of March 31, 2016. This information is current as Expression To Suppress Monocyte Adhesion 2 Integrin β Conjugated Linoleic Acid Targets William G. James and Orina Belton Monica de Gaetano, Eugene Dempsey, Simone Marcone, http://www.jimmunol.org/content/191/8/4326 doi: 10.4049/jimmunol.1300990 September 2013; 2013; 191:4326-4336; Prepublished online 18 J Immunol Material Supplementary 0.DC1.html http://www.jimmunol.org/content/suppl/2013/09/18/jimmunol.130099 References http://www.jimmunol.org/content/191/8/4326.full#ref-list-1 , 14 of which you can access for free at: cites 49 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2013 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on March 31, 2016 http://www.jimmunol.org/ Downloaded from by guest on March 31, 2016 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of March 31, 2016.This information is current as
Expression To Suppress Monocyte Adhesion2 IntegrinβConjugated Linoleic Acid Targets
William G. James and Orina BeltonMonica de Gaetano, Eugene Dempsey, Simone Marcone,
http://www.jimmunol.org/content/191/8/4326doi: 10.4049/jimmunol.1300990September 2013;
2013; 191:4326-4336; Prepublished online 18J Immunol
MaterialSupplementary
0.DC1.htmlhttp://www.jimmunol.org/content/suppl/2013/09/18/jimmunol.130099
Referenceshttp://www.jimmunol.org/content/191/8/4326.full#ref-list-1
, 14 of which you can access for free at: cites 49 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2013 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Conjugated Linoleic Acid Targets b2 Integrin Expression ToSuppress Monocyte Adhesion
Monica de Gaetano,1 Eugene Dempsey,1 Simone Marcone, William G. James, and
Orina Belton
Chronic recruitment of monocytes and their subsequent migration through the activated endothelium contribute to atheroscle-
rotic plaque development. Integrin-mediated leukocyte adhesion is central to this process. Conjugated linoleic acid (CLA) has the
unique property of inducing regression of pre-established murine atherosclerosis via modulation of monocyte/macrophage func-
tion. Understanding the mechanisms through which CLA mediates its atheroprotective effect may help to identify novel pathways
that limit or reverse atherosclerosis. In this study, we identified a novel mechanism through which CLA alters monocyte function.
We show that CLA inhibits human peripheral blood monocyte cell adhesion to activated endothelial cells via loss of CD18
expression, the b2 chain of LFA-1 and Mac-1 integrins. In addition, using a static-adhesion assay, we provide evidence that
CLA prevents monocytes from binding to ICAM-1 and subsequently reduces the capacity of these cells to polarize. CXCL12–
CXCR4 interactions induce a conformational change in b2 integrins, facilitating leukocyte adhesion. In this study, we demonstrate
that CLA inhibits CXCR4 expression, resulting in a failure of monocytes to directionally migrate toward CXCL12. Finally, using
intravital microscopy, we show that, during CLA-induced regression of pre-established atherosclerosis in ApoE2/2 mice, there is
reduced leukocyte adhesion and decreased CD18 expression on Gr1+/CD115+ proinflammatory monocytes. In summary, the data
presented describe a novel functional role for CLA in the regulation of monocyte adhesion, polarization, and migration. The
Journal of Immunology, 2013, 191: 4326–4336.
Atherosclerosis, the underlying cause of heart disease andstroke is a complex progressive disease with multiplegenetic and environmental contributions. In addition to
the role of lipids, atherosclerosis has many features of an in-flammatory disease, including infiltration of monocytes to the ves-sel wall and a proliferative response of smooth muscle cells, forexample. Early events in the pathogenesis of the disease involvedisturbed fluid shear stress, particularly at branch points orbifurcations, such as the aortic arch and iliac arteries, resultingin dysfunction of the endothelium on blood vessels (1–3) and,subsequently, increased vascular permeability to macromolecules,including cholesterol containing low-density lipoprotein (LDL).Subendothelial deposits of LDL become oxidized to produceminimally oxidized LDL, which induces local inflammation andsubsequent infiltration of immune cells. Although many cell types,including dendritic cells, T cells, endothelial cell, and smoothmuscle cells (4, 5), are documented to play a role in atherogenesis,the chronic recruitment of monocytes to the developing plaque
and their differentiation to macrophages are central elements inthe continual progression of the disease (6, 7).The recruitment of monocytes to inflammatory sites is a highly
orchestrated process in which integrins play a central role. Integrinsare a large family of multifunctional heterodimeric transmembraneproteins consisting of an a- and b-chain that mediate rolling andadhesion of leukocytes. Integrins on circulating leukocytes aretypically present in low-affinity states; however, a mechanisminitiated by chemokines, such as IL-8 and CXCL12 (8, 9), termed“inside out” signaling, induces their conformational change tointermediate or high-affinity states that results in increased bind-ing affinity for their respective ligands (10). The b2 integrinsLFA-1 and Mac-1 share a common b-chain (CD18), which is non-covalently linked with a unique a-chain: CD11a (aL) or CD11b(aM), respectively. Both b2 integrins bind ICAM-1 (11), whichis upregulated on endothelial cells in the presence of proin-flammatory cytokines, such as TNF-a. Sumagin et al. (12) re-cently demonstrated the differential roles of LFA-1 and Mac-1 onmonocyte crawling and showed that LFA-1 is used by monocytesto crawl on unactivated endothelial cells; however, during TNF-a–induced inflammation, Mac-1 becomes the more important integrinfor attachment and crawling.Most of the cellular and molecular mechanisms involved in
the development of atherosclerotic lesions have been identified;however, there has been no defined pathway that would explainhow this process could be reversed or even if such pathways exist.Conjugated linoleic acid (CLA) is a generic term denoting a groupof naturally occurring isomers of linoleic acid (18:2, n6), whichdiffer in the position and geometry of their double bonds. CLAhas been investigated as a potential prophylactic interventionfor multiple inflammatory diseases, including obesity, hyper-insulinemia, and hypertension (13, 14). Additionally, CLA wasidentified to have antiatherogenic effects in a number of animalmodels of atherosclerosis (15–17). The most potent immuno-modulatory CLA isomers are cis-9,trans-11 CLA and trans-10,
School of Biomolecular and Biomedical Science, Conway Institute, University Col-lege Dublin, Dublin 4, Ireland
1M.d.G. and E.D. contributed equally to this work.
Received for publication April 11, 2013. Accepted for publication August 6, 2013.
This work was funded by the Health Research Board, Dublin, Ireland.
Address correspondence and reprint requests to Dr. Orina Belton, School of Biomo-lecular and Biomedical Science, Conway Institute, University College Dublin, Bel-field, Dublin 4, Ireland. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: CD, 1% cholesterol chow diet; CLA, conjugatedlinoleic acid; FAK, focal adhesion kinase; HAEC, human aortic epithelial cell;HPBMC, human peripheral blood monocyte cell; IVM, intravital microscopy;LDL, low-density lipoprotein; MFI, median fluorescence intensity; ND, normal cho-lesterol chow diet; OA, oleic acid; PPAR, peroxisome proliferator–activated receptor;PSGl-1, P-selectin glycoprotein ligand-1; SFM, serum-free medium; TROG, trogli-tazone.
Copyright� 2013 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/13/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1300990
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

cis-12 CLA (18). The two isomers have differing physiologicalactivities; however, when used in combination they can elicitdistinct biological effects (19). We showed previously that dietaryadministration of a 1% CLA blend (80:20, cis-9,trans-11/trans-10,cis-12 CLA) induces regression of pre-established atherosclerosisin the ApoE2/2 mouse model, despite continuing a high choles-terol challenge (20). In addition, we showed that the monocyte/macrophage is the cellular target through which CLA mediates itsatheroprotective effect (21). Therefore, understanding the mech-anisms through which CLA mediates regression may help toidentify endogenous pathways or mechanisms that limit or reversethe disease.The exact mechanism through which CLA mediates its bio-
logical effects has not been fully elucidated, although regulation ofperoxisome proliferator–activated receptor (PPAR)g is proposedto be central to its mode of action (22–24). However, in ourprevious studies (21), we showed that CLA mediates the inhibitionof monocyte migration in both a PPARg-dependent and -inde-pendent manner. Importantly, although activation of PPARg inhib-its the development of atherosclerosis, it has no effect on regressionin vivo (25), which is in contrast to the regression seen with CLAsupplementation (20).In this study, we aimed to define the mechanisms through which
CLA alters monocyte function and confers an atheroprotectivephenotype. To this end, we examined the effect of the two mostabundant CLA isomers, cis-9,trans-11 CLA and trans-10,cis-12CLA, either individually or as an 80:20 ratio blend, on monocyteadhesion. We demonstrate that CLA inhibits human peripheralblood monocyte cell (HPBMC) adhesion to activated endothelialcells. We also show that this is mediated by inhibition of b2integrins via a PPARg-dependent mechanism. Additionally, wedemonstrate that CLA also inhibits the expression of CXCR4, a Gprotein–coupled receptor, leading to a failure of monocytes todirectionally migrate toward the chemokine CXCL12. Finally,using intravital microscopy (IVM), we show that CLA-supple-mented ApoE2/2 mice have a reduction in the attachment ofleukocytes in areas of vascular inflammation in vivo. The datapresented in this article describe a novel functional role for CLAin the regulation of monocyte adhesion, polarization, and migra-tion.
