Vol. 30, No. 3 JOURNAL OF VIROLOGY, June 1979, p. 839-846 0022-538X/79/06-0839/08$02.00/0 Comparison of the Genomes of Simian, Bovine, and Human Rotaviruses by Gel Electrophoresis and Detection of Genomic Variation Among Bovine Isolates SUZANNE M. RODGER* AND IAN H. HOLMES Department of Microbiology, University of Melbourne, Parkville, Victoria, 3052 Australia Received for publication 30 November 1978 By co-electrophoresis in polyacrylamide gels, the segmented double-stranded RNA genome of the simian rotavirus, SA 11, was compared with those of human and bovine rotaviruses. A comparison between SA 11 virus and the Northern Ireland cell culture adapted bovine virus showed that the electrophoretic mobil- ities of each of the 11 corresponding segments differed. In other comparisons, four to seven segment variations were more common. When the genomes of various bovine rotaviruses were compared, eight different electropherotypes were detected. Four of these electropherotypes were obtained from one property during a single outbreak of disease. In view of such genetic diversity, a scheme for the systematic designation of different rotavirus samples is proposed. The significance of the variations in relation to the molecular epidemiology of bovine rotavirus infections is discussed. Biochemical and biophysical studies have shown that the rotavirus genome consists of eleven segments of double-stranded RNA (12, 15, 20, 22; S. M. Rodger and I. H. Holmes, submitted for publication). However, in spite of the close serological relationship between rota- viruses obtained from different animal species (19), quite obvious differences in the relative electrophoretic mobility of corresponding ge- nome segments have been noted, thus making them readily distinguishable (4, 5, 13, 21; Rodger and Holmes, submitted for publication). To fa- cilitate comparisons between rotavirus samples in different laboratories, some form of standard- ization has become necessary so, as proposed by Rodger and Holmes (submitted for publication), the genomes of some Australian human and bovine rotaviruses have been co-electrophoresed with the genome of a simian rotavirus, SA 11. Initially, it was generally assumed that gel electrophoresis of genomes would provide a sim- ple means of distinguishing rotaviruses from dif- ferent hosts (6, 12, 15). More recently, genomic variations have been found among rotaviruses obtained from single species (1, 6, 22), but the diversity we are now reporting is much greater than was previously suspected. At this stage, care must be exercised in the interpretation of RNA profiles for identification of the species of origin of new rotaviruses; nevertheless, electro- phoresis is an excellent tool for molecular epi- demiological studies of rotavirus infections. MATERIAIS AND METHODS Viruses and cells. Simian rotavirus SA 11 was propagated as described previously (13). The cell culture-adapted strain of Northern Ireland bovine rotavirus (kindly supplied by M. S. McNulty) was propagated in the CV-1 monkey kidney cell line (Cercopithecus aethiops) in the same way as SA 11 virus. Field samples of bovine rotavirus were extracted from single stools collected from calves on properties in Western Victoria during the winters of 1974, 1975, 1976, and 1977. Some of these samples were kindly supplied by the Victorian Department of Agriculture. Also, a single stool containing human rotavirus, col- lected during the winter of 1976, was kindly supplied by R. D. Schnagl. Purification of viruses. Preparations containing large numbers of virus particles were purified as de- scribed previously (12). Smaller samples were. only clarified by differential centrifugation. Deproteinization of viral nucleic acid. Virus preparations were deproteinized with phenol and so- dium dodecyl sulfate (SDS) as described (12). Prepa- rations still containing material which interfered with electrophoresis and staining (producing wavy bands and fluorescent backgrounds) were subsequently ex- tracted with chloroform and isoamyl alcohol as de- scribed by Oda and Joklik (9). Pelleted RNA was suspended in 100 Al of Laemmli sample buffer (7). Polyacrylamide gel electrophoresis of RNA. Electrophoresis of deproteinized RNA was conducted in 10% polyacrylamide slab gels with a 3% stacking gel, using the discontinuous buffer system described by Laemmli (7). Electrophoresis was conducted at room temperature for 4 to 5 h, at a constant current of 40 839 on February 2, 2019 by guest http://jvi.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 30, No. 3JOURNAL OF VIROLOGY, June 1979, p. 839-8460022-538X/79/06-0839/08$02.00/0
Comparison of the Genomes of Simian, Bovine, and HumanRotaviruses by Gel Electrophoresis and Detection of Genomic
Variation Among Bovine IsolatesSUZANNE M. RODGER* AND IAN H. HOLMES
Department ofMicrobiology, University ofMelbourne, Parkville, Victoria, 3052 Australia
Received for publication 30 November 1978
By co-electrophoresis in polyacrylamide gels, the segmented double-strandedRNA genome of the simian rotavirus, SA 11, was compared with those of humanand bovine rotaviruses. A comparison between SA 11 virus and the NorthernIreland cell culture adapted bovine virus showed that the electrophoretic mobil-ities of each of the 11 corresponding segments differed. In other comparisons,four to seven segment variations were more common. When the genomes ofvarious bovine rotaviruses were compared, eight different electropherotypes weredetected. Four of these electropherotypes were obtained from one property duringa single outbreak of disease. In view of such genetic diversity, a scheme for thesystematic designation of different rotavirus samples is proposed. The significanceof the variations in relation to the molecular epidemiology of bovine rotavirusinfections is discussed.
