Biological Psychology 92 (2013) 125–134 Contents lists available at SciVerse ScienceDirect Biological Psychology journa l h o me page: www.elsevier.com/locate/biopsycho Contingent capture can occur at specific feature values: Behavioral and electrophysiological evidence Jun Jiao a , Guang Zhao b , Qi Wang c , Kan Zhang a , Hong Li d , Hong-jin Sun e,∗∗ , Qiang Liu d,∗ a Key Laboratory of Behavioral Science, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China b Key Laboratory of Cognition and Personality, Ministry of Education, Southwest University, Chongqing 400715, China c Department of Psychology, Tufts University, 490 Boston Ave., Medford, MA 02155, USA d Research Center of Psychological Development and Education, Liaoning Normal University, Dalian 116029, China e Department of Psychology, Neuroscience and Behaviour, McMaster University, Hamilton, Ontario, L8S 4K1, Canada a r t i c l e i n f o Article history: Received 20 December 2011 Accepted 16 September 2012 Available online 13 October 2012 Keywords: Attentional capture Top–down control Specific feature value N2pc effect a b s t r a c t The notion that attentional top–down control can be tuned to a stimulus feature is widely accepted. Although previous studies suggested that the stimulus-driven attentional capture could be contingent on top–down attentional control settings, it was uncertain whether contingent capture can occur at a specific feature value. Three experiments were conducted to address this issue using both behavioral and ERPs measures. Participants were required to respond to one color singleton in the search display (target) but refrain from responding to the search display containing another color singleton (nontarget). When target and nontarget belonged to different color categories (Experiment 1), only the target-color cue and within category irrelevant-color cue elicited the significant cue validity effect (i.e. RTs were shorter when the target was presented at the same location as the preceding cue rather than at a different location); they also lead to a robust N2pc effect, indicative of attention-capture. In addition, these two cue types had similar attention-capturing capacity. However, when target and nontarget belonged to the same color category (Experiments 2 and 3), only the target-color cue elicited the significant cue validity effect and the robust N2pc effect. The same within category irrelevant-color cue no longer elicited the cue validity effect, and the N2pc effect was also attenuated. Present findings suggest that contingent capture can occur at a specific feature value. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Over the past two decades, there have been heated debates concerning the extent to which attentional capture is controlled either by certain stimulus properties in an involuntary, bottom- up fashion or by behavioral goals of the observer in a voluntary, top–down fashion. It has been argued that perceptually salient stimuli can automatically capture attention irrespective of the observer’s intentions or goals. For instance, Theeuwes (1992; see also 1991, 1994), using the additional singleton paradigm – in which participants searched for a singleton target among several dis- played items (e.g., a green circle among all green diamonds), found that the presence of a task-irrelevant singleton (e.g., a red diamond) ∗ Corresponding author at: Research Center of Psychological Development and Education, Liaoning Normal University, Dalian 116029, China. Tel.: +86 13332220573. ∗∗ Corresponding author. Tel.: +1 905 5259140x24367; fax: +1 905 529 6225. E-mail addresses: [email protected] (H.-j. Sun), [email protected] (Q. Liu). among displayed items resulted in slowed responses. He there- fore suggested the “pure-capture hypothesis”, which stated that attentional capture is determined by bottom-up salience of the stimulus, irrespective of whether or not the observer is actually looking for it. However, Folk et al. (1992), using a spatial cuing paradigm – in which the target display was preceded by a cue dis- play, found that the spatially uninformative color singleton cue produced a cue validity effect, i.e. reaction times (RTs) were faster for targets at cued versus uncued locations (evidence of attentional capture), in blocks where target itself was also a color singleton but not an onset singleton. Similarly, the spatially nonpredictive onset singleton cue produced the cue validity effect in blocks where target itself was also the onset singleton but not the color single- ton. On the basis of such findings, they proposed the “contingent capture hypothesis”, which stated that salient stimuli automati- cally capture an observer’s attention only when their properties match the top–down control settings. The classic findings of Folk et al. (1992) have been replicated many times in subsequent exper- iments (Al-Aidroos et al., 2010; Folk and Remington, 1999; Folk et al., 1994; Gibson and Amelio, 2000; Gibson and Kelsey, 1998; Lien et al., 2010a). 0301-0511/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.biopsycho.2012.09.010

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ce
Ja
b
c
d
e
a
ARAA
KATSN
1
ceutsoappt
ET
(
0h
Biological Psychology 92 (2013) 125– 134
Contents lists available at SciVerse ScienceDirect
Biological Psychology
journa l h o me page: www.elsev ier .com/ locate /b iopsycho
ontingent capture can occur at specific feature values: Behavioral andlectrophysiological evidence
un Jiaoa, Guang Zhaob, Qi Wangc, Kan Zhanga, Hong Lid, Hong-jin Sune,∗∗, Qiang Liud,∗
Key Laboratory of Behavioral Science, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, ChinaKey Laboratory of Cognition and Personality, Ministry of Education, Southwest University, Chongqing 400715, ChinaDepartment of Psychology, Tufts University, 490 Boston Ave., Medford, MA 02155, USAResearch Center of Psychological Development and Education, Liaoning Normal University, Dalian 116029, ChinaDepartment of Psychology, Neuroscience and Behaviour, McMaster University, Hamilton, Ontario, L8S 4K1, Canada
r t i c l e i n f o
rticle history:eceived 20 December 2011ccepted 16 September 2012vailable online 13 October 2012
eywords:ttentional captureop–down controlpecific feature value2pc effect
a b s t r a c t
The notion that attentional top–down control can be tuned to a stimulus feature is widely accepted.Although previous studies suggested that the stimulus-driven attentional capture could be contingenton top–down attentional control settings, it was uncertain whether contingent capture can occur at aspecific feature value. Three experiments were conducted to address this issue using both behavioral andERPs measures. Participants were required to respond to one color singleton in the search display (target)but refrain from responding to the search display containing another color singleton (nontarget). Whentarget and nontarget belonged to different color categories (Experiment 1), only the target-color cue andwithin category irrelevant-color cue elicited the significant cue validity effect (i.e. RTs were shorter whenthe target was presented at the same location as the preceding cue rather than at a different location);
they also lead to a robust N2pc effect, indicative of attention-capture. In addition, these two cue types hadsimilar attention-capturing capacity. However, when target and nontarget belonged to the same colorcategory (Experiments 2 and 3), only the target-color cue elicited the significant cue validity effect andthe robust N2pc effect. The same within category irrelevant-color cue no longer elicited the cue validityeffect, and the N2pc effect was also attenuated. Present findings suggest that contingent capture canoccur at a specific feature value.. Introduction
Over the past two decades, there have been heated debatesoncerning the extent to which attentional capture is controlledither by certain stimulus properties in an involuntary, bottom-p fashion or by behavioral goals of the observer in a voluntary,op–down fashion. It has been argued that perceptually salienttimuli can automatically capture attention irrespective of thebserver’s intentions or goals. For instance, Theeuwes (1992; seelso 1991, 1994), using the additional singleton paradigm – in which
articipants searched for a singleton target among several dis-layed items (e.g., a green circle among all green diamonds), foundhat the presence of a task-irrelevant singleton (e.g., a red diamond)∗ Corresponding author at: Research Center of Psychological Development andducation, Liaoning Normal University, Dalian 116029, China.el.: +86 13332220573.∗∗ Corresponding author. Tel.: +1 905 5259140x24367; fax: +1 905 529 6225.
E-mail addresses: [email protected] (H.-j. Sun), [email protected]. Liu).
301-0511/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.biopsycho.2012.09.010
© 2012 Elsevier B.V. All rights reserved.
among displayed items resulted in slowed responses. He there-fore suggested the “pure-capture hypothesis”, which stated thatattentional capture is determined by bottom-up salience of thestimulus, irrespective of whether or not the observer is actuallylooking for it. However, Folk et al. (1992), using a spatial cuingparadigm – in which the target display was preceded by a cue dis-play, found that the spatially uninformative color singleton cueproduced a cue validity effect, i.e. reaction times (RTs) were fasterfor targets at cued versus uncued locations (evidence of attentionalcapture), in blocks where target itself was also a color singletonbut not an onset singleton. Similarly, the spatially nonpredictiveonset singleton cue produced the cue validity effect in blocks wheretarget itself was also the onset singleton but not the color single-ton. On the basis of such findings, they proposed the “contingentcapture hypothesis”, which stated that salient stimuli automati-cally capture an observer’s attention only when their propertiesmatch the top–down control settings. The classic findings of Folk
et al. (1992) have been replicated many times in subsequent exper-iments (Al-Aidroos et al., 2010; Folk and Remington, 1999; Folket al., 1994; Gibson and Amelio, 2000; Gibson and Kelsey, 1998;Lien et al., 2010a).
