Amphibia-Reptilia 32 (2011): 399-411 Contemporary gene flow and weak genetic structuring in Rococo toad (Rhinella schneideri) populations in habitats fragmented by agricultural activities Maurício P. Arruda 1,* , Eliana Morielle-Versute 1 , Artur Silva 2 , Maria Paula C. Schneider 2 , Evonnildo C. Gonçalves 2 Abstract. The reduced vagility and philopatric behaviour of most amphibians make them especially vulnerable to the effects of habitat fragmentation, in particular the loss of genetic variation. However, almost no data are available on the effects of agricultural practices on populations of Neotropical amphibians. Here, the genetic diversity of Rococo toad (Rhinella schneideri) populations in the highly disturbed landscape of the north-western region of the Brazilian state of São Paulo was analysed using microsatellite markers. Two areas were sampled – one dominated by open pastures (four populations) and the other by sugar cane plantations (two populations) – in an attempt to evaluate the possible influence of the type of anthropogenic matrix on genetic variability and gene flow (dispersion). The populations presented a relatively uniform genetic stock, with low levels of inbreeding (Fis) and high levels of admixture between localities (Fst,Rst, STRUCTURE) indicating no genetic subdivision. The results indicated relatively high levels of recent migration among sites (m) and no isolation by distance. The analyses also found that historical and contemporary rates of migration among populations were broadly similar. Overall, then, neither type of matrix appeared to have an effect on the connectivity of the Rococo toad populations. This suggests that the species has a considerable capacity for dispersal, allowing it to maintain a relatively homogeneous population, even under intense human pressure. Keywords: anthropogenic disturbance, gene flow, genetic diversity, high dispersion, microsatellite, population structure. Introduction The destruction and fragmentation of natural habitats are the primary cause of the loss of biodiversity throughout the world (Cushman, 2006). The configuration of anthropogenic land- scapes may have an especially marked effect on the genetic variation of populations of amphib- ians, which are generally considered to have reduced mobility (Blaustein, Wake and Sousa, 1994). Amphibian populations also tend to ex- hibit strong breeding philopatry (Beebee, 2005). These characteristics may result in reduced gene 1 - Laboratório de Chiroptera, Departamento de Zoologia e Botânica, Universidade Estadual Paulista (UNESP), Rua Cristóvão Colombo 2265, Jardim Nazareth CEP 15061-000, São José do Rio Preto, São Paulo State, Brazil 2 - Laboratório de Polimorfismo de DNA, Instituto de Ciên- cias Biológicas, Universidade Federal do Pará (UFPA), Augusto Corrêa 1, CEP 66075-900, Belém, Pará State, Brazil * Corresponding author; e-mail: [email protected] flow and consequently, population structuring. Anthropogenic habitat disturbance will likely exacerbate these effects. In Brazil, human activities have led to the conversion of continuous tracts of native habi- tat into a landscape of relatively small and iso- lated fragments. The north-western region of São Paulo state is characterised by typical Cer- rado savannah, interspersed with areas of sea- sonally semideciduous forest, although more than 90% of the original vegetation has been re- placed with pastures, plantations and urban ar- eas. Extensive plantations of sugar cane have been present in the region for more than 80 years, although habitat conversion has increased considerably in recent decades (CETEC, 2008), and the region is now São Paulo’s most heav- ily deforested, and also has the lowest concen- tration of protected areas (Kronka et al., 1993). These characteristics make the region an appro- priate model for the assessment of fragmented populations. © Koninklijke Brill NV, Leiden, 2011. DOI:10.1163/017353711X588182

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Amphibia-Reptilia 32 (2011): 399-411
Contemporary gene flow and weak genetic structuringin Rococo toad (Rhinella schneideri) populationsin habitats fragmented by agricultural activities
Maurício P. Arruda1,*, Eliana Morielle-Versute1, Artur Silva2,
Maria Paula C. Schneider2, Evonnildo C. Gonçalves2
Abstract. The reduced vagility and philopatric behaviour of most amphibians make them especially vulnerable to the effectsof habitat fragmentation, in particular the loss of genetic variation. However, almost no data are available on the effectsof agricultural practices on populations of Neotropical amphibians. Here, the genetic diversity of Rococo toad (Rhinellaschneideri) populations in the highly disturbed landscape of the north-western region of the Brazilian state of São Paulowas analysed using microsatellite markers. Two areas were sampled – one dominated by open pastures (four populations)and the other by sugar cane plantations (two populations) – in an attempt to evaluate the possible influence of the type ofanthropogenic matrix on genetic variability and gene flow (dispersion). The populations presented a relatively uniform geneticstock, with low levels of inbreeding (Fis) and high levels of admixture between localities (Fst, Rst, STRUCTURE) indicatingno genetic subdivision. The results indicated relatively high levels of recent migration among sites (m) and no isolationby distance. The analyses also found that historical and contemporary rates of migration among populations were broadlysimilar. Overall, then, neither type of matrix appeared to have an effect on the connectivity of the Rococo toad populations.This suggests that the species has a considerable capacity for dispersal, allowing it to maintain a relatively homogeneouspopulation, even under intense human pressure.
Keywords: anthropogenic disturbance, gene flow, genetic diversity, high dispersion, microsatellite, population structure.
Introduction
The destruction and fragmentation of naturalhabitats are the primary cause of the loss ofbiodiversity throughout the world (Cushman,2006). The configuration of anthropogenic land-scapes may have an especially marked effect onthe genetic variation of populations of amphib-ians, which are generally considered to havereduced mobility (Blaustein, Wake and Sousa,1994). Amphibian populations also tend to ex-hibit strong breeding philopatry (Beebee, 2005).These characteristics may result in reduced gene
1 - Laboratório de Chiroptera, Departamento de Zoologiae Botânica, Universidade Estadual Paulista (UNESP),Rua Cristóvão Colombo 2265, Jardim Nazareth CEP15061-000, São José do Rio Preto, São Paulo State,Brazil
2 - Laboratório de Polimorfismo de DNA, Instituto de Ciên-cias Biológicas, Universidade Federal do Pará (UFPA),Augusto Corrêa 1, CEP 66075-900, Belém, Pará State,Brazil*Corresponding author; e-mail: [email protected]
flow and consequently, population structuring.Anthropogenic habitat disturbance will likelyexacerbate these effects.
In Brazil, human activities have led to theconversion of continuous tracts of native habi-tat into a landscape of relatively small and iso-lated fragments. The north-western region ofSão Paulo state is characterised by typical Cer-rado savannah, interspersed with areas of sea-sonally semideciduous forest, although morethan 90% of the original vegetation has been re-placed with pastures, plantations and urban ar-eas. Extensive plantations of sugar cane havebeen present in the region for more than 80years, although habitat conversion has increasedconsiderably in recent decades (CETEC, 2008),and the region is now São Paulo’s most heav-ily deforested, and also has the lowest concen-tration of protected areas (Kronka et al., 1993).These characteristics make the region an appro-priate model for the assessment of fragmentedpopulations.
© Koninklijke Brill NV, Leiden, 2011. DOI:10.1163/017353711X588182

400 M.P. Arruda et al.
Data on the effects of agricultural activitiesare vital to the understanding of communitystructure. The majority of studies investigatingthe effects of changes in habitat structure onamphibian populations have been conducted inNorth America (Gardner, Barlow and Peres,2007), emphasising the need for research intropical and subtropical regions, which not onlyhave the greatest diversity of amphibian species,but also suffer the highest rates of habitat loss.
Few data are available on the effects ofhuman disturbance on amphibian populations(Urbina-Cardona, Olivarez-Pérez and Reynoso,2006), and there are contradictory patterns(Gray, Smith and Brenes, 2004). Knutson et al.(1999) recorded both positive and negative ef-fects on the abundance and diversity of frogsand toads, given that some habitat remnants inagricultural landscapes may provide refuges forcertain species. However, despite the relativeabundance of some species in disturbed areas,the overall probability of local extinctions ishigher than that in pristine environments (Gray,Smith and Brenes, 2004).
