Conifer and Angiosperm Fine-Root Biomass in Relation to Stand Age and Site Productivity in Douglas-Fir Forests Author(s): Kristiina A. Vogt, Daniel J. Vogt, Erin E. Moore, Babatunde A. Fatuga, Mark R. Redlin and Robert L. Edmonds Source: Journal of Ecology, Vol. 75, No. 3 (Sep., 1987), pp. 857-870 Published by: British Ecological Society Stable URL: http://www.jstor.org/stable/2260210 . Accessed: 26/02/2014 17:40 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . British Ecological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal of Ecology. http://www.jstor.org This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PM All use subject to JSTOR Terms and Conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Conifer and Angiosperm Fine-Root Biomass in Relation to Stand Age and Site Productivity inDouglas-Fir ForestsAuthor(s): Kristiina A. Vogt, Daniel J. Vogt, Erin E. Moore, Babatunde A. Fatuga, Mark R.Redlin and Robert L. EdmondsSource: Journal of Ecology, Vol. 75, No. 3 (Sep., 1987), pp. 857-870Published by: British Ecological SocietyStable URL: http://www.jstor.org/stable/2260210 .
Accessed: 26/02/2014 17:40
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
British Ecological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal ofEcology.
http://www.jstor.org
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

Journal of Ecology (1987), 75, 857-870
CONIFER AND ANGIOSPERM FINE-ROOT BIOMASS IN RELATION TO STAND AGE AND SITE
PRODUCTIVITY IN DOUGLAS-FIR FORESTS
KRISTIINA A. VOGT, DANIEL J. VOGT, ERIN E. MOORE, BABATUNDE A. FATUGA, MARK R. REDLIN AND
ROBERT L. EDMONDS
College of Forest Resources, AR-JO, University of Washington, Seattle, Washington 98195, U.S.A.
SUMMARY
(1) Conifer and angiosperm fine-root biomass were determined in an age sequence (range 13-160 years) of eight high-productivity and seven low-productivity Douglas-fir stands in western Washington, U.S.A.
(2) No significant changes in total (conifer plus angiosperm) root biomass occurred among the different stand ages within each productivity class. Of the total fine-root biomass, however, the proportion of angiosperm to conifer roots varied significantly when stands were in the 'tree initiation' and 'competitive tree-stem exclusion' stages of stand development.
(3) Conifer fine-root biomass reached a peak at canopy closure in both low- and high- productivity stands. After canopy closure, conifer fine-root and foliage biomass levelled off as basal area continued to increase in the low-productivity stands; however, conifer fine-root biomass decreased as basal area increased and foliage biomass levelled off in the high-productivity stands.
(4) At all stand ages, the low-productivity stands had significantly more total (conifer plus angiosperm) fine-root biomass than those of high-productivity. At canopy closure and later stages of stand development, a significantly higher conifer fine-root biomass was maintained in low-productivity compared to high-productivity stands.
(5) Conifer fine-root biomass was positively related to litterfall lignin content, litterfall lignin: nitrogen ratio and forest-floor nitrogen content in low-, but not in high- productivity stands; these patterns followed an asymptotic exponential curve.
INTRODUCTION
Stand development in forests after a disturbance can be divided into four general stages: (i) tree initiation, angiosperm dominance; (ii) tree dominance with competitive tree-stem exclusion, angiosperms reduced; (iii) tree dominance, angiosperms redevelop as gaps form in tree canopy; and (iv) tree-height growth stops. Although several structural and functional changes occurring with stand development through these stages have been extensively studied, research has focused almost exclusively on above-ground compo- nents. For example, forest managers have for years been measuring changes in bole biomass to predict tree growth based on site quality and stand age (Daniel, Helms & Baker 1979).
Based only on above-ground components, some generalities on structural changes with stand development have been shown. In most forests, stand basal area increases until trees are approximately 100 years old (Daniel, Helms & Baker 1979). While other aspects of stand development such as foliage biomass, litterfall, net primary production and nutrient
857.
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

858 Fine-root biomass in Douglas-fir forests
accumulation in above-ground tree components tend to reach a plateau at the 'competitive tree stem exclusion' stage (Kira & Shidei 1967; Gessel & Turner 1976; MacLean & Wein 1977; Tadaki et al. 1977; Oliver 1981; Singh 1982; Sprugel 1985). This stage begins at crown closure and can occur anywhere from 5 to 60 years after 'tree initiation' depending upon the plant species and stand density. The closing of the overstorey canopy generally coincides with a dramatic decrease in above-ground understorey biomass associated with reduced penetration of light (MacLean & Wein 1977; Alaback 1982).
Recent research has inferred the importance of understorey angiosperm plants in ecosystem-level processes at different stages of stand development (Carlisle, Brown & White 1967; Turner & Long 1975; MacLean & Wein 1977,1978; Roberts et al. 1980; Yarie 1980; Black & Spittlehouse 1981; Alaback 1982; Davison & Forman 1982; Hough 1982; Martin, Sharick & Smith 1982). Yet only a few of these studies (Turner & Long 1975; MacLean & Wein 1977; Roberts et al. 1980; Black & Spittlehouse 1981; Alaback 1982) actually examined changes in both understorey and overstorey above-ground biomass across a series of different-aged stands to allow quantitative evaluation of the understorey importance in carbon, nutrient and water budgets.
Even fewer researchers have examined how the below-ground component changes as a stand develops or as a function of stand nutritional status (site quality). Karizumi (1968), Berish (1982) and Vogt et al. (1983a) showed that fine roots of trees reach a maximum biomass at the time of canopy closure. Vogt et al. (1983a), however, only considered roots of trees in the forest floor. None of these studies evaluated root biomass of understorey shrubs. Also unknown is the effect of site productivity on fine-root biomass across stand age. Some stand-age studies have suggested that fine-root biomass will vary in response to changes in nutrient availability (Ericsson & Persson 1980; Persson 1980a; Keyes & Grier 1981; Linder & Rook 1984; Vogt et al. 1985).
This study was initiated to examine several hypotheses related to the growth of conifers on fertile and infertile sites. The first hypothesis states that stands growing on infertile sites have slower above-ground growth rates resulting from more photosynthate translocation to roots for sequestering limiting nutrients required for growth than stands growing on better, more fertile sites. If this hypothesis is correct, a smaller fine-root biomass should be maintained and a higher basal area should be reached at a younger stand age for trees growing on fertile versus infertile sites. The second hypothesis states that tree foliage and fine-root biomass are in equilibrium once canopy has maximized, with any reductions in foliage biomass resulting in concomitant decreases in root biomass in older stands. This equilibrium between foliage and roots occurs at an earlier age for trees growing on more fertile sites. The third hypothesis states that maximum root occupancy occurs at all stages of stand development, with angiosperms contributing more root biomass at earlier stages and conifers at later stages of stand development.
