J. Moll. Stud. (2002), 68, 111–126 © The Malacological Society of London 2002 INTRODUCTION The Baicaliidae Clessin, 1880 and the Benedictiidae Clessin, 1880 (or subfamily Benedictiinae Clessin, 1878 of Hydro- biidae) are exclusively endemic to Lake Baikal, Russia (Sitnik- ova, 1995). Although the systematic status, biogeography, and phylogenetic relationships of the Baicaliidae and Benedictiidae have been investigated for more than a century, these remain subjects for continuing debate and research. Dybowski (1875) and Lindholm (1909) described most of the known species of Baicaliidae and Benedictiidae, and pro- vided data on the teleoconch and anatomy, including some information on the radula for various species. The first Baicaliidae to be described were included by W. Dybowski (1875) in his new genus Limnorea, but that name proved to be preoccupied and Martens (1876) subsequently proposed a new name for the genus Baicalia. Dall (1877) included these species within the genus Tryonia Stimpson, 1865 (Pleistocene–Recent of North America) assigning them to three subgenera—Baicalia Martens, 1876 [T. (B.) carinata], Liobaicalia Martens, 1876 [T. (L.) stiedae], and Dybowskia Dall, 1877 [T. (D.) ciliata, T. (D.) duthiersi, now Teratobaikalia ciliata and T. duthiersi]. The most recent classification by Starobogatov & Sitnikova (1998) is mainly based on Sitnikova (1991) who focused on anatomy of the female reproductive organs (but compare Sitnikova, Röpstorf & Riedel, 2001), and subdivides the baicaliids into eight genera. This classification is used in the present account. Clessin (1880) was the first to recognize a dis- crete family, the Baicaliidae, but also suggested that this group probably had a close relationship to the Hydrobiidae (see also Lindholm, 1909; Kozhov, 1936). Similarity of the teleoconch characters led Thiele (1929) to synonymize Turricaspia COMPARATIVE SPERM ULTRASTRUCTURE OF BAIKALIAN ENDEMIC PROSOBRANCH GASTROPODS PETER RÖPSTORF 1 , JOHN M. HEALY 2 , FRANK RIEDEL 1 AND TATIANA Y. SITNIKOVA 3 1 Institute of Geological Sciences, Freie Universität, D-12249, Berlin, Germany, 2 Centre for Marine Studies, University of Queensland, 4072, Brisbane, Australia, and 3 Limnological Institute, Russian Academy of Sciences, 664033, Irkutsk, Russia (Received 19 April 2001; accepted 7 September 2001) ABSTRACT Mature euspermatozoan ultrastructure is described for seven species of the rissooidean family Baicaliidae (endemic to Lake Baikal, Russia)—Liobaicalia stiedae, Teratobaikalia ciliata, T. macrostoma, Baicalia carinata, Pseudobaikalia pulla, Maackia bythiniopsis, M. variesculpta, and M. herderiana. For com- parison with these species and previously investigated Rissooidea, two species of the Lake Baikal endemic genus Benedictia (B. cf. fragilis and B. baicalensis; Hydrobiidae: Benedictiinae of some authors, Benedictiidae of other authors) in addition to Lithoglyphus naticoides (Hydrobiidae: Lithoglyphinae) and Bythinella austriaca (Hydrobiidae: Bythinellinae) were also investigated. Paraspermatozoa were not observed in any of the species examined, supporting the view that these cells are probably absent in the Rissooidea. In general, the euspermatozoa of all species examined resemble those of many other caenogastropods (basally invaginated acrosomal vesicle, mid-piece with 7–13 helical mitochon- dria, an annulus, glycogen piece with nine peri-axonemal tracts of granules). However, the presence of a completely flattened acrosomal vesicle and a specialized peri-axonemal membranous sheath (a scroll-like arrangement of 4–6 double membranes) at the termination of the mid-piece, clearly indicates a close relationship between the Baicaliidae and other rissooidean families possessing these features (Bithyniidae, Hydrobiidae, Pyrgulidae, and Stenothyridae). Euspermatozoa of Benedictia, Lithoglyphus, Bythinella, and Pyrgula all have a solid nucleus, which exhibits a short, posterior invagina- tion (housing the centriolar complex and proximal portion of the axoneme). Among the Rissooidea, this form of nucleus is known to occur in the Bithyniidae, Hydrobiidae, Truncatellidae, Pyrgulidae, Iravadiidae, Pomatiopsidae, and Stenothyridae. In contrast, the euspermatozoa of the Baicaliidae all have a long, tubular nucleus, housing not only the centriolar derivative, but also a substantial portion of the axoneme. Among the Rissooidea, a tubular nuclear morphology has previously been seen in the Rissoidae, which could support the view, based on anatomical grounds, that the Baicaliidae may have arisen from a different ancestral source than the Hydrobiidae. However, the two styles of nuclear morphology (short, solid versus long, tubular) occur widely within the Caenogastropoda, and some- times both within a single family, thereby reducing the phylogenetic importance of nuclear differ- ences within the Rissooidea. More significantly, the occurrence of the highly unusual membranous sheath within the mid-piece region in the Baicaliidae appears to tie this family firmly to the Bithyniidae Hydrobiidae Stenothyridae Pyrgulidae assemblage. Eusperm features of Benedictia spp. strongly resemble those of hydrobiids and bithyniids, and neither support recognition of a dis- tinct family Benedictiidae (at best this is a subfamily of Hydrobiidae) nor any close connection with the hydrobiid subfamily Lithoglyphinae. Correspondence: P. Röpstorf, Institute of Geological Sciences, Freie Universität, D-12249, Berlin, Germany. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Moll. Stud. (2002), 68, 111–126 © The Malacological Society of London 2002
INTRODUCTION
The Baicaliidae Clessin, 1880 and the Benedictiidae Clessin,1880 (or subfamily Benedictiinae Clessin, 1878 of Hydro-biidae) are exclusively endemic to Lake Baikal, Russia (Sitnik-ova, 1995). Although the systematic status, biogeography, andphylogenetic relationships of the Baicaliidae and Benedictiidaehave been investigated for more than a century, these remainsubjects for continuing debate and research.
Dybowski (1875) and Lindholm (1909) described most ofthe known species of Baicaliidae and Benedictiidae, and pro-vided data on the teleoconch and anatomy, including someinformation on the radula for various species.
The first Baicaliidae to be described were included by W.Dybowski (1875) in his new genus Limnorea, but that name
proved to be preoccupied and Martens (1876) subsequentlyproposed a new name for the genus Baicalia. Dall (1877)included these species within the genus Tryonia Stimpson, 1865(Pleistocene–Recent of North America) assigning them to threesubgenera—Baicalia Martens, 1876 [T. (B.) carinata], LiobaicaliaMartens, 1876 [T. (L.) stiedae], and Dybowskia Dall, 1877 [T. (D.)ciliata, T. (D.) duthiersi, now � Teratobaikalia ciliata and T.duthiersi]. The most recent classification by Starobogatov &Sitnikova (1998) is mainly based on Sitnikova (1991) whofocused on anatomy of the female reproductive organs (butcompare Sitnikova, Röpstorf & Riedel, 2001), and subdividesthe baicaliids into eight genera. This classification is used in thepresent account. Clessin (1880) was the first to recognize a dis-crete family, the Baicaliidae, but also suggested that this groupprobably had a close relationship to the Hydrobiidae (see alsoLindholm, 1909; Kozhov, 1936). Similarity of the teleoconchcharacters led Thiele (1929) to synonymize Turricaspia
COMPARATIVE SPERM ULTRASTRUCTURE OF BAIKALIANENDEMIC PROSOBRANCH GASTROPODS
PETER RÖPSTORF1, JOHN M. HEALY2, FRANK RIEDEL1 AND TATIANA Y. SITNIKOVA3
1Institute of Geological Sciences, Freie Universität, D-12249, Berlin, Germany, 2Centre for Marine Studies, University of Queensland, 4072, Brisbane, Australia,and 3Limnological Institute, Russian Academy of Sciences, 664033, Irkutsk, Russia
(Received 19 April 2001; accepted 7 September 2001)
ABSTRACT
Mature euspermatozoan ultrastructure is described for seven species of the rissooidean familyBaicaliidae (endemic to Lake Baikal, Russia)—Liobaicalia stiedae, Teratobaikalia ciliata, T. macrostoma,Baicalia carinata, Pseudobaikalia pulla, Maackia bythiniopsis, M. variesculpta, and M. herderiana. For com-parison with these species and previously investigated Rissooidea, two species of the Lake Baikalendemic genus Benedictia (B. cf. fragilis and B. baicalensis; Hydrobiidae: Benedictiinae of some authors,Benedictiidae of other authors) in addition to Lithoglyphus naticoides (Hydrobiidae: Lithoglyphinae)and Bythinella austriaca (Hydrobiidae: Bythinellinae) were also investigated. Paraspermatozoa werenot observed in any of the species examined, supporting the view that these cells are probably absentin the Rissooidea. In general, the euspermatozoa of all species examined resemble those of manyother caenogastropods (basally invaginated acrosomal vesicle, mid-piece with 7–13 helical mitochon-dria, an annulus, glycogen piece with nine peri-axonemal tracts of granules). However, the presenceof a completely flattened acrosomal vesicle and a specialized peri-axonemal membranous sheath (a scroll-like arrangement of 4–6 double membranes) at the termination of the mid-piece, clearlyindicates a close relationship between the Baicaliidae and other rissooidean families possessing thesefeatures (Bithyniidae, Hydrobiidae, Pyrgulidae, and Stenothyridae). Euspermatozoa of Benedictia,Lithoglyphus, Bythinella, and Pyrgula all have a solid nucleus, which exhibits a short, posterior invagina-tion (housing the centriolar complex and proximal portion of the axoneme). Among the Rissooidea,this form of nucleus is known to occur in the Bithyniidae, Hydrobiidae, Truncatellidae, Pyrgulidae,Iravadiidae, Pomatiopsidae, and Stenothyridae. In contrast, the euspermatozoa of the Baicaliidae allhave a long, tubular nucleus, housing not only the centriolar derivative, but also a substantial portionof the axoneme. Among the Rissooidea, a tubular nuclear morphology has previously been seen inthe Rissoidae, which could support the view, based on anatomical grounds, that the Baicaliidae mayhave arisen from a different ancestral source than the Hydrobiidae. However, the two styles of nuclearmorphology (short, solid versus long, tubular) occur widely within the Caenogastropoda, and some-times both within a single family, thereby reducing the phylogenetic importance of nuclear differ-ences within the Rissooidea. More significantly, the occurrence of the highly unusual membranoussheath within the mid-piece region in the Baicaliidae appears to tie this family firmly to theBithyniidae � Hydrobiidae � Stenothyridae � Pyrgulidae assemblage. Eusperm features of Benedictiaspp. strongly resemble those of hydrobiids and bithyniids, and neither support recognition of a dis-tinct family Benedictiidae (at best this is a subfamily of Hydrobiidae) nor any close connection withthe hydrobiid subfamily Lithoglyphinae.