1 sychol
ccms((tdiwgclpsctTvwt
stcfte(r(tcwsiim
a2matdtt(Mp
csteettuv
vbti
26 J. Jiao et al. / Biological P
Bacon and Egeth (1994) offered an explanation for theontroversy between the pure-capture hypothesis and theontingent-capture hypothesis. They argued that specific searchode adopted by the observer is what determines whether or not
alient stimuli captured attention. Specifically, in the Theeuwes1992) study, the target was a singleton in a given dimensione.g., a circle among diamonds), and participants might chooseo adopt the singleton-detection mode, in which singletons in allimensions would capture attention. Thus, the irrelevant singleton
ncreased response time to the target. While, when participantsere required to search for a specific feature that was not a sin-
leton, they were forced to use the feature search mode. As aonsequence, the presence of the same irrelevant singleton noonger captured attention and affected search performance. In sup-ort of this claim, Bacon and Egeth (1994), using the additionalingleton paradigm, showed that interference from an irrelevantolor singleton was observed when the target was a shape single-on (participants adopted singleton-detection mode), replicatingheeuwes’s finding. In contrast, no such distracting effect of theery same irrelevant color singleton was found when shape targetsere not unique in the shape dimension (participants were forced
o adopt feature-search mode).Combing behavioral and electrophysiological approach, a recent
tudy by Eimer and Kiss (2010) provided compelling evidence forhe existence of two search modes. They found that when twoolor singletons were defined as targets (Experiment 1, e.g., searchor red and green singleton targets), both the target-color single-on cue and the irrelevant-color singleton cue elicited cue validityffects and an N2pc (short for N2-posterior-contralateral) effectindicating attentional capture). However, when participants wereequired to refrain from responding to one type of color singletonsExperiment 2, e.g., ignore green singletons while attending to redargets), the cue validity effect was only observed for target-colorue condition (e.g., red color singleton cue); no cue validity effectsere seen for the nontarget-color cue condition (e.g., green color
ingleton cue) and the irrelevant-color cue condition. These find-ngs indicated that participants adopted the singleton search moden Experiment 1 and the feature-specific search mode in Experi-
ent 2.The study by Eimer and Kiss (2010) and also others (e.g., Eimer
nd Kiss, 2008; Eimer et al., 2009; Lamy and Egeth, 2003; Lien et al.,010b) showed that attentional capture depends critically on theatch between the stimulus feature and participants’ top–down
ttentional set. However, it is not clear how precise the match needso be for attentional capture to take place. For example, if the target-efining feature was “redness”, could all shades of “redness” cuehat matched the target on a broad color category capture atten-ion or would only the cue with one particular level of “redness”one which exactly matched the target’s shade) capture attention?
ore importantly, under what conditions would these effects takelace?
A study by Ansorge and Heumann (2003), also using the spatialuing paradigm, showed that when participants were instructed toearch for one abrupt onset target with specific color (e.g., green),he cue which had a different feature value but the same color cat-gory as the target (e.g., bluish green) triggered a stronger cuingffect than the cue from a different color category than that of thearget (e.g., yellowish red). Based on these results we might arguehat contingent capture could occur at a category level. But it is stillnclear whether contingent capture can occur at a specific featurealue.
It has been suggested by Navalpakkam and Itti (2006), using a
isual search paradigm, that attentional top–down task sets cane finely tuned to the specific feature value. They instructed par-icipants to search for a known single interval target among threentervals (LOW, MID, and HIGH intervals) distracters. The resultsogy 92 (2013) 125– 134
showed that participants could selectively saccade to relevantinterval in contrast to irrelevant intervals.
A recent study by Kiss and Eimer (2011) examined contingentcapture that occurs at size dimension. They found that small or largesize singleton cues triggered cue validity effects and N2pc compo-nents only when these cues matched the current target definingfeature. Specifically, they found that when participants’ task was torespond to small targets, small cues elicited the N2pc and behav-ioral spatial cue validity effects, but not for large cues; while, whenparticipants were instructed to search for large targets, the N2pcand behavioral spatial cue validity effects were observed for largecues, but not for small cues. In their study, search displays containedsix gray bars and the target bar was either smaller or larger than theother five bars, while cue displays contained six gray items com-posed of four dots and one item was either smaller or larger thanthe others. Because shapes of stimulus items in the cue and searchdisplays were not the same, the size of stimulus items in the cueand search displays could not be easily compared. Instead, the sizeinformation of stimulus items in cue and search displays was avail-able in relative terms (in contrast to other stimulus elements in thesame display). Therefore, it is reasonable to argue that participantscould categorically discriminate relative small vs. large, rather thanpay attention to the absolute value of the size. Thus, in their study,singleton items in cue displays and target displays could match onthe category, namely relatively small or large size category, ratherthan on the exact value of size dimension. Based on their findings,strictly speaking, it is safer to conclude that contingent capture canoccur at a broad size category rather than feature value.
In summary, whether contingent capture can occur at the spe-cific feature value remains inconclusive. The present study wasdesigned to shed additional light on this issue, by focusing on colorattribute of the stimulus where both the color category and shade ofcolor could be part of the attentional task set. It has been well docu-mented that color perception exhibits characteristics of categoricalperception (Kay and Kempton, 1984). This has been demonstratedby faster or more accurate performance on between-category dis-crimination (e.g. comparing green to blue) than on within-categorydiscrimination (e.g. comparing two different shades of green). Inthe context of color, the hierarchy of a category (e.g., green) anda specific feature value within a category (e.g., a shade of green)has already been established. Moreover, by having the color cate-gory and/or the shade of color match exactly between the cue andtarget/nontarget, our stimulus allows a direct comparison of theabsolute value of the color feature between cue and search dis-plays. Thus, this approach offered a direct test of whether, and inwhat condition, attentional capture can be extended from featurecategory to specific value of the features.
We employed a similar spatial cuing paradigm used by Eimerand Kiss (2010, Experiment 2). Similar to Eimer and Kiss (2010),we also combined behavioral and electrophysiological measures ofattentional capture. We expected to obtain the N2pc component inthe ERP waveform. The N2pc is an ERP component characterizedby an increased negativity over posterior scalp contralateral to anattended stimulus observed between 180 and 300 ms after displayonset, and is assumed to reflect the allocation of attention to anobject in a visual display (Eimer, 1996; Hickey et al., 2006; Jolicoeuret al., 2006; Luck and Hillyard, 1994; Mazza et al., 2007).
