Original article Conservation of remnant populations of Colchicum autumnale – The relative importance of local habitat quality and habitat fragmentation Dries Adriaens a, *, Hans Jacquemyn b , Olivier Honnay b , Martin Hermy a a University of Leuven, Division of Forest, Nature and Landscape, Celestijnenlaan 200 E, B-3001 Leuven, Belgium b University of Leuven, Biology Department, Plant Ecology, Kasteelpark Arenberg 31, B-3001 Leuven, Belgium article info Article history: Received 13 February 2008 Accepted 20 August 2008 Published online 21 October 2008 Keywords: Population size Plant reproductive capacity Habitat isolation Clonal growth Habitat restoration Microsite availability abstract Semi-natural habitat is extremely vulnerable to habitat fragmentation and degradation since its socio-economic value has decreased substantially during the last century in most parts of Europe. We evaluated the relative effects of habitat fragmentation and local environmental conditions on population structure and reproductive performance of the long-lived corm geophyte Colchicum autumnale in 17 highly fragmented populations. Habitat isolation did not affect patch occupancy, population structure or plant performance. In contrast, population size and local environment strongly affected population structure and reproductive performance. Densities of all life stages increased with increasing population size. Large populations also showed a higher reproductive performance and a larger proportion of new recruits. Relationships with local growth conditions pointed towards the importance of an open grassland sward for flower and fruit set and the presence of microsites for successful sexual recruitment. These results suggest that the distribution of C. autumnale consists of an assemblage of basically unconnected populations that are remnants of formerly larger populations. This is in accordance with the species’ ability to grow clonally, allowing long-term persistence under deteriorating conditions that occurred during a long period of habitat fragmentation. In conclusion, our results indicate that local habitat and population size are more important than habitat fragmentation (i.e. calcareous grassland isolation and surface area) and argue in favour of a management that is primarily focused on local habitat restoration. This is preferentially accomplished by reintroducing grazing practices, complemented by regular setback of spontaneous succession towards forest. ª 2008 Elsevier Masson SAS. All rights reserved. 1. Introduction In most western-European countries, habitat fragmentation is a widespread phenomenon that has profound consequences for biodiversity. Due to socio-economic changes during the 19th and 20th century, large and species-rich grassland areas, once extensively managed by man, were abandoned and converted to more profitable land uses with mainly agricultural, forestry or * Corresponding author. Tel.: þ32 16 32 97 36; fax: þ32 16 32 97 60. E-mail addresses: [email protected] (D. Adriaens), [email protected] (H. Jacquemyn), olivier.honnay@bio. kuleuven.be (O. Honnay), [email protected] (M. Hermy). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/actoec 1146-609X/$ – see front matter ª 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.actao.2008.08.003 acta oecologica 35 (2009) 69–82

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2
ava i lab le at www.sc ienced i rec t . com
journa l homepage : www. e lsev ier . com/ loca te /ac toec
Original article
Conservation of remnant populations of Colchicum autumnale –The relative importance of local habitat quality and habitatfragmentation
Dries Adriaensa,*, Hans Jacquemynb, Olivier Honnayb, Martin Hermya
aUniversity of Leuven, Division of Forest, Nature and Landscape, Celestijnenlaan 200 E, B-3001 Leuven, BelgiumbUniversity of Leuven, Biology Department, Plant Ecology, Kasteelpark Arenberg 31, B-3001 Leuven, Belgium
a r t i c l e i n f o
Article history:
Received 13 February 2008
Accepted 20 August 2008
Published online 21 October 2008
Keywords:
Population size
Plant reproductive capacity
Habitat isolation
Clonal growth
Habitat restoration
Microsite availability
* Corresponding author. Tel.: þ32 16 32 97 36E-mail addresses: [email protected]
kuleuven.be (O. Honnay), martin.hermy@ee1146-609X/$ – see front matter ª 2008 Elsevdoi:10.1016/j.actao.2008.08.003
a b s t r a c t
Semi-natural habitat is extremely vulnerable to habitat fragmentation and degradation
since its socio-economic value has decreased substantially during the last century in most
parts of Europe. We evaluated the relative effects of habitat fragmentation and local
environmental conditions on population structure and reproductive performance of the
long-lived corm geophyte Colchicum autumnale in 17 highly fragmented populations. Habitat
isolation did not affect patch occupancy, population structure or plant performance. In
contrast, population size and local environment strongly affected population structure and
reproductive performance. Densities of all life stages increased with increasing population
size. Large populations also showed a higher reproductive performance and a larger
proportion of new recruits. Relationships with local growth conditions pointed towards the
importance of an open grassland sward for flower and fruit set and the presence of
microsites for successful sexual recruitment. These results suggest that the distribution of
C. autumnale consists of an assemblage of basically unconnected populations that are
remnants of formerly larger populations. This is in accordance with the species’ ability to
grow clonally, allowing long-term persistence under deteriorating conditions that occurred
during a long period of habitat fragmentation. In conclusion, our results indicate that local
habitat and population size are more important than habitat fragmentation (i.e. calcareous
grassland isolation and surface area) and argue in favour of a management that is
primarily focused on local habitat restoration. This is preferentially accomplished by
reintroducing grazing practices, complemented by regular setback of spontaneous
succession towards forest.
ª 2008 Elsevier Masson SAS. All rights reserved.
1. Introduction for biodiversity. Due to socio-economic changes during the 19th
In most western-European countries, habitat fragmentation is
a widespread phenomenon that has profound consequences
; fax: þ32 16 32 97 60.om (D. Adriaens), hans.j
s.kuleuven.be (M. Hermy)ier Masson SAS. All rights
and 20th century, large and species-rich grassland areas, once
extensively managed by man, were abandoned and converted
tomore profitable landuses with mainlyagricultural, forestry or
[email protected] (H. Jacquemyn), olivier.honnay@bio..
reserved.

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 270
urbanization purposes. As a consequence, many semi-natural
grasslands were destroyed and the size of the remaining areas
further diminished, while the distances between remnants
increased. In many cases, their habitat quality also seriously
deteriorated due to the cessation of traditional management
practices and the onset of natural succession, driving many
species to local extinction (e.g. Fischer and Stocklin, 1997; Maes
and Van Dyck, 2001; Wenzel et al., 2006).
The consequences of the different components of habitat
fragmentation (pure area loss, decline in fragment area and
increasing isolation) may be far-reaching. While the result of
pure area loss is obvious, small habitat remnants can only
harbour small populations that suffer from increased envi-
ronmental, demographic and genetic stochasticity (Matthies
et al., 2004; Munzbergova, 2006). In general, the impact of
severe environmental disturbance is most pronounced,
whereas demographic stochasticity only affects very small
populations (Menges and Dolan, 1998; Holsinger, 2000;
Matthies et al., 2004). Inbreeding and loss of genetic variation
can further lead to reduced fitness and evolutionary potential,
eventually causing further population decline (Ellstrand and
Elam, 1993; Newman and Pilson, 1997; Fischer and Matthies,
1998). Moreover, the inflow of genes and seeds by pollinators
or seed dispersers is hampered by increasing isolation, pre-
venting small populations from being ‘rescued’ and extinct
habitat patches from being recolonized (Brown and Kodric-
Brown, 1977; Hanski, 1998). Not only rare species but also
common species have been found to suffer from habitat
fragmentation (Honnay and Jacquemyn, 2007).
However, the impact of habitat fragmentation may take
years to become manifest (Ellstrand and Elam, 1993). Plant
species may respond slowly to changing landscape configu-
ration. They can still be present as remnant populations
despite current conditions that do no longer fulfill their
ecological requirements, contributing to a so-called extinction
debt (Tilman et al., 1994; Helm et al., 2006; Vellend et al., 2006).
This is especially true for long-lived and slow-growing species
that can reproduce vegetatively. Because habitat fragmenta-
tion often implies a decline in habitat quality, it is also
important to discriminate between the effects of habitat
fragmentation per se and local environmental conditions on
long-term population viability, to ensure that management
actions are deployed as efficiently as possible to sustain and
reinforce fragmented populations. These effects can be eval-
uated by measuring changes in vegetative and reproductive
capacity, population structure or long-term demography.
Although results from long-term demographic monitoring are
likely to be most informative, gathering these data is labour-
intensive and requires many years. A single-time census of
population structure, i.e. the relative proportion of different
ontogenetic stages in a species’ life cycle, combined with
measuring various life-cycle characteristics, has been pro-
posed as a more feasible way to quickly assess population
viability (Oostermeijer et al., 1994), in particular when it was
shown that population demographic structures are strongly
correlated with population growth rates obtained from demo-
graphic monitoring (e.g. Oostermeijer et al., 1996; Colling and
Matthies, 2006). As a result, assessing the population structure
has repeatedly been used to evaluate the success of mana-
gement interventions by changing habitat characteristics
(Oostermeijer et al., 1994; Hegland et al., 2001), to evaluate the
performance of species in different habitat types (Endels et al.,
2004; Mroz, 2006) and to measure the impact of habitat isola-
tion and population size on species persistence (Hooftman
and Diemer, 2002; Lienert et al., 2002).
We sampled the population structure and plant perfor-
mance of the long-lived Colchicum autumnale in calcareous
grasslands that have had a long history of habitat fragmen-
tation and cover a range of habitat characteristics. The impact
of population isolation and size, and local environmental
conditions was studied in order to quantify the relative
importance of habitat fragmentation and local environment,
respectively. Since many of its primary habitat has been
destroyed, the remaining populations in calcareous grass-
lands are important for regional species survival.