Materials and MethodsAnimals and diets
Homozygous ApoE-deficient mice (002052) (C57BL/6J-ApoEtm1Unc) werepurchased from The Jackson Laboratory (Bar Harbor, ME). All animalexperiments were conducted in conformity with institutional guidelinesand in compliance with international laws. An 80:20 cis-9,trans-11CLA:trans-10,cis-12 CLA blend was supplied by Stepan Lipid Nutrition(Wormerveer, Holland) and incorporated into 1% cholesterol chow diet(CD; Special Dietary Services, Essex, U.K.). ApoE2/2 mice were ran-domized at 40 d to receive CD for 12 wk or CD for 12 wk during which thefinal 4 wk was supplemented with 1% CLA blend. This diet protocol wasshown to promote regression of pre-established atherosclerotic lesions inthis model (20). ApoE2/2 mice maintained on a normal cholesterol chowdiet (ND) for the 12-wk period were also examined.
Isolation of HPBMCs
Whole blood from healthy volunteers was drawn into heparin-coatedvacutainers (BD - UK/Ireland). All volunteers were nonsmoking, aged25–30 y, and free from medication for $10 d. Platelet-rich plasma wasisolated by centrifugation (190 3 g for 15 min), diluted 1:3 with PBSbefore addition to Lymphoprep (AXIS-SHIELD PoC AS, Oslo, Norway),and centrifuged at 450 3 g for 30 min. Buffy coats were recovered usinga Pasteur pipette, washed twice with PBS, and resuspended in 10 mlserum-free medium (SFM) M-199 (Thermo Scientific), supplemented withL-glutamine (6.8 mM), antibiotics (100 U/ml penicillin, 100 mg/ml strep-tomycin), and 10 ng/ml polymyxin B sulfate (Sigma-Aldrich, Dublin,
Ireland). Monocytes were purified by plastic adherence in SFM for 2 h.Attached monocytes were washed and gently scraped from the surfacebefore use in subsequent assays.
HPBMC culture and treatments
Prior to specific assays, monocytes were incubated at 37˚C in SFM andtreated with 10 mM cis-9,trans-11:trans-10,cis-12 (80:20) CLA blend,oleic acid (OA; used as fatty acid control), the PPARg agonist troglitazone(TROG; 5 mM), or DMSO for 18 h. CLA and OA were purchased fromCayman Chemicals (Ann Arbor, MI); TROG was from Sigma-Aldrich.
Static-adhesion assay of monocytes to immobilized humanaortic endothelial cells
Human aortic endothelial cells HAECs (Cascade Biologics, Invitrogen,Carlsbad, CA) were grown in endothelial cell culturemediaMV plus growthsupplements from Promo Cell (Heidelberg, Germany), supplemented with100 U/ml penicillin and 100 mg/ml streptomycin. For experimental anal-ysis, HAECs were incubated in growth media with 1% FCS, 0.4% endo-thelial cell growth supplements, 90 mg/ml heparin, 100 U/ml penicillin,and 100 mg/ml streptomycin. Cells were grown in a humidified atmosphereof 5% CO2 at 37˚C.
For static-adhesion assays, 4 3 104 HAECs were seeded in a 96-wellplate for 24 h, followed by stimulation with TNF-a (10 ng/ml) for 6 h, andwashing in media. Peripheral blood monocytes were labeled with fluo-rescein using 1 mg/ml BCEFC-AM (Calbiochem), as per the manu-facturer’s instructions. Subsequently 1 3 105 labeled monocytes wereadded to activated HAECs and coincubated for 30 min at 37˚C. Cells werewashed three times with media, and fluorescence was measured in aSpectramax M2 (Molecular Devices, Sunnyvale, CA) plate reader with485-nm excitation and 585-nm emission wavelength, before and afterwashing. A fluorescent intensity unit/cell was derived by dividing the totalfluorescent signal before washing by the number of cells seeded. The finalnumber of adherent cells was obtained by multiplying this unit by the totalfluorescent signal after washing.
RNA extraction and gene-expression analysis
For gene-expression experiments, monocytes were washed twice with ice-cold PBS prior to addition of 200 ml RLT buffer (QIAGEN, Manchester,U.K.). Total RNA was isolated from cell lysates using the RNeasy kit(QIAGEN), as per the manufacturer’s instructions. Reverse transcriptionwas carried out on 1 mg total RNA using Superscript II Reverse Tran-scriptase (Invitrogen), according to the manufacturer’s instructions. Rela-tive gene expression quantification by real-time PCR was performed on anABI Prism 7700 Sequence Detection System (Applied Biosystems, War-rington, U.K.). b1 and b2 integrins, PSGL-1, and CXCR4 expression wasexamined using specific TaqMan assays (Applied Biosystems) and nor-malized to 18S rRNA.
Western blot and protein-expression analysis
Whole-cell lysates were prepared from monocytes in lysis buffer (0.2%Nonidet P-40, 150 mM NaCl, 50 mM Tris-HCl [pH 7.2]) and then clearedby centrifugation (10,000 3 g for 10 min at 4˚C). Lysates were separatedusing 4–20% SDS-polyacrylamide gel and transferred to nitrocellulose(Hybond-C Extra; Amersham Biosciences, Buckinghamshire, U.K.). Mem-branes were blocked with 3% BSA in TBST (20 mm Tris [pH 7.6],150 mm NaCl, 0.05% Tween 20) at room temperature prior to overnightincubation with primary Abs at 4˚C with gentle shaking. Detection wasperformed using an HRP-conjugated secondary Ab (1 h at room temper-ature) and ECL detection system (Amersham Biosciences). mAb againstb-actin (1:1000) and polyclonal Abs against CD11a (1:500) and CD11b(1:500) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA).mAb against CD18 (RE72B; 1:3000) was a kind gift from Prof. CarlGahmberg’s laboratories (Division of Biochemistry, Department of Bio-sciences, University of Helsinki, Helsinki, Finland).
Flow cytometry analysis
To block unspecific binding, monocytes were incubated in 1% BSA in PBSfor 15 min prior to Ab staining. FITC-CD14 (clone UCHM-1; Sigma-Aldrich), PE–LFA-1 (BioLegend, London, U.K.), allophycocyanin–Mac-1(BioLegend), and AF700-CD18 (EXBIO) Abs were used to label mono-cytes. All Ab incubations were for 30 min at room temperature. Cellswere washed and fixed in 2% paraformaldehyde. Forward and side scattergates were established to exclude cell debris from the analysis. The meanfluorescence intensity of 1 3 105 cells was analyzed in each sample.Autofluorescence signals generated by unlabeled cells were used as neg-
The Journal of Immunology 4327
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

ative controls in each experiment. Flow cytometric analysis was performedon an Accuri C6 instrument and analyzed with CFlow Software (Accuri,Ann Arbor, MI).
Static-adhesion assay of monocytes to immobilized ICAM-1
Ninety-six–well glass-bottom imaging plates (Zell-Kontakt, Norten-Hardenberg, Germany) were coated overnight with recombinant humanICAM-1 protein (5 mg/ml; R&D Systems), washed with Ca2+ and Mg2+–free PBS (Life Technologies, Invitrogen), and blocked with 2% BSA(Sigma-Aldrich) for 2 h at 37˚C. Following washes in Ca2+ and Mg2+–freePBS, 1 3 105 fluorescein-labeled (using 1 mg/ml BCEFC-AM; Calbio-chem) HPBMCs were treated with EGTA (1 mM) and MgCl2 (5 mM) toinduce the active conformation of surface integrins. Cells were rapidlyadded to ICAM-1–coated wells and incubated for 30 min at 37˚C. Poly-L-lysine (0.01%; Sigma-Aldrich) coated wells were used as a positive con-trol. Changes in adhesion were calculated as previously described.
Immunocytochemistry
To visualize adhesion and polarization of monocytes, 1 3 105 cells werecultured and treated in a 96-well imaging glass plate (Zell-Kontakt). Ad-herent monocytes (activated with either EGTA/MgCl2 or CXCL12) werefixed in 4% formaldehyde, permeabilized with Triton X-100 (Sigma-
Aldrich), and labeled with Alexa Fluor 568–phalloidin (Invitrogen) to stainF-actin and with DAPI (Sigma-Aldrich) to visualize nucleic acids. Imagesof stained monocytes were captured using a Zeiss AxioVert 200M InvertedFluorescent Microscope, and images were captured using an AxioCam HR(Zeiss) camera - CCD Basic resolution; 103(air)/0.25 and 633(oil)/1.40lenses were used at room temperature. When needed, immersion oil, typeDF (Cargille Laboratories) was used.
