Comparison of molecular techniques for the typing of Mycoplasma hyopneumoniae isolates Tim Stakenborg a, * , Jo Vicca b , Dominiek Maes b , Johan Peeters a , Aart de Kruif b , Freddy Haesebrouck b , Patrick Butaye a a Veterinary and Agrochemical Research Centre, Groeselenberg 99, 1180 Brussels, Belgium b Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium Received 5 September 2005; received in revised form 6 December 2005; accepted 7 December 2005 Available online 3 February 2006 Abstract In this study, we compared the potential of amplified fragment length polymorphism (AFLP), random amplified polymorphic DNA (RAPD) analysis, restriction fragment length polymorphism (RFLP) of the gene encoding lipoprotein P146, and the variable number of tandem repeats (VNTR) of the P97 encoding gene, as possible methods for typing an international collection of Mycoplasma hyopneumoniae isolates. All techniques showed a typeability of 100% and high intraspecific diversity. However, the discriminatory power of the different techniques varied considerably. AFLP (N 0.99) and PCR-RFLP of the P146 encoding gene (N 0.98) were more discriminatory than RAPD (0.95) and estimation of the VNTR of P97 (b 0.92). Other, preferentially well spread, tandem repeat regions should be included in order for this latter technique to become valuable for typing purposes. RAPD was also found to be a less interesting typing technique because of its low reproducibility between different runs. Nevertheless, all molecular techniques showed overall more resemblance between strains isolated from different pigs from the same herd. On the other hand, none of the techniques was able to show a clear relationship between the country of origin and the fingerprints obtained. We conclude that AFLP and an earlier described PFGE technique are highly reliable and discriminatory typing techniques for outlining the genomic diversity of M. hyopneumoniae isolates. Our data also show that RFLP of a highly variable gene encoding P146 may be an equally useful alternative for demonstrating intraspecific variability, although the generation of sequence variability of the gene remains unclear and must be further examined. D 2005 Elsevier B.V. All rights reserved. Keywords: Amplified fragment length polymorphism; Random amplified polymorphic DNA analysis; Restriction fragment length polymorphism; Variable number of tandem repeats; Mycoplasma hyopneumoniae 1. Introduction Mycoplasma hyopneumoniae is the primary cause of enzootic pneumonia in pigs. Although vaccines have been developed, infections are still hard to control (Maes et al., 1999) and, even in countries aiming to eradicate enzootic pneumonia, reinfections occur fre- quently (Hege et al., 2002). The disease is not associ- ated with a high mortality rate, but the severity may vary greatly between different herds (Vicca et al., 2003). Farm management is considered essential (Done, 1991), but also the intrinsic virulence of circu- lating M. hyopneumoniae strains has been proven to be an important cause for this variation (Vicca et al., 2003). The underlying mechanism to explain these 0167-7012/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.mimet.2005.12.002 * Corresponding author. Tel.: +32 2 3790437; fax: +32 2 3790690. E-mail address: [email protected] (T. Stakenborg). Journal of Microbiological Methods 66 (2006) 263 – 275 www.elsevier.com/locate/jmicmeth

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/jmicmeth
Journal of Microbiological Methods 6
Comparison of molecular techniques for the typing of
Mycoplasma hyopneumoniae isolates
Tim Stakenborg a,*, Jo Vicca b, Dominiek Maes b, Johan Peeters a,
Aart de Kruif b, Freddy Haesebrouck b, Patrick Butaye a
a Veterinary and Agrochemical Research Centre, Groeselenberg 99, 1180 Brussels, Belgiumb Faculty of Veterinary Medicine, Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium
Received 5 September 2005; received in revised form 6 December 2005; accepted 7 December 2005
Available online 3 February 2006
Abstract
In this study, we compared the potential of amplified fragment length polymorphism (AFLP), random amplified polymorphic
DNA (RAPD) analysis, restriction fragment length polymorphism (RFLP) of the gene encoding lipoprotein P146, and the variable
number of tandem repeats (VNTR) of the P97 encoding gene, as possible methods for typing an international collection of
Mycoplasma hyopneumoniae isolates. All techniques showed a typeability of 100% and high intraspecific diversity. However, the
discriminatory power of the different techniques varied considerably. AFLP (N0.99) and PCR-RFLP of the P146 encoding gene
(N0.98) were more discriminatory than RAPD (0.95) and estimation of the VNTR of P97 (b0.92). Other, preferentially well spread,
tandem repeat regions should be included in order for this latter technique to become valuable for typing purposes. RAPD was also
found to be a less interesting typing technique because of its low reproducibility between different runs. Nevertheless, all molecular
techniques showed overall more resemblance between strains isolated from different pigs from the same herd. On the other hand,
none of the techniques was able to show a clear relationship between the country of origin and the fingerprints obtained. We
conclude that AFLP and an earlier described PFGE technique are highly reliable and discriminatory typing techniques for outlining
the genomic diversity of M. hyopneumoniae isolates. Our data also show that RFLP of a highly variable gene encoding P146 may
be an equally useful alternative for demonstrating intraspecific variability, although the generation of sequence variability of the
gene remains unclear and must be further examined.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Amplified fragment length polymorphism; Random amplified polymorphic DNA analysis; Restriction fragment length polymorphism;
Variable number of tandem repeats; Mycoplasma hyopneumoniae
1. Introduction
Mycoplasma hyopneumoniae is the primary cause of
enzootic pneumonia in pigs. Although vaccines have
been developed, infections are still hard to control
(Maes et al., 1999) and, even in countries aiming to
0167-7012/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.mimet.2005.12.002
* Corresponding author. Tel.: +32 2 3790437; fax: +32 2 3790690.