More specifically, we addressed the following questions:
– How are populations of C. autumnale structured and how
do they perform in a fragmented and heterogeneous
landscape?
– What is the relative importance of habitat fragmentation
vs. local variables in determining Colchicum population
structure?
– If local environment is important, which variables most
influence plant performance and population structure?
– What management can be the most beneficial for the
long-term survival of Colchicum in calcareous grasslands?
2. Materials and methods
2.1. Study area
The study area (80 km2) is located in the south-western part of
an elongated belt of calcareous outcrops in south-western Bel-
gium and consists of hilly areas surrounded by small valleys
(Fig. 1). After a period of socio-economic change in the second
half of the 20th century, traditional management was aban-
doned. Of the 4000 ha of calcareous grasslands that existed in
1775, only 70 ha remain today, as 70 small and isolated frag-
ments in the mainly agricultural and forested landscape matrix.
Most fragments are actively managed as nature reserves by
rotational sheep and goat grazing. We refer to Adriaens et al.
(2006) for a more detailed description of the study area.
2.2. Study species
C. autumnale or meadow saffron is an iteroparous and peren-
nial geophyte with a corm. Preferring moist and fertile valley
meadows, it also occurs in dry calcareous semi-natural
grasslands. Nowadays, the latter have become crucial for
regional survival of Colchicum because its optimal habitat has
become very scarce due to agricultural intensification and
conversion, and because C. autumnale has been regarded as
a pest and toxic plant species in meadows (Butcher, 1954).
Colchicum rapidly disappears after intensive mowing, grazing,
treading, draining or after ploughing and heavy fertilization
(Mroz, 2006). Colchicum also rarely occurs in road verges or
forest margins within the study area. However, these sites are
characterised by very low numbers of reproducing

Fig. 1 – Overview of the calcareous grasslands in the study area. Fragments that are occupied by Colchicum autumnale are
indicated in black. Fragments that were sampled for this study bear their respective label (see Table 1). The other occupied
fragments are marked with an asterisk.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 71
individuals. In terms of ecological connectivity, their impor-
tance is thus considered minimal (Mroz, 2006).
Meadow saffron has a hysteranthous life cycle: flowers
(1–5) appear from August to October and are followed by 2–5
leaves only in early spring (March), together with the fruits
that ripen until June. In mid-July, the leaves wither and only
the dry fruits remain visible aboveground (Butcher, 1954).
Flowers produce nectar and are pollinated by flies and
butterflies. Autogamy is possible if cross-pollination has not
taken place (Butcher, 1954). Seed dispersal is myrmecocho-
rous due to the presence of an elaiosome (Persson, 1993;
personal observation). Vegetative reproduction occurs by
daughter corms that occasionally split off the mother corm
(Jaehn and Roux, 1986; Frankova et al., 2004). The dispersal of
both vegetative and regenerative offspring is thus very limited
and results mainly in locally clumped distributions of indi-
viduals. Plants grown from seed rarely flower before the fifth
year, while vegetative offspring can be reproductive already in
the first year (Poutaraud and Champay, 1995). Seed banks are
transient (Thompson et al., 1997).
2.3. Data collection
Patch occupancy (i.e. the number of occupied calcareous
grassland fragments) of C. autumnale was determined based on
complete species lists of 64 grassland fragments and further
field observations (see Adriaens et al., 2006). In total, 58 plots,
varying in size from 1 to 4 m2, were randomly established in
populations of 17 grassland fragments (Table 1). All individuals
within a grassland fragment were considered to belong to
a single population, given the mainly clumped occurrence of C.
autumnale within the grassland fragments. The number of plots
was determined by the size and spread of the population. To
determine stage structure, i.e. plant density (number of indi-
viduals divided by plot area) and proportion of each different
life stage, we used a classification that discriminated between
seedlings and juveniles (S&J; single leaf), and vegetative (VA)
and reproductive adults (RA; two or more leaves, the latter
flowering in autumn and possibly fruiting in spring) (cf Mroz,
2006). Proportions among life stages were calculated.
In autumn 2004, the exact location of all reproductive
adults within each plot was measured with high-precision
differential GPS (up to the nearest cm) and the number of
flowers per plant was counted. During June 2005, all plots were
revisited to count and locate seedlings, juveniles and vegeta-
tive adults. An individual (S, J, VA or RA) could be easily
discriminated by the flowers or leaf/leaves originating from
a single sheath at ground level. Fruits (i.e. seed pods) of
a maximum of five adults were collected in the neighbour-
hood of the plots to estimate seed production within each plot.
Seed mass was determined as average seed mass after drying
to constant mass at 30 �C. From these measurements,
7 quantitative plant performance variables that represent
reproductive capacity were calculated for each plot: flower,
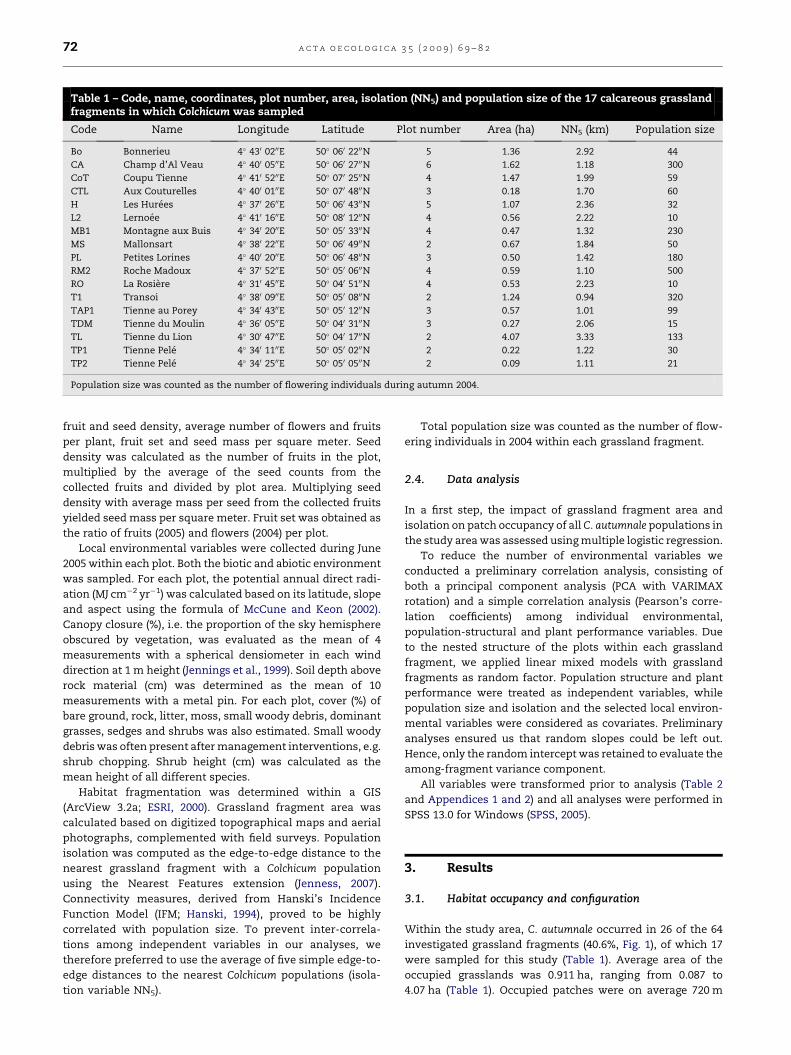
Table 1 – Code, name, coordinates, plot number, area, isolation (NN5) and population size of the 17 calcareous grasslandfragments in which Colchicum was sampled
Code Name Longitude Latitude Plot number Area (ha) NN5 (km) Population size
Bo Bonnerieu 4� 430 0200E 50� 060 2200N 5 1.36 2.92 44
CA Champ d’Al Veau 4� 400 0500E 50� 060 2700N 6 1.62 1.18 300
CoT Coupu Tienne 4� 410 5200E 50� 070 2500N 4 1.47 1.99 59
CTL Aux Couturelles 4� 400 0100E 50� 070 4800N 3 0.18 1.70 60
H Les Hurees 4� 370 2600E 50� 060 4300N 5 1.07 2.36 32
L2 Lernoee 4� 410 1600E 50� 080 1200N 4 0.56 2.22 10
MB1 Montagne aux Buis 4� 340 2000E 50� 050 3300N 4 0.47 1.32 230
MS Mallonsart 4� 380 2200E 50� 060 4900N 2 0.67 1.84 50
PL Petites Lorines 4� 400 2000E 50� 060 4800N 3 0.50 1.42 180
RM2 Roche Madoux 4� 370 5200E 50� 050 0600N 4 0.59 1.10 500
RO La Rosiere 4� 310 4500E 50� 040 5100N 4 0.53 2.23 10
T1 Transoi 4� 380 0900E 50� 050 0800N 2 1.24 0.94 320
TAP1 Tienne au Porey 4� 340 4300E 50� 050 1200N 3 0.57 1.01 99
TDM Tienne du Moulin 4� 360 0500E 50� 040 3100N 3 0.27 2.06 15
TL Tienne du Lion 4� 300 4700E 50� 040 1700N 2 4.07 3.33 133
TP1 Tienne Pele 4� 340 1100E 50� 050 0200N 2 0.22 1.22 30
TP2 Tienne Pele 4� 340 2500E 50� 050 0500N 2 0.09 1.11 21
Population size was counted as the number of flowering individuals during autumn 2004.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 272
fruit and seed density, average number of flowers and fruits
per plant, fruit set and seed mass per square meter. Seed
density was calculated as the number of fruits in the plot,
multiplied by the average of the seed counts from the
collected fruits and divided by plot area. Multiplying seed
density with average mass per seed from the collected fruits
yielded seed mass per square meter. Fruit set was obtained as
the ratio of fruits (2005) and flowers (2004) per plot.