The Rococo toad (Rhinella schneideri) is themost widespread South American amphibian,with a geographic range that includes much ofBrazil, Paraguay and Bolivia, and neighbouringareas of Argentina, and Uruguay (Bastos et al.,2003). Despite this, little is known of the ecol-ogy of the species, which is considered to bea habitat generalist with a preference for openvegetation (Silvano and Pimenta, 2003; Vascon-cellos and Colli, 2009; Batista et al., 2011).Widely distributed and abundant species such asR. schneideri are generally the most appropri-ate models for the investigation of the effects ofhabitat loss and fragmentation (Whiteley, Spru-ell and Allendorf, 2006).
This study – the first of its kind – analysedthe genetic variability of R. schneideri popula-tions inhabiting fragmented ecosystems usingpolymorphic DNA microsatellite loci to detectpopulation differentiation and patterns of geneflow. These molecular markers have been ap-plied in genetic studies of amphibians for the
inference of dispersal patterns and functionalconnectivity in populations inhabiting impactedlandscapes (Johansson et al., 2005; Purrenhage,Niewiarowski and Moore, 2008). The main ob-jective was to determine the impact of differ-ent types of anthropogenic matrix (pastures andplantations) on the population structure and ge-netic variability of the Rococo toad.
Materials and methods
Study area and specimen collection
The study area encompasses six sites in north-westernSão Paulo state, Brazil (fig. 1), which represent two riverbasins characterised by distinct patterns of land use. Exten-sive cattle ranching predominates in the lower Tietê riverbasin located downstream, where four populations weresampled – Votuporanga (20◦30′17.36′′S, 50◦05′42.67′′W),Macaubal (20◦44′44.34′′S, 49◦56′17.64′′W), Santo An-tônio do Aracanguá, or simply Aracanguá (20◦55′47.25′′S,50◦21′16.34′′W), and Planalto (20◦59′31.28′′S,49◦58′48.59′′W). By contrast, the Tietê Batalha basin isdominated by dense plantations of sugar cane (monocul-ture plantations). Two R. schneideri populations were sam-pled here, in Sales (21◦24′39.73′′S, 49◦28′32.64′′W) andNovo Horizonte (21◦30′08.16′′S, 49◦18′32.35′′W). Speci-mens were collected between September, 2006, and March,2007, from reservoirs, lakes, and swamps in areas of transi-tion between the Cerrado and the tropical Atlantic Forest.
Specimen collection for scientific research was autho-rised by the Brazilian Institute of Environment and Renew-able Natural Resources (IBAMA) through special licencenumber 13923-1. Ten specimens were collected at each site,resulting in a total sample of 60 individuals.
Extraction of genomic DNA
The animals were sacrificed according to the protocolrecommended for amphibians by the Australia and NewZealand Council for the Care of Animals in Research andTeaching (2001), which involves the cutaneous absorptionof a lethal dose of benzocaine. The DNA was extracted frommuscle fragments according to the Sambrook, Fritsch andManiatis (1989) protocol for the preparation of the DNAtemplate.
Microsatellite markers and the evaluation of genotypes
The specimens were genotyped using nine polymorphic mi-crosatellite loci – Rschµ1, Rschµ3, Rschµ4, Rschµ6, Rschµ8,Rschµ9, Rschµ10, Rschµ12, and Rschµ14, which were de-veloped and standardised according to the Molecular Ecol-ogy Resources Primer Development Consortium (2009) andassumed to be neutral markers. All these microsatellites arecomposed of dinucleotide repeat motifs, with the excep-tion of Rschµ4 and Rschµ6, a di-tetra complex and a repeat

Genetic variability of Rococo toad 401
Figure 1. Sites at which specimens of R. schneideri were collected in the lower Tietê and Tietê Batalha river basins innorth-western São Paulo, Brazil.
tetranucleotide, respectively. An M13-tail was added to the5′ end of the each forward primer of the identical sequenceof the universal primer labelled with Cy5, and the PCR am-plification conditions were developed according to the pro-cedure described by Arruda et al. (2010). The PCR productswere separated by vertical electrophoresis in 6% denaturingpolyacrylamide gels in an ALF Express II automatic ana-lyzer (Amersham Biosciences, Freiburg, Germany). Allelelength was estimated using the Allele Locator 1.03 applica-tion (Amersham Biosciences).
Intrapopulation variability
Intrapopulation variability was quantified according to ob-served and expected heterozygosity and the effective num-ber of alleles estimated in POPGENE 1.32 (Yeh, Yang andBoylet, 1997). Allele richness was calculated by FSTAT2.9.3.2 (Goudet, 1997). The significance of deviations fromthe Hardy-Weinberg equilibrium was evaluated on the basisof the inbreeding coefficient generated by GENEPOP 4.0.9(Rousset, 2008), with significance levels calculated by theMarkov chain method, with 10 000 dememorization steps,and 100 batches with 5000 iterations per batch. This pro-gram also calculated the inbreeding coefficient (Fis) basedon allele identity.
Genetic structure of populations. Levels of genetic differ-entiation among all populations and between pairs of popu-lation were estimated in GENEPOP 4.0.9 (Rousset, 2008)and ARLEQUIN 3.11 (Excoffier and Schneider, 2006),based on the Fst and Rst indices, respectively. The Rst
values assume a stepwise mutation model of evolution formicrosatellite markers. The number of populations repre-sented in the samples (k) was estimated using the Bayesianapproach in STRUCTURE 2.3.1 (Pritchard, Stephens andDonnelly, 2000). The values of k (the putative number ofpopulations) varied from 1 to 10. Three runs were per-formed for each k value with a burn-in length of 100 000 anda Markov chain Monte Carlo (MCMC) of 200 000, usingthe admixture model assuming independent and correlatedallele frequency, without any prior population information.Spatial discontinuities in the genetic structure (breaks ingene flow) were analysed using the spatial Bayesian cluster-ing algorithm with a hidden Gaussian random field (HGRF)implemented in TESS 2.3.1 (Chen et al., 2007) and withMCMC computations in GENELAND 3.2.4 (Guillot, San-tos and Estoup, 2008). The TESS analysis was based onthe conditional autoregressive (CAR) admixture model withthe interaction parameter (�) fixed at 0.6, as described byChen et al. (2007). Ten independent runs were conductedfor each inferred k value (2-10), with 100 000 sweeps anda burn-in period of 20 000 sweeps to identify the maxi-mum population number (Kmax), which was selected whenthe deviance information criterion (DIC) reached a plateauand/or the Q-matrix of individual posterior probabilitiesstabilised. Average assignments of individuals to popula-tions (Q values) for the selected Kmax were then anal-ysed in CLUMPP 1.1.2b (Jakobsson and Rosenberg, 2007)to correct possible discrepancies between runs. The resultswere displayed in DISTRUCT 1.1 (Rosenberg, 2004). Inthe TESS and GENELAND analyses, it was assumed thatthe specimens were collected close to their breeding sites,

402 M.P. Arruda et al.
and the uncertainty associated with spatial coordinates wasset to 1 km, the mean radius of a R. schneideri territory.The GENELAND analysis was run independently 10 times,with 500 000 MCMC iterations and thinning of 50 000 withk values ranging from 1 to 10 and the maximum number ofnuclei in the Poisson-Voronoi tessellation fixed at 200. TheDirichlet distribution was tested as a model for allelic fre-quencies. The number of subpopulations was then inferredfrom the modal value of k based on the run that producedthe highest average posterior density probability.