E-mail address: [email protected] (T. Stakenborg).
eradicate enzootic pneumonia, reinfections occur fre-
quently (Hege et al., 2002). The disease is not associ-
ated with a high mortality rate, but the severity may
vary greatly between different herds (Vicca et al.,
2003). Farm management is considered essential
(Done, 1991), but also the intrinsic virulence of circu-
lating M. hyopneumoniae strains has been proven to be
an important cause for this variation (Vicca et al.,
2003). The underlying mechanism to explain these
6 (2006) 263–275
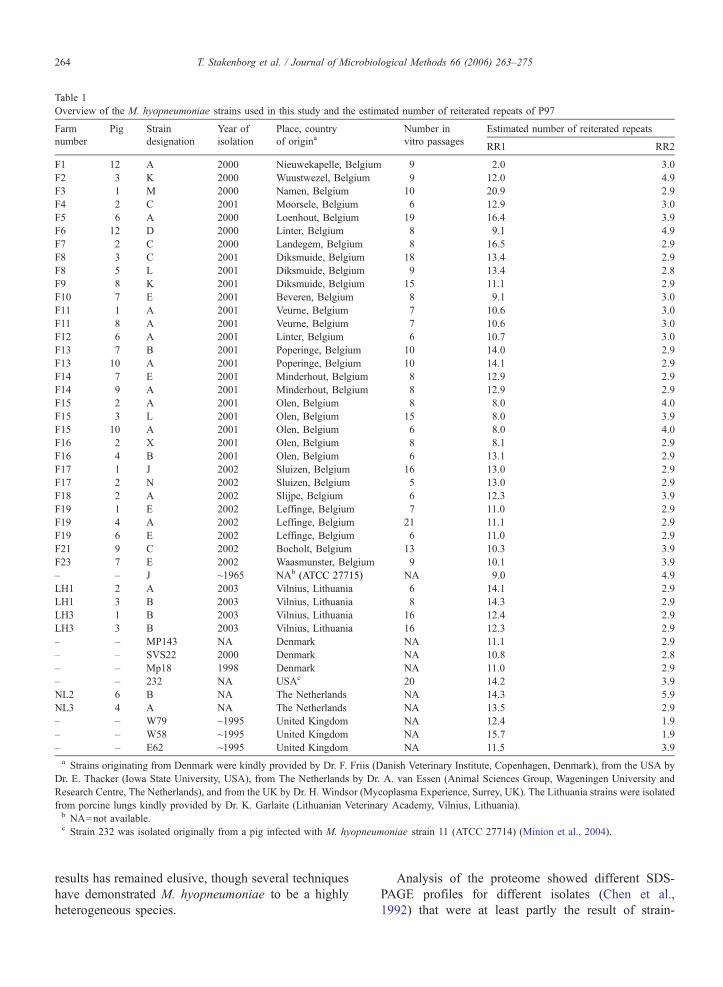
Table 1
Overview of the M. hyopneumoniae strains used in this study and the estimated number of reiterated repeats of P97
Farm Pig Strain Year of Place, country Number in Estimated number of reiterated repeats
number designation isolation of origina vitro passagesRR1 RR2
F1 12 A 2000 Nieuwekapelle, Belgium 9 2.0 3.0
F2 3 K 2000 Wuustwezel, Belgium 9 12.0 4.9
F3 1 M 2000 Namen, Belgium 10 20.9 2.9
F4 2 C 2001 Moorsele, Belgium 6 12.9 3.0
F5 6 A 2000 Loenhout, Belgium 19 16.4 3.9
F6 12 D 2000 Linter, Belgium 8 9.1 4.9
F7 2 C 2000 Landegem, Belgium 8 16.5 2.9
F8 3 C 2001 Diksmuide, Belgium 18 13.4 2.9
F8 5 L 2001 Diksmuide, Belgium 9 13.4 2.8
F9 8 K 2001 Diksmuide, Belgium 15 11.1 2.9
F10 7 E 2001 Beveren, Belgium 8 9.1 3.0
F11 1 A 2001 Veurne, Belgium 7 10.6 3.0
F11 8 A 2001 Veurne, Belgium 7 10.6 3.0
F12 6 A 2001 Linter, Belgium 6 10.7 3.0
F13 7 B 2001 Poperinge, Belgium 10 14.0 2.9
F13 10 A 2001 Poperinge, Belgium 10 14.1 2.9
F14 7 E 2001 Minderhout, Belgium 8 12.9 2.9
F14 9 A 2001 Minderhout, Belgium 8 12.9 2.9
F15 2 A 2001 Olen, Belgium 8 8.0 4.0
F15 3 L 2001 Olen, Belgium 15 8.0 3.9
F15 10 A 2001 Olen, Belgium 6 8.0 4.0
F16 2 X 2001 Olen, Belgium 8 8.1 2.9
F16 4 B 2001 Olen, Belgium 6 13.1 2.9
F17 1 J 2002 Sluizen, Belgium 16 13.0 2.9
F17 2 N 2002 Sluizen, Belgium 5 13.0 2.9
F18 2 A 2002 Slijpe, Belgium 6 12.3 3.9
F19 1 E 2002 Leffinge, Belgium 7 11.0 2.9
F19 4 A 2002 Leffinge, Belgium 21 11.1 2.9
F19 6 E 2002 Leffinge, Belgium 6 11.0 2.9
F21 9 C 2002 Bocholt, Belgium 13 10.3 3.9
F23 7 E 2002 Waasmunster, Belgium 9 10.1 3.9
– – J ~1965 NAb (ATCC 27715) NA 9.0 4.9
LH1 2 A 2003 Vilnius, Lithuania 6 14.1 2.9
LH1 3 B 2003 Vilnius, Lithuania 8 14.3 2.9
LH3 1 B 2003 Vilnius, Lithuania 16 12.4 2.9
LH3 3 B 2003 Vilnius, Lithuania 16 12.3 2.9
– – MP143 NA Denmark NA 11.1 2.9
– – SVS22 2000 Denmark NA 10.8 2.8
– – Mp18 1998 Denmark NA 11.0 2.9
– – 232 NA USAc 20 14.2 3.9
NL2 6 B NA The Netherlands NA 14.3 5.9
NL3 4 A NA The Netherlands NA 13.5 2.9
– – W79 ~1995 United Kingdom NA 12.4 1.9
– – W58 ~1995 United Kingdom NA 15.7 1.9
– – E62 ~1995 United Kingdom NA 11.5 3.9
a Strains originating from Denmark were kindly provided by Dr. F. Friis (Danish Veterinary Institute, Copenhagen, Denmark), from the USA by
Dr. E. Thacker (Iowa State University, USA), from The Netherlands by Dr. A. van Essen (Animal Sciences Group, Wageningen University and
Research Centre, The Netherlands), and from the UK by Dr. H. Windsor (Mycoplasma Experience, Surrey, UK). The Lithuania strains were isolated
from porcine lungs kindly provided by Dr. K. Garlaite (Lithuanian Veterinary Academy, Vilnius, Lithuania).b NA=not available.c Strain 232 was isolated originally from a pig infected with M. hyopneumoniae strain 11 (ATCC 27714) (Minion et al., 2004).
