Construction of a prototype two-component system from the phosphorelay system TodS/TodT Hortencia Silva-Jime ´nez, Juan Luis Ramos and Tino Krell 1 Department of Environmental Protection, Estacio ´n Experimental del Zaidı ´n, Consejo Superior de Investigaciones Cientı ´ficas, C/ Prof. Albareda, 1, 18008 Granada, Spain 1 To whom correspondence should be addressed. E-mail: [email protected] Received June 29, 2011; revised December 15, 2011; accepted December 30, 2011 Edited by Feng Ni Two-component systems (TCSs) play key roles in the adaptation of bacteria to environmental changes. In prototype TCSs a single phosphoryl transfer between the sensor kinase and response regulator occurs, whereas phosphorelay TCSs are characterised by a His1–Asp1– His2 – Asp2 phosphorylation cascade. The TodS/TodT TCS controls the expression of a toluene degradation pathway and the TodS sensor kinase operates by a three- step internal phosphorelay. Based on TodS we report the construction of a minimal form of TodS, termed as Min- TodS, that contains only three of the seven TodS domains. Min-TodS is composed of the N-terminal PAS sensor domain as well as the C-terminal dimerisation/ phosphotransfer domain and catalytic domain of TodS. We have conducted a comparative analysis of the phos- phorelay TCS with its prototypal derivative. We demon- strate that Min-TodS binds effector molecules with affinities comparable with those observed for TodS. Min- TodS forms a TCS with TodT and toluene increases the amount of TodT-P. In contrast to TodS, toluene does not stimulate Min-TodS autophosphorylation. The half-life of Min-TodS-P was significantly increased as compared with TodS. Analysis of TodSD500A revealed that the hydroly- sis of the acylphosphate of the receiver domain is respon- sible for the reduced half-life of TodS. The regulation of P todX expression by Min-TodS/TodT and TodS/TodT in response to different effectors are compared. The Min- TodS/TodT system was characterized by a higher basal activity but a lower magnitude of response. Data will be discussed in the context that the phosphorelay system appears to be better suited for the control of a degrad- ation pathway for toxic compounds. Keywords: chimeric protein/phosphorelay/sensor kinase/ signal transduction/two-component system Introduction Bacteria have to constantly adapt to changes in the environment to guarantee survival. An important mechanism by which bacteria sense environmental signals is through the action of two-component systems (TCSs) that contain a sensor kinase (SK) and a response regulator (RR) as basic elements. Bacteria contain a large number of TCSs and were found to harbour on average 52 TCS genes (Cock and Whitworth, 2007). TCSs are involved in the regulation of virtually all types of processes including virulence, sporulation, metabolism, quorum sensing, chemotaxis, transport or nitrogen fixation (reviewed in Mascher et al., 2006; Krell et al., 2010). Within the family of TCSs, prototype and phosphorelay systems can be distinguished. Typically, a prototype SK is composed of a sensor domain and an autokinase (AK) module. This module is composed of a dimerisation and phosphotransfer domain (DHp) that harbours the phosphory- laccepting histidine residue and a catalytic domain (CA) to which ATP binds and which catalyses the phosphorylation of the histidine residue. The phosphorylated SK donates phos- phoryl groups to the RR receiver domain, which in turn alters the functional properties of the RR output domain. This domain often possesses DNA-binding activity and alters gene expression by binding to promoter regions. Other RR output domains exert their activity by binding to proteins or possess enzymatic activities (Galperin, 2010). Apart from the SK–RR phosphotransfer activity, the majority of SKs possess kinase as well as phosphatase activ- ities towards the RR (reviewed by Huynh and Stewart, 2011) Well-studied examples of SKs with dual-kinase/phosphatase activity are RcsB (Majdalani and Gottesman, 2005; Fredericks et al., 2006), DesK (Albanesi et al., 2009), FixL (Lois et al., 1993), AbsA1 (Sheeler et al., 2005) or PhoQ (Chamnongpol et al., 2003). In a number of cases it has been demonstrated that the signal modulates both activities, which is exemplified by the concerted modulation of both activities in FixL or DesK by oxygen or temperature, respectively (Lois et al., 1993; Albanesi et al., 2009). Only a few SKs have kinase activities only and in those cases auxiliary phos- phatases were found to actively dephosphorylate the RR (Huynh and Stewart, 2011). Instead of the simple His–Asp transphosphorylation of prototype systems, phosphorelay TCSs use a His1–Asp1– His2–Asp2 phosphorylation cascade (Zhang and Shi, 2005, Fig. 1). His1 forms part of an AK module and Asp1 and Asp2 are present on receiver domains. Genome analyses (Williams and Whitworth, 2010) indicate that there might be two major classes of phosphorelay systems depending on whether His2 forms part of a histidine-containing phospho- transfer domain (systems termed as TRPR) or part of a second AK module (systems termed as TRTR). Phosphorelay systems of the TRPR family have been characterised exten- sively and representative examples are the ArcBA, TorSR and EvgSA systems of Escherichia coli (Perraud et al., 1999; Malpica et al., 2006), the BvgST system of Bordetella # The Author 2012. Published by Oxford University Press. All rights reserved. For Permissions, please e-mail: [email protected] 159 Protein Engineering, Design & Selection vol. 25 no. 4 pp. 159–169, 2012 Published online February 3, 2012 doi:10.1093/protein/gzs001 at CSIC on April 8, 2012 http://peds.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Construction of a prototype two-component systemfrom the phosphorelay system TodS/TodT
Hortencia Silva-Jimenez, Juan Luis Ramosand Tino Krell1
Department of Environmental Protection, Estacion Experimental del Zaidın,Consejo Superior de Investigaciones Cientıficas, C/ Prof. Albareda, 1,18008 Granada, Spain
1To whom correspondence should be addressed.E-mail: [email protected]
Received June 29, 2011; revised December 15, 2011;accepted December 30, 2011
Edited by Feng Ni
Two-component systems (TCSs) play key roles in theadaptation of bacteria to environmental changes. Inprototype TCSs a single phosphoryl transfer between thesensor kinase and response regulator occurs, whereasphosphorelay TCSs are characterised by a His1–Asp1–His2–Asp2 phosphorylation cascade. The TodS/TodTTCS controls the expression of a toluene degradationpathway and the TodS sensor kinase operates by a three-step internal phosphorelay. Based on TodS we report theconstruction of a minimal form of TodS, termed as Min-TodS, that contains only three of the seven TodSdomains. Min-TodS is composed of the N-terminal PASsensor domain as well as the C-terminal dimerisation/phosphotransfer domain and catalytic domain of TodS.We have conducted a comparative analysis of the phos-phorelay TCS with its prototypal derivative. We demon-strate that Min-TodS binds effector molecules withaffinities comparable with those observed for TodS. Min-TodS forms a TCS with TodT and toluene increases theamount of TodT-P. In contrast to TodS, toluene does notstimulate Min-TodS autophosphorylation. The half-life ofMin-TodS-P was significantly increased as compared withTodS. Analysis of TodSD500A revealed that the hydroly-sis of the acylphosphate of the receiver domain is respon-sible for the reduced half-life of TodS. The regulation ofPtodX expression by Min-TodS/TodT and TodS/TodT inresponse to different effectors are compared. The Min-TodS/TodT system was characterized by a higher basalactivity but a lower magnitude of response. Data will bediscussed in the context that the phosphorelay systemappears to be better suited for the control of a degrad-ation pathway for toxic compounds.Keywords: chimeric protein/phosphorelay/sensor kinase/signal transduction/two-component system
Introduction
Bacteria have to constantly adapt to changes in the environmentto guarantee survival. An important mechanism by which
bacteria sense environmental signals is through the action oftwo-component systems (TCSs) that contain a sensor kinase(SK) and a response regulator (RR) as basic elements. Bacteriacontain a large number of TCSs and were found to harbour onaverage 52 TCS genes (Cock and Whitworth, 2007). TCSs areinvolved in the regulation of virtually all types of processesincluding virulence, sporulation, metabolism, quorum sensing,chemotaxis, transport or nitrogen fixation (reviewed in Mascheret al., 2006; Krell et al., 2010).