CXCL12-induced crawling assay
For transmission light microscopy acquisition of live cells, freshly isolatedperipheral blood monocytes were treated as described above and incubatedat 37˚C for 18 h in a 96-well uncoated bare plastic plate (Greiner Bio-One,Cruinn Diagnostics, Dublin, Ireland). CXCL12 was added to one side ofthe well using either 1 ng/ml for THP-1 (in 1 ml) or 100 ng/ml for primaryhuman monocytes (in 10 ml) to induce directional crawling. Immediatelyafter addition of CXCL12, images were captured from the opposite side ofthe well at regular intervals over a period of 5–10 min, using a NIKON-TMS microscope, 203(air)/0.4 lens, at room temperature. Pictures werecaptured using a 3-CCD Color Video Camera (JVC), KY-F55B. MatrixIntellicam acquisition software was used. Image sequences were analyzed,with x and y coordinates being assigned to individual cells; cell tracks weresubsequently visualized using rosette plots.
FIGURE 1. CLA inhibits HPBMC
adhesion to activated endothelial
cells. (A) HAECs were activated fol-
lowing stimulation with 10 ng/ml
TNF-a for 6 h. HPBMCswere treated
with 10 mM cis-9,trans-11 CLA (c9,
t11), trans-10,cis-12 CLA (t10,c12),
CLA blend, OA, 5 mM TROG, or
DMSO vehicle control (VC) for 18 h.
Data are mean 6 SEM. (B) MTT
assay showing viability of HPBMCs
following treatment with a dose range
of c9,t11 and t10,c12 CLA. A total of
10 mM of each isomer was used for
all subsequent experimental analyses.
*p , 0.05, **p , 0.01 versus TNF-
a–activated DMSO vehicle, $$$p ,0.001 versus resting DMSO control.
4328 CONJUGATED LINOLEIC ACID INHIBITS MONOCYTE ADHESION
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

Intravital microscopy
Leukocyte recruitment within the femoral artery was imaged by IVM.Animals from each diet regimen were randomly allocated into two treat-ment groups, a control saline injection or an acute dose of TNF-a (1 mg), aspreviously described (26). Briefly, 200 ml saline or TNF-a (1 mg in 200 mlsaline) was administered i.p. 4 h prior to IVM imaging. Animals wereanesthetized by i.p. injection of a mixture of xylazine (10 mg/kg) andketamine hydrochloride (150 mg/kg) (C&M Vetlink, Limerick, Ireland). Acatheter was inserted into the right jugular vein to administer anestheticand fluorescent dye. To visualize leukocytes within the vasculature, 50 mlRhodamine 6G (Sigma-Aldrich) (0.05% in saline) was administered i.v.prior to surgical exposure of the femoral artery (27). Animals were placedin a supine position on a temperature-controlled heating mat to maintaincore temperature at 37˚C.
Skin was incised along the interior surface of the thigh, and the femoralartery on each leg was exposed in turn for imaging. PBS superfusion wasapplied to the tissue. Fluorescent emission from Rhodamine 6G was vi-sualized by epifluorescent microscopy via a 546/12-nm excitation filter anda 575–640-nm emission filter (Filter#20; Carl Zeiss U.K.). The femoralartery was imaged using a Carl Zeiss Axioskop 2 Fs-mot intravital mi-croscope with a 203 water-immersion objective lens (203/0.5 NA; CarlZeiss U.K.). A Baxall CDX9742 monochrome camera (Carl Zeiss U.K.)collected the emitted light, which was displayed on a video monitor andrecorded on a hard-drive recorder (JVC SR-DVM70) for playback analy-sis. The femoral arteries (150–190 mm in diameter) of both legs wereimaged in the direction from the knee toward the body, with ∼10–12randomly selected fields-of-view recorded per artery. The CCD camerawas rotated to position the artery in a horizontal orientation on the monitorto produce a standardized vessel length of 278 mm for each recording.Parameters analyzed in each recording included the leukocyte rolling flux(rolling cells/min), leukocyte rolling velocity (mm/sec), and the numberof leukocytes adherent to the artery endothelium (adherent cells/vessellength). Leukocytes were deemed to be stably adhered if they remainedstationary for $30 s. Because of the relatively large diameter of thefemoral artery, depth of imaging was restricted to the outer endothelialsurface.
MTT assay
For this colorimetric assay, the following dose responsewas applied for eachCLA isomer (cis-9,trans-11 and trans-10,cis-12): 0, 1, 10, 25, or 50 mM.After 18 h of treatment, 5 mg/ml MTT (Sigma Aldrich) solution was addedand incubated at 37˚C for 3.5 h. After cell lysing with DMSO, absorbancewas read spectrophotometrically at 570 nm.
Statistical analysis
Results are expressed as mean 6 SEM or fold change relative to vehiclecontrol. All data are from a minimum of three independent experiments(n = 3). Statistical comparisons between controls and treated groups weremade using the unpaired Student t test, assuming unequal variance. Ap value ,0.05 was considered significant.
ResultsCLA inhibits human monocyte adhesion to endothelial cells
We demonstrated previously that dietary administration of 1%CLA blend (80:20, cis-9,trans-11:trans-10,cis-12) induces regres-sion of pre-established atherosclerosis in the ApoE2/2 mouse (20)and that modulation of monocyte function plays a central rolein CLA-induced regression (21, 28). To examine this further, weinvestigated the effect of CLA on monocyte adhesion to activatedendothelial cells. MTT assays were used to determine the opti-mum concentration of CLA isomers for all assays; 10 mM of cis-9,trans-11 CLA and trans-10,cis-12 CLA was the lowest dose ofeach isomer that did not affect cell viability (Fig. 1) but is knownto have biological effects (21); hence, this concentration was usedfor all subsequent experimental analyses. Activation of HAECswith TNF-a induced a significant increase in the number of ad-herent monocytes (Fig. 1). Monocytes that were pretreated withcis-9,trans-11 CLA and 80:20 cis-9,trans-11:trans-10,cis-12 CLA
FIGURE 2. Effect of CLA on b1, b2 integrins and PSGL-1 mRNA expression in HPBMCs. Peripheral blood monocytes were treated as previously
described and analyzed for mRNA expression of the common b subunit (CD18) of the b2 integrin receptor (A), LFA-1 a-chain (CD11a) (B), Mac-1 a-chain
(CD11b) (C), b-chain (CD29) (D) and a-chain (CD49d) of the a4b1 integrin (E), and PSGL-1 (F). cis-9,trans-11 CLA and the CLA blend inhibit CD18
expression; trans-10,cis-12 CLA has no effect on CD18 but reduces PSGL-1 expression; and TROG inhibits b1, b2, and PSGL-1. Data are mean 6 SEM.
*p , 0.05, **p , 0.01, ***p , 0.001 versus vehicle control (VC).
The Journal of Immunology 4329
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
blend had an attenuated ability to bind to activated endothelium,displaying a 63% (p , 0.01) and 68% (p , 0.01) decrease in thenumber of adherent cells, respectively. Treatment with trans-10,cis-12 CLA also had an inhibitory effect on adhesion, displayinga modest, but significant, 38% decrease in adhesion (p , 0.05). Incontrast, OAwas used as a fatty acid control and had no significanteffect on monocyte adhesion. PPARg is a known target of CLA(24); TROG, a PPARg agonist, mimicked the CLA treatments byinhibiting adhesion by 58% (p , 0.05), suggesting that CLA in-hibition of monocyte adhesion may be mediated via a PPARg-dependent mechanism.