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275264
results has remained elusive, though several techniques
have demonstrated M. hyopneumoniae to be a highly
heterogeneous species.
Analysis of the proteome showed different SDS-
PAGE profiles for different isolates (Chen et al.,
1992) that were at least partly the result of strain-

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 265
specific post-translational modifications (Djordjevic
et al., 2004). At the genomic level, an enormous hete-
rogeneity has been demonstrated by various typing
techniques such as random amplified polymorphic
DNA (RAPD) (Artiushin and Minion, 1996), amplified
fragment length polymorphism (AFLP) (Kokotovic
et al., 1999), and pulsed-field gel electrophoresis
(PFGE) (Stakenborg et al., 2005). Moreover, the num-
ber of a yet unassigned insertion-like sequence varied
between different strains (Harasawa et al., 1995) and
differences in the reiterated regions of a P97 adhesin
encoding gene have been reported for different iso-
lates (Hsu et al., 1997; Hsu and Minion, 1998b). All
these different techniques may prove useful in future
epidemiological studies to trace strains or to visualize
infection patterns. To perform such epidemiological
studies, the choice of the typing technique is essential.
However, in the case of M. hyopneumoniae, the value
of different typing techniques has never been assessed.
As long as whole genome sequencing is not easily
attainable, typing techniques, which ideally represent
the true phylogenetic relation between strains, are
bound to their own intrinsic limitations. In this study,
we compared the use of formerly described techniques
(RAPD and AFLP) and newly PCR-based techniques
(PCR-RFLP of the P146 gene and the VNTR of the
P97 gene) as possible methods for studying the diver-
sity of M. hyopneumoniae strains. For each typing
technique, the discriminatory power, reproducibility
and ease of performance were compared using an iden-
tical set of strains. The results obtained were discussed
in detail and compared with PFGE data on a similar set
of isolates described earlier by our group (Stakenborg
et al., 2005).
2. Materials and methods
2.1. Bacterial isolates, media, and DNA extraction
A total of 43 M. hyopneumoniae isolates were used,
together with reference strains J (ATCC 25934), USA
232 (Minion et al., 2004), and Mycoplasma flocculare
Ms42 (ATCC 27399). All Belgian and Lithuanian field
isolates were derived from lung samples of pigs at
slaughter. These isolates received a name of the format
dF1.2AT, where F1 represents the number of the herd,
2 indicates the number of the pig and A is an arbitrary
letter representing the isolate. Isolates from different
pigs from the same herd were obtained from lung
samples collected at the same moment. For strains
that were received from other laboratories, the genuine
strain designation was kept unchanged. For reasons of
clarity, the international code representing the country
of origin was always indicated between parentheses
after the isolate’s name. Further information about the
included strains is listed in as much detail as possible
in Table 1.
Friis’ broth was used to grow both the M. flocculare
and the M. hyopneumoniae strains (Friis, 1975).
Purified, genomic DNA was prepared using a phenol/
chloroform extraction method (Bashiruddin, 1998).
2.2. RAPD
For RAPD analyses, 45 cycles (1V94 8C; 1V36 8C;and 2V72 8C) were run on a GeneAmp 9600 Thermal
Cycler (Perkin Elmer, Ma, USA) using 20 pmol of
a primer OPA-3 (5V AGTCAGCCAC) described by
Artiushin and Minion (1996), and exactly 30 ng of
purified, genomic DNA as a template. To minimize
the variability between different runs, Ready-To-Go
RAPD Beads (Amersham Biosciences, Germany)
were used and all samples were run simultaneously
during one single PCR. After amplification, 10 Al ofthe PCR mixture were analyzed by electrophoresis
(120 V, 90V) on 1% agarose gel (Sigma, UK). The
DNA fragments were visualized using a GeneGenius
gel documentation system (Westburg, The Netherlands)
and exported to Bionumerics (V3.5, Applied-Maths,
Belgium) for further analysis. Bands annotated by the
software were visually controlled and fragments smaller
than 500 bp were omitted for further analysis. Calcu-
lation of similarity coefficients was performed using the
Dice algorithm. The unweighted pair group method
with arithmetic means (UPGMA) was used for cluster-
ing with a band position tolerance and optimization
setting of 1%.
2.3. AFLP
AFLP was performed in similarity to an earlier
report (Kokotovic et al., 1999). Briefly, 200 ng ge-
nomic DNA was diluted in 20 Al restriction buffer
(SuRE/Cut Buffer M, Roche, Switzerland) and restrict-
ed with 10 U BglII (Roche) and 10 U MfeI (Fermentas,
Lithuania) for 3 h at 37 8C. After incubation for 15Vat65 8C, a 20 Al ligation reaction was set up using 5 Al ofthe digested DNA, 2 pmol of the BGL adapter, 20 pmol
of the MFE adapter (Kokotovic et al., 1999), 1 U T4
ligase (Amersham), 2 Al restriction buffer (Amersham),
and 8 Al restriction buffer (Amersham). Ligation was
carried out overnight at 16 8C. The succeeding ampli-
fication reaction was performed as noted in Table 2
using 2 Al of the 10-fold diluted ligation product as

Table 2
Primers and cycle conditions used in this study
Target sequence Sequence (5VY3V) Number of cycles (cycle conditions)a
Primer
AFLP PCRb 30 (1V 94 8C; 1V 54 8C; and 90W 72 8C)BGL-2F* (D4*)GAGTACACTGTCGATCT
MFE-1 GAGAGCTCTTGGAATTG
P146 (pre-amplification) 30 (15W 94 8C; 30W 51 8C; and 1V72 8C)P146 cFOR CATTAGTAACAGCAACAGCCATTG
P146 cREV TACCTCGCCGCCTTAGCAG
P146 (amplification) 25 (15W 94 8C; 30W 52,5 8C; and 1V72 8C)P146 FOR TTAGTAACAGCAACAGCCATTG
P146 REV CCCTTAAGTGGACAATTTTAGC
P97 (repeat region 1) 30 (30W 94 8C; 30W 53,7 8C; and 1V72 8C)RR1 FOR GAAGCTATCAAAAAAGGGGAAACTA
RR1 REV GGTTTATTTGTAAGTGAAAAGCCAG
P97 (repeat region 2) 30 (1V94 8C; 1V50,3 8C; and 45W 72 8C)RR2 FOR AGCGAGTATGAAGAACAAGAA
RR2 REV TTTTTACCTAAGTCAGGAAGG
a All PCRs, unless stated otherwise (see footnote b of this table), were performed using 3U of recombinant Taq polymerase (Invitrogen), 5 Al ofPCR buffer (Invitrogen) including 2 mM MgCl2, 0.2 mM of each dNTP and 10 pmol of both forward and reverse primer.b The AFLP-PCR was performed using 2U of AmpliTaq polymerase (Amersham Biosciences), 5 Al of PCR buffer II (Amersham Biosciences)
supplemented with 2.5 mM MgCl2, 0.2 mM of each dNTP and 10 pmol of both forward and reverse primer.