Within the family of TCSs, prototype and phosphorelaysystems can be distinguished. Typically, a prototype SK iscomposed of a sensor domain and an autokinase (AK)module. This module is composed of a dimerisation andphosphotransfer domain (DHp) that harbours the phosphory-laccepting histidine residue and a catalytic domain (CA) towhich ATP binds and which catalyses the phosphorylation ofthe histidine residue. The phosphorylated SK donates phos-phoryl groups to the RR receiver domain, which in turnalters the functional properties of the RR output domain.This domain often possesses DNA-binding activity and altersgene expression by binding to promoter regions. Other RRoutput domains exert their activity by binding to proteins orpossess enzymatic activities (Galperin, 2010).
Apart from the SK–RR phosphotransfer activity, themajority of SKs possess kinase as well as phosphatase activ-ities towards the RR (reviewed by Huynh and Stewart, 2011)Well-studied examples of SKs with dual-kinase/phosphataseactivity are RcsB (Majdalani and Gottesman, 2005;Fredericks et al., 2006), DesK (Albanesi et al., 2009), FixL(Lois et al., 1993), AbsA1 (Sheeler et al., 2005) or PhoQ(Chamnongpol et al., 2003). In a number of cases it has beendemonstrated that the signal modulates both activities, whichis exemplified by the concerted modulation of both activitiesin FixL or DesK by oxygen or temperature, respectively(Lois et al., 1993; Albanesi et al., 2009). Only a few SKshave kinase activities only and in those cases auxiliary phos-phatases were found to actively dephosphorylate the RR(Huynh and Stewart, 2011).
Instead of the simple His–Asp transphosphorylation ofprototype systems, phosphorelay TCSs use a His1–Asp1–His2–Asp2 phosphorylation cascade (Zhang and Shi, 2005,Fig. 1). His1 forms part of an AK module and Asp1 andAsp2 are present on receiver domains. Genome analyses(Williams and Whitworth, 2010) indicate that there might betwo major classes of phosphorelay systems depending onwhether His2 forms part of a histidine-containing phospho-transfer domain (systems termed as TRPR) or part of asecond AK module (systems termed as TRTR). Phosphorelaysystems of the TRPR family have been characterised exten-sively and representative examples are the ArcBA, TorSRand EvgSA systems of Escherichia coli (Perraud et al.,1999; Malpica et al., 2006), the BvgST system of Bordetella
# The Author 2012. Published by Oxford University Press. All rights reserved.
For Permissions, please e-mail: [email protected]
159
Protein Engineering, Design & Selection vol. 25 no. 4 pp. 159–169, 2012Published online February 3, 2012 doi:10.1093/protein/gzs001
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

sp. (Beier and Gross, 2008) or the sporulation phosphorelayof Bacillus subtilis (Burbulys et al., 1991). Although thegenetic abundance of TRPR and TRTR systems is similar(Williams and Whitworth, 2010), the wealth of informationavailable on TRPR systems contrasts with the scarceness ofdata on the TRTR systems.
Work in our laboratory on the TodS/TodT system has led tothe first functional characterisation of a TRTR-type phosphor-elay (Lacal et al., 2006; Busch et al., 2007, 2009). The TodS/TodT TCS modulates the expression of the tod (toluene diox-ygenase) operon (Lau et al., 1997), which encodes theenzymes of the TOD pathway that permits the mineralisationof benzene, toluene and ethylbenzene in strains ofPseudomonas putida. The tod operon is under control of thepromoter PtodX (Zylstra and Gibson, 1989; Mosqueda et al.,1999). Although the TOD pathway uses only three substrates(toluene, benzene and ethylbenzene), TodS/TodT mediatesthe regulation of PtodX activity in response to a wide range ofdifferent aromatic compounds, including different methyl-,amino-, nitro- and halogen-substituted benzene and toluenederivatives (Lacal et al., 2006). The TodS SK is unusuallylarge (108 kDa) and composed of the following sevendomains (Fig. 1): PAS sensor 1, DHp 1, CA 1, receiver, PASsensor 2, DHp2 and CA 2. Effector molecules like toluenebind to the N-terminal PAS domain, which stimulates thekinase activity of the N-terminal AK module (Busch et al.,2009). The phosphoryl group is then transferred to Asp500 ofthe receiver domain and subsequently to the C-terminal AKmodule (Fig. 1). The phosphoryl group then passes fromHis760 of TodS to Asp57 of the TodT RR (Busch et al.,2009). We have shown that TodT binds to five different sitesat PtodX in a sequential and cooperative manner (Lacal et al.,2008a,b). In addition, we have demonstrated that the activityof TodS/TodT is mediated by agonists and antagonists (Buschet al., 2007). Both classes of compounds compete for bindingat the PAS sensor 1 domain, but only agonists cause anincrease in the TodS phosphorylation state. Owing to theircompetition for binding, the presence of antagonists reduces
the magnitude of agonist-mediated upregulation of geneexpression. Agonists and antagonists are structurally verysimilar (i.e. toluene and o-xylene, respectively) and are bothpresent in mixtures such as petroleum. A major limitation inbiodegradation consists in the insufficient induction ofdegradation pathway expression in bacteria exposed tocomplex mixtures (Cases and de Lorenzo, 2005). The datapresented (Busch et al., 2007) are consistent with the notionthat the action of antagonists might be at least partiallyresponsible for this reduced pathway induction. The TodS/TodT system is the best-characterised member of a proteinfamily. Other family members are the TCSs StyS/StyR(Velasco et al., 1998; Leoni et al., 2003), TutC/TutB(Coschigano and Young, 1997) and TmoS/TmoT(Ramos-Gonzalez et al., 2002), which are all involved in theregulation of degradation pathways. In all cases the SKs sharethe same domain arrangement as TodS.
There is now evidence that phosphorelay TCSs haveevolved from a fusion of prototype TCSs (Zhang and Shi,2005; Whitworth and Cock, 2009). The fusion of prototypeTCS to phosphorelay TCSs was found to be a continuousevolutionary process. Phosphorelay systems can be found inmost of the phylogenetic clades of TCS and are present inone-third of bacterial genomes (Zhang and Shi, 2005;Whitworth and Cock, 2009). The physiological role of phos-phorelay TCSs is still unclear but it was suggested that amodulation of the additional phosphorylation steps mightoffer the possibility of integrating additional environmentalstimuli leading to a fine tuning of the final response(Burbulys et al., 1991). Lau et al. (1997) have suggested thatone reason for the complexity of TodS consists in thecapacity to integrate additional signals. Based on a weaksequence similarity of the PAS2 domain with the FixLsensor domain the authors proposed that TodS might beable to sense oxygen. However, so far no evidence whichsupports this hypothesis has been reported.
It remains poorly understood as to why some cellular func-tions are controlled by prototype systems whereas others by
Fig. 1. Domain organisation and mode of action of TodS and Min-TodS. (A) Domain organisation of the prototypal TCS PhoQ/PhoP. For further informationrefer to Prost and Miller (2008). (B) Domain organisation and mode of action of the TodS/TodT TCS. The mode of transphosphorylation is illustrated. (C)Domain organisation of the TCS composed of Min-TodS and TodT. Tm, transmembrane region; PAS, PAS domain; AK, autokinase module; RRR, responseregulator receiver domain; DNA-b, DNA-binding domain; DHp, dimerisation and phosphotransfer domain; Ca; catalytic domain. As indicated an AK moduleis composed of a DHp and a Ca domain. The phosphorylgroup accepting residues are labelled.
H.Silva-Jimenez et al.
160
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

phosphorelay TCS. Based on the known mechanism of actionof TodS, namely the fact that aromatic effectors bind to thePAS1 domain, that there is an intramolecular phosphorelayand that phosphorylation of TodT occurs exclusively from theAK2 module, we have constructed a minimal version ofTodS, termed as Min-TodS (Fig. 1), which possesses onlythree of the seven TodS domains and which can be consideredas a prototypal SK. Min-TodS was found to form a TCS withTodT that is capable of regulating expression from PtodX inresponse to agonists. Different in vitro and in vivo analyses,however, revealed that the Min-TodS/TodT system differsfrom the TodS/TodT system in a number of features.
Materials and methods
Strains and plasmidsThe strains and plasmids used in this study are listed inTable I.