Local environmental variables were collected during June
2005 within each plot. Both the biotic and abiotic environment
was sampled. For each plot, the potential annual direct radi-
ation (MJ cm�2 yr�1) was calculated based on its latitude, slope
and aspect using the formula of McCune and Keon (2002).
Canopy closure (%), i.e. the proportion of the sky hemisphere
obscured by vegetation, was evaluated as the mean of 4
measurements with a spherical densiometer in each wind
direction at 1 m height (Jennings et al., 1999). Soil depth above
rock material (cm) was determined as the mean of 10
measurements with a metal pin. For each plot, cover (%) of
bare ground, rock, litter, moss, small woody debris, dominant
grasses, sedges and shrubs was also estimated. Small woody
debris was often present after management interventions, e.g.
shrub chopping. Shrub height (cm) was calculated as the
mean height of all different species.
Habitat fragmentation was determined within a GIS
(ArcView 3.2a; ESRI, 2000). Grassland fragment area was
calculated based on digitized topographical maps and aerial
photographs, complemented with field surveys. Population
isolation was computed as the edge-to-edge distance to the
nearest grassland fragment with a Colchicum population
using the Nearest Features extension (Jenness, 2007).
Connectivity measures, derived from Hanski’s Incidence
Function Model (IFM; Hanski, 1994), proved to be highly
correlated with population size. To prevent inter-correla-
tions among independent variables in our analyses, we
therefore preferred to use the average of five simple edge-to-
edge distances to the nearest Colchicum populations (isola-
tion variable NN5).
Total population size was counted as the number of flow-
ering individuals in 2004 within each grassland fragment.
2.4. Data analysis
In a first step, the impact of grassland fragment area and
isolation on patch occupancy of all C. autumnale populations in
the study area was assessed using multiple logistic regression.
To reduce the number of environmental variables we
conducted a preliminary correlation analysis, consisting of
both a principal component analysis (PCA with VARIMAX
rotation) and a simple correlation analysis (Pearson’s corre-
lation coefficients) among individual environmental,
population-structural and plant performance variables. Due
to the nested structure of the plots within each grassland
fragment, we applied linear mixed models with grassland
fragments as random factor. Population structure and plant
performance were treated as independent variables, while
population size and isolation and the selected local environ-
mental variables were considered as covariates. Preliminary
analyses ensured us that random slopes could be left out.
Hence, only the random intercept was retained to evaluate the
among-fragment variance component.
All variables were transformed prior to analysis (Table 2
and Appendices 1 and 2) and all analyses were performed in
SPSS 13.0 for Windows (SPSS, 2005).
3. Results
3.1. Habitat occupancy and configuration
Within the study area, C. autumnale occurred in 26 of the 64
investigated grassland fragments (40.6%, Fig. 1), of which 17
were sampled for this study (Table 1). Average area of the
occupied grasslands was 0.911 ha, ranging from 0.087 to
4.07 ha (Table 1). Occupied patches were on average 720 m

Table 2 – Factor loadings of the four principal components (eigenvalue > 1) for 13 local environmental variables of 58 plotsof Colchicum after VARIMAX rotation
Transformation PC1 PC2 PC3 PC4
Eigenvalue 2.896 2.223 1.591 1.234
Variance explained 22.275 17.098 12.236 9.494
PotDirRad (MJ cm�2 yr�1) – �0.382 �0.123 0.113 �0.492
Soil depth (cm) x2 �0.146 �0.107 0.837 �0.089
Bare ground (%) x�1/2 0.235 �0.708 �0.200 �0.163
Rock (%) x�1/2 �0.198 �0.134 �0.779 �0.151
Moss (%) sinh(x1/2 ) 0.124 0.637 �0.137 0.034
Litter (%) – 0.102 0.572 0.320 �0.193
Small woody debris (%) x�1/2 0.120 �0.625 0.274 0.040
Vegetation height (cm) – 0.064 0.584 0.049 0.601
Grasses (%) – �0.441 0.576 0.285 0.039
Sedges (%) x�1/2 �0.172 �0.154 0.091 0.804
Canopy closure (%) sin�1(x1/2 ) 0.826 0.005 0.053 �0.032
Shrub height (cm) log(x) 0.754 0.027 0.123 �0.013
Shrub cover (%) log(x) 0.850 �0.212 �0.098 0.058
Loadings in italic indicate significant ( p� 0.05) correlations between variables and principal components. Each variable is attributed to a single
principal component based on the most significant correlation, indicated in bold.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 73
apart from each other. Average distance to the nearest 5
occupied patches (NN5 avg) was 1572 m.
Patch occupancy was not determined by the area of the
grassland fragment (c2¼ 0.379, p¼ 0.538), neither by its isola-
tion (c2¼ 0.481; p¼ 0.488) nor by their interaction (c2¼ 0.679;
p¼ 0.410).
0
100
200
300
400
500
600
P2 TL P1 DM B1 PL RO L2 P1 M2
MS H T1 Bo oT CA TL
Po
pu
latio
n size
0
0.5
1
1.5
2
2.5
3
3.5
Iso
latio
n (km
)
Population sizeNN5 (km)
3.2. Environmental gradients
The first environmental gradient was characterised by
decreasing local light availability due to increasing shrub
cover (Table 2), and to a lesser extent by decreasing global light
interception (PotDirRad, intrinsic to local physiography).
Increasing amount of soil cover by woody debris, litter,
mosses and dominant grasses defined the second principal
component. The first two principal components explained
39.3% of the variance among the 13-variable dataset. A third
component was highly correlated with soil depth and super-
ficial rock cover and added 12.2% to the explained variance.
Although highly informative to understand the major
environmental gradients that were present in the sampled
population of C. autumnale, the PCs did not explain any vari-
ation in the mixed model analyses and were therefore
replaced by the individual environmental variables that
proved to explain most of the variance in the dataset: shrub
cover, cover of small woody debris and soil depth. These
variables showed both highest factor loadings on each of the
first three orthogonal PCA axes with eigenvalues exceeding 1
and exhibited the highest correlations with population
structure and plant performance. These 3 variables were
therefore used in further analyses.
T C T T M TA R C
Grassland fragment
Fig. 2 – Size (number of flowering adults) and isolation
(average of the 5 nearest distances to neighbour
populations, NN5) of the 17 sampled Colchicum autumnale
populations (r [ L0.099; p [ 0.705).
3.3. Population size and structure of C. autumnale
Population size, measured as the number of flowering individ-
uals, variedfrom10to500 (mean:123,SD:139) (Fig.2).Population
size was not affected by grassland area (r¼ 0.395, p¼ 0.117,
n¼ 17) or landscape isolation (r¼�0.448, p¼ 0.071, n¼ 17), but
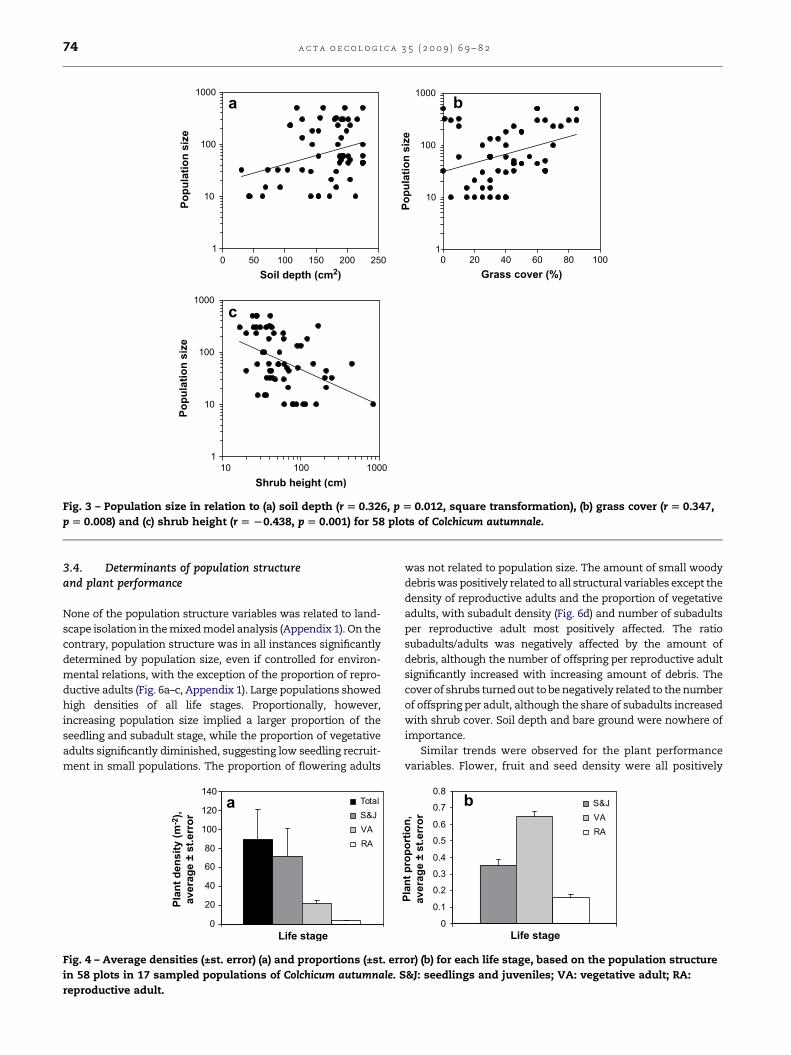
was significantly correlated with average soil depth (r¼ 0.326,
p¼ 0.012, n¼ 58), grass cover (r¼ 0.347, p¼ 0.008, n¼ 58) and
shrub height (r¼�0.438, p¼ 0.001, n¼ 58) (Fig. 3). No relation-
ships between isolation and environment were observed.