Biochemical and biophysical studies haveshown that the rotavirus genome consists ofeleven segments of double-stranded RNA (12,15, 20, 22; S. M. Rodger and I. H. Holmes,submitted for publication). However, in spite ofthe close serological relationship between rota-viruses obtained from different animal species(19), quite obvious differences in the relativeelectrophoretic mobility of corresponding ge-nome segments have been noted, thus makingthem readily distinguishable (4, 5, 13, 21; Rodgerand Holmes, submitted for publication). To fa-cilitate comparisons between rotavirus samplesin different laboratories, some form of standard-ization has become necessary so, as proposed byRodger and Holmes (submitted for publication),the genomes of some Australian human andbovine rotaviruses have been co-electrophoresedwith the genome of a simian rotavirus, SA 11.
Initially, it was generally assumed that gelelectrophoresis of genomes would provide a sim-ple means of distinguishing rotaviruses from dif-ferent hosts (6, 12, 15). More recently, genomicvariations have been found among rotavirusesobtained from single species (1, 6, 22), but thediversity we are now reporting is much greaterthan was previously suspected. At this stage,care must be exercised in the interpretation ofRNA profiles for identification of the species oforigin of new rotaviruses; nevertheless, electro-phoresis is an excellent tool for molecular epi-demiological studies of rotavirus infections.
MATERIAIS AND METHODSViruses and cells. Simian rotavirus SA 11 was
propagated as described previously (13).The cell culture-adapted strain of Northern Ireland
bovine rotavirus (kindly supplied by M. S. McNulty)was propagated in the CV-1 monkey kidney cell line(Cercopithecus aethiops) in the same way as SA 11virus.
Field samples of bovine rotavirus were extractedfrom single stools collected from calves on propertiesin Western Victoria during the winters of 1974, 1975,1976, and 1977. Some of these samples were kindlysupplied by the Victorian Department of Agriculture.Also, a single stool containing human rotavirus, col-lected during the winter of 1976, was kindly suppliedby R. D. Schnagl.
Purification of viruses. Preparations containinglarge numbers of virus particles were purified as de-scribed previously (12). Smaller samples were. onlyclarified by differential centrifugation.
Deproteinization of viral nucleic acid. Viruspreparations were deproteinized with phenol and so-dium dodecyl sulfate (SDS) as described (12). Prepa-rations still containing material which interfered withelectrophoresis and staining (producing wavy bandsand fluorescent backgrounds) were subsequently ex-tracted with chloroform and isoamyl alcohol as de-scribed by Oda and Joklik (9). Pelleted RNA wassuspended in 100 Al of Laemmli sample buffer (7).
Polyacrylamide gel electrophoresis of RNA.Electrophoresis of deproteinized RNA was conductedin 10% polyacrylamide slab gels with a 3% stacking gel,using the discontinuous buffer system described byLaemmli (7). Electrophoresis was conducted at roomtemperature for 4 to 5 h, at a constant current of 40
839
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

840 RODGER AND HOLMES
mA and in the presence of 0.1% SDS. Gel slabs were1.5 mm thick, and electrophoresis was carried out inan apparatus based on the design described by Studier(17). At the completion of electrophoresis, the gel slaband the reservoir buffer were distinctly warm. Suchwarming did not appear to be harmful to the resolu-tion. Slabs were stained with ethidium bromide at afinal concentration of 1 ,ug/ml for 30 to 60 min, orovernight, before photography.
Comparisons of the various isolates were made bymixing and then co-electrophoresing the RNA of ap-propriate genomes. Aggregation did not appear to bea problem.
RESULTSNaming of rotaviruses. In view of the re-
sults reported here, a systematic scheme for thenaming of rotaviruses is clearly needed. It isproposed that a scheme based on the nomencla-ture used in naming influenza viruses be used.The system utilizes a cryptogram which shouldcontain the following information.