Construction of plasmids containing min-todSPlasmids pMIR66-Min-TodS and pET28b-Min-TodS wereconstructed for b-galactosidase measurements and proteinexpression, respectively. The gene coding for Min-TodS was
constructed by three different polymerase chain reactions(PCRs). The initial two PCR reactions were carried out usingpMIR66 (Table I) as template, which contains the todSsequence cloned into the BamHI and NheI sites ofthe vector. The first PCR reaction (amplification of theN-terminal fragment of Min-TodS) involved an amplificationof a fragment covering the BamHI restriction site of pMIR66until the codon encoding amino acid 182 of TodS usingprimers PAS1f and PAS1r (Table II). The latter primer con-tains an extension (shown in italics in Table II) that iscomplementary to the sequence encoding amino acids 753–757; the initial sequence of the C-terminal fragment ofMin-TodS. In a second PCR (amplification of the C-terminalfragment of Min-TodS) a fragment comprising amino acids753–978 and covering the NheI site of plasmid pMIR66 wasamplified using primers AK2f and AK2r. Both PCR productsoverlap by 15 nucleotides. A third PCR contained equimolaramounts of the first two PCR products as well as primersPAS1f and AK2r. This reaction produced a DNA fragmentcoding for Min-TodS flanked by extensions containing theBamHI and NheI restriction sites, which permitted cloninginto pMIR66 linearised with the same enzymes. The result-ing plasmid was named pMIR66-Min-TodS.
Table I. Bacterial strains and plasmids used
Strain/plasmid Relevant characteristics Reference
StrainsE.coli BL21 (DE3) F2, ompI, hsdSB (r2
B m2B ) Studier and Moffat (1986)
P.putida DOT-T1E Tolþ, wild type Ramos et al. (1995)P.putida DOT-T1E DtodST DOT-T1E, todST::Km, Tol2 Ramos-Gonzalez et al. (2002)PlasmidspMIR66 GmR, containing the todST genes Ramos-Gonzalez et al. (2002)pMIR66-Min-TodS pMIR66 derivative, containing the min-TodS gene instead of the todS gene This workpMIR77 TcR, PtodX ::0lacZ Ramos-Gonzalez et al. (2002)pET28b Protein expression plasmid NovagenpET28b-Min-TodS pET28b derivative containing the min-TodS This workpTodS pET28b derivative containing todS Lacal et al. (2006)pAB2 pET28b derivative containing todS mutant D500A Busch et al. (2009)pJLTodT pET28b derivative containing todT Lacal et al. (2006)pJLC1 pMIR66, todS::aphA3 Lacal et al. (2006)pET28b-PAS1long pET28b containing TodS amino acids 1-146 This workpET28b-PAS1short pET28b containing TodS amino acids 15-146 This workpET28b-PAS2 pET28b containing TodS amino acids 609-729 This workpET28b-AK1 pET28b containing TodS amino acids 147-449 This work
Table II. Oligonucleotides used in this study
Name Sequence plasmid
PAS1f 50-TAGAACTAGTGGATCCTCTAGAGTCG-30 pMIR66-Min-TodSPAS1r 50-ATAGGCTGGAAATTCACTCTTCGCATT-30 pMIR66-Min-TodSAK2f 50-GAATTTCCAGCCTATATTGCACAC-30 pMIR66-Min-TodSAK2r 50-TCAGGAATACGCTAGCGGATGA-30 pMIR66-Min-TodSPAS1f-1 50-GTGAGTCATTACATATGAGCTCCTTGG-30 pET28b-Min-TodSAK2r-1 50-AGATGCCCGGATCCTCATGTG-30 pET28b-Min-TodSPAS1longf 50-GTGAGTCATTACATATGAGCTCCTTGG-30 pET28b-PAS1longPAS1longr 50-TCGGTAATGGATCCTCATTCCGCAAGAAGG-30 pET28b-PAS1longPAS1shortf 50-GCCTCAAAATAGACATATGAATAATTATT-30 pET28b-PAS1shortPAS1shortr 50-TATCGGTAATGGATCCCTATTCCGCAAGA-30 pET28b-PAS1shortPAS2f 50-GCAGAGTAACATCCATATGTCCGCCTCGG-30 pET28b-PAS2PAS2r 50-GTTAAGATTTTCGGATCCCTATTTCTTCTCAG-30 pET28b-PAS2AK1f 50-TTACCTTCTTCATATGAGGGCGCAATA-30 pET28b-AK1AK1r 50-CGATTAGCACGGATCCCTAGTCGGATTGAAATGG-30 pET28b-AK1
Restriction sites are highlighted in bold. Complementary and overlapping regions in the construction of Min-TodS are shown in italics.
Conversion of phosphorelay into prototype TCS
161
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

A similar strategy was used to generate pET28b-Min-TodS, except that primers PAS1f and AK2r were substitutedby primers PAS1f-1 and AK2r-1 that contain NdeI andBamHI cloning sites, respectively, permitting cloning intopET28b(þ) (Novagen) linearised with the same enzymes.The resulting plasmid was named pET28b-Min-TodS. Allplasmids constructed were verified by DNA sequencing ofthe insert and flanking regions. Protein expression from thisplasmid gives rise to a fusion protein of Min-TodS flankedby the N-terminal sequence MGSSHHHHHHSSGLVPRGSHcontaining the histidine tag.
Construction of expression plasmids containing the individualPAS domains and the AK1 moduleThe following four expression plasmids were constructedcontaining the individual PAS domains or the AK1 modules:pET28b-PAS1long (TodS amino acids 1–146),pET28b-PAS1short (TodS amino acids 15–146),pET28b-PAS2 (TodS amino acids 609–729) andpET28b-AK1 (TodS amino acids 147–449). In all cases thecorresponding fragments were amplified by PCR usingpMIR66 as the template. Primers were designed to introduceNdeI and BamHI restriction sites, which permits cloning ofthe resulting products into pET28b(þ) (Novagen) linearisedwith the same enzymes. The following primers were used forthe PCR reactions: plasmid pET28b-PAS1long: PAS1longfand PAS1longr; plasmid pET28b-PAS1short: PAS1shortfand PAS1longr; plasmid pET28b-PAS2: PAS2f and PAS2r;plasmid pET28b-AK1: AK1f and AK1r. Protein expressionfrom this plasmid gives rise to a fusion protein of eachfragment flanked by an N-terminal sequenceMGSSHHHHHHSSGLVPRGSH containing a histidine tag.All plasmids constructed were verified by DNA sequencingof the insert and flanking regions.
Overexpression and purification of min-TodS undernon-denaturing conditionsEscherichia coli BL21 (DE3) was transformed with plasmidpET28b-Min-TodS. Cultures were grown in 2 l erlenmeyerflasks containing 500 ml of LB medium supplemented with50 mg/ml kanamycin at 308C until an OD660 of 0.6, at whichpoint protein production was induced by adding 0.1 mMisopropyl-beta-D-thiogalactopyranoside. Growth was contin-ued at 168C overnight prior to cell harvest by centrifugationat 10 000 � g for 30 min. Cell pellets were resuspended inbuffer A (20 mM Tris, 0.1 mM EDTA, 500 mM NaCl,10 mM imidazole, 5 mM b-mercaptoethanol and 5% (vol/vol) glycerol, pH 8.0) and broken using a French press at1000 psi. After centrifugation at 20 000 � g for 1 h, thesupernatant was loaded onto a 5 ml HisTrap column(Amersham Bioscience), washed with 10 column volumes ofbuffer A and eluted with an imidazole gradient of 45–500 mM in buffer A. Fractions containing Min-TodS weredialysed against analysis buffer 50 mM Tris pH 7.5, 300 mMKCl, 2 mM MgCl2, 0.1 mM EDTA, 2 mM DTT and 10%(vol/vol) glycerol, for immediate analysis.
Purification of Min-TodS under denaturing conditionsand protein refoldingMin-TodS was expressed and purified as described above,except that the both purification buffers were supplementedwith 6 M Guanidine-HCl. Eluted protein was pooled, diluted
to 0.2 mg/ml and dialysed against analysis buffer containing2 M urea and subsequently against two changes of analysisbuffer. Protein was then filtered and concentrated for imme-diate analysis.
Overexpression and purification of TodS, TodSD500Aand TodTTodS and TodS D500A were purified as described in Lacalet al. (2006). TodT was expressed and purified as describedin Lacal et al. (2008a).