For cue displays, we employed four different types of colorsingleton cue. Three (target-color cue, irrelevant-color cue,nontarget-color cue) of them had been used in Experiment 2 inEimer and Kiss (2010). The irrelevant-color cue in our study was across-category irrelevant-color cue. The fourth one was a within-
category irrelevant-color cue with the same color category (but adifferent RGB value) as that of the target.In addition to cue types, we also manipulated target andnon-target stimulus properties. In Experiment 1, color categories
sychology 92 (2013) 125– 134 127
o2stNcvwoawwi
2
2
2
mtw
2
aTdfivdtwTfpwc21brh(F
2
ddatw
ctwooIbtbgktt
twsw
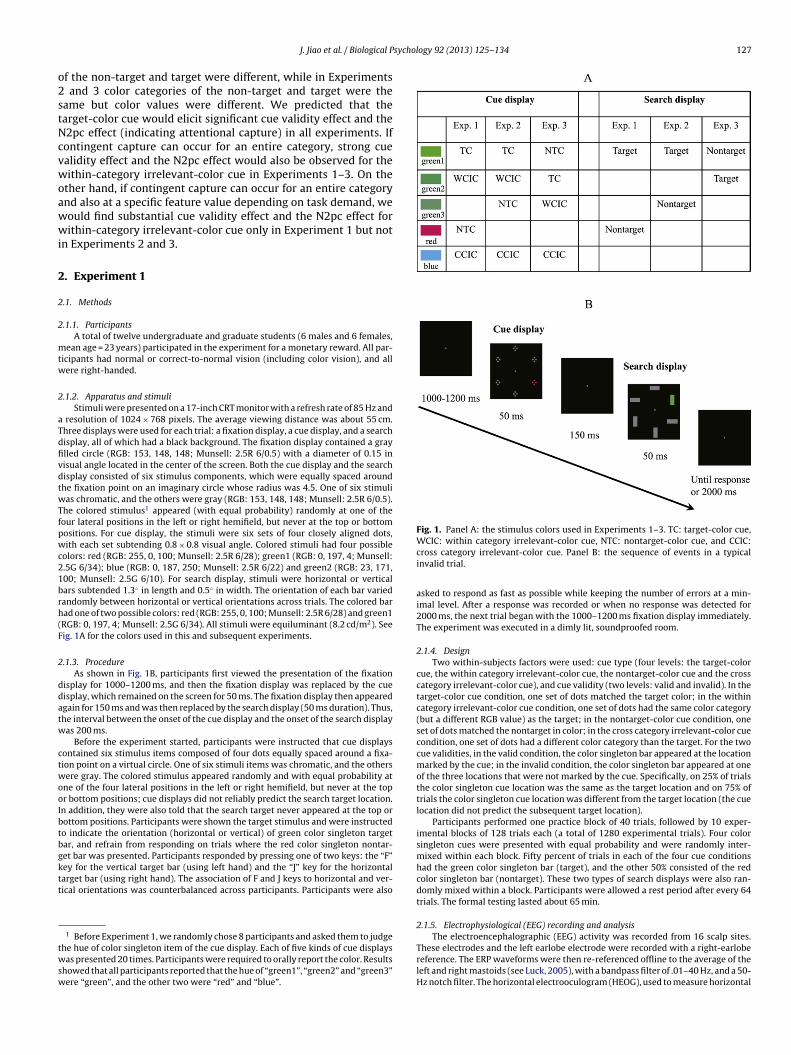
Fig. 1. Panel A: the stimulus colors used in Experiments 1–3. TC: target-color cue,
J. Jiao et al. / Biological P
f the non-target and target were different, while in Experiments and 3 color categories of the non-target and target were theame but color values were different. We predicted that thearget-color cue would elicit significant cue validity effect and the2pc effect (indicating attentional capture) in all experiments. Ifontingent capture can occur for an entire category, strong cuealidity effect and the N2pc effect would also be observed for theithin-category irrelevant-color cue in Experiments 1–3. On the
ther hand, if contingent capture can occur for an entire categorynd also at a specific feature value depending on task demand, weould find substantial cue validity effect and the N2pc effect forithin-category irrelevant-color cue only in Experiment 1 but not
n Experiments 2 and 3.
. Experiment 1
.1. Methods
.1.1. ParticipantsA total of twelve undergraduate and graduate students (6 males and 6 females,
ean age = 23 years) participated in the experiment for a monetary reward. All par-icipants had normal or correct-to-normal vision (including color vision), and allere right-handed.
.1.2. Apparatus and stimuliStimuli were presented on a 17-inch CRT monitor with a refresh rate of 85 Hz and
resolution of 1024 × 768 pixels. The average viewing distance was about 55 cm.hree displays were used for each trial: a fixation display, a cue display, and a searchisplay, all of which had a black background. The fixation display contained a graylled circle (RGB: 153, 148, 148; Munsell: 2.5R 6/0.5) with a diameter of 0.15 inisual angle located in the center of the screen. Both the cue display and the searchisplay consisted of six stimulus components, which were equally spaced aroundhe fixation point on an imaginary circle whose radius was 4.5. One of six stimulias chromatic, and the others were gray (RGB: 153, 148, 148; Munsell: 2.5R 6/0.5).
he colored stimulus1 appeared (with equal probability) randomly at one of theour lateral positions in the left or right hemifield, but never at the top or bottomositions. For cue display, the stimuli were six sets of four closely aligned dots,ith each set subtending 0.8 × 0.8 visual angle. Colored stimuli had four possible
olors: red (RGB: 255, 0, 100; Munsell: 2.5R 6/28); green1 (RGB: 0, 197, 4; Munsell:.5G 6/34); blue (RGB: 0, 187, 250; Munsell: 2.5R 6/22) and green2 (RGB: 23, 171,00; Munsell: 2.5G 6/10). For search display, stimuli were horizontal or verticalars subtended 1.3◦ in length and 0.5◦ in width. The orientation of each bar variedandomly between horizontal or vertical orientations across trials. The colored barad one of two possible colors: red (RGB: 255, 0, 100; Munsell: 2.5R 6/28) and green1RGB: 0, 197, 4; Munsell: 2.5G 6/34). All stimuli were equiluminant (8.2 cd/m2). Seeig. 1A for the colors used in this and subsequent experiments.
.1.3. ProcedureAs shown in Fig. 1B, participants first viewed the presentation of the fixation
isplay for 1000–1200 ms, and then the fixation display was replaced by the cueisplay, which remained on the screen for 50 ms. The fixation display then appearedgain for 150 ms and was then replaced by the search display (50 ms duration). Thus,he interval between the onset of the cue display and the onset of the search displayas 200 ms.
Before the experiment started, participants were instructed that cue displaysontained six stimulus items composed of four dots equally spaced around a fixa-ion point on a virtual circle. One of six stimuli items was chromatic, and the othersere gray. The colored stimulus appeared randomly and with equal probability at
ne of the four lateral positions in the left or right hemifield, but never at the topr bottom positions; cue displays did not reliably predict the search target location.n addition, they were also told that the search target never appeared at the top orottom positions. Participants were shown the target stimulus and were instructedo indicate the orientation (horizontal or vertical) of green color singleton targetar, and refrain from responding on trials where the red color singleton nontar-
et bar was presented. Participants responded by pressing one of two keys: the “F”ey for the vertical target bar (using left hand) and the “J” key for the horizontalarget bar (using right hand). The association of F and J keys to horizontal and ver-ical orientations was counterbalanced across participants. Participants were also1 Before Experiment 1, we randomly chose 8 participants and asked them to judgehe hue of color singleton item of the cue display. Each of five kinds of cue displaysas presented 20 times. Participants were required to orally report the color. Results
howed that all participants reported that the hue of “green1”, “green2” and “green3”ere “green”, and the other two were “red” and “blue”.
WCIC: within category irrelevant-color cue, NTC: nontarget-color cue, and CCIC:cross category irrelevant-color cue. Panel B: the sequence of events in a typicalinvalid trial.
asked to respond as fast as possible while keeping the number of errors at a min-imal level. After a response was recorded or when no response was detected for2000 ms, the next trial began with the 1000–1200 ms fixation display immediately.The experiment was executed in a dimly lit, soundproofed room.
2.1.4. DesignTwo within-subjects factors were used: cue type (four levels: the target-color
cue, the within category irrelevant-color cue, the nontarget-color cue and the crosscategory irrelevant-color cue), and cue validity (two levels: valid and invalid). In thetarget-color cue condition, one set of dots matched the target color; in the withincategory irrelevant-color cue condition, one set of dots had the same color category(but a different RGB value) as the target; in the nontarget-color cue condition, oneset of dots matched the nontarget in color; in the cross category irrelevant-color cuecondition, one set of dots had a different color category than the target. For the twocue validities, in the valid condition, the color singleton bar appeared at the locationmarked by the cue; in the invalid condition, the color singleton bar appeared at oneof the three locations that were not marked by the cue. Specifically, on 25% of trialsthe color singleton cue location was the same as the target location and on 75% oftrials the color singleton cue location was different from the target location (the cuelocation did not predict the subsequent target location).
Participants performed one practice block of 40 trials, followed by 10 exper-imental blocks of 128 trials each (a total of 1280 experimental trials). Four colorsingleton cues were presented with equal probability and were randomly inter-mixed within each block. Fifty percent of trials in each of the four cue conditionshad the green color singleton bar (target), and the other 50% consisted of the redcolor singleton bar (nontarget). These two types of search displays were also ran-domly mixed within a block. Participants were allowed a rest period after every 64trials. The formal testing lasted about 65 min.