METHODS
Study plots Fifteen stands, all dominated by naturally regenerated Douglas-fir (Pseudotsuga
menziesii Mirb.), were selected in the foothills of the Cascade Mountains in western Washington (47?N, 122?W). Description of stand locations are given in greater detail in Vogt et al. (1 983a). An age sequence of eight nutrient-rich, high-productivity (class II) and
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
K. A. VOGT et al. 859
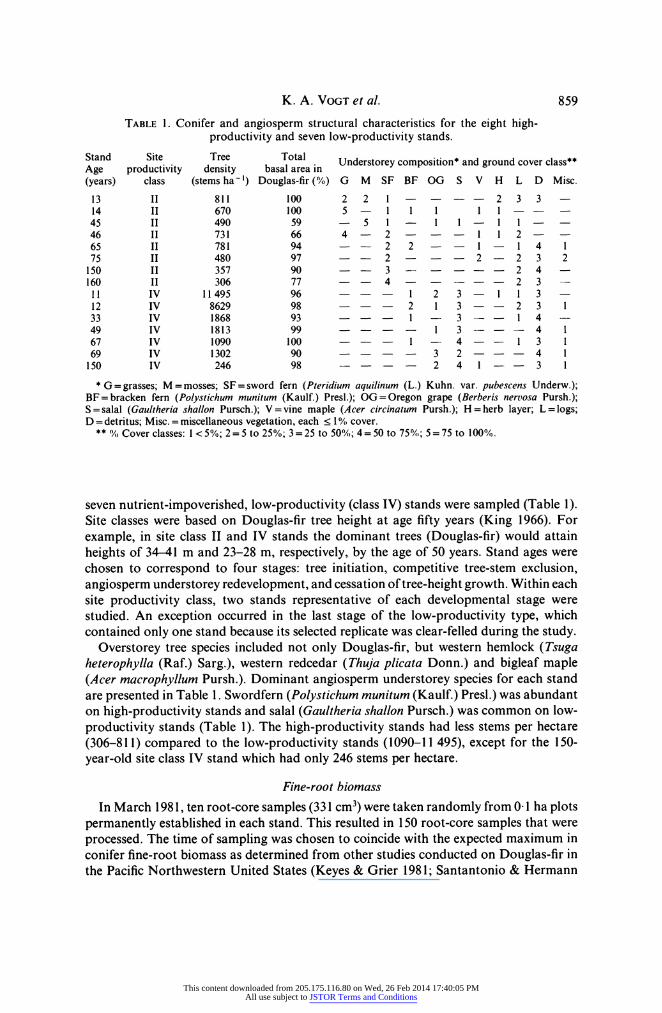
TABLE 1. Conifer and angiosperm structural characteristics for the eight high- productivity and seven low-productivity stands.
Stand Site Tree Total Understoreycomposition* and groundcoverclass" Age productivity density basal area in (years) class (stems ha-') Douglas-fir (0%) G M SF BF OG S V H L D Misc. 13 II 811 100 2 2 1 2 3 3 14 II 670 100 5 -1 1 1 11-- 45 II 490 59 - 51- 1 1 11- -
46 II 731 66 4 - 2 ---1 1 2- 65 II 781 94 -- 2 2 --I- 1 4 1 75 II 480 97 -- 2 --- 2 - 2 3 2
150 II 357 90 -- 3 2 4 160 II 306 77 -- 4 2 3 - 11 IV 11495 96 - - - 1 2 3 - 1 1 3 12 IV 8629 98 --- 2 1 3 -- 2 3 1 33 IV 1868 93 --- 1 - 3 - - 1 4 -
49 IV 1813 99 ---- 1 3 - - 4 1 67 IV 1090 100 --- 1- 4 -- 1 3 1 69 IV 1302 90 ---- 3 2 --- 4 1 150 IV 246 98 - --- 2 4 1 -- 3 1
* G =grasses; M=mosses; SF=sword fern (Pteridium aquilinum (L.) Kuhn. var. pubescens Underw.); BF=bracken fern (Polystichum munitum (Kaulf.) Presl.); OG=Oregon grape (Berberis nertosa Pursh.); S=salal (Gaultheria shallon Pursch.); V=vine maple (Acer circinatum Pursh.); H=herb layer; L=logs; D =detritus; Misc. = miscellaneous vegetation, each < 1 /O, cover.
** 0/ Cover classes: 1 < 5 0%; 2 = 5 to 25 0%; 3 = 25 to 50%/,; 4= 50 to 75 0%; 5 = 75 to 100%/,.
seven nutrient-impoverished, low-productivity (class IV) stands were sampled (Table 1). Site classes were based on Douglas-fir tree height at age fifty years (King 1966). For example, in site class II and IV stands the dominant trees (Douglas-fir) would attain heights of 34-41 m and 23-28 m, respectively, by the age of 50 years. Stand ages were chosen to correspond to four stages: tree initiation, competitive tree-stem exclusion, angiosperm understorey redevelopment, and cessation of tree-height growth. Within each site productivity class, two stands representative of each developmental stage were studied. An exception occurred in the last stage of the low-productivity type, which contained only one stand because its selected replicate was clear-felled during the study.
Overstorey tree species included not only Douglas-fir, but western hemlock (Tsuga heterophylla (Raf.) Sarg.), western redcedar (Thuja plicata Donn.) and bigleaf maple (Acer macrophyllum Pursh.). Dominant angiosperm understorey species for each stand are presented in Table 1. Swordfern (Polystichum munitum (Kaulf.) Presl.) was abundant on high-productivity stands and salal (Gaultheria shallon Pursch.) was common on low- productivity stands (Table 1). The high-productivity stands had less stems per hectare (306-811) compared to the low-productivity stands (1090-11 495), except for the 150- year-old site class IV stand which had only 246 stems per hectare.
Fine-root biomass In March 1981, ten root-core samples (331 cm3) were taken randomly from 0.1 ha plots
permanently established in each stand. This resulted in 150 root-core samples that were processed. The time of sampling was chosen to coincide with the expected maximum in conifer fine-root biomass as determined from other studies conducted on Douglas-fir in the Pacific Northwestern United States (Keyes & Grier 1981; Santantonio & Hermann
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

860 Fine-root biomass in Douglas-fir forests
1985; Vogt et al. 1985). Details on stand design, root sampling (along with processing procedures) and biomass calculations are described in other publications (Vogt et al. 1981, 1983a, b). Root cores consisted of the forest floor (ranging from 1 1 to 7 4-cm depths, Vogt et al. 1983a) and soil collected to a 15-cm depth. Root-core samples were sorted into organic and mineral horizons and separately bagged in the field. Fine-root biomass included roots < 2 mm in diameter.
Distinction between conifer overstorey and angiosperm understorey fine roots was relatively simple because of their distinctly different morphologies. Fine roots (both conifer and angiosperm) were separated into live and dead categories using previously described criteria (Vogt et al. 1981) and also with the use of a potassium-iodide solution which stains starch granules black. Dead roots contain no starch granules, senescing roots have reduced starch granules, and live roots contain abundant granules. Only live-root biomass will be presented in this paper.
Above-ground measurements For qualitative comparison, percentage angiosperm understorey ground cover was
estimated visually for nine separate, permanently marked (10 m x 10 m) quadrats within each stand and then averaged together to obtain a stand mean. Percentage plant cover was divided into five classes: 1 = <5%, 2=5-25%, 3 =25-50%, 4=50-75% and 5 = 75- 100% cover of the ground surface.