(i) Species of animal from which the rotaviruswas obtained, including human (HU), bovine(BO), porcine (PO), murine (MU), simian (SI),ovine (OV), lapine (LA), equine (EQ), canine(CN), caprine (CP), etc. In the event that, forexample, a human rotavirus is obtained from acalf, the abbreviation for the animal from whichthe virus was obtained should be followed, inparentheses, by the abbreviation for the type ofvirus, e.g., BO (HU).
(ii) Geographical origin of virus, e.g., country,state, city, etc.
(iii) Strain identification number, if appropri-ate.
(iv) Year in which the virus was obtained.(v) Serotype number: because several differ-
ent methods are currently in use for serotyping(18, 23) and there is disagreement, a distinctionshould be made as to which procedure was used.For example, types identified by complementfixation, serum neutralization, or hemagglutina-tion-inhibition should be listed as Ci, C2 ... Ni,N2 ... Hi, H2 ... etc., a"d others identified byradioimmunoassay or enzyme-linked immuno-sorbent assay listed as Ri, 2 ... El, 2 ... etc.
(vi) Electropherotype: it is proposed that vi-ruses of differing electropherotypes be identifiedby a capital letter.Cryptograms for the viruses used in this study
(Table 1) were produced by the above proce-dure.Co-electrophoresis of the simian rotavi-
rus genome with those of human and Aus-tralian and Northern Ireland bovine rota-viruses. By co-electrophoresis, the genome ofsimian rotavirus (SI/S. Africa/H96/58) (8) wascompared with the genomes of an Australian
human rotavirus (HU/W. Australia/76) andthree bovine rotaviruses (BO/Victoria/75/B,BO/Victoria/77/C and BO/N. Ireland/76/F).Figure 1 shows the fractionation pattern ob-tained by co-electrophoresing the genome ofsimian rotavirus with that of rotavirus HU/W.Australia/76. Although the RNA of the twoviruses migrated in a similar pattern, differencesin the mobility of six segments, i.e., 1, 2, 3, 4, 5,and 8, were observed. In the region of segments7, 8, and 9, segments 7 and 8 of SI/S. Africa/H96/58 comigrated with segment 7 of HU/W.Australia/76, and segment 9 of the former co-
TABLE 1. Proposed nomenclature for rotavirusesElectropherotype Proposed name
A BO/Victoria/185/74/AB BO/Victoria/75/BC BO/Victoria/77/CD BO/Victoria/38/74/DE BO/Victoria/75/74/EF BO/N. Ireland/76/FG BO/Victoria/76/GH BO/Victoria/47/74/H
SA 11 SI/S. Africa/H96/58Human rotavirus HU/W. Australia/76
FIG. 1. Comparison of the RNA migration pat-terns of a bovine, a simian, and a human rotavirus,electrophoresed on one 10% Laemmlipolyacrylamidegel slab. Row I (BOIC) is BO/Victoria/77/C; row 2(C+ SI) isBO/Victoria/77/Cco-electrophoresed withSI/S. Africa/H96/58; row 3 (SI/58) is SI/S. AfricalH96/58; row 4 (SI + HU) is SI/S. Africa/H96/58 co-electrophoresed with HU/W. Australia/76; and row5 (HU/76) is HUIW. Australia/76. Migration wasfrom top to bottom. RNA segments are numberedfrom the top, and in the case ofSA 1I virus, segments7 and 8 have comigrated.