Correspondence: P. Röpstorf, Institute of Geological Sciences, Freie Universität, D-12249, Berlin, Germany. E-mail: [email protected]

Dybowski & Grochmalicki, 1917 and Micromelania Brusina, 1874,which has a fossil type. Moreover, he united the Baicaliidaewith his Micromelaniidae (now � Pyrgulidae). A more detailedcomparison between baicaliid and pyrgulid taxa by Kozhov(1951) revealed several significant anatomical differences suchas the absence of a metapodial commissure, multiple gill leaf-lets, absence of an outgrowth on the penis, absence of a bursacopulatrix, and presence of multiple seminal receptaculae.Collectively, these differences led Kozhov to conclude thatthere existed a greater phylogenetic distance between theBaicaliidae and Pyrgulidae than between the Baicaliidae andHydrobiidae. Kozhov (1951) emphasized the distinctness ofthe Baicaliidae from the Hydrobiidae, as well as from the Pyr-gulidae, but was aware that these taxa were probably moreclosely related to each other than to any other families withinthe Rissooidea. In contrast, Starobogatov (1970) interpretedthe similarities between baicaliids and pyrgulids as the result ofconvergence. The Baicaliidae were maintained by Boss (1982)and, more recently, by Vaught (1989) as a distinct rissooideanfamily in their classifications. In contrast, Ponder (1988) pre-ferred to regard them as only a subfamily of the Hydrobiidaeand Ponder & Warén (1988) formally listed baicaliids as such intheir classification of higher prosobranchs, but without anyindication of specific reasons.
Dybowski (1875) regarded Benedictia as belonging to theHydrobiidae. Soon after, Clessin (1878) placed his new subfam-ily, the Benedictiinae, within the Rissoidae, but later recog-nized a separate family Benedictiidae (Clessin, 1880). Fischer(1887) considered the genus Benedictia as belonging to the sub-family Lithoglyphinae of the family Hydrobiidae. Lindholm(1909) included two endemic genera – Benedictia W. Dybowski1875 and Kobeltocochlea Lindholm, 1909 in the Benedictiidae.Kozhov (1945, 1963) studying the anatomy of these taxa foundsimilarities to the Lithoglyphinae and especially to the NorthAmerican genus Fluminicola Stimpson, 1865. Additional simi-larities also centre on radula characters and the sculpture ofthe protoconch (Thompson, 1984; P. Röpstorf, T. Y. Sitnikova& F. Riedel, unpublished data). The present study on spermultrastructure was conducted partly to find additional charac-ters to clarify the taxonomic status of the Benedictiidae withinthe Rissooidea.
Ultrastructural studies of molluscan euspermatozoa in several supra-specific taxa have provided useful characters fortaxonomic and phylogenetic discussion especially at and abovethe family level (Giusti, 1971; Thompson, 1973; Healy,1982a–c, 1983a,c, 1986a,b, 1988a,b, 1989, 1990, 1992, 1993,1996a,b; Healy & Willan, 1984; Kohnert & Storch, 1984, Koike,1985; Hodgson & Bernard, 1988; Hodgson, Heller & Bernard,1990; Hodgson, Bernard & Lindley, 1991; Jamieson, Hodgson& Bernard, 1991; Hodgson & Foster, 1992; Hodgson & Chia,1993; Hodgson, Ridgway, Branch & Hawkins, 1996; Hodgson,Healy & Tunnicliffe, 1997; Hodgson & Healy, 1998). A numberof previous studies have dealt with sperm ultrastructure (andoften also spermiogenesis) in various rissooideans includingrepresentatives of the Hydrobiidae (Hydrobia acuta (Drapar-naud), Bythinella cf. opaca (see Giusti 1971), Pseudamnicola sp.,Belgrandia caprai Giusti (Giusti, 1969, 1971), Hydrobia sp.(Kohnert & Storch, 1984), Truncatellidae (Truncatella sub-cylindrica (Linn.); Giusti & Mazzini, 1973), Bithyniidae(Bithynia tentaculata Linn., B. leachi Sheppard; Reader, 1973;Kohnert, 1980; Kohnert & Storch, 1984), Stenothyridae(Stenothyra sp.; Healy, 1983b, 1988a), Rissoidae (Rissoa ventri-cosa Desmarest, R. violacea Desmarest, Alvania lineata Risso;Kohnert & Storch, 1984), Pomatiopsidae (Oncomelania hupensisquadrasi; Claveria & Etges, 1988) and Pyrgulidae (Pyrgula annu-lata (Linn.); Riedel, Healy, Röpstorf & Sitnikova, 2001).
Although spermatogenesis of Benedictia has been investi-gated with light microscopy by Kozhov (1945), euspermato-
zoan ultrastructure in this genus or in any species of theBaicaliidae has until now not been documented. The aims ofthe present study are:
(1) to describe mature sperm features of the Baicaliidae andBenedictia;
(2) to use this information, and that available in the literature(listed above), to probe relationships between the Bai-caliidae, and the Hydrobiidae and the Pyrgulidae, andbetween Benedictia and undisputed members of the Hydro-biidae, especially of the subfamily Lithoglyphinae.
As a better understanding of relationships of the Baikalian taxa with hydrobiids is needed, we have included in this studyrepresentatives of other hydrobiid genera, Lithoglyphus [Litho-glyphus naticoides (C. Pfeiffer, 1828), Lithoglyphinae] andBythinella [Bythinella austriaca (Frauenfeld, 1857), Bythinel-linae].
MATERIAL AND METHODS
Species of Baicaliidae (Table 1) were gathered by SCUBA diversfrom various localities in Lake Baikal (see Figure 1) using aboat and also during expeditions of the research vesselsVereshchagin (12–21 of September 1996) and Obruchev (26–28September 1997). Benedictia cf. fragilis was gathered by trawltaken by Professor Zerbst in Chivyrkuj Bay (Figure 1). All speci-mens collected were identified by Sitnikova at the LimnologicalInstitute of the Siberian Branch of the Russian Academy ofSciences. Voucher specimens are deposited in the AustralianMuseum, Sydney (AMS registration numbers are given in Table 1).
Baikalian species (all Baicaliidae and Benedictia cf. fragilis)were collected in autumn, because spermatozoa were reportedto be mature at this time (Kozhov, 1945). Lithoglyphus naticoides(C. Pfeiffer, 1828; Hydrobiidae, Lithoglyphinae) was sampledin May 1998 from the Spree River in Berlin, Germany. Bythinella
P. RÖPSTORF ET AL .
112
Table 1. Baicaliidae and ‘Benedictiidae’ from Lake Baikal examined in thisstudy.
Species Locality Depth
Registration number Number on map Substratum
(Baicaliidae)
Liobaicalia stiedae (W. Dybowski, 1875) Kultuk 18 m
AMS C398666 1 sand
Baicalia carinata (W.Dybowski, 1875) Cape Tolsty 10 m
AMS C398660 2 sand
Teratobaikalia ciliata (W. Dybowski, 1875) Bolshiye Koty 15 m
AMS C398659 3 rock, stones
T. macrostoma (Lindholm, 1909) Kaltygej 2–3 m
AMS C398673 4 stones
Maackia variesculpta (Lindholm, 1909) Ayaya Bay 2–3 m
AMS C398662 8 stones
M. herderiana (Lindholm, 1909) Bolshiye Koty 15 m
AMS C 398656 3 rock, stones
M. bythiniopsis (Lindholm, 1909) Murinskaya Banka 18 m
AMS C 398658 5 stones
Pseudobaikalia pulla (W. Dybowski, 1875) Murinskaya Banka 18 m
AMS C398670 5 sand
‘Benedictiidae’
Benedictia cf. fragilis (W. Dybowski, 1875) Chivyrkuj Bay 20 m
AMS C398654 6 sand
B. baicalensis (Gerstfeldt, 1859) Baikalsk 15 m
AMS C398674 7 sand

austriaca (Frauenfeld, 1857; Hydrobiidae, Bythinellinae) wascollected in May 1998 from a cold spring in Dolina Eliaszowki,about 20 km NW of Krakow, Poland.
For all taxa, the complete visceral mass or sperm-ducts werefixed in 2.5 or 3.5% glutaraldehyde prepared in filtered andsterile commercial water. For investigation the samples werewashed three times in 0.05 M cacodylate buffer (pH � 7.0) overa period of 2 days. Tissues were post-fixed in 1% osmium tetrox-ide in 0.05 M cacodylate buffer for 90 min. After washing threetimes in 0.05 M cacodylate buffer the fixed parts were dehy-drated through a graded series of ethanol and embedded in Spurr’s epoxy resin. Semi-thin (1–2 �m) and ultra-thin(60–90 nm) sections were cut with a Reichert microtomeUltracut E. In order to enhance contrast, ultra-thin sectionswere stained with lead citrate (5 min), uranyl acetate (60–90min), and again with lead citrate (5 min; a modification of themethod of Daddow, 1986). A Philips transmission electronmicroscope operated at 60 or 80 kV (Freie Universität Berlin,most observations and photography) and a Hitachi 300 trans-mission electron microscope operated at 75 kV (University ofQueensland, confirmatory observations and some photo-graphy) were used to examine the sections.
For SEM investigation the glutaraldehyde-fixed sperm-ductswere washed in 0.05 M cacodylate buffer (three times for 1minute) and once in tap water. The ducts were opened, andthe sperm mass was pipetted onto a mounting-base and air-dried. After sputter-coating (Bio-Rad) the examination was carried out using a Cambridge S360 SEM.
Measurements of ultrastructural features were made usingTEM data, those of whole sperm, nucleus, mid-piece, andglycogen piece were made by light microscopy (Zeiss, AxioskopPlan-Neofluar 100). Ten sperm (15 nuclei) from two speci-
mens for each species were measured by light microscopy and10 sperm of Maackia herderiana by SEM. For light microscopyglutaraldehyde-fixed spermatozoa were transferred onto a glassslide. After air-drying the glass-mounted samples were washedgently with water and then air-dried again.
RESULTS
Paraspermatozoa were not observed in any species included inthis study. The following descriptions apply only to eusperma-tozoa (fertile sperm); n refers to the number of observations ofthe eusperm or eusperm feature under consideration.
