
Their results were consistent with those of Nicklas and Staehly (1967) and extended the demonstration that bivalents were mechanically attached to the spindle by their spindle fibers. Using the piezoelectric micromanipulator with a polarization microscope, Begg and Ellis (1979b) showed that abolishing the birefringent spindle fiber with colcemid and vinblastine resulted in the abolition of the chromosome-spindle attach- ment. In addition, by placing the microneedle next to a spindle fiber and tugging it laterally, they were able to displace the attached bivalent (Begg and Ellis, 1979a). Besides demonstrating mechanical attachment, this experiment also showed the flexibility of the spindle fiber, which could be bent at nearly a 90” angle without disruption. This flexibility was also evident when pushing an anaphase half-bivalent poleward. The birefringence of the spindle fiber attachment was not eliminated, but became diffuse after the chromosome was pushed to the pole, suggesting that only its parallel alignment was affected (Begg and Ellis, 1979a). Indeed, the displaced chromosome remained attached to the pole (Nicklas and Staehly, 1967) and continued anaphase movement once the other chromosomes reached the pole in living cells (Begg and Ellis, 1979a). It was concluded that the spindle fibers of both manipulated and unmanipulated chromosomes were shortening at the same rate and, once the slack of its spindle fiber was taken up, the manipulated chromosome moved poleward (Begg and Ellis, 1979a). Together, the experiments of Nicklas and Staehly (1967) and of Begg and Ellis (1979a, b) show that the spindle fibers attach chromosomes to the spindle. They also show that spindle fibers are flexible, but can support a tensile force that can stretch chromosomes up to 8 times their original length.
Nicklas (1967) investigated the relationship of kinetochore orientation, and reorienta- tion, during prometaphase to chromosome distribution at anaphase in M. differentialis spermatocytes. He found that, with increased force from a microneedle, it was possible to detach a bivalent from a spindle during prometaphase and metaphase without killing the cell, and that these detached bivalents would, in a short time, reattach. This allowed him to place detached bivalents in various positions within the cell and observe their reattachment to the spindle. Nicklas (1967) discovered that the direction in which a kinetochore faced was more important than polar distance in determining the pole to which the kinetochore will orient, i.e. the more a kinetochore faced a particular pole, the more likely it would be to orient to that pole, even when that pole was the farthest away. He was able, by positioning the bivalent, to direct the orientation and subsequent segregation of any given half-bivalent to a predetermined pole. This discovery illustrated the biological significance of the intrinsic structure of a bivalent (and of a mitotic chromosome). The kinetochores of both bivalents and mitotic chromosomes usually face in opposite directions (Schrader, 1936). Hence, when one kinetochore orients to a pole, its sister kinetochore is in position to interact with the opposite pole (bstergren, 1951). Nicklas (1967) induced mal-orientation by positioning both kinetochores of a bivalent to the same pole. Like natural mal-orientations, these induced mal-orientations invariably reoriented into the proper bipolar orientation prior to anaphase, resulting in normal chromosome segregation. Chromosome nondisjunction could only be induced in late anaphase.
Dietz (1956), while observing the behavior of bivalents on the spindle in living crane fly spermatocytes, concluded that only bipolar orientations were stable and all other orientations were unstable (see also Bauer et al., 1961). Dietz (1958) later suggested that perhaps the tension, demonstrated by the stretching of bipolarly oriented bivalents,
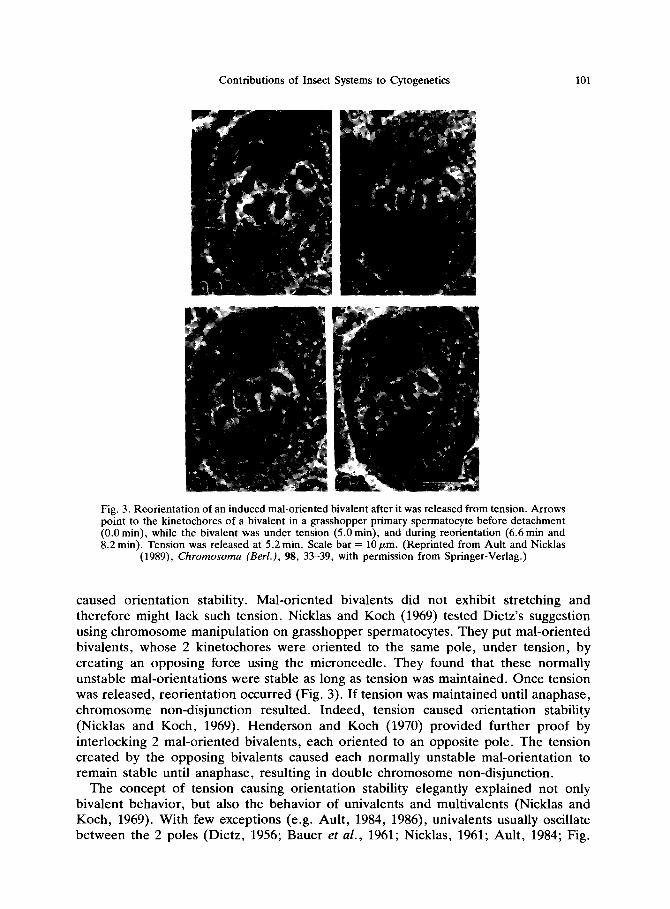
Contributions of Insect Systems to Cytogenetics 101
Fig. 3. Reorientation of an induced mal-oriented bivalent after it was released from tension. Arrows point to the kinetochores of a bivalent in a grasshopper primary spermatocyte before detachment (0.0 min), while the bivalent was under tension (5.0 min), and during reorientation (6.6 min and 8.2 min). Tension was released at 5.2 min. Scale bar = 10 pm. (Reprinted from Ault and Nicklas
(1989), Chromosoma (Berl.), 98, 33-39, with permission from Springer-Verlag.)
