Hydrobiologia 354: 165–175, 1997. 165 c 1997 Kluwer Academic Publishers. Printed in Belgium. Contribution to a revision of Floscularia Cuvier, 1798 (Rotifera: Monogononta): notes on some Neotropical taxa Hendrik Segers Laboratory of Animal Ecology, Zoogeography and Nature Conservation, Department M.S.E., University of Ghent, K.L. Ledeganckstraat 35, B-9000 Gent, Belgium Received 19 June 1997; in revised form 19 August 1997; accepted 28 September 1997 Key words: Taxonomy, Rotifera, Floscularia, biogeography, autecology Abstract The taxonomy of some Neotropical representatives of the genus Floscularia Cuvier, 1798 (Rotifera: Monogononta) is discussed, based on SEM studies of trophi and external morphology. Floscularia armata n.sp. and F. bifida n. sp. are described, and F. conifera (Hudson, 1886) is restored at full species rank. A hypothesis on the functional relevance of differences in trophi morphology for niche segregation is formulated. Introduction In the most recent taxonomic key including floscu- lariid Rotifera (Koste, 1978), the genus Floscularia Cuvier, 1798 is listed as containing six valid morphos- pecies and one subspecies. Most of these have been described from the northern hemisphere, and have subsequently been recorded from almost all parts of the world. Exceptions are Floscularia noodti Koste, 1972, recorded only from the Neotropical region (see Koste, 1978; De Ridder & Segers, 1997), and F. deco- ra Edmondson, 1940, recorded only from the northern hemisphere. This situation is symptomatic for Rotifera, as it reflects present-day taxonomy as commented upon by, amongst others, Dumont (1983) and Koste & Shiel (1989). Poor taxonomic resolution has resulted in the hypothesis of potential cosmopolitanism for all Rotifera (Ruttner-Kolisko in Dumont, 1980). It is now clear that this view needs revision. Pejler (1977), De Ridder (1981), Dumont (1983) and Segers (1996a) have demonstrated a wide variety of distribution pat- terns in rotifers. A study on the genus Scaridium Ehren- berg, 1830, a group with a highly characteristic general body size, revealed that detailed morphological studies are required to distinguish morphotaxa with circum- scribed distributions (Segers, 1995; 1996b). The situa- tion in Floscularia is reminiscent of Scaridium. Here, identification of relatively large animals relies mostly on the structure of the tube they inhabit, and on some other, easily appreciable features; trophi morphology has never been considered. Therefore, it did not come as a surprise to find several undescribed morphospecies of Floscularia in samples from a subtropical Brazilian reservoir, and to find that these are most reliably char- acterized by their trophi morphology, in addition to the features considered to date. In this contribution I report on the four Floscularia morphospecies that co-occurred in Broa reservoir (Iti- rapira, SP, Brazil) on 13 February 1996, and comment on the relevance of the differences in trophus structure among the four taxa. Material and methods The material treated here was collected in Broa reser- voir, Itirapira, S˜ ao Paulo, Brazil. Living specimens were collected from submerged macrophytes and stud- ied at CRHEA, Universidade de S˜ ao Paulo, Brazil. Pre- served animals contained in net samples (mesh width 50 m) collected on 19 January or 13 February 1996 and fixed in formaldehyde (4%) were studied using a Wild M10 dissection microscope and a Olympus CH- 2 microscope with drawing tube. Scanning electron microscopy (S.E.M.) was performed using a Jeol JSM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hydrobiologia 354: 165–175, 1997. 165c 1997Kluwer Academic Publishers. Printed in Belgium.
Contribution to a revision of Floscularia Cuvier, 1798 (Rotifera:Monogononta): notes on some Neotropical taxa
Hendrik SegersLaboratory of Animal Ecology, Zoogeography and Nature Conservation, Department M.S.E.,University of Ghent, K.L. Ledeganckstraat 35, B-9000 Gent, Belgium
Received 19 June 1997; in revised form 19 August 1997; accepted 28 September 1997
Key words:Taxonomy, Rotifera,Floscularia, biogeography, autecology
Abstract
The taxonomy of some Neotropical representatives of the genusFlosculariaCuvier, 1798 (Rotifera: Monogononta)is discussed, based on SEM studies of trophi and external morphology.Floscularia armatan.sp. andF. bifida n.sp. are described, andF. conifera(Hudson, 1886) is restored at full species rank. A hypothesis on the functionalrelevance of differences in trophi morphology for niche segregation is formulated.