Contemporary and historical migration rates. TheBAYESASS 1.3 program (Wilson and Rannala, 2003) wasused to evaluate current migration rates. This analysis isbased on a Bayesian approach under the assumption of non-equilibrium and MCMC resampling for estimating contem-porary immigration rates over the last few generations, po-tentially during the peak of anthropogenic impacts. This ap-proach incorporates the fact that the genotypes of immi-grants (or their recent descendants) will present transientdeviation from Hardy-Weinberg equilibrium, thus permit-ting their identification. A total of 3 000 000 iterations wererun in triplicate, with a burn-in of 1 000 000 and a samplingfrequency of 2000. The delta values for migration rate (m),allele frequencies (P ), and inbreeding (F ) were adjusted(m = 0.15; P = 0.29 and F = 0.35) to ensure that 40-60%of the total changes were accepted. In the case of F , how-ever, accepted changes could not be reduced to below 73%.An assignment test, implemented in GENECLASS 2.0 (Piryet al., 2004), was also employed to identify possible recentimmigrants in the putative populations. This program wasalso used to identify first generation migrants, based on twoapproaches – the Lhome : Lmax ratio and the Lhome val-ues. The Lhome : Lmax ratio is the most informative param-eter when all source population have been sampled (Paetkauet al., 2004), while the Lhome values is more appropriatewhen not all these populations have been sampled (Paetkauet al., 2004; Piry et al., 2004). For more precise computing,all procedures run in GENECLASS were inferred using thepartial Bayesian criterion of Rannala and Mountain (1997)and the MCMC resampling method of Paetkau et al. (2004),with a total of 100 000 individuals simulated and a thresholdof 0.01.
Coalescence approaches were used to estimate long-term migration (approximately 1000 years) and historicaleffective population size, in MIGRATE 3.2.7 (Beerli andFelsentein, 2001). Migration was estimated based on themutation-scaled immigration rate or M, defined as m/μ
(m = migration rate, μ = mutation rate) and effectivepopulation size � (= 4Ne·μ). A Brownian motion modelwas assumed for the analysis, and the settings consistedof 10 short chains with 2000 samples, 400 records and 3long chains with 10 000 samples and 3000 records usingmaximum-likelihood inference.
Contemporary and historic migration rates were com-pared using the values of m obtained from BAYESASS,and estimated from the values of M (=m/μ) detected byMIGRATE, with all M values being divided by an estimatedmutation rate of 5·10−4 (Garza and Williamson, 2001). Thecorrelation between contemporary and historic migrationrates (m) was assessed statistically using a Mantel test withztmantel implemented in MANTEL TESTER (Bonnet andVan de Peer, 2002) with 100 000 permutations.
Bottlenecks and isolation by distance and landscape resis-tance. Evidence of a recent population bottleneck (excessof heterozygosity) or founder event, was assessed for eachpopulation in BOTTLENECK 1.2.0.02 program (Cornuetand Luikart, 1996), using a one-tailed Wilcoxon signed-ranktest. The test was conducted with 100 000 replications us-ing an infinite allele mutation model and two-phase muta-tional models, with the latter incorporating 70% and 90% ofthe stepwise mutation model, which covers the most proba-ble range for amphibian microsatellites (Schlötterer, 2000).The degree of isolation by distance (Mantel test) was evalu-ated by the correlation between the genetic (Fst and Rst) andgeographic distances (Euclidean distance) between samplesites. The test was run in IBD 1.52 (Bohonak, 2002) with10 000 randomizations, for the full set of samples, and thoseof the lower Tietê basin only. The sequential Bonferroni cor-rection was applied to adjust the level of significance formultiple comparisons (Rice, 1989).
The isolation by landscape resistance model was testedin CIRCUITSCAPE 2.2 (McRae, 2006). This software in-corporates an algorithm that predicts total landscape re-sistance between sample sites based on all possible path-ways and complements least-cost approaches. To generatethe land cover data sets and identify absolute barriers to dis-persal, a 2.27 km pixel map of land use in the study area(north-western São Paulo) was obtained, based on the agri-cultural census of 2006 (IBGE, 2010). Four general cate-gories of land use were defined according to their potentialeffects on the dispersal of R. schneideri, and each categorywas assigned a conductance rating (higher values indicatelower resistance to dispersal): bodies of water (>2 km indiameter) = 0, urban areas = 2 (R. schneideri is found inurban areas), monoculture plantations = 4, other uses or ar-eas of transition (Cerrado and semideciduous forest) = 5,pasture or Cerrado = 6. In the absence of specific dataon R. schneideri, this classification was based on empiri-cal studies of other toad species adapted to open habitats(Stevens et al., 2006) and was modified according to thespecific characteristics of R. schneideri. In particular, as thespecies is adapted to open habitats (Batista et al., 2011),it was assumed that plantations offer more resistance todispersal than pasture. These values must nevertheless beconsidered arbitrary for R. schneideri. The resistance dis-tance was calculated according to the permeability of thelandscape separating localities (each pixel of the map beingcounted as a specific cost to dispersal according to its clas-sification). Finally, the correlation between resistance (CIR-CUITSCAPE outputs) and genetic distances (Fst and Rst)was used to check the influence of the composition of thelandscape (land use) on genetic differentiation among thepopulations. This was tested statistically based on a Manteltest using ztmantel implemented in Mantel Tester (Bonnetand Van de Peer, 2002) with 10 000 permutations. Geneticdistances were linearised using the Fst/(1 − Fst) transfor-mation (Rousset, 1997). The correlation between resistancedistance and contemporary gene flow (the sum of migrationrates in both directions) was evaluated concomitantly, due tothe fact that the map used for analysis was based on recentland cover.

Genetic variability of Rococo toad 403
Results
Intrapopulation genetic variability was esti-mated using six different parameters (table 1).None of the populations deviated significantlyfrom Hardy-Weinberg equilibrium. All indicesof genetic variability were relatively similaramong the populations analysed. The inbreed-ing coefficient was low (< 0.12) and not sig-nificant (p > 0.005) in all the populations,indicating reduced endogamy rates at the in-trapopulation level. A relatively small numberof private alleles were recorded in the popula-tions (2.96%, on average), with a uniform dis-tribution among them (3.45-3.70%) except forPlanalto, which had no private alleles whatso-ever.
The values for the fixation indices indicatedweak genetic structuring among all populations(Fst = 0.040; Rst = 0.021), with no well-defined genetic structure (Fst and Rst) in anypairwise comparison between populations (ta-ble 2). The lowest values were recorded for
Table 1. Levels of neutral genetic variability withinpopulations.
Putative Ho He PHWE Ne AR Fispopulation
Votuporanga 0.47 0.47 0.984 1.93 3.11 0.011Macaubal 0.47 0.46 0.787 1.93 3.11 −0.022Aracanguá 0.53 0.52 0.996 2.16 3.22 −0.019Planalto 0.41 0.45 0.693 1.84 3.67 0.094Sales 0.40 0.45 0.612 2.02 3.00 0.120Novo Horizonte 0.46 0.48 0.824 2.17 3.22 0.055
Ho = observed heterozygosity; He = expected heterozy-gosity; PHWE = probability of deviation from Hardy-Weinberg equilibrium (p value); Ne = effective number ofalleles; AR = allelic richness; Fis = inbreeding coefficient.
the comparisons between Sales and Novo Hori-zonte (Fst and Rst equal to zero), and Votupo-ranga and Aracanguá, and between Macaubaland both Sales and Novo Horizonte (table 2).