caused orientation stability. Mal-oriented bivalents did not exhibit stretching and therefore might lack such tension. Nicklas and Koch (1969) tested Dietz’s suggestion using chromosome manipulation on grasshopper spermatocytes. They put mal-oriented bivalents, whose 2 kinetochores were oriented to the same pole, under tension, by creating an opposing force using the microneedle. They found that these normally unstable mal-orientations were stable as long as tension was maintained. Once tension was released, reorientation occurred (Fig. 3). If tension was maintained until anaphase, chromosome non-disjunction resulted. Indeed, tension caused orientation stability (Nicklas and Koch, 1969). Henderson and Koch (1970) provided further proof by interlocking 2 mal-oriented bivalents, each oriented to an opposite pole. The tension created by the opposing bivalents caused each normally unstable mal-orientation to remain stable until anaphase, resulting in double chromosome non-disjunction.
The concept of tension causing orientation stability elegantly explained not only bivalent behavior, but also the behavior of univalents and multivalents (Nicklas and Koch, 1969). With few exceptions (e.g. Ault, 1984, 1986), univalents usually oscillate between the 2 poles (Dietz, 1956; Bauer et al., 1961; Nicklas, 1961; Ault, 1984; Fig.

102 J. G. Ault
Fig. 4. Typical univalent behavior on the spindle in a grasshopper primary spermatocyte. During prometaphase I, two small autosomal univalents (a and b) oscillate between the poles. The large arrow points to the X chromosome. Time is in minutes. Scale bar = 10 pm. (Reprinted from Ault
(1984) Chromosoma (Berl.), 89, 201-205, with permission from Springer-Verlag.)
4). The instability of their unipolar attachments and their continued reorientations are the result of no opposing force to generate tension and therefore to ensure orientation stability. In the case of multivalents, attachments are formed to both poles, but only certain configurations persist (e.g. Virkki, 1968). These stable configurations are those that appear to distribute tension equally among the spindle attachments (Nicklas and Koch, 1969).
SPINDLE ULTRASTRUCTURE
High-resolution observations of the spindle are achieved only with the use of the electron microscope. Whereas most information about the ultrastructure of the mitotic spindle comes from observations of cultured mammalian cells and Huemanthus endosperm cells, information about the meiotic spindle comes mostly from insect spermatocytes (Fuge, 1977; Rieder et al., 1993). The fine structure associated with specific types of chromosome behavior on the meiotic spindle has been studied almost exclusively in orthopteran and dipteran insects. Because of the specificity of this review, I will focus on the male insect meiotic spindle and its contributions to our understanding of the fine structure associated with chromosome behavior. The unique ultrastructure of lepidopteran spindles will not be discussed, for it has recently been reviewed extensively by Wolf (1994).

Contributions of Insect Systems to Cytogenetics 103
Electron microscopy suggests that the spindle consists mostly of microtubules (MTs). Membranes are also prevalent, either surrounding the spindle proper (e.g. Drosophila spermatocytes; Fig. 5) or interspersed among the MTs (e.g. Xenon peckii oocytes- Rieder and Nowogrodzki, 1983; for a recent review of spindle membranes, see Wolf, 1995). At each spindle pole of a primary spermatocyte is a centrosome, which consists of a pair of centrioles surrounded by pericentriolar material (Fig. 6). The 2 centrosomes are the primary microtubule-organizing centers (MTOCs) of the spindle (for a current review of MTOCs, see Kimble and Kuriyama, 1992).
Microtubules are generally classified by their location and associations. MTs penetrating the kinetochore are referred to as kinetochore microtubules (kMTs), whereas all others are referred to as non-kinetochore microtubles (nkMTs) (Fig. 7). The nkMTs can be further classified as: (1) polar MTs, which have one end at the pole and the other end free in the spindle; (2) aster MTs, which have one end at the pole and the other end outside the spindle proper; (3) interdigitating MTs, which have one end at the pole and the other end laterally associated with a MT from the opposite pole; (4) continuous MTs, which span the spindle with an end at each pole; and (5) free MTs, which have both ends free (Heath, 1981; McDonald, 1989). The majority of nkMTs appear to be either polar or interdigitating MTs. There are usually very few continuous or free MTs. It should be emphasized that these classifications are made from static images of fixed material. Because of the dynamic nature of MTs (Mitchison and Kirschner, 1984a, b), it is more than likely that individual MTs will fit more than one classification during the course of their existence. For example, kinetochores capture MTs (Hayden et al., 1990; Rieder et al., 1990). Therefore, captured polar nkMTs become kMTs. Another example is polar nkMTs that detach from the pole (Belmont et al., 1990). These MTs become free MTs until their depolymerization.
The microtubules of the spindle fiber, particularly kMTs, have been the most carefully characterized. They have long been implicated in chromosome attachment to the spindle and chromosome movement (reviewed in Nicklas, 1971; McIntosh and Hering, 1991; McIntosh, 1994). The most useful information has come from reconstructions from serial sections. Two-dimensional and partial 3-dimensional reconstructions have provided general descriptions of the distribution of spindle fiber MTs (Fuge, 1974a, 1980, 1984, 1985; Steffen, 1986a, b; Steffen and Fuge, 1982), while complete computer-generated 3-dimensional reconstructions have provided specific information on MT spacing and the interactions of different classes of MTs (McDonald et al., 1991, 1992). Serial-section EM studies of cells observed in life, up to the time of fixation, have provided important information about the fine structure associated with certain types of chromosome behaviors (Nicklas et al., 1979, 1982; Church and Lin, 1985; Nicklas and Kubai, 1985; Ault, 1986; Steffen, 1986a, b; Ault and Nicklas, 1989). Most ultrastructural observations of meiotic spindle fibers have been done on insect primary spermatocytes, particularly those of crane flies, grasshoppers and fruit flies.