Introduction
In the most recent taxonomic key including floscu-lariid Rotifera (Koste, 1978), the genusFlosculariaCuvier, 1798 is listed as containing six valid morphos-pecies and one subspecies. Most of these have beendescribed from the northern hemisphere, and havesubsequently been recorded from almost all parts ofthe world. Exceptions areFloscularia noodtiKoste,1972, recorded only from the Neotropical region (seeKoste, 1978; De Ridder & Segers, 1997), andF. deco-ra Edmondson, 1940, recorded only from the northernhemisphere. This situation is symptomatic for Rotifera,as it reflects present-day taxonomy as commented uponby, amongst others, Dumont (1983) and Koste & Shiel(1989).
Poor taxonomic resolution has resulted in thehypothesis of potential cosmopolitanism for allRotifera (Ruttner-Koliskoin Dumont, 1980). It is nowclear that this view needs revision. Pejler (1977), DeRidder (1981), Dumont (1983) and Segers (1996a)have demonstrated a wide variety of distribution pat-terns in rotifers. A study on the genusScaridiumEhren-berg, 1830, a group with a highly characteristic generalbody size, revealed that detailed morphological studiesare required to distinguish morphotaxa with circum-scribed distributions (Segers, 1995; 1996b). The situa-tion in Floscularia is reminiscent ofScaridium. Here,
identification of relatively large animals relies mostlyon the structure of the tube they inhabit, and on someother, easily appreciable features; trophi morphologyhas never been considered. Therefore, it did not comeas a surprise to find several undescribedmorphospeciesof Flosculariain samples from a subtropical Brazilianreservoir, and to find that these are most reliably char-acterized by their trophi morphology, in addition to thefeatures considered to date.
In this contribution I report on the fourFlosculariamorphospecies that co-occurred in Broa reservoir (Iti-rapira, SP, Brazil) on 13 February 1996, and commenton the relevance of the differences in trophus structureamong the four taxa.
Material and methods
The material treated here was collected in Broa reser-voir, Itirapira, Sao Paulo, Brazil. Living specimenswere collected from submerged macrophytes and stud-ied at CRHEA, Universidadede Sao Paulo, Brazil. Pre-served animals contained in net samples (mesh width50�m) collected on 19 January or 13 February 1996and fixed in formaldehyde (4%) were studied using aWild M10 dissection microscope and a Olympus CH-2 microscope with drawing tube. Scanning electronmicroscopy (S.E.M.) was performed using a Jeol JSM
PIPS Nr.: 150556 BIO2KAPhy4115.tex; 11/05/1998; 10:03; v.7; p.1

166
Table 1. Summary of diagnostic characters in fourFlosculariaspecies
F. ringens F. decora F. armata F. bifida
(Linneus) (Hudson) n.sp. n.sp.
characters:
tube pellets round, brownish (with bullet-shaped, yellowish round, brownish (with round, brownish
detritus), densily packed to dark brown, densily detritus), loosely (with detritus),
packed packed closely packed
foot peduncle short short short long
anterior region smooth smooth with five humps smooth
(retracted)
dorsal projection two smoothly curved two abruptly separated, two variable two bifid hooks;
hooks; basal part small small hooks; basal part mucrones, basal part small
large occasionally with
accessory tubercules;
basal part large, with
three teeth
trophi unci teeth weakly unci teeth weakly unci teeth strongly unci teeth strongly
differentiated differentiated differentiated differentiated
length: 33–38�m length: 33–36�m length: 34–37�m length: 42–47�m
largest tooth diameter ca. largest tooth diameter ca. largest tooth diameter largest tooth
1.5�m 1.5�m ca. 3�m diameter 6–9�m
840 microscope on critical point-dried complete spec-imens or on trophi processed following a method afterSegers (1993) and Segers & Dumont (1993).