CLA inhibits the expression of adhesion molecules inperipheral blood monocytes
To identify a possible mechanism by which CLA inhibits monocyteadhesion to activated endothelium, we analyzed the mRNA ex-pression of a number of integrin and selectin receptors, includingthe b1 integrin, VLA-4 (a4b1), b2 integrins (LFA-1 [aLb2] andMac-1 [aMb2], and P-selectin glycoprotein ligand-1 (PSGL-1),in CLA-treated human primary monocytes. Primary monocytestreated with either cis-9,trans-11 CLA or CLA blend experienceda significant decrease in the expression of CD18 by 1.92-fold(p , 0.05) and 1.53-fold (p , 0.001), respectively (Fig. 2A),whereas neither treatment significantly influenced the expression
of CD11a, CD11b, CD29, or CD49d (Fig. 2B–E). Although thetrans-10,cis-12 CLA isomer reduced the expression of CD18 by1.47-fold, this was not significant. Indeed, with the exception ofPSGL-1, which showed a significant 1.37-fold decrease (p ,0.05), trans-10,cis-12 CLA had no significant effect on any of theother mRNA transcripts examined. In contrast to CLA, the fattyacid control OA had no effect on the expression on b2 integrinsubunits and, in fact, increased the expression of CD49d andPSGL-1 by 1.3-fold (p , 0.05) and 1.5-fold (p , 0.05), respec-tively. TROG had a modest effect on most of the transcripts ex-amined; however, it was a potent suppressor of CD18, resulting ina 4.35-fold (p , 0.001) decrease in expression.UsingWestern blot analysis, we confirmed that the CLA-induced
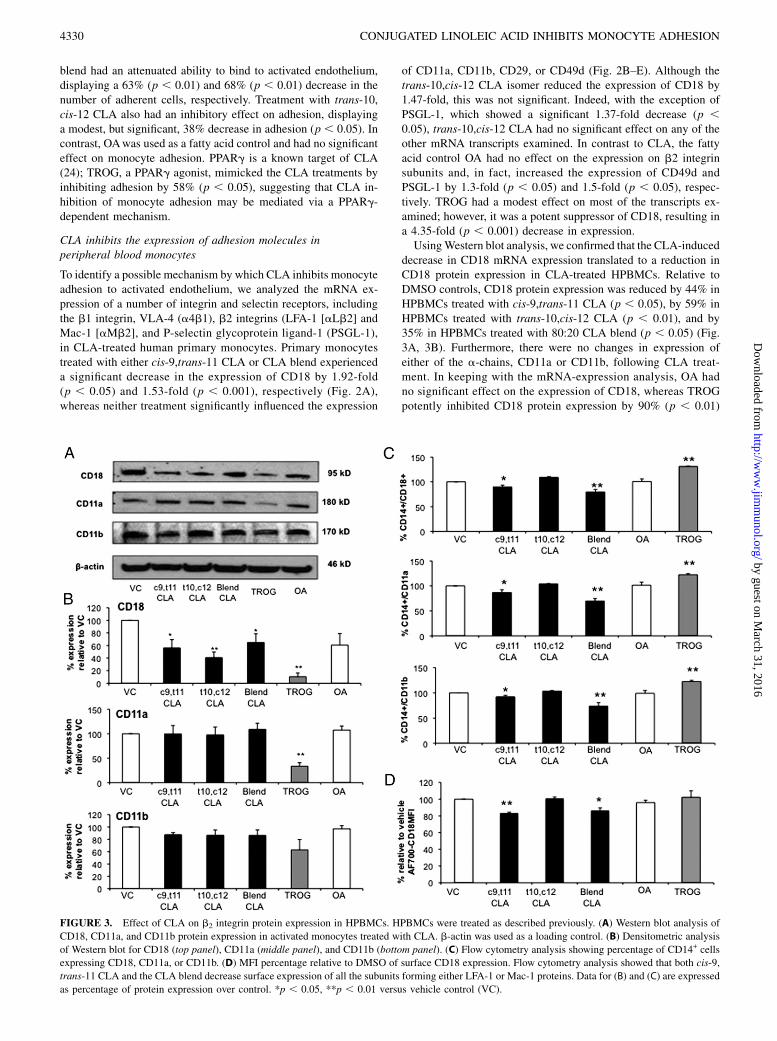
decrease in CD18 mRNA expression translated to a reduction inCD18 protein expression in CLA-treated HPBMCs. Relative toDMSO controls, CD18 protein expression was reduced by 44% inHPBMCs treated with cis-9,trans-11 CLA (p , 0.05), by 59% inHPBMCs treated with trans-10,cis-12 CLA (p , 0.01), and by35% in HPBMCs treated with 80:20 CLA blend (p , 0.05) (Fig.3A, 3B). Furthermore, there were no changes in expression ofeither of the a-chains, CD11a or CD11b, following CLA treat-ment. In keeping with the mRNA-expression analysis, OA hadno significant effect on the expression of CD18, whereas TROGpotently inhibited CD18 protein expression by 90% (p , 0.01)
FIGURE 3. Effect of CLA on b2 integrin protein expression in HPBMCs. HPBMCs were treated as described previously. (A) Western blot analysis of
CD18, CD11a, and CD11b protein expression in activated monocytes treated with CLA. b-actin was used as a loading control. (B) Densitometric analysis
of Western blot for CD18 (top panel), CD11a (middle panel), and CD11b (bottom panel). (C) Flow cytometry analysis showing percentage of CD14+ cells
expressing CD18, CD11a, or CD11b. (D) MFI percentage relative to DMSO of surface CD18 expression. Flow cytometry analysis showed that both cis-9,
trans-11 CLA and the CLA blend decrease surface expression of all the subunits forming either LFA-1 or Mac-1 proteins. Data for (B) and (C) are expressed
as percentage of protein expression over control. *p , 0.05, **p , 0.01 versus vehicle control (VC).
4330 CONJUGATED LINOLEIC ACID INHIBITS MONOCYTE ADHESION
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
relative to controls. To confirm a PPARg-dependent mechanismfor the CLA-mediated regulation of CD18 expression, monocyteswere treated with 10 mM of the PPARg antagonist GW9662 for2 h prior to the addition of CLA for 18 h (28). In the presence of thePPARg antagonist, the effect of CLA on CD18 gene expression islost, confirming a PPARg-dependent mechanism for the regulationof CD18 gene expression (Supplemental Fig. 1).Because integrins are integral membrane proteins, in the next
series of experiments we used flow cytometry to identify whetherthere was a loss of cell surface integrin expression in CLA-treatedmonocytes. By gating on CD14+ monocytes, we assessed the ex-pression of the b2 integrin subunits. Because of the potential forinterindividual variability between three human donors, we cal-culated both the percentage median fluorescence intensity (MFI)relative to DMSO and the percentage of monocytes expressing theb2 integrin subunits. Compared with controls, cis-9,trans-11 CLAand CLA blend decreased the number of monocytes expressingthe b-chain by 10% (p , 0.05) and 20% (p , 0.01), respectively(Fig. 3C). This was confirmed by analysis of the MFI percentagerelative to DMSO control of cellular AF700-CD18 (Fig. 3D).Additionally, the cis-9,trans-11 isomer decreased expression ofboth CD11a (by 13%, p , 0.05) and CD11b (by 8%, p , 0.05).Furthermore, the 80:20 blend followed the same trend as did cis-9,trans-11 CLA, but to a greater degree; CD11a-expressing cellswere decreased by 30% (p , 0.01), whereas CD11b-expressingcells were decreased by 27% (p , 0.01). Neither the trans-10,cis-12 isomer nor OA had any effect on CD18, CD11a, or CD11bexpression. In contrast to the mRNA-quantification and Westernblot data, TROG induced a significant increase in the number ofcells expressing b2 integrin subunits on their cell surface (CD18:36%, p , 0.01; CD11a: 29%, p , 0.01; CD11b: 27%, p , 0.01).
Nonetheless, when these data are taken together, they suggestthat CLA-mediated inhibition of monocyte adhesion to HAECs isdue to a loss of expression of CD18, the b2 chain of both LFA-1 orMac-1 integrins, and is potentially mediated via a PPARg-inde-pendent mechanism.
