7162 European Review for Medical and Pharmacological Sciences 2021; 25: 7162-7184 A.A. RABAAN 1,2,3 , A.A. MUTAIR 4,5,6 , Z.A. ALAWI 7 , S. ALHUMAID 8 , M.A. MOHAINI 9,10 , J. ALDALI 11 , R. TIRUPATHI 12,13 , A.A. SULE 14 , T. KORITALA 15 , R. ADHIKARI 16 , M. BILAL 17 , M. DHAWAN 18,19 , R.K. MOHAPATRA 20 , R. TIWARI 21 , S.A. SAMI 22 , S. MITRA 23 , M.K. PANDEY 24 , H. HARAPAN 25,26,27 , T.B. EMRAN 28 , K. DHAMA 29 1 Molecular Diagnostic Laboratory, Johns Hopkins Aramco Healthcare, Dhahran, Saudi Arabia 2 Department of Public Health and Nutrition, The University of Haripur, Haripur, Pakistan 3 College of Medicine, Alfaisal University, Riyadh, Saudi Arabia 4 Research Center, Almoosa Specialist Hospital, Al-Ahsa, Saudi Arabia 5 College of Nursing, Princess Norah Bint Abdulrahman University, Riyadh, Saudi Arabia 6 School of Nursing, Wollongong University, Wollongong, NSW, Australia 7 Division of Allergy and Immunology, College of Medicine, King Faisal University, Al-Ahsa, Saudi Arabia 8 Administration of Pharmaceutical Care, Al-Ahsa Health Cluster, Ministry of Health, Al-Ahsa, Saudi Arabia 9 Basic Sciences Department, College of Applied Medical Sciences, King Saud bin Abdulaziz University for Health Sciences, Al-Ahsa, Saudi Arabia 10 King Abdullah International Medical Research Center, Al-Ahsa, Saudi Arabia 11 Pathology Organization, Imam Mohammed Ibn Saud Islamic University, Riyadh, Saudi Arabia 12 Department of Medicine Keystone Health, Penn State University School of Medicine, Hershey, PA, USA 13 Department of Medicine Wellspan Chambersburg and Waynesboro Hospitals, Penn State Chambersburg, PA, USA 14 Department of Informatics and Outcomes, St. Joseph Mercy Oakland Pontiac, MI, USA 15 Department of Internal Medicine, Mayo Clinic Health System Mankato, Mayo Clinic College of Medicine and Science, MN, USA 16 Department of Hospital Medicine, Franciscan Health Lafayette, IN, USA 17 School of Life Science and Food Engineering, Huaiyin Institute of Technology, Huaian, China 18 Department of Microbiology, Punjab Agricultural University, Ludhiana, India 19 The Trafford Group of Colleges, Manchester, UK 20 Department of Chemistry, Government College of Engineering, Keonjhar, Odisha, India 21 Department of Veterinary Microbiology and Immunology, College of Veterinary Sciences, Uttar Pradesh Pandit DeenDayal Upadhyaya PashuChikitsa Vigyan Vishwavidyalaya Evam Go AnusandhanSansthan (DUVASU), Mathura, India 22 Department of Pharmacy, Faculty of Biological Sciences, University of Chittagong, Chittagong, Bangladesh 23 Department of Pharmacy, Faculty of Pharmacy, University of Dhaka, Dhaka, Bangladesh 24 Department of Translational Medicine Center, All India Institute of Medical Sciences, Bhopal, Madhya Pradesh, India 25 Medical Research Unit, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia 26 Tropical Diseases Centre, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia 27 Department of Microbiology, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia 28 Department of Pharmacy, BGC Trust University Bangladesh, Chittagong, Bangladesh 29 Division of Pathology, ICAR-Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, India Corresponding Authors: Kuldeep Dhama, MVSc, Ph.D; e-mail: [email protected] Talha Bin Emran, Ph.D; e-mail: [email protected] Comparative pathology, molecular pathogenicity, immunological features, and genetic characterization of three highly pathogenic human coronaviruses (MERS-CoV, SARS-CoV, and SARS-CoV-2)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

7162
European Review for Medical and Pharmacological Sciences 2021; 25: 7162-7184