One question about the spindle fiber that has been answered decisively by insect studies has been what is the relationship between kMT numbers and chromosome size within the same genome. Because chromosomes come in various sizes, it was suggested that the attachment and movement of large chromosomes may require more MT attachments than those of smaller chromosomes (Fuge, 1974b). Indeed, a positive correlation between kMT numbers and chromosome size appears to exist when one compares the average kMT numbers of different species (reviewed in Fuge, 1977).

104 J. G. Ault
Fig. 5. Spindle regions within (a) primary and (b) secondary spermatocytes in Drosophila melanogaster. Arrows point to membranes surrounding the spindles. Scale bar = 2 pm.

Contributions of Insect Systems to Cytogenetics 105
Fig. 6. Spindle pole of a Drosophila melunogasfer primary spermatocyte. The two centrioles of the centrosome are numbered. The arrow points to a chromosome kinetochore. Scale
bar = 1 pm.
Species with small chromosomes tend, on average, to have fewer kMTs per chromosome (e.g. 1 or 2 kMTs for small Physarum and Thruustotheca chromosomes) than those with large chromosomes (e.g. 120 kMTs for large Huemunthus chromosomes; reviewed in Fuge, 1974b, 1977). Moens (1979), however, counted the kMTs and determined the volumes of different-sized chromosomes within 3 species of orthopteran insects, and found no correlation between kMT number and chromosome size within the same genome. Lin et al. (1981) also found similar results in the dipteran D. melunoguster. Despite up to a lo-fold difference in chromosome size in D. melunoguster, kMT numbers remained similar between different chromosomes at any given stage (Lin et al., 1981).
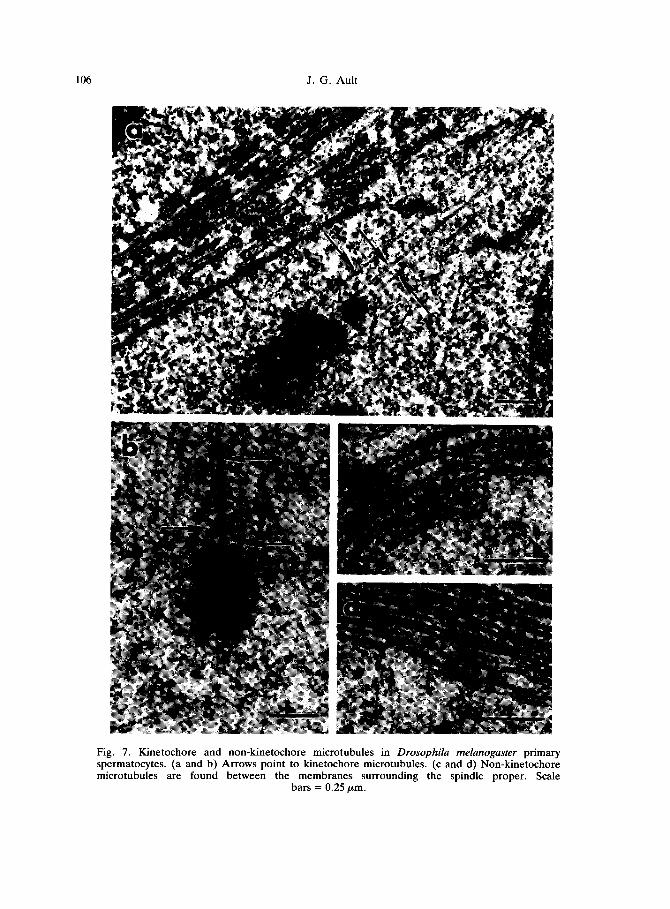
106 J. G. Ault
Fig. 7. Kinetochore and non-kinetochore microtubules in Drosophila melanogaster primary spermatocytes. (a and b) Arrows point to kinetochore microtubules. (c and d) Non-kinetochore microtubules are found between the membranes surrounding the spindle proper. Scale
bars = 0.25 pm.

Contributions of Insect Systems to Cytogenetics 107
Moens (1979) suggests that, “in addition to the mechanical demands of force, load, and viscosity, kMT numbers may be governed by cell type and evolutionary history of the karyotype”. This statement is supported in part by the observation that some large Robertsonian fusion chromosomes have about twice as many kMTs than other chromosomes in the genome and appear to be functional dicentric chromosomes (Moens, 1978, 1979).
Church and Lin (1982) may have been the first to follow and classify every MT within a meiotic spindle fiber of a higher eukaryote. They reconstructed completely 14 prometaphase-I spindle fibers of the dipteran D. melunoguster. Their study was especially important for 2 reasons. Firstly, they found that the majority of the MTs (54%) extended from the polar region towards, but did not end in, the kinetochore proper. These MTs either ended short of the kinetochore, grazed it, or passed through it. This suggested that the majority of spindle fiber MT’s were of polar origin and that spindle fiber formation occurred by the kinetochore capturing, rather than nucleating, MTs. Secondly, they observed that a kinetochore need not face directly the pole to which it is oriented (attached). For example, they found that a kinetochore directly facing one pole may have MT connections with both poles and that a kinetochore facing perpendicular to the spindle axis may have MT connections only to one pole. This helped to visualize how kinetochore reorientation could occur, in that it showed that a kinetochore oriented to one pole could interact with MTs from the opposite pole, a pole toward which it usually did not face. Previously, such interactions seemed important in kinetochore reorientation, but were hard to imagine after the experimental results of Nicklas (1967).