CLA-treated peripheral blood monocytes fail to adhere toICAM-1 or develop a migratory phenotype
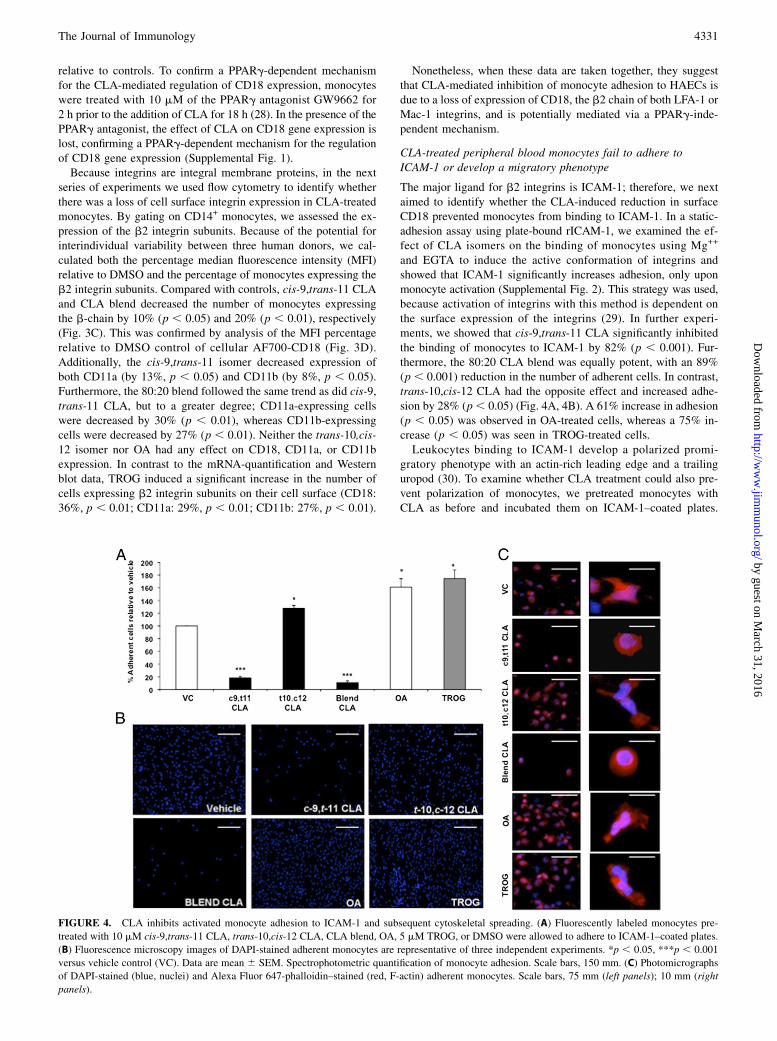
The major ligand for b2 integrins is ICAM-1; therefore, we nextaimed to identify whether the CLA-induced reduction in surfaceCD18 prevented monocytes from binding to ICAM-1. In a static-adhesion assay using plate-bound rICAM-1, we examined the ef-fect of CLA isomers on the binding of monocytes using Mg++
and EGTA to induce the active conformation of integrins andshowed that ICAM-1 significantly increases adhesion, only uponmonocyte activation (Supplemental Fig. 2). This strategy was used,because activation of integrins with this method is dependent onthe surface expression of the integrins (29). In further experi-ments, we showed that cis-9,trans-11 CLA significantly inhibitedthe binding of monocytes to ICAM-1 by 82% (p , 0.001). Fur-thermore, the 80:20 CLA blend was equally potent, with an 89%(p , 0.001) reduction in the number of adherent cells. In contrast,trans-10,cis-12 CLA had the opposite effect and increased adhe-sion by 28% (p, 0.05) (Fig. 4A, 4B). A 61% increase in adhesion(p , 0.05) was observed in OA-treated cells, whereas a 75% in-crease (p , 0.05) was seen in TROG-treated cells.Leukocytes binding to ICAM-1 develop a polarized promi-
gratory phenotype with an actin-rich leading edge and a trailinguropod (30). To examine whether CLA treatment could also pre-vent polarization of monocytes, we pretreated monocytes withCLA as before and incubated them on ICAM-1–coated plates.
FIGURE 4. CLA inhibits activated monocyte adhesion to ICAM-1 and subsequent cytoskeletal spreading. (A) Fluorescently labeled monocytes pre-
treated with 10 mM cis-9,trans-11 CLA, trans-10,cis-12 CLA, CLA blend, OA, 5 mM TROG, or DMSO were allowed to adhere to ICAM-1–coated plates.
(B) Fluorescence microscopy images of DAPI-stained adherent monocytes are representative of three independent experiments. *p , 0.05, ***p , 0.001
versus vehicle control (VC). Data are mean 6 SEM. Spectrophotometric quantification of monocyte adhesion. Scale bars, 150 mm. (C) Photomicrographs
of DAPI-stained (blue, nuclei) and Alexa Fluor 647-phalloidin–stained (red, F-actin) adherent monocytes. Scale bars, 75 mm (left panels); 10 mm (right
panels).
The Journal of Immunology 4331
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

Cells were subsequently stained for F-actin and analyzed. Mono-cytes treated with either cis-9,trans-11 CLA or the CLA blend
maintained a rounded phenotype, whereas all other treatment con-ditions allowed the monocytes to polarize (Fig. 4C). Interestingly,
FIGURE 5. CLA prevents ICAM-1–induced polarization on CXCL12-activated HPBMCs. (A) Following activation of HPBMCs for 1, 5, or 10 min with
100 ng/ml CXCL12, cells were fixed and stained. Fluorescent microscopy images of monocytes labeled with DAPI (blue) and Alexa Fluor 647–phalloidin
(red). Images are representative of three independent experiments. Scale bars, 10 mm. (B) RT-PCR analysis of mRNA expression of CXCR4 in HPBMCs.
(C) Cell tracking of CXCL12-induced crawling in THP-1 monocytes. Plots show individual tracks from 15 cells normalized to their starting position. cis-9,
trans-11 CLA–treated, CLA blend–treated, and unstimulated vehicle control (VC)-treated cells do not spread on ICAM-1–coated surface. Data are
expressed as fold change expression over control. *p , 0.05, **p , 0.01, versus vehicle.
4332 CONJUGATED LINOLEIC ACID INHIBITS MONOCYTE ADHESION
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

cells treated with TROG also maintained their ability to polarize,which is consistent with the results from the static ICAM-1–bindingassay and the flow cytometry data. Taken together, these resultsdemonstrate that CLA prevents monocyte binding to ICAM-1 andreduces the capacity of these cells to polarize via a PPARg-inde-pendent mechanism.
CLA reduces chemokine sensitivity in primary monocytes
To gain further insights into the effect of CLA on inside out ac-tivation of integrins, we examined the effect of CXCL12, whichbinds to the G protein–coupled receptor CXCR4, on CLA-treatedmonocytes. The signaling pathways initiated by CXCL12 andCXCR4 interactions induce a conformational change in b2 integ-rins, facilitating adhesion to ICAM-1 and subsequent polari-zation (28). To examine polarization, we pretreated human pri-mary monocytes with CLA and subsequently exposed them to100 ng/ml CXCL12. Shape change was monitored using F-actinstaining. As with the previous experiments, cells treated withcis-9,trans-11 CLA or the CLA blend retained a rounded pheno-type compared with all other treatments that induced polarization(Fig. 5A). Furthermore, we examined the mRNA expression of theCXCL12 receptor CXCR4. Both cis-9,trans-11 CLA and the CLAblend significantly inhibited CXCR4 expression by 1.88-fold (p .0.05) and 1.49-fold (p . 0.01), respectively (Fig. 5B). TROG hadno effect on its expression, indicating a likely PPARg-independentmechanism through which CLA regulates CXCR4.We extended these studies to establish whether CLA-treated
monocytes failed to directionally migrate toward CXCL12 dueto decreased CXCR4 expression. Using the THP-1 monocyte cellline, we tracked the movement of live cells toward 1 ng/ml CXCL12over a 5-min period (Supplemental Fig. 3). We extended this toinvestigate the effect in HPBMCs. Plotting individual cell tracksover time revealed that cis-9,trans-11 CLA and the CLA blendinhibited both the distance traveled and the general directionalmovement toward CXCL12. These data demonstrate that cis-9,trans-11 CLA and the atheroprotective CLA blend reduce mono-cyte crawling and impair the ability of monocytes to migrate to-ward CXCL12 (Fig. 5C).
CLA-fed mice display reduced leukocyte adhesion to activatedendothelium
To further elucidate whether CLA-mediated inhibition of mono-cyte adhesion and crawling plays a functional role in atheropro-tection in vivo, we investigated the effect of CLA on monocytefunction in the ApoE2/2 model. We established a model of CLA-induced regression of pre-established atherosclerosis in vivo (31).ApoE2/2 mice were randomized at 40 d of age to receive eitherND (n = 12) or CD (n = 12) for 12 wk or to receive CD for 12 wk,with 1% CLA blend supplementation for the last 4 wk (n = 12).Leukocyte recruitment, including leukocyte rolling flux, rollingvelocity, and adhesion, was quantified in the femoral artery byIVM. At the time of IVM imaging, ApoE2/2 mice from each di-etary regimen received an injection of TNF-a (1 mg, 4 h) to in-duce endothelial cell activation (n = 6) or an injection of saline(control; n = 6).The basal level of leukocyte adhesion (saline treated) measured
in the femoral artery was increased significantly (p = 0.011) inmice fed CD compared with ND. A significant decrease in basalleukocyte adhesion was observed (p, 0.05) in mice fed the CLA-supplemented diet compared with mice fed CD. As expected uponactivation of adhesion with TNF-a, there was increased leukocyteadhesion across all animal groups on all three dietary regimens.Although there was no difference between mice fed ND or CDfollowing administration of TNF-a, it is important to note that
mice on a CLA-supplemented diet displayed a significantly (p ,0.05) reduced level of adhesion under activated conditions com-
FIGURE 6. CLA inhibits leukocyte adhesion in regression of pre-
established atherosclerosis. IVM analysis of leukocyte recruitment was
performed in the femoral artery of ApoE2/2 mice. ApoE2/2 mice were fed
ND (n = 12) or CD for 12 wk (n = 12) or CD for 12 wk, with the final 4 wk
supplemented with 1% CLA blend (CD + CLA) (n = 12). Leukocyte ad-
hesion (A) and leukocyte velocity (B) were quantified following injection
of TNF-a (1 mg, 4 h), to induce endothelial cell activation, or saline
control. CLA significantly inhibits leukocyte adhesion under basal and
activated conditions compared with cholesterol-fed mice. CD-fed mice
displayed decreased basal leukocyte rolling velocity compared with those
fed ND. Mice supplemented with CLA had significantly faster rolling
velocities compared with mice fed CD, making them more comparable
with mice fed ND. (C) CD18 expression on monocyte subsets was deter-
mined by flow cytometry. Monocytes were separated based on their Gr1
and CD115 status, and the expression of CD18 was quantified. Circulating
proinflammatory Gr1+/CD115+ monocytes in mice fed CD had a higher
percentage of CD18 expression compared with mice fed ND. CLA sup-
plementation significantly downregulated CD18 expression on the proin-
flammatory monocyte subset. **p , 0.01 versus CD.