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275266
template. One Al of the amplified PCR products was
diluted in 40 Al sample loading solution (Beckman,
UK) supplemented with CEQ Size-standard 600 (Beck-
man) and run on a CEQ8000 Genetic Analysis System
(Beckman) for separation and visualization. The raw
data obtained were subsequently exported to Bionu-
merics (Applied-Maths) and converted to gel images.
After normalization, fragments between 60 and 560 bp
were defined. Clustering analysis of the fingerprints
obtained was performed with UPGMA on the basis
of a similarity matrix with calculated Jaccard’s similar-
ity coefficients. For clustering, the tolerance and opti-
misation level was set to 0.7%.
To determine the reproducibility of the AFLP pro-
cedure, three independent DNA samples of 10 arbitrari-
ly chosen strains were analyzed on different days.
2.4. PCR-RFLP analysis of the P146 encoding gene
For the amplification of the P146 gene, a PCR was
performed using primers and reaction conditions noted
in Table 2. Some strains yielded a faint non-specific
PCR fragment of about 900 bp in size, and for these
strains the PCR was repeated in nested format using
a pre-amplification step as noted in Table 2. After
PCR, about 100 ng of the final PCR product was
digested for 3 h at 37 8C in restriction buffer (SuRE/
Cut Buffer A, Roche) containing 10 U of restriction
enzyme AluI (Roche). Restricted fragments were
separated for 2 h at 120 V on a 2% Nusieve agar
(Cambrex Bioscience). The 50 bp O’RangeRuler (Fer-
mentas) was used as size standard and was loaded at
least twice for every 10 samples. After electrophoresis,
DNA fragments were visualized using a GeneGenius
gel documentation system (Westburg). The digital
images were exported to Bionumerics (Applied-
Maths) for standardization and annotation of the
bands. Fragments smaller than 175 bp were omitted
from the analysis. Levels of similarity between finger-
prints were calculated employing the Dice algorithm. In
order to attain a complete match between strains ana-
lyzed in duplicate, the tolerance and optimization levels
were both set to 1%. Cluster analysis was performed
with UPGMA.
To verify the accuracy of the technique, the observed
in vitro restriction patterns of isolates F7.2C and USA
232 were compared with those calculated in silico
based on the P146 gene sequences (see further). To
check whether in vitro cultivation influenced the
results, the test was repeated on three strains after 5,
10, and 15 in vitro subcultivation steps.
2.5. VNTR present in the P97 encoding gene
Two different reiterated repeat regions (RR1 and
RR2) have been described for the P97 adhesin gene
of M. hyopneumoniae (Hsu et al., 1997). For each
strain, two PCRs were performed to selectively amplify

Table 3
The by PCR estimated number of RR repeats of the P97 gene of strain USA232 and 10 arbitrarily chosen isolates compared with the actual number
of RR repeats determined by sequence analysis
Strain RR1 RR2
Estimated
length
Actual
length
Estimated
RR1 repeatsaActual
RR1 repeats
Number
TN repeatsbEstimated
length
Actual
length
Estimated
RR2 repeatscActual
RR2 repeats
F1.12K 184 185 1.9 2 3 284 284 3.0 3
F5.6B 403 401 16.5 16 4 312 314 3.0 3
F6.12D 292 288 9.1 8 5 341 344 3.9 4
F7.2C 399 397 16.3 15 6 282 284 4.9 5
F9.8K 322 320 11.1 11 3 280 284 2.9 3
F12.6A 316 320 10.7 11 3 285 284 3.0 3
F13.7B 365 368 14.0 13 6 282 284 2.9 3
F15.2A 275 275 8.0 8 3 317 314 4.1 4
J 290 290 9.0 9 3 340 344 4.9 5
MP143 322 317 11.1 11 3 280 284 2.9 3
USA232 368 368 14.2 15 1 312 314 3.9 4
a The estimated number of RR1 repeats was calculated assuming 3 TN repeats (i.e., the estimated length of the PCR product minus 155 bp and
divided by the 15 bp of one repeat unit).b The RR1 repeat region is followed by a repeat region of format GCT(ACTAAT)nACT on sequence level (or A(TN)nT on amino acid level), in
which dnT represents a number between 1 and 6.c The estimated number of RR2 repeats is determined by subtracting 194 (i.e., the number of amplified base pairs not included in the repeat
region) from the estimated length of the amplified PCR product and dividing this result by 30 (i.e., the length of one reiterated repeat unit).
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 267
the RR regions using the primers and cycle conditions
stated in Table 2. Amplified fragments were separated
on a 2% Nusieve agar (Cambrex Bioscience) for 2 h at
120V and visualized using a GeneGenius gel documen-
tation system (Westburg). Based on a 50 bp O’Ran-
geRuler (Fermentas), which was loaded at least twice
for every 10 samples, the sizes of amplified fragments
were estimated using Kodak digital science 1D soft-
ware (V3.0, Kodak Company, NY, USA). The accuracy
of the technique for estimation of the number of repeats
was examined by sequence analysis of the repeat
regions for 10 arbitrarily chosen isolates (see further).
In addition, the standard deviation of the technique was
calculated by comparing the expected size of the am-
plified RR2 region (i.e., number of RR2 repeats times
30 bp plus 194 bp) with the size of the amplification
products observed on gel. To check whether the strains
could be safely grown in the laboratory, the number of
repeats was compared for three arbitrarily chosen
strains that were subcultivated 5, 10, and 15 times in
vitro.
2.6. Discriminatory power
The Simpson’s index of diversity was calculated for
each technique (Hunter and Gaston, 1988). Since the
dependency between isolates originating from a single
herd was unknown, two different discriminatory
indexes were calculated, one including all M. hyopneu-
moniae strains and one excluding all isolates that had
an identical fingerprint and originated from a single
herd. This implies that in the event some isolates of
the same herd represent an identical clone, the true
value of the Simpson’s index of diversity should fall
between these two estimates.