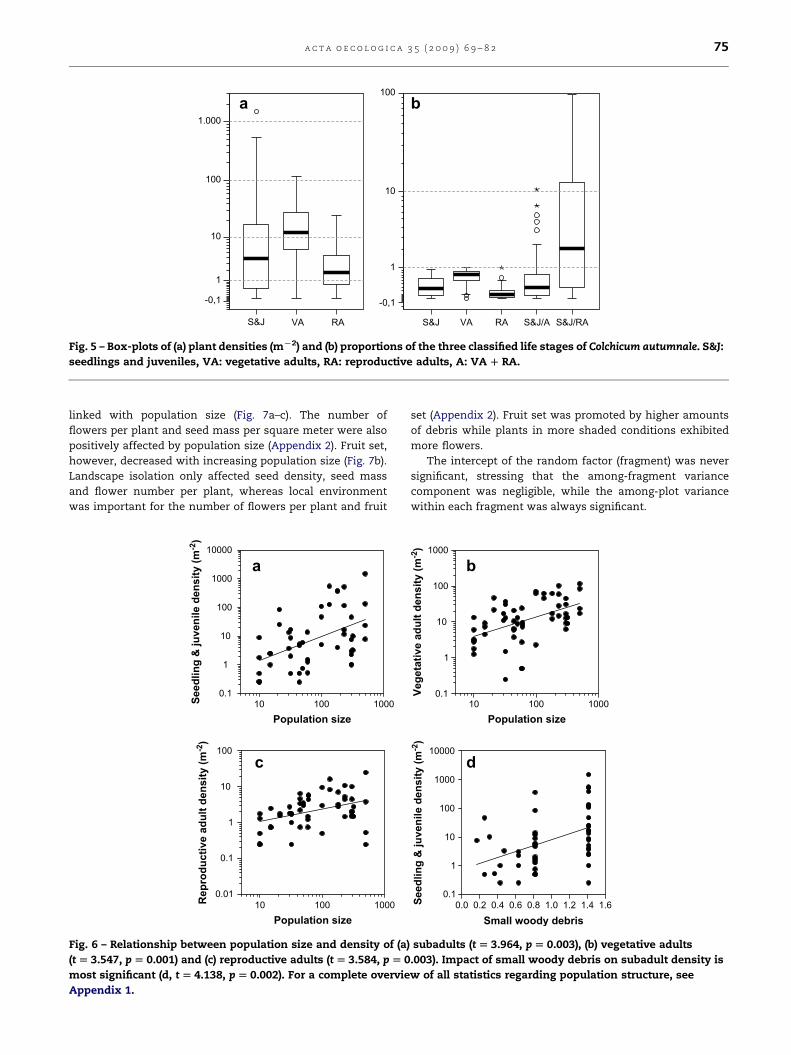
Average plant density varied from 3.31 reproductive adults
(RA) per square meter to densities of 21.87 and 72.11 m�2 for
vegetative adults (VA) and subadults (seedlings and juveniles,
S&J), respectively (Fig. 4a). However, population structure was
highly variable. Especially the number of subadults showed
a high coefficient of variation of 309%, and values of 118% and
128% for vegetative and reproductive adults, respectively. The
skewed distribution of each of the life stages is revealed by the
medians and quartiles. In 75% of the plots subadult densities
did not exceed 16.72 m�2. For RA and VA these densities were
4.25 and 26.87 m�2, respectively (Fig. 5). Thus, proportionally
and on average, vegetative adults were the most abundant
(64.7%), followed by the subadult (35.1%) and reproductive
stage (15.4%) (Fig. 4b).
Soil depth (cm2)
Po
pu
latio
n size
1
10
100
1000
Grass cover (%)
0 50 100 150 200 250 0 20 40 60 80 100
Po
pu
latio
n size
1
10
100
1000
Shrub height (cm)
10 100 1000
Po
pu
latio
n size
1
10
100
1000
ba
c
Fig. 3 – Population size in relation to (a) soil depth (r [ 0.326, p [ 0.012, square transformation), (b) grass cover (r [ 0.347,
p [ 0.008) and (c) shrub height (r [ L0.438, p [ 0.001) for 58 plots of Colchicum autumnale.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 274
3.4. Determinants of population structureand plant performance
None of the population structure variables was related to land-
scape isolation in the mixed model analysis (Appendix 1). On the
contrary, population structure was in all instances significantly
determined by population size, even if controlled for environ-
mental relations, with the exception of the proportion of repro-
ductive adults (Fig. 6a–c, Appendix 1). Large populations showed
high densities of all life stages. Proportionally, however,
increasing population size implied a larger proportion of the
seedling and subadult stage, while the proportion of vegetative
adults significantly diminished, suggesting low seedling recruit-
ment in small populations. The proportion of flowering adults
0
20
40
60
80
100
120
140
Life stage
Pla
nt d
en
sity
(m
-2),
av
era
ge
± s
t.e
rro
r
TotalS&JVARA
a
Fig. 4 – Average densities (±st. error) (a) and proportions (±st. err
in 58 plots in 17 sampled populations of Colchicum autumnale. S
reproductive adult.
was not related to population size. The amount of small woody
debris was positively related to all structural variables except the
density of reproductive adults and the proportion of vegetative
adults, with subadult density (Fig. 6d) and number of subadults
per reproductive adult most positively affected. The ratio
subadults/adults was negatively affected by the amount of
debris, although the number of offspring per reproductive adult
significantly increased with increasing amount of debris. The
cover of shrubs turned out to be negatively related to the number
of offspring per adult, although the share of subadults increased
with shrub cover. Soil depth and bare ground were nowhere of
importance.
Similar trends were observed for the plant performance
variables. Flower, fruit and seed density were all positively
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Life stage
Pla
nt p
ro
po
rtio
n,
av
era
ge
± s
t.e
rro
r
S&JVARA
b
or) (b) for each life stage, based on the population structure
&J: seedlings and juveniles; VA: vegetative adult; RA:
RAVAS&J
1.000
100
10
1
-0,1
S&J/RAS&J/ARAVAS&J
100
10
1
-0,1
a b
Fig. 5 – Box-plots of (a) plant densities (mL2) and (b) proportions of the three classified life stages of Colchicum autumnale. S&J:
seedlings and juveniles, VA: vegetative adults, RA: reproductive adults, A: VA D RA.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 75
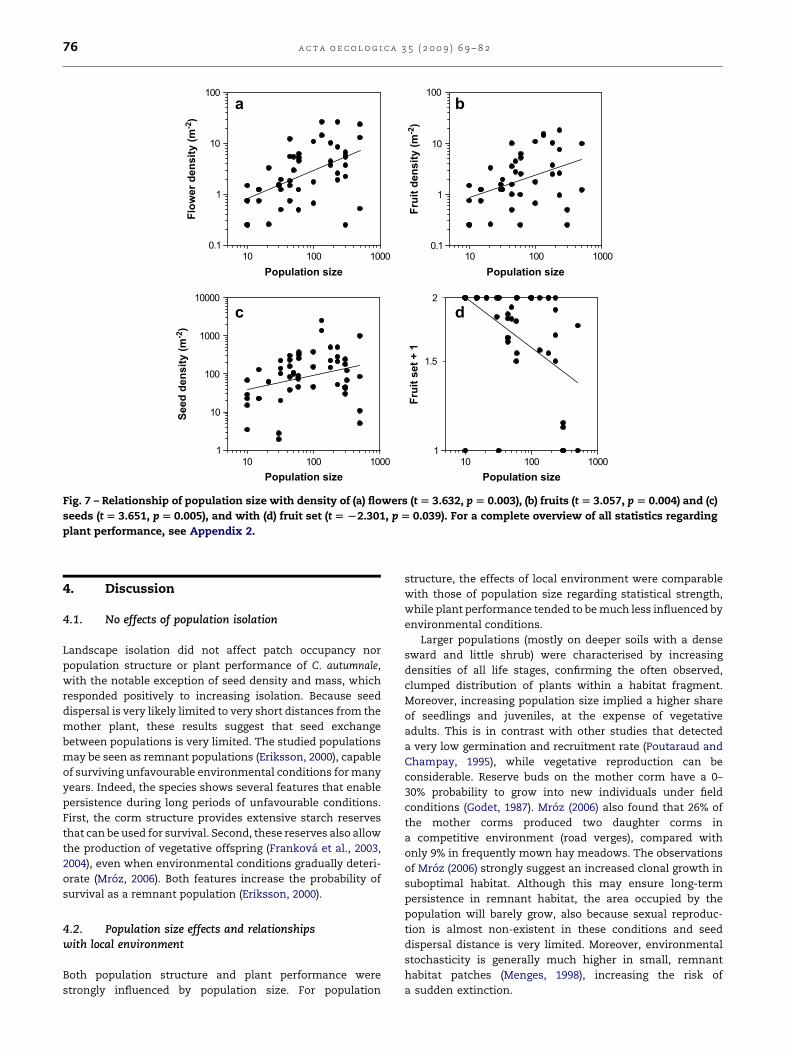
linked with population size (Fig. 7a–c). The number of
flowers per plant and seed mass per square meter were also
positively affected by population size (Appendix 2). Fruit set,
however, decreased with increasing population size (Fig. 7b).