Nicklas and Kubai (1985) used correlative living-cell/serial-section electron microscopy on grasshopper spermatocytes to study the relationship between MTs and the movement of chromosomes detached from the spindle by micromanipulation. Recently detached chromosomes lack or at most have only a single short MT, and do not move. Nicklas and Kubai (1985) showed that continuation of chromosome movement was always correlated with at least one kMT running from the kinetochore to the polar region. These initial kMT attachments were most likely the result of kinetochore capture rather than nucleation because no kMTs of intermediate length were ever observed prior to movement, and no kMTs were associated with the sister kinetochore facing away from the spindle at time of movement. Nicklas and Kubai (1985) also found that, no matter how unusual the movement, it could be explained by poleward forces acting parallel to the kMT axis. Such findings were also observed in D. melanogaster spermatocytes (Church and Lin, 1985). Kinetochore reorientation was observed in both studies (Church and Lin, 1985; Nicklas and Kubai, 1985) and appeared to involve the loss of MT attachments from one pole and the establishment of new MT attachments to the opposite pole. This led Church and Lin (1985) to hypothesize “that the majority of short kMTs are remnants of previous MT-mediated associations between a kinetochore and a pole”.
Ault and Nicklas (1989)) using correlative living-cell/serial-section electron microscopy on grasshopper spermatocytes, studied the sequence of events during kinetochore reorientation and the effects that tension-induced stability has on kMTs and the proper distribution of chromosomes. Mal-oriented bivalents having both kinetochores oriented to one pole were induced by micromanipulation. Such mal-orientations are stable while the bivalent is under tension by a force created using a microneedle but unstable after tension is released (Nicklas and Koch, 1969). Unstable bivalents always reorient with

J. G. Ault
Fig. 8. Reorientation of a large grasshopper bivalent. Arrows point to both kinetochores before detachment (0.0 min) and while the bivalent was under tension (4.6 min). Tension was released at 4.7 min. The reorienting kinetochore moved toward the upper pole (6.0 min and 10.7 min), while the other kinetochore turned away from the lower pole (6.4 min, 8.6 min, 10.2 min and 11.0 min). Scale bar = 10 pm. (Reprinted from Ault and Nicklas (1989), Chromosoma (Berl.), 98,33-39, with
permission from Springer-Verlag.)
movement of one kinetochore toward the opposite pole. Ault and Nicklas (1989) found that MTs associated with stably oriented bivalents, whether they are mal-oriented or in normal bipolar orientation, were arranged in orderly parallel bundles running from each kinetochore toward the pole. After tension was released, these orderly bundles continued until kinetochore movement, at which time they became more splayed. A reorienting kinetochore started out with MTs running toward both poles. Though few kMTs ran to the pole to which the kinetochore was reorienting, they were enough to move the chromosome. This was because all other kMTs of the reorienting kinetochore had lost their anchorage to the original pole and were being dragged passively as the kinetochore moved toward the other pole (Figs. 8 and 9). Ault and Nicklas (1989) showed that “the stable anchorage of kMTs to the spindle is associated with tension force and unstable anchorage with the absence of tension”. They hypothesized that “the effect of tension is readily explained if force production and anchorage are both produced by mitotic motors, which link MTs to the spindle as they generate tension forces”.
THE LOCATION AND FORCE OF THE CHROMOSOME MOTOR DURING
ANAPHASE
Most current evidence suggests that chromosome movement is at least in part the result of molecular motors located somewhere within the kinetochore (Ault and Rieder,

Contributions of Insect Systems to Cytogenetics 109
Fig. 9. Three-dimensional reconstruction (stereo pair) of the reorienting bivalent in Fig. 8. Distal ends of the kinetochore microtubules are boxed. The kinetochore microtubule of the reorienting kinetochore that extended farthest toward the upper pole (arrows and boxes at both ends) ran through the kinetochore. Two kinetochore microtubules of the non-reorienting kinetochore that ended near the lower pole (centriole and pericentriolar material are outlined) are indicated by single arrowheads and a curvilinear one by a double arrowhead. Scale bar = 2 pm. (Reprinted from Ault
and Nicklas (1989), Chromosoma (Bed.), 98, 33-39, with permission from Springer-Verlag.)
1994). Before numerous motor proteins were hailed as the putative mitotic motor that moved chromosomes poleward during anaphase, the location of this motor was determined decisively to be in or near the kinetochore (Gorbsky et al., 1987, 1988; Koshland et al., 1988; Nicklas, 1989). Of these elegant studies, that by Nicklas (1989), using an insect model system, was the most direct. Using a microneedle, he cut through the spindle of grasshopper spermatocytes between the chromosomes and one pole, and swept the polar region away. These cuts were clean and completely severed the spindle (Nicklas et al., 1989). Despite the removal of the pole and part of the spindle fiber, chromosome movement continued, sometimes within a micrometer of the cut edge (Nicklas, 1989). This left little doubt that the mitotic motor for poleward chromosome movement during anaphase was in or near the kinetochore.
The maximum force that the spindle can generate during chromosome movement at anaphase was also first measured in grasshopper primary spermatocytes. Again, Nicklas and his skill at chromosome micromanipulation did the job. Using a flexible microneedle calibrated in dynes per micrometer of tip deflection, Nicklas (1983) applied just enough opposing force to stop the poleward movement of an anaphase chromosome. The deflection of the needle determined the maximum force produced by the spindle to be of the order of 7 x 10m5 dynes. Interestingly, this is 10,000 times greater than previous calculations, which considered only the force needed to overcome the viscous resistance to chromosome movement (Nicklas, 1965; Taylor, 1965). This means that the spindle

110 J. G. Ault
has the force potential to overcome increases in resistance to chromosome movement, caused either by large genomic changes or by problems associated with chromosome disjunction.