2.7. Sequence analysis
Sequencing of the gene encoding lipoprotein P146
of strain F7.2C and part of the genes encoding P97 of
10 arbitrarily chosen isolates (Table 3) was performed
on PCR products. The samples were purified with
QIAquick spin columns (Qiagen, Germany) and se-
quenced on a CEQ8000 Genetic Analysis System
(Beckman, UK) by using the Quickstart kit (Beckman)
according to the manufacturer’s instructions. The
sequences obtained were exported to VectorNTI (V9,
Informax, Invitrogen) for assemblage and further anal-
ysis. The sequence of the P146 gene of isolate F7.2C
was submitted to Genbank (accession nr. DQ088147).
3. Results
3.1. RAPD
Since the observed RAPD patterns were not repro-
ducible between different runs (data not shown), even
not with the use of an as much as possible standardised
method, the analysis of all samples was carried out
during one single run. A limited number of fragments

Fig. 1. RAPD patterns of theM. hyopneumoniae isolates and theM. flocculare strain Ms42. Cluster analysis was performed with UPGMA using the
Dice coefficient and a tolerance and optimization level of 1%. Bands below 500 bp were omitted for analysis.
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275268

Fig. 2. Dendrogram of the obtained AFLP fragments from 60 to 560 bp in size. Cluster analysis was performed with UPGMA using the Jaccard’s
coefficient and a tolerance and optimization level of 0.7%. The dashed line represents the cut-off value (92%) for similarity determined by analysis
of replicates. Patterns with a higher similarity value are considered indistinguishable. The M. flocculare strain Ms42 that is included served as
an outgroup.
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 269

Fig. 3. PCR-RFLP patterns of the P146 gene the M. hyopneumoniae isolates. Cluster analysis was performed with UPGMA using the Dice
coefficient and a tolerance and optimization level of 1%. Bands below 175 bp were omitted from the analysis.
T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275270

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 271
(two to eight) were observed for each isolate. All iso-
lates showed a band of about 1300 bp in size and for
most M. hyopneumoniae isolates another band of about
550 bp was observed. For the M. flocculare Ms42
strain, an intense band of about 750 bp was observed,
but it could not be used for species differentiation as
fragments of a similar size were observed for some
Danish M. hyopneumoniae field isolates as well
(Fig. 1). The intensity of many bands between non-
identical patterns varied and complicated the analysis.
Isolates originating from the same herd had identical
RAPD patterns, with the exception of isolates from
herds F8 (BE), F16 (BE), F19 (BE) and LH3 (LT).
On the other hand, many strains originating from dif-
ferent herds also had identical profiles, resulting in
a discriminatory index of 0.95 (for both calculated
indexes).
3.2. AFLP
All M. hyopneumoniae isolates generated about 100
clearly separated fragments, with the exception of the
isolates of herd F19 (BE), F21 (BE) and F23 (BE),
which showed more bands, and of the isolate F2.3K
(BE), which showed considerably fewer bands. The M.
flocculare Ms42 strain showed a clearly different and
less complex pattern and formed the root of the den-
drogram. Reproducibility tests showed similar peak
profiles, though peak intensities often varied. After
normalization and band annotation, all replicates
showed similarity values of at least 92% (data not
shown). This value was used as a cut-off value to
differentiate between isolates. Despite this cut-off
value, only the multiple isolates originating from herd
F17 (BE), F19 (BE), and LH1 (LT) were indistinguish-
able. The AFLP patterns of F21.9C (BE) and F23.7E
(BE) were also considered identical. All other
isolates had similarity values below the cut-off value
(Fig. 2). This corresponded to a discriminatory index
that was calculated to be higher than 99% (both with
and without the inclusion of multiple isolates of the
same herd).
3.3. PCR-RFLP analysis of the P146 encoding gene
Restriction analysis with AluI showed an extensive
variation in the P146 gene of different isolates. This
variation was further illustrated by the high Simpson’s
index of diversity, which was calculated to be higher
than 0.98 without and higher than 0.97 with the inclu-
sion of isolates originating from the same farm. In
contrast to this enormous variation, isolates originating
from the same herd had identical profiles in 6 out of 10
cases (Fig. 3). The restriction profiles of three strains
that were subcultured up to 15 times in the laboratory
were also identical (data not shown).
Restriction patterns calculated in silico for the
determined sequence of the P146 encoding gene of
strain F7.2K (BE) and strain USA232 (Minion et al.,
2004) corresponded to those observed on gel. Upon
comparison of the two DNA sequences, several highly
variable repeat regions were observed, mainly in the
C-terminal part of the gene. These regions included
a poly-serine chain of variable length, a repeat region
rich in proline and glutamine residues of variable
length that could be represented by the following
format [Q]n[(P/S)Q]m, and a variable poly-alanine
chain situated directly before the stop codon of the
gene.
3.4. VNTR present in the P97 encoding gene
Both the RR1 (15 bp in length) and the RR2
repeat (30 bp in length) have been described in detail
before (Hsu et al., 1997; Wilton et al., 1998). As
shown in Table 1, the estimated number of RR1
repeats ranged for most strains from 8 to 16 copies.
However, two extremes were noted, isolate F1.12
(BE) with only 2 copies, and isolate F3.1M (BE)
with 21 copies of the RR1 repeat. The number of
RR2 repeats was less diverse and ranged from 2 to
6 copies, with most isolates having 3 copies. The
number of repeat regions of isolates originating from
the same herd was identical, except for the two
isolates of herd F16 (BE) where a difference between
the number of RR1 repeats was noted. This resulted
in a discriminatory power as low as 0.90 for RR1
and 0.59 for RR2 when excluding replicates, or 0.88
for RR1 and 0.53 for RR2 when including all isolates
of the same herd as well. When the two repeats were
combined, the discriminatory power rose to 0.91 with
and 0.94 without the inclusion of multiple isolates
per herd.
The calculated standard deviation of the estimated
size was 3.0 bp, while the maximum error observed
between the expected fragment size and the size de-
termined on gel was 7 bp. Therefore, the technique
can be used to exactly determine the number of RR2
repeats. In the event of RR1 repeats, and as verified
by sequence analyses, the copy number is merely an
estimate of the true value. Apparently, RR1 repeats are
followed by another repeat region of the format
GCT(ACTAAT)nACT, where n represents a number
from 1 to 6 (Table 3). Since the region consists of

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275272
repeated threonine and asparagine residues, the repeat
is further referred to as a TN repeat.