J. VIROL.
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

GENOMIC VARIATION AMONG ROTAVIRUSES 841
migrated with segment 9 of the latter.Comparison of the simian rotavirus genome
with rotavirus BO/Victoria/75/B is shown inFig. 2, row 7, and variation in the electrophoreticmobility of 7 segments, i.e., 1, 3, 6, 7, 8, 9, and 10was observed. Segment 9 of the simian virusappeared to comigrate with segment 8 of BO/Victoria/75B. Similarly, co-electrophoresis ofthe simian rotavirus genome with that of BO/Victoria/77/C rotavirus revealed variations inthe mobility of segments 1, 3, 6, 7, 8, andl 10 (Fig.1, row 2). In this case, segment 9 of SI/S. Africa/H96/58 comigrated with segment 9 of BO/Vic-toria/77. (BO/Victoria/75/B and BO/Victoria/77C rotaviruses have different RNA profiles; seebelow.)The simian rotavirus genome was also com-
pared with the genome of the cell cultureadapted rotavirus BO/N. Ireland/76/F (Fig. 2,row 5). In this case, all corresponding genomesegments were observed to migrate differently.Although it is difficult to resolve the differenceinvolving segments 10 and 11 photographically,
they were visible on the freshly stained gel.Segments 7, 8, and 9 of BO/N. Ireland/76/Fvirus were unresolved and migrated in the spacebetween the comigrating segments 7, and 8, andsegment 9 of SI/S. Africa/H96/58 virus (Table2).Diversity ofthe rotavirus genome as seen
in specimens of bovine origin. To examinethe degree of diversity of the rotavirus genome,the RNA of rotaviruses of bovine origin was
compared by polyacrylamide gel electrophore-sis. By using this technique, eight different re-
producible fractionation patterns were obtained(Fig. 3). Where material was available, RNApreparations showing fractionation patternsidentical to those originally obtained were suc-
cessfully reisolated from original fecal samples(Fig. 3A, B, C, D). In the case of the cell culture-adapted strain of calf rotavirus from NorthemIreland (Fig. 3F), the pattem was successfullyreproduced even after passage of the virus in cellculture, indicating that the virus genome is sta-ble. To further examine which segments were
FIG. 2. Electrophoretic fractionation patterns of the RNA of a simian and various bovine rotaviruses, onthe one 10% Laemmli polyacrylamide gel slab. Row I (BO/B) is BO/Victoria/75/B; row 2 (B + F) is BO/Victoria/75/B co-electrophoresed with BO/N. Ireland/76/F; row 3 (F + C) BO/N. Ireland/76/F co-electro-phoresed with BO/Victoria/77/C; row 4 (BO/F) BO/N. Ireland/76/F; row 5 (F + SI) is BO/N. Ireland/76/Fco-electrophoresed with SI/S. Africa/H96/58; row 6 (SI/58) is SI/S. Africa/H96/58; row 7 (B + SI) is BO/Victoria/75/B co-electrophoresed with SI/S. Africa/H96/58; and row 8 (BO/B) is BO/Victoria/75/B. Migra-tion was from top to bottom. RNA segments are numbered from the top, and in the case ofBO/F, segments 2and 3 have comigrated, as have segments 7, 8, and 9.
TABLE 2. Comparison of the genome ofa simian rotavirus with those obtained from other animal species
Rotaviruses co-electrophoresed Segment variations Total varia-tionsSI/S. Africa/H96/58 + BO/N. Ireland/76/F 1 2 3 4 5 6 7 8 9 10 11 11SI/S. Africa/H96/58 + BO/Victoria/75/B 1 3 6 7 8 9 10 7SI/S. Africa/H96/58 + BO/Victoria/77/C 1 3 6 7 8 10 6SI/S. Africa/H96/58 + HU/W. Australia/76 1 2 3 4 5 8 6
VOL. 30, 1979
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

842 RODGER AND HOLMES
A B C CL E F G H
FIG. 3. Eight different bovine rotavirus RNA electropherotypes. Types A, D, and H were electrophoresedon one gel slab, types E and were electrophoresed on another slab, and the remainder were each