2.1.5. Electrophysiological (EEG) recording and analysis
The electroencephalographic (EEG) activity was recorded from 16 scalp sites.These electrodes and the left earlobe electrode were recorded with a right-earlobereference. The ERP waveforms were then re-referenced offline to the average of theleft and right mastoids (see Luck, 2005), with a bandpass filter of .01–40 Hz, and a 50-Hz notch filter. The horizontal electrooculogram (HEOG), used to measure horizontal

128 J. Jiao et al. / Biological Psychology 92 (2013) 125– 134
Experiment 1 Experiment 2 Experiment 3
25
30
750
800
Erro
r(%
RT
(ms)
15
20
650
700
%)
)
5
10
550
600
C diti
0500
TC WCIC NT C CCIC TC WCIC NTC CCIC TC WCI C NTC CCIC
RT Error s
I lid
Cue con dition
Invali d
Vali d
F tion ofo : nont
eefCaoetrc
tialotnatafct
2
2
cnctp
nvcpyMtp6wn
ig. 2. Mean reaction times (RTs; line graphs) and error rates (bar graphs) as a funcf the means. TC: target-color cue, WCIC: within category irrelevant-color cue, NTC
ye movements, was bipolarly recorded from electrodes at the outer canthi of bothyes. The vertical electrooculogram (VEOG), used to detect eye blinks, was recordedrom electrodes above and below the right eye. Data were sampled at 500 Hz usingogNeuro amplifiers (PSYCHE-ARK, Inc.). All impedances were kept below 5 k�. Tri-ls with horizontal eye movements (horizontal EOG amplitude exceeding ±25 �V),r eye blinks (vertical EOG amplitude exceeding ±100 �V), and other artifacts (anylectrode exceeding ±80 �V) were excluded from analysis. An average of 8.2% ofrials were rejected because of artifacts (range = 0.6–20.5%). Trials with incorrectesponses and with RTs to targets that exceeded the mean RT for this specific trialondition by more than 3 SDs were excluded from all analyses.
ERPs response to cue displays were epoched into 400 ms periods from −100 mso 300 ms relative to cue onset for each cue type (target-color cue; within categoryrrelevant-color cue; nontarget-color cue and cross category irrelevant-color cue)nd cue position (left vs. right hemifield) collapsed across all four possible targetocations. Data were analyzed using the PO7 and PO8 electrodes only. To quantify theverall magnitude of the N2pc effect, we focused on the time window from 150 mso 250 ms after cue onset, since this is the time range during which the N2pc compo-ent is typically detected. For the PO7 and PO8 electrodes, the waveform observedt the left-sided electrode when cue was presented on the right was averaged withhe waveform for the right-sided electrode when cue was on the left, yielding theverage waveform contralateral to the cue. We also computed the average wave-orm ipsilateral to the cue at lateral posterior electrodes PO7 and PO8. Finally, weomputed the N2pc difference wave by subtracting the ipsilateral waveform fromhe contralateral waveform.
.2. Results
.2.1. Behavioral dataThe behavioral data were analyzed as a function of cue type
ondition (target-color cue; within category irrelevant-color cue;ontarget-color cue and cross category irrelevant-color cue) andue validity (valid and invalid). Mean RTs (line graphs) on correctrials and error rates (bar graphs) for each of these conditions wereresented in Fig. 2 (left panel).
RT analyses showed that the main effect of cue type was sig-ificant [F(3, 33) = 6.71, p < .01], and so was the main effect of cuealidity [F(1, 11) = 8.63, p < .05]. In addition, the interaction betweenue type and cue validity was also significant [F(3, 33) = 63.38,
< .001]. To further explore this interaction, simple effects anal-ses of cue validity were conducted on each cue type condition.ean correct RTs were significantly faster on valid than on invalid
rials for the target-color cue [661 vs. 698 ms, F(1, 11) = 69.72,
< .001] and within the category irrelevant-color cue [653 vs.82 ms, F(1, 11) = 61.50, p < .001]. In addition, mean correct RTsere significantly slower on valid than on invalid trials for theontarget-color cue [709 vs. 698 ms, F(1, 11) = 5.20, p < .05] and thecue validity and cue type in Experiments 1–3. Error bars represent standard errorsarget-color cue, and CCIC: cross category irrelevant-color cue.
cross category irrelevant-color cue [688 vs. 672 ms, F(1, 11) = 11.54,p < .01]. In addition, we also compared the cue validity effect for thetarget-color cue (37 ms) with the cue validity effect for the withincategory irrelevant-color cue (29 ms). There was a tendency towarda larger cue validity effect for the target-color cue compared tothe within category irrelevant-color cue, but this difference did notapproach significance [t(11) = 1.72, p > .05].
Error rates were generally low. ANOVA revealed no significanteffects or interactions.
2.2.2. ERP dataFig. 3A shows grand-average ERP waveforms elicited in the
300 ms post-cue interval at electrodes PO7/8 contralateral andipsilateral to color singleton cue location, and Fig. 3B shows themean difference amplitudes obtained by subtracting event-relatedpotential (ERP) waveforms at ipsilateral posterior electrode fromthat recorded at contralateral site, separately for trials with thetarget-color cue, the within category irrelevant-color cue, thenontarget-color cue and the cross category irrelevant-color cue.The analysis of mean amplitudes of the N2pc component wascarried out across a time window of 150–250 ms after the onsetof the cue display. A 4 (cue type: target-color, within categoryirrelevant-color, nontarget-color, and cross category irrelevant-color cue) × 2 (electrode laterality: contralateral and ipsilateral)repeated-measure ANOVA was performed. There was a reli-able main effect of laterality [F(1, 11) = 19.81, p < .01], indicatingthe presence of the N2pc for color singleton cues. The interac-tion between cue type and laterality was also significant [F(3,33) = 25.58, p < .001]. To further explore this interaction, we sepa-rately analyzed pairwise combinations of two cue conditions, withcue type now a two-level factor. When ERPs in response to thetarget-color cue and the within category irrelevant-color cue wereanalyzed together, the interaction between cue type and lateral-ity was not significant [F(1, 11) = .47, p > .05], reflecting that N2pcamplitudes did not differ significantly between these two cue typeconditions. Similarly, when ERPs to the nontarget-color cue andthe cross category irrelevant-color cue were analyzed together, theinteraction between cue type and laterality was also not significant
[F(1, 11) = .82, p > .05]. However, a significant interaction was foundfor the other four pairwise combinations of two cue conditions, thatis, the “target-color cue vs. nontarget-color cue”, the “target-colorcue vs. cross category irrelevant-color cue”, the “within category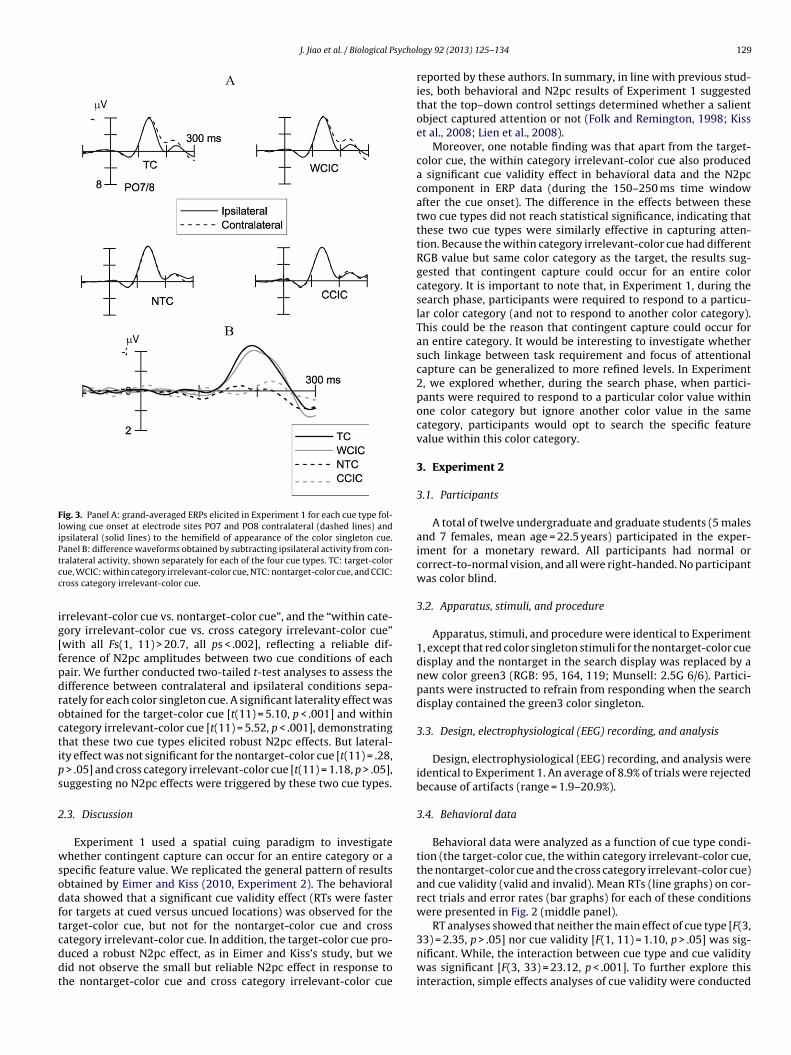
J. Jiao et al. / Biological Psychol
Fig. 3. Panel A: grand-averaged ERPs elicited in Experiment 1 for each cue type fol-lowing cue onset at electrode sites PO7 and PO8 contralateral (dashed lines) andipsilateral (solid lines) to the hemifield of appearance of the color singleton cue.Panel B: difference waveforms obtained by subtracting ipsilateral activity from con-tcc
ig[fpdroctips
2
wsodftcddt
ralateral activity, shown separately for each of the four cue types. TC: target-colorue, WCIC: within category irrelevant-color cue, NTC: nontarget-color cue, and CCIC:ross category irrelevant-color cue.