Baicaliidae (Figures 2 and 3)
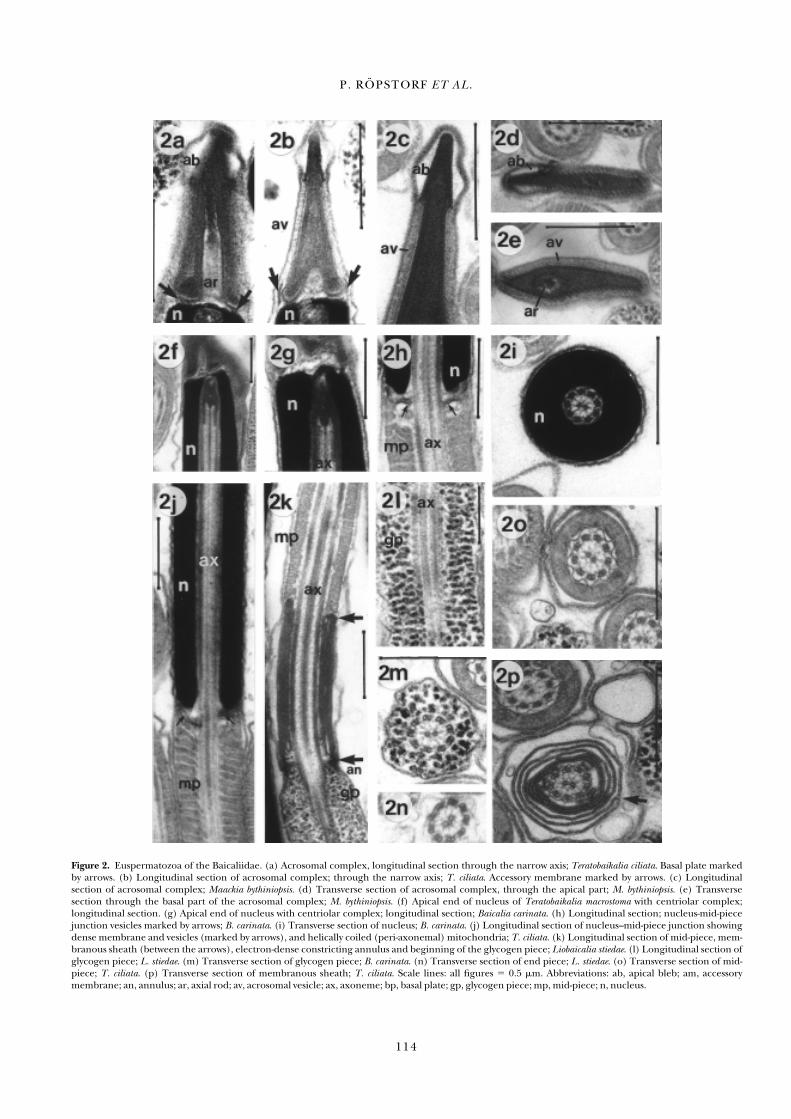
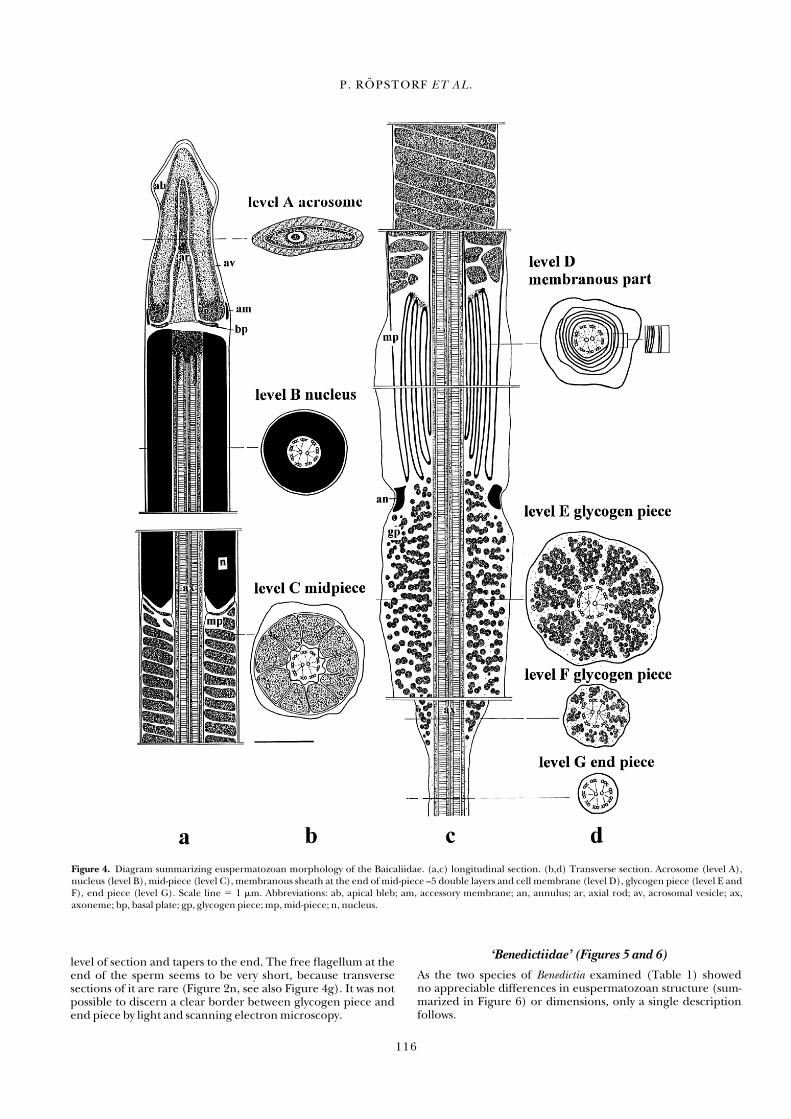
As the mature euspermatozoa of all examined baicaliid species(Table 1) show essentially the same ultrastructural features,only a single description follows. The features of a generalizedbaicaliid euspermatozoon are summarized diagrammatically in Figure 4. The total length of the sperm is about 210 �m (n � 80).
Acrosomal complex. The acrosomal vesicle is membrane-bound,approximately 1 �m long and strongly compressed laterallyresulting in a spatula-like profile (Figures 2a,d,e and 3a,d,e).The contents of the acrosomal vesicle are differentiated into adense outer layer (studded with angularly-inclined, periodically-spaced plates) and a less dense core region lacking such plates(Figures 2d,e). At the anterior tip of the acrosomal vesicle is anelectron-lucent space, the apical bleb, formed from the closespatial association of the plasma membrane, and a localizedswelling of the acrosomal vesicle membrane (Figures 2a–c).The base of the acrosomal vesicle is flanked by a short, acces-sory membrane (Figure 2b, arrows). An axial rod occupiesmost of the basal invagination of the acrosomal vesicle (Figures2a,e). A thin, centrally perforate basal plate lies between thebasal rim of the acrosomal vesicle and the nuclear apex (Figure2a).
Nucleus. The nucleus is about 12.5 �m long in all examinedspecies (n � 15 each species) and tubular, sheathing the anterior-most portion of the axoneme (Figures 2f–j and 3a,e).The diameter of the nucleus changes from a maximum of 0.6 �m basally to a minimum of 0.38 (M. bythiniopsis, P. pulla, T. ciliata) or 0.44 �m (B. carinata) in the vicinity of the acroso-mal vesicle (Figure 2f–i). A centriolar complex lies at the apexof the nucleus and is continuous with the axoneme (Figure2f,g). Dense material from this complex occurs between the central tubules and the nine doublets of the axoneme(Figure 2f,g).
Mid-piece. At the junction of the nucleus and the mid-piecedense membranes can be seen associated with apparently empty,poorly defined vesicles adjoining the mid-piece mitochondria(Figure 2h). The mid-piece is elongate (Figures 2a,b), measur-ing 125–140 �m in length (n � 80). Ultrastructurally, thisregion consists principally of the axoneme, and 7–9 continuousand elongate mitochondria, which spiral helically in a tightsheath around the axoneme (Figures 2h,j,o). A single plasmamembrane of the mitochondrial sheath encloses the axoneme.Although possibly an artefact of fixation, this membrane is folded to form a series of pockets, one associated with eachmicrotubular doublet of the axoneme the membrane formingout-pockets associated with each axonemal doublet (Figure2o). The mid-piece has a relatively constant diameter (about0.5 �m). In the distal part of the mid-piece the helical mito-chondria terminate and are replaced by a scroll-like arrange-ment of double membranes—the membranous sheath of Healy
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
113
12345678910123456789201234567893012345678940123456789501234567896012345678
Figure 1. Map of Lake Baikal (Russia) showing collection sites of materialused in this study.
P. RÖPSTORF ET AL .
114
Figure 2. Euspermatozoa of the Baicaliidae. (a) Acrosomal complex, longitudinal section through the narrow axis; Teratobaikalia ciliata. Basal plate markedby arrows. (b) Longitudinal section of acrosomal complex; through the narrow axis; T. ciliata. Accessory membrane marked by arrows. (c) Longitudinal section of acrosomal complex; Maackia bythiniopsis. (d) Transverse section of acrosomal complex, through the apical part; M. bythiniopsis. (e) Transverse section through the basal part of the acrosomal complex; M. bythiniopsis. (f) Apical end of nucleus of Teratobaikalia macrostoma with centriolar complex; longitudinal section. (g) Apical end of nucleus with centriolar complex; longitudinal section; Baicalia carinata. (h) Longitudinal section; nucleus-mid-piecejunction vesicles marked by arrows; B. carinata. (i) Transverse section of nucleus; B. carinata. (j) Longitudinal section of nucleus–mid-piece junction showingdense membrane and vesicles (marked by arrows), and helically coiled (peri-axonemal) mitochondria; T. ciliata. (k) Longitudinal section of mid-piece, mem-branous sheath (between the arrows), electron-dense constricting annulus and beginning of the glycogen piece; Liobaicalia stiedae. (l) Longitudinal section ofglycogen piece; L. stiedae. (m) Transverse section of glycogen piece; B. carinata. (n) Transverse section of end piece; L. stiedae. (o) Transverse section of mid-piece; T. ciliata. (p) Transverse section of membranous sheath; T. ciliata. Scale lines: all figures � 0.5 �m. Abbreviations: ab, apical bleb; am, accessory membrane; an, annulus; ar, axial rod; av, acrosomal vesicle; ax, axoneme; bp, basal plate; gp, glycogen piece; mp, mid-piece; n, nucleus.

(1983b)—around the axoneme (Figure 2k,p). This sheath,which measures approximately 1.2 �m in length and 0.6–0.9�m in width, lies in contact with electron dense material at itsanterior and posterior extremities (Figure 2k) and also in con-tact with the annulus. In transverse section it can be observedthat the membranes may fuse at specific points (for example,see Figure 2p, arrow).
Glycogen piece and free flagellum. The junction between the glyco-gen piece and the membranous sheath of the mid-piece is
marked by a constricting, highly electron-dense ring, the annulus (Figure 2k). The glycogen piece is approximately 70 �m in length based on SEM observations (n � 80, Figure3b,c). Immediately posterior to this structure, the axoneme issheathed by nine longitudinal tracts of putative glycogen gran-ules, one tract associated with each of the axonemal doublets(Figure 2k–m). Although the tracts of granules are discerniblein transverse sections, they are rather poorly defined (Figure2m). The granules are not densely packed. The diameter of theglycogen piece may vary from 0.3–0.7 �m, depending on the
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
115
12345678910123456789201234567893012345678940123456789501234567896012345678
Figure 3. SEM of euspermatozoa of the baicaliid Maackia herderiana. (a–c) SEM of an euspermatozoon. (a) Anterior part with acrosome, nucleus, and mid-piece, partly lying in a loop. (b) End of mid-piece with membranous sheath and glycogen piece. (c) Posterior part of glycogen piece and end piece. (d) Acrosome and anterior part of nucleus. Note the spatula-like shape of acrosome. (e) Acrosome and nucleus. (f) Transition between mid-piece and glycogen piece. Scale lines: 3a–c,e � 5 �m; 3d,f � 2 �m. Abbreviations: a, acrosome; gp, glycogen piece; mp, mid-piece; ms, membranous sheath; n, nucleus.
level of section and tapers to the end. The free flagellum at theend of the sperm seems to be very short, because transverse sections of it are rare (Figure 2n, see also Figure 4g). It was notpossible to discern a clear border between glycogen piece andend piece by light and scanning electron microscopy.
‘Benedictiidae’ (Figures 5 and 6)
As the two species of Benedictia examined (Table 1) showed no appreciable differences in euspermatozoan structure (sum-marized in Figure 6) or dimensions, only a single descriptionfollows.