Conifer overstorey basal area was determined from circumference measurements taken at breast height ([ 37 m) for each tree within a plot and converted to an area basis. Regression equations developed by Gholz et al. (1979) were used to estimate tree component biomass with diameter at breast height (dbh). However the estimation of current and total foliage biomass using dbh in regressions is thought to be subject to potential error especially in more mature stands (Grier & Waring 1974; Snell & Brown 1978; Marshall & Waring 1986). Total foliage biomass was therefore estimated using the cross-sectional sapwood area method to calculate leaf area (Whitehead 1978; Kaufmann & Troendle 1981; Long, Smith & Scott 1981; Waring, Schroeder & Oren 1982; Pearson, Fahey & Knight 1984). Coefficients of leaf area: sapwood area (0 47 m2 cm-2 at breast height or, if the tree taper was known, 0 54 m2 cm-2 at the base of the crown) (Waring, Schroeder & Oren 1982) were multiplied by the sapwood area to obtain projected leaf area. Leaf area was then divided by its specific leaf area (SLA) to convert it to leaf biomass. All closed-canopy stands in this study were assumed to have a SLA of 7-39 m2 kg-' (Gholz et al. 1976). However, for the three open-canopy stands, site-specific SLAs were developed and used (stand age 14 = 7 03 and stand ages 11 and 12 = 7 25 m2 kg- ').
Conifer overstorey litterfall was assumed to equal current foliage biomass (Keyes & Grier 1981). However, the sapwood-area method only estimates total foliage biomass. Even though it was found that using dbh in regressions (Gholz et al. 1979) overestimated total foliage, it was assumed that the ratio between current and total foliage was not affected. The current foliage biomass was therefore estimated by multiplying the proportion of current: total foliage biomass, derived from the regressions presented by Gholz et al. (1979), by the total foliage derived from the sapwood method. Within-stand variation was presented by dividing each 0- 1 -ha plot into equal thirds. Standard deviation of conifer foliage biomass was then derived using n = 3.
Some needle litterfall was collected in October 1979 from all stands. Three subsamples of needles from each stand were analysed chemically for total nitrogen and lignin. Total nitrogen was determined using a digestion method described by Parkinson & Allen (1975)
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

K. A. VOGT et al. 861
and analysed on a Technicon Autoanalyzer II. Lignin concentration was determined using the Klason (acid-insoluble) lignin method (Tappi Standard 1974).
In each stand, average forest-floor mass was determined from ten samples (two from five randomly selected subplots within each stand). A circular metal corer (16-cm inside diameter) was used for sampling the forest floor. All woody material greater than 2-mm diameter was removed from the forest floor samples prior to drying at 70 ?C; the dry weights were converted to an ash-free basis by measuring the weight loss of subsample after ignition (450 C). The two forest-floor samples collected from each subplot were combined and analysed for total nitrogen in the same manner as the litterfall.
Statistical procedures A one-way analysis of variance was used to determine if significant root biomass
differences existed for each independent variable (age and productivity class). When significant differences were observed, a Student-Newman-Keuls test (at P < 0 10) was used to differentiate root biomass means within the different stand ages or productivity classes (Snedecor & Cochran 1967). Statistical analyses were conducted using the 'Statistical Package for the Social Sciences' (SPSS) software (Nie et al. 1975; Hull & Nie 1981).
An iterative non-linear curve fitting program was used to develop the curves between conifer fine-root biomass and litterfall lignin content, litterfall lignin: nitrogen ratio, and forest-floor N content.
RESULTS
Conifer and angiospermfine-root biomass
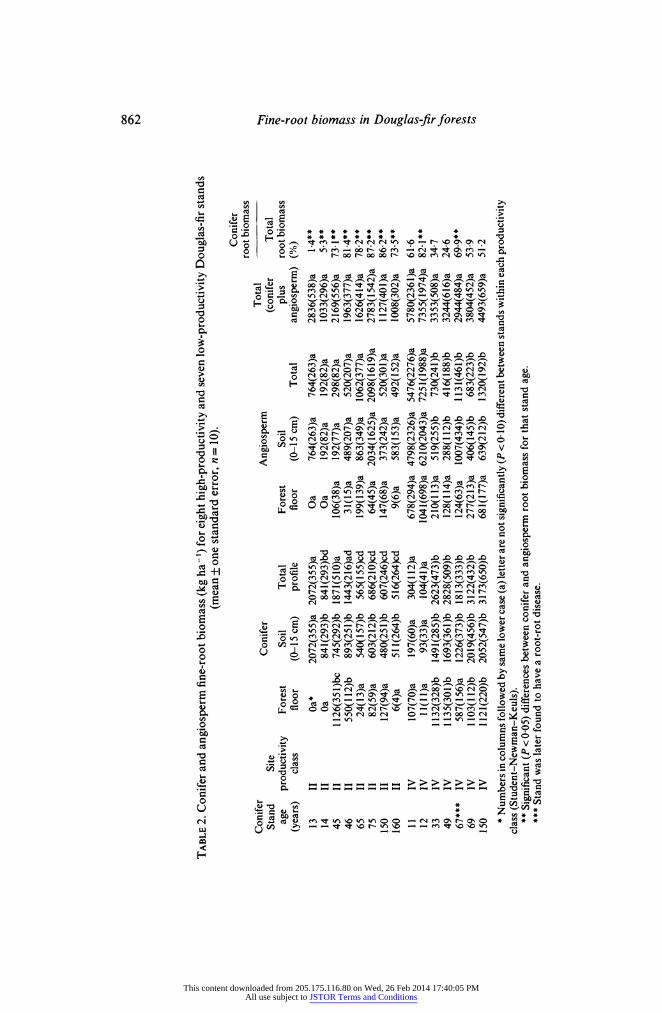
In the high-productivity stands, slightly more fine-root biomass of conifers was observed in the combined forest floor and soil (to 15 cm depth) in the 13-46-year-old stands than in the older stands, although these were not statistically significant (Table 2). In the low-productivity stands, significantly higher angiosperm and lower conifer fine- root biomass were measured in the forest floor plus soil of the 1 1- and 12-year-old stands than in the remaining stand ages.
In the high-productivity stands, significantly higher conifer fine-root biomass was measured in the forest floors of the 45- and 46-year-old stands than in the remaining stand ages (Table 2). Except for the 13-year-old stand, there were no significant differences in conifer fine-root biomass in the soil (Table 2). There were no significant differences in angiosperm fine-root biomass between stands in either the forest floor or soil.
Low-productivity stands generally had significantly lower conifer fine-root biomass in both the forest floor and soil of the 1 1- and 12-year-old stands than in other stand ages (Table 2). A significantly higher angiosperm fine-root biomass was measured in the soil of the 1 1- and 12-year-old stands than all the remaining stand ages (Table 2).
No significant differences were found in the combined conifer plus angiosperm fine- root biomass among different-aged stands within the same productivity class (Table 2). Comparison of total fine-root biomass by productivity class, however, showed that the high-productivity stands had significantly (P<0 10) less combined conifer and angio- sperm root biomass than the low-productivity stands.