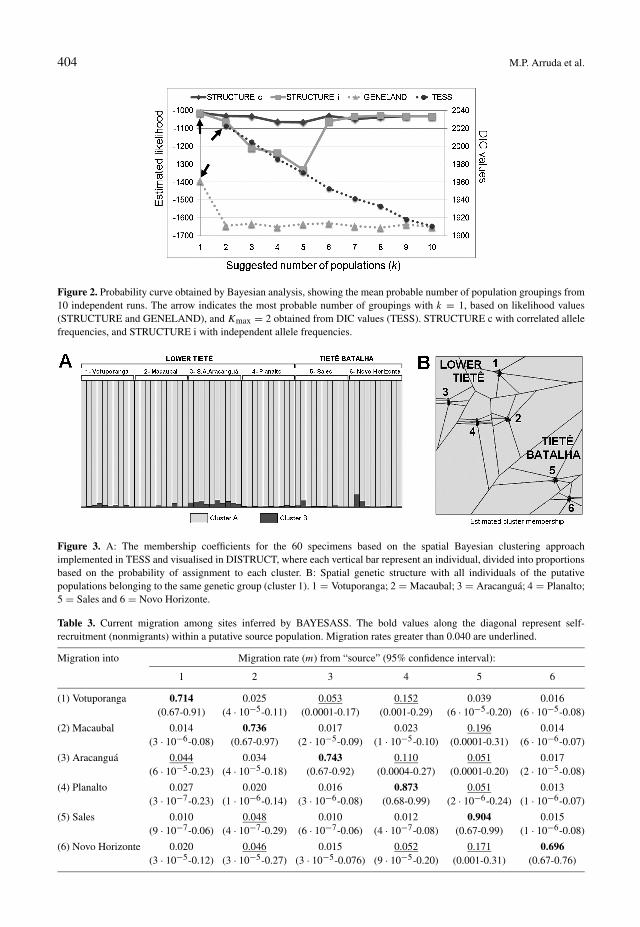
The STRUCTURE Bayesian analysis ar-ranged the R. schneideri populations in a sin-gle group (fig. 2), whether applying either theindependent or the correlated allele frequencymodels. The Bayesian analyses that incorpo-rated geographic data assigned all the individ-uals to a single group in the GENELAND anal-ysis (fig. 2), while in TESS, the analysis re-vealed an optimal Kmax of 2 (fig. 2), althoughall putative populations were assigned to samecluster (Q values > 0.963 to one cluster), andthe assignment of all individuals (membershipcoefficients) to this cluster was also stronglysupported (0.90 to 0.99; fig. 3A). The TESSanalysis found no spatial structuring amongsites (fig. 3B), and the CLUMPP analysis re-turned highly consistent estimates of Kmax (= 2)among the 10 runs (H ′ = 0.991).
Contemporary immigration rates (m) intoeach population, estimated from BAYESASS,are shown in table 3. Most pairwise compar-isons between localities detected recent migra-tions, indicated by m values above 0.032 (dis-tinguishable from the values generated by thesimulation with no information in the data set),and in eleven cases, the migration rate wasgreater than 0.04, which indicates the occur-rence of substantial contemporary gene flow(fig. 4). Migration rates were especially high(m > 0.1, which indicates that more than10% of the population are recent immigrants)in some cases, i.e. Planalto to Votuporanga,
Table 2. Estimates of Fst and Rst (in parentheses) for pairwise comparisons between populations.
Putative population
Votuporanga Macaubal Aracanguá Planalto SalesMacaubal 0.057 (0.061)Aracanguá 0.015 (0.015) 0.042 (0.044)Planalto 0.030 (0.031) 0.090* (0.099) 0.068* (0.072)Sales 0.043 (0.045) 0.021 (0.021) 0.057* (0.061) 0.051 (0.054)Novo Horizonte 0.033 (0.035) 0.013 (0.013) 0.058* (0.062) 0.039 (0.041) 0.000 (0.000)
∗ = significant P values after Bonferroni correction (p < 0.005).
404 M.P. Arruda et al.
Figure 2. Probability curve obtained by Bayesian analysis, showing the mean probable number of population groupings from10 independent runs. The arrow indicates the most probable number of groupings with k = 1, based on likelihood values(STRUCTURE and GENELAND), and Kmax = 2 obtained from DIC values (TESS). STRUCTURE c with correlated allelefrequencies, and STRUCTURE i with independent allele frequencies.
Figure 3. A: The membership coefficients for the 60 specimens based on the spatial Bayesian clustering approachimplemented in TESS and visualised in DISTRUCT, where each vertical bar represent an individual, divided into proportionsbased on the probability of assignment to each cluster. B: Spatial genetic structure with all individuals of the putativepopulations belonging to the same genetic group (cluster 1). 1 = Votuporanga; 2 = Macaubal; 3 = Aracanguá; 4 = Planalto;5 = Sales and 6 = Novo Horizonte.
Table 3. Current migration among sites inferred by BAYESASS. The bold values along the diagonal represent self-recruitment (nonmigrants) within a putative source population. Migration rates greater than 0.040 are underlined.
Migration into Migration rate (m) from “source” (95% confidence interval):
1 2 3 4 5 6
(1) Votuporanga 0.714 0.025 0.053 0.152 0.039 0.016(0.67-0.91) (4 · 10−5-0.11) (0.0001-0.17) (0.001-0.29) (6 · 10−5-0.20) (6 · 10−5-0.08)
(2) Macaubal 0.014 0.736 0.017 0.023 0.196 0.014(3 · 10−6-0.08) (0.67-0.97) (2 · 10−5-0.09) (1 · 10−5-0.10) (0.0001-0.31) (6 · 10−6-0.07)
(3) Aracanguá 0.044 0.034 0.743 0.110 0.051 0.017(6 · 10−5-0.23) (4 · 10−5-0.18) (0.67-0.92) (0.0004-0.27) (0.0001-0.20) (2 · 10−5-0.08)
(4) Planalto 0.027 0.020 0.016 0.873 0.051 0.013(3 · 10−7-0.23) (1 · 10−6-0.14) (3 · 10−6-0.08) (0.68-0.99) (2 · 10−6-0.24) (1 · 10−6-0.07)
(5) Sales 0.010 0.048 0.010 0.012 0.904 0.015(9 · 10−7-0.06) (4 · 10−7-0.29) (6 · 10−7-0.06) (4 · 10−7-0.08) (0.67-0.99) (1 · 10−6-0.08)
(6) Novo Horizonte 0.020 0.046 0.015 0.052 0.171 0.696(3 · 10−5-0.12) (3 · 10−5-0.27) (3 · 10−5-0.076) (9 · 10−5-0.20) (0.001-0.31) (0.67-0.76)
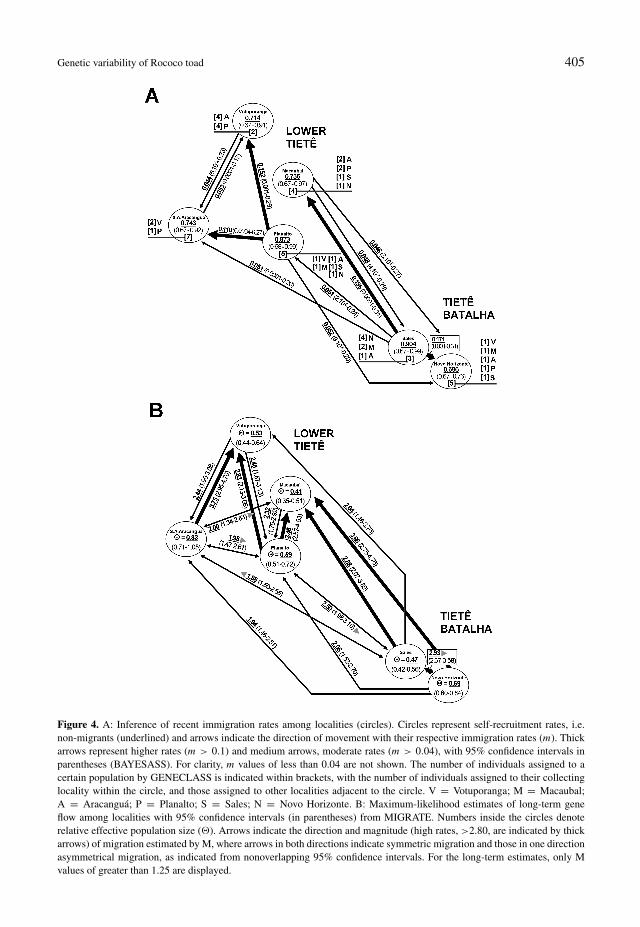
Genetic variability of Rococo toad 405
Figure 4. A: Inference of recent immigration rates among localities (circles). Circles represent self-recruitment rates, i.e.non-migrants (underlined) and arrows indicate the direction of movement with their respective immigration rates (m). Thickarrows represent higher rates (m > 0.1) and medium arrows, moderate rates (m > 0.04), with 95% confidence intervals inparentheses (BAYESASS). For clarity, m values of less than 0.04 are not shown. The number of individuals assigned to acertain population by GENECLASS is indicated within brackets, with the number of individuals assigned to their collectinglocality within the circle, and those assigned to other localities adjacent to the circle. V = Votuporanga; M = Macaubal;A = Aracanguá; P = Planalto; S = Sales; N = Novo Horizonte. B: Maximum-likelihood estimates of long-term geneflow among localities with 95% confidence intervals (in parentheses) from MIGRATE. Numbers inside the circles denoterelative effective population size (�). Arrows indicate the direction and magnitude (high rates, >2.80, are indicated by thickarrows) of migration estimated by M, where arrows in both directions indicate symmetric migration and those in one directionasymmetrical migration, as indicated from nonoverlapping 95% confidence intervals. For the long-term estimates, only Mvalues of greater than 1.25 are displayed.