Landscape isolation only affected seed density, seed mass
and flower number per plant, whereas local environment
was important for the number of flowers per plant and fruit
Population size
10 100 1000Seed
lin
g &
ju
ven
ile d
en
sity (m
-2)
0.1
1
10
100
1000
10000
Population size
10 100 1000Rep
ro
du
ctive ad
ult d
en
sity (m
-2)
0.01
0.1
1
10
100
a
c
Fig. 6 – Relationship between population size and density of (a)
(t [ 3.547, p [ 0.001) and (c) reproductive adults (t [ 3.584, p [ 0
most significant (d, t [ 4.138, p [ 0.002). For a complete overvie
Appendix 1.
set (Appendix 2). Fruit set was promoted by higher amounts
of debris while plants in more shaded conditions exhibited
more flowers.
The intercept of the random factor (fragment) was never
significant, stressing that the among-fragment variance
component was negligible, while the among-plot variance
within each fragment was always significant.
Population size
10 100 1000
Veg
etative ad
ult d
en
sity (m
-2)
0.1
1
10
100
1000
Small woody debris
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6Seed
lin
g &
ju
ven
ile d
en
sity (m
-2)
0.1
1
10
100
1000
10000
b
d
subadults (t [ 3.964, p [ 0.003), (b) vegetative adults
.003). Impact of small woody debris on subadult density is
w of all statistics regarding population structure, see
Population size
10 100 1000
Flo
wer d
en
sity (m
-2)
0.1
1
10
100
Population size
10 100 1000
Fru
it d
en
sity (m
-2)
0.1
1
10
100
Population size
10 100 1000
Seed
d
en
sity (m
-2)
1
10
100
1000
10000
Population size
10 100 1000
Fru
it set +
1
1.5
2
1
a b
dc
Fig. 7 – Relationship of population size with density of (a) flowers (t [ 3.632, p [ 0.003), (b) fruits (t [ 3.057, p [ 0.004) and (c)
seeds (t [ 3.651, p [ 0.005), and with (d) fruit set (t [ L2.301, p [ 0.039). For a complete overview of all statistics regarding
plant performance, see Appendix 2.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 276
4. Discussion
4.1. No effects of population isolation
Landscape isolation did not affect patch occupancy nor
population structure or plant performance of C. autumnale,
with the notable exception of seed density and mass, which
responded positively to increasing isolation. Because seed
dispersal is very likely limited to very short distances from the
mother plant, these results suggest that seed exchange
between populations is very limited. The studied populations
may be seen as remnant populations (Eriksson, 2000), capable
of surviving unfavourable environmental conditions for many
years. Indeed, the species shows several features that enable
persistence during long periods of unfavourable conditions.
First, the corm structure provides extensive starch reserves
that can be used for survival. Second, these reserves also allow
the production of vegetative offspring (Frankova et al., 2003,
2004), even when environmental conditions gradually deteri-
orate (Mroz, 2006). Both features increase the probability of
survival as a remnant population (Eriksson, 2000).
4.2. Population size effects and relationshipswith local environment
Both population structure and plant performance were
strongly influenced by population size. For population
structure, the effects of local environment were comparable
with those of population size regarding statistical strength,
while plant performance tended to be much less influenced by
environmental conditions.
Larger populations (mostly on deeper soils with a dense
sward and little shrub) were characterised by increasing
densities of all life stages, confirming the often observed,
clumped distribution of plants within a habitat fragment.
Moreover, increasing population size implied a higher share
of seedlings and juveniles, at the expense of vegetative
adults. This is in contrast with other studies that detected
a very low germination and recruitment rate (Poutaraud and
Champay, 1995), while vegetative reproduction can be
considerable. Reserve buds on the mother corm have a 0–
30% probability to grow into new individuals under field
conditions (Godet, 1987). Mroz (2006) also found that 26% of
the mother corms produced two daughter corms in
a competitive environment (road verges), compared with
only 9% in frequently mown hay meadows. The observations
of Mroz (2006) strongly suggest an increased clonal growth in
suboptimal habitat. Although this may ensure long-term
persistence in remnant habitat, the area occupied by the
population will barely grow, also because sexual reproduc-
tion is almost non-existent in these conditions and seed
dispersal distance is very limited. Moreover, environmental
stochasticity is generally much higher in small, remnant
habitat patches (Menges, 1998), increasing the risk of
a sudden extinction.

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 77
Little is known about the flower and seed ecology of
C. autumnale. Until now, most research concentrated on
seed production and seed alkaloid content for medicinal
purposes (Poutaraud and Girardin, 2005, 2006). Results
point towards increasing seed yield under competitive-free
growth conditions, caused by a yearly vegetative pop-
ulation increase of about 50%, despite a slight decrease in
seed yield per plant due to increasing intraspecific
competition among ramets (Poutaraud and Girardin, 2003).
It is thus suggested that generative reproduction has
a tendency to decline after long periods of vegetative
growth. Despite the fact that population growth is lower in
grassland (about 15% yr�1), although with considerably
more fluctuations during population build-up, an analo-
gous reduction of sexual reproduction can be expected
here as well, as confirmed by our results. First, our data
showed that competition for light significantly reduced the
number of flowers per plant, which is confirmed by other
studies (Godet, 1987; Mroz, 2006). Secondly, fruit set was
higher in open vegetation, as quantified by high amounts
of woody debris associated with recent shrub removal
management interventions. Thus, our results suggest
a significant decrease in sexual reproduction when grass-
land habitat quality deteriorates. Surprisingly, fruit set was
higher in small populations. Normally, smaller populations
receive less pollinator attention, resulting in lower fruit
and seed set (e.g. Jennersten, 1988). Although higher visi-
tation rates by pollinators can be observed in smaller
populations (e.g. Bosch and Waser, 1999; Mustajarvi et al.,
2001; Goverde et al., 2002), this generally promotes
inbreeding (Richards, 2000) that negatively affects offspring
fitness (Lennartsson, 2002). However, no data are available
on the influence of selfing and outcrossing rates on fruit
set, seed production and offspring performance of C.
autumnale. It must be noted, however, that pollinator
activity during anthesis of C. autumnale is generally low
due to the often unfavourable weather conditions during
September and October, probably promoting autonomous
self-pollination. Despite this uncertainty, the lower fruit set
in large populations was compensated for by a higher
density of reproductive adults (Appendix 1) each having
more flowers. Consequently, seed production per unit area
significantly increased (Appendix 2).
We observed high variation in recruitment (Fig. 5), even
between plots within fragments. Subadult density was
highly promoted by the cover of small woody debris.
Probably, small woody debris provides shelter and assures
sufficient soil surface humidity to enable germination.
Therefore, we assume that the availability of very specific
microsites might be limiting successful germination and
establishment, possibly in combination with deteriorating
seed quality in small populations. However, a more
detailed study of the seed and recruitment ecology would
be of interest.
4.3. Management guidelines
Although small fragments did not necessarily harbour small
populations, and no effects of landscape isolation were found,
it is likely that small and isolated fragments are especially
prone to deterioration of habitat quality because of rapid
shrub encroachment during natural succession towards
forest. Moreover, these fragments often lack conservation
protection status, making them even more vulnerable to this
threat. C. autumnale can survive beneath a dense canopy, but
the number of flowering and vegetative adults steadily
decreases and fruit set is significantly diminished, leading to
a shrinking population size and a lack of sexual reproduction.
Long-term survival may thus strongly depend on recurrent
phases with sexual reproduction, given that (1) population
genetic variability may be low due to long periods of stasis or
gradual population regression during which genotypes
disappear, (2) dispersal capacity of daughter ramets is
extremely low, (3) whole genotypes can be destroyed at once
due to the clumped nature of mortality in response to
disturbance and (4) C. autumnale does not form a persistent
seed bank. This was also observed in a detailed long-term
study of the bulb species Allium tricoccum in the forests of
Quebec (Nault and Gagnon, 1993).
Since highest densities of subadults were observed after
recent shrub removal, with the resulting small woody debris
still present, cyclic management with shrub removal followed
by recurrent regrowth might favour C. autumnale populations.
This treatment indeed allows for phases of both vegetative
(during encroachement) and sexual (directly after clearance)
population growth. Management has to look for a balance
between both modes of reproduction in order to ensure long-
term survival. However, to ensure open grassland conditions
during which seedlings can establish and reach the adult
state, periodic shrub removal should be accompanied by
extensive grazing or late-summer mowing, i.e. when plants
are dormant, to reduce excessive competition. Grazing can
occur almost year-round because plants are avoided by cattle
due to their poisonous nature. Excessive trampling has to be
avoided though, especially during anthesis. Additionally,
small disturbances by animal treading can also increase
microsite availability, stimulating seed germination and
seedling establishment.