The proportion of fine-root biomass composed of conifer roots for the total sampled profile is given in Table 2. Angiosperm roots comprised a significantly higher proportion
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
862 Fine-root biomass in Douglas-fir forests
cn
co c
< c - E,
u 0 0 SC _TO enj - ooT C'4 C4 C1 W) D - 1~ N ON d 0<2
0 -00 r- 00 00 r- 1< 00 cen C14 1z0 tO} tO}
S_ co co co co co
cti? b t o co -A A co co co co
C C - ~ ~~~~~~~~ oo =c _c r--> t _
r- oo N o t
> ci 00 en C (Z l- -- O- or ) ) en CN ?O - ,0 or
O- t O' -go
N _ (O
N _ _O M
,- 0 O 1r M ' IT IT
~~~~~~~~~~~e en10 I'D CA 00 Ctfi 0t 0 or) 4n DR ^R ^ 3~W =~~~~~~~~~C _0 Oe - (Z _~ _ - oo- 00 - M en C4( 0
0 CI C1 00I 00 - or) _- en en C, en 't _ D
ct~~~~~~~~~~~~~~~i CO CI
en, I d - ON Ch "I- e.O 000 ? V
-v 11 C O ? Ch Ch 00 ? o- C14l - 00 C - -00 en CA
> * . --d 00 00C 0 - M t t- C - 9'T C1 - tD
z~~~~~~~~~F ,t ^' O o 5< oo 5- Q 5 5 5 en _ '= _ _ -r [,L =~~~~~~~1~ ? Z ? Z -C14 1 t ( C14(O 00- or) en b- en 0"
Ct - _ m ^ O ? m~~~~~~~~ O "ct "ct 1Y en S.^ i
cn E ~~~~~~~~n0 I.- I.- t- 0 t C1 q m W) C1 _q W) cl O t ^
g
ce ~~~~~~~~D D1 D- D DIc D~ DIc D n D en 'i D
*- v_ - ~~~~~~~Its "> "r cON en It en en 00 o e 00 r- 1N (Z 1 3 c , - ~ ~~~~~~~~~~~ ( ZN -Z 0 1^ en r- 00 - Z - - 0 O O en w 3?
o S: c;T o 0 oo en oo m - " W ? " C> o, ,C S.~~~~~ ~~~ C1 "C 'I t cis
zC ~ ~~~~~~~~ 00 WI * or _ 0 - - (Z t - en ? _ - C. N ' * ;
ci co en ? ? ct O . d . "D l-4 l - _ - O . X~~~~~~~~ I' ( N oot r (O-N oo O 00 "c r- C
C1 O n Z 0 It - " "$ W . , ~ ~~~~~~~~~~~~~~~~~~~~~~ ., - - V, W~
C1 ) C' 0Cw t z cis* >~~~~~~~~c *0 Ot t4O 1r)@- O q >- t?* *ItC4M(Z -
z~~~~~~~~~~~r ( Cth r - 1~ 00 d ?- O O -1 1 - d1 r- W5}
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
K. A. VOGT et al. 863
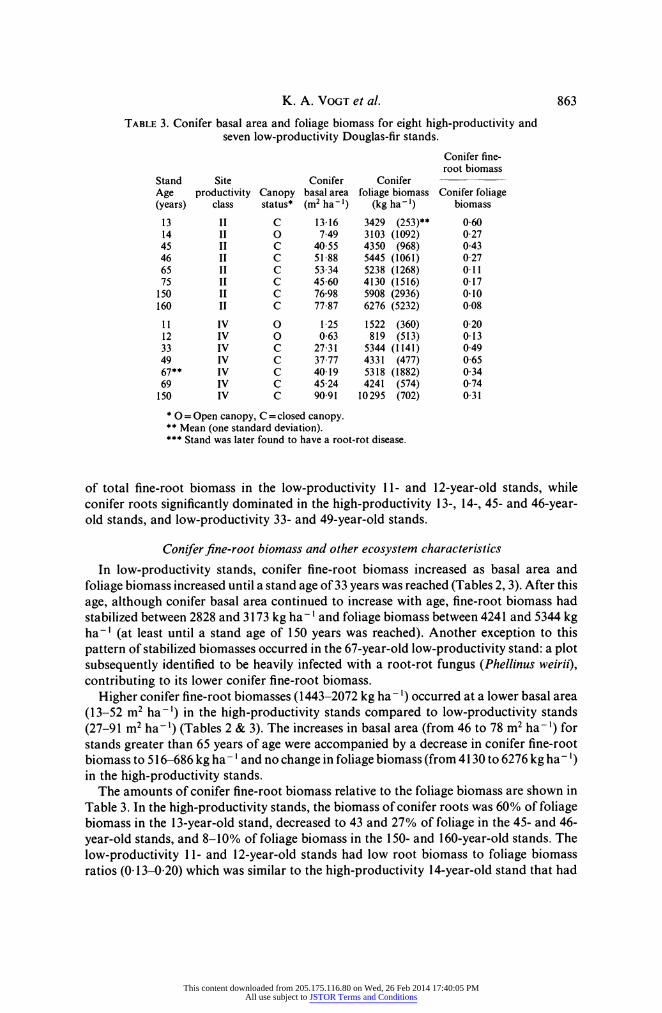
TABLE 3. Conifer basal area and foliage biomass for eight high-productivity and seven low-productivity Douglas-fir stands.
Conifer fine- root biomass
Stand Site Conifer Conifer Age productivity Canopy basal area foliage biomass Conifer foliage (years) class status* (m2 ha- 1) (kg ha 1) biomass 13 II C 13 16 3429 (253)** 060 14 II 0 7 49 3103 (1092) 0 27 45 II C 40 55 4350 (968) 0 43 46 II C 51 88 5445 (1061) 027 65 II C 53 34 5238 (1268) 0 11 75 II C 4560 4130 (1516) 0 17 150 II C 76-98 5908 (2936) 0 10 160 II C 77 87 6276 (5232) 0 08 11 IV 0 1 25 1522 (360) 020 12 IV 0 063 819 (513) 0 13 33 IV C 27 31 5344 (1141) 049 49 IV C 37 77 4331 (477) 0 65 67** IV C 40 19 5318 (1882) 034 69 IV C 45 24 4241 (574) 0 74 150 IV C 90 91 10295 (702) 0 31
*0 = Open canopy, C = closed canopy. ** Mean (one standard deviation). *** Stand was later found to have a root-rot disease.
of total fine-root biomass in the low-productivity 11- and 12-year-old stands, while conifer roots significantly dominated in the high-productivity 13-, 14-, 45- and 46-year- old stands, and low-productivity 33- and 49-year-old stands.
Coniferfine-root biomass and other ecosystem characteristics
In low-productivity stands, conifer fine-root biomass increased as basal area and foliage biomass increased until a stand age of 33 years was reached (Tables 2, 3). After this age, although conifer basal area continued to increase with age, fine-root biomass had stabilized between 2828 and 3173 kg ha- ' and foliage biomass between 4241 and 5344 kg ha-' (at least until a stand age of 150 years was reached). Another exception to this pattern of stabilized biomasses occurred in the 67-year-old low-productivity stand: a plot subsequently identified to be heavily infected with a root-rot fungus (Phellinus weirii), contributing to its lower conifer fine-root biomass.
Higher conifer fine-root biomasses (1443-2072 kg ha-1) occurred at a lower basal area (13-52 m2 ha-') in the high-productivity stands compared to low-productivity stands (27-91 m2 ha-') (Tables 2 & 3). The increases in basal area (from 46 to 78 m2 ha-') for stands greater than 65 years of age were accompanied by a decrease in conifer fine-root biomass to 516-686 kg ha- ' and no change in foliage biomass (from 4130 to 6276 kg ha-') in the high-productivity stands.