406 M.P. Arruda et al.
Planalto to Aracanguá (in the lower Tietê basin),Sales to Novo Horizonte (Tietê Batalha basin)and even between basins (Sales to Macaubal).The lowest self-recruitment rate was recordedat Novo Horizonte and the highest at Sales, bothlocated within the Tietê Batalha basin.
The assignment test indicated 26 correctly-assigned individuals with an accuracy ap-proaching 43%. The results of this test coin-cided with those of BAYESASS (fig. 4A), inwhich the localities identified as recipients ofcontemporary gene flow attributed many in-dividuals to other localities, identified as thesource of gene flow. For example, two speci-mens from Aracanguá were assigned to Votupo-ranga, four specimens from Votuporanga wereassigned to Planalto, and two from Sales wereassigned to Macaubal.
The output from GENECLASS also identi-fied four individuals as first-generation migrants(p < 0.01) according to the Lhome : Lmaxratio (from Planalto to Votuporanga p =0.007, Planalto-Macaubal p = 0.004, NovoHorizonte-Sales p = 0.000, Sales-Novo Hor-izonte p = 0.007). Similar results were ob-tained for Lhome, except for the migrant iden-tified in Macaubal (Planalto-Votuporanga p =0.005, Planalto-Macaubal p = 0.003, NovoHorizonte-Sales p = 0.000, Sales-Novo Hori-zonte p = 0.001). These results were also con-gruent with those of the BAYESASS analysis,with the exception of the recent gene flow de-tected by GENECLASS between Novo Hori-zonte to Sales (including in the assignment test).
The analysis of immigration rates accord-ing to the coalescence theory in MIGRATE re-corded values of M ranging from 0.52 (0.26-0.91) from Aracanguá to Novo Horizonte to3.75 (2.95-4.70) from Aracanguá to Votupo-ranga (fig. 4B). The model of symmetric mi-gration occurred predominantly within the twobasins, but not between the basins, which wascharacterised by significant asymmetric migra-tion (nonoverlapping 95% confidence intervalsfor pairwise comparisons). MIGRATE also de-tected past routes of gene flow that exhibited
similarities with those indicated by BAYESASS(e.g. from Planalto to Votuporanga, bidirection-ally between Votuporanga and Aracanguá, andfrom Sales to Novo Horizonte and Sales toMacaubal). Estimated effective population sizefor Aracanguá was larger than those of otherlocalities. An important convergence was ob-served (rmantel = 0.441077, p = 0.0041) be-tween the outputs obtained from BAYESASS(contemporary migration) and MIGRATE (his-torical migration) in the MANTEL TESTERanalysis.
The Mantel test found no significant asso-ciation between genetic and geographic dis-tance based on Rst for all localities (rmantel =0.335798; p = 0.15) or Fst (rmantel = 0.000740;p = 0.45). The same pattern was observedat sites within the lower Tietê basin (Rstrmantel = −0.82199; p = 0.96; Fst rmantel =0.077532; p = 0.55). This is further sup-ported by the results of the BAYESASS anal-ysis, which indicated that migration rates werenot necessarily higher between geographicallycloser localities. The correlation between ge-netic divergence (Fst, Rst) and resistance dis-tance (CIRCUITSCAPE outputs) was not sig-nificant (Fst rmantel = −0.1591, p = 0.2889;Rst rmantel = −0.1461, p = 0.3028), and asimilar result was obtained for migration rates(rmantel = −0.2868, p = 0.1556).
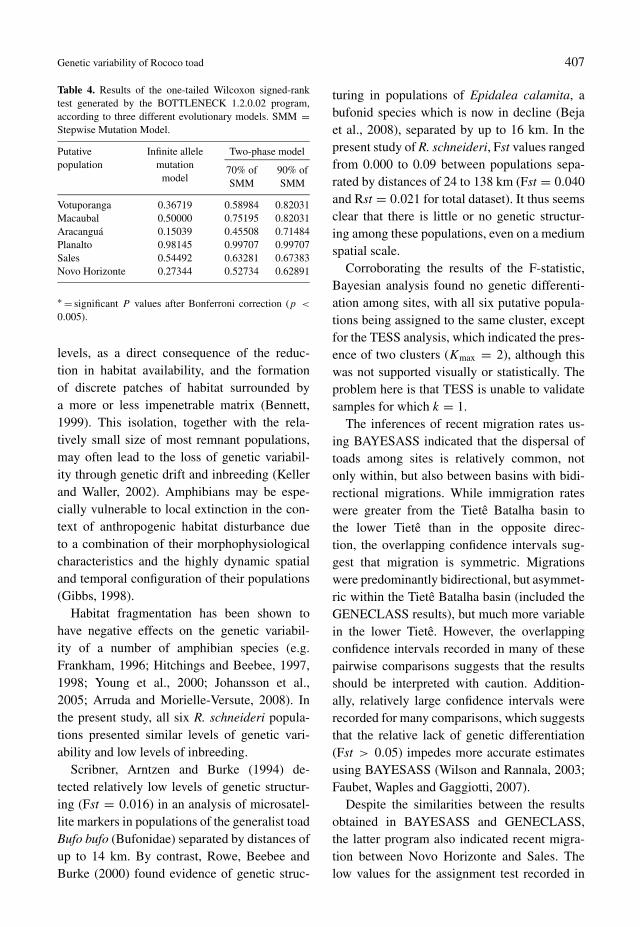
With the exception of Rschµ9, which wasunimodal, all allelic frequencies exhibited a bi-or multimodal distribution. Distributions of thistype are more appropriate for an infinite allelemutation model, although the results with bothmodels were convergent. No evidence of recentpopulation bottlenecks (or founder events) wasfound in any of the six study populations, usingany of the three evolutionary models employed(table 4).
Discussion
Anthropogenic disturbance of natural habitatsappears to be a primary factor in the decline ofamphibian populations at both local and global
Genetic variability of Rococo toad 407
Table 4. Results of the one-tailed Wilcoxon signed-ranktest generated by the BOTTLENECK 1.2.0.02 program,according to three different evolutionary models. SMM =Stepwise Mutation Model.
Putativepopulation
Infinite allelemutation
model
Two-phase model
70% ofSMM
90% ofSMM
Votuporanga 0.36719 0.58984 0.82031Macaubal 0.50000 0.75195 0.82031Aracanguá 0.15039 0.45508 0.71484Planalto 0.98145 0.99707 0.99707Sales 0.54492 0.63281 0.67383Novo Horizonte 0.27344 0.52734 0.62891
∗ = significant P values after Bonferroni correction (p <
0.005).
levels, as a direct consequence of the reduc-tion in habitat availability, and the formationof discrete patches of habitat surrounded bya more or less impenetrable matrix (Bennett,1999). This isolation, together with the rela-tively small size of most remnant populations,may often lead to the loss of genetic variabil-ity through genetic drift and inbreeding (Kellerand Waller, 2002). Amphibians may be espe-cially vulnerable to local extinction in the con-text of anthropogenic habitat disturbance dueto a combination of their morphophysiologicalcharacteristics and the highly dynamic spatialand temporal configuration of their populations(Gibbs, 1998).