electrophoresed on -a different slab gel. (A) BO/Victoria/185/74/A; (B) BO/Victoria/75/B; (C) BOlVictorial77/C; (D) BO/Victoria/38/74/D; (E) BO/Victoria/75/74/E; (F) BOIN. Ireland/76/F; (G) BO/Victoria/76/G;and (H) BO/Victoria/47/74/H. Migration was fr-om top to bottom on 10% Laemmlipolyacrylamide slab gels.RNA segments are numbered fr-om the top.
involved in the variations, the RNAs were co-
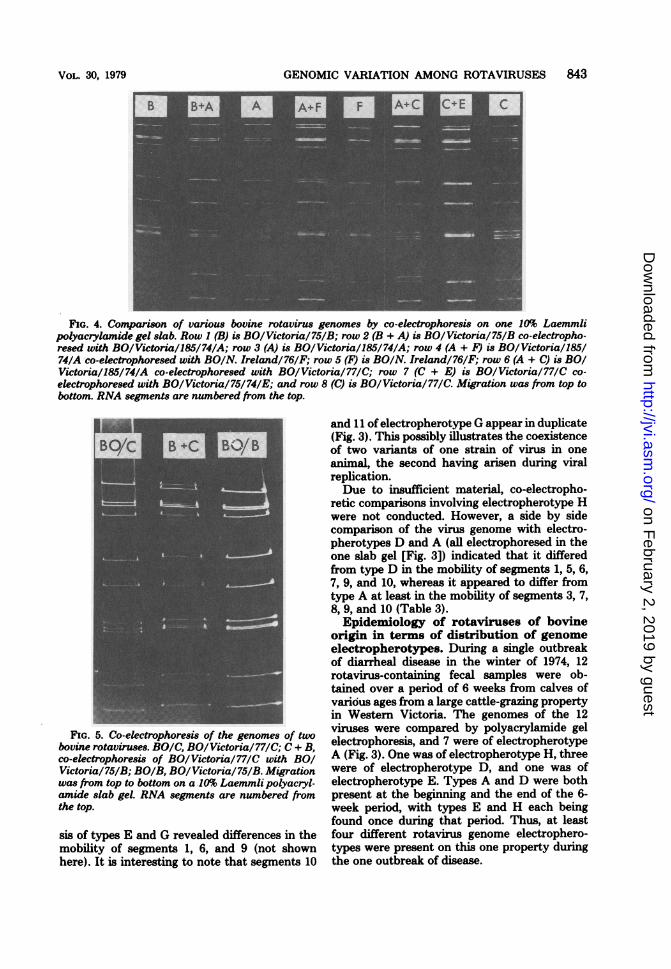
electrophoresed, and typical profiles are seen inFig. 2, 4, and 5. Co-electrophoresis of electro-pherotypes A and F (Fig. 4, row 4) revealeddifferences in the mobility of segments 1, 2, 4, 5,6, 7, 8, 9, and 10. Segments 2 and 3 of type Fappeared to comigrate with segment 3 of typeA, and segments 7, 8, and 9 of the former mi-grated in the space between segments 7 and 9 ofthe latter. The possibility of a difference involv-ing segment 8 cannot be excluded.When the genomes of electropherotypes A
and B were compared (Fig. 4, row 2), all genomesegments were found to migrate identically, ex-
cept in the case of segment 1. This result indi-cated a very close relationship between the twoviruses.
Co-electrophoresis ofelectropherotypes A andC (Fig. 4, row 6) revealed differences in themobility of 4 segments, i.e., 2, 5, 6, and 10,whereas comparison of type C with type E (Fig.4, row 7) revealed differences in the mobility ofsegments 1, 2, 7, and 9.Although the fractionation patterns of types
B and C appear similar (Fig. 3), co-electropho-resis of their genomes revealed differences in the
migration of segments 1, 2, 5, 6, 8, and 10 (Fig.5). The differences involving segment 8 were
difficult to reproduce photographically and onlyappeared as a fuzzy band.
Electropherotype F was co-electrophoresedwith electropherotype E (not shown), and differ-ences in the mobility of segments 1, 4, 6; and 10were observed, whereas co-electrophoresis oftype F with type B (Fig. 2, row 2) revealeddifferences in the mobility of all segments except3 and 11. The band representing segments 2 and3 of type F comigrated with the band represent-ing segment 3 of type B, thus indicating a differ-ence in the mobility of segment 2. Similarly, theband representing segments 7, 8, and 9 of typeF migrated just ahead of segment 8 of type B,indicating that the mobility of segments 7, 8, and9 of the two viruses differed.Comparison of electropherotype F with elec-
tropherotype C (Fig. 2, row 3) showed differ-ences in the mobility of six segments, i.e., 2, 4, 6,7, 9, and 10. In this case, segments 7, 8, and 9 oftype F comigrated with segment 8 of type C.
Co-electrophoresis ofelectropherotypes A andD revealed differences in the mobility of 3 seg-ments, i.e., 1, 5, and 6, whereas co-electrophore-
J. VIROL.
I4
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
GENOMIC VARIATION AMONG ROTAVIRUSES 843
+A+A
FIG. 4. Comparison of various bovine rotavirus genomes by co-electrophoresis on one 10% Laemmlipolyacrylamide gel slab. Row 1 (B) is BO/Victoria/75/B; row 2 (B + A) is BO/Victoria/75/B co-electropho-resed with BO/Victoria/185/74/A; row 3 (A) is BO/Victoria/185/74/A; row 4 (A + F) is BO/Victoria/185/74/A co-electrophoresed with BO/N. Ireland/76/F; row 5 (F) is BO/N. Ireland/76/F; row 6 (A + C) is BOIVictoria/185/74/A co-electrophoresed with BO/Victoria/77/C; row 7 (C + E) is BO/Victoria/77/C co-electrophoresed with BO/Victoria/75/74/E; and row 8 (C) is BO/Victoria/77/C. Migration was from top tobottom. RNA segments are numbered from the top.