The Journal of Immunology 4333
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

pared with mice fed CD, indicating that there was an impairmentin their ability to bind to activated endothelium (Fig. 6A). In thenext series of experiments, we measured the rolling velocities ofleukocytes in the femoral artery both in the presence and absenceof TNF-a. In general, leukocytes from mice on CD displayeda significantly decreased basal rolling velocity compared withthose fed ND (p , 0.01). Moreover, mice supplemented withCLA had significantly faster rolling velocities compared with thosefed CD, making them more comparable to mice fed ND. However,treatment with TNF-a blunted the effect of CLA on leukocyterolling velocity. This is not surprising, because rolling velocity ismore dependent on expression of endothelial selectins and theircorresponding ligands, rather than integrin expression, which, asidentified through our in vitro experiments, is regulated by CLA.Finally, CLA had no effect on rolling leukocyte flux, suggestingthat its effect is not due to a decreased number of leukocytesrecruited. These data provide evidence that CLA inhibits leuko-cyte adhesion to the endothelium in vivo during regression ofatherosclerosis.
CLA preferentially downregulates CD18 expression onGr1+/CD115+ monocytes
Having shown that mice on a CLA-supplemented diet had impairedleukocyte adhesion compared with those fed CD, we aimed toidentify whether there were changes in CD18 expression onmonocyte subsets. Leukocytes were isolated from mice on the CDor CLA-supplemented diet and phenotyped using flow cytometry.Gr1+/CD115+ monocytes are described as inflammatory mono-cytes, whereas Gr12/CD115+ monocytes are described as patrol-ling monocytes (32). Monocytes were separated based on theirGr1 and CD115 status, and the expression of CD18 was quanti-fied. Circulating Gr1+/CD115+ monocytes in mice fed CD dis-played a higher percentage of CD18 expression (37.7%) compared
with mice fed ND (21.4%), although this difference was not sig-nificant. However, the percentage of CD18 expression on thismonocyte subset in CLA-supplemented mice was significantlydownregulated (4.2%, p = 0.012) compared with those fed CD (Fig.6C). Interestingly, CD18 expression on Gr12/CD115+ monocyteswas not significantly different among the groups fed ND (42.8%),CD (41.1%), or CLA-supplemented diet (38.2%). Thus, CLA spe-cifically targets CD18 expression on inflammatory Gr1+/CD115+
monocytes in vivo. Together, these data show that CLA mediatesan effect on firm adhesion, cell polarization, and chemotactic mi-gration of HPBMCs. (Fig. 7).
DiscussionCLA has the unique property of inducing regression of pre-established atherosclerosis in vivo (20, 31). The predominantbioactive CLA forms are the cis-9,trans-11 and trans-10,cis-12isomers, and a 80:20 ratio blend approximates their relative levelsduring dietary intake (33). Our previous studies showed that a80:20 cis-9,trans-11:trans-10,cis-12 CLA blend induces regres-sion of pre-established atherosclerosis in the ApoE2/2 murinemodel, despite continued high cholesterol challenge. Recently, weshowed that the monocyte/macrophage cell is the cellular targetthrough which CLA mediates this profound effect (31). Un-derstanding the mechanism(s) through which CLA mediatesmonocyte function in the context of regression may facilitate theidentification of novel mechanisms or pathways that limit pre-established disease.In this study, we demonstrated that cis-9,trans-11 CLA and the
atheroprotective CLA blend, but not trans-10,cis-12 CLA, inhibitsthe adhesion of monocytes to activated endothelial cells throughthe regulation of CD18 expression. For our experimental analysiswe used 10 mM of CLA isomers in accordance with our previouslypublished data and based on the results from the MTT assays
FIGURE 7. Effects of CLA on firm adhesion, cell polarization, and chemotactic migration of HPBMCs. CLA isomers and the 80:20 blend (cis-9,trans-
11:trans-10,cis-12), acting via a PPARg-dependent mechanism, decrease the expression of CD18 (b-chain) by b2 integrins, decreasing complex formation
between a and b subunits in the Golgi apparatus and reducing the number of integrins expressed on the external surface. As a consequence, fewer LFA-1
and Mac-1 molecules adhere to endothelial ICAM-1. In addition, CLA acts via a PPARg-independent manner on the adhesive/migratory CXCR4/CXCL12
pathway, significantly inhibiting CXCR4 expression and subsequently decreasing the interaction with CXCL12 cytokine, leading to a minor initiation of an
“inside out” pathway. As result, partially, but incompletely, activated b2 integrins influence the interaction between CD18 of both LFA-1 and Mac-1, with
actin filaments necessary for cell movement. In this way, cytoskeletal rearrangement of the monocyte is also inhibited, preventing monocyte adhesion,
polarization, and, consequently, chemokine-targeted migration.
4334 CONJUGATED LINOLEIC ACID INHIBITS MONOCYTE ADHESION
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

described above. Indeed, to the best of our knowledge, 10 mM isthe lowest concentration of CLA isomers used in experimentalanalysis of HPBMCs. For example, Platt and El-Sohemy (34)treated primary human CD14+ monocytes with 50 mM of CLAisomers, whereas other investigators used 30 mM of those twoCLA isomers to treat both human primary and THP-1 monocytes(35). The differential effects of the two most abundant CLA iso-mers on the inhibition of monocyte adhesion to activated endo-thelial cells are consistent with previous studies (36) that identifiedthe divergent effects of the two CLA isomers in atherosclerosis. Arecent study (37) demonstrated that cis-9,trans-11 CLA decreasesMac-1 expression on macrophages through the suppression ofCD11b. However, that study did not investigate potential changesin CD18 expression. The biosynthesis of b2 integrins occurs withthe assembly of the a2 and b-chains in the endoplasmic retic-ulum, after which they are transported to the cell surface via theGolgi apparatus. In the absence of a pairing b-chain, the a-chainis not shuttled to the cell membrane (38). Our data are consistentwith this paradigm of integrin assembly. We showed that althoughthe mRNA and protein expression of both a-chains are unaf-fected, both are reduced on the cell surface as a consequenceof CD18 inhibition (Fig. 7). It should be noted that HPBMCSwere isolated using adhesion onto plastic, rather than a negative-selection method. Although our method ensures a pure popula-tion of monocytes, the method used may induce a state of pre-activation.CLAwas reported to mediate its effect, in part, via activation of
PPARg. However, our previous work showed that CLA inhibitsmonocyte migration via a PPARg-dependent and -independentmechanism (21). In the current study, the PPARg agonist TROGreduced CD18 mRNA and total protein expression. We confirmedthat CLA mediates its effect on CD18 expression via a PPARg-dependent mechanism because pretreatment with the PPARg an-tagonist GW9662 abolished the effect of CLA on CD18 mRNAexpression. Paradoxically, flow cytometry analysis showed thatTROG increased the levels of surface CD18 expression. Althoughthis may appear contradictory, our data are in keeping with pre-vious studies. For example, Hirasawa et al. (39) demonstrated thatTROG could reduce the binding of eosinophils to ICAM-1, but itdid not downregulate the surface expression of CD11a, CD11b, orCD18, indicating that TROG may influence the activity of b2 viaan alternative mechanism.The binding of CXCL12 to the G protein receptor CXCR4 is an
important mediator in the initial stages of leukocyte adhesion andextravasation to sites of inflammation (40). CXCL12 is present athigh concentrations in atherosclerotic lesions where it is releasedby smooth muscle cells, endothelial cells, and macrophages (41).Engagement of CXCL12 with CXCR4 triggers a signaling path-way that induces integrins to switch from a low- to a high-affinitystate. Hence, the activities of CXCR4 and b2 integrins are inex-tricably linked in the process of trafficking monocytes to sites ofinflammation. We show that CLA inhibits CXCR4 expression andresults in a failure of monocytes to migrate toward CXCL12. Fromour analysis, CLA-regulated expression of CXCR4 is not underthe direct control of PPARg, because the agonist TROG had noeffect on its expression, highlighting the complexity of CLA sig-naling. However, although the data for CLA are consistent, ininstances where CLA decreases adhesion to activated endothe-lial cells, reduces CXCR4 expression, and hence, chemotaxis toCXCL12 (Fig. 5), the data for TROG are conflicting. Although thePPARg agonist TROG had no effect on CXCR4 expression, itinhibited chemotaxis to CXCL12. This may be due to the fact thatTROG was shown to inhibit the phosphorylation of focal adhesionkinase (FAK), which is central in the induction of cell motility.