The number of repeats did not appear to change
easily over in vitro passages, since bands of identical
size were observed for three strains subcultivated 5, 10
and 15 times in vitro (data not shown).
4. Discussion
In this study, M. hyopneumoniae isolates were
differentiated by several typing techniques, including
some newly proposed techniques and some already
described by other authors (Artiushin and Minion,
1996; Kokotovic et al., 1999; Wilton et al., 1998).
Though multi-locus sequence typing (MLST) has been
proposed as a key technique for typing and character-
izing strains of many bacterial species (Urwin and
Maiden, 2003), it has not been worked out in detail
for M. hyopneumoniae. Moreover, MLST is still
rather expensive to be used routinely (Olive and
Bean, 1999) and other molecular typing techniques
may be favorable. However, until this study, these
different techniques had never been compared to each
other for the typing of M. hyopneumoniae. Based on
the results described here, we conclude that all the
different techniques typed all strains, but they
showed differing levels of discriminatory power and
reproducibility.
RAPD has been used as an easily performable and
highly discriminatory test to type strains of many
Mycoplasma species (Rawadi, 1998). In our study, as
well, the discriminatory power was satisfactory, despite
the relatively low number of fragments using the primer
described. On the other hand, RAPD lacked reproduc-
ibility, which made the comparison with new isolates
possible only by reanalyzing all isolates again in a
single experiment. Such a low reproducibility has
been described before (e.g., Penner et al., 1993; Van
Looveren et al., 1999) and even for a species like
Mycoplasma gallisepticum, where RAPD has often
been successfully used for epidemiological studies,
variation between gels and different runs has been
reported (Hong et al., 2005). Contrary to RAPD,
AFLP yielded more complex banding patterns and
was much more reproducible. Although AFLP was
reported fully reproducible for mycoplasmas (Kokoto-
vic et al., 1999), in our study the conversion of AFLP
patterns to gel images and subsequent analysis in Bio-
numerics (Applied-Maths) yielded similarity values of
92% or higher for replicates analyzed on different days.
Similar cut-off values have been reported for several
other bacterial species (e.g., Duim et al., 1999; van
Eldere et al., 1999). This did not influence the discrim-
inatory power of the technique (N99%).
As demonstrated earlier (Stakenborg et al., 2005),
PFGE also proved to be a reproducible and highly
discriminatory typing technique. The use of two differ-
ent restriction enzymes, SalI and ApaI, was evaluated
and yielded complementary results. The Simpson’s
index of diversity for this technique was calculated to
be at least as high as 0.98, which is comparable
to AFLP.
In this study, we additionally describe some molec-
ular techniques that have never been evaluated for the
typing M. hyopneumoniae. The VNTR of the P97
encoding gene were assessed. Compared to the other
methods, the estimation of the number of repeats in
the P97 encoding gene may be a fast and easily per-
formable technique. Since it is PCR based, theoretically
no culture steps are necessary and the technique may
give an initial indication about possible variation be-
tween two strains. However, a major drawback of
the technique is its low discriminatory power. Even
when including the combined data of the two repeats,
the discriminatory power merely rose above 0.91.
Moreover, the number of repeats can abruptly change
and more similar repeat units, preferably well-spread
over the genome, should be included before any pos-
tulations about the relation between strains can be
made. With the raising number of fully sequenced
genomes and revelation of new regions with tandem
repeats, VNTR typing has been evaluated for and ap-
plied to several bacterial species (e.g., Ramisse et al.,
2004; Scott et al., 2005; Yazdankhah et al., 2005). This
technique may be especially useful for mycoplasmas
in general, which are fastidious to cultivate and carry
many repeats (Rocha and Blanchard, 2002). However,
in the case of M. hyopneumoniae, the number of
genes containing tandem repeat regions appears to be
limited (Minion et al., 2004). Moreover, before setting
up a VNTR typing scheme, the stability must be firmly
validated since, in contrast to our results for P97, many
mycoplasma-specific proteins were reported to change
between different passages (Rosengarten and Yogev,
1996).
Despite the low discriminatory power, the use of P97
repeats in typing may be an indication of the colonizing
capacities of the isolates. A direct link between the
number of RR1 tandem repeats and adhesion has
been demonstrated and at least seven RR1 repeats
seem to be necessary to allow a strain to adhere to
sodium dodecyl sulfate-solubilized porcine tracheal
cells in vitro (Hsu and Minion, 1998a). Strain F1.2A
(BE) only contained two RR1 repeat regions. Still, the

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 273
isolate appears to be able to colonize the respiratory
tract, as it was isolated from a lung sample collected in
the slaughterhouse and it was able to cause lesions in an
experimental study (Vicca et al., 2003). This might
indicate that, besides P97 repeats, other colonization
factors may be present on M. hyopneumoniae strains.
The P146 lipoprotein of M. hyopneumoniae
shows a strong homology to the LppS lipoprotein of
Mycoplasma conjunctivae, which was shown to be
involved in in vitro adhesion (Belloy et al., 2003). In
addition, the N-terminus region of P146 also shows a
strong homology to the P97 adhesin (Minion et al.,
2004) and possesses a strong hydrophobic region
(amino acid 7-29), indicating a transmembrane region
and suggesting that the protein is expressed on the
surface of M. hyopneumoniae cells. The enormous
intraspecific diversity shown for the P146 encoding
gene was at least partly the result of differences be-
tween several repeat regions present in the gene, most
notably a polyserine chain of variable length and a
[Q]n[(P/S)Q]m repeat region. Polyserine chains often
function as a spacer region in proteins involved in
complex carbohydrate degradation (Howard et al.,
2004), while sequences rich in both proline and gluta-
mine are not uncommon and can form a conformation
known as a polyproline II helix (Kay et al., 2000;
Rucker and Creamer, 2002). Such proline rich
sequences are often involved in binding processes and
are highly immunogenic (Kay et al., 2000). Interesting-
ly, Bencina et al. (2001) hypothesized a correlation
between the length of a proline-rich region in the
pvpA gene of Mycoplasma synoviae and virulence.
Still, as long as the function of the P146 protein remains
unknown, correlations with virulence or adhesion are
speculative and need further investigation.