rrelevant-color cue vs. nontarget-color cue”, and the “within cate-ory irrelevant-color cue vs. cross category irrelevant-color cue”with all Fs(1, 11) > 20.7, all ps < .002], reflecting a reliable dif-erence of N2pc amplitudes between two cue conditions of eachair. We further conducted two-tailed t-test analyses to assess theifference between contralateral and ipsilateral conditions sepa-ately for each color singleton cue. A significant laterality effect wasbtained for the target-color cue [t(11) = 5.10, p < .001] and withinategory irrelevant-color cue [t(11) = 5.52, p < .001], demonstratinghat these two cue types elicited robust N2pc effects. But lateral-ty effect was not significant for the nontarget-color cue [t(11) = .28,
> .05] and cross category irrelevant-color cue [t(11) = 1.18, p > .05],uggesting no N2pc effects were triggered by these two cue types.
.3. Discussion
Experiment 1 used a spatial cuing paradigm to investigatehether contingent capture can occur for an entire category or a
pecific feature value. We replicated the general pattern of resultsbtained by Eimer and Kiss (2010, Experiment 2). The behavioralata showed that a significant cue validity effect (RTs were fasteror targets at cued versus uncued locations) was observed for thearget-color cue, but not for the nontarget-color cue and cross
ategory irrelevant-color cue. In addition, the target-color cue pro-uced a robust N2pc effect, as in Eimer and Kiss’s study, but weid not observe the small but reliable N2pc effect in response tohe nontarget-color cue and cross category irrelevant-color cueogy 92 (2013) 125– 134 129
reported by these authors. In summary, in line with previous stud-ies, both behavioral and N2pc results of Experiment 1 suggestedthat the top–down control settings determined whether a salientobject captured attention or not (Folk and Remington, 1998; Kisset al., 2008; Lien et al., 2008).
Moreover, one notable finding was that apart from the target-color cue, the within category irrelevant-color cue also produceda significant cue validity effect in behavioral data and the N2pccomponent in ERP data (during the 150–250 ms time windowafter the cue onset). The difference in the effects between thesetwo cue types did not reach statistical significance, indicating thatthese two cue types were similarly effective in capturing atten-tion. Because the within category irrelevant-color cue had differentRGB value but same color category as the target, the results sug-gested that contingent capture could occur for an entire colorcategory. It is important to note that, in Experiment 1, during thesearch phase, participants were required to respond to a particu-lar color category (and not to respond to another color category).This could be the reason that contingent capture could occur foran entire category. It would be interesting to investigate whethersuch linkage between task requirement and focus of attentionalcapture can be generalized to more refined levels. In Experiment2, we explored whether, during the search phase, when partici-pants were required to respond to a particular color value withinone color category but ignore another color value in the samecategory, participants would opt to search the specific featurevalue within this color category.
3. Experiment 2
3.1. Participants
A total of twelve undergraduate and graduate students (5 malesand 7 females, mean age = 22.5 years) participated in the exper-iment for a monetary reward. All participants had normal orcorrect-to-normal vision, and all were right-handed. No participantwas color blind.
3.2. Apparatus, stimuli, and procedure
Apparatus, stimuli, and procedure were identical to Experiment1, except that red color singleton stimuli for the nontarget-color cuedisplay and the nontarget in the search display was replaced by anew color green3 (RGB: 95, 164, 119; Munsell: 2.5G 6/6). Partici-pants were instructed to refrain from responding when the searchdisplay contained the green3 color singleton.
3.3. Design, electrophysiological (EEG) recording, and analysis
Design, electrophysiological (EEG) recording, and analysis wereidentical to Experiment 1. An average of 8.9% of trials were rejectedbecause of artifacts (range = 1.9–20.9%).
3.4. Behavioral data
Behavioral data were analyzed as a function of cue type condi-tion (the target-color cue, the within category irrelevant-color cue,the nontarget-color cue and the cross category irrelevant-color cue)and cue validity (valid and invalid). Mean RTs (line graphs) on cor-rect trials and error rates (bar graphs) for each of these conditionswere presented in Fig. 2 (middle panel).
RT analyses showed that neither the main effect of cue type [F(3,
33) = 2.35, p > .05] nor cue validity [F(1, 11) = 1.10, p > .05] was sig-nificant. While, the interaction between cue type and cue validitywas significant [F(3, 33) = 23.12, p < .001]. To further explore thisinteraction, simple effects analyses of cue validity were conducted
130 J. Jiao et al. / Biological Psychol
Fig. 4. Panel A: grand-averaged ERPs elicited in Experiment 2 for each cue type fol-lowing cue onset at electrode sites PO7 and PO8 contralateral (dashed lines) andipsilateral (solid lines) to the hemifield of appearance of the color singleton cue.Panel B: difference waveforms obtained by subtracting ipsilateral activity from con-tcc
oRvivAiiccp
e
3
3impttn
woc
ralateral activity, shown separately for each of the four cue types. TC: target-colorue, WCIC: within category irrelevant-color cue, NTC: nontarget-color cue, and CCIC:ross category irrelevant-color cue.
n each cue type condition. For target-color cue, mean correctTs were significantly faster on valid than on invalid trials [684s. 722 ms, F(1, 11) = 37.25, p < .001]. However, for within categoryrrelevant-color cue, mean correct RTs were significantly slower onalid than on invalid trials [715 vs. 705 ms, F(1, 11) = 11.50, p < .01].
marginally significant inverted cue validity effect was presentn the nontarget-color cue condition [720 vs. 708 ms for valid andnvalid trials; F(1, 11) = 4.18, p = .066]. In contrast, no significantue validity effect was presented for the cross category irrelevant-olor cue [707 vs. 700 ms for valid and invalid trials; F(1, 11) = 2.76,
> .05].Error rates were generally low. ANOVA revealed no significant
ffects or interactions.
.5. ERP data
Fig. 4A shows grand-average ERP waveforms elicited in the00 ms post-cue interval at electrodes PO7/8 contralateral and
psilateral to the color singleton cue location, and Fig. 4B showsean difference amplitudes obtained by subtracting event-related
otential (ERP) waveforms at ipsilateral posterior electrode fromhat recorded at contralateral site, separately for trials with thearget-color cue, the within category irrelevant-color cue, theontarget-color cue and the cross category irrelevant-color cue.