5. Concluding remarks
We found evidence that meadow saffron did not suffer
from a long history of habitat fragmentation. In combina-
tion with its high clonal growth capacity, these results
suggest that C. autumnale currently occurs as a collection of
basically unconnected, remnant populations within the
landscape. Therefore, survival of these population
remnants is especially conditional upon local growth
conditions, as suggested by the strong effects of population
size and environment on population structure and repro-
ductive plant performance. Since reproductive success is
especially dependent on open vegetation with sufficient
microsite availability for seed germination and establish-
ment, extensive grazing is suggested as the most beneficial
management for population expansion. However, to
unravel the ultimate reasons behind the impact of pop-
ulation size on reproductive success and to assess future
response to changing environment, further insight into the
reproductive mechanisms of C. autumnale is needed.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 278
Acknowledgements
This study was funded by a grant from the Flemish Fund for
Scientific Research (FWO) and the BIOCORE project EV/01/26A,
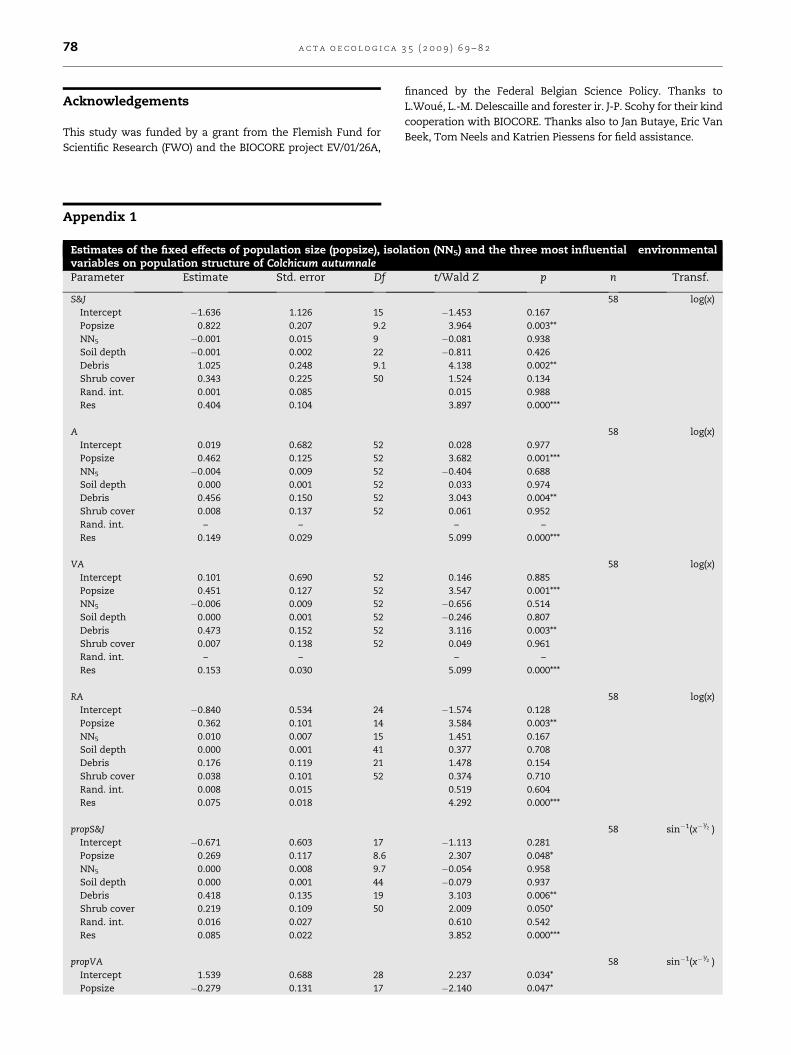
Appendix 1
Estimates of the fixed effects of population size (popsize), isolvariables on population structure of Colchicum autumnaleParameter Estimate Std. error Df
S&J
Intercept �1.636 1.126 15
Popsize 0.822 0.207 9.2
NN5 �0.001 0.015 9
Soil depth �0.001 0.002 22
Debris 1.025 0.248 9.1
Shrub cover 0.343 0.225 50
Rand. int. 0.001 0.085
Res 0.404 0.104
A
Intercept 0.019 0.682 52
Popsize 0.462 0.125 52
NN5 �0.004 0.009 52
Soil depth 0.000 0.001 52
Debris 0.456 0.150 52
Shrub cover 0.008 0.137 52
Rand. int. – –
Res 0.149 0.029
VA
Intercept 0.101 0.690 52
Popsize 0.451 0.127 52
NN5 �0.006 0.009 52
Soil depth 0.000 0.001 52
Debris 0.473 0.152 52
Shrub cover 0.007 0.138 52
Rand. int. – –
Res 0.153 0.030
RA
Intercept �0.840 0.534 24
Popsize 0.362 0.101 14
NN5 0.010 0.007 15
Soil depth 0.000 0.001 41
Debris 0.176 0.119 21
Shrub cover 0.038 0.101 52
Rand. int. 0.008 0.015
Res 0.075 0.018
propS&J
Intercept �0.671 0.603 17
Popsize 0.269 0.117 8.6
NN5 0.000 0.008 9.7
Soil depth 0.000 0.001 44
Debris 0.418 0.135 19
Shrub cover 0.219 0.109 50
Rand. int. 0.016 0.027
Res 0.085 0.022
propVA
Intercept 1.539 0.688 28
Popsize �0.279 0.131 17
financed by the Federal Belgian Science Policy. Thanks to
L.Woue, L.-M. Delescaille and forester ir. J-P. Scohy for their kind
cooperation with BIOCORE. Thanks also to Jan Butaye, Eric Van
Beek, Tom Neels and Katrien Piessens for field assistance.
ation (NN5) and the three most influential environmental
t/Wald Z p n Transf.
58 log(x)
�1.453 0.167
3.964 0.003**
�0.081 0.938
�0.811 0.426
4.138 0.002**
1.524 0.134
0.015 0.988
3.897 0.000***
58 log(x)
0.028 0.977
3.682 0.001***
�0.404 0.688
0.033 0.974
3.043 0.004**
0.061 0.952
– –
5.099 0.000***
58 log(x)
0.146 0.885
3.547 0.001***
�0.656 0.514
�0.246 0.807
3.116 0.003**
0.049 0.961
– –
5.099 0.000***
58 log(x)
�1.574 0.128
3.584 0.003**
1.451 0.167
0.377 0.708
1.478 0.154
0.374 0.710
0.519 0.604
4.292 0.000***
58 sin�1(x�1/2 )
�1.113 0.281
2.307 0.048*
�0.054 0.958
�0.079 0.937
3.103 0.006**
2.009 0.050*
0.610 0.542
3.852 0.000***
58 sin�1(x�1/2 )
2.237 0.034*
�2.140 0.047*
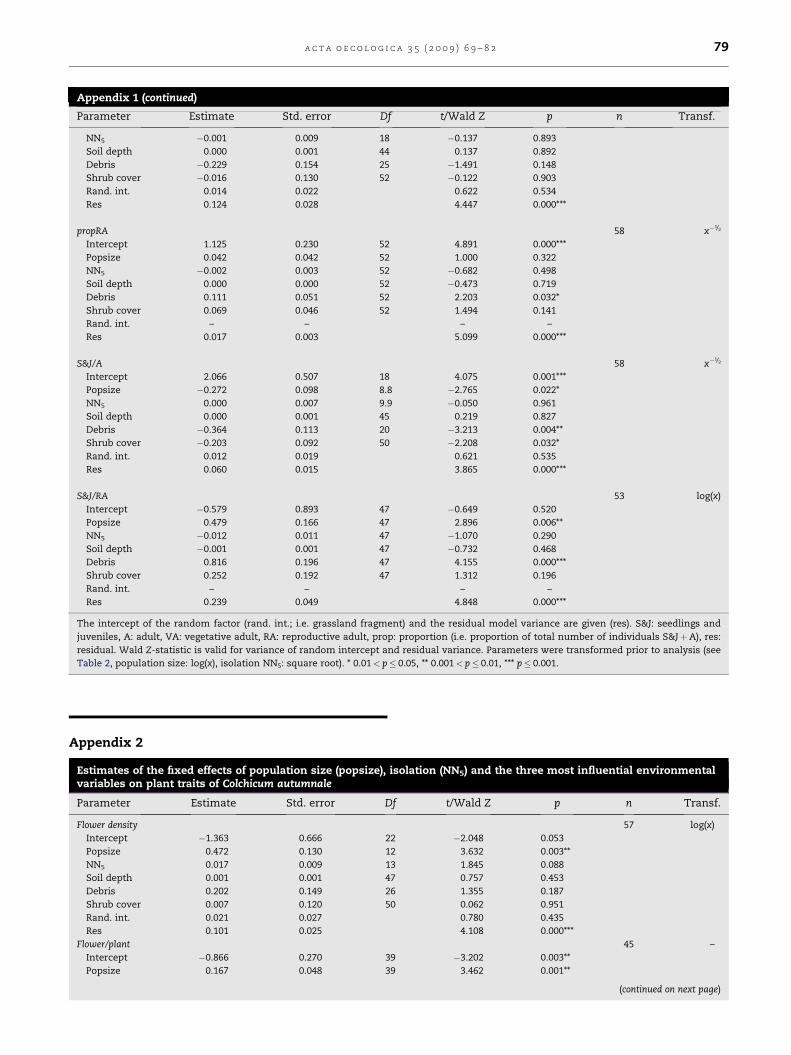
Appendix 1 (continued)
Parameter Estimate Std. error Df t/Wald Z p n Transf.