FAK can mediate its effect downstream of integrin signaling toinduce polymerization of actin. Indeed, it was shown that thePPARg antagonist GW9662 has no effect on the ability of TROGto regulate FAK, highlighting that TROG itself can mediatePPARg-independent effects in cells (42). Thus, it is feasible tosuggest that the inability of TROG-treated monocytes to migratetoward CXCL12 is due to a general effect on cell motility and isindependent of CXCR4. Further studies will be required to elu-cidate the complete mechanism behind CXCR4 regulation; how-ever, CLAwas shown to regulate other transcription factors beyondthe PPARs, including ERK (43) and NF-kB (44).Murine monocytes can be classified into two main functional
subtypes, inflammatory monocytes (Gr1+/CD115+) and resident/patrolling monocytes (Gr12/CD115+), which have distinct prop-erties (45). Resident monocytes patrol blood vessels in steadystates but can also be recruited to inflammatory tissue (46) wherethey are thought to differentiate toward an M2 macrophage phe-notype with wound healing and anti-inflammatory properties (7).Inflammatory monocytes are significantly increased during hy-percholesterolemia and give rise to macrophages present in ath-erosclerotic plaques (47). Inflammatory monocytes preferentiallydifferentiate toward M1 macrophages and TNF-a/iNOS–produc-ing dendritic cells (7). Although both M1 and M2 macrophagesare present in plaques, they have different distribution patterns(48); however, a balance between the two subtypes is an importantfactor in plaque progression (49). We showed that CLA has theeffect of downregulating CD18 on inflammatory monocytes in vivoand, thus, this may have a dramatic influence on the microenvi-ronment within established plaques by shifting the balance towardthe recruitment of resident monocytes.To conclude, in this study we demonstrated that CD18 ex-
pression is downregulated by CLA, resulting in monocytes witha deficit in their ability to adhere to activated endothelium throughICAM-1. In addition, the observed increased leukocyte rollingvelocity within CLA-fed mice indicates that leukocytes are un-dergoing weaker and more fleeting interactions with the vascularendothelium, translating to decreased opportunities for leukocytesto interact with endothelial integrins and convert to firmly adherentcells. Mice fed a CD supplemented with the 80:20 blend of CLAdemonstrates that CLA has a greater influence on CD18 expressionin GR1+/CD115+ inflammatory monocytes than other leukocytesubsets. Further studies will be required to determine the overallrelationship between downregulating b2 integrins on one mono-cyte subset versus another, as well as how this alters the micro-environment within plaques, leading to regression.
AcknowledgmentsWe thank Prof. Carl G. Gahmberg for the generous gift of anti-CD18 Ab
(R2E7B). We also thank Dr. Alfonso Blanco Fernandez, Director of the
Flow Cytometry Core Facility (Conway Institute, University College Dub-
lin) for assistance with flow cytometry.
DisclosuresThe authors have no financial conflicts of interest.
References1. Cunningham, K. S., and A. I. Gotlieb. 2005. The role of shear stress in the
pathogenesis of atherosclerosis. Lab. Invest. 85: 9–23.2. Mehdizadeh, A., and A. Norouzpour. 2009. New insights in atherosclerosis:
endothelial shear stress as promoter rather than initiator. Med. Hypotheses 73:989–993.
3. Hahn, C., and M. A. Schwartz. 2009. Mechanotransduction in vascular physi-ology and atherogenesis. Nat. Rev. Mol. Cell Biol. 10: 53–62.
4. Jonasson, L., J. Holm, O. Skalli, G. Bondjers, and G. K. Hansson. 1986. Re-gional accumulations of T cells, macrophages, and smooth muscle cells in thehuman atherosclerotic plaque. Arteriosclerosis 6: 131–138.
The Journal of Immunology 4335
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

5. Hansson, G. K., L. Jonasson, B. Lojsthed, S. Stemme, O. Kocher, andG. Gabbiani. 1988. Localization of T lymphocytes and macrophages in fibrousand complicated human atherosclerotic plaques. Atherosclerosis 72: 135–141.
6. Moore, K. J., and I. Tabas. 2011. Macrophages in the pathogenesis of athero-sclerosis. Cell 145: 341–355.
7. Woollard, K. J., and F. Geissmann. 2010. Monocytes in atherosclerosis: subsetsand functions. Nat Rev Cardiol 7: 77–86.
8. Gerszten, R. E., E. A. Garcia-Zepeda, Y. C. Lim, M. Yoshida, H. A. Ding,M. A. Gimbrone, Jr., A. D. Luster, F. W. Luscinskas, and A. Rosenzweig. 1999.MCP-1 and IL-8 trigger firm adhesion of monocytes to vascular endotheliumunder flow conditions. Nature 398: 718–723.
9. Chan, J. R., S. J. Hyduk, and M. I. Cybulsky. 2001. Chemoattractants inducea rapid and transient upregulation of monocyte alpha4 integrin affinity for vas-cular cell adhesion molecule 1 which mediates arrest: an early step in the processof emigration. J. Exp. Med. 193: 1149–1158.
10. Luo, B. H., C. V. Carman, and T. A. Springer. 2007. Structural basis of integrinregulation and signaling. Annu. Rev. Immunol. 25: 619–647.
11. Diamond, M. S., D. E. Staunton, A. R. de Fougerolles, S. A. Stacker, J. Garcia-Aguilar, M. L. Hibbs, and T. A. Springer. 1990. ICAM-1 (CD54): a counter-receptor for Mac-1 (CD11b/CD18). J. Cell Biol. 111: 3129–3139.
12. Sumagin, R., H. Prizant, E. Lomakina, R. E. Waugh, and I. H. Sarelius. 2010. LFA-1and Mac-1 define characteristically different intralumenal crawling and emigrationpatterns for monocytes and neutrophils in situ. J. Immunol. 185: 7057–7066.
13. Park, Y., J. M. Storkson, K. J. Albright, W. Liu, and M. W. Pariza. 1999. Evi-dence that the trans-10,cis-12 isomer of conjugated linoleic acid induces bodycomposition changes in mice. Lipids 34: 235–241.
14. Nagao, K., N. Inoue, Y. M. Wang, and T. Yanagita. 2003. Conjugated linoleicacid enhances plasma adiponectin level and alleviates hyperinsulinemia andhypertension in Zucker diabetic fatty (fa/fa) rats. Biochem. Biophys. Res. Commun.310: 562–566.
15. Navarro, V., A. Zabala, M. T. Macarulla, A. Fernandez-Quintela, V. M. Rodrıguez,E. Simon, and M. P. Portillo. 2003. Effects of conjugated linoleic acid on body fataccumulation and serum lipids in hamsters fed an atherogenic diet. J. Physiol.Biochem. 59: 193–199.
16. Kritchevsky, D., S. A. Tepper, S. Wright, P. Tso, and S. K. Czarnecki. 2000.Influence of conjugated linoleic acid (CLA) on establishment and progression ofatherosclerosis in rabbits. J. Am. Coll. Nutr. 19: 472S–477S.
17. Lee, K. N., D. Kritchevsky, and M. W. Pariza. 1994. Conjugated linoleic acidand atherosclerosis in rabbits. Atherosclerosis 108: 19–25.
18. O’Shea, M., J. Bassaganya-Riera, and I. C. Mohede. 2004. Immunomodulatoryproperties of conjugated linoleic acid. Am. J. Clin. Nutr. 79(6, Suppl.)1199S–1206S.
19. Churruca, I., A. Fernandez-Quintela, and M. P. Portillo. 2009. Conjugatedlinoleic acid isomers: differences in metabolism and biological effects. Bio-factors 35: 105–111.
20. Toomey, S., B. Harhen, H. M. Roche, D. Fitzgerald, and O. Belton. 2006.Profound resolution of early atherosclerosis with conjugated linoleic acid. Ath-erosclerosis 187: 40–49.