P. RÖPSTORF ET AL .
116
Figure 4. Diagram summarizing euspermatozoan morphology of the Baicaliidae. (a,c) longitudinal section. (b,d) Transverse section. Acrosome (level A),nucleus (level B), mid-piece (level C), membranous sheath at the end of mid-piece –5 double layers and cell membrane (level D), glycogen piece (level E andF), end piece (level G). Scale line � 1 �m. Abbreviations: ab, apical bleb; am, accessory membrane; an, annulus; ar, axial rod; av, acrosomal vesicle; ax,axoneme; bp, basal plate; gp, glycogen piece; mp, mid-piece; n, nucleus.
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
117
12345678910123456789201234567893012345678940123456789501234567896012345678
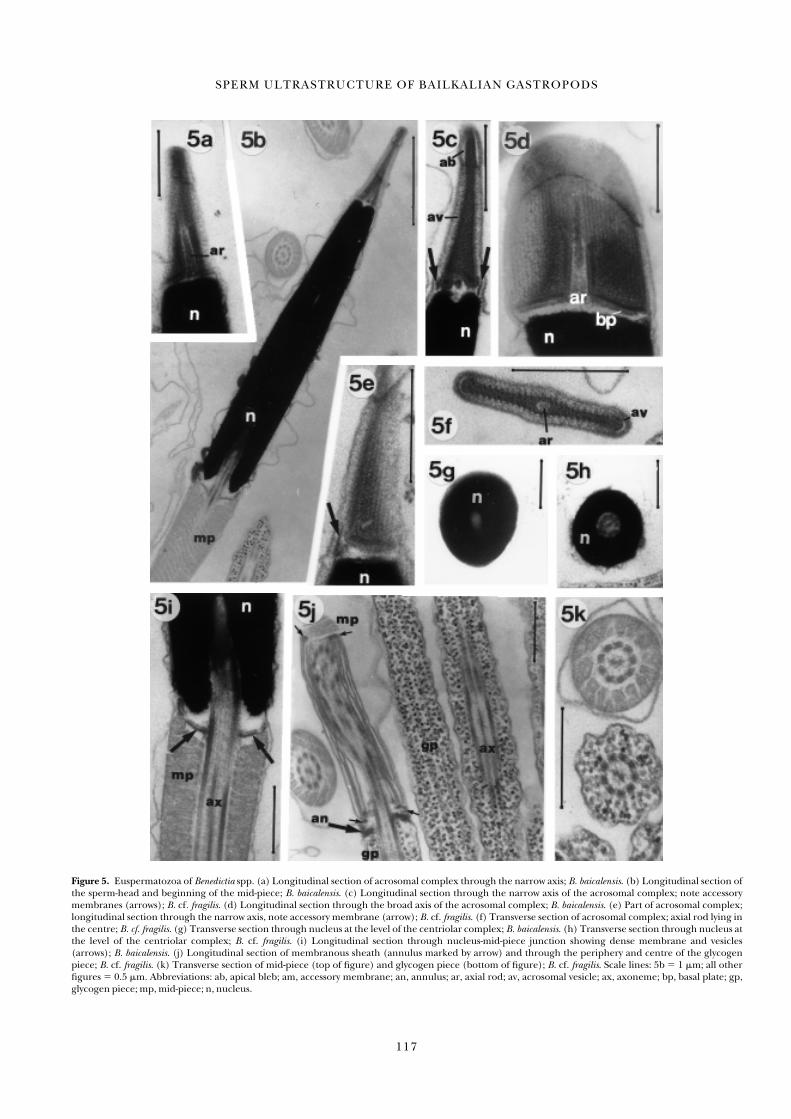
Figure 5. Euspermatozoa of Benedictia spp. (a) Longitudinal section of acrosomal complex through the narrow axis; B. baicalensis. (b) Longitudinal section ofthe sperm-head and beginning of the mid-piece; B. baicalensis. (c) Longitudinal section through the narrow axis of the acrosomal complex; note accessorymembranes (arrows); B. cf. fragilis. (d) Longitudinal section through the broad axis of the acrosomal complex; B. baicalensis. (e) Part of acrosomal complex;longitudinal section through the narrow axis, note accessory membrane (arrow); B. cf. fragilis. (f) Transverse section of acrosomal complex; axial rod lying inthe centre; B. cf. fragilis. (g) Transverse section through nucleus at the level of the centriolar complex; B. baicalensis. (h) Transverse section through nucleus atthe level of the centriolar complex; B. cf. fragilis. (i) Longitudinal section through nucleus-mid-piece junction showing dense membrane and vesicles(arrows); B. baicalensis. (j) Longitudinal section of membranous sheath (annulus marked by arrow) and through the periphery and centre of the glycogenpiece; B. cf. fragilis. (k) Transverse section of mid-piece (top of figure) and glycogen piece (bottom of figure); B. cf. fragilis. Scale lines: 5b � 1 �m; all otherfigures � 0.5 �m. Abbreviations: ab, apical bleb; am, accessory membrane; an, annulus; ar, axial rod; av, acrosomal vesicle; ax, axoneme; bp, basal plate; gp,glycogen piece; mp, mid-piece; n, nucleus.

Acrosome. The acrosomal complex (acrosomal vesicle, axial rod,and basal plate) is very similar to that observed in theBaicaliidae, but even more laterally compressed. The acro-somal vesicle is membrane-bound, about 1 �m long, spatulate,and approximately 0.9 �m wide basally (Figure 5a–f). A deep,but narrow basal invagination of the vesicle is largely filled byan axial rod (Figure 5 a,d,f). A prominent, electron-lucent,
apical bleb is present at the anterior end of the acrosomal vesicle (Figure 5c), while posteriorly a short, highly electron-dense accessory membrane surrounds the basal rim of the vesicle and in places touches the basal plate (Figure 5c).Angularly-inclined, periodically spaced plate structures runlengthwise through the outer layer of the acrosomal vesicle(Figure 5d–f). The basal plate lying between the base of the
P. RÖPSTORF ET AL .
118
Figure 6. Diagram summarizing euspermatozoan morphology of Benedictia spp. (a,c,e) Longitudinal section. (b,d) Transverse section. Acrosome (level Aand B), nucleus (level C and D), mid-piece (level E), membranous part at the end of mid-piece—five double layers and cell membrane (level F), glycogenpiece (level G). Scale line � 1 �m. Abbreviations: ab, apical bleb; am, accessory membrane; an, annulus; ar, axial rod; av, acrosomal vesicle; ax, axoneme; bp, basal plate; gp, glycogen piece; mp, mid-piece; n, nucleus.

acrosomal vesicle and nuclear apex is centrally perforate(Figure 5c,d).
Nucleus. The nucleus is highly electron-dense, relatively short(3.8–4.0 �m length), and solid (Figure 5b,g,h). Towards theacrosome the nucleus is slightly compressed laterally, giving anoval profile in transverse section, whereas basally the profile isround in transverse sections (Figure 5g). The maximum basaldiameter of the nucleus is approximately 0.9 �m. At the base ofthe nucleus a short invagination accommodates an electron-dense centriolar complex and the attached proximal portionof the axoneme (Figure 5b,i).
Mid-piece. The mid-piece morphology in both species ofBenedictia is essentially as described for the Baicaliidae—mito-chondria (n � 10 sperms) are wound helically around a 9 � 2microtubular pattern axoneme (Figure 5i–k). The number ofmitochondria is about 13. The dense membranes associatedwith vesicles at the nucleus-mid-piece junction are very clearlyapparent in longitudinal sections (Figure 5i, arrows). Themembranous sheath positioned between the terminal regionof the mitochondria and the annulus is similar to that of theBaicaliidae (Figure 5j), and is about 1.7 �m long. The anteriorand posterior extremities of this sheath are associated withdense material (Figure 5j, little arrows).
Glycogen piece and free flagellum. At the junction of mid-piece andglycogen piece an electron-dense annulus is present (Figure5j). This structure contacts both the plasma membrane and theaxoneme and also appears to contact the terminal edge of themembranous sheath. The glycogen granules are arranged intonine, longitudinally continuous tracts, one associated with eachaxonemal doublet (Figure 5j,k). The diameter throughoutmost of the glycogen piece varies from 0.45 to 0.52 �m (Figure5k), but posteriorly it tapers. The free flagellum region was notobserved.
Hydrobiidae, Lithoglyphinae (Figure 7)
Acrosomal complex. The acrosomal vesicle of Lithoglyphus nati-coides is membrane-bound, about 0.9 �m in length and spatu-late (Figure 7a,c). Contents of the vesicle are differentiatedinto a markedly electron-dense outer layer, and a granular, lessdense inner region (Figure 7b,c). Tangential longitudinal sections indicate the presence of longitudinal plates within theouter layer of the vesicle (Figure 7c). An apical bleb is very welldeveloped, and occasionally exhibits a small vesicle which ispossibly a fixation artefact (Figure 7b). At the base of the vesicle, the broad axis measures 0.46 �m and the narrow axis0.32 �m. A short posterior invagination of the acrosomal vesicle is partly filled by an equally short axial rod (Figure 7b).The basal plate and the nuclear apex it contacts are both slightly convex in longitudinal profile (Figure 7b,c). Surround-ing the basal rim of the acrosomal vesicle is a short accessorymembrane, which appears to connect with the basal plate(Figure 7b, arrow).
Nucleus. The nucleus is solid and measures about 10 �m inlength and 0.5 �m in maximum diameter (Figure 7a). Thenucleus is round in transverse section and only slightly com-pressed laterally towards the acrosome. At the posterior of thenucleus a short (0.9 �m deep) invagination houses a hollow-conical centriolar complex and the initial portion of theaxoneme (Figure 7d).
Mid-piece. Nine or ten mitochondria are arranged helicallyaround the 9 � 2 pattern axoneme to form the mid-pieceregion (Figure 7e,f). The plasma membrane and mitochon-
drial membrane form a double layer externally, but only themitochondrial membrane sheaths the axoneme (Figure 7d,f).Transverse sections through the mid-piece reveal that the mito-chondrial membrane surrounding the axoneme exhibits nineout-pockets, one associated with each doublet (Figure 7e). Themitochondria each exhibit highly electron-dense bodies withinthe matrix (Figure 7d–f). There does not appear to be anydefined pattern of distribution of these intra-mitochondrialbodies, although they are always clearly apparent in transverseand longitudinal sections and are easily differentiated from themitochondrial cristae (Figure 7f). At the termination of themitochondria there is a well developed membranous sheath,approximately 1.7 �m long, and consisting of four or five layersof double membranes (Figure 7g). The membranes are in con-tact with electron-dense material at both ends (Figure 7g, smallarrows; Figure 7h, arrow).
Glycogen piece and free flagellum. An electron-dense annulus clearly distinguishes the beginning of the glycogen piece fromthe mid-piece (Figure 7h). The glycogen granules are arrangedinto tracts ordered to the nine microtubule doublets (Figure7h), but these tracts are often somewhat obscured in transversesections (Figure 7i). The diameter of glycogen piece through-out most of its length varies from 0.48 to 0.54 �m, but tapersposteriorly. A free flagellum at the end was not observed.
Hydrobiidae, Bythinellinae (Figure 8)
Acrosomal complex. The acrosomal vesicle of Bythinella austriaca ismembrane-bound, measures approximately 1.2 �m in lengthand is spatulate (Figure 8a–d). At the base of the vesicle, thebroad axis measures about 0.75 �m, while the narrow axis isabout 0.35 �m. A prominent, electron-lucent apical bleb is pre-sent on the acrosomal vesicle (Figure 8a,c). The contents of thevesicle are very clear differentiated into an electron-denseouter layer (featuring angularly inclined, periodically repeat-ing, longitudinal plates) and a less dense, granular innerregion (Figure 8b–d). As demonstrated in Figure 8d, the periodically arranged plates also line the margin of acrosomalvesicle invagination. An axial rod occupies most of this invagination (Figure 8d).