The amounts of conifer fine-root biomass relative to the foliage biomass are shown in Table 3. In the high-productivity stands, the biomass of conifer roots was 60% of foliage biomass in the 13-year-old stand, decreased to 43 and 27% of foliage in the 45- and 46- year-old stands, and 8-10% of foliage biomass in the 150- and 160-year-old stands. The low-productivity 11- and 12-year-old stands had low root biomass to foliage biomass ratios (0 13-0 20) which was similar to the high-productivity 14-year-old stand that had
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
864 Fine-root biomass in Douglas-fir forests
not reached crown closure. Low-productivity stands 33-, 49- and 69-years old maintained high conifer root: foliage biomass ratios (0 49-074). High-productivity stands 65 years old or greater had less than 20% root biomass relative to the foliage biomass in contrast to 31-74% in the low productivity stands.
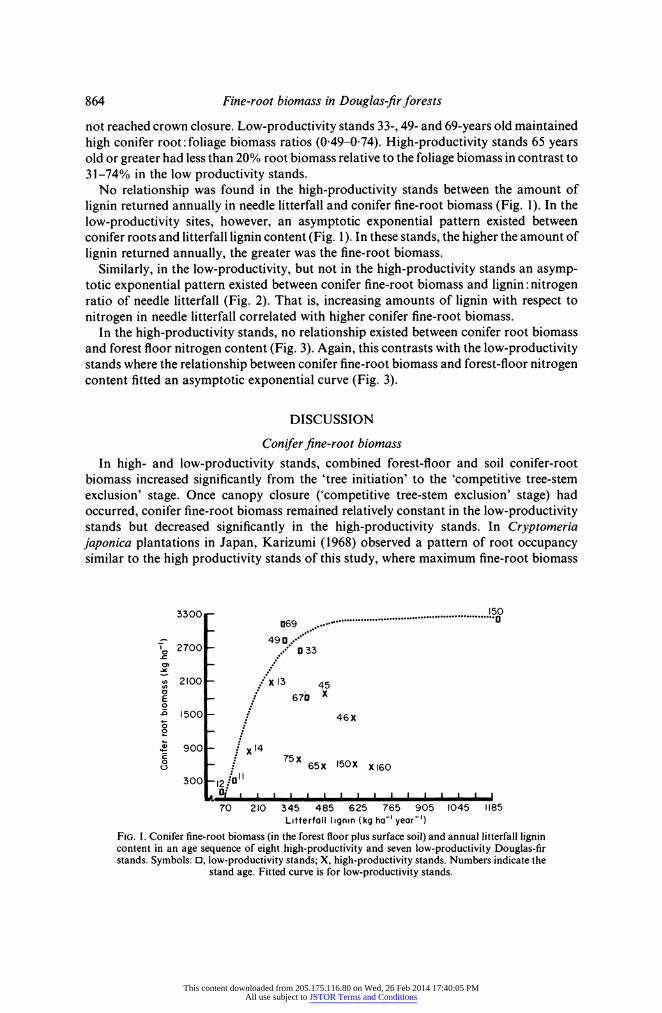
No relationship was found in the high-productivity stands between the amount of lignin returned annually in needle litterfall and conifer fine-root biomass (Fig. 1). In the low-productivity sites, however, an asymptotic exponential pattern existed between conifer roots and litterfall lignin content (Fig. 1). In these stands, the higher the amount of lignin returned annually, the greater was the fine-root biomass.
Similarly, in the low-productivity, but not in the high-productivity stands an asymp- totic exponential pattern existed between conifer fine-root biomass and lignin: nitrogen ratio of needle litterfall (Fig. 2). That is, increasing amounts of lignin with respect to nitrogen in needle litterfall correlated with higher conifer fine-root biomass.
In the high-productivity stands, no relationship existed between conifer root biomass and forest floor nitrogen content (Fig. 3). Again, this contrasts with the low-productivity stands where the relationship between conifer fine-root biomass and forest-floor nitrogen content fitted an asymptotic exponential curve (Fig. 3).
DISCUSSION
Coniferfine-root biomass In high- and low-productivity stands, combined forest-floor and soil conifer-root
biomass increased significantly from the 'tree initiation' to the 'competitive tree-stem exclusion' stage. Once canopy closure ('competitive tree-stem exclusion' stage) had occurred, conifer fine-root biomass remained relatively constant in the low-productivity stands but decreased significantly in the high-productivity stands. In Cryptomeria japonica plantations in Japan, Karizumi (1968) observed a pattern of root occupancy similar to the high productivity stands of this study, where maximum fine-root biomass
3300 150 0369 ...0
490 T 2700 . 0 33
2100 x13 45
E - 670 X
D 1500 46X 0
900 14
'~~? ~ 75x ~65X 150 X X 160
0: 300 12..o 1 0 t ' ' ' I I I I I
70 210 345 485 625 765 905 1045 1185 Litterfall lignin (kg ha-' year-')
FIG. 1. Conifer fine-root biomass (in the forest floor plus surface soil) and annual litterfall lignin content in an age sequence of eight high-productivity and seven low-productivity Douglas-fir stands. Symbols: 0, low-productivity stands; X, high-productivity stands. Numbers indicate the
stand age. Fitted curve is for low-productivity stands.
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
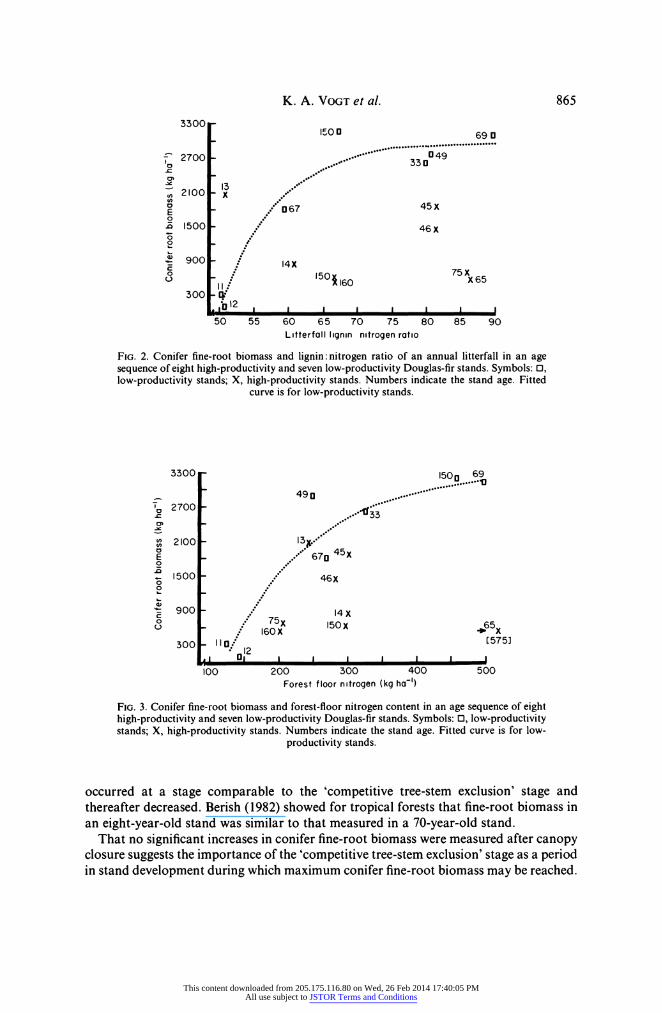
K. A. VOGT et al. 865
3300 Ir00 690
....... .......................................