CONCLUSION
We have learned much about chromosome behavior on the spindle by studying insects and no doubt will learn much more in the future. In this era of molecular biology, the dipteran D. melunogaster remains an important model system. Insect primary sper- matocytes will always remain ideal material to observe directly chromosome behavior on the male meiotic spindle using any of a number of optical technologies, including correlative living-cell/serial-section electron microscopy (Ault and Nicklas, 1989). Also, insect eggs and embryos have been, for the first time, coupled with recent technological advances to allow adequate observations of chromosome behavior on the female meiotic spindle and on the early mitotic spindles after fertilization (Theurkauf and Hawley, 1992; McDonald and Morphew, 1993; McDonald, 1994). Such studies will provide important structural information about mitosis and female meiosis, and in the case of D. melunogaster with its many known mitotic and meiotic mutants, will provide important insights into the function of many known genes that are essential to mitosis and meiosis.
One important contribution of insects that was not covered in this review is the abundance of unorthodox spindles, and of odd chromosome behavior, found in this class (e.g. see Kubai and Wise, 1981; Kubai, 1982, 1987). These natural experiments allow us to test the tenets that have been put forth about chromosome behavior on the spindle (Kubai, 1975; Heath, 1980; Wise, 1988). The early work of Sally Hughes-Schrader on coccids (e.g. Hughes-Schrader, 1948a) and mantids (e.g. Hughes-Schrader, 1943b, 1948b, 1950, 1969, 1979) points to gold veins yet to be mined by modern technology. And to think that the number of different insects studied is minuscule when compared with the number of insects which have yet to be studied, makes predicting the number of new gold mines impossible. Indeed, insects will remain important teachers of mitosis and meiosis to cytogeneticists for years to come.
REFERENCES Ault, J. G. 1984. Unipolar orientation stability of the sex univalent in the grasshopper (Melunoplus
sanguinipes) . Chromosoma 89: 20145. Ault, J. G. 1986. Stable versus unstable orientations of sex chromosomes in two grasshopper species.
Chromosoma 93: 298-304. Ault, J. G. and R. B. Nicklas. 1989. Tension, microtubule rearrangements, and the proper distribution of
chromosomes in mitosis. Chromosoma 98: 33-9. Ault, J. G. and C. L. Rieder. 1994. Centrosome and kinetochore movement during mitosis. Cur. Opin. Cell
Biol. 6: 41-9. Bauer, H., R. Dietz and C. Robbelen. 1961. Die Spermatocytenteilungen der Tipuliden. III. Mitt.
Das Bewegungsverhalten der Chromosomen in Translokationsheterozygoten von Tip& olerucea. Chromosoma 12: 116-89.
Beams, H. W. and R. L. King. 1936. The effects of ultracentrifuging upon chick embryonic cells, with special reference to the “resting” nucleus and the mitotic spindle. Biol. Bull. (Woods Hole) 71: 188-98.
Begg, D. A. and G. W. Ellis. 1979a. Micromanipulation studies of chromosome movement. I. Chromosome- spindle attachment and the mechanical properties of chromosomal spindle fibers. J. Cell Biol. 82: 52841.
Begg, D. A. and G. W. Ellis. 197913. Micromanipulation studies of chromosome movement. II. Birefringent

Belai,
Contributions of Insect Systems to Cytogenetics 111
chromosomal fibers and the mechanical attachment of chromosomes to the swindle. J. Cell Biol. 82: 542-54. K. 1929. Beitrlge zur Kausalanalyse der Mitose. II. Untersuchungen an den Spermatocyten von Chorthippus (Stenobothrus) lineatus Panz. Wilhelm Roux’ Arch. Entwicklungsmech. Org. 118: 3.59-480.
Belmont, L. D., A. A. Hyman, K. E. Sawin and T. J. Mitchison. 1990. Real-time visualization of cell cycle-dependent changes in microtubule dynamics in cytoplasmic extracts. Cell 62: 579-89.
Bridges, C. B. 1916. Non-disjunction as proof of the chromosome theory of heredity. Genetics 1: l-52, 107-63.
Carlson, J. G. 1952. Microdissection studies of the dividing neuroblast of the grasshopper, Chortophaga viridifasciata (De Geer). Chromosoma 5: 199-220.
Carothers, E. E. 1913. The Mendelian ratio in relation to certain Orthopteran chromosomes. J. Morphol. 24: 487-509.
Chambers, R. Jr. 1914. Some physical properties of the cell nucleus. Science (Wash., D.C.) 40: 824-27. Church, K. and H.-P. P. Lin. 1982. Meiosis in Drosophila melanogaster. II. The prometaphase-I kinetochore
microtubule bundle and kinetochore orientation in males. J. Cell Biol. 93: 365-73. Church, K. and H.-P. P. Lin. 1985. Kinetochore microtubules and chromosome movement during
prometaphase in Drosophila melanogaster spermatocytes studied in life and with the electron microscope. Chromosoma 92: 273-82.
Cooper, K. W. 1941. Visibility of the primary spindle fibers and the course of mitosis in the living blastomeres of the mite, Pediculopsis graminum Reut. Proc. Nat1 Acad. Sci. USA 21: 48&83.
Dietz, R. 1956. Die Spermatocytenteilungen der Tipuliden. II. Graphische Analyse der Chromosomen- bewegung wlhrend der Prometaphase I im Leben. Chromosoma 8: 183-211.