A.A. RABAAN1,2,3, A.A. MUTAIR4,5,6, Z.A. ALAWI7, S. ALHUMAID8, M.A. MOHAINI9,10, J. ALDALI11, R. TIRUPATHI12,13, A.A. SULE14, T. KORITALA15, R. ADHIKARI16, M. BILAL17, M. DHAWAN18,19, R.K. MOHAPATRA20, R. TIWARI21, S.A. SAMI22, S. MITRA23, M.K. PANDEY24, H. HARAPAN25,26,27, T.B. EMRAN28, K. DHAMA29
1Molecular Diagnostic Laboratory, Johns Hopkins Aramco Healthcare, Dhahran, Saudi Arabia2Department of Public Health and Nutrition, The University of Haripur, Haripur, Pakistan3College of Medicine, Alfaisal University, Riyadh, Saudi Arabia4Research Center, Almoosa Specialist Hospital, Al-Ahsa, Saudi Arabia5College of Nursing, Princess Norah Bint Abdulrahman University, Riyadh, Saudi Arabia6School of Nursing, Wollongong University, Wollongong, NSW, Australia7Division of Allergy and Immunology, College of Medicine, King Faisal University, Al-Ahsa, Saudi Arabia8Administration of Pharmaceutical Care, Al-Ahsa Health Cluster, Ministry of Health, Al-Ahsa, Saudi Arabia9Basic Sciences Department, College of Applied Medical Sciences, King Saud bin Abdulaziz University for Health Sciences, Al-Ahsa, Saudi Arabia10King Abdullah International Medical Research Center, Al-Ahsa, Saudi Arabia11Pathology Organization, Imam Mohammed Ibn Saud Islamic University, Riyadh, Saudi Arabia12Department of Medicine Keystone Health, Penn State University School of Medicine, Hershey, PA, USA13Department of Medicine Wellspan Chambersburg and Waynesboro Hospitals, Penn StateChambersburg, PA, USA14Department of Informatics and Outcomes, St. Joseph Mercy Oakland Pontiac, MI, USA15Department of Internal Medicine, Mayo Clinic Health System Mankato, Mayo Clinic College ofMedicine and Science, MN, USA16Department of Hospital Medicine, Franciscan Health Lafayette, IN, USA17School of Life Science and Food Engineering, Huaiyin Institute of Technology, Huaian, China18Department of Microbiology, Punjab Agricultural University, Ludhiana, India19The Trafford Group of Colleges, Manchester, UK20Department of Chemistry, Government College of Engineering, Keonjhar, Odisha, India21Department of Veterinary Microbiology and Immunology, College of Veterinary Sciences, Uttar Pradesh Pandit DeenDayal Upadhyaya PashuChikitsa Vigyan Vishwavidyalaya Evam Go AnusandhanSansthan (DUVASU), Mathura, India22Department of Pharmacy, Faculty of Biological Sciences, University of Chittagong, Chittagong, Bangladesh23Department of Pharmacy, Faculty of Pharmacy, University of Dhaka, Dhaka, Bangladesh24Department of Translational Medicine Center, All India Institute of Medical Sciences, Bhopal, Madhya Pradesh, India25Medical Research Unit, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia26Tropical Diseases Centre, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia27Department of Microbiology, School of Medicine, Universitas Syiah Kuala, Banda Aceh, Aceh, Indonesia28Department of Pharmacy, BGC Trust University Bangladesh, Chittagong, Bangladesh29Division of Pathology, ICAR-Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, India
Corresponding Authors: Kuldeep Dhama, MVSc, Ph.D; e-mail: [email protected] Talha Bin Emran, Ph.D; e-mail: [email protected]
Comparative pathology, molecular pathogenicity, immunological features, and genetic characterization of three highly pathogenic human coronaviruses (MERS-CoV, SARS-CoV, and SARS-CoV-2)

Comparative review of three human coronaviruses
7163