The analysis of the mean amplitudes of the N2pc componentas carried out across a time window of 150–250 ms after the
nset of the cue display. A 4 (cue type: target-color, withinategory irrelevant-color, nontarget-color, and cross category
ogy 92 (2013) 125– 134
irrelevant-color cue) × 2 (electrode laterality: contralateral vs.ipsilateral) repeated-measure ANOVA was performed. Therewas a reliable main effect of laterality [F(1, 11) = 15.49, p < .01],indicating the presence of the N2pc to color singleton cues, andan interaction between cue type and laterality [F(3, 33) = 26.13,p < .001]. To explore this interaction, we separately analyzedpairwise combinations of two cue conditions, with cue type nowa two-level factor. When ERPs to the within category irrelevant-color cue and the nontarget-color cue were analyzed together,the cue type × laterality interaction effect was not significant[F(1, 11) = .004, p > .05]. However, a reliable difference in N2pcamplitudes was found for other five pairwise combinations of twocue conditions, namely, the “target-color cue vs. within categoryirrelevant-color cue”, the “target-color cue vs. cross categoryirrelevant-color cue”, the “target-color cue vs. nontarget-colorcue”, the “cross category irrelevant-color cue vs. nontarget-colorcue”, and the “within category irrelevant-color cue vs. cross cate-gory irrelevant-color cue” [with all Fs(1, 11) > 10, all ps < .01]. Wefurther conducted two-tailed t-test analyses to assess the differ-ence between contralateral and ipsilateral conditions separatelyfor each cue condition. A significant laterality effect was obtainedfor the target-color cue [t(11) = 5.74, p < .001], the within categoryirrelevant-color cue [t(11) = 3.17, p < .01], and the nontarget-colorcue [t(11) = 3.09, p < .05], demonstrating that these three cue typeselicited the N2pc effect. But the laterality effect was not significantfor cross category irrelevant-color cue [t(11) = .05, p > .05], whichsuggested that the cue triggered no N2pc effect.
3.6. Discussion
As in Experiment 1, neither the behavioral cue validity effect norERP measures of the N2pc effect was found for the cross categoryirrelevant-color cue, indicating no attentional capture. The target-color cue again captured attention (reflected by behavioral cuevalidity effect and the observation of the N2pc). Results providedfurther evidence for the contingent capture hypothesis developedby Folk and colleagues (e.g., Folk and Remington, 1998; Folk et al.,1992).
A small N2pc effect was elicited by the nontarget-color cueand the within category irrelevant-color cue, suggesting that thesetwo types of cues did, to some extent, capture attention. How-ever, behavioral cue validity effect was not present for either ofthem. Eimer and Kiss (2010) offered a viable explanation for theseseemingly inconsistent results between behavioral and ERP meas-ures. According to their explanation, some no-capture trials (e.g.,trials which elicited no reliable N2pc) elicited inverted cue valid-ity effects, and these inverted cue validity effects reconciled cuevalidity effect triggered by capture trials (see Section 5 for a moredetailed debate).
The main purpose of Experiment 2 was to further investigatewhether contingent capture could occur for an entire category orfor the specific feature value, when participants had to ignore a cer-tain shade of green while attending to another shade of green. Theresults showed that the N2pc effect was present for the target-colorcue, the within category irrelevant-color cue, and the nontarget-color cue. In contrast to the Experiment 1, where no significantdifference in the N2pc effect was obtained between the target-colorcue and the within category irrelevant-color cue, N2pc amplitudesto the target-color cue were significantly larger compared to thewithin category irrelevant-color cue and the nontarget-color cuein Experiment 2. Furthermore, behavioral cue validity effect was
present for the target-color cue, and no such behavioral cue valid-ity effect was elicited by the within category irrelevant-color cue.Taken together, present results suggested that contingent capturecould occur at the specific feature value.
sychology 92 (2013) 125– 134 131
ssIerctaifut
4
cftttcti
2gaaE
4
afnw
4
2tsgcsbc
4
ib
4
dnctp
Fig. 5. Panel A: grand-averaged ERPs elicited in Experiment 3 for each cue type fol-lowing cue onset at electrode sites PO7 and PO8 contralateral (dashed lines) andipsilateral (solid lines) to the hemifield of appearance of the color singleton cue.Panel B: difference waveforms obtained by subtracting ipsilateral activity from con-tralateral activity, shown separately for each of the four cue types. TC: target-colorcue, WCIC: within category irrelevant-color cue, NTC: nontarget-color cue, and CCIC:
J. Jiao et al. / Biological P
Before the conclusion that contingent capture could occur at thepecific feature value can be firmly established, we need to con-ider whether results from the current design can be generalized.n particular, we need to confirm that the choice of color for certainxperimental conditions used in our Experiment 2 did not bias theesults in some way. For example, in Experiment 2, if the withinategory irrelevant-color cue was perceived to be more similar tohe nontarget than to the target (e.g., based on saturation or otherttributes), the lack of attentional capture for the within categoryrrelevant-color cue condition would have to be interpreted dif-erently. To rule out alternative explanations, in Experiment 3, wesed the same cue colors used in Experiment 2 but assigned themo different cue conditions.
. Experiment 3
In Experiment 3, participants’ task was to respond to the green2olor singleton bar and ignore the green1 color singleton bar. There-ore, in Experiment 3, the green2 color singleton cue (which washe within category irrelevant-color cue in Experiment 2) was nowhe target-color cue; the green1 color singleton cue (which washe target-color cue in Experiment 2) was now the nontarget-olor cue; consequently the green3 color singleton cue (which washe nontarget color cue in Experiment 2) was the within categoryrrelevant-color cue.
If the results pattern of Experiment 3 replicates Experiment results, this will further confirm the conclusion that contin-ent capture could occur at the specific feature value. If not,n alternative interpretation of results would have to be gener-ted from the properties of the specific set of simuli we used inxperiment 2.
.1. Participants
A total of twelve undergraduate and graduate students (7 malesnd 5 females, mean age = 21 years) participated in the experimentor a monetary reward. All participants had normal or correct-to-ormal vision, and all were right-handed. None of the participantsere color blind.
.2. Apparatus, stimuli, and procedure
Apparatus, stimuli, and procedure were identical to Experiment, except that participants were asked to search for a green single-on of a certain shade of color green (green2) only and to ignore aingleton of another shade of color green (green1). Therefore, thereen 2 color singleton cue was the target-color cue, the green 1olor singleton cue was the nontarget-color cue, the green 3 coloringleton cue was the within category irrelevant-color cue, and thelue color singleton cue was the cross category irrelevant-colorue.
.3. Design, electrophysiological (EEG) recording, and analysis
Design, electrophysiological (EEG) recording, and analysis weredentical to Experiment 2. An average of 7.8% of trials were rejectedecause of artifacts (range = 0.8–21.1%).
.4. Behavioral data
The behavioral data were analyzed as a function of cue type con-ition (the target-color cue, within category irrelevant-color cue,
ontarget-color cue and cross category irrelevant-color cue) andue validity (valid and invalid). Mean RTs (line graphs) on correctrials and error rates (bar graphs) for each of these conditions wereresented in Fig. 2 (right panel).cross category irrelevant-color cue.
There was a significant main effect of cue type [F(3, 33) = 7.22,p < .01], but not cue validity [F(1, 11) = .17, p > .05]. In addition,interaction between cue type and cue validity was significant [F(3,33) = 4.40, p < .05]. To further explore this interaction, simple effectsanalyses of cue validity were conducted on each cue type condition.Mean correct RTs were significantly faster on valid than on invalidtrials for the target-color cue [734 vs. 760 ms, F(1, 11) = 5.10, p < .05].However, for the nontarget-color cue, mean correct RTs were sig-nificantly slower on valid than on invalid trials [737 vs. 711 ms,F(1, 11) = 4.93, p < .05]. In contrast, no significant cue validity effectwas presented for the within category irrelevant-color cue [709 vs.711 ms for valid and invalid trials; F(1, 11) = .13, p > .05] and thecross category irrelevant-color cue [710 vs. 712 ms for valid andinvalid trials; F(1, 11) = .06, p > .05].
Error rates were generally low. ANOVA revealed no significanteffects or interactions.
4.5. ERP data
Fig. 5A shows grand-average ERP waveforms elicited in the300 ms post-cue interval at electrodes PO7/8 contralateral andipsilateral to color singleton cue location, and Fig. 5B showsmean difference amplitudes obtained by subtracting event-related
potential (ERP) waveforms at ipsilateral posterior electrode fromthat recorded at contralateral site, separately for trials with thetarget-color cue, the within category irrelevant-color cue, thenontarget-color cue and the cross category irrelevant-color cue.