Dietz, R. 1958. Multiple Geschlechtschromosomen bei den Ostracoden, ihre Evolution und ihr Teilungsver- halten. Chromosoma 9: 359440.
Doncaster, L. and G. H. Raynor. 1906. Breeding experiments with Lepidoptera. Proc. Zool. Sot. Land. 1: 125-33.
Ellis, G. W. 1962. Piezoelectric micromanipulators. Science (Wash., D.C.) 138: 84-91. Flemming, W. 1879. Contributions to the knowledge of the cell and its life appearance. Arch. Mikrosk. Anat.
16: 302406. Fuge, H. 1974a. The arrangement of microtubules and the attachment of chromosomes to the spindle during
anaphase in tipulid spermatocytes. Chromosoma 45: 245-60. Fuge, H. 1974b. Ultrastructure and function of the spindle apparatus microtubules and chromosomes during
nuclear division. Protoplasma 82: 289-320. Fuge, H. 1977. Ultrastructure of the mitotic spindle. ht. Rev. Cytol. Suppl. 6: l-58. Fuge, H. 1980. Microtubule disorientation in anaphase half-spindles during autosome segregation in crane
fly spermatocytes. Chromosoma 76: 309-28. Fuge, H. 1984. The three-dimensional architecture of chromosome fibres in the crane fly. I. Syntelic autosomes
in meiotic metaphase and anaphase I. Chromosoma 90: 32331. Fuge, H. 1985. The three-dimensional architecture of chromsome fibres in the crane fly. II. Amphitelic sex
univalents in meiotic anaphase I. Chromosoma 91: 322-28. Gorbsky, G. J., P. J. Sammak and G. G. Borisy. 1987. Chromosomes move poleward in anaphase along
stationary microtubules that coordinately disassemble from their kinetochore ends. J. Cell Biol. 104: 9-18.
Gorbsky, G. J., P. J. Sammak and G. G. Borisy. 1988. Microtubule dynamics and chromosome motion visualized in living anaphase cells. J. Cell Biol. 106: 1185-92.
Hayden, J. H., S. S. Bowser and C. L. Rieder. 1990. Kinetochores capture astral microtubules during chromosome attachment to the mitotic spindle: direct visualization in live newt lung cells. J. Cell Biol. 111: 103945.
Heath, I. B. 1980. Variant mitoses in lower eukaryotes: indicators of the evolution of mitosis? Znt. Rev. Cytol. 64: l-80.
Heath, I. B. 1981. Mitosis through the electron microscope, pp. 245-75. In A. M. Zimmerman and A. Forer (eds.) MitosislCytokinesis. Academic Press, New York.
Henderson, S. A. and C. A. Koch. 1970. Co-orientation stability by physical tension: a demonstration with experimentally interlocked bivalents. Chromosoma 29: 207-16.
Henking, H. 1891. Untersuchungen iiber die ersten Entwicklungsvorgange in den Eiem der Insekten. Z. Wiss. Zool. 51: 685-736.
Hughes-Schrader, S. 1943a. Polarization, kinetochore movements, and bivalent structure in the meiosis of male mantids. Biol. Bull. (Woods Hole) 85: 265-300.
Hughes-Schrader, S. 1943b. Meiosis without chiasmata in diploid and tetraploid spermatocytes of the mantid Callimantis anttllarum Saussure. J. Morphol. 73: 11141.
Hughes-Schrader, S. 1948a. Cytology of coccids (Cocco’idea-Homoptera). Adv. Genet. 2: 127-203.

112 J. G. Ault
Hughes-Schrader, S. 1948b. Expulsion of the sex chromosome from the spindle in spermatocytes of a mantid. Chromosoma 3: 257-70.
Hughes-Schrader, S. 1950. The chromosomes of mantids (Orthoptera : Manteidae) in relation to taxonomy. Chromosoma 4: l-55.
Hughes-Schrader, S. 1969. Distance segregation and compound sex chromosomes in mantispids (Neurop- tera : Mantispidae). Chromosoma 27: 109-29.
Hughes-Schrader, S. 1979. Diversity of chromosomal segregational mechanisms in mantispids (Neurop- tera : Mantispidae). Chromosoma 75: l-17.
Hughes-Schrader, S. and H. Ris. 1941. The diffuse spindle attachment of coccids, verified by the mitotic behavior of induced chromosome fragments. J. Exp. Zool. 87: 429-56.
InouC, S. 1952a. Studies on depolarization of light at microscope lens surfaces. I. The origin of stray light by rotation at the lens surfaces. Exp. Cell Res. 3: 199-208.
Inoue, S. 1952b. Improvements of the polarization microscope for biological purposes. Trans. Amer. Microsc. Sot. 71: 311-12.
InouC, S. 1953. Polarization optical studies of the mitotic spindle. I. The demonstration of spindle fibers in living cells. Chromosoma 5: 487-500.
Kimble, M. and R. Kuriyama. 1992. Functional components of microtubule-organizing centers. Int. Rev. Cytol. 136: l-50.
Kite, G. L. and R. Chambers Jr. 1912. Vital staining of chromosomes and the function and structure of the nucleus. Science (Wash., D. C.) 36: 63wl.
Koshland, D. E., T. J. Mitchison and M. W. Kirschner. 1988. Polewards chromosome movement driven bv microtubule depolymerization in vitro. Nature (Lond.) 331: 499-504.