Isothermal titration calorimetryIsothermal titration calorimetry (ITC) experiments wereconducted using freshly purified protein and aVP-microcalorimeter (Microcal, Amherst, MA) at 258C.Protein was dialysed into analysis buffer (50 mM Tris-HCl,300 mM KCl, 2 mM MgCl2, 2 mM DTT, 0.1 mM EDTA,10% glycerol, pH 7.5) and placed into the sample cell.Typically, 5–10 mM Min-TodS was titrated with 350–500 mM effector solutions. Since these compounds are vola-tile and hydrophobic, these solutions were made in glassvessels immediately before use. The mean enthalpiesmeasured from the injection of effectors into the buffer weresubtracted from raw titration data prior to data analysis withthe MicroCal version of ORIGIN. Data were fitted with the‘One binding site model’ of ORIGIN.
Autophosphorylation assayTen micromolar of TodS or Min-TodS in 50 mM Tris-HCl,pH 7.5, 300 mM KCl, mM MgCl2, 0.1 mM EDTA, 10%(vol/vol) glycerol, 2 mM DTT were incubated at 48C with200 mM ATP containing 4 mCi [g32P]ATP in the absence orpresence of 100 mM toluene. At time intervals, samples weretaken, the reaction being stopped by the addition of 4 � SDSsample buffer and then stored at 2208C. After the comple-tion of the time course samples were analysed bySDS–PAGE using 10% (wt/vol) gels. Protein-associatedradioactive phosphorylgroups were visualised on aphosphoimager.
Transphosphorylation assayDuplicate samples of Min-TodS were autophosphorylated inthe absence or presence of 100 mM toluene. This reactionwas carried out in 50 mM Tris-HCl, pH 7.5, 300 mM KCl,mM MgCl2, 0.1 mM EDTA, 10% (vol/vol) glycerol and2 mM DTT at 48C. The reaction volume was 10 ml and con-tained 10 mM of purified Min-TodS. Reactions were initiatedby the addition of radio-labelled ATP (200 mM ATP contain-ing 4 mCi [g32P]ATP). After 50 min of autophosphorylation6 ml of 20 mM purified TodT was added to one of the dupli-cate samples, whereas the corresponding amount of bufferwas added to the remaining sample. After 10 min, the reac-tion was stopped by adding 4 � SDS sample buffer. Sampleswere then treated as detailed for the autophosphorylationassay. All manipulations were done rapidly and at 48C.
Dephosphorylation assayFor dephosphorylation experiments 10 mM of protein in thebuffer used for autophosphorylation (see above) was incu-bated with 200 mM ATP containing 4 mCi [g-32P-ATP] inthe absence or presence of 100 mM toluene for 50 min. A200-fold molar excess of unlabelled ATP was added, and
H.Silva-Jimenez et al.
162
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from
samples were removed at time intervals to be treated in thesame manner as described for the transphosphorylationassay.
b-Galactosidase measurementsPseudomonas putida DOT-T1E DtodST bearing pMIR77(containing a PtodX::lacZ fusion) and pMIR66 (containingtodST) or pMIR66-Min-TodS (containing min-todST) wereanalysed in parallel. Cells were grown overnight in LBmedium supplemented with 25 mg/ml kanamycin, 10 mg/mltetracycline and 100 mg/ml gentamycin. Cultures werediluted 100 times with the same medium and the differentcompounds were provided in the gas phase or for the compe-tition assays in the presence of various effector molecules(Supplementary Fig. S4) were added to the medium at aconcentration of 1 mM. When the cultures reached an OD600
of 0.8+ 0.05, b-galactosidase activity was determined inpermeabilised cells as described in Ramos-Gonzalez et al.(2002).
Results
The two PAS domains and AK modules of TodS belongto different subfamiliesPAS domains are among the most frequent sensor domainsin bacteria (Ulrich et al., 2005). According to Pfam, thesuperfamily of PAS domains can be divided into sevenfamilies, which are each identified by a hidden Markovmodel. TodS contains two domains that belong to the PASsuperfamily. However, each domain belongs to a differentfamily. The PAS1 domain is recognised by model PF08448(which defines the PAS4 subfamily), whereas the PAS2domain belongs to the family defined by PF00989 (whichdefines the PAS family). The fact that both domains belongto different families is also supported by a sequence align-ment of both domains (Supplementary Fig. S1), whichreveals an amino acid identity of 14%. This value is close tothe random sequence identity obtained by the alignment ofunrelated proteins (Doolittle, 1981).
The histidine protein kinases also form a superfamily thatcan be divided into 11 different families (Grebe and Stock,1999). As reported in Busch et al. (2009) the N-terminal AKmodule of TodS belongs to the subfamily HPK1a whereasthe C-terminal module forms part of the HPK4 family. Inanalogy to the PAS domains the sequence alignment of bothAK modules also reveals a low degree of sequence identityof 22% (Supplementary Fig. S1). These results are consistentwith the notion that TodS is the result of a gene fusion eventof two TCSs rather than gene duplication.
Design and purification of Min-TodSWe have provided evidence that effectors bind solely to theN-terminal PAS domain of TodS, that this binding stimulatesexclusively the N-terminal AK module and that TodT trans-phosphorylation occurs only from the C-terminal AK module(Busch et al., 2009). These observations form the basis forthe design of a minimal version of TodS (Min-TodS) that iscomposed of the PAS1 and the AK2 module (Fig. 1).Min-TodS was constructed by fusing amino acids 1–182 ofthe full-length TodS (containing the PAS1 domain and a 24amino acid linker) with amino acids 753–978 (see Materials
and methods). A central fragment of 570 amino acids wasthus removed from TodS. Min-TodS consists of 408 aminoacids and has a molecular weight of 44.8 kDa. The codingsequence of Min-TodS was cloned into the expression vectorpET28 and the protein was expressed in E.coli. Min-TodSwas purified from the soluble fraction of the E.coli lysate byaffinity chromatography. Purified Min-TodS was subjected toa number of analyses, which were conducted using theexperimental conditions that were used to study the TodS/TodT system (Lacal et al., 2006; Busch et al., 2007, 2009).
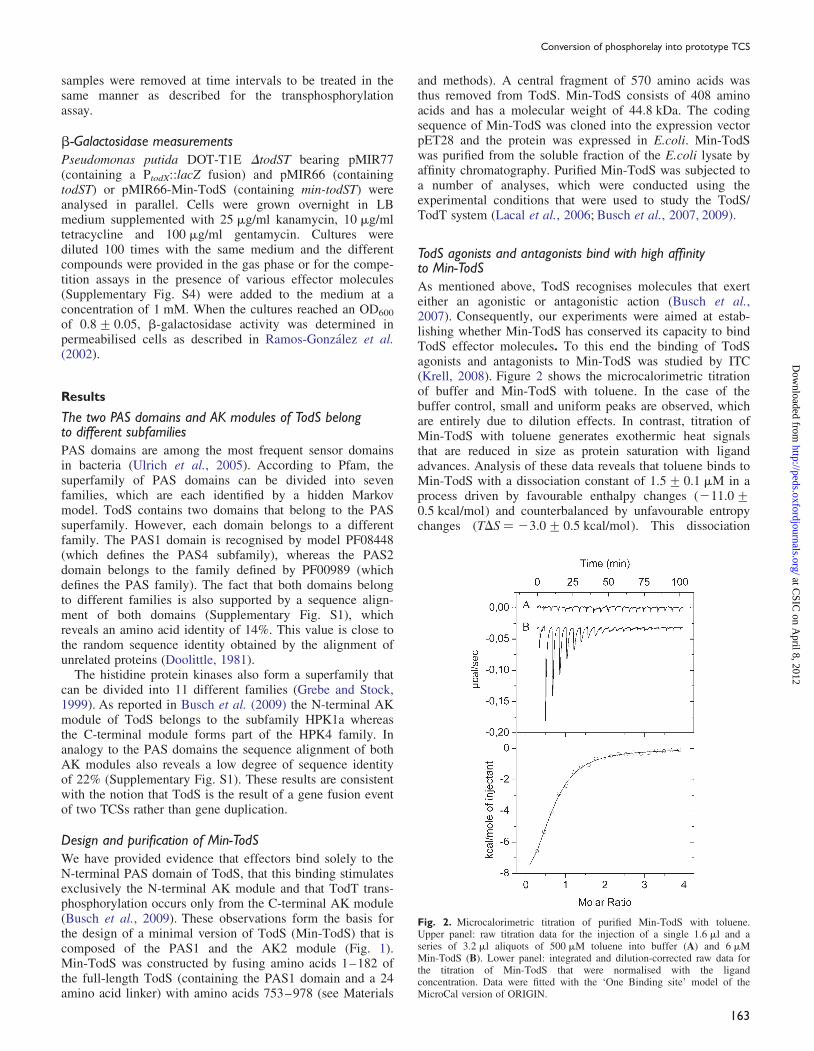
TodS agonists and antagonists bind with high affinityto Min-TodSAs mentioned above, TodS recognises molecules that exerteither an agonistic or antagonistic action (Busch et al.,2007). Consequently, our experiments were aimed at estab-lishing whether Min-TodS has conserved its capacity to bindTodS effector molecules. To this end the binding of TodSagonists and antagonists to Min-TodS was studied by ITC(Krell, 2008). Figure 2 shows the microcalorimetric titrationof buffer and Min-TodS with toluene. In the case of thebuffer control, small and uniform peaks are observed, whichare entirely due to dilution effects. In contrast, titration ofMin-TodS with toluene generates exothermic heat signalsthat are reduced in size as protein saturation with ligandadvances. Analysis of these data reveals that toluene binds toMin-TodS with a dissociation constant of 1.5+ 0.1 mM in aprocess driven by favourable enthalpy changes (211.0+0.5 kcal/mol) and counterbalanced by unfavourable entropychanges (TDS ¼ 23.0+ 0.5 kcal/mol). This dissociation
Fig. 2. Microcalorimetric titration of purified Min-TodS with toluene.Upper panel: raw titration data for the injection of a single 1.6 ml and aseries of 3.2 ml aliquots of 500 mM toluene into buffer (A) and 6 mMMin-TodS (B). Lower panel: integrated and dilution-corrected raw data forthe titration of Min-TodS that were normalised with the ligandconcentration. Data were fitted with the ‘One Binding site’ model of theMicroCal version of ORIGIN.