21. McClelland, S., C. Cox, R. O’Connor, M. de Gaetano, C. McCarthy, L. Cryan,D. Fitzgerald, and O. Belton. 2010. Conjugated linoleic acid suppresses themigratory and inflammatory phenotype of the monocyte/macrophage cell. Ath-erosclerosis 211: 96–102.
22. Yu, Y., P. H. Correll, and J. P. Vanden Heuvel. 2002. Conjugated linoleic aciddecreases production of pro-inflammatory products in macrophages: evidencefor a PPAR gamma-dependent mechanism. Biochim. Biophys. Acta 1581: 89–99.
23. Stachowska, E., J. Kijowski, V. Dziedziejko, A. Siennicka, and D. Chlubek.2011. Conjugated linoleic acid regulates phosphorylation of PPARg by modu-lation of ERK 1/2 and p38 signaling in human macrophages/fatty acid-ladenmacrophages. J. Agric. Food Chem. 59: 11846–11852.
24. Perdomo, M. C., J. E. Santos, and L. Badinga. 2011. Trans-10, cis-12 conjugatedlinoleic acid and the PPAR-g agonist rosiglitazone attenuate lipopolysaccharide-induced TNF-a production by bovine immune cells. Domest. Anim. Endocrinol.41: 118–125.
25. Nakaya, H., B. D. Summers, A. C. Nicholson, A. M. Gotto, Jr., D. P. Hajjar, andJ. Han. 2009. Atherosclerosis in LDLR-knockout mice is inhibited, but not re-versed, by the PPARgamma ligand pioglitazone. Am. J. Pathol. 174: 2007–2014.
26. Bernhagen, J., R. Krohn, H. Lue, J. L. Gregory, A. Zernecke, R. R. Koenen,M. Dewor, I. Georgiev, A. Schober, L. Leng, T. Kooistra, et al. 2007. MIF isa noncognate ligand of CXC chemokine receptors in ininflammatory and ath-erogenic cell recruitment. Nat. Med. 13: 587–596.
27. Kuligowski, M. P., A. R. Kitching, and M. J. Hickey. 2006. Leukocyte recruit-ment to the inflamed glomerulus: a critical role for platelet-derived P-selectin inthe absence of rolling. J Immunol. 176: 6991–6999.
28. McCarthy, C., P. O’Gaora, W. G. James, M. de Gaetano, S. McClelland,D. J. Fitzgerald, and O. Belton. 2010. SorLA modulates atheroprotective prop-erties of CLA by regulating monocyte migration. Atherosclerosis 213: 400–407.
29. Dransfield, I., C. Cabanas, A. Craig, and N. Hogg. 1992. Divalent cation regu-lation of the function of the leukocyte integrin LFA-1. J. Cell Biol. 116: 219–226.
30. Verma, N. K., E. Dempsey, M. Freeley, C. H. Botting, A. Long, D. Kelleher, andY. Volkov. 2011. Analysis of dynamic tyrosine phosphoproteome in LFA-1triggered migrating T-cells. J. Cell. Physiol. 226: 1489–1498.
31. McCarthy, C., M. M. Duffy, D. Mooney, W. G. James, M. D. Griffin,D. J. Fitzgerald, and O. Belton. 2013. IL-10 mediates the immunoregulatoryresponse in conjugated linoleic acid-induced regression of atherosclerosis.FASEB J. 27: 499–510.
32. Geissmann, F., S. Jung, and D. R. Littman. 2003. Blood monocytes consist oftwo principal subsets with distinct migratory properties. Immunity 19: 71–82.
33. Evans, M., J. Brown, and M. McIntosh. 2002. Isomer-specific effects of conju-gated linoleic acid (CLA) on adiposity and lipid metabolism. J. Nutr. Biochem.13: 508.
34. Platt, I., and A. El-Sohemy. 2009. Effects of 9cis,11trans and 10trans,12cis CLAon osteoclast formation and activity from human CD14+ monocytes. LipidsHealth Dis. 8: 15–18.
35. Stachowska, E., M. Ba�skiewicz-Masiuk, V. Dziedziejko, I. Gutowska,I. Baranowska-Bosiacka, M. Marchlewicz, B. Dołegowska, B. Wiszniewska,B. Machali�nski, and D. Chlubek. 2008. Conjugated linoleic acid increases in-tracellular ROS synthesis and oxygenation of arachidonic acid in macrophages.Nutrition 24: 187–199.
36. Arbones-Mainar, J. M., M. A. Navarro, S. Acın, M. A. Guzman, C. Arnal,J. C. Surra, R. Carnicer, H. M. Roche, and J. Osada. 2006. Trans-10, cis-12- andcis-9, trans-11-conjugated linoleic acid isomers selectively modify HDL-apolipoprotein composition in apolipoprotein E knockout mice. J. Nutr. 136:353–359.
37. Stachowska, E., A. Siennicka, M. Ba�skiewcz-Hałasa, J. Bober, B. Machalinski,and D. Chlubek. 2012. Conjugated linoleic acid isomers may diminish humanmacrophages adhesion to endothelial surface. Int. J. Food Sci. Nutr. 63: 30–35.
38. Huang, C., and T. A. Springer. 1997. Folding of the beta-propeller domain of theintegrin alphaL subunit is independent of the I domain and dependent on thebeta2 subunit. Proc. Natl. Acad. Sci. USA 94: 3162–3167.
39. Hirasawa, H., T. Chiba, S. Ueki, Y. Kamada, W. Ito, M. Takeda, M. Fujita,H. Kato, H. Kayaba, and J. Chihara. 2008. The synthetic PPARgamma agonisttroglitazone inhibits eotaxin-enhanced eosinophil adhesion to ICAM-1-coatedplates. Int. Arch. Allergy Immunol. 146(Suppl. 1): 11–15.
40. Bleul, C. C., R. C. Fuhlbrigge, J. M. Casasnovas, A. Aiuti, and T. A. Springer.1996. A highly efficacious lymphocyte chemoattractant, stromal cell-derivedfactor 1 (SDF-1). J. Exp. Med. 184: 1101–1109.
41. Abi-Younes, S., A. Sauty, F. Mach, G. K. Sukhova, P. Libby, and A. D. Luster.2000. The stromal cell-derived factor-1 chemokine is a potent platelet agonisthighly expressed in atherosclerotic plaques. Circ. Res. 86: 131–138.
42. Wang, P. S., F. S. Chou, L. Porchia, M. Saji, and J. J. Pinzone. 2008. Tro-glitazone inhibits cell migration, adhesion, and spreading by modulatingcytoskeletal rearrangement in human breast cancer cells. Mol. Carcinog. 47:905–915.
43. Stachowska, E., J. Kijowski, V. Dziedziejko, A. Siennicka, and D. Chlubek.2011. Conjugated linoleic acid regulates phosphorylation of PPARg by modu-lation of ERK 1/2 and p38 signaling in human macrophages/fatty acid-ladenmacrophages. J. Agric. Food Chem. 59: 11846–11852 .
44. Cheng, W. L., C. K. Lii, H. W. Chen, T. H. Lin, and K. L. Liu. 2004. Contri-bution of conjugated linoleic acid to the suppression of inflammatory responsesthrough the regulation of the NF-kappaB pathway. J. Agric. Food Chem. 52: 71–78.
45. Geissmann, F., C. Auffray, R. Palframan, C. Wirrig, A. Ciocca, L. Campisi,E. Narni-Mancinelli, and G. Lauvau. 2008. Blood monocytes: distinct subsets,how they relate to dendritic cells, and their possible roles in the regulation of T-cell responses. Immunol. Cell Biol. 86: 398–408.
46. Auffray, C., D. Fogg, M. Garfa, G. Elain, O. Join-Lambert, S. Kayal, S. Sarnacki,A. Cumano, G. Lauvau, and F. Geissmann. 2007. Monitoring of blood vesselsand tissues by a population of monocytes with patrolling behavior. Science 317:666–670.
47. Swirski, F. K., P. Libby, E. Aikawa, P. Alcaide, F. W. Luscinskas, R. Weissleder,and M. J. Pittet. 2007. Ly-6Chi monocytes dominate hypercholesterolemia-associated monocytosis and give rise to macrophages in atheromata. J. Clin.Invest. 117: 195–205.
48. Stoger, J. L., M. J. Gijbels, S. van der Velden, M. Manca, C. M. van der Loos,E. A. Biessen, M. J. Daemen, E. Lutgens, and M. P. de Winther. 2012. Distri-bution of macrophage polarization markers in human atherosclerosis. Athero-sclerosis 225: 461–468.
49. Mantovani, A., C. Garlanda, and M. Locati. 2009. Macrophage diversity andpolarization in atherosclerosis: a question of balance. Arterioscler. Thromb. Vasc.Biol. 29: 1419–1423.
4336 CONJUGATED LINOLEIC ACID INHIBITS MONOCYTE ADHESION
by guest on March 31, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