Nucleus. The nucleus is solid and measures 3.8 �m in lengthand, depending on the level of section, between 0.34–0.65 �min width (Figure 8a,b). The nucleus is oval in transverse profilebasally, but markedly flattened towards the acrosome. A basalinvagination of about 0.6 �m depth contains an electron-densecentriolar complex, as well as the initial portion of the axo-neme (Figure 8f).
Mid-piece. The mid-piece measures 0.45–0.50 �m in diameterand exhibits the same structural pattern of helical, peri-axonemal mitochondria as observed in Benedictia and theBaicaliidae (Figure 8a,e). The borders of about 16 mito-chondria are not always clearly seen. A dense membrane andvesicles occur immediately posterior to the nucleus (Figure 8f).A well-developed membranous sheath (length about 2 �m)composed of about four or five double membranes in additionto the plasma membrane, encloses the axoneme following termination of the mitochondria (Figure 8g,h).
Glycogen piece and free flagellum. The annulus is well developedand appears to have a collar-like shape (Figure 8h). The glyco-gen granules are arranged distinctly into nine tracts associatedclosely with the nine axonemal doublets (Figure 8i). Diameterof the glycogen piece varies from 0.38 to 0.50 �m and tapers tothe end, leaving the axoneme of the free flagellum to beenclosed by a loose-fitting plasma membrane (Figure 8j).
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
119
12345678910123456789201234567893012345678940123456789501234567896012345678

DISCUSSION
Results of this study and of previous studies on spermatozoa ofthe Rissooidea (Giusti, 1969, 1971; Giusti & Mazzini, 1973;Kohnert, 1980; Healy, 1983b, 1988a; Kohnert & Storch, 1984;Claveria & Etges, 1988) suggest that paraspermatozoa areabsent within the superfamily. Given that paraspermatozoa arepresent in many other groups of caenogastropods including allbasal taxa (for discussion and further references see Nishiwaki,1964; Buckland-Nicks, Williams, Chia & Fontaine, 1982; Giusti& Selmi, 1982; Healy & Jamieson, 1981; Healy, 1986a,b, 1988a,1993, 1996a; Hodgson, 1997; Buckland-Nicks, 1998) it seems
likely that these cells have been lost within the Rissooidea. Thephenomenon of euspermatozoal dimorphism, known in onespecies of Bithyniidae (Bithynia tentaculata; see Kohnert, 1980),has not been observed in other Rissooidea including thosespecies dealt with in the present study. Such dimorphism hasalso been demonstrated in one species of Neritoidea (Tochi-moto, 1967), two species of Janthinoidea (Nishiwaki & Tochi-moto, 1969) and one cerithioidean (Healy, 1982c), but itsexact significance is, as yet, impossible to gauge, given its sporadic occurrence among the Caenogastropoda. It may, assuggested by various authors (Ankel, 1933; Nishiwaki, 1964;Tochimoto, 1967; Jamieson, 1987) represent an initial step
P. RÖPSTORF ET AL .
120
Figure 7. Euspermatozoa of Lithoglyphus naticoides. (a) Longitudinal section of head (acrosomal complex, nucleus) and beginning of mid-piece. (b) Longi-tudinal section through the narrow axis of acrosomal complex; note accessory membranes marked by arrows. (c) Longitudinal section through the broad axisof acrosomal complex. (d) Transverse section through nucleus at the level of the centriolar complex. (e) Transverse section of mid-piece; mitochondria withdense intra-mitochondrial bodies. (f) Longitudinal section of mid-piece; mitochondria with dense intra-mitochondrial bodies. (g) Longitudinal section ofmembranous part and beginning of the glycogen piece. (h) Longitudinal section through periphery of annulus and glycogen piece. (i) Transverse section ofglycogen piece. Scale lines: 7a � 1 �m; all other figures � 0.5 �m. Abbreviations: a, acrosome; ab, apical bleb; am, accessory membrane; an, annulus; ar, axialrod; av, acrosomal vesicle; ax, axoneme; bp, basal plate; gp, glycogen piece; mp, mid-piece; n, nucleus; ?, inclusion in apical bleb—probably artefact.

toward profound sperm polymorphism (for further discussionof paraspermatozoan evolution, see Jamieson, 1987).
Acrosomal complexTypically most prosobranchs have a truly conical acrosomalvesicle, although there is substantial variation between taxa asto its length and the depth of the basal invagination (seeKohnert & Storch, 1984; Koike, 1985; Healy, 1988a, 1996a,b).
The spatula-like shape of the acrosomal vesicle appears to becommon to other rissooideans such as the Hydrobiidae (Giusti,1969, 1971; Kohnert & Storch, 1984), Bithyniidae (Kohnert,1980; Kohnert & Storch, 1984), Pyrgulidae (Riedel, Healy,Röpstorf & Sitnikova, 2001) and Stenothyridae (Healy, 1983b),but elsewhere within the Caenogastropoda has also beenrecorded in the Cerithioidea (Giusti, 1971; Hachiri & Higashi,1971; Healy, 1983a; Kohnert & Storch, 1984; Koike, 1985)
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
121
12345678910123456789201234567893012345678940123456789501234567896012345678
Figure 8. Euspermatozoa of Bythinella austriaca. (a) Longitudinal section through the narrow axis of acrosomal complex and nucleus; accessory membranesmarked by arrow. (b) Longitudinal section through the broad axis of acrosomal complex and nucleus. (c) Longitudinal section through the narrow axis ofacrosomal complex. (d) Transverse section of acrosomal complex near the base. (e) Transverse section of the mid-piece. (f) Longitudinal section throughnucleus-mid-piece junction vesicles (arrows). (g) Longitudinal section through the membranous part at the end of the mid-piece. (h) Longitudinal sectionthrough the membranous part, the annulus (arrow) and the beginning of the glycogen piece. (i) Transverse section of glycogen piece, granules distinctlyarranged in nine tracts. (j) End piece in transverse section. Scale lines: 8a,b � 1 �m; all other figures � 0.5 �m. Abbreviations: ab, apical bleb; am, accessorymembrane; an, annulus; ar, axial rod; av, acrosomal vesicle; ax, axoneme; bp, basal plate; gp, glycogen piece; mp, mid-piece; n, nucleus.

and Cyclophoroidea (Selmi & Giusti, 1980; Koike, 1985). Inmany caenogastropods (including some Rissooidea–Rissoidae)only the apical area of the acrosomal vesicle is flattened (seeKohnert & Storch, 1984, and Koike, 1985, for examples). Inthose rissooidean families showing a flattened acrosomal vesicle(Baicaliidae, Benedictiidae, Pyrgulidae, Hydrobiidae, Bithyn-iidae, Stenothryidae) the transverse profile may differ betweenfamilies or subfamilies. Of these families, the Stenothyridaeunquestionably show the most complex acrosomal vesicle(Healy, 1983b). Within the Hydrobiidae, the basal invaginationof the acrosomal vesicle is flattened in Pseudamnicola sp.(Hydrobiinae, Giusti, 1971) and oval in Bythinella austriaca(Bythinellinae, present study). The profile of the basal invagi-nation is circular in the ‘Benedictiidae’ (present study), Bithyn-iidae (Kohnert, 1980), Stenothyridae (Healy, 1983b), andBaicaliidae and Pyrgulidae, and almost certainly is the plesio-morphic state for this feature.
The plate-like structures within the acrosomal vesicle appearto be typical of the acrosomes of all caenogastropod proso-branchs (Healy, 1996a). Various degrees of differentiation ofthe acrosomal vesicle contents occur in ‘archaeogastropod’prosobranchs, notably zonation of the contents, e.g. in severalvetigastropods (Hodgson & Foster, 1992; Healy, Beames &Barclay, 1998) and patellogastropods (Hodgson & Bernard,1988). In heterodont bivalves, the relatively electron-densebasal ring zone of the acrosomal vesicle physically binds withexposed microvillar tufts on the egg surface as a necessary stepin the fertilization process (Hylander & Summers, 1977).Possibly the plate-like structures of rissooideans and othercaenogastropods play an important role in the pre-fertilizationphenomenon known as the acrosome reaction, but as yet thisevent has never been examined in any internally fertilizing gastropod.
The presence of an apical bleb in the acrosomal vesicle iswidespread within the Caenogastropoda, but evidently absentin the Cerithioidea, Cyclophoroidea, and Ampullarioidea (seeHealy, 1996a, and references therein). The function of thisbleb is not known, but its position at the apex of the spermato-zoon suggests that it may have a role in the acrosome reactionor even in direct sperm-egg contact.
The electron-dense, accessory membrane (‘ragged mem-brane’ of some authors) here observed in all Baicaliidae and‘Benedictiidae‘, has been reported before in many caeno-gastropod superfamilies (e.g. the Campaniloidea, Littorin-oidea, Loxonematoidea, Stromboidea, Vermetoidea, Cyprae-oidea, Janthinoidea, Muricoidea, Conoidea; Walker &MacGregor, 1968; Koike & Nishiwaki, 1980; Buckland-Nicks et al., 1982; Koike, 1985; Healy, 1986a,b, 1988a,b, 1989, 1992;Healy & Jamieson, 1993). As yet, no function has been ascribedto this membrane, but its positioning would suggest some rolein the acrosome reaction. Although not previously identified ineuspermatozoa of any member of the Rissooidea, an accessorymembrane was also found in the hydrobiids Bythinella austriaca,Lithoglyphus naticoides (this paper), as well as in the pyrgulidPyrgula annulata (Riedel, Healy, Röpstorf & Sitnikova, 2001).This feature can be clearly seen also on a micrograph ofTruncatella subcylindrica presented by Giusti & Mazzini (1973,figure 6). In another micrograph from Giusti & Mazzini’saccount (1973, figure 5) the possible precursor of the accessorymembrane in a late euspermatid is referred to as the ‘remainsof Golgi cisterns’. As we have not investigated spermiogenesisof the Baicaliidae or the ‘Benedictiidae‘ we cannot evaluate this suggestion, but it does at least seem feasible and certainlyworthy of further investigation. The accessory membrane of all investigated baicaliids and benedictiids is poorly developed,as is also the case in some other neotaenioglossans (Strom-boidea; Koike & Nishiwaki, 1980; Koike, 1985) and manyneogastropods (see Koike, 1985; Healy, 1988a). In several
caenogastropod taxa (e.g. the Campaniloidea, Littorinoidea,Vermetoidea, Cypraeoidea, Janthinoidea) the membrane isvery well developed, and sheaths between one-third and one-half of the acrosomal vesicle (Koike, 1985; Healy, 1986a,b,1988a,b, 1996a,b).