FIG. 5. Co-electrophoresis of the genomes of twobovine rotaviruses. BO/C, BO/Victoria/77/C; C + B,co-electrophoresis of BO/Victoria/77/C with BOQVictoria/75/B; BO/B, BO/Victoria/75/B. Migrationwas from top to bottom on a .10% Laemmli polyacryl-amide slab gel. RNA segments are numbered fromthe top.
sis of types E and G revealed differences in themobility of segments 1, 6, and 9 (not shownhere). It is interesting to note that segments 10
and 11 of electropherotype G appear in duplicate(Fig. 3). This possibly illustrates the coexistenceof two variants of one strain of virus in oneanimal, the second having arisen during viralreplication.Due to insufficient material, co-electropho-
retic comparisons involving electropherotype Hwere not conducted. However, a side by sidecomparison of the virus genome with electro-pherotypes D and A (all electrophoresed in theone slab gel [Fig. 3]) indicated that it differedfrom type D in the mobility of segments 1, 5, 6,7, 9, and 10, whereas it appeared to differ fromtype A at least in the mobility of segments 3, 7,8, 9, and 10 (Table 3).Epidemiology of rotaviruses of bovine
origin in terms of distribution of genomeelectropherotypes. During a single outbreakof diarrheal disease in the winter of 1974, 12rotavirus-containing fecal samples were ob-tained over a period of 6 weeks from calves ofvarious ages from a large cattle-grazing propertyin Westem Victoria. The genomes of the 12viruses were compared by polyacrylamide gelelectrophoresis, and 7 were of electropherotypeA (Fig. 3). One was of electropherotype H, threewere of electropherotype D, and one was ofelectropherotype E. Types A and D were bothpresent at the beginning and the end of the 6-week period, with types E and H each beingfound once during that period. Thus, at leastfour different rotavirus genome electrophero-types were present on this one property duringthe one outbreak of disease.
VOL. 30, 1979
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

844 RODGER AND HOLMES
TABLE 3. Comparison of the genomes ofrwtavirusesobtained from calves
Rotavi- Totalruses CO- Segment variations vari-
electropho- ationsresed
A+D 1 5 6 3A+E 1 2 4 5 6 7 8 9 10 9A+B 1 1A+C 2 5 6 10 4E+C 1 2 7 9 4B + C 1 2 5 6 8 10 6E+G 1 6 9 3E+F 1 4 6 10 4E + B 1 2 4 5 6 7 8 9 10 9C + F 2 4 6 7 9 10 6
In another study, a rotavirus of electrophero-type B was obtained from a scouring calf on asmall (130-acre [ca. 52.61-ha]) dairy farm insouthwestern Victoria during an outbreak of"white scours" in the winter of 1975. In 1976, theproperty appeared to be free of disease, but inthe winter of 1977, a rotavirus of electrophero-type C was obtained from a scouring calf. Al-though other calves were also affected, no more
samples were collected. It is therefore not knownwhether either of these electropherotypes or anyother electropherotype was present on this prop-erty at a time other than that specified; i.e., it isnot known whether electropherotype C was alsopresent in 1975, and likewise, whether electro-pherotype B was present in 1977.Although the genomes of rotaviruses appear
to vary at one locality, whether in the same or
different years, they also vary from place toplace. For instance, electropherotypes G and Fwere obtained during 1976 in Australia andNorthem Ireland, respectively. Considering thegeographical separation of these two viruses,they appear to be closely related. This degree ofclose similarity of genomes is also seen in rota-viruses obtained from different places, but alsoat different times. For example, electrophero-types A and B, shown to differ in segment 1only, were obtained during the winters of 1974and 1975, respectively, on properties approxi-mately 100 miles (ca. 160.93 km) apart. Simi-larly, electropherotypes E and G, with smalldifferences in three segments, were obtainedduring the winters of 1974 and 1976 on propertiesapproximately 60 miles (ca. 96.56 km) apart.Four segments were observed to vary when elec-tropherotypes E and C were compared. Theseviruses were obtained in 1974 and 1977, respec-tively, and the properties from which they wereobtained are located 100 miles apart. Althoughthese differences involve only a few segmentsand indicate close relatedness, it must not be
J. VIROL.
forgotten that the viruses are in fact different.Only in one case, apart from those from a singleproperty, was a virus obtained with a genomethat comigrated with a previously recognizedelectropherotype. In 1975, a virus of electro-pherotype B was obtained from a property lo-cated approximately 60 miles away from anotherproperty where electropherotype B was also ob-tained at about the same time.