Repositories of type material are as follows:K.B.I.N.: royal Belgian Institute for Natural Sciences,Brussels, Belgium; I.N.P.A.: Instituto Nacional dePesquisas da Amazonia, Manaus, Brazil; R.U.G.: Uni-versity of Gent, Belgium; R.U.C.A.: Department ofBiology, F.E.P., University of Antwerp, Belgium (col-lection W.H. De Smet).
Results
Four representatives of the genusFloscularia wereencountered in Broa reservoir. These are as follows:
Floscularia ringens(Linneus, 1758)Figures 1–4, 9–11, 13, 40, 41
Serpula ringensLinneus, 1758Melicerta ringens(Linneus) Schrank, 1803Floscularia ringens(Linneus) Harring, 1913
Synonyms:Brachionus tubifexPallas, 1766;Tubico-laria tetrapetalaCuvier, 1817.
Differential diagnosis (Table 1):Floscularia ringens
can be distinguished from its congeners by the tubeconstructed of round, densely packed pellets contain-ing debris; dorsal projection in the neck region bearinga pair of smoothly curved hooks (Figures 2–3, 10–11)and trophus with weakly differentiated unci teeth (Fig-ures 4, 13, 41).
Measurements: Total length up to 1900�m, tube lengthup to 1600�m (Koste, 1978; Tiefenbacher, 1972: tubelength up to 2455�m), antenna length 38–50�m. Tro-phus length 33–38�m, width 55�m, uncus length18–20�m, width 18–20�m, largest tooth diameter1.5�m.
Distribution: F. ringensis the commonest species ofthe genus. It is reportedly cosmopolitan.
Floscularia conifera(Hudson, 1886)Figures 5–8, 12, 14, 38
Melicerta coniferaHudson, 1886Floscularia conifera(Hudson, 1886) Harring, 1913Floscularia ringens conifera: Koste, 1978
Differential diagnosis:Floscularia coniferais charac-terized by its tube being constructed of densely packed
hy4115.tex; 11/05/1998; 10:03; v.7; p.2

167
Figures 1–8. Figures 1–4:Floscularia ringens(Linneus). 1: habitus of contracted specimen, lateral, 2: apical region, lateral, 3: apical region,frontal, 4: trophus, lateral. Figures 5–8:Floscularia conifera(Hudson). 5: habitus of contracted specimen, lateral, 6: apical region, lateral, 7:apical region, frontal, 8: trophus, ventral.
bullet-shaped pellets without debris; dorsal projectionwith separated, downwardly pointed mucrones (Fig-ures 6–7, 12) and trophus with weakly differentiatedunci teeth (Figures 8, 14). The first two features easilydistinguishF. coniferafrom F. ringens.
Measurements: Total length up to 2000�m (Koste,1978; Edmondson, 1945: 3550�m), tube length up to1900�m, antenna length 40–44�m. Trophus length33–36�m, width 43–45�m, uncus length 15–18�m,width 6–18�m, largest tooth diameter 1.5�m.
Distribution: F. conifera is less common thanF. rin-gens, but has been recorded from all parts of the world.
Comments. This taxon is treated as a subspecies ofFloscularia ringensby Koste (1978). Considering thatthe two co-occur, this subspecific status cannot beretained. As the two are clearly morphologically dis-tinct, F. coniferais reinstated as a separate morphos-pecies.
hy4115.tex; 11/05/1998; 10:03; v.7; p.3

168
Fig
ure
s9–14.
Fig
ures
9–11
,13
:Flo
scula
riaringe
ns(
Lin
neus
).9:
apic
alre
gion
,la
tera
l,10
:po
ster
ior
proj
ectio
n,la
tera
l,11
:po
ster
ior
proj
ectio
nw
ithdo
rsal
ante
nna,
fron
tal1
3:tr
ophu
s,ve
ntra
l.F
igur
es12
,14:F
losc
ular
iaco
nife
ra(H
udso
n).1
2:po
ster
ior
proj
ectio
nan
dla
tera
lant
enna
e,la
tera
l,14
:tro
phus
,ven
tral
.S.E
.M.p
hoto
grap
hs,
alls
cale
bars
:5
�
m.
hy4115.tex; 11/05/1998; 10:03; v.7; p.4

169
Figures 15–24. Floscularia armatan. sp. 15: habitus of contracted specimen, lateral, 16: apical region, lateral, 17: apical region, frontal, 18–20:dorsal projection, lateral, 21–23: dorsal projection, ventral (different specimens), 24: trophus, ventral.