NN5 �0.001 0.009 18 �0.137 0.893
Soil depth 0.000 0.001 44 0.137 0.892
Debris �0.229 0.154 25 �1.491 0.148
Shrub cover �0.016 0.130 52 �0.122 0.903
Rand. int. 0.014 0.022 0.622 0.534
Res 0.124 0.028 4.447 0.000***
propRA 58 x�1/2
Intercept 1.125 0.230 52 4.891 0.000***
Popsize 0.042 0.042 52 1.000 0.322
NN5 �0.002 0.003 52 �0.682 0.498
Soil depth 0.000 0.000 52 �0.473 0.719
Debris 0.111 0.051 52 2.203 0.032*
Shrub cover 0.069 0.046 52 1.494 0.141
Rand. int. – – – –
Res 0.017 0.003 5.099 0.000***
S&J/A 58 x�1/2
Intercept 2.066 0.507 18 4.075 0.001***
Popsize �0.272 0.098 8.8 �2.765 0.022*
NN5 0.000 0.007 9.9 �0.050 0.961
Soil depth 0.000 0.001 45 0.219 0.827
Debris �0.364 0.113 20 �3.213 0.004**
Shrub cover �0.203 0.092 50 �2.208 0.032*
Rand. int. 0.012 0.019 0.621 0.535
Res 0.060 0.015 3.865 0.000***
S&J/RA 53 log(x)
Intercept �0.579 0.893 47 �0.649 0.520
Popsize 0.479 0.166 47 2.896 0.006**
NN5 �0.012 0.011 47 �1.070 0.290
Soil depth �0.001 0.001 47 �0.732 0.468
Debris 0.816 0.196 47 4.155 0.000***
Shrub cover 0.252 0.192 47 1.312 0.196
Rand. int. – – – –
Res 0.239 0.049 4.848 0.000***
The intercept of the random factor (rand. int.; i.e. grassland fragment) and the residual model variance are given (res). S&J: seedlings and
juveniles, A: adult, VA: vegetative adult, RA: reproductive adult, prop: proportion (i.e. proportion of total number of individuals S&JþA), res:
residual. Wald Z-statistic is valid for variance of random intercept and residual variance. Parameters were transformed prior to analysis (see
Table 2, population size: log(x), isolation NN5: square root). * 0.01< p� 0.05, ** 0.001< p� 0.01, *** p� 0.001.
Appendix 2
Estimates of the fixed effects of population size (popsize), isolation (NN5) and the three most influential environmentalvariables on plant traits of Colchicum autumnale
Parameter Estimate Std. error Df t/Wald Z p n Transf.
Flower density 57 log(x)
Intercept �1.363 0.666 22 �2.048 0.053
Popsize 0.472 0.130 12 3.632 0.003**
NN5 0.017 0.009 13 1.845 0.088
Soil depth 0.001 0.001 47 0.757 0.453
Debris 0.202 0.149 26 1.355 0.187
Shrub cover 0.007 0.120 50 0.062 0.951
Rand. int. 0.021 0.027 0.780 0.435
Res 0.101 0.025 4.108 0.000***
Flower/plant 45 –
Intercept �0.866 0.270 39 �3.202 0.003**
Popsize 0.167 0.048 39 3.462 0.001**
(continued on next page)
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 79
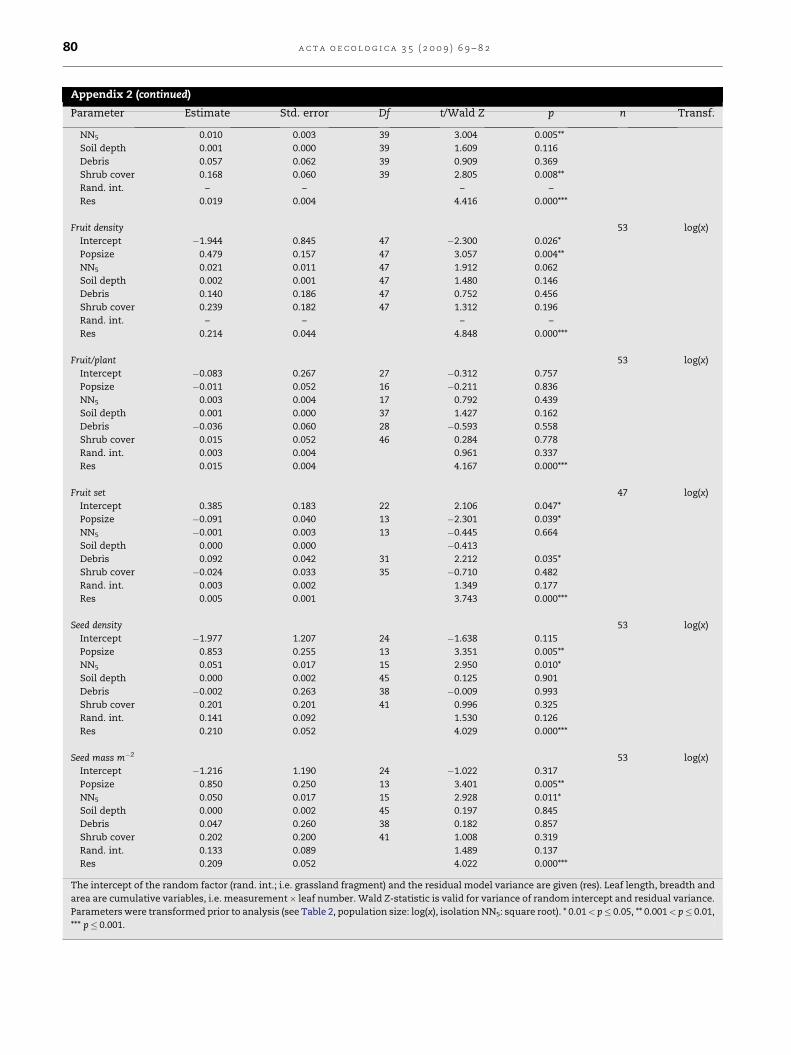
Appendix 2 (continued)
Parameter Estimate Std. error Df t/Wald Z p n Transf.
NN5 0.010 0.003 39 3.004 0.005**
Soil depth 0.001 0.000 39 1.609 0.116
Debris 0.057 0.062 39 0.909 0.369
Shrub cover 0.168 0.060 39 2.805 0.008**
Rand. int. – – – –
Res 0.019 0.004 4.416 0.000***
Fruit density 53 log(x)
Intercept �1.944 0.845 47 �2.300 0.026*
Popsize 0.479 0.157 47 3.057 0.004**
NN5 0.021 0.011 47 1.912 0.062
Soil depth 0.002 0.001 47 1.480 0.146
Debris 0.140 0.186 47 0.752 0.456
Shrub cover 0.239 0.182 47 1.312 0.196
Rand. int. – – – –
Res 0.214 0.044 4.848 0.000***
Fruit/plant 53 log(x)
Intercept �0.083 0.267 27 �0.312 0.757
Popsize �0.011 0.052 16 �0.211 0.836
NN5 0.003 0.004 17 0.792 0.439
Soil depth 0.001 0.000 37 1.427 0.162
Debris �0.036 0.060 28 �0.593 0.558
Shrub cover 0.015 0.052 46 0.284 0.778
Rand. int. 0.003 0.004 0.961 0.337
Res 0.015 0.004 4.167 0.000***
Fruit set 47 log(x)
Intercept 0.385 0.183 22 2.106 0.047*
Popsize �0.091 0.040 13 �2.301 0.039*
NN5 �0.001 0.003 13 �0.445 0.664
Soil depth 0.000 0.000 �0.413
Debris 0.092 0.042 31 2.212 0.035*
Shrub cover �0.024 0.033 35 �0.710 0.482
Rand. int. 0.003 0.002 1.349 0.177
Res 0.005 0.001 3.743 0.000***
Seed density 53 log(x)
Intercept �1.977 1.207 24 �1.638 0.115
Popsize 0.853 0.255 13 3.351 0.005**
NN5 0.051 0.017 15 2.950 0.010*
Soil depth 0.000 0.002 45 0.125 0.901
Debris �0.002 0.263 38 �0.009 0.993
Shrub cover 0.201 0.201 41 0.996 0.325
Rand. int. 0.141 0.092 1.530 0.126
Res 0.210 0.052 4.029 0.000***
Seed mass m�2 53 log(x)
Intercept �1.216 1.190 24 �1.022 0.317
Popsize 0.850 0.250 13 3.401 0.005**
NN5 0.050 0.017 15 2.928 0.011*
Soil depth 0.000 0.002 45 0.197 0.845
Debris 0.047 0.260 38 0.182 0.857
Shrub cover 0.202 0.200 41 1.008 0.319
Rand. int. 0.133 0.089 1.489 0.137
Res 0.209 0.052 4.022 0.000***
The intercept of the random factor (rand. int.; i.e. grassland fragment) and the residual model variance are given (res). Leaf length, breadth and
area are cumulative variables, i.e. measurement� leaf number. Wald Z-statistic is valid for variance of random intercept and residual variance.