1 sychol
Tcoicrmecicitgicfpwtcce1stbetitspg
4
fstia(cgaa
tsccEmt
5
ttpsaw
32 J. Jiao et al. / Biological P
he analysis of mean amplitudes of the N2pc component wasarried out across a time window of 150–250 ms after the onsetf the cue display. A 4 (cue type: target-color, within categoryrrelevant-color, nontarget-color, and cross category irrelevant-olor cue) × 2 (electrode laterality: contralateral vs. ipsilateral)epeated-measure ANOVA was performed. There was a reliableain effect of laterality [F(1, 11) = 5.06, p < .05], indicating the pres-
nce of the N2pc to color singleton cues, and an interaction betweenue type and laterality [F(3, 33) = 5.33, p < .05]. To explore thisnteraction, we separately analyzed pairwise combinations of twoue conditions, with cue type now a two-level factor. A signif-cant interaction was found for three pairwise combinations ofwo cue conditions, that is, the “target-color cue vs. within cate-ory irrelevant-color cue”, the “target-color cue vs. cross categoryrrelevant-color cue”, and the “target-color cue vs. nontarget-colorue” [with all Fs (1, 11) > 9.94, all ps < .01], reflecting a reliable dif-erence of N2pc amplitudes between two cue conditions of eachair. However, the interaction between cue type and lateralityas not significant for the other three pairwise combinations of
wo cue conditions, that is, the “cross category irrelevant-colorue vs. nontarget-color cue”, the “within category irrelevant-colorue vs. cross category irrelevant-color cue”, and the “within cat-gory irrelevant-color cue vs. nontarget-color cue” [with all Fs (1,1) < .49, all ps > .05], reflecting that N2pc amplitudes did not differignificantly between two cue conditions of each pair. We fur-her conducted two-tailed t-test analyses to assess the differenceetween the contralateral and ipsilateral conditions separately forach cue condition. A significant laterality effect was obtained forhe target-color cue [t(11) = 5.28, p < .001], and the within categoryrrelevant-color cue [t(11) = 3.67, p < .01], demonstrating that thesewo cue types elicited N2pc effects. But the laterality effect was notignificant for the cross category irrelevant-color cue [t(11) = 1.12,
> .05], and the nontarget-color cue [t(11) = .04, p > .05]; this sug-ested that these two cue types triggered no N2pc effects.
.6. Discussion
Results of Experiment 3 were generally in line with what weound in Experiment 2. When participants were asked to ignore aingleton of a certain shade of the color green (green1) and attendo another shade of the color green (green2; task set green2), asn Experiments 1 and 2, significant behavioral cue validity effectnd the robust N2pc effect was observed for the target-color cueindicating attentional capture), while we did not find behavioralue validity effect and the N2pc effect for both the cross cate-ory irrelevant-color cue and the nontarget-color cue (indicating nottentional capture). These findings once again demonstrated thatttentional capture was contingent on top–down control setting.
In addition, although the within category irrelevant-color cueriggered the N2pc effect, N2pc amplitudes were significantlymaller relative to the target-color cue. Furthermore no behavioralue validity effect was elicited by the within category irrelevant-olor cue. These findings replicated the pattern of results fromxperiment 2. Therefore, the present study, together with Experi-ent 2 strongly suggested that contingent capture could occur at
he specific feature value.
. General discussion
Although previous studies have demonstrated that attentionalop–down control could be tuned to “features”, it is not clear whathis “feature” could be and circumstances in which those features
lay a role in attentional capture. Specifically, in this study, weet out to determine whether contingent capture can occur forn entire category or a specific feature value. Three experimentsere conducted to explore this issue. We used a spatial cueingogy 92 (2013) 125– 134
paradigm, in which a spatially uninformative cue display was fol-lowed by a search display, and combined behavioral measures (e.g.,cue validity effects on RT) and electrophysiological measures (e.g.,N2pc effects) of attentional capture. One unique feature in this cue-ing paradigm was that participants were required to respond to atarget color singleton bar in the search display but ignore the searchdisplay containing the nontarget-color singleton bar.
Behavioral results of Experiment 1 showed that when the targetand nontarget were from different color categories, both the target-color cue and the within category irrelevant-color cue produced cuevalidity effect and the magnitude of the effect for these two cuetypes was comparable, reflecting that these two cue types eliciteda similar magnitude of attentional capture. In the meantime, no cuevalidity effect was found for the nontarget-color cue and the crosscategory irrelevant-color cue. Moreover, the ERP data also providedimportant converging evidence. The target-color cue produced arobust N2pc effect, and the within category irrelevant-color cuetriggered the N2pc effect of similar magnitude as well. However,the nontarget-color cue and the cross category irrelevant-color cueelicited no reliable N2pc effect. Thus, our findings were in linewith previous observations by Ansorge and Heumann (2003), thatis, color singleton cues would capture attention only when theymatched the target color category. Our study provided new evi-dence for the hypothesis that contingent capture could occur foran entire category.
In Experiment 2, the target and nontarget in the search displayhad the same color category, but their specific feature values weredifferent. Results showed that the target-color cue produced thebehavioral cue validity effect and a robust N2pc effect, while nobehavioral cue validity effect was obtained by the within categoryirrelevant-color cue and the N2pc effect was significantly reduced.Similar results were obtained in Experiment 3 where the same cuesfrom Experiment 2 were used but were assigned to different cueconditions, suggesting that results from Experiment 2 were notan artifact of the specific relation between color properties andconditions.
It is important to point out that the most noteworthy resultin this study was the result difference between Experiment 1 andExperiments 2/3 for the within category irrelevant-color cue condi-tion. While the within category irrelevant-color cue produced a cuevalidity effect in Experiment 1, the within category irrelevant-colorcue could no longer elicit such cue validity effects in Experiments2/3. This difference was the result of the single methodological dif-ference between Experiments 1 and 2: the non-target color in thesearch display. Results of Experiments 2/3 provided clear evidencefor the hypothesis that contingent capture could occur at a specificfeature value.
In Experiment 1, the color singleton target and the color single-ton non-target in the search display not only had different specificfeature value, but also belonged to different color categories. It ispossible that there may be two separable search modes, a categor-ical search mode and a feature-value search mode. In Experiment1, both the categorical search mode and the feature-value searchmode were available to participants. They could finish the task byusing either the color category or the specific feature value. It islikely that use of information from the color category imposedless memory demand and required less cognitive resources andthus would serve as a default mode. Our findings were consis-tent with the conclusion made by Eimer and Kiss (2010) thatwhen two search modes (the singleton-detection mode and thefeature-search mode) were available, participants tended to choosethe singleton-detection mode, which would involve less cognitive
demand and working memory load. Attention would be set for thebroad color category (green). In this case, both the target-color cueand within category irrelevant-color cue would capture attention,because they matched task set on target’s category.
sychol
etcwtatte
sgaac3e(scwm
a1cnciagreitaarevvbeb(dwotTcaicfwactacevNb
J. Jiao et al. / Biological P
However, in Experiments 2 and 3, participants had to differ-ntiate different shades of green color between the target andhe non-target during search phase, thus selectivity for the spe-ific shade of green for the target color in the search displayas required. In that case, participants would be forced to adopt
he feature-value search mode. Attention will be set narrowlyround the specific feature value of the target. Consequentlyhe within category irrelevant-color cue, which did not matchhe observer’s control settings, would not elicit the cue validityffect.