Conversion of phosphorelay into prototype TCS
163
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

constant is thus comparable with that measured for TodS,which was of 0.68+ 0.02 mM (Lacal et al., 2006).
Subsequently, Min-TodS was titrated with four TodS ago-nists (benzene, ethylbenzene, p-xylene and m-chlorotoluene)and two antagonists (o-xylene, o-chlorotoluene). All thesecompounds were found to bind to Min-TodS and the derivedthermodynamic parameters are summarised in Table III.Affinities range from 270 nM in the case of m-chlorotolueneto 2.8 mM for o-chlorotoluene. With the exception ofm-chlorotoluene, which binds 30 times tighter to Min-TodSas compared with TodS, the effector affinities for Min-TodSare comparable with those obtained for TodS (last column inTable III). In addition, binding of 1,3,5-trimethylbenzene(1,3,5-TMB) was studied. This compound, although structur-ally similar to the other effector molecules tested, was foundto interact with TodS with a high dissociation constant of133+ 22 mM, which corresponds to the highest dissociationconstant of any TodS ligand analysed (Busch et al., 2007).Similarly, 1,3,5-TMB failed to bind to Min-TodS (notshown). These data show that Min-TodS has largely pre-served its capacity to bind TodS agonists and antagonistswith high affinity.
TodS agonists and antagonists reduce Min-TodS AK activityWe then investigated the influence of TodS agonists andantagonists on the AK activity. Toluene is one of the threeTOD pathway substrates and the most potent inducer of todoperon expression in vivo (Lacal et al., 2006). In the case ofTodS the presence of saturating concentrations of tolueneincreased the phosphorylation state of the protein by a factorof �5 (Fig. 3, Lacal et al., 2006). However, this stimulationof AK activity observed might account only partially for the�50-fold increase in gene expression (Lacal et al., 2006).
Min-TodS was subjected to autophosphorylation assays.The basal phosphorylation activity (in the absence oftoluene) of Min-TodS was largely superior to the basal phos-phorylation of TodS (Fig. 3). However, the addition oftoluene to Min-TodS resulted in a reduction of the phosphor-ylation state of the protein by �30–40% (Fig. 3). Thisexperiment was then repeated in the presence of two otherTodS agonists (benzene and m-chlorotoluene) as well as withtwo antagonists (o-xylene and o-chlorotoluene). As shown inFig. 3, all of these compounds caused a reduction in thephosphorylation state of TodS. However, this reduction wasless pronounced in the presence of agonists than antagonists
(Fig. 3). This suggests that agonists and antagonists have adifferential effect on Min-TodS autophosphorylation.Compared with the effect on TodS the regulatory effect ofagonists and antagonists on Min-TodS appears to be off-setinto the negative range. To explain the unexpectedly highphosphorylation state of Min-TodS in the absence of effec-tors, we hypothesised that tightly binding signal moleculesmight co-purify with Min-TodS, causing this elevated autop-hosphorylation in the absence of toluene. To this endMin-TodS was purified under denaturing conditions and thenrefolded (for further detail see Materials and methods).Autophosphorylation assays with refolded protein and withprotein purified under non-denaturing conditions were con-ducted in parallel. These assays reveal an almost identicallevel of autophosphorylation in the absence of toluene and asimilar decrease in autophosphorylation in the presence ofthe above effectors (not shown). These data show that theelevated basal autophosphorylation level and the reduction inprotein phosphorylation in the presence of effectors is aninternal property of Min-TodS.
Min-TodS forms a TCS with TodT and mediates an increasein the amount of phosphorylated TodT in response totolueneWe have shown previously that the phosphorylated form ofTodT stimulates expression from PtodX (Lacal et al., 2006).Central to the study of Min-TodS was to establish whether itforms a TCS with TodT and whether it maintains thecapacity to modulate the amount of phosphorylated TodT inresponse to agonists. To this end Min-TodS and TodT werepurified and subjected to transphosphorylation studies in thepresence and absence of toluene (Fig. 4). These assaysinvolved the exposure of proteins to [g32P] ATP for 10 min,followed by an electrophoretic separation by SDS–PAGEand autoradiographic detection of protein-associated 32P,indicative of the amount of protein phosphorylation. Asshown in lane 1 of Fig. 4, exposure of Min-TodS to ATPresulted in a strong band indicative of efficient autophosphor-ylation of the protein in the absence of toluene. When thisexperiment is repeated in the presence of TodT, the band cor-responding to Min-TodS is significantly reduced, which isconsistent with a transphosphorylation to TodT. However, nophosphorylated TodT could be visualised, which is partlydue to the reduced phosphorylation half-life of TodT. Theaddition of toluene caused a slight reduction in the
Table III. Thermodynamic parameters of ligand binding derived from microcalorimetric titrations of purified Min-TodS with different effector molecules
Effector Effect on TodSa KD (mM) KA (M21) DH (kcal/mol) TDS (kcal/mol) KDTodS/KDMinTodSb
Toluene Agonist 1.5+0.1 (6.6+0.4) 105 211.0+0.5 23.0+0.5 0.46Ethylbenzene Agonist 2.3+0.3 (4.4+0.7) 105 24.6+1 3.0+1 1.34Benzene Agonist 1.0+0.2 (1.0+0.2) 106 25.7+0.8 2.5+0.8 0.76p-Xylene Agonist 1.2+0.1 (8.0+1) 105 210.1+1.0 22.04+1.0 0.63m-Chlorotoluene Agonist 0.27+0.02 (3.6+0.3) 106 21.7+0.1 7.2+0.1 30o-Xylene Antagonist 0.5+0.1 (2.0+0.3) 106 25.2+0.4 3.4+0.4 1.2o-Chlorotoluene Antagonist 2.8+0.5 (3.6+0.7) 105 28.0+2.3 20.38+2.2 0.261,3,5-TMBc Very weak binding No binding
The last column shows the ratio of dissociation constants determined for TodS and the corresponding values determined in this work.aThe effect of these compounds on TodS activity has been described in Busch et al. (2007).bDissociation constants for TodS are taken from Busch et al. (2007).c1,3,5-Trimethylbenzene.
H.Silva-Jimenez et al.