NucleusTwo main types of nuclear morphology occur within theCaenogastropoda (and also the Heterobranchia)—the short,solid nucleus (with shallow basal invagination for the insertionof the centriole/axoneme complex) and the long, tubularnucleus (with centriole/axoneme complex effectively travers-ing the full length of the nucleus; for several examples seeKohnert & Storch, 1984; Koike, 1985; Healy, 1988a, 1990,1996a). Typically, most caenogastropod families show one orthe other style of nuclear shape, but it is known that in certainfamilies both styles may occur in a single family (e.g. Thiaridae,Littorinidae, Eulimidae; see Healy, 1996b, for discussion),which to some extent diminishes the significance of nuclearmorphology as a taxonomically important character at andabove the family level. The precise reason for the dichotomy innuclear shape within the Caenogastropoda has yet to be identi-fied, but Healy (1996b) has suggested that sperm storage con-straints may be a controlling factor. Within the Rissooidea, theshort, solid nucleus occurs in the Hydrobiidae, Bithyniidae,Pyrgulidae, Truncatellidae, Stenothyridae, Pomatiopsidae, andIravadiidae (Giusti, 1969, 1971; Giusti & Mazzini, 1973; Kohnert,1980; Kohnert & Storch, 1984; Claviera & Etges, 1988; Healy,1983b; present study), whereas the long, tubular nucleusoccurs in the Rissoidae and Baicaliidae (Kohnert & Storch,1984; present study).
Mid-pieceIn almost all the rissooideans examined to date, as in a largeproportion of other caenogastropods, the peri-axonemal mito-chondria are helical and contain irregularly arranged, tubularcristae (Walker & MacGregor, 1968; Giusti, 1969, 1971; Giusti& Mazzini, 1973; Buckland-Nicks, 1973; Buckland-Nicks &Chia, 1976; Buckland-Nicks et al., 1982; Kohnert & Storch,1984; Koike, 1985; Claveria & Etges, 1988; Healy, 1986b,1988a,b, 1996b). There are a number of exceptions, the mostnotable being that group of superfamilies with precision-arranged arrays of parallel crystal plates, namely the Cerithi-oidea (with four straight mitochondria; Guisti, 1971; Healy &Jamieson, 1981; Healy, 1982c, 1983a, 1996a; Afzelius & Dallai,1983; Kohnert & Storch 1984; Koike 1985; Hodgson & Heller,1990), the Cyclophoroidea (with seven or eight straight mito-chondria with paracrystalline deposits; Selmi & Guisti, 1980;Healy, 1988a, 1996a) and the Vivipariidae (of the Ampul-larioidea, with four helical mitochondria; Ishizaki & Kato, 1958;Kaye, 1958; Gall, 1961; Hachiri & Higashi, 1972; Griffond, 1980;Healy, 1996a).
Within the Rissooidea there are dissappointingly few mid-piece differences between the various investigated families, thenotable exception being the Stenothyridae, which show helicalkeels, helical compartments, and linear ‘paracrystalline’ struc-tures (helical fibrils) between the mitochondria (Healy,1983b). Among the species examined in the present study, themid-piece mitochondria of Lithoglyphus naticoides (Hydro-biidae, Lithoglyphinae) alone exhibit prominent, electron-dense, intra-mitochondrial bodies. This character, togetherwith the difference in shape of the nucleus, suggests a closerrelationship between Benedictia and Hydrobia (see Kohnert &Storch, 1984) than between Benedictia and Lithoglyphus. Micro-graphs presented by Giusti (1971) suggest the possible pres-ence of these dense bodies in Bythinella cf. opaca (Hydrobiidae,Bythinellinae), but no evidence of such structures could beseen in our material of Bythinella austriaca.
P. RÖPSTORF ET AL .
122

The membranous sheath at the end of the mid-piece occursin all the Baicaliidae, ‘benedictiids’ and hydrobiids examinedherein, and in the pyrgulid species Pyrgula annulata (Riedel,Healy, Röpstorf & Sitnikova, 2001). According to Kohnert(1980) the sheath in Bithynia tentaculata (Bithyniidae) arisesbetween two dense ring structures late in euspermiogenesis,and is a product of mitochondrial metamorphosis and, hence,strictly a feature of the mid-piece, rather than a discrete eusper-matozoan zone. Our data would support this view, but note that the tendency of the membranes to fuse at anterior and pos-terior extremities makes it difficult to clearly demarcate membranes from the dense rings in mature euspermatozoa.However, TEM micrographs of Kohnert (1980) unequivocallydemonstrate the presence of two rings in late euspermatids(the anterior ring being best developed), and there are nogrounds for suspecting that this is not typical of the Rissooideain general. While the posterior ring can be accepted as the‘annulus’ observed in other caenogastropods, the origin, andtherefore the homology of the anterior ring remains uncertain,and can only be determined by detailed studies of lateeuspermiogenesis.
We believe the membranous sheath to be a taxonomicallysignificant feature. It has previously been recorded in anotherhydrobiid (Hydrobia sp.; Kohnert & Storch, 1984), the Bithyni-idae (Bithynia tentaculata, B. leachi; Kohnert, 1980; Kohnert &Storch, 1984) and the Stenothyridae (Stenothyra sp.; Healy,1983b). The sheath does not occur in the Truncatellidae (seeGiusti & Mazzini, 1973) and was not reported by Kohnert &Storch (1984) in any of the three species of Rissoidae studiedby them. There is every reason to believe that the membranesheath occurs in only one subsection of the Rissooidea, present-ly consisting of the Baicaliidae � ‘Benedictiidae’ � Hydro-biidae � Pyrgulidae � Bithyniidae � Stenothyridae, and weregard this feature as a synapomorphy of this assemblage(other families may join this group when adequate sperm dataare available). As with several other distinctive features (helicalcompartments, secondary keels and inter-mitochondrial fibrils;see Healy, 1983b), the Stenothyridae is distinguishable fromthe Baicaliidae, ‘Benedictiidae’, Hydrobiidae, Pyrgulidae, andBithyniidae by having the membrane sheath enclosed also bythe mitochondrial sheath (in the other five families the mito-chondria do not enclose the membrane sheath).
Regarding the function of the sheath, nothing can as yet bededuced. It may perhaps have some role in enhancing the flexi-bility of the glycogen piece by essentially extending the dis-tance between the end of the mitochondrial sheath and thebeginning of the glycogen piece, although in the Stenothyridaethe sheath is enclosed by the mitochondria (see Healy, 1983b).
It has been suggested that the annulus plays an importantrole in the locomotion of Littorina sperm (Buckland-Nicks &Chia, 1981). Although the membranous sheath appears to liein contact with the annulus, it is unlikely that the sheath itself(which consists only of membranes) is directly associated withaxonemal motility.
Glycogen piece and free flagellumExtensive peri-axonemal glycogen deposits are characteristicnot only of Caenogastropoda, but also many Heterobranchia(see Giusti, 1971; Thompson, 1973; Maxwell, 1980; Healy,1988a, 1996a) and certain cephalopods (Octopoda; see Longo& Anderson, 1970; Maxwell, 1974; Healy, 1996a) and is pre-sumably associated with the metabolic requirements of long-term sperm storage (Maxwell, 1980; but see Giusti & Selmi,1985, for further discussion). In most caenogastropods and afew heterobranchs (some Architectonicidae; see Healy &Jamieson, 1991), the glycogen granules are arranged in ninetracts correlated with the nine axonemal doublets, which maysuggest a direct correlation with sperm motility. The rather
indistinct nature of the nine granular tracts in most of thespecies examined by us is not especially significant, as this hasbeen observed in several other groups of caenogastropods (seeKohnert & Storch, 1984; Koike, 1985). However, in Truncatellasubcylindrica discrete granular tracts are not distinguishable andGiusti & Mazzini (1973) utilized this feature, together with the short conical acrosome and the helically-keeled nucleus ofT. subcylindrica, to further differentiate the Truncatellidae fromthe Hydrobiidae. Similarly, the Stenothyridae can be disting-uished from other Rissooidea (and all other gastropods) bytheir unique organization of the glycogen piece in which theaxoneme is enveloped by a thick, continuous sheath of glyco-gen rosettes and a thin, outer sheath of glycogen rosettes (separ-ated from the thick sheath by a membrane; Healy, 1983b).
Giusti (1971) demonstrated the presence of nine well devel-oped coarse fibres associated with the nine axonemal doubletsin the terminal or free flagellum region in the hydrobiidsBelgrandia caprai and Hydrobia acuta. Similar coarse fibres werealso observed by Healy (1983b) in Stenothyra sp. (Stenothyri-dae), but in the neck region (basal invagination of nucleus andanterior-most region of mid-piece). Although we did notobserve axoneme-associated coarse fibres in the present study,this may be due in part to the lack of favourable sections throughthis region or quite possibly the highly localized nature of thefibres within the euspermatozoa.
Taxonomic and phylogenetic considerationsResults of the present study and those of previous authors(Giusti, 1969, 1971; Giusti & Mazzini, 1973; Kohnert, 1980;Healy, 1983b; Kohnert & Storch, 1984; Claveria & Etges, 1988)have demonstrated that comparative euspermatozoan ultra-structure sheds new light on taxonomic and phylogenetic questions within the Rissooidea.
Using nuclear similarities it could be argued that theBaicaliidae are more closely affiliated with the Rissoidae (alsoshowing elongate, tubular eusperm nucleus), than with theshort-nuclei rissooidean families (e.g. Hydrobiidae, ‘Benedicti-idae’, Bithyniidae, Pyrgulidae, Stenothyridae, Truncatellidae).However, given the limitations of nuclear morphology as a taxonomically reliable feature at the family level (discussedabove) and, more importantly, the occurrence of the highlyunusual, membranous sheath within the mid-piece of theBaicaliidae (associating this family with the Hydrobiidae,‘Benedictiidae’, Bithyniidae, Pyrgulidae, and Stenothyridae,but not with the Truncatellidae; see discussion below), abaicaliid link with the Rissoidae seems unsupportable. Never-theless, the consistent presence of the long tubular nucleus inall species of the Baicaliidae, together with radular and nervoussystem features (Kozhov, 1945, 1951) does at least offer someadditional support for the continued recognition of this family.Concerning the possibility of a close relationship between thePyrgulidae and the Baicaliidae (e.g. see Thiele, 1929), it canonly be stated that the two families differ profoundly in nuclearmorphology, i.e. a short, solid, compressed profile in Pyrgula(Riedel, Healy, Röpstorf & Sitnikova, 2001) versus a long, tubular, circular profile in Baicaliidae; present study), but otherwise do not show marked differences. Keeping in mindthe variability of nuclear morphology in some caenogastropodfamilies (e.g. see Healy, 1996b), the precise nature of the Pyrgulidae–Baicaliidae relationship remains uncertain. As for the taxonomic placement of Benedictia, eusperm ultra-structure does not provide any defining feature at either family(Benedictiidae) or subfamily level (Benedictiinae). This is also true for the Bithyniidae (compare with results of Kohnert,1980; Kohnert & Storch, 1984) and Pyrgulidae (Riedel, Healy, Röpstorf & Sitnikova, 2001), but for the present we suggest continued recognition of at least the subfamilyBenedictiinae within the Hydrobiidae. By contrast, differences
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
123
12345678910123456789201234567893012345678940123456789501234567896012345678

in eusperm nuclear and mid-piece structure between Litho-glyphus naticoides and other investigated hydrobiids, includingBenedictia spp. (Giusti, 1969, 1971; Kohnert & Storch, 1984; present study) suggest that the Lithoglyphinae may eventuallyprove worthy of familial status.