Abstract. – The last two decades have wit-nessed the emergence of three deadly coronavi-ruses (CoVs) in humans: severe acute respiratory syndrome coronavirus (SARS-CoV), Middle East respiratory syndrome coronavirus (MERS-CoV), and severe acute respiratory syndrome corona-virus 2 (SARS-CoV-2). There are still no reliable and efficient therapeutics to manage the devas-tating consequences of these CoVs. Of these, SARS-CoV-2, the cause of the currently ongoing coronavirus disease 2019 (COVID-19) pandem-ic, has posed great global health concerns. The COVID-19 pandemic has resulted in an unprec-edented crisis with devastating socio-econom-ic and health impacts worldwide. This highlights the fact that CoVs continue to evolve and have the genetic flexibility to become highly patho-genic in humans and other mammals. SARS-CoV-2 carries a high genetic homology to the previously identified CoV (SARS-CoV), and the immunological and pathogenic characteristics of SARS-CoV-2, SARS-CoV, and MERS contain key similarities and differences that can guide therapy and management. This review presents salient and updated information on compara-tive pathology, molecular pathogenicity, immu-nological features, and genetic characterization of SARS-CoV, MERS-CoV, and SARS-CoV-2; this can help in the design of more effective vaccines and therapeutics for countering these pathogen-ic CoVs.
Key Words: Pathology, Immunology, Genetic characterization,
Coronaviruses, MERS-CoV, SARS-CoV, SARS-CoV-2, COVID-19.
Introduction
The devastating fact that zoonotic diseases at-tributed to coronavirus (CoV) strains can result in pandemics came to public attention in 2003 after a severe acute respiratory syndrome coro-navirus (SARS-CoV) outbreak. Since this re-alization, scientists and public health officials have raised concerns over health threats posed to the human population by the three coronavi-ruses (CoVs) SARS-CoV, Middle East respira-tory syndrome coronavirus (MERS-CoV), and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2)1-7. Among at least six strains of human-infecting CoVs that have been identified by studies, these three have proved to be highly pathogenic as they trigger severe pneumonia and systemic symptoms in humans5,8-13. CoVs are a complex and diverse family of enveloped, posi-tive-sense, single-stranded RNA viruses and are
divided into four genera: alpha, beta, gamma, and delta CoV8,14,15. Of these, beta CoVs have drawn the most attention due to their ability to cross an-imal-human barriers and act as significant global infectious agents2,6,8,16. SARS-CoV, SARS-CoV-2, and MERS-CoV have been identified as the most important and evolving beta CoVs, and their mo-lecular biology and immunological features re-main to be investigated in detail1,4,5,8,17-19. Seasonal variations have been observed in the pattern of these viruses: SARS-CoV-2 outbreak occurs in the winter, in contrast to MERS-CoV and SARS-CoV outbreaks and triggering severe pneumo-nia18. Moreover, these three viruses show similar genomic composition, clinical manifestations, and route of transmission1,4,20. The current pan-demic of coronavirus disease 2019 (COVID-19) caused by SARS-CoV-2 has apparent similari-ties with SARS9,10,21, including disease progres-sion, escape from the host immune system, and subsequent acute respiratory distress syndrome (ARDS). The International Committee on Tax-onomy of Viruses (ICTV) designated the causal agent of COVID-19 as “SARS-CoV-2” due to its similarities with SARS-CoV19,22-24.
During the COVID-19 pandemic, the world has experienced unprecedented challenges, with over 4.9 million deaths and more than 243 mil-lion confirmed cases of SARS-CoV-2 infection in over 225 countries with a fatality rate ranging be-tween 1.5% and 5% as of October 26, 202117,22,25. A high case fatality rate of about 49%9,26 has been reported in patients with an acute disease requir-ing ventilator support and Intensive Care Unit (ICU) admission. There have been significant breakthroughs in vaccine development, with sev-eral vaccines administered globally for protection against SARS-CoV-2. In addition, many effective drugs and therapeutic candidates are being eval-uated, such as antivirals, monoclonal antibodies, cytokine inhibitors, and immunosuppressants27-32.