164
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

phosphorylation state of Min-TodS (lane 3 in Fig. 4).However, when all three components, Min-TodS, TodT andtoluene, are present in the reaction mixture, a strong band atthe level of Min-TodS is observed indicative of sustainedautophosphorylation. Most importantly, a band representingphosphorylated TodT is clearly visible (Fig. 4, lane 4). Theintensity of this band was comparable with the correspondingband in the TodS–TodT transphosphorylation assays asreported earlier (Lacal et al., 2006). From these experimentsit can be concluded that Min-TodS has preserved its capacityto recognise and phosphorylate TodT and to mediate anincrease of TodT-P in response to toluene.
Phosphorylation half-life of Min-TodS is significantlyincreased compared to TodSA characteristic feature of a SK is its phosphorylation half-life. We have reported previously (Lacal et al., 2006) that thephosphorylation half-life of TodS is of 71 min and the corre-sponding gel is shown as a reference in Fig. 5. The identicalprocedure was used to assess the phosphorylation half-life ofMin-TodS. Purified Min-TodS is phosphorylated with200 mM ATP containing 4 mCi [g32P]ATP for 50 min,
Fig. 3. Autophosphorylation of TodS and Min-TodS in the absence andpresence of agonists or antagonists. Ten micromolars of TodS (A) orMin-TodS (B) were incubated with 200 mM ATP containing 4 mCi[g32P]ATP in the absence or presence of 100 mM the effector indicated.Samples were taken at the time intervals indicated and then submitted toSDS–PAGE analysis. (C) Densitometric analysis of the gels shown in panelB. Continuous line: buffer control; dotted line: in the presence of TodSagonists: filled cross indicates toluene; filled circle indicates benzene; filleddiamond indicates m-chlorotoluene; dashed line indicates the presence ofTodS antagonists: filled square indicates o-chlorotoluene; and plus indicateso-xylene.
Fig. 4. Transphosphorylation of Min-TodS to TodT in the absence andpresence of toluene. Transphosphorylation between Min-TodS and TodT inthe absence and presence of toluene. Min-TodS was autophosphorylated inthe presence and absence of toluene for 50 min, prior to the addition ofTodT or buffer. Reactions were stopped after another 10 min.
Fig. 5. Dephosphorylation of Min-TodS, TodS and TodS mutant D500A.(A) Dephosphorylation of TodS, Min-TodS and TodS D500A. Aspartate 500is the phosphoryl group accepting residue in the RRR domain of TodS (seeFig. 1). Proteins were autophosphorylated in the presence of 200 mM ATPcontaining 4 mCi [g32P]ATP for 50 min prior to the addition of a 200-foldexcess of unlabelled ATP. Samples were taken at time intervals and analysedon SDS–PAGE (A). (B) Densitometric analysis of the upper panel.Triangles indicate TodS; diamonds indicate Min-TodS; and circles indicateTodS D500A. The dephosphorylation half-life of TodS has been reported inLacal et al. (2006).
Conversion of phosphorelay into prototype TCS
165
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

subsequently a 200-fold excess of unlabelled ATP is addedto dilute out radioactive ATP and samples are taken at timeintervals. Phosphorylation at Min-TodS was remarkablystable and only an �15% decrease in phosphorylation wasobserved after 6 h of incubation (Fig. 5). This indicates thatthe phosphorylation half-life of Min-TodS is much higherthan that of TodS.
TodS operates by a phosphorelay involving His190–Asp500 and His760 of TodS and Asp57 of TodT (Buschet al., 2009). We hypothesised that the absence of the RRreceiver (RRR) domain (harbouring Asp500) in Min-TodSmight be the reason for this increased half-life. To this end,dephosphorylation experiments with the TodS mutantD500A were conducted (Fig. 5). The phosphorylation half-life of this mutant was comparable with that of Min-TodSsince phosphorylation decreased by �30% after 6 h. Thisindicates that the hydrolysis of the acylphosphate at Asp500is the primary cause for the comparatively low phosphoryl-ation half-life of TodS, which explains the stability ofMin-TodS-P.
The individual PAS domains and AK modules cannot beobtained as structured, recombinant proteinsSubsequent studies were aimed at producing the two PASdomains and the AK1 modules as recombinant proteins andto study their mutual interaction by calorimetry as well as tostudy effector binding to the individual PAS1 domain. Tothis end the coding sequence for the PAS1 and PAS2domains as well as for the AK1 module were cloned into theexpression vector pET28 and the resulting plasmid wereintroduced into several E.coli strains for overexpression (seeMaterials and methods). Despite the use of various E.colistrains and a wide range of expression conditions no proteinexpression was observed in the case of PAS1 and AK1. Inthe case of PAS2, a significant amount of pure protein wasobtained. However, a 10–1298C upscan using differentialscanning calorimetry (Krell, 2008) did not show any unfold-ing transitions (data not shown), indicating that the proteinwas unstructured. This conclusion is supported by theabsence of secondary structure elements as revealed by farUV circular dichroism spectroscopy (data not shown). Thesedata are consistent with the idea that the individual domainsare thermodynamically unstable and do not permit furtheranalyses.
Min-TodS modulates expression from PtodX in responseto effector moleculesTo complement data obtained by in vitro experimentation,we have analysed the capacity of the Min-TodS/TodT TCSto modulate gene expression from PtodX. For these experi-ments plasmid pMIR66-Min-TodS was constructed. Theparent plasmid pMIR66 corresponds to a pBBR1MCS-1derivative that contains the todST genes (Ramos-Gonzalezet al., 2002). In plasmid pMIR66-Min-TodS the todS genewas replaced by the coding sequence of Min-TodS. Theplasmid was introduced into P.putida DOT-T1E DtodST(Ramos-Gonzalez et al., 2002) harbouring plasmid pMIR77containing the PtodX::lacZ fusion. The resulting strain wasused for b-galactosidase measurements in the presence andabsence of effector molecules. In the absence of effectormolecules, a b-galactosidase activity of �900 Miller Units(MU) was measured. Subsequently, the responses to four
TodS agonists (toluene, benzene, m-chlorotoluene andp-xylene), two TodS antagonists (o-xylene ando-chlorotoluene) and a structurally related compound thatdoes not bind to Min-TodS (1,3,5-TMB) were determined(Fig. 6A). The two TOD pathway substrates toluene andbenzene stimulated PtodX expression by factors of 4 and 2.5,respectively. A slight, but reproducible, increase in promoteractivity was also observed for m-, o-chlorotoluene andp-xylene. It should be noted that toluene was found to be themost potent effector in the TodS-mediated increase in PtodX
expression (Lacal et al., 2006). Finally, no significantincrease in gene expression was observed in the presence of1,3,5-TMB. This is not surprising since we have shown
Fig. 6. Expression from PtodX mediated by Min-TodS/TodT (A) and TodS/TodT (B) in response to different effector molecules. b-Galactosidasemeasurements of P.putida DOT-T1E DtodST harbouring plasmids pMIR77and pMIR66-Min-TodS (A) or pMIR77 and pMIR66 (B). Plasmid pMIR66harbours full-length TodS whereas pMIR66-Min-TodS harbours Min-TodS.Plasmid pMIR77 contains a fusion of promoter PtodX with the lacZ gene.Data are the means and corresponding standard errors derived from at leastthree independent assays, each done in triplicate.
H.Silva-Jimenez et al.
166
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

above that it does not bind to Min-TodS. This is initialevidence that the modulation of gene expression is modu-lated by Min-TodS/TodT and not due to any potential pleio-tropic effects on gene expression that might have beencaused by the addition of these toxic aromatic compounds tothe bacterial culture. These measurements were taken at anOD of 0.8. We have then analysed promoter expression atODs of 0.5 and 1.2, which showed similar results(Supplementary Fig. S2).
To entirely exclude the possibility that upregulationobserved is mediated by potential alternative mechanisms,b-galactosidase measurements were made using P.putidaDOT-T1E DtodST harbouring only pMIR77 (containingPtodX::lacZ) but lacking pMIR66-Min-TodS (encoding theMin-TodS/TodT). The addition of toluene did not cause anysignificant modulation of gene expression providing evidencethat gene upregulation reported above is mediated byMin-TodS/TodT (data not shown). In a second controlexperiment plasmid pJLC1 containing the todT gene wastransferred into P.putida DOT-T1E DtodST harbouring onlypMIR77. Promoter expression studies were conducted in theabsence of effector and in presence of toluene,m-chlorotoluene and o-chlorotoluene. In all cases, obtainedactivities were indistinguishable and close to zero(Supplementary Fig. S3). This indicates that promoteractivity observed is mediated by Min-TodS and is not due toa secondary effect and also suggests that there is noadditional path to phosphorylate TodT.