The scroll-like arrangement of double membranes (mem-brane sheath) positioned between the terminal portion of themitochondria and the annulus was first detected by Kohnert(1980) in the Bithyniidae (see also Kohnert & Storch, 1984),and subsequently recorded in the Stenothyridae (Healy, 1983b)and Hydrobiidae (Hydrobiinae; Kohnert & Storch, 1984). Ourstudy has demonstrated the presence of this specialized featurein the Baicaliidae and further members of the Hydrobiidae(subfamilies Bythinellinae, Lithoglyphinae and Benedictiinae,which is here recognized as such) and it also occurs in thePyrgulidae (Riedel, Healy, Röpstorf & Sitnikova, 2001). As thissheath is unknown elsewhere within the Gastropoda (includingrissooidean families Truncatellidae, Rissoidae, and possiblyPomatiopsidae; Giusti & Mazzini, 1973; Kohnert & Storch,1984; Claviera & Etges, 1988), we consider it a very usefulsynapomorphy for the assemblage of families that exhibit it(Hydrobiidae � Bithyniidae � Stenothyridae � Pyrgulidae �Baicaliidae).
ACKNOWLEDGEMENTS
The authors would like to thank: the Director of the Limno-logical Institute of the Siberian Branch of the Russian Academyof Sciences, Mikhail Aleksandrovich Grachev, for organizingthe expedition; Professor Dr Klaus Hausmann, for the use ofhis laboratory, TEM, and ultramicrotomes; and MonikaBulang-Lörcher, for preparing the drawings. This investigationwas mainly financed by the Deutsche Forschungsgemeinschaft(grant Ri 809/1–2) and the Russian Foundation for BasicResearch (grants 96–04–63024 and 98–04–49276). A SeniorResearch Fellowship and research grant from the AustralianResearch Council made possible the participation of JMH inthis project.
REFERENCESAFZELIUS, B.A. & DALLAI, R. 1983. The paired spermatozoa of the
marine snail, Turritella communis Lamarck (Mollusca, Mesogastro-poda). Journal of Ultrastructure Research, 85: 311–319.
ANKEL, W.A. 1933. Untersuchungen über Keimzellenbildung undBefruchtung bei Bithynia tentaculata L. II. Gibt es in der Spermato-genese von Bithynia tentaculata eine Polymegakie? Zeitschrift fürZellforschung und Mikroskopische Anatomie, 17: 160–198.
BOSS, K.J. 1982. Mollusca. In: Synopsis and Classification of Living Organ-isms (S.P. Parker, ed.), Vol. 2, 945-1166. McGraw-Hill, New York.
BUCKLAND-NICKS, J.A. 1973. The fine structure of the spermatozoanof Littorina (Gastropoda: Prosobranchia), with special reference tosperm motility. Zeitschrift für Zellforschung, 144: 111–129.
BUCKLAND-NICKS, J.A. 1998. Prosobranch parasperm: sterile germcells that promote paternity? Micron, 29: 267–280.
BUCKLAND-NICKS, J.A. & CHIA, F.S. 1976. Spermatogenesis of a marine snail, Littorina sitkana. Cell and Tissue Research, 170:455–475.
BUCKLAND-NICKS, J.A. & CHIA, F.S. 1981. Locomotion of the fili-form spermatozoan of Littorina (Gastropoda, Prosobranchia). Celland Tissue Research, 219: 27–39.
BUCKLAND-NICKS, J.A., WILLIAMS, D., CHIA, F.S. & FONTAINE, A.1982. The fine structure of the polymorphic spermatozoa ofFusitriton oregonensis (Mollusca: Gastropoda), with notes on the cyto-chemistry of the internal secretions. Cell and Tissue Research, 227:235–255.
CLAVERIA, F.G. & ETGES, F.J. 1988. Spermatogenesis in Oncomelaniahupensis quadrasi, a molluscan host of Schistosoma japonicum. Malaco-logia, 28: 81–94.
CLESSIN, S. 1878. Referat über Dybowski, Die Gasteropodenfauna desBaikalsees. Malacozoologische Blätter, 25: 132–139.
CLESSIN, S. 1880. Studien über die Familie der Paludinen. Malaco-zoologische Blätter, NF, 2: 161–196.
DADDOW, L.Y.M. 1986. An abbreviated method of double lead staintechnique. Journal of Submicroscopic Cytology, 18: 221–224.
DALL, W.H. 1877. Note on ‘Die Gasteropodenfauna des Baikal-Sees’.Proceedings of the Boston Society of Natural History, 19: 43–47.
DYBOWSKI, W. 1875. Die Gasteropodenfauna des Baikal-Sees, Anato-misch und systematisch bearbeitet. Memoires de L´ Academie Imperialedes Sciences de St. Petersbourg, 22/8: 1–73.
FISCHER, P. 1887. Manuel de Conchyliologie et de Paléontologie Conchy-liologique ou Histoire Naturelle des Mollusques vivants et fossiles. F. Savy,Paris.
GALL, J.G. 1961. Centriole replication. A study of spermatogenesis inthe snail Viviparus. Journal of Biophysical and Biochemical Cytology, 10:163–193.
GIUSTI, F. 1969. The spermatozoon of a freshwater prosobranch mol-lusc. Journal of Submicroscopic Cytology, 1: 263–273.
GIUSTI, F. 1971. L’ultrastruttura dello spermatozoo nella filogenesi enella sistematica dei molluschi gasteropodi. Atti della Societa Italianadi Scienze Naturali e Museo Civico di Storia Naturale Milano, 112:381–402.
GUISTI, F. & MAZZINI, M. 1973. The spermatozoon of Truncatella(s. str.) subcylindrica (L.) (Gastropoda: Prosobranchia). MonitoreZoologica Italiano, 7: 181–201.
GIUSTI, F. & SELMI, M.G. 1982. The atypical sperm in the proso-branch molluscs. Malacologia, 22: 171–181.
GIUSTI, F. & SELMI, M.G. 1985. The seminal receptacle and spermstorage in Cochlostoma montanum (Issel) (Gastropoda: Proso-branchia). Journal of Morphology, 184: 121–133.
GRIFFOND, B. 1980. Étude ultrastructurale de la spermatogenèse typique de Viviparus viviparus (L.), Mollusque Gastéropode. Archivesde Biologie (Bruxelles), 91: 445–462.
HACHIRI, S. & HIGASHI, S. 1971. Spermiogenesis in the melaniansnails Semisulcospira decipiens and Semisulcospira niponica. Memoirs ofthe Faculty of Education, Shiga University, 21: 43–51.
HACHIRI, S. & HIGASHI, S. 1972. Utilization of glycogen in spermato-zoa of pond snails, Sinotaia histrica and Heterogen longispira. Memoirsof the Faculty of Education, Shiga University, 22: 43–57.
HEALY, J.M. 1982a. An ultrastructural examination of developing andmature euspermatozoa in Pyrazus ebeninus (Mollusca, Gastropoda,Potamididae). Zoomorphology, 100: 157–175.
HEALY, J.M. 1982b. Ultrastructure of the spermiogenesis of Philippia(Psilaxis) oxytropis with special reference to the taxonomic positionof the Architectonicidae (Gastropoda). Zoomorphology, 101:197–214.
HEALY, J.M. 1982c. Ultrastructure of paraspermatozoa, euspermatozoaand eusperm-like spermatozoa of Obtortio cf. fulva (Prosobranchia:Cerithiacea). Helgoländer Meeresuntersuchungen, 35: 489–500.
HEALY, J.M.1983a. Ultrastructure of euspermatozoa of cerithiaceangastropods (Prosobranchia: Mesogastropoda). Journal of Morphology,178: 57–75.
HEALY, J.M. 1983b. Ultrastructure of euspermiogenesis in the meso-gastropod Stenothyra sp. (Prosobranchia, Rissoacea, Stenothyridae).Zoologica Scripta, 12: 203–214.
HEALY, J.M. 1983c. An ultrastructural study of basommatophoranspermatozoa (Mollusca, Gastropoda). Zoologica Scripta, 12: 57–66.
HEALY, J.M. 1986a. Euspermatozoa and paraspermatozoa of a relictcerithiacean gastropod Campanile symbolicum (Prosobranchia, Meso-gastropoda, Campanilidae). Helgoländer Meeresuntersuchungen, 40:201–218.
HEALY, J.M. 1986b. An ultrastructural study of euspermatozoa, para-spermatozoa and nurse cells of the cowrie Cypraea errones (Gastro-poda, Prosobranchia, Cypraeidae). Journal of Molluscan Studies, 52:125–137.
HEALY, J.M. 1988a. Sperm morphology and its systematic importancein the Gastropoda. In: Prosobranch Phylogeny (ed. W. F. Ponder).Malacological Review (Supplement), 4: 251–266.
P. RÖPSTORF ET AL .
124

HEALY, J.M. 1988b. Sperm morphology in Serpulorbis and Dendropomaand its relevance to the systematic position of the Vermetidae(Gastropoda). Journal of Molluscan Studies, 54: 295–308.
HEALY, J.M. 1989. Spermatozeugmata of Abyssochrysos: ultrastructure,development and relevance to the systematic position of the Abysso-chrysidae (Prosobranchia, Caenogastropoda). Bulletin du MuséumNational D’Histoire Naturelle. 4. e sér. (Sect.A, No. 3), 11: 509–533.
HEALY, J.M. 1990. Spermatozoa and spermiogenesis of Cornirostra,Valvata and Orbitestella (Gastropoda: Heterobranchia) with a discus-sion of valvatoidean sperm morphology. Journal of Molluscan Studies,56: 557–566.
HEALY, J.M. 1992. Dimorphic spermatozoa of the hydrothermal ventprosobranch Alviniconcha hessleri: systematic importance and com-parison with other caenogastropods. Bulletin du Muséum Nationald’Histoire Naturelle (Paris) 4 sér. A, 14: 273–291.
HEALY, J.M. 1993. Transfer of the gastropod family Plesiotrochidae tothe Campaniloidea based on sperm ultrastructural evidence. Journalof Molluscan Studies, 59: 135–147.
HEALY, J.M. 1996a. Molluscan sperm ultrastructure: correlation withtaxonomic units within the Gastropoda, Cephalopoda and Bivalvia.In: Origin and evolutionary radiation of the Mollusca (ed. J. Taylor),99–113. Oxford University Press.
HEALY, J.M. 1996b. An ultrastructural study of euspermatozoa inBembicium auratum including a comparison with other Caeno-gastropoda, especially Littorinoidea. Journal of Molluscan Studies, 62:57–63.
HEALY, J.M. & JAMIESON, B.G. 1981. An ultrastructural examinationof developing and mature paraspermatozoa in Pyrazus ebeninus(Mollusca: Gastropoda, Potamididae). Zoomorphology, 98: 101–119.
HEALY, J.M. & JAMIESON, B.G.M. 1991. Ultrastructure of spermio-genesis in the gastropod Heliacus variegatus (Architectonicidae)with description of a banded peri-axonemal helix. Marine Biology,109: 67–77.
HEALY, J.M. & JAMIESON, B.G.M. 1993. Euspermatozoa, parasperma-tozoa and spermatozeugmata of Littorina (Palustorina) articulata(Prosobranchia: Caenogastropoda) with special reference to thepseudotrich. Acta Zoologica (Stockholm), 74: 321–330.
HEALY, J.M. & WILLAN, R.C. 1984. Ultrastructure and phylogeneticsignificance of notaspidean spermatozoa (Mollusca, Gastropoda,Opisthobranchia). Zoologica Scripta, 13: 107–120.
HEALY, J.M., BEAMES, K.P. & BARCLAY, D.B. 1998. Spermatozoa ofthe Australian ´greenlip´ abalone Haliotis laevigata Donovan: ultra-structure and comparison with other gastropods, especially otherHaliotidae (Vetigastropoda, Mollusca). Invertebrate Reproduction andDevelopment, 34: 197–206.