DISCUSSIONPolyacrylamide gel electrophoresis proved to
be a useful tool for examining the genetic diver-sity of the rotavirus genome. The fact that all 11corresponding segments migrated differentlywhen the genomes of SI/S. Africa/H96/58 andBO/N. Ireland/76 were co-electrophoresed par-alleled the degree of diversity recognized amongother members of the family Reoviridae (16, 14).Although diversity of this magnitude is previ-ously unreported for rotavirus, variations involv-ing fewer segments appeared to be more com-mon. Such genetic diversity led to the exami-nation of the epidemiology of bovine rotavirusesat a molecular level, and although our resultssupported those of Verly and Cohen (22), theyalso showed that several electropherotypes cancoexist on a single fann during one outbreak ofdisease and, possibly, that different electropher-otypes are responsible for successive outbreaksof white scours. It has also revealed that veryclosely similar electropherotypes can cause out-breaks of disease in different areas in differentyears. The possibility that several rotavirus elec-tropherotypes may be serologically indistin-guishable, as observed among reoviruses (11)and influenza viruses (2, 10), could simplify theseemingly complex situation. The existence ofhuman rotavirus serotypes has been recognizedrecently (18, 23, 24), but as yet a relationshipbetween electropherotypes and serotypes re-mains to be established. In the meantime, it ishoped that a molecular approach can be used tofollow the epidemiology and evolution of indi-vidual rotavirus strains. A similar approach wasrecently adopted by Hinshaw et al. (2) in epi-demiological studies of influenza viruses, after itwas observed that the genomes of serologicallyidentical isolates may have different electropho-retic mobility profiles. In the case of influenza,a single-stranded RNA virus, the differences inmobility of corresponding genome segmentsfrom different isolates do not necessarily reflectsize differences, but differences in their basecompositions (R. Barry, personal communica-tion), and it is possible that a similar situationapplies for rotaviruses. A study of the molecularepidemiology of a large number of human rota-
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

GENOMIC VARIATION AMONG ROTAVIRUSES 845
viruses is currently under way.Although the bovine rotavirus genome ap-
pears quite diverse, the different electrophero-types closely resemble each other in overall seg-ment distribution. This allowed the viruses to begrouped most readily by considering the mobil-ity and spatial relationships between segments2 and 3, and the "7, 8, 9 complex." On this basis,electropherotypes A, B, and C form one group,E and F form another group, and D, G, and Heach represent other groups. In a current studyofhuman rotavirus electropherotypes, such sub-grouping became a necessity (Rodger andHolmes, manuscript in preparation).As yet, the coding relationship between ge-
nome segments and polypeptides is unknown.However, in comparisons between rotaviruses ofdifferent species origins, segment 3 varied ineach case, indicating that this segment may codefor a "species-specific" component. This obser-vation was further supported when it was seenthat segment 3 was invariable when bovine ro-tavirus genomes were compared. It is also ofinterest to note that the Northern Ireland bovinerotavirus differed from the Australian bovinerotaviruses in segment 4, whereas this segmentappeared to be reasonably constant among theAustralian viruses studied. It is possible thatvariations in segment 4 are geographically ori-ented, and may be useful in epidemiologicalstudies. Our results do not provide any insightas to which segment(s) may code for the viralantigen(s). Kalica et al. (5) proposed that seg-ment 5 codes for the viral antigen on the basisthat rotaviruses obtained from consecutive in-fections in one child differed only in segment 5.If, in fact, this is correct, then 5 of the 8 bovinerotaviruses observed in this study could be se-rologically distinct. On the other hand, we alsofound that segment 5 of SI/S. Africa/H96/58 co-electrophoresed with segment 5 of BO/Victoria/75/B and BO/Victoria/77/C, so it is unlikelythat segment 5 is species specific. However, itmust be remembered that even if two segmentscomigrate, they may not necessarily code foridentical polypeptides. In any case, on the basisof the genome patterns observed, the possibilitythat rotaviruses may be as serologically diverseas rhinoviruses (6) cannot be excluded.At this stage, the epidemiology and distribu-
tion of rotavirus types can be studied readily ata molecular level by gel electrophoresis of thevirus genome. The procedure described here isquick and easy to perform, gives excellent reso-lution, and in agreement with Espejo et al. (1),is as sensitive as electron microscopy in terms ofthe quantity of material required for analysisand diagnosis. In the event of the establishmentof a relationship between serotype and electro-
pherotype, both diagnosis and strain identifica-tion can be accomplished at once. To standardizethe procedure, we suggest that the Laemmli slabgel be used, because the electrophoretic mobilityofsome segments varies with the type of electro-phoresis buffer system used (11; Rodger andHolmes, submitted for publication).
ACKNOWLEDGMENTS
We thank Anneke Veenstra, Andrew Dettmer, and RobynPark for their very capable technical assistance. We alsoparticularly thank Andrew Dettmer for preparation of thephotographs.
Part of this work was completed while S. M. Rodger was aholder of a Postgraduate Scholarship awarded by the DairyingResearch Committee, Melbourne. We thank the NationalHealth and Medical Research Council of Australia for theirfinancial support.
LITERATURE CITED1. Espejo, R. T., E. Calderon, and N. Gonzalez. 1977.
Distinct reovirus-like agents associated with acute in-fantile gastroenteritis. J. Clin. Microbiol. 6:502-506.