2700 049 330
- ~ 13 ,2100 X ow /- 067 45X E 0 D 1500 _ 46 X o: 0
900 . 14 X o - . 1501 75 X
o 11, 'J" 1*60 X65 300 _ ,
,12
50 55 60 65 70 75 80 85 90 Litterfall lignin nitrogen ratio
FIG. 2. Conifer fine-root biomass and lignin: nitrogen ratio of an annual litterfall in an age sequence of eight high-productivity and seven low-productivity Douglas-fir stands. Symbols: 0, low-productivity stands; X, high-productivity stands. Numbers indicate the stand age. Fitted
curve is for low-productivity stands.
3300 _500 69
490 T 2700
u 2100 _3 E .. * 670
1500 46x 0
900 4 _ . 14 X 0 . 75X 150X
160 X 300 * 12 575i 12
1 01 I I I I I 100 200 300 400 500
Forest floor n i trogen (kg ha-')
FIG. 3. Conifer fine-root biomass and forest-floor nitrogen content in an age sequence of eight high-productivity and seven low-productivity Douglas-fir stands. Symbols: 0, low-productivity stands; X, high-productivity stands. Numbers indicate the stand age. Fitted curve is for low-
productivity stands.
occurred at a stage comparable to the 'competitive tree-stem exclusion' stage and thereafter decreased. Berish (1982) showed for tropical forests that fine-root biomass in an eight-year-old stand was similar to that measured in a 70-year-old stand.
That no significant increases in conifer fine-root biomass were measured after canopy closure suggests the importance of the 'competitive tree-stem exclusion' stage as a period in stand development during which maximum conifer fine-root biomass may be reached.
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

866 Fine-root biomass in Douglas-fir forests
This maximum would be expected, because trees fully occupy the site at this time and therefore are growing in the highly competitive environment that exists in a closed forest. Rooting density will also be restricted at this time unless new substrates become available for roots, such as increased forest floor accumulation (Vogt et al. 1983b) or greater use of deeper soil horizons (McQueen 1968).
Conifer and angiosperm fine-root biomass A complementary relationship existed between conifer and angiosperm contribution to
total fine-root biomass in the low- and high-productivity stands. Total fine-root biomass did not vary significantly with stand age within each productivity class, but the proportion of conifer to angiosperm roots fluctuated within this total. This occurred despite the fact that the contribution of conifer or angiosperm root biomass to the total fluctuated significantly in some stand ages.
Angiosperm roots contributed substantially to total fine-root biomass during the early stages of 'tree initiation' and later during 'angiosperm redevelopment' stage when gaps form in the canopy. These patterns of high angiosperm root biomass are similar to the cycles of high aboveground angiosperm understorey biomass recorded with stand development by MacLean & Wein (1977), Yarie (1980) and Alaback (1982). During the 'angiosperm redevelopment' and 'cessation of tree-height growth' stages, the high- productivity stands had a higher proportion of angiosperm roots (46-75%) to total fine- root biomass than did the low-productivity stands (18-38%). The higher proportion of angiosperm to total root biomass in the high-productivity stands at these developmental stages are not reflected in the visual determination of percentage ground cover by angiosperm understorey vegetation.
It has to be kept in mind that the contribution of angiosperm roots to total root biomass will vary considerably between stands based on the density and basal area of the conifer vegetation. For instance, the two Scots pine stands studied by Persson (1980b, c) in Sweden appeared to be more open and showed a high dominance of the rooting space by the shrub understorey. In a young (15-20-year-old) Scots pine stand, an estimated 79% of the total live fine-root biomass was contributed by understorey vegetation while a value of 64% was estimated for the mature (120-year-old) stand (Persson 1980b, c). Those stands contrasted with the high density Abies amabilis stands studied by Vogt et al. (1982, 1983b) in Washington, U.S.A., where angiosperm roots only contributed from 24 to 2% of total root biomass in a 23- and 180-year-old stand, respectively (Vogt et al. 1982, 1983b).
Total fine-root (angiosperm and conifer) biomass in this study was significantly greater in the low-productivity than high-productivity stands. These data reflect a general pattern that fine-root biomass is greater in naturally poor than rich sites (Keyes & Grier 1981) and that decreases in fine-root biomass may occur when a poor site is fertilized (Linder & Rook 1984; Vogt et al. 1985).
Basal area, fine roots andfoliage biomass The time of canopy closure determined the basal area at which maximum root biomass
was reached in low- and high-productivity stands. Maximum conifer fine-root biomass was reached at a lower basal area and younger stand age in the high-productivity (13 m2 ha- ') than in the low-productivity stands (45 m2 ha -' ). These differences are explained by the fact that high-productivity stands reached canopy closure at an earlier stand age than low-productivity stands. In fact, canopy closure had already occurred in the 13-year-old high-productivity stand, but did not occur until 33 and 49 years in the low-productivity
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

K. A. VOGT et al. 867
stands. These differences by productivity class illustrate that low- and high-productivity stands go through the same developmental stages, but the high-productivity stands go through each stage at an earlier stand age (Daniel, Helms & Baker 1979).
After canopy closure, conifer fine-root biomass either levelled off or decreased with increasing conifer basal area depending on the productivity class. In the low-productivity stands, no increase in conifer root biomass occurred between basal areas of 45-91 m2 ha- '. The one stand that had less fine-root biomass for its basal area was the 67-year-old low-productivity stand which was affected by root disease. Unlike stands of low- productivity, high-productivity stands had a decrease in conifer fine-root biomass between basal areas of 45 and 78 m2 ha- '. After canopy closure, both the low- and high- productivity stands had similar increases in basal area but the low-productivity stands supported about four times greater conifer fine-root biomass.
Prior to canopy closure, high- and low-productivity stands maintained a greater foliage biomass compared to fine-root biomass: fine-root biomass was 13-27% of foliage biomass. At this stage in stand development, more carbon appeared to be allocated to producing photosynthetic tissues than for tissue involved in nutrient acquisition. At canopy closure, the biomasses of fine roots and foliage maintained by a stand were more similar (fine-root biomass was 49-65% of foliage biomass). In general, foliage and root biomasses had both equilibrated by the 'competitive tree-stem exclusion' and 'angio- sperm understorey redevelopment' stages for the low-productivity, but not for the high- productivity stands. While foliage biomass had equilibrated by the 'competitive tree-stem exclusion' stage in the high-productivity stands, fine-root biomass decreased significantly at the 'angiosperm understorey redevelopment' stage.