In this study, the presence of size-variable regions
increased the discriminatory power of the P146 PCR-
RFLP technique, which turned out to be very high
(N0.98). Moreover, the typing technique is reproducible
and easy to perform. Similar to estimation of the VNTR
of P97, the technique is PCR-based and thus in princi-
ple does not need pure cultures. The most important
drawback for epidemiological studies is the limited
genomic region under investigation. The molecular
clock of the gene (i.e., the rate at which mutations
are included) is unknown and does not appear to be
constant over the entire gene. The presence of the
highly variable regions seems in sharp contrast with
the observed stability of the gene after several in vitro
passages and with the fact that several isolates of
the same herd but of different pigs had identical restric-
tion patterns.
Apart from differences in reproducibility, complex-
ity and discriminatory power, the different techniques
yielded largely different clusters. Multiple isolates
of herds LH1 (LT), F15 (BE) and F17 (BE) were
indistinguishable with all described typing techniques,
including the above described PFGE analyses (Staken-
borg et al., 2005), and likely represent a single clone. In
addition, isolates F21.9C (BE) and F23.7E (BE) were
considered identical for all tests, although the farms are
from different geographical locations. Interestingly, the
different isolates of F16 were largely diverse for all
techniques applied in this study, while they appeared
almost identical on the basis of PFGE analysis (Staken-
borg et al., 2005).
Although most techniques clustered the other iso-
lates of the same farm in close proximity of each other,
not one technique uniquely clustered isolates according
to their geographical origin. Since an extensive vari-
ability was observed even for small geographical
regions, an association between country and isolate is
unlikely, even by including more international isolates.
This complicates the comparison of different methods,
since validation of typing data is most effective when it
is based on profound epidemiological knowledge and a
known phylogenetic relationship between isolates. In
the case of M. hyopneumoniae, such a relationship
between strains is hard to attain. Isolates are abundantly
present in nature and highly diverse, and they often
cause infections that remain subclinical. In order to
include at least some closely related strains in our
study, multiple isolates originating from the same
herd were used. Our data do indeed indicate that mainly
one clone is circulating at a specific point in time in a
single herd. It may be interesting to investigate the
variability of isolates of the same farm over a longer
period of time to better understand and to model the
transmission patterns of M. hyopneumoniae clones. As
demonstrated, various typing techniques with high dis-
criminatory power are available to perform such stud-
ies, although the use of more than one technique may
be essential to detect differences between closely
related strains.
In conclusion, the typing of M. hyopneumoniae can
easily be performed with high discriminatory power
and reproducibility by restriction analysis of the highly
variable gene encoding a P146 lipoprotein. However,
the limited region under investigation may be insuffi-
cient to visualize many genomic rearrangements.
Therefore, AFLP or PFGE, although much more labor
intensive, may be preferred. RAPD lacks reproducibil-
ity, while determination of the number of repeats in the
gene encoding P97 may only be used if other epidemi-

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275274
ological markers are included as well. The different
techniques described are useful for modeling the epi-
demiology of M. hyopneumoniae, which may be help-
ful for developing precautionary measures in order to
control enzootic pneumonia in the future. Finally, a
typing scheme containing P97 VNTR analysis and
P146 variability may be valuable for direct application
on clinical samples and might provide information on
the adherence capacities or virulence of the strains. This
should be further investigated, however.
Acknowledgements
This study was supported by a grant from the Fed-
eral Public Service Health, Food Chain Safety and
Environment (Grant number S-6136).
The authors are grateful to Dr. F. Friis, Dr. E.
Thacker, Dr. A. van Essen, Dr. H. Windsor and Dr. K.
Garlaite for providing us with international M. hyop-
neumoniae isolates or lung samples. The authors would
also like to thank Sara Tistaert for her skilful techni-
cal assistance.
References
Artiushin, S., Minion, F.C., 1996. Arbitrarily primed PCR analysis of
Mycoplasma hyopneumoniae field isolates demonstrates genetic
heterogeneity. Int. J. Syst. Bacteriol. 46, 324–328.
Bashiruddin, J.B., 1998. Extraction of DNA from mycoplasmas.
Methods Mol. Biol. 104, 141–144.
Belloy, L., Vilei, E.M., Giacometti, M., Frey, J., 2003. Characteriza-
tion of LppS, an adhesin of Mycoplasma conjunctivae. Microbi-
ology 149, 185–193.
Bencina, D., Drobnic-Valic, M., Horvat, S., Narat, M., Kleven, S.H.,
Dovc, P., 2001. Molecular basis of the length variation in the N-
terminal part of Mycoplasma synoviae hemagglutinin. FEMS
Microbiol. Lett. 203, 115–123.
Chen, J.W., Zhang, L., Song, J., Hwang, F., Dong, Q., Liui, J.,
Qian, Y., 1992. Comparative analysis of glycoprotein and gly-
colipid composition of virulent and avirulent strain membranes
of Mycoplasma hyopneumoniae. Curr. Microbiol. 24, 189–192.
Djordjevic, S.P., Cordwell, S.J., Djordjevic, M.A., Wilton, J., Minion,
F.C., 2004. Proteolytic processing of the Mycoplasma hyopneu-
moniae cilium adhesin. Infect. Immun. 72, 2791–2802.
Done, S.H., 1991. Environmental factors affecting the severity of
pneumonia in pigs. Vet. Rec. 128, 582–586.
Duim, B., Wassenaar, T.M., Rigter, A., Wagenaar, J., 1999. High-
resolution genotyping of Campylobacter strains isolated from
poultry and humans with amplified fragment length polymor-
phism fingerprinting. Appl. Environ. Microbiol. 65, 2369–2375.
Friis, N.F., 1975. Some recommendations concerning primary isola-
tion of Mycoplasma suipneumoniae and Mycoplasma flocculare a
survey. Nord. Vet. Med. 27, 337–339.
Harasawa, R., Asada, K., Kato, I., 1995. A novel repetitive
sequence from Mycoplasma hyopneumoniae. J. Vet. Med. Sci.
57, 557–558.
Hege, R., Zimmermann, W., Scheidegger, R., Stark, K.D., 2002.
Incidence of reinfections with Mycoplasma hyopneumoniae and
Actinobacillus pleuropneumoniae in pig farms located in respira-
tory-disease-free regions of Switzerland. Identification and quan-
tification of risk factors. Acta Vet. Scand. 43, 145–156.
Hong, Y., Garcia, M., Levisohn, S., Savelkoul, P., Leiting, V.,
Lysnyansky, I., Ley, D.H., Kleven, S.H., 2005. Differentiation
of Mycoplasma gallisepticum strains using amplified fragment
length polymorphism and other DNA-based typing methods.