PtodX expression studies by the TodS/TodT system wereperformed previously (Lacal et al., 2006), but using adifferent experimental strategy as compared with this study.In the initial study, P.putida DOT-T1E bearing pMIR77 wasused and TodS and TodT were of genomic origin. In thepresent study the P.putida DOT-T1E mutant strain DtodSTwas used and Min-TodS/TodT were encoded by the mediumcopy number plasmid pMIR66-Min-TodS. To compare themagnitude of promoter activity of the TodS/TodT TCS withthe Min-TodS/TodT system, b-galactosidase measurementsof P.putida DOT-T1E DtodST containing pMIR77 andpMIR66 (harbouring todST) were conducted (Fig. 6B). In theabsence of effector molecules, a basal activity of �50 MUwas observed and the addition of toluene resulted in anactivity of �12 300 MU, which corresponds to an increaseby a factor of �250. In agreement with the published data,the three remaining agonists benzene, m-chlorotoluene andp-xylene induced an activity of 3000–8000 MU whereas thevalues for the antagonists o-xylene and o-chlorotoluene werecomparable with the control.
The last set of experiments involved expression experi-ments with multiple effector molecules. In the experimentsso far reported, the effector molecules were provided in thegas phase. The competition experiments have been con-ducted using a protocol similar to that described by Buschet al. (2007) in which effectors were added directly to thegrowth medium in order to precisely control its concentra-tions. As shown in Supplementary Fig. S4 toluene was themost potent inducer followed by o-chlorotoluene andm-chlorotoluene. The difference in the potency of both effec-tors was also reflected when they were added to toluene-containing samples (for experimental details please refer tothe legend of Supplementary Fig. S4). Promoter activities inthe presence of toluene and o-chlorotoluene were comparable
with that in the presence of toluene alone. However, amixture of m-chlorotoluene with toluene caused a reductionin promoter expression as compared with the toluene sample.These data are consistent with the notion that effectors sharethe same binding site at Min-TodS.
Discussion
Bacterial signal transduction is mediated primarily by theconcerted action of several protein families, namely TCSs,one-component systems, methylaccepting chemotaxis pro-teins, serine/threonine kinases, adenylate and guanylatecyclases and phosphodiesterases (Galperin, 2005; Ulrich andZhulin, 2005). These protein families are characterised by amodular arrangement containing input (sensing) and outputdomains. Interestingly, protein families share the same typeof sensor modules. For example, the PAS (Moglich et al.,2009b) and TarH domains (Ulrich and Zhulin, 2005) werefound to be present in many different protein families. Thisindicates that during evolution sensor domains have beenrecombined with other output modules.
This evolutionary evidence is also supported by experi-mental data and there are several reports on the recombin-ation of sensor domains with output modules of differentproteins. The chimeric Taz protein is a fusion of theTarH-type sensor domain of the Tar chemoreceptor with theAK module of EnvZ. The Tar chemoreceptor mediates achemotactic response towards aspartate whereas the EnvZ/OmpR system controls the expression of genes encodingporins. It was shown that the Taz/OmpR system controlsporin expression in response to aspartate (Utsumi et al.,1989). The Moffat group has constructed various chimericproteins based on the photosensor YtvA and the FixL SK.YtvA has a single PAS domain that senses blue light,whereas FixL has two PAS domains, of which one (PAS A)is of unknown function whereas the other (PAS B) sensesoxygen. A FixL derivative in which both PAS domains(Moglich et al., 2009a) or the PAS A domain (Moglichet al., 2010) are replaced by the YtvA sensor domain, wasfound to be active and to mediate responses to blue light.
To our knowledge, our study corresponds to the firstconstruction of a prototypal SK using elements of a phos-phorelay kinase. Our data confirm the apparent ease bywhich sensor domains can be recombined with differentoutput modules. The comparison of TodS/TodT with MinTodS/TodT has revealed several issues. Firstly, TodS andMin-TodS recognise effector molecules with similar affinity(Table III). In addition, for both proteins effector moleculescan be classified into agonists and antagonists (Fig. 6). Thesedata indicate that the molecular determinants for effectorrecognition and their either agonistic or antagonistic actionare located in the PAS1 domain. This conclusion isconsistent with our proposition (Busch et al., 2007) that theagonistic or antagonistic action of effector molecules may becaused by two different modes of ligand binding to thePAS1 domain.
Secondly, significant differences in the phosphorylationhalf-lifes of TodS and Min-TodS were observed. For TodS aphosphorylation half-life of 71 min was determined (Lacalet al., 2006), whereas the phosphorylated form of Min-TodSwas stable over at least 6 h, indicating a significant increasein its phosphorylation half-life (Fig. 5). Experiments with the
Conversion of phosphorelay into prototype TCS
167
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

TodS mutant D500A demonstrated that the hydrolysis of theacylphosphate at the aspartate residue of the TodS receiverdomain is the primary cause for the comparatively low half-life of TodS-P. It can thus be concluded that the differencesin half-lives enable the TodS/TodT system to return faster tothe non-activated state than its prototypal derivative. Thisrapidity of reaction could be an advantage in the context ofenvironmental changes that involve a sudden absence ofaromatic hydrocarbons.
Thirdly, there were differences at the basal activities ofpromoter expression. Promoter PtodX was almost silent(50 MU) in the native TodS system, whereas a basal activityof �900 MU was observed for the Min-TodS system. Wehave shown previously (Busch et al., 2009) that TodSmutants in which the phosphorylaccepting amino acidsH190, D500 and H760 are replaced by alanine, possess a lowbasal AK activity that is comparable with the wild-typeprotein. As a result promoter activities are very low in theabsence of effectors. This demonstrates that the low basalactivity occurs irrespective of there being a functionalphosphorelay. However, the removal of the centralAK1-RRR-PAS2 fragment causes an increase in the basalpromoter activity. We hypothesise that intra-TodS contactsbetween the AK1-RRR-PAS2 fragment with the remainingprotein might cause this reduction in basal activity. This verylow basal activity of the TodS system is advantageous com-pared to the Min-TodS system since the TOD pathway actsexclusively on three substrates and the tod operon expressionin the absence of these compounds is a futile event.
Fourthly, the magnitude of gene expression mediated bytoluene and benzene, the two primary pathway substrates,was significantly higher for the phosphorelay system as com-pared with its prototypal derivative. For example, the TodS/TodT-mediated gene expression was of �12 000 and8000 MU for toluene and benzene, respectively, whereas thecorresponding values for the Min-TodS/TodT system were of3500 and 2000 MU. Since the phosphorylated form of TodTcan be associated with the increase in gene expression, it canbe suggested that the amount of TodT-P generated by theaction of TodS is superior to that generated by Min-TodS.
Fifthly, despite the very-low-sequence identity betweenthe PAS1 and PAS2 domains and the AK1 and AK2modules of TodS, the successful construction of Min-TodSunderlines that PAS sensor domains and AK modules can berecombined with apparent ease. To understand this apparentplasticity in domain interaction, the 3D structure of theThkA SK (Yamada et al., 2009) can provide potentialreasons. The cytosolic ThkA is composed of a PAS domainfollowed in sequence by an AK module, which correspondsto the Min-TodS domain organisation. This structure revealsdetails of the PAS domain–AK module interaction(Supplementary Fig. S5). The surface of domain interactionis surprisingly small and restricted to a simple interactionbetween strand b3 of the PAS b-sheet and strand b6 of thecatalytic domain of the AK module, which creates an inter-domain b-sheet. Interestingly, the interdomain b-sheet ismediated exclusively by main chain interactions. It is likelythat the domain communication between the two domains inMin-TodS occurs in a similar manner as in ThkA. The factthat the area of interaction between both domains is minimaland that this interaction is mediated by main chain atomsmight indicate that there is only a modest evolutionary
pressure on both domains to conserve amino acids involvedin interaction.