Fine roots and nutrient availability indices Some research has shown an inverse relationship between nutrient availability and
carbon allocation to roots in mature stands: the higher the nitrogen availability of a site, the lower the fine-root biomass (Keyes & Grier 1981; Linder & Rook 1984; Vogt et al. 1985). In this study, the importance of nutrient availability in also influencing the amount of conifer fine-root biomass in some stands is suggested by the positive relationship obtained between conifer root biomass and chemical quality of needle litterfall and forest- floor nitrogen content in the low-productivity stands. That these relationships did not exist in the high-productivity stands may be explained either by nutrient availability not being limiting for tree growth in these stands or factors other than litter chemical composition having a stronger influence on needle litter decay rates. When the chemical composition of litterfall does not limit tree growth or affect nutrient availability, other factors such as climate may have a greater influence on decomposition rates (Meente- meyer 1978).
The positive relationship between lignin and lignin:N ratio of needle litterfall and conifer fine-root biomass probably reflects the importance of litter chemical composition in influencing nutrient availability on poor sites. The initial lignin or nitrogen contents (or both) of litter have been identified as important factors in controlling litter decay rates (Minderman 1968; Fogel & Cromack 1977; Berg & Staaf 1980; Melillo, Aber & Muratore 1982; Pandey & Singh 1982; Berg & Ekbohm 1983).
A positive relationship was observed between forest-floor nitrogen content and root biomass for the nutrient poor sites of this study. This again reflects the increases in root biomass that may occur as nitrogen availability decreases. Slow decomposition rates of litter usually result in N accumulating in the forest floor because of low mineralization
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

868 Fine-root biomass in Douglas-fir forests
rates and, therefore, reduced nutrient availability (Turner 1977; Powers 1980; Vogt, Grier & Vogt 1986). Other studies have also shown N accumulating in the forest floor to be correlated with plant uptake of N (measured as foliar-N concentrations) and stand growth potential (Zottle 1960; Le Tacon 1972; Powers 1980; Peterson Ryan & Gessel 1984).
ACKNOWLEDGMENTS
This research was supported by the National Science Foundation. The iterative non- linear curve fitting program used was written by Dr Douglas Sprugel (University of Washington). We acknowledge Dr Loveday Conquest (University of Washington) for statistical consultation.
REFERENCES
Alaback, P. B. (1982). Dynamics of understorey biomass in Sitka spruce-western hemlock forests of southeast Alaska. Ecology, 63, 1932-1948.
Berg, B. & Ekbohm, G. (1983). Nitrogen immobilization in decomposing needle litter at variable carbon: nitro- gen ratios. Ecology, 64, 63-67.
Berg, B. & Staaf, H. (1980). Decomposition rate and chemical changes of Scots pine needle litter. II. Influence of chemical composition. Structure and Function of Northern Coniferous Forests-an Ecosystem Study (Ed by T. Persson). Ecological Bulletin, Stockholm, 32, 373-390.
Berish, C. W. (1982). Root biomass and surface area in three successional tropical forests. Canadian Journal of Forest Research, 12, 699-704.
Black, T. A. & Spittlehouse, D. L. (1981). Modelling the water balance for watershed management. Interior West Watershed Management (Ed by D. M. Baumgartner), pp. 117-129. Washington State University, Pullman.
Carlisle, A., Brown, A. H. F. & White, E. J. (1967). The nutrient contents of tree stem flow and ground flora litter and leachates in a sessile oak (Quercus petraea) woodlands. Journal of Ecology, 55, 615-627.
Daniel, T. W., Helms, J. A. & Baker, F. S. (1979). Principles of Silviculture, 2nd edn. McGraw-Hill Book Company, New York.
Davison, S. E. & Forman, R. T. T. (1982). Herb and shrub dynamics in a mature oak forest: a thirty-year study. Bulletiti of the Torrey Botanical Club, 109, 64-73.
Ericsson, A. & Persson, H. (1980). Seasonal changes in starch reserves and growth of fine roots of 20-year-old Scots pines. Structure and Function of Northern Coniferous Forests-an Ecosystem Study (Ed by T. Persson). Ecological Bulletin, Stockholm, 32, 239-250.
Fogel, R. & Cromack, K. Jr (1977). Effect of habitat and substrate quality on Douglas-fir litter decomposition in western Oregon, Canadian Journal of Botany, 55, 1632-1640.
Gessel, S. P. & Turner, J. (1976). Litter production in western Washington Douglas-fir stands. Forestry, 49,63- 72.
Gholz, H. L., Fitz, F. K. & Waring, R. H. (1976). Leaf area differences associated with old-growth forest communities in the western Oregon Cascades. Canadian Journal of Forest Research, 6, 49-57.
Gholz, H. L., Grier, C. C., Campbell, A. G. & Brown, A. T. (1979). Equations for estimating biomass and leaf area of plants in the Pacific Northwest. Forest Research Laboratory, Oregon State University, Corvallis, Oregon, Research Paper 41.
Grier, C. C. & Waring, R. H. (1974). Conifer foliage mass related to sapwood area. Forest Science, 20,205-206. Hough, W. A. (1982). Phytomass and nutrients in the understorey and forest floor of slash/longleaf pine stands.
Forest Science, 28, 359-372. Hull, C. H. & Nie, N. H. (1981). SPSS update 7-9. McGraw-Hill Book Company, New York. Karizumi, N. (1968). Estimation of root biomass in forests by the soil block sampling. Methods of Productivity
Studies in Root Systems and Rhizosphere Organisms (Ed by M. S. Ghilarow, V. A. Kovda, L. N. Novichkova-Ivanova, L. E. Rodin & V. M. Sveshnikova), pp. 79-86. Publishing House 'Nauka', Leningrad.
Kaufmann, M. R. & Troendle, C. A. (1981). The relationship of leaf area and foliage biomass to sapwood conducting area in four subalpine forest tree species. Forest Science, 27, 477-482.
Keyes, M. R. & Grier, C. C. (1981). Above- and below-ground net production in 40-year-old Douglas-fir stands on low and high productivity sites. Canadian Journal of Forest Research, 11, 599-605.
King, J. E. (1966). Site index curves for Douglas-fir in the Pacific Northwest. Weyerhaeuser Forestry Paper, 8, 1- 49.
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

K. A. VoGTetal. 869
Kira, T. & Shidei, T. (1967). Primary productivity and turnover of organic matter in different forest ecosystems of the western Pacific. Japanese Journal of Ecology, 17, 70-87.
Linder, S. & Rook, D. A. (1984). Effects of mineral nutrition on carbon dioxide exchange and partitioning of carbon in trees. Nutrition of Forest Trees in Plantations (Ed by G. D. Bowen, & E. K. S. Nambiar), pp. 21 1- 236. Academic Press, London.
Le Tacon, F. (1972). Disponibilite de l'azole nitrique et ammoniacal dans certains sol de l'est de la France. Influence sur la nutrition et la croissance de l'epicea commun (Picea abies Karst.). Annales des Sciences Forestieres, 30, 183-203.
Long, J. N., Smith, F. W. & Scott, D. R. M. (1981). The role of Douglas-fir stem sapwood and heartwood in the mechanical and physiological support of crowns and development of stem form. Canadian Journal of Forest Research, 11, 459-464.
MacLean, D. A. & Wein, R. W. (1977). Nutrient accumulation for postfire jack pine and hardwood succession patterns in New Brunswick. Canadian Journal of Forest Research, 7, 562-578.