Avian Dis. 49, 43–49.
Howard, M.B., Ekborg, N.A., Taylor, L.E., Hutcheson, S.W., Weiner,
R.M., 2004. Identification and analysis of polyserine linker
domains in prokaryotic proteins with emphasis on the
marine bacterium Microbulbifer degradans. Protein Sci. 13,
1422–1425.
Hsu, T., Minion, F.C., 1998a. Identification of the cilium binding
epitope of the Mycoplasma hyopneumoniae P97 adhesin. Infect.
Immun. 66, 4762–4766.
Hsu, T., Minion, F.C., 1998b. Molecular analysis of the P97
cilium adhesin operon of Mycoplasma hyopneumoniae. Gene
214, 13–23.
Hsu, T., Artiushin, S., Minion, F.C., 1997. Cloning and functional
analysis of the P97 swine cilium adhesin gene of Mycoplasma
hyopneumoniae. J. Bacteriol. 179, 1317–1323.
Hunter, P.R., Gaston, M.A., 1988. Numerical index of the discrimi-
natory ability of typing systems: an application of Simpson’s
index of diversity. J. Clin. Microbiol. 26, 2465–2466.
Kay, B.K., Williamson, M.P., Sudol, M., 2000. The importance of
being proline: the interaction of proline-rich motifs in signaling
proteins with their cognate domains. FASEB J. 14, 231–241.
Kokotovic, B., Friis, N.F., Jensen, J.S., Ahrens, P., 1999. Amplified-
fragment length polymorphism fingerprinting of Mycoplasma
species. J. Clin. Microbiol. 37, 3300–3307.
Maes, D., Deluyker, H., Verdonck, M., Castryck, F., Miry, C., Vrijens,
B., de Kruif, A., 1999. Risk indicators for the seroprevalence of
Mycoplasma hyopneumoniae, porcine influenza viruses and
Aujeszky’s disease virus in slaughter pigs from fattening pig
herds. Zentralbl. Veterinarmed. [B] 46, 341–352.
Minion, F.C., Lefkowitz, E.J., Madsen, M.L., Cleary, B.J., Swartzell,
S.M., Mahairas, G.G., 2004. The genome sequence of Mycoplas-
ma hyopneumoniae strain 232, the agent of swine mycoplasmosis.
J. Bacteriol. 186, 7123–7133.
Olive, D.M., Bean, P., 1999. Principles and applications of methods
for DNA-based typing of microbial organisms. J. Clin. Microbiol.
37, 1661–1669.
Penner, G.A., Bush, A., Wise, R., Kim, W., Domier, L., Kasha, K.,
Laroche, A., Scoles, G., Molnar, S.J., Fedak, G., 1993. Repro-
ducibility of random amplified polymorphic DNA (RAPD) anal-
ysis among laboratories. PCR Methods Appl. 2, 341–345.
Ramisse, V., Houssu, P., Hernandez, E., Denoeud, F., Hilaire, V.,
Lisanti, O., Ramisse, F., Cavallo, J.D., Vergnaud, G., 2004. Var-
iable number of tandem repeats in Salmonella enterica subsp.
enterica for typing purposes. J. Clin. Microbiol. 42, 5722–5730.
Rawadi, G.A., 1998. Characterization of Mycoplasmas by RAPD
fingerprinting. Methods Mol. Biol. 104, 179–187.
Rocha, E.P., Blanchard, A., 2002. Genomic repeats, genome plasticity
and the dynamics of Mycoplasma evolution. Nucleic. Acids Res.
30, 2031–2042.
Rosengarten, R., Yogev, D., 1996. Variant colony surface antigenic
phenotypes within Mycoplasma strain populations: implications
for species identification and strain standardization. J. Clin.
Microbiol. 34, 149–158.

T. Stakenborg et al. / Journal of Microbiological Methods 66 (2006) 263–275 275
Rucker, A.L., Creamer, T.P., 2002. Polyproline II helical structure in
protein unfolded states: lysine peptides revisited. Protein Sci. 11,
980–985.
Scott, A.N., Menzies, D., Tannenbaum, T.N., Thibert, L., Kozak, R.,
Joseph, L., Schwartzman, K., Behr, M.A., 2005. Sensitivities and
specificities of spoligotyping and mycobacterial interspersed re-
petitive unit-variable-number tandem repeat typing methods for
studying molecular epidemiology of tuberculosis. J. Clin. Micro-
biol. 43, 89–94.
Stakenborg, T., Vicca, J., Butaye, P., Maes, D., Peeters, J., de Kruif,
A., Haesebrouck, F., 2005. The diversity of Mycoplasma hyop-
neumoniae within and between herds using pulsed-field gel elec-
trophoresis. Vet. Microbiol. 109, 29–36.
Urwin, R., Maiden, M.C., 2003. Multi-locus sequence typing: a tool
for global epidemiology. Trends Microbiol. 11, 479–487.
van Eldere, J., Janssen, P., Hoefnagels-Schuermans, A., van Lierde,
S., Peetermans, W.E., 1999. Amplified-fragment length polymor-
phism analysis versus macro-restriction fragment analysis for
molecular typing of Streptococcus pneumoniae isolates. J. Clin.
Microbiol. 37, 2053–2057.
Van Looveren, M., Ison, C.A., Ieven, M., Vandamme, P., Martin, I.M.,
Vermeulen, K., Renton, A., Goossens, H., 1999. Evaluation of the
discriminatory power of typing methods for Neisseria gonor-
rhoeae. J. Clin. Microbiol. 37, 2183–2188.
Vicca, J., Stakenborg, T., Maes, D., Butaye, P., Peeters, J., de
Kruif, A., Haesebrouck, F., 2003. Evaluation of virulence of
Mycoplasma hyopneumoniae field isolates. Vet. Microbiol. 97,
177–190.
Wilton, J.L., Scarman, A.L., Walker, M.J., Djordjevic, S.P., 1998.
Reiterated repeat region variability in the ciliary adhesin gene of
Mycoplasma hyopneumoniae. Microbiology 144, 1931–1943.
Yazdankhah, S.P., Lindstedt, B.A., Caugant, D.A., 2005. Use of
variable-number tandem repeats to examine genetic diversity of
Neisseria meningitidis. J. Clin. Microbiol. 43, 1699–1705.
Related Documents