In a pond near Pirasununga (SP., Brazil, coll.27 January 1997), a colony of 14 specimens ofF.coniferawas found, onto which severalPtygura pilula(Cubitt) were attached.
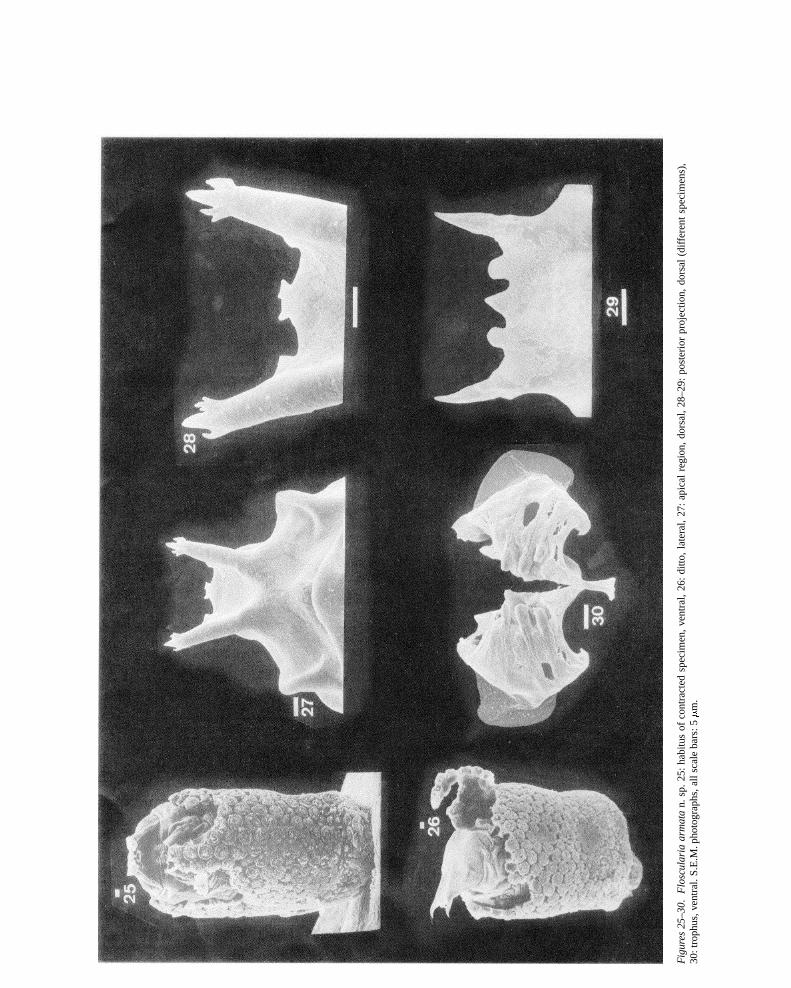
Floscularia armatanew speciesFigures 15–30, 39, 42
Holotype in K.B.I.N. (IG 28485 RIR 90)
Paratypes: two in K.B.I.N. (IG 28485 RIR 91–92),
one paratype in I.N.P.A. and R.U.C.A. each, three (onea trophus preparation) in R.U.G. All specimens fromthe type locality, Broa reservoir, Itirapira, Sao Paulo,Brazil.
Differential diagnosis:Floscularia armatan. sp. isunique by the shape of its dorsal projection and byits trophus having strongly differentiated unci teeth.It closely resemblesF. ringensand F. bifurca n. sp.by tube architecture. Its large dorsal projection bear-
hy4115.tex; 11/05/1998; 10:03; v.7; p.5
170
Fig
ure
s25–30.