Kubai, D. F. 1975. The evolution of the mitotic soindle. Znt. Rev. Cvtol. 43: 167-227. Kubai, D. F. 1982. Meiosis in Sciara coprophila: &ucture of the spindle and chromosome behavior during
the first meiotic division. J. Cell Biol. 93: 655-69. Kubai, D. F. 1987. Nonrandom chromosome arrangements in germ line nuclei of Sciara coprophila males:
the basis for nonrandom chromosome segregation on the meiosis I spindle. .Z. Cell Biol. 105: 2433-46.
Kubai, D. F. and D. Wise. 1981. Nonrandom chromosome segregation in Neocurtilla (Gryllotalpa) hexadacryla: an ultrastructural study. J. CeN Biol. 88: 281-93.
Lin, H.-P., J. G. Ault and K. Church. 1981. Meiosis in Drosophila melanogaster. I. Chromosome identification and kinetochore microtubule numbers during the first and second meiotic divisions in males. Chromosoma 83: 507-21.
Mazia, D. 1961. Mitosis and the physiology of cell division, pp. 77412. In J. Brachet and A. E. Mirsky (eds.) The CeN. Academic Press, New York.
McClung, C. E. 1902. The accessory chromosome-sex determinant? Biol. Bull. (Woods Hole) 3: 43-84. McDonald, K. 1994. Electron microscopy and EM immunocytochemistry. Methods in Cell Biol. 44:
41144. McDonald, K. 1989. Mitotic spindle ultrastructure and design, pp. l-38. In J. S. Hyams and B. R. Brinkley
(eds) Mitosis: Molecules and Mechanisms. Academic Press, New York. McDonald, K. and M. K. Morphew. 1993. Improved preservation of ultrastructure in difficult-to-fix organisms
by high pressure freezing and freeze substitution: I. Drosophila melanogaster and Strongylocentrotus purpuratus embryos. Microsc. Res. Tech. 24: 465-73.
McDonald, K., D. Mastronarde, E. O’Toole, R. Ding and J. R. McIntosh. 1991. Computer-based tools for morphometric analysis of mitotic spindles and other microtubule systems. EMSA Bull. 21: 47-53.
McDonald, K., E. O’Toole, D. Mastronarde and J. R. McIntosh. 1992. Kinetochore microtubules in PTK cells. J. Cell Biol. 118: 369-83.
McIntosh, J. R. 1994. The roles of microtubules in chromosome movement, pp. 413-34. In J. S. Hyams and C. W. Lloyd (eds) Microtubules. Wiley-Liss, Inc., New York.
McIntosh, J. R. and G. E. Hering. 1991. Spindle fiber action and chromosome movement. Annu. Rev. Cell Biol. 7: 403-26.
Mendel, G. 1865. Verhandlungen des naturforschenden Vereines in Brtlnn 4. Abhandlungen, pp. 3-47. (English translation published in .Z. Royal Horticultural Society of London, 1901.)
Mitchison, T. J. and Kirschner, M. W. 1984a. Microtubule assemblv nucleated bv isolated centrosomes. Nature (Lond.) 312: 232-37.
Mitchison, T. J. and Kirschner, M. W. 1984b. Dynamic instability of microtubule growth. Nature (Land.) 312: 237-42.
Moens, P. B. 1978. Kinetochores of grasshoppers with Robertsonian fusions. Chromosoma 67: 41-54. Moens, P. B. 1979. Kinetochore microtubule numbers of different sized chromosomes. Z. Cell Biol. 83:
556-61. Montgomery, T. H. Jr. 1901. A study of the chromosomes of the germ cells of metazoa. Trans. Amer. Z’hilos.
Sot. 20: 154-230.

Contributions of Insect Systems to Cytogenetics 113
Moore, J. A. 1972. Heredity and Development. 2nd edition. Oxford University Press, New York. Morgan, T. H. 1910. Sex-limited inheritance in Drosophila. Science (Wash., D.C.) 32: 120-22. Morgan, T. H. 1911. Random segregation versus coupling in Mendelian inheritance. Science (Wash., D.C.)
34: 384. Nicklas, R. B. 1961. Recurrent pole-to-pole movements of the sex chromosome during prometaphase I in
Melanoplus differential& spermatocytes. Chromosoma 12: 97-115. Nicklas, R. B. 1965. Chromosome velocity during mitosis as a function of chromosome size and position.
J. Cell Biol. 25: 119-35. Nicklas, R. B. 1967. Chromosome micromanipulation. II. Induced reorientation and the experimental control
of segregation in meiosis. Chromosoma 21: 17-50. Nicklas, R. B. 1971. Mitosis. Adv. Cell Biol. 2: 225-94. Nicklas, R. B. 1974. Chromosome segregation mechanisms. Genetics 78: 205-13. Nicklas, R. B. 1983. Measurements of the force produced by the mitotic spindle in anaphase. J. Cell Biol.
97: 542-48. Nicklas, R. B. 1985. Mitosis in eukaryotic cells: an overview of chromosome distribution, pp. 18%94. In V.
L. Dellarco, P. E. Voytek and A. Hollaender (eds) Aneuploidy. Plenum, New York. Nicklas, R. B. 1989. The motor for poleward chromosome movement in anaphase is in or near the
kinetochore. J. Cell Biol. 109: 2245-55. Nicklas, R. B., B. R. Brinkley, D. A. Pepper, D. F. Kubai and G. K. Rickards. 1979. Electron microscopy
of spermatocytes previously studied in life: methods and some observations on micromanipulated chromosomes. J. Cell Sci. 35: 87-104.
Nicklas, R. B. and C. A. Koch. 1969. Chromosome micromanipulation. III. Spindle fiber tension and the reorientation of mal-oriented chromosomes. J. Cell Biol. 43: 40-50.
Nicklas, R. B. and D. F. Kubai. 1985. Microtubules, chromosome movement, and reorientation after chromosomes are detached from the spindle by micromanipulation. Chromosoma 92: 313-24.