In addition, the results in our study also showed a small yetignificantly inverted cue validity effect (RTs were slower for tar-ets at cued versus uncued locations) for the nontarget-color cuend the cross category irrelevant-color cue (Experiment 1), as wells for the nontarget-color cue and the within category irrelevant-olor cue (Experiment 2), and the nontarget-color cue (Experiment). Previous studies also reported similar findings (e.g., Eimert al., 2009; Eimer and Kiss, 2010; Lamy et al., 2004). Lamy et al.2004), for instance, found that RTs were significantly slower atame-location than different-location in the nontarget-singletonondition (Experiments 1 and 2). They argued that color singletonshose features did not match the currently top–down task settingight induce a transient location-specific inhibition.It is interesting to point out the discrepancy between behavioral
nd ERP results observed in all three experiments. In Experiment, RT results showed inverted cue validity effect for the nontarget-olor cue and the cross category irrelevant-color cue but there wereo N2pc effects. In Experiments 2 and 3, RT results showed invertedue validity effect (Experiment 2) or no cue validity effect (Exper-ment 3) for the within category irrelevant-color cue. However,
small but significant N2pc was observed for the within cate-ory irrelevant-color cue. The study by Eimer and Kiss (2010) alsoeported similar results where RTs showed inverted cue validityffect for the nontarget-color cue or no cue validity effect for therrelevant-color cue. However, the ERP results showed that thesewo cues did trigger a small but significant N2pc. They offered
viable explanation for these seemingly inconsistent behavioralnd ERP results. They suggested that cues that did not match cur-ent top–down control settings would be inhibited and wouldlicit no N2pc. However, such inhibition mechanism cannot pre-ent attentional capture in all trials. Thus, behavioral spatial cuealidity effects triggered on capture trials were compensated fory inverted spatial cue validity effects on no-capture trials. How-ver, the N2pc elicited by capture trials could only be reducedut could not be counteracted in the process of ERP data analysissee Eimer and Kiss, 2010, p. 960 and p. 961 for a more detailediscussion). In our Experiment 1, the categorical search mode,hich would involve less cognitive demand and working mem-
ry load, might be adopted to finish the task. Correspondingly,here may be more resources available for an inhibition mechanism.herefore, we inferred that the nontarget-color cue and the crossategory irrelevant-color cue, which did not match the currentlyctive task set, would be strongly inhibited in almost all trials. And,nverted cue validity effect but no N2pc was observed by these twoues. While, in Experiments 2 and 3, participants might adopt theeature-value search mode, which would require more resourceshen completing the task, and there may be fewer resources avail-
ble for inhibition mechanism. In this context, although the withinategory irrelevant-color cue did not match the currently activeask set and would be inhibited, a few trails still could capturettention and trigger the N2pc. So, it is possible that for the withinategory irrelevant-color cue, inverted cue validity effects, which
licited on some trials without capture, reconciled positive cuealidity effects triggered by capture trials (e.g., trials elicited robust2pc). This possibility finally resulted in discrepancy betweenehavioral and ERP results.ogy 92 (2013) 125– 134 133
6. Conclusions
The current study provided further evidence for the hypothesisthat attentional capture is contingent on current top–down tasksets. Both behavioral and electrophysiological data in the presentstudy suggested that contingent capture could occur for an entirecategory and could also occur at a specific feature value. When bothsearch modes (a categorical versus a feature-value search mode)were available, the choice of a specific search strategy may bedirected by the goal of minimizing attention control and workingmemory load in the processing of visual search.
Acknowledgements
This study was supported by awards from the National NaturalScience Foundation of China (NSFC 31170982), The authors thankthe four anonymous reviewers for their helpful comments.
References
Al-Aidroos, N., Harrison, S., Pratt, J., 2010. Attentional control settings prevent abruptonsets from capturing visual spatial attention. Quarterly Journal of ExperimentalPsychology 63, 31–41.
Ansorge, U., Heumann, M., 2003. Top–down contingencies in peripheral cueing:the roles of color and location. Journal of Experimental Psychology: HumanPerception and Performance 29, 937–948.
Bacon, W.F., Egeth, H.E., 1994. Overriding stimulus-driven attentional capture. Per-ception and Psychophysics 55, 485–496.
Eimer, M., 1996. The N2pc component as an indicator of attentional selectivity.Electroencephalography and Clinical Neurophysiology 99, 225–234.
Eimer, M., Kiss, M., 2008. Involuntary attentional capture is determined by task set:evidence from event-related brain potentials. Journal of Cognitive Neuroscience20, 1423–1433.
Eimer, M., Kiss, M., 2010. Top–down search strategies determine attentional cap-ture in visual search: behavioral and electrophysiological evidence. AttentionPerception & Psychophysics 72, 951–962.
Eimer, M., Kiss, M., Press, C., Sauter, D., 2009. The roles of feature-specific taskset and bottom-up salience in attentional capture: an ERP study. Journalof Experimental Psychology: Human Perception and Performance 35, 1316–1328.
Folk, C.L., Remington, R., 1998. Selectivity in distraction by irrelevant featural sin-gletons: evidence for two forms of attentional capture. Journal of ExperimentalPsychology: Human Perception and Performance 24, 847–858.
Folk, C.L., Remington, R., 1999. Can new objects override attentional control settings?Attention, Perception, & Psychophysics 61, 727–739.
Folk, C.L., Remington, R.W., Johnston, J.C., 1992. Involuntary covert orienting is con-tingent on attentional control settings. Journal of Experimental Psychology:Human Perception and Performance 18, 1030–1044.
Folk, C.L., Remington, R.W., Wright, J.H., 1994. The structure of attentional control:contingent attentional capture by apparent motion, abrupt onset, and color.Journal of Experimental Psychology: Human Perception and Performance 20,317–329.
Gibson, B.S., Amelio, J., 2000. Inhibition of return and attentional control settings.Attention, Perception, & Psychophysics 62, 496–504.
Gibson, B.S., Kelsey, E.M., 1998. Stimulus-driven attentional capture is contingenton attentional set for displaywide visual features. Journal of Experimental Psy-chology: Human Perception and Performance 24, 699–706.
Hickey, C., McDonald, J.J., Theeuwes, J., 2006. Electrophysiological evidenceof the capture of visual attention. Journal of Cognitive Neuroscience 18,604–613.
Jolicoeur, P., Sessa, P., Dell’Acqua, R., Robitaille, N., 2006. Attentional control and cap-ture in the attentional blink paradigm: evidence from human electrophysiology.European Journal of Cognitive Psychology 18, 560–578.
Kay, P., Kempton, W., 1984. What is the Sapir–Whorf hypothesis? American Anthro-pologist 86, 65–79.
Kiss, M., Eimer, M., 2011. Attentional capture by size singletons is determined bytop–down search goals. Psychophysiology 48, 784–787.
Kiss, M., Jolicaeur, P., Dell’Acqua, R., Eimer, M., 2008. Attentional capture by visualsingletons is mediated by top–down task set: new evidence from the N2pccomponent. Psychophysiology 45, 1013–1024.
Lamy, D., Egeth, H.E., 2003. Attentional capture in singleton-detection and feature-search modes. Journal of Experimental Psychology: Human Perception andPerformance 29, 1003–1020.
Lamy, D., Leber, A., Egeth, H.E., 2004. Effects of task relevance and stimulus-driven
salience in feature-search mode. Journal of Experimental Psychology: HumanPerception and Performance 30, 1019–1031.Lien, M.C., Ruthruff, E., Cornett, L., 2010a. Attentional capture by singletons iscontingent on top–down control settings: evidence from electrophysiologicalmeasures. Visual Cognition 18, 682–727.

1 sychol
L
L
L
L
34 J. Jiao et al. / Biological P
ien, M.C., Ruthruff, E., Goodin, Z., Remington, R.W., 2008. Contingent attentionalcapture by top–down control settings: converging evidence from event-relatedpotentials. Journal of Experimental Psychology: Human Perception and Perfor-mance 34, 509–530.
ien, M.C., Ruthruff, E., Johnston, J.C., 2010b. Attentional capture with rapidly chang-ing attentional control settings. Journal of Experimental Psychology: HumanPerception and Performance 36, 1–16.
uck, S.J., 2005. An Introduction to the Event-Related Potential Technique. MIT Press,Cambridge, MA.
uck, S.J., Hillyard, S.A., 1994. Spatial filtering during visual search: evidence fromhuman electrophysiology. Journal of Experimental Psychology: Human Percep-tion and Performance 20, 1000–1014.
ogy 92 (2013) 125– 134
Mazza, V., Turatto, M., Umilta, C., Eimer, M., 2007. Attentional selection and identifi-cation of visual objects are reflected by distinct electrophysiological responses.Experimental Brain Research 181, 531–536.
Navalpakkam, V., Itti, L., 2006. Top–down attention selection is fine grained. Journalof Vision 6, 1180–1193.
Theeuwes, J., 1991. Exogenous and endogenous control of attention: the effect ofvisual onsets and offsets. Attention, Perception, & Psychophysics 49, 83–90.
Theeuwes, J., 1992. Perceptual selectivity for color and form. Attention, Perception,& Psychophysics 51, 599–606.
Theeuwes, J., 1994. Stimulus-driven capture and attentional set: selective searchfor color and visual abrupt onsets. Journal of Experimental Psychology: HumanPerception and Performance 20, 799–806.
Related Documents