Flo
scula
riaarm
ata
n.sp
.25
:ha
bitu
sof
cont
ract
edsp
ecim
en,
vent
ral,
26:
ditto
,la
tera
l,27
:ap
ical
regi
on,
dors
al,
28–2
9:po
ster
ior
proj
ectio
n,do
rsal
(diff
eren
tsp
ecim
ens)
,30
:tro
phus
,ven
tral
.S.E
.M.p
hoto
grap
hs,
alls
cale
bars
:5
�
m.
hy4115.tex; 11/05/1998; 10:03; v.7; p.6

171
Figures 31–34. Floscularia bifidan. sp. 31: habitus of contracted specimen, lateral, 32: apical region, ventral, 33: apical region, lateral, 34:trophus, ventral.
ing several accessory teeth is highly distinctive. Thetrophus teeth ofF. armatan. sp. are more strongly dif-ferentiated than those ofF. ringens, but the major teethare less robust and the trophus smaller than inF. bifida.
Description (of contracted specimens): Tube con-structed of loosely packed spherical pellets containingdebris. Body and foot elongate. Foot retractile (not ful-ly retracted in Figure 15), attachment peduncle short.
Head region with shallow lobes, a dorsal projectionbearing sharp mucrones and the dorsal antenna and apair of lateral antenna. Two pairs of shallow lobes, anda single dorsal lobe present (Figures 16–17). Dorsalprojection large, variable (Figures 18–23, 28–29) withtwo strong lateral spines, these slightly incurved ornearly straight and pointed ventrad terminally, smooth(Figure 29) or with accessory projections near the tips(e.g., Figure 28). Basis of the dorsal projection large,
hy4115.tex; 11/05/1998; 10:03; v.7; p.7

172
Fig
ure
s35–40.
Flo
scula
riabifi
da
n.sp
.35
:po
ster
ior
proj
ectio
n,do
rsal
,36
:po
ster
ior
proj
ectio
n,la
tera
l,37
:tr
ophu
s,ve
ntra
lvi
ew.
Fig
ures
38–4
0:tr
ophi
ofF
losc
ula
ria
spp.
,ve
ntra
l,m
agni
ficat
ion
asin
Fig
ure
37,3
8:F.co
nife
ra,3
9:F.
arm
ata
n.sp
.,40
:F.r
inge
ns.
S.E
.M.p
hoto
grap
hs,a
llsc
ale
bars
:5�
m.
hy4115.tex; 11/05/1998; 10:03; v.7; p.8

173
Figures 41–43. Flosculariaspp., detail of major unci teeth and rami, 41:F. ringens, 42:F. armatan.sp., 43:F. bifidan. sp. S.E.M. photographs,all scale bars: 5�m.
midventral anterior margin with a pair of lateral andan intermediate teeth, occasionally bearing denticles(Figure 28). Dorsal antenna situated ventrally on basisof dorsal projection (Figures 18, 19). Lateral antennae(Figure 17) elongate, situated slightly anterior to theretracted pellet-producing organ.
Trophus malleoramate (Figures 24, 30, 39). Unciasymmetrical, with large number of teeth. Teeth dif-ferentiated, the three proximal ones robust, the othersreduced. Second and third tooth strongest of right, resp.left uncus. Unci teeth webbed. Rami with serrated rein-forced ridge medially and proximally, opposed to therobust teeth (Figure 42).
Measurements: Tube length up to 1100�m. Body(contracted) 250–280�m; attachment peduncle 8–15 �m, dorsal projection 34–46�m, antenna length27–33 �m. Trophus length 34–37�m, width 44–59 �m, uncus length 19–20�m, width 19–20�m,maximum tooth diameter 2.9–3.0�m.
Etymology. The species name is an adjective, andrefers to the armature of the dorsal projection which is
the strongest of all congeners known to date.
Distribution.Floscularia armatan. sp. is only knownfrom the type locality, where it was the commonest ofall Floscularia.
Floscularia bifidanew speciesFigures 31–37, 43
Holotype in K.B.I.N. (IG 28485 RIR 87)
Paratypes: two (one a trophus preparation) in K.B.I.N.(IG 28485 RIR 88–89), three paratypes in I.N.P.A.,one in R.U.C.A., three (one a trophus preparation)in R.U.G. All specimens from the type locality, Broareservoir, Itirapira, Sao Paulo, Brazil.