Nicklas, R. B., D. F. Kubai and T. S. Hays. 1982. Spindle microtubules and their mechanical associations after micromanipulation in anaphase. J. Cell Biol. 95: 91-104.
Nicklas, R. B., G. M. Lee, C. L. Rieder and G. Rupp. 1989. Mechanically cut mitotic spindles: clean cuts and stable microtubules. J. Cell Sci. 94: 415-23.
Nicklas, R. B. and C. A. Staehly. 1967. Chromosome micromanipulation. I. The mechanics of chromosome attachment to the spindle. Chromosoma 21: 1-16.
Nicklas. R. B. and S. C. Ward. 1994. Elements of error correction in mitosis: microtubule capture, release, and tension. J. Cell Biol. 126: 1241-53.
Gstereren. G. 1951. The mechanism of co-orientation in bivalents and multivalents. The theory of orientation ” by’pulling. Hereditas 37: 85-156.
Rieder, C. L., S. P. Alexander and G. Rupp. 1990. Kinetochores are transported poleward along a single astral microtubule during chromosome attachment to the spindle in newt lung cells. J. Cell Biol. 110: 81-95.
Rieder, C. L., J. G. Ault, U. Eichenlaub-Ritter and G. Sluder. 1993. Morphogenesis of the mitotic and meiotic spindle: conclusions obtained from one system are not necessarily applicable to the other, pp. 183-97. In B. K. Vig (ed.) Chromosome Segregation and Aneuploidy, NATO AS1 Series, Subseries H, Vol. 72. Springer-Verlag, Berlin.
Rieder, C. L. and R. Nowogrodzki. 1983. Intranuclear membranes and the formation of the first meiotic spindle in Xenos peckii (Acroschismus wheeleri) oocytes. J. Cell Biol. 97: 1144-55.
Schmidt, W. J. 1939. Doppelbrechung der Kernspindel und Zugfasertheorie der Chromosomenbewegung. Chromosoma 1: 253-64.
Schneider, A. 1873. Untersuchungen iiber Platyhelminthen. Ober. Ges. Nat. Heilkund. 14: 69-140. Schrader, F. 1935. On the reality of spindle fibers. Biol. Bull. (Woods Hole) 67: 519-53. Schrader, F. 1936. The kinetochore or spindle fiber locus in Amphiuma tridactylum. Biol. Bull. (Woods Hole)
70: 484-98. Schrader, F. 1953. Mitosis. The Movements of Chromosomes in Cell Division. Columbia University Press,
New York. Shimamura, T. 1940. The mechanism of nuclear division and chromosome arrangement. VI. Studies on the
effect of the centrifugal force upon nuclear division. Cytologia 11: 186-216. Steffen, W. 1986a. Three-dimensional architecture of chromosome fibres in the crane fly: co-oriented
autosomal bivalents and amphitelic sex univalents during prometaphase. Chromosoma 94: 107-14. Steffen, W. 1986b. Relationship between the arrangement of microtubules and chromosome behaviour of
syntelic autosomal univalents during prometaphase in crane fly spermatocytes. Chromosoma 94: 412-18.
Steffen, W. and H. Fuge. 1982. Dynamic changes in autosomal spindle fibers during prometaphase in crane fly spermatocytes. Chromosoma 87: 36S71.
Stevens, N. M. 1908. A study of the germ cells of certain Diptera, with reference to the heterochromosomes

114 J. G. Ault
and the phenomena of synapsis. J. Exp. Zool. 5: 359-83. Sutton, W. S. 1902. On the morphology of the chromosome group in Brachystola magna. Biol. Bull. (Woods
Hole) 4: 24-39. Sutton, W. S. 1903. The chromosomes in heredity. Biol. Bull. (Woods Hole) 4: 231-51. Taylor, E. W. 1965. Brownian and saltatory movements of cytoplasmic granules and the movement of
anaphase chromosomes, pp. 175-91. In A. L. Copley (ed.) Proceedings of the Fourth International Congress on Rheology, Part 4, Symposium on Biorheology. Interscience, New York.
Theurkauf, W. E. and R. S. Hawley. 1992. Meiotic spindle assembly in Drosophila females: behavior of nonexchange chromosomes and the effects of mutations in the nod kinesin-like protein. J. Cell Biol. 116: 116780.
van Beneden, E. 1883. Recherches sur la maturation de l’oeuf et la fecondation. Arch. Biol. 4: 265-640. Virkki, N. 1968. A chiasmate sex quadrivalent in the male of an alticid beetle, Cyrsylus volkameriae. Can.
J. Genet. Cytol. 10: 898-907. Voeller, B. R. 1968. The Chromosome Theory of Inheritance. Appleton-Century-Crofts, New York. Wada, B. 1935. Mikrurgische Untersuchungen lebender Zellen in der Teilung. II. Das Verhalten der
Spindelfigur und einige ihrer physikalischen Eigenschaften in den somatischen Zellen. Cytologia 6: 381-406.
Wilson, E. B. 1905. The chromosomes in relation to the determination of sex in insects. Science (Wash., D.C.) 22: 500-02.
Wilson, E. B. 1928. The Cell in Development and Heredity. Macmillan, New York. Wise, D. 1988. The diversity of mitosis: the value of evolutionary experiments. Biochem. Cell Biol. 66:
515-29. Wolf, K. W. 1994. The unique structure of Lepidopteran spindles. Znt. Rev. Cytol. 152: 148. Wolf, K. W. 1995. Spindle membranes and spindle architecture in invertebrates. Micron 26: 69-98.
Related Documents