Habitat fragmentation has been shown tohave negative effects on the genetic variabil-ity of a number of amphibian species (e.g.Frankham, 1996; Hitchings and Beebee, 1997,1998; Young et al., 2000; Johansson et al.,2005; Arruda and Morielle-Versute, 2008). Inthe present study, all six R. schneideri popula-tions presented similar levels of genetic vari-ability and low levels of inbreeding.
Scribner, Arntzen and Burke (1994) de-tected relatively low levels of genetic structur-ing (Fst = 0.016) in an analysis of microsatel-lite markers in populations of the generalist toadBufo bufo (Bufonidae) separated by distances ofup to 14 km. By contrast, Rowe, Beebee andBurke (2000) found evidence of genetic struc-
turing in populations of Epidalea calamita, abufonid species which is now in decline (Bejaet al., 2008), separated by up to 16 km. In thepresent study of R. schneideri, Fst values rangedfrom 0.000 to 0.09 between populations sepa-rated by distances of 24 to 138 km (Fst = 0.040and Rst = 0.021 for total dataset). It thus seemsclear that there is little or no genetic structur-ing among these populations, even on a mediumspatial scale.
Corroborating the results of the F-statistic,Bayesian analysis found no genetic differenti-ation among sites, with all six putative popula-tions being assigned to the same cluster, exceptfor the TESS analysis, which indicated the pres-ence of two clusters (Kmax = 2), although thiswas not supported visually or statistically. Theproblem here is that TESS is unable to validatesamples for which k = 1.
The inferences of recent migration rates us-ing BAYESASS indicated that the dispersal oftoads among sites is relatively common, notonly within, but also between basins with bidi-rectional migrations. While immigration rateswere greater from the Tietê Batalha basin tothe lower Tietê than in the opposite direc-tion, the overlapping confidence intervals sug-gest that migration is symmetric. Migrationswere predominantly bidirectional, but asymmet-ric within the Tietê Batalha basin (included theGENECLASS results), but much more variablein the lower Tietê. However, the overlappingconfidence intervals recorded in many of thesepairwise comparisons suggests that the resultsshould be interpreted with caution. Addition-ally, relatively large confidence intervals wererecorded for many comparisons, which suggeststhat the relative lack of genetic differentiation(Fst > 0.05) impedes more accurate estimatesusing BAYESASS (Wilson and Rannala, 2003;Faubet, Waples and Gaggiotti, 2007).
Despite the similarities between the resultsobtained in BAYESASS and GENECLASS,the latter program also indicated recent migra-tion between Novo Horizonte and Sales. Thelow values for the assignment test recorded in

408 M.P. Arruda et al.
GENECLASS for many individuals (57%) maybe related to sample size. Moreover, the resultsof the assignment test indicated low levels of ge-netic differentiation across the putative popula-tions, given that most individuals sampled at agiven site were assigned to other localities (e.g.Votuporanga, Macaubal and Sales).
Contemporary gene flow, including first-generation immigrants (in GENECLASS) weredetected in both downstream (e.g. from Salesto Planalto) and upstream directions (e.g. fromSales to Novo Horizonte), and no markedtendency for downstream dispersal (i.e. TietêBatalha → lower Tietê) was recorded in thepresent study. This contrasts with the pattern re-corded in other amphibian species (Kraaijeveld-Smit et al., 2005). Predictably, first generationimmigrants were found in the geographicallyclosest populations (24 to 56 km). Funk et al.(2005) recorded relatively high rates of dis-persal in frogs over longer distances (approxi-mately 5 km), but this contradicts the generalpattern of low vagility in amphibians (Blaustein,Wake and Sousa, 1994; Palo et al., 2004). Giventhis, the results of the present study may notrepresent true first generation migrations, evenassuming high vagility for R. schneideri, butrather movements from intermediate sites orpossibly even a reflection of the considerablehomogeneity in genotype frequencies observedamong sites.
Field studies have reported poor dispersal ca-pacity and a high degree of breeding philopa-try in amphibians (e.g. Driscoll, 1997; Gillette,2003), and molecular studies have indicated lowlevels of gene flow between populations (e.g.Scribner et al., 2001; Kraaijeveld-Smit et al.,2005; Cabe et al., 2007). However, such pat-terns may not be typical of all species (Marshand Ternham, 2001; Berry, 2001; Lampert et al.,2003), given that some genetic studies, includ-ing the present one, have found high levels ofgene flow between geographically distant popu-lations (Seppä and Laurila, 1999; Newman andSquire, 2001). Species vary in their toleranceof habitat fragmentation, and the most vulnera-
ble to disturbance include those with low popu-lation density and fecundity, relatively volatilepopulations, reduced capacity for dispersal andadaptation to specific habitats (Henle et al.,2004). By contrast, R. schneideri is widely-distributed in South America, where it occu-pies a broad variety of habitats (Aquino etal., 2004), and is relatively abundant in an-thropogenic landscapes (Silvano and Pimenta,2003).
The comparison of contemporary and his-torical migration rates indicated no significantrecent shifts in gene flow or migration rates,which might be related to recent anthropogenicprocesses. Moreover, no correlation was ob-served between landscape resistance and ge-netic distance (Fst, Rst) or current gene flow(m), indicating that patterns of genetic differen-tiation and migration among populations are notrelated to distinct forms of land use.
The absence of isolation by distance is inter-preted as a reflection of high gene flow over longdistances (Seppä and Laurila, 1999) or very lowgene flow, even at short distances (Olsen et al.,2003). In the present study, the absence of iso-lation by distance and the relatively high lev-els of migration between populations suggestthat R. schneideri has a considerable capacityfor dispersal, which is responsible for the ge-netic homogenization of populations. Further-more, there was no indication of any decline inthe genetic variability of the populations withinthe study area. Given this, the results of thepresent study indicate that the anthropogenicimpacts within the study area have not had anysignificant effect on the dynamics of the local R.schneideri population, even in the Tietê Batalhabasin, where the dense sugar cane plantationsmight be expected to represent an effective bar-rier to dispersal. While other factors, such asallelic homoplasy or shared ancestral polymor-phisms (or their synergistic effects), may ex-plain such homogeneity, this would have beenlimited in the present case by the relatively largenumber of loci analysed.

Genetic variability of Rococo toad 409
Haddad and Abe (1999) have concluded thatthe geographic range of R. schneideri has beenexpanding in recent years due to deforesta-tion, which makes this species a particularlyuseful model for the study of founder events.However, the analyses presented here failed toidentify any recent event of this type, which in-dicates that the populations represent the orig-inal geographical distribution of R. schneiderior have a relatively long history in the region.The latter hypothesis is consistent with studiesof the Holocene biogeography of central Brazil(Pessenta et al., 1998; Salgado-Labouriau et al.,1998; Behing, 2003), which indicate successiveexpansions and contractions of savannah andforest habitats associated with the climate fluc-tuations of the Quaternary period (Ledru, 2002).On the other hand, the results of this study maysimply reflect an expansion of the populationso recent, that insufficient time has elapsed forthe occurrence of significance divergence in mi-crosatellite loci. This study also indicates weakfidelity to breeding sites, although additionalfield data will be required to evaluate this pat-tern more systematically.
Acknowledgements. This study was supported by theSão Paulo State Research Foundation (FAPESP) and theBrazilian National Research Council (CNPq). CNPq alsosupported E.C. Gonçalves through grant 152757/2007-4.
References
Aquino, L., Reichle, S., Colli, G., Scott, N., Lavilla, E.,Langone, J. (2004): Rhinella schneideri. In: IUCN 2009.IUCN Red List of Threatened Species. Version 2010.4.www.iucnredlist.org. Accessed on 30 March 2010.