Differential diagnosis:Floscularia bifidan. sp. is dis-tinct from F. armatan.sp. andF. ringensby the bifidmucrones on its dorsal projection and its large tro-phus, bearing strongly differentiated unci teeth. Its longattachment stalk is reminiscent ofF. pedunculata(Joli-et), but the latter has a differently shaped dorsal pro-
hy4115.tex; 11/05/1998; 10:03; v.7; p.9

174
jection with smooth, inwardly curved mucrones and asmall median projection (see Figure 33 in Edmondson,1940), and has characteristic quadridigitate cuticularprocesses laterally, near the basis of the antennae.
Description (of contracted specimens): Tube con-structed of loosely packed spherical pellets containingdebris. Body and foot elongate (note fully contract-ed in Figure 31). Foot retractile, attachment peduncleelongate. Corona relatively large. Head (Figures 32–33) smooth, with a dorsal projection bearing sharpmucrones and the dorsal antenna, and a pair of later-al antenna. Dorsal projection (Figure 32, 35–36) withtwo bifid, smoothly curved lateral spines, each withone anteriad and one inwardly pointed tooth. Basis ofthe dorsal projection weakly differentiated, midven-tral anterior margin straight. Dorsal antenna situatedventrally on basis of dorsal projection. Lateral anten-nae (Figure 32) elongate, situated slightly anterior tothe retracted pellet-producing organ, sensorial organretractile.
Trophus malleoramate (Figures 34, 37). Unciasymmetrical, with large number of teeth. Teethstrongly differentiated, the three proximal ones robust,the others reduced. Second tooth the strongest on rightuncus, third tooth strongest on left uncus. Unci teethwebbed. Rami with knobby projections medially andproximally, opposed to the robust teeth (Figure 43).
Measurements: Tube length up to 1400�m, Body(contracted) 242–370�m; attachment peduncle (64)129–300�m, dorsal projection 26–36�m. Trophuslength 42–47�m, width 52–82�m, uncus length 21–23 �m, width 28–29�m, maximum tooth diameter6.3–8.7�m.
Etymology. The species namebifida is an adjective,and refers to the bifid lateral spines of the dorsal pro-jection.
Distribution. Floscularia bifidan.sp. is only knownfrom the type locality.
Discussion
Of the characters classically used in the taxonomy ofFloscularia, the shape of the dorsal projection appearsespecially useful for the taxonomic differentiation ofmorphospecies. The function of this projection is notclear. I observed that while feeding,Flosculariaspec-
imens have their corona extended, and the projectionis covered. During contraction, the corona is retractedinside the body. The dorsal projection moves abruptlyfrom dorsal to ventral, with the spines pointed in thedirection of the movement. By this, a potential preda-tor will be struck by this fairly rigid projection, whichis probably sufficient to deter many. Hence, it seemslikely that the posterior protection acts as a defenceagainst predators.
To date, reports on trophus morphology inFloscu-laria have been poorly detailed (e.g., see Koste, 1978).Our comparative S.E.M. analysis reveals many inter-specific differences in trophus morphology, as illustrat-ed in Figures 37–40, whereas intraspecific variation isnegligible. The trophi ofF. ringens(Figure 40) andF.conifera(Figure 38) are similar with regards to size andstructure of the unci. InF. armatan.sp. (Figure 39),the trophus is more or less similar in size to that ofF. ringensand of F. conifera, but the unci teeth arestrongly differentiated: three pairs of proximal teethare enlarged, and are more robust than the correspond-ing teeth inF. ringensandF. conifera. The rest of theteeth appear reduced when compared to those ofF. rin-gensandF. conifera. The unci teeth ofF. bifidan. sp.(Figure 37) are even more strongly differentiated thanthose ofF. armatan. sp., while knobby projectionsopposed to the main unci teeth have developed on therami (Figure 43). Such rami projections are representedby a reinforced ridge inF. armata(Figure 42), and areonly indicated inF. ringens(Figure 41) andF. conifera.In addition, the trophus ofF. bifida is noticeably largerthan that of the other species.