Sixthly, toluene causes in both systems an increase in theconcentration of TodT-P (Lacal et al., 2006, Fig. 4).However, in the case of TodS toluene was found to stimulateits AK activity (Lacal et al., 2006), whereas the exposure ofMin-TodS to toluene caused a slight reduction in its AKactivity (Fig. 3). To explain this apparent discrepancy wewould like to refer to a survey of TCS, which has revealedthat the large part of SKs is bifunctional proteins that possessboth kinase and phosphatase activities towards its RR(Huynh and Stewart, 2011). It was suggested that the kinaseactive and the phosphatase active states of a SK correspondto two mutually exclusive conformations of the SK trans-mitter module, which implies a reciprocal character of bothactivities associated (Stewart, 2010). Both SK conformationsare thought to be present in equilibrium and the stimulusresulting from signal molecule recognition is thought to shiftthis equilibrium between both conformations to one or theother side (Huynh and Stewart, 2011). In analogy to thisgeneral model for bifunctional SKs, we propose that tolueneincreases TodS AK activity and at the same time reduces itsphosphatase activity towards TodT. In Min-TodS the capacityto upregulate its AK activity in response to toluene has beeneliminated and the regulatory effect of the Min-TodS/TodTsystem is therefore likely due to the regulation of its phos-phatase activity. However, all attempts to characterise TodSor Min-TodS phosphatase activities were unsuccessful.
In summary, the reduced basal activity, the larger magni-tude of gene induction in response to toluene as well as thereduced half-life of phosphorylated TodS as compared withMin-TodS appear to be features which make the phosphore-lay system better suited to regulate expression of a degrad-ation pathway of toxic compounds.
Supplementary data
Supplementary data are available at PEDS online.
Acknowledgements
We thank Cristina Garcıa-Fontana for technical assistance. The work wasfinanced by grants from the BBVA Foundation, the Andalusian regional gov-ernment Junta de Andalucıa [grant P09-RNM-4509] and the SpanishMinistry for Science and Innovation [grant Bio2010-16937].
ReferencesAlbanesi,D., Martın,M., Trajtenberg,F., Mansilla,M.C., Haouz,A.,
Alzari,P.M., de Mendoza,D. and Buschiazzo,A. (2009) Proc. Natl. Acad.Sci. USA, 106, 16185–16190.
Beier,D. and Gross,R. (2008) Adv. Exp. Med. Biol., 631, 149–160.Burbulys,D., Trach,K.A. and Hoch,J.A. (1991) Cell, 64, 545–552.Busch,A., Guazzaroni,M.E., Lacal,J., Ramos,J.L. and Krell,T. (2009) J. Biol.
Chem., 284, 10353–10360.Busch,A., Lacal,J., Martos,A., Ramos,J.L. and Krell,T. (2007) Proc. Natl
Acad. Sci. USA, 104, 13774–13779.Cases,I. and de Lorenzo,V. (2005) Int. Microbiol., 8, 213–222.Chamnongpol,S., Cromie,M. and Groisman,E.A. (2003) J. Mol. Biol., 325,
795–807.Cock,P.J. and Whitworth,D.E. (2007) Mol. Biol. Evol., 24, 2355–2357.Coschigano,P.W. and Young,L.Y. (1997) Appl. Environ. Microbiol., 63,
652–660.Doolittle,R.F. (1981) Science, 214, 149–159.
H.Silva-Jimenez et al.
168
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from

Fredericks,C.E., Shibata,S., Aizawa,S., Reimann,S.A. and Wolfe,A.J. (2006)Mol. Microbiol., 61, 734–747.
Galperin,M.Y. (2005) BMC Microbiol., 5, 35–53.Galperin,M.Y. (2010) Curr. Opin. Microbiol., 13, 150–159.Grebe,T.W. and Stock,J.B. (1999) Adv. Microb. Physiol., 41, 139–227.Huynh,T.N. and Stewart,V. (2011) Mol. Microbiol., 82, 275–286.Krell,T. (2008) Microb. Biotechnol., 1, 126–136.Krell,T., Lacal,J., Busch,A., Guazzaroni,M.E., Silva-Jimenez,H. and
Ramos,J.L. (2010) Annu. Rev. Microbiol., 64, 539–559.Lacal,J., Busch,A., Guazzaroni,M.E., Krell,T. and Ramos,J.L. (2006) Proc.
Natl Acad. Sci. USA, 103, 8191–8196.Lacal,J., Guazzaroni,M.E., Busch,A., Krell,T. and Ramos,J.L. (2008a)
J. Mol. Biol., 376, 325–337.Lacal,J., Guazzaroni,M.E., Gutierrez del Arroyo,P., Busch,A., Velez,M.,
Krell,T. and Ramos,J.L. (2008b) J. Mol. Biol., 384, 1037–1047.Lau,P.C., Wang,Y., Patel,A., Labbe,D., Bergeron,H., Brousseau,R.,
Konishi,Y. and Rawlings,M. (1997) Proc. Natl Acad. Sci. USA, 94,1453–1458.
Leoni,L., Ascenzi,P., Bocedi,A., Rampioni,G., Castellini,L. and Zennaro,E.(2003) Biochem. Biophys. Res. Commun., 303, 926–931.
Lois,A.F., Weinstein,M., Ditta,G.S. and Helinski,D.R. (1993) J. Biol. Chem.,268, 4370–4375.
Majdalani,N. and Gottesman,S. (2005) Annu. Rev. Microbiol., 59, 379–405.Malpica,R., Sandoval,G.R., Rodrıguez,C., Franco,B. and Georgellis,D.
(2006) Antioxid. Redox Signal, 8, 781–795.Mascher,T., Helmann,J.D. and Unden,G. (2006) Microbiol. Mol. Biol. Rev.,
70, 910–938.Moglich,A., Ayers,R.A. and Moffat,K. (2009a) J. Mol. Biol., 385,
1433–1444.Moglich,A., Ayers,R.A. and Moffat,K. (2009b) Structure, 17, 1282–1294.Moglich,A., Ayers,R.A. and Moffat,K. (2010) J. Mol. Biol., 400, 477–486.Mosqueda,G., Ramos-Gonzalez,M.I. and Ramos,J.L. (1999) Gene, 232,
69–76.Perraud,A.L., Weiss,V. and Gross,R. (1999) Trends Microbiol., 7, 115–120.Prost,L.R. and Miller,S.I. (2008) Cell Microbiol., 10, 576–582.Ramos,J.L., Duque,E., Huertas,M.J. and Haıdour,A. (1995) J. Bacteriol.,
177, 3911–3916.Ramos-Gonzalez,M.I., Olson,M., Gatenby,A.A., Mosqueda,G.,
Manzanera,M., Campos,M.J., Vıchez,S. and Ramos,J.L. (2002)J. Bacteriol., 184, 7062–7067.
Sheeler,N.L., MacMillan,S.V. and Nodwell,J.R. (2005) J. Bacteriol., 187,687–696.
Stewart,R.C. (2010) Curr. Opin. Microbiol., 13, 133–141.Studier,F.W. and Moffat,B.A. (1986) J. Mol. Biol., 189, 113–130.Ulrich,L.E. and Zhulin,I.B. (2005) Bioinformatics, 21(Suppl 3), iii45–48.Ulrich,L.E., Koonin,E.V. and Zhulin,I.B. (2005) Trends Microbiol., 13,
52–56.Utsumi,R., Brissette,R.E., Rampersaud,A., Forst,S.A., Oosawa,K. and
Inouye,M. (1989) Science, 245, 1246–1249.Velasco,A., Alonso,S., Garcıa,J.L., Perera,J. and Dıaz,E. (1998)
J. Bacteriol., 180, 1063–1071.Whitworth,D.E. and Cock,P.J. (2009) Amino Acids, 37, 459–466.Williams,R.H. and Whitworth,D.E. (2010) BMC Genomics, 11, 720.Yamada,S., Sugimoto,H., Kobayashi,M., Ohno,A., Nakamura,H. and
Shiro,Y. (2009) Structure, 17, 1333–1344.Zhang,W. and Shi,L. (2005) Microbiology, 151, 2159–2173.Zylstra,G.J. and Gibson,D.T. (1989) J. Biol. Chem., 264, 14940–14946.
Conversion of phosphorelay into prototype TCS
169
at CSIC
on April 8, 2012
http://peds.oxfordjournals.org/D
ownloaded from
Related Documents