SARS-CoV-2, when observed under an electron microscope, has a structure similar to a crown (co-rona). The mechanism of virus entry into the host is identical to that of SARS-CoV, which binds to the human angiotensin-converting enzyme 2 (ACE2) receptor via its protein receptor-binding domain (RBD)33-35. In contrast, MERS-CoV binds to the DPP4 receptor to enter host cells. Genomic analysis data has revealed that the genome sequence simi-larity of SARS-CoV-2 to SARS-CoV and MERS-CoV is 80% and 50%, respectively21,36. While ex-ploring the evolutionary potential of SARS-CoV-2, studies have found that its genome exhibits 96%

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7164
similarity to that of bat-derived CoV isolated in 201337,38. SARS-CoV-2 and SARS-CoV have 380 amino acid (AA) substitution sites. It has been hy-pothesized that any substitution in the AA sequence could lead to a possible novel viral protein function with unclear pathogenesis39. The spike (S) protein and the nucleocapsid protein are linked to higher transmission capability and lower pathogenicity in SARS-CoV-2. However, the mutations in the S protein are especially crucial because the S protein is key for the first step of viral transmission: entry into the cell by binding to the ACE2 receptor40-44. SARS-CoV-2 steadily mutates during continuous transmission among humans, and naturally occur-ring S mutations can reduce or enhance cell entry via the ACE2 receptor44,45. According to a recent study46, six AA residues (D480, T487, Y442, N479, L472, and Y4911 for SARS-CoV, and Q493, S494, L455, F486, Y505, and N501 for SARS-CoV-2) are essential for binding to the human ACE2 receptor. Among these six SARS-CoV-2 AA residues, the lack of similarity of five residues to those of SARS-CoV may be attributed to the deletions, insertions, or mutations in the S1 and S2 regions, which are responsible for evolutionary changes46,47. The nov-el strain has an evolutionary path different from those of MERS-CoV and SARS-CoV, with lin-eage similarity to previously evaluated bat-derived CoV. However, there are proteomic and genomic differences between the bat and human CoVs, in-dicating a unique immune invasion mechanism and a distinct immunopathology associated with host response48. The common clinical symptoms of COVID-19 are similar to those of SARS: dry cough (67.7%), fever (87.9%), myalgia (34.8%), fa-tigue (69.6%), hypoxia, and progressive dyspnea followed by damage to multiple organs. In contrast to SARS-CoV, SARS-CoV-2 is more transmissi-ble, but the overall mortality rate is lower than that for SARS-CoV infection. Like MERS and SARS, COVID-19 is likely to be more severe in elderly people and those suffering underlying comorbid-ities, including many chronic health conditions. Here, we present salient and updated information on comparative pathology, molecular pathogenic-ity, immunological features, and genetic charac-terization of SARS-CoV, MERS-CoV, and SARS-CoV-2. As the current pandemic remains ongoing, this review can contribute to the design of more effective vaccines and therapeutics.
Early Phase of Viral InfectionIn the early stages, SARS infection causes
non-specific symptoms such as myalgia, fever,
headache, and severe fatigue49. These symptoms tend to diminish in seven days. Sequential naso-pharyngeal aspirate samples from SARS patients indicate a direct relationship between clinical pro-gression and viral load50. After its peak, viral load usually decreases rapidly, with IgG seroconver-sion serving as an indicator of specific immunity development. However, some patients’ clinical conditions can worsen during this period, creat-ing inconsistencies with viral clearance observa-tions. Delay in viral peak can indicate absence or hindrance of host antiviral responses necessary to enhance viral clearance51.