Trophus structure is closely connected with feeding(e.g., Salt et al., 1978). It is likely that the observedinterspecific differences reflect differences in the typeof food eaten by different species. The malleoramatetrophi of Flosculariaceae act by grinding of the unciteeth against the rami, so macerating food particlescollected by the corona. It appears logical to assumethat the size and robustness of the functional teeth isrelated to the size and hardness of the food particles.I hypothesize thatF. ringensandF. coniferafeed onminute particles (see Tiefenbacher, 1972), whereasF.armatan. sp. feeds on larger and stiffer particles. Thelarge and robust trophi ofF. bifida n. sp. enable thistaxon to feed on particles of a size and sturdity that isbeyond the reach of its congeners. This differentiationmay be a key factor to explain niche segregation inthese species. It is the largest interspecific variabilityso far reported in any genus with malleoramate trophi.
hy4115.tex; 11/05/1998; 10:03; v.7; p.10

175
The discovery of two new species on a total of fourFlosculariaspecies in samples from a single artificial,subtropical reservoir indicate that much remains to bedone in this field. Hence, it is likely that many moreFloscularia await discovery, especially from tropicalregions. Also, the present views on biogeography ofFloscularia, and of sessile Rotifera in general, need tobe revised.
Acknowledgements
This study was supported by grants from N.F.W.O.(Belgium) and CNPq (Brazil). The staff of CRHEA(USP), especially Prof. Dr Maria do Carmo Calijuri(Director), Dr A. Rietzler and A. L. de Oliveiro-Netoare acknowledged for the hospitality during my studyvisit to Broa reservoir. Part of this work was done whilethe author was a post-doctoral research fellow at theAcademy of Natural Sciences of Philadelphia (John J.and Anna H. Gallagher fellowship). I thank R. J. Shieland R. L. Wallace for reviewing the manuscript of thispaper.
References
De Ridder, M., 1981. Some considerations on the geographical dis-tribution of rotifers. Hydrobiologia 85: 209–225.
De Ridder, M. & H. Segers, 1997. Rotifera Monogononta in sixzoogeographical regions after publications between 1960–1992.Studiedocumenten van het Koninklijk Belgisch Instituut voorNatuurwetenschappen (J. Van Goethem ed.) 87: 481 pp.
Dumont, H. J., 1980. Workshop on taxonomy and biogeography.Hydrobiologia 73: 205–206.
Dumont, H. J., 1983. Biogeography of rotifers. Hydrobiologia 104(Dev. Hydrobiol. 14): 19–30.
Edmondson, W. T., 1940. New species of Rotatoria, with notes onheterogonic growth. Trans. Am. Microsc. Soc. 58: 459–472.
Edmondson, W. T., 1945. Ecological studies of sessile Rotatoria, 2.Dynamics of populations and social structure. Ecol. Monogr. 15:141–172.
Koste, W., 178. Rotatoria. Die Radertiere Mitteleuropas. Born-traeger, Berlin, 2 vols: 673 pp., 234 plates.
Koste, W. & R. J. Shiel, 1989. Classical taxonomy and modernmethodology. Hydrobiologia 186/187: 279–284.
Pejler, B., 1977. On the global distribution of the family Brachion-idae. Arch. Hydrobiol./Suppl. 53: 255–306.
Salt, G. W., G. F. Sabbadini & M. L. Commins, 1978. Trophi mor-phology relative to food habits in six species of rotifers (Asplanch-nidae). Trans. am. Micr. Soc. 97: 469–485.
Segers, H., 1993. Rotifera of some lakes in the floodplain of the RiverNiger (Imo State, Nigeria). I. New species and other taxonomicconsiderations. Hydrobiologia 250: 39–61.
Segers, H., 1995. A reappraisal of the Scaridiidae (Rotifera: Mono-gononta). Zool. Scr. 24: 91–100.
Segers, H., 1996a. The biogeography of littoralLecaneRotifera.Hydrobiologia 323: 169–197.
Segers, H., 1996b.Scaridium elongatumn.sp., a new monogonontrotifer from Brazil. Belg. J. Zool. 126: 57–63.
Segers, H. & H. J. Dumont, 1993. Rotifera from Arabia, with descrip-tions of two new species. Fauna of Saudi-Arabia 13: 3–26.
Tiefenbacher, L., 1972. Beitrage zur Biologie undOkologie sessilerRotatorien unter besonderer Berucksichtigung des Gegausebauesund der Regeneration-fahigkeit. Arch. Hydrobiol. 71: 31–78.
hy4115.tex; 11/05/1998; 10:03; v.7; p.11
Related Documents