Arruda, M.P., Morielle-Versute, E. (2008): Cytogenetic andrandom amplified polymorphic DNA analysis of Lep-todactylus species from rural and urban environments(Anura, Amphibia). Genet. Mol. Res. 7 (1): 161-176.
Arruda, M.P., Gonçalves, E.G., Schneider, M.P.C., Silva,A.L., Morielle-Versute, E. (2010): An alternative geno-typing method using dye-labeled universal primer toreduce unspecific amplifications. Mol. Biol. Rep. 37:2031-2036.
Australia and New Zealand Council for the Care of Ani-mals in Research and Teaching (ANZCCART) (2001):Euthanasia of Animals Used for Scientific Purposes, 2nd
Edition. Reilly, J.S., Ed., ANZCCART, Australia andNew Zealand.
Bastos, R.P., Motta, J.A.O., Lima, L.P., Guimarães, L.D.(2003): Anfíbios da Floresta Nacional de Silvânia. Goiâ-nia, Estado de Goiás, Brazil.
Batista, R.C., De-Carvalho, C.B., Freitas, E.B., Franco,S.C., Batista, C.C., Coelho, W.A., Faria, R.G. (2011):Diet of Rhinella schneideri (Werner, 1984) (Anura: Bu-fonidae) in the Cerrado, Central Brazil. HerpetologyNotes 4: 17-21.
Beebee, T.J.C. (2005): Conservation genetics of amphib-ians. Heredity 95: 423-427.
Behling, H. (2003): Late glacial and Holocene vegetation,climate and fire history inferred from Lagoa Nova in thesoutheastern Brazilian lowland. Veg. Hist. Archaeobot.12 (4): 263-270.
Beja, P., Kuzmin, S., Beebee, T., Denoël, M., Schmidt,B., Tarkhnishvili, D., Ananjeva, N., Orlov, N., Nys-tröm, P., Ogrodowczyk, A., Ogielska, M., Bosch,J., Miaud, C., Tejedo, M., Lizana, M., Martínez-Solano, I. (2008): Epidalea calamita. In: IUCN 2010.IUCN Red List of Threatened Species. Version 2010.4.www.iucnredlist.org. Accessed on 14 December 2010.
Bennett, A.F. (1999): Linkages in the Landscape: The Roleof Corridors and Connectivity in Wildlife Conservation.IUCN, Gland, Switzerland and Cambridge, UK.
Berry, O. (2001): Genetic evidence for wide dispersal by thesand frog, Heleioporus psammophilus (Anura: Myoba-trachidae), in Western Australia. J. Herpetol. 35: 136-141.
Blaustein, A.R., Wake, D.B., Sousa, W.P. (1994): Amphib-ian declines: judging stability, persistence, and suscep-tibility of populations to local and global extinctions.Conserv. Biol. 8: 60-71.
Bohonak, A.J. (2002): IBD (isolation by distance): a pro-gram for analyses of isolation by distance. J. Hered. 93:154-155.
Cabe, P.R., Page, R.B., Hanlon, T.J., Alrich, M.E., Con-nors, L., Marsh, D.M. (2007): Fine-scale populationdifferentiation and gene flow in a terrestrial salaman-der (Plethodon cinereus) living in continuous habitat.Heredity 98: 53-60.
CETEC (2008): Relatório de situação dos recursos hídricosda bacia hidrográfica do Tietê-Batalha. São Paulo, Es-tado de São Paulo, Brazil.
Chen, C., Durand, E., Forbes, F., François, O. (2007):Bayesian clustering algorithms ascertaining spatialpopulation structure: a new computer program and acomparison study. Mol. Ecol. Notes 7: 747-756.
Cornuet, J.M., Luikart, G. (1996): Description and poweranalysis of two tests for detecting recent populationbottlenecks from allele frequency data. Genetics 144:2001-2014.
Cushman, S.A. (2006): Effects of habitat loss and fragmen-tation on amphibians: a review and prospectus. Biol.Conserv. 128: 231-240.
Driscoll, D.A. (1997): Mobility and metapopulation struc-ture of Geocrinia alba and Geocrinia vitellina, twoendangered frog species from southwestern Australia.Aust. J. Ecol. 22: 185-195.

410 M.P. Arruda et al.
Excoffier, I.G., Schneider, S. (2006): Arlequin 3.1: Anintegrated software package for population genetics dataanalysis. Evolutionary Bioinformatics Online 1: 47-50.
Faubet, P., Waples, R.W., Gaggiotti, O.E. (2007): Evaluatingthe performance of a multilocus Bayesian method for theestimation of migration rates. Mol. Ecol. 16: 1149-1166.
Frankham, R. (1996): Relationship of genetic variation topopulation size in wildlife. Conserv. Biol. 10: 1500-1508.
Funk, W.C., Greene, A.E., Corn, P.S., Allendorf, F.W.(2005): High dispersal in a frog species suggests that it isvulnerable to habitat fragmentation. Biol. Lett. 1: 13-16.
Gardner, T.A., Barlow, J., Peres, C.A. (2007): Paradox, pre-sumption and pitfalls in conservation biology: The im-portance of habitat change for amphibians and reptiles.Biol. Conserv. 138: 166-179.
Gibbs, J. (1998): Distribution of woodland amphibiansalong a forest fragmentation gradient. Landscape Ecol.13: 263-268.
Gillette, J.R. (2003): Population ecology, social behavior,and intersexual differences in a natural population ofred-backed salamanders: a long-term field study. Ph.D.thesis, University of Louisiana.
Goudet, J. (1997): FSTAT: a program to estimate and testgene diversities and fixation indices. Version 2.8. Uni-versité de Lausanne, Switzerland.
Gray, M.J., Smith, L.M., Brenes, R. (2004): Effects ofagricultural cultivation on demographics of southernhigh plains amphibians. Conserv. Biol. 18: 1368-1377.
Guillot, G., Santos, F., Estoup, A. (2008): Analysing geo-referenced population genetics data with GENELAND:a new algorithm to deal with null alleles and a friendlygraphical user interface. Bioinformatics 24: 1406-1407.
Haddad, C.F.B., Abe, A.S. (1999): Anfíbios e Répteis. In:Workshop Mata Atlântica e Campos Sulinos.
Henle, K., Davies, K.F., Kleyer, M., Margules, C., Settele,J. (2004): Predictors of species sensitivity to fragmenta-tion. Biodivers. Conserv. 13: 207-251.
Hitchings, S.P., Beebee, T.J.C. (1997): Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations: implications forbiodiversity conservation. Heredity 79: 117-121.
Hitchings, S.P., Beebee, T.J.C. (1998): Loss of genetic di-versity and fitness in common toad (Bufo bufo) popula-tions isolated by inimical habitat. J. Evolution. Biol. 11:269-283.
IBGE (2010): Mapa da cobertura e uso da terra. Brasília,Distrito Federal, Brazil.
Jakobsson, M., Rosenberg, N. (2007): CLUMPP: a clustermatching and permutation program for dealing with la-bel switching and multimodality in analysis of popula-tion structure. Bioinformatics 23: 1801-1806.
Johansson, M., Primmer, C.R., Sahlsten, J., Merila, J.(2005): The influence of landscape structure on occur-rence, abundance and genetic diversity of the commonfrog, Rana temporaria. Glob. Change Biol. 11: 1664-1679.
Keller, L.F., Waller, D.M. (2002): Inbreeding effects in wildpopulations. Trends Ecol. Evol. 17: 230-241.
Knutson, M.G., Sauer, J.R., Olsen, D.A., Mossman, M.J.,Hemesath, L.M., Lannoo, M.J. (1999): Effects of land-scape composition and wetland fragmentation on frogand toad abundance and species richness in Iowa andWisconsin, U.S.A. Conserv. Biol. 13 (6): 1437-1446.