2. Hinshaw, V. S., W. J. Bean, Jr., R. G. Webster, andB. C. Easterday. 1978. The prevalence of influenzaviruses in swine and the antigenic and genetic related-ness of influenza viruses from man and swine. Virology84:51-62.
3. Holmes, I. H., S. M. Rodger, R. D. Schnagl, B. J.Ruck, I. D. Gust, R. F. Bishop, and G. L. Barnes.1976. Is lactose the receptor and uncoating enzyme forinfantile enteritis (rota) viruses? Lancet i: 1387-1389.
4. Jackson, G. G., and R. L. Muldoon. 1973. Virusescausing common respiratory infections in man. I. Rhi-noviruses. J. Infect. Dis. 127:328-355.
5. Kalica, A. R., C. F. Garon, R. G. Wyatt, C. A. Mebus,D. H. Van Kirk, R. M. Chanock, and A. Z. Kapikian.1976. Differentiation of human and calf reovirus-likeagents associated with diarrhea using polyacrylamidegel electrophoresis of RNA. Virology 74:86-92.
6. Kalica, A. R., M. M. Sereno, R. G. Wyatt, C. A. Mebus,R. M. Chanock, and A. Z. Kapikian. 1978. Compari-son of human and animal rotavirus strains by gel elec-trophoresis of viral RNA. Virology 87:247.
7. Laemmli, U. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (London) 227:680-685.
8. Malherbe, H. H., and M. Strickland-Cholmley. 1967.Simian virus SA 11 and the related 0 agent. Arch.Gesamte Virusforsch. 22:235-245.
9. Oda, K.-I., and W. K. Joklik. 1967. Hybridization andsedimentation studies on "early" and "late" vacciniamessenger RNA. J. Mol. Biol. 27:395-419.
10. Palese, P., and J. L. Schulman. 1976. Differences inRNA patterns of influenza A viruses. J. Virol. 17:876-884.
11. Ramig, R. F., R. K. Cross, and B. N. Fields. 1977.Genome RNAs and polypeptides of reovirus serotypes1, 2, and 3. J. Virol. 22:726-733.
12. Rodger, S. M., R. D. Schnagl, and I. H. Holmes. 1975.Biochemical and biophysical characteristics of diarrheaviruses of human and calf origin. J. Virol. 16:1229-1235.
13. Rodger, S. M., R. D. Schnagl, and I. H. Holmes. 1977.Further biochemical characterization, including the de-tection of surface glycoproteins, of human, calf, andsimian rotaviruses. J. Virol. 24:91-98.
14. Schnagl, R. D., and I. H. Holmes. 1975. Polyacrylamidegel electrophoresis of the genomes of two orbiviruses:D'Aguilar and Eubenangee. Intervirology 5:300-304.
15. Schnagl, R. D., and I. H. Holmes. 1976. Characteristics
VOL. 30, 1979
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

846 RODGER AND HOLMES
of the genome of human infantile enteritis virus (rota-virus). J. Virol. 19:267-270.
16. Spandidos, D. A., and A. F. Graham. 1976. Physicaland chemical characterization of an avian reovirus. J.Virol. 19:968-976.
17. Studier, F. W. 1973. Analysis of bacteriophage 77 earlyRNA and protein on slab gels. J. Mol. Biol. 79:237-248.
18. Thouless, M. E., A. S. Bryden, and T. H. Flewett.1978. Serotypes of human rotavirus. Lancet i:39.
19. Thouless, M. E., A. S. Bryden, T. H. Flewett, G. N.Woode, J. C. Bridger, D. R. Snodgrass, and J. A.Herring. 1977. Serological relationships between rota-viruses from different species as studies by complementfixation and neutralization. Arch. Virol. 53:287-294.
20. Todd, D., and M. S. McNulty. 1976. Characterization of
J. VIROL.
pig rotavirus RNA. J. Gen. Virol. 33:147-150.21. Todd, D., and M. S. McNulty. 1977. Biochemical studies
on a reovirus-like agent (rotavirus) from lambs. J. Virol.21:1215-1218.
22. Verly, E., and J. Cohen. 1977. Demonstration of sizevariation of RNA segments between different isolatesof calf rotavirus. J. Gen. Virol. 35:583-586.
23. Yolken, R. H., B. Barbour, R. G. Wyatt, A. R. Kalica,A. Z. Kapikian, and R. M. Chanock. 1978. Enzyme-linked immunosorbent assay for identification of rota-viruses from different animal species. Science 201:259-262.
24. Zissis, G., and J. P. Lambert. 1978. Different serotypesof human rotavirus. Lancet i:38-39.
on February 2, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
Related Documents