Parameters were transformed prior to analysis (see Table 2, population size: log(x), isolation NN5: square root). * 0.01< p� 0.05, ** 0.001< p� 0.01,
*** p� 0.001.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 280

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 2 81
r e f e r e n c e s
Adriaens, D., Honnay, O., Hermy, M., 2006. No evidence of a plantextinction debt in highly fragmented calcareous grasslands inBelgium. Biological Conservation 133, 212–224.
Bosch, M., Waser, N.M., 1999. Effects of local density onpollination and reproduction in Delphinium nuttallianum andAconitum columbianum (Ranunculaceae). American Journal ofBotany 86, 871–879.
Brown, J.H., Kodric-Brown, A., 1977. Turnover rates in insularbiogeography – effect of immigration on extinction. Ecology58, 445–449.
Butcher, R.W., 1954. Colchicum autumnale L. Journal of Ecology 42,249–257.
Colling, G., Matthies, D., 2006. Effects of habitat deterioration onpopulation dynamics and extinction risk of an endangered,long-lived perennial herb (Scorzonera humilis). Journal ofEcology 94, 959–972.
Ellstrand, N.C., Elam, D.R., 1993. Population geneticconsequences of small population size – implications forplant conservation. Annual Review of Ecology andSystematics 24, 217–242.
Endels, P., Adriaens, D., Verheyen, K., Hermy, M., 2004. Populationstructure and adult plant performance of forest herbs in threecontrasting habitats. Ecography 27, 225–241.
Eriksson, O., 2000. Functional roles of remnant plant populationsin communities and ecosystems. Global Ecology andBiogeography 9, 443–449.
ESRI, 2000. ArcView 3.2a. Enviroinmental Systems ResearchInstitute Inc., Redlands.
Fischer, M., Matthies, D., 1998. Effects of population size onperformance in the rare plant Gentianella germanica. Journal ofEcology 86, 195–204.
Fischer, M., Stocklin, J., 1997. Local extinctions of plants inremnants of extensively used calcareous grasslands 1950–1985. Conservation Biology 11, 727–737.
Frankova, L., Komjathyova, H., Boka, K., Gasparıkova, O.,Psenak, M., 2003. Biochemical and physiological aspects ofdevelopmental cycle of Colchicum autumnale L. BiologiaPlantarum 47, 509–516.
Frankova, L., Cibırova, K., Boka, K., Gasparıkova, O., Psenak, M.,2004. The role of the roots in the life strategy of Colchicumautumnale. Biologia, Bratislava 59, 87–93.
Godet X., 1987. Biologie du colchique (Colchicum autumnale).Multiplication vegetative par voie traditionnelle et in vitro. Ph.D.thesis, University of Blaise Pascal, Clermont-Ferrand.
Goverde, M., Schweizer, K., Baur, B., Erhardt, A., 2002. Small-scalehabitat fragmentation effects on pollinator behaviour:experimental evidence from the bumblebee Bombus veteranuson calcareous grasslands. Biological Conservation 104, 293–299.
Hanski, I., 1998. Metapopulation dynamics. Nature 396, 41–49.Hanski, I., 1994. A practical model of metapopulation dynamics.
Journal of Animal Ecology 63, 151–162.Hegland, S.J., Van Leeuwen, M., Oostermeijer, J.G.B., 2001.
Population structure of Salvia pratensis in relation tovegetation and management of Dutch dry floodplaingrasslands. Journal of Applied Ecology 38, 1277–1289.
Helm, A., Hanski, I., Partel, M., 2006. Slow response of plantspecies richness to habitat loss and fragmentation. EcologyLetters 9, 72–77.
Holsinger, K.E., 2000. Demography and extinction in smallpopulations. In: Young, A.G., Clarke, G.M. (Eds.), Genetics,Demography and Viability of Fragmented Populations.Cambridge University Press, Cambridge, pp. 55–74.
Honnay, O., Jacquemyn, H., 2007. Susceptibility of common andrare plant species to the genetic consequences of habitatfragmentation. Conservation Biology 21, 823–831.
Hooftman, D.A.P., Diemer, M., 2002. Effects of small habitat sizeand isolation on the population structure of common wetlandspecies. Plant Biology 4, 720–728.
Jaehn, F., Roux, J., 1986. Architecture and annual cycle of theAutumn crocus (Colchicum autumnale L.). Bulletin de laSociete Botanique de France-Lettres Botaniques 133,225–233.
Jenness, J., 2007. Nearest Features. Jenness Enterprises.Jennersten, O., 1988. Pollination in Dianthus deltoides
(Caryophyllaceae) – effects of habitat fragmentation onvisitation and seed set. Conservation Biology 2, 359–366.
Jennings, S.B., Brown, N.D., Sheil, D., 1999. Assessing forestcanopies and understorey illumination: canopy closure,canopy cover and other measures. Forestry 72, 59–73.
Lienert, J., Diemer, M., Schmid, B., 2002. Effects of habitatfragmentation on population structure and fitnesscomponents of the wetland specialist Swertia perennisL. (Gentianaceae). Basic and Applied Ecology 3, 101–114.
Lennartsson, T., 2002. Extinction thresholds and disrupted plant–pollinator interactions in fragmented plant populations.Ecology 83, 3060–3072.
Maes, D., Van Dyck, H., 2001. Butterfly diversity loss in Flanders(North Belgium): Europe’s worst case scenario? BiologicalConservation 99, 263–276.
Matthies, D., Brauer, I., Maibom, W., Tscharntke, T., 2004.Population size and the risk of local extinction: empiricalevidence from rare plants. Oikos 105, 481–488.
McCune, B., Keon, D., 2002. Equations for potential annual directincident radiation and heat load. Journal of Vegetation Science13, 603–606.
Menges, E.S., 1998. Evaluating extinction risks in plantpopulations. In: Fiedler, P.L., Kareiva, P.M. (Eds.), ConservationBiology for the Coming Decade. Chapman and Hall, New York,pp. 49–65.
Menges, E.S., Dolan, R.W., 1998. Demographic viability ofpopulations of Silene regia in midwestern prairies:relationships with fire management, genetic variation,geographic location, population size and isolation. Journal ofEcology 86, 63–78.
Mroz, L., 2006. Variation in stage structure and fitness traitsbetween road verge and meadow populations of Colchicumautumnale (Liliaceae): effects of habitat quality. Acta SocietatisBotanicorum 75, 69–78.
Munzbergova, Z., 2006. Effect of population size on the prospectof species survival. Folia Geobotanica 41, 137–150.
Mustajarvi, K., Siikamaki, P., Rytkonen, S., Lammi, A., 2001.Consequences of plant population size and density for plant–pollinator interactions and plant performance. Journal ofEcology 89, 80–87.
Nault, A., Gagnon, D., 1993. Ramet demography of Alliumtricoccum, a spring ephemeral, perennial forest herb. Journal ofEcology 81, 101–119.
Newman, D., Pilson, D., 1997. Increased probability of extinction dueto decreased genetic effective population size: experimentalpopulations of Clarkia pulchella. Evolution 51, 354–362.
Oostermeijer, J.G.B., Vaneijck, M.W., Dennijs, J.C.M., 1994.Offspring fitness in relation to population size and geneticvariation in the rare perennial plant species Gentianapneumonanthe (Gentianaceae). Oecologia 97, 289–296.
Oostermeijer, J.G.B., Brugman, M.L., De Boer, E.R., den Nijs, J.C.M.,1996. Temporal and spatial variation in the demography ofGentiana pneumonanthe, a rare perennial herb. Journal ofEcology 84, 153–166.
Persson, K., 1993. Reproductive strategies and evolution inColchicum. In: Proceedings of the Fifth OPTIMA Meeting,Istanbul, 8–15 September 1986, pp. 397–414.
Poutaraud, A., Champay, N., 1995. Le Colchique: une plantemedicinale a domestiquer. Revue Suisse d’Agriculture 27, 93–100.

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 6 9 – 8 282
Poutaraud, A., Girardin, P., 2006. Agronomical and chemicalvariability of Colchicum autumnale accessions. CanadianJournal of Plant Science 86, 547–555.
Poutaraud, A., Girardin, P., 2005. Influence of chemicalcharacteristics of soil on mineral and alkaloid seed contents ofColchicum autumnale. Environmental and Experimental Botany54, 101–108.
Poutaraud, A., Girardin, P., 2003. Seed yield and components ofalkaloid of Meadow saffron (Colchicum autumnale) in naturalgrassland and under cultivation. Canadian Journal of PlantScience 83, 23–29.
Richards, C.M., 2000. Inbreeding depression and genetic rescue ina plant metapopulation. American Naturalist 155, 383–394.
SPSS, 2005. SPSS 13.0 for Windows. SPSS Inc., Chicago.
Thompson, K., Bakker, J.P., Bekker, R.M., 1997. The Soil SeedBanks of North-West Europe: Methodology, Density andLongevity. Cambridge University Press, Cambridge.
Tilman, D., May, R.M., Lehman, C.L., Nowak, M.A., 1994.Habitat destruction and the extinction debt. Nature 371,65–66.
Vellend, M., Verheyen, K., Jacquemyn, H., Kolb, A., Van Calster, H.,Peterken, G., Hermy, M., 2006. Extinction debt of forest plantspersists for more than a century following habitatfragmentation. Ecology 87, 542–548.
Wenzel, M., Schmitt, T., Weitzel, M., Seitz, A., 2006. The severedecline of butterflies on western German calcareousgrasslands during the last 30 years: a conservation problem.Biological Conservation 128, 542–552.
Related Documents