Kraaijeveld-Smit, F.J.L., Beebee, T.J.C., Griffiths, R.A.,Moore, R.D., Schley, L. (2005): Low gene flow but highgenetic diversity in the threatened Mallorcan midwifetoad Alytes muletensis. Mol. Ecol. 14: 3307-3315.
Kronka, F.J.N., Matsukuma, C.K., Nalon, M.A., Aurelio,M. (1993): Inventário florestal do Estado de São Paulo.Instituto Florestal, Estado de São Paulo, Brazil.
Lampert, K.P., Rand, A.S., Mueller, U.G., Ryan, M.J.(2003): Fine-scale genetic pattern and evidence for sex-biased dispersal in the túngara frog, Physalaemus pustu-losus. Mol. Ecol. 12: 3325-3334.
Ledru, M.P. (2002): Late Quaternary history and evolutionof the cerradões as revealed by palynological records. In:The Cerrado of Brazil: Ecology and Natural History of aNeotropical Savanna, p. 33-50. Oliveira, P.S., Marquis,R.J., Eds, Columbia University Press, New York.
Marsh, D.M., Trenham, P.C. (2001): Metapopulation dy-namics and amphibian conservation. Conserv. Biol. 15:40-49.
McRae, B.H. (2006): Isolation by resistance. Evolution 60:1551-1561.
Molecular Ecology Resources Primer Development Con-sortium (2009): Permanent genetic resources added toMolecular Ecology Resources Database 1 May 2009-31July. Mol. Ecol. Resour. 9 (6): 1460-1466.
Newman, R.A., Squire, T. (2001): Microsatellite variationand finescale population structure in the wood frog(Rana sylvatica). Mol. Ecol. 10: 1087-1100.
Olsen, J.B., Miller, S.J., Spearman, W.J., Wenburg, J.K.(2003): Patterns of intra- and inter-population genetic di-versity in Alaskan coho salmon: Implications for conser-vation. Conserv. Genet. 4: 557-569.
Paetkau, D., Slade, R., Burden, M., Estoup, A. (2004): Ge-netic assignment methods for the direct, real-time esti-mation of migration rate: a simulation-based explorationof accuracy and power. Mol. Ecol. 13: 55-65.
Palo, J.U., Schmeller, D., Laurila, A., Primmer, C.R.,Kuzmins, S.I., Merila, J. (2004): High degree of popula-tion subdivision in a widespread amphibian. Mol. Ecol.13: 2631-2644.
Pessenda, L.C.R., Gomes, B.M., Aravena, R., Ribeiro, A.S.,Boulet, R., Gouveia, S.E.M. (1998): The carbon isotoperecord in soils along a Forest-cerrado ecosystem tran-sect: implication for vegetation changes in the Rondo-nia state, southwestern Brazilian Amazon region. TheHolocene 8 (5): 599-603.
Piry, S., Alapetite, A., Cornuet, J.M., Paetkau, D., Bau-douin, L., Estoup, A. (2004): Geneclass 2: a softwarefor genetic assignment and firstgeneration migrant de-tection. J. Hered. 95: 536-539.
Pritchard, J.K., Stephens, M., Donnelly, P. (2000): Inferenceof population structure using multilocus genotype data.Genetics 155: 945-959.

Genetic variability of Rococo toad 411
Purrenhage, J.L., Niewiarowski, P.H., Moore, F.B.G.(2009): Population structure of spotted salamanders(Ambystoma maculatum) in a fragmented landscape.Mol. Ecol. 18 (2): 235-247.
Rannala, B., Mountain, J.L. (1997): Detecting immigrationby using multilocus genotypes. Proc. Natl. Acad. Sci.USA 94: 9197-9221.
Rice, W.R. (1989): Analyzing tables of statistical tests.Evolution 43: 223-225.
Rosenberg, N.A. (2004): Distruct: a program for the graph-ical display of population structure. Mol. Ecol. Notes 4:137-138.
Rousset, F. (1997): Genetic differentiation and estimation ofgene flow from F -statistics under isolation by distance.Genetics 145: 1219-1228.
Rousset, F. (2008): GENEPOP’007: a complete re-implementation of the genepop software for Windowsand Linux. Mol. Ecol. Resour. 8: 103-106.
Rowe, G., Beebee, T.J.C., Burke, T. (2000): A microsatelliteanalysis of natterjack toad, Bufo calamita, metapopula-tions. Oikos 88: 641-651.
Salgado-Labouriau, M.L., Barbieri, M., Ferrazvicentini,K.R., Parizii, M.G. (1998): A dry climatic event duringthe late Quaternary of tropical Brazil. Rev. Palaeobot.Palyno. 99: 115-129.
Sambrook, J., Fritsch, E.F., Maniatis, T. (1989): MolecularCloning – A Laboratory Manual, 2nd Edition. ColdSpring Harbor Laboratory Press, USA.
Schlötterer, C. (2000): Evolutionary dynamics of mi-crosatellite DNA. Chromosoma 109: 365-371.
Scribner, K.T., Arntzen, J.W., Burke, T. (1994): Compar-ative analysis of intra- and interpopulation genetic di-versity in Bufo bufo, using allozyme, single-locus mi-crosatellite, minisatellite, and multilocus minisatellitedata. Mol. Biol. Evol. 11: 737-748.
Scribner, K.T., Arntzen, J.W., Burke, T., Cruddace, N.,Oldham, R.S. (2001): Environmental correlates of toadabundance and population genetic diversity. Biol. Con-serv. 98: 201-210.
Seppä, P., Laurila, A. (1999): Genetic structure of islandpopulations of the anurans Rana temporaria and Bufobufo. Heredity 82: 309-317.
Silvano, D.L., Pimenta, B.V.S. (2003): Diversidade e dis-tribuição de anfíbios anuros na Mata Atlântica do sul daBahia. In: Corredor de Biodiversidade na Mata Atlân-tica do Sul da Bahia. Prado, P.I., Landau, E.C., Moura,R.T., Pinto, L.P.S., Fonseca, G.A.B., Alger, K., Orgs,IESB/CI/CABS/UFMG/UNICAMP, Ilhéus, Brazil. Pub-lished on CD-ROM.
Stevens, V.M., Leboulengé, E., Wesselingh, R.A., Baguette,M. (2006): Quantifying functional connectivity: experi-mental assessment of boundary permeability for the nat-terjack toad (Bufo calamita). Oecologia 150: 161-171.
Urbina-Cardona, J.N., Olivarez-Pérez, M., Reynoso, V.H.(2006): Herpetofauna diversity and microenvironmentcorrelates across a pasture-edge-interior ecotone in trop-ical rainforest fragments in the Los Tuxtlas BiosphereReserve of Veracruz, México. Biol. Conserv. 132: 61-75.
Vasconcellos, M.M., Colli, G.R. (2009): Factors affectingthe populations dynamics of two toads (Anura: Bu-fonidae) in a seasonal neotropical savanna. Copeia 2009(2): 266-276.
Whiteley, A.R., Spruell, P., Allendorf, F.W. (2006): Cancommon species provide valuable information for con-servation? Mol. Ecol. 35: 2767-2786.
Wilson, G., Rannala, B. (2003): Bayesian inference of re-cent migration rates using multilocus genotypes. Gene-tics 163: 1177-1191.
Yeh, F.C., Yang, R., Boylet, T. (1997): POPGENE Version1.32: Software Microsoft Window-Based Freeware forPopulation Genetic Analysis. University of Alberta, Al-berta, Canada.
Young, A.G., Brown, A.D.H., Murray, B.G., Thrall, P.H.,Miller, C.H. (2000): Genetic erosion, restricted matingand reduced viability in fragmented populations of theendangered grassland herb Rutidosis leptorrynchoides.In: Genetics, Demography and Viability of FragmentedPopulations, p. 335-359. Young, A.G., Clarke, G.M.,Eds, Cambridge University Press, Cambridge, UK.
Received: January 16, 2011. Accepted: July 7, 2011.
Related Documents