HODGSON, A.N. 1997. Paraspermiogenesis in gastropod molluscs.Invertebrate Reproduction and Development, 31: 31–38.
HODGSON, A.N. & BERNARD, R.T.F. 1988. A comparison of thestructure of the spermatozoa and spermatogenesis of 16 species ofpatellid limpets (Mollusca: Gastropoda: Archaeogastropoda).Journal of Morphology, 195: 205–223.
HODGSON, A.N. & CHIA, F-S. 1993. Spermatozoon structure of someNorth American prosobranchs from the families Lottiidae (Patello-gastropoda) and Fissurellidae (Archaeogastropoda). Marine Biology,116: 97–101.
HODGSON, A.N. & FOSTER, G.G. 1992. Structure of the sperm ofsome South African archaeogastropods (Mollusca) from the super-families Haliotoidea, Fissurelloidea and Trochoidea. Marine Biology,113: 89–97.
HODGSON, A.N. & HEALY, J.M. 1998. Comparative sperm morph-ology of the pulmonate limpets Trimusculus costatus, T. reticulatus(Trimusculidae) and Burnupia stenochorias and Ancylus fluviatilis(Ancylidae). Journal of Molluscan Studies, 64: 447–460.
HODGSON, A.N. & HELLER, J. 1990. Spermatogenesis and spermstructure of the normally parthenogenetic freshwater snail Melan-oides tuberculata. Israel Journal of Zoology, 37: 31–50.
HODGSON, A.N., HELLER, J. & BERNARD, R.T.F. 1990. Ultra-structure of the sperm and spermatogenesis in five South Africanspecies of the trochid genus Oxystele (Mollusca, Prosobranchia).Molecular Reproduction and Development, 25: 263–271.
HODGSON, A.N., BERNARD, R.T.F. & LINDLEY, D.S. 1991. Com-parative spermatology of four sympatric species of Siphonaria (Pul-monata: Basommatophora). Journal of Molluscan Studies, 57: 309–322.
HODGSON, A.N., RIDGWAY, S., BRANCH, G.M. & HAWKINS, S.J.1996. Spermatozoan morphology of 19 species of prosobranchlimpets (Patellogastropoda) with a discussion of patellid relation-ships. Philosophical Transactions of the Royal Society of London, B, 351:339–347.
HODGSON, A.N., HEALY, J.M. & TUNNICLIFFE, V. 1997. Spermato-genesis and sperm structure of the hydrothermal vent prosobranchgastropod Lepetodrilus fucensis (Lepetodrilidae, Mollusca). Inverte-brate Reproduction and Development, 31: 87–97.
HYLANDER, B.L. & SUMMERS, R.G. 1977. Ultrastructural analysis ofthe gametes and early fertilization in two bivalve molluscs Chamamacerophylla and Spisula solidissima with special reference to gametebinding. Cell and Tissue Research, 182: 469–489.
ISHIZAKI, T. & KATO, K. 1958. The fine structure of atypical sperma-tozoa of the pond snail Viviparus malleatus. Zoological Magazine(Tokyo), 67: 286–295.
JAMIESON, B.G.M. 1987. A biological classification of sperm types,with special reference to annelids and molluscs, and an example ofspermiocladistics. In: New Horizons of Sperm Cell Research (ed. H.Mohri), 311–332. Japanese Scientific Society Press, Tokyo.
JAMIESON, B.G.M., HODGSON, A.N. & BERNARD, R.T.F. 1991.Phylogenetic trends and variation in the ultrastructure of the spermatozoa of sympatric species of South African patellid limpets(Archaeogastropoda, Mollusca). Invertebrate Reproduction and Develop-ment, 20: 137–146.
KAYE, J.S. 1958. Changes in the fine structure of mitochondria duringspermatogenesis. Journal of Morphology, 102: 347–369.
KOHNERT, R. 1980. Zum Spermiendimorphismus der Prosobranch-ier: Spermiogenese und ultrastruktureller Aufbau der Spermienvon Bithynia tentaculata (L.). Zoologischer Anzeiger, 205: 145–161.
KOHNERT, R. & STORCH, V. 1984. Vergleichend-ultrastrukturelleUntersuchungen zur Morphologie eupyrener Spermien der Monotocardia (Prosobranchia). Zoologische Jahrbücher Anatomie, 111:51–93.
KOIKE, K. 1985. Comparative ultrastructural studies on the spermato-zoa of the Prosobranchia (Mollusca: Gastropoda). Science Report ofthe Faculty of Education, Gunma University, 34: 33–153.
KOIKE, K. & NISHIWAKI, S. 1980. The ultrastructure of dimorphicspermatozoa in two species of the Strombidae (Gastropoda, Proso-branchia). Venus, 38: 259–274.
KOZHOV, M.M. 1936. Mollyuski ozera Baikala (Molluscs of LakeBaikal). Trudy Baikalskoy Limnol Stantsii, 8: 1–320. [In Russian, partin German.]
KOZHOV, M.M. 1945. K morfologii endemichnykh mollyuskov ozeraBaikal (Contributions to the morphology of endemic molluscs ofLake Baikal). 1. Benedictiinae Gastropoda, Mesogastropoda. 2.Male reproductive organs. Zoologicheskiy Zhurnal, 24/ 5: 277–290.[In Russian.]
KOZHOV, M.M. 1951. K morfologii i istorii baikal´skikh endemichnikhmollyuskov sem. Baicaliidae Gastropoda, Prosobranchia (On themorphology and history of the Baikal endemic molluscs of the family Baicaliidae). Trudy Baikalskoy Limnol. Stantsii., 13: 93–119. [InRussian.]
KOZHOV, M.M. 1963. Lake Baikal and its Life. Monographiae Biologicae,11: 1–344.
LINDHOLM, W.A. 1909. Die Mollusken des Baikalsees. Wissenschaft-liche Ergebnisse einer Zoologischen Expedition nach dem Baikal-See (Kiewund Berlin), 4: 1–104.
LONGO, F.J. & ANDERSON, E. 1970. Structural and cytochemical fea-tures of the sperm of the cephalopod Octopus bimaculatus. Journal ofUltrastructure Research, 32: 94–106.
MARTENS, E., VON. 1876. Über “W. Dybowski, Die Gasteropoden-Fauna des Baikal-Sees”. Jahrbücher der deutschen malakozoologischenGesellschaft, 3: 181–184.
MAXWELL, W.L. 1974. Spermiogenesis of Eledone cirrhosa Lamarck(Cephalopoda, Octopoda). Proceedings of the Royal Society of London,B, 186: 181–190.
SPERM ULTRASTRUCTURE OF BAILKALIAN GASTROPODS
125
12345678910123456789201234567893012345678940123456789501234567896012345678

MAXWELL, W.L. 1980. Distribution of glycogen deposits in two euthy-neuran sperm tails. International Journal of Invertebrate Reproduction, 2:245–249.
NISHIWAKI, S. 1964. Phylogenetical study on the type of the dimor-phic spermatozoa in Prosobranchia. Science Reports of the TokyoKyoiku Daigaku, Section B, 11: 237–275.
NISHIWAKI, S. & TOCHIMOTO, T. 1969. Dimorphism in typical andatypical spermatozoa forming two types of spermatozeugmata intwo epitoniid prosobranchs. Venus (Japanese Journal of Malacology),28: 37–49.
PONDER, W.F. 1988. The truncatelloidean (� rissooidean) radia-tion—a preliminary phylogeny. In: Prosobranch Phylogeny (W. F.Ponder, ed.), Malacological Review Supplement, 4: 129–164.
PONDER, W. & WARÉN, A. 1988. Classification of the Caenogastro-poda and Heterostropha—a list of the family-group names andhigher taxa. In: Prosobranch Phylogeny (W. F. Ponder, ed.), Malaco-logical Review Supplement, 4: 288–326.
READER, T.A.J. 1973. Histological and ultrastructural studies on thetestis of Bithynia tentaculata (Mollusca: Gastropoda), and the effectsof Cercaria helvetica XII (Trematoda: Digenea) on the host organ.Journal of Zoology, London, 171: 541–561.
RIEDEL, F., HEALY, J. M., RÖPSTORF, P. & SITNIKOVA, T. Y. 2001.Ecology, shell morphology, anatomy and sperm ultrastructure ofthe caenogastropod Pyrgula annulata, with a discussion of the rela-tionship between the ‘Pyrgulidae’ and Caspian and BaikalianRissooideans. Limnologica, 31: 289–302.
SELMI, M.G. & GIUSTI, F. 1980. Structure and function in typical andatypical spermatozoa of Prosobranchia (Mollusca). 1. Cochlostomamontanum (Issel) (Mesogastropoda) (1). In: Atti IV Congresso DellaSocietà Malacologica Italiana, Siena, October 1978 (F. Giusti, ed.),115–147. Atti Accademia Fisiocritici, Siena.
SITNIKOVA, T.Y. 1991. Novaya struktura Baikal´skogo endemichnogosemejstva Baicaliidae (New structure of Baikal endemic molluscs of
the family Baicaliidae). In: Morfologiya i evolyutsiya bezpozvonochnykh(Morphology and Evolution of Invertebrates). Fauna of Lake Baikal,1991: 266–281. Nauka, Novosibirsk. [In Russian.]
SITNIKOVA, T.Y. 1995. Guide to Recent molluscs of Northern Eurasia.5. Gastropods of the family Benedictiidae from Lake Baikal.Ruthenica, 5: 77–90.
SITNIKOVA, T.Y., RÖPSTORF, P. & RIEDEL, F. 2001. Reproduction,duration of embryogenesis, egg capsules and protoconchs of gastro-pods of the family Baicaliidae (Caenogastropoda) endemic to LakeBaikal. Malacologia, 43: 59–85.
STAROBOGATOV, Y.I. 1970. Fauna Mollyuskov i zoogeograficheskoye rayonirovaniye kontinental ‘nikh vodoemov. (Malakofauna and zoogeo-graphic classification of the continental water bodies). Nauka,Leningrad. [In Russian.]
STAROBOGATOV, Y.I. & SITNIKOVA, T.Y. 1998. List of Molluscaspecies. In: Lake Baikal, Evolution and Biodiversity (O. M. Kozhova &L. R. Izmest́eva, eds), 404–414. Backhuys Publishers, Leiden.
THIELE, J. 1929. Handbuch der systematischen Weichtierkunde. GustavFischer, Jena.
THOMPSON, F.G. 1984. North American freshwater snail genera ofhydrobiid subfamily Lithoglyphinae. Malacologia, 25: 109–141.
THOMPSON, T.E. 1973. Euthyneuran and other molluscan spermato-zoa. Malacologia, 14: 167–206 (plus addendum, 443–444).
TOCHIMOTO, T. 1967. Comparative histochemical study on thedimorphic spermatozoa of the Prosobranchia with special refer-ence to polysaccharides. Science Report of the Tokyo Kyoiku Daigaku, 13:75–109.
VAUGHT, K. 1989. A Classification of the Living Mollusca (R. T. Abbott &K. J. Boss, eds). American Malacologists Inc., Melbourne, Florida.
WALKER, M.H. & MACGREGOR, H.C. 1968. Spermatogenesis and thestructure of the mature sperm in Nucella lapillus (L.). Journal of CellScience, 3: 95–104.
P. RÖPSTORF ET AL .
126
Related Documents