MacLean, D. A. & Wein, R. W. (1978). Litter production and forest floor nutrient dynamics in pine and hardwood stands of New Brunswick, Canada. Holarctic Ecology, 1, 1-15.
Marshall, J. D. & Waring, R. H. (1986). Comparison of methods of estimating leaf-area index in old-growth Douglas-fir. Ecology 67, 975-979.
Martin, W. L., Sharik, T. L. & Smith, D. W. (1982). Phytomass and nutrient levels in the understory of an Appalachian oak forest in southwest Virginia. Bulletin of the Torrey Botanical Club, 109, 330-343.
McQueen, D. R. (1968). The quantitative distribution of absorbing roots of Pinus sylvestris and Fagus sylvatica in a forest succession. Oecologia Plantarum, 3, 83-99.
Meentemeyer, V. (1978). Macroclimate and lignin control of litter decomposition rates. Ecology, 59, 465- 472.
Melillo, J. M., Aber, J. D. & Muratore, J. F. (1982). Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology, 63, 621-626.
Minderman, G. (1968). Addition, decomposition and accumulation of organic matter in forests. Journal ol Ecology, 56, 355-362.
Nie, N. H., Hull, C. H., Jenkins, J. G., Steinbrenner, K. & Bent, D. H. (Eds) (1975). Statistical Package for the Social Sciences (SPSS). McGraw-Hill, Inc., New York.
Oliver, C. D. (1981). Forest development in North America following minor disturbances. Forest Ecology and Management, 3, 153-168.
Pandey, U. & Singh, J. S. (1982). Leaf-litter decomposition in an oak-conifer forest in Himalaya: the effects of climate and chemical composition. Forestry, 55, 47-59.
Parkinson, J. A. & Allen, S. E. (1975). A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Communications Soil Science and Plant Analysis, 6, 1-11.
Pearson, J. A., Fahey, T. J. & Knight, D. H. (1984). Biomass and leaf area in contrasting lodgepole pine forests. Canadian Journal of Forest Research, 14, 259-265.
Persson, H. (1980a). Fine-root dynamics in a Scots pine stand with and without near-optimum nutrient and water regimes. Acta Phytogeographica Suecica, 68, 10 1-1 10.
Persson, H. (1980b). Death and replacement of fine roots in a mature Scots pine stand. Structure and Function of Northern Coniferous Forests - an Ecosystem Study (Ed by T. Persson). Ecological Bulletin, Stockholm, 32, 251-260.
Persson, H. (1980c). Spatial distribution of fine-root growth, mortality and decomposition in a young Scots pine stand in central Sweden. Oikos, 34, 77-87.
Peterson, C. E., Ryan, P. J. & Gessel, S. P. (1984). Response of northwest Douglas-fir to urea: Correlations with forest soil properties. Soil Science Society of America Journal, 48, 162-169.
Powers, R. F. (1980). Mineralizable soil nitrogen as an index of nitrogen availability to forest trees. Soil Science Society of America Journal, 44, 1314-1320.
Roberts, J., Pymar, C. F., Wallace, J. S. & Pitman, R. M. (1980). Seasonal changes in leaf area, stomatal and canopy conductances and transpiration from bracken fern (Pteridium aquilinum (L.) Kuhn) below a forest canopy. Journal of Applied Ecology, 17, 409-422.
Santantonio, D. & Hermann, R. K. (1985). Standing crop, production, and turnover of fine roots on dry, moderate, and wet sites of mature Douglas-fir in western Oregon. Annales des Sciences Foresti&res, 42, 113- 142.
Singh, B. (1982). Nutrient content of standing crop and biological cycling in Pinus patula ecosystem. Forest Ecology and Management, 4, 317-332.
Snedecor, G. W. & Cochran, W. G. (1967). Statistical Methods. The Iowa State University Press, Ames, Iowa. Snell, J. A. K. & Brown, J. K. (1978). Comparison of tree biomass estimators-dbh and sapwood area. Forest
Science, 24, 455-457. Sprugel, D. G. (1985). Changes in biomass components through stand development in wave-regenerated balsam
fir forests. Canadian Journal of Forest Research, 15, 269-278. Tadaki, Y., Sato, A., Sakurai, S., Takeuchi, I. & Kawahara, T. (1977). Studies on the production structure of
Forest. XVIII. Structure and primary production in subalpine 'dead trees strips' Abies forest near Mt. Asahi. Japanese Journal of Ecology, 27, 83-90.
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions

870 Fine-root biomass in Douglas-fir forests
Tappi Standard (1974). Acid insoluble lignin in wood and pulp. T222 OS-74. Technical Association of Pulp and Paper Industry.
Turner, J. (1977). Effect of nitrogen availability on nitrogen cycling in a Douglas-fir stand. Forest Science, 23, 307-316.
Turner, J. & Long, J. N. (1975). Accumulation of organic matter in a series of Douglas-fir stands. Canadian Journal of Forest Research, 5, 681-690.
Vogt, K. A., Edmonds, R. L. & Grier, C. C. (1981). Seasonal changes in biomass and vertical distribution of mycorrhizal and fibrous-textured conifer fine roots in 23- and 180-year-old subalpine Abies amabilis stands. Canadian Journal of Forest Research, 11, 223-229.
Vogt, K. A., Grier, C. C., Meier, C. E. & Edmonds, R. L. (1982). Mycorrhizal role in net primary production and nutrient cycling in Abies amabilis ecosystems in western Washington. Ecology, 63, 370-380.
Vogt, K. A., Grier, C. C., Meier, C. E. & Keyes, M. R. (1983b). Organic matter and nutrient dynamics in forest floors of young and mature Abies amabilis stands in western Washington, as affected by fine-root input. Ecological Monographs, 53, 139-157.
Vogt, K. A., Grier, C. C. & Vogt, D. J. (1986). Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests. Advances in Ecological Research, 15, 303-377.
Vogt, K. A., Moore, E. E., Vogt, D. J., Redlin, M. J. & Edmonds, R. L. (1983a). Conifer fine root and mycorrhizal root biomass within the forest floors of Douglas-fir stands of different ages and site productivities. Canadian Journal of Forest Research, 13, 429-437.
Vogt, K. A., Vogt, D. J., Moore, E. E., Littke, W., Grier, C. C. & Leney, L. (1985). Estimating Douglas-fir fine root biomass and production from living bark and starch. Canadian Journal of Forest Research, 15, 177- 179.
Waring, R. H., Schroeder, P. E. & Oren, R. (1982). Application of the pipe-model theory to predict canopy leaf area. Canadian Journal of Forest Research, 12, 556-560.
Whitehead, D. (1978). The estimation of foliage area from sapwood basal area in Scots pine. Forestry, 51, 137- 149.
Yarie, J. (1980). The role of understory vegetation in the nutrient cycle of forested ecosystems in the mountain hemlock biogeoclimatic zone. Ecology, 61, 1498-1514.
Zottle, V. H. (1960). Die mineral stickstoffanlieferung in Fichten- und Kiefernbestanden Bayerns. Forstwissens- chaftiches Centralblatt, 79, 221-236.
(Received 17 September 1986)
This content downloaded from 205.175.116.80 on Wed, 26 Feb 2014 17:40:05 PMAll use subject to JSTOR Terms and Conditions
Related Documents