A retrospective study evaluating the cause of worsening clinical condition after viral load re-duction highlighted the underlying association be-tween viral clearance, immune dysregulation, and disease development52. The host hyper-inflamma-tory response, not the cytopathic effect of the vi-rus, may be responsible for this phenomenon36,51. To some extent, rapid viral load elevation could be the contributing factor for disease pathology. Clinical features such as diarrhea, oxygen desat-uration, hepatic dysfunction, and fatality indicate that high viral load may contribute to direct organ dysfunction49,53. Clinical specimens of various anatomic sites of organ dysfunction have yielded virus. For instance, stool specimen was highly re-lated to diarrhea, with viral particles detected in ileum and colon biopsies observed under an elec-tron microscope54. There is extensive evidence regarding the relationship between pathological effects, viremia, and viral loads from these find-ings. Strong evidence exists of high viral loads associated with massive infiltration of the inflam-matory immune cells being significantly linked to worse clinical outcomes in patients54. Patients with elevated viral load at an early stage were also likely to have higher mortality55,56. Therefore, it is essential to address the molecular pathology, immunological characteristics, pathogenicity, and genetic sequence of MERS-CoV, SARS-CoV, SARS-CoV-2, and other CoVs. A few of the gen-eral characteristics of MERS-CoV, SARS-CoV and SARS-CoV-2 are presented in Table I.
Genetic Similarities of MERS-CoV, SARS-CoV, and SARS-CoV-2
Among the CoV subtypes, beta CoVs cause severe and fatal diseases in humans, while al-pha CoVs cause mild infections. The genom-ic sequences of MERS-CoV, SARS-CoV, and SARS-CoV-2 are quite similar, but SARS-CoV-2 displays significant differences in genome com-

Comparative review of three human coronaviruses
7165
position compared to its predecessors57. Genom-ic analysis suggests that SARS-CoV-2 is closely related to pangolin CoV (86%-92%) and bat CoV (96%), which further suggests bats as the prima-ry reservoir43,58-60. Furthermore, the outbreak of SARS-CoV-2 is thought to be linked to trading practices in Wuhan’s wet market, and due to the genetic identities between SARS-CoV-2 and BatCoV RaTG13 (a bat-CoV), it has been hy-pothesized that bats could be the natural source of SARS-CoV-28,43,61. A plethora of research ev-idence shows that pangolins may be the inter-mediate host—there is 99% homology between SARS-CoV-2 and the CoV strain originating from pangolins—but bats are the natural reservoir for the virus62,63. Bats are generally recognized as potential primary reservoirs for most of the RNA viruses64. The genome of SARS-CoV-2 showed 96.2% homology to that of the bat CoV (RaTG13) collected in the Yunnan province of China43. The SARS-CoV-2 genome is closely related (88%) to zoonotic bat viruses, bat-SL-CoVZXC45, and bat-SL-CoVZXC2165. The most commonly identified sequence similarity between these bat and human viruses is in the E gene, and the least common-ly identified similarity is in the S gene. Multiple SARS-CoV-2 proteins have the same sequence as the bat-SL-CoVZC45 and bat-SL-CoVZXC21, except for the S protein and protein 1366. A team of researchers concluded that pangolin-CoV is a
highly associated descendant of SARS-CoV-2, suggesting that pangolins could be the natural reservoirs for SARS-CoV-2 and bat CoV67. The se-quence similarity (89.2%) between SARS-CoV-2 and RaTG13, in terms of the RBD, is less than the sequence similarity (97.2%) between SARS-CoV-2 and pangolin-CoV. Additionally, the latter contains six complete identical RBD residues, whereas the former contains only one identical amino acid residue43. Notably, pangolins in Chi-na are categorized as endangered due to their decreasing numbers, which are close to the point of extinction; this reduces the likelihood of pan-golins acting as an intermediate host of SARS-CoV-2. The selling of pangolins is against the law, and they have not been spotted in Wuhan’s wet markets in recent times68. Through the use of the optimized random forest model for human se-quences of MERS-CoV and SARS-CoV, interme-diate hosts (Camelids and Carnivores) were con-firmed based on evolutionary signatures. With the same method, SARS-CoV-2 evolutionary signa-tures identified bats as hosts, further confirming bats as the suspected origin of the present pan-demic69. Furthermore, a recent study70 based on genetic similarities proposed that snakes may be intermediate hosts, as there are similarities in co-dons among SARS-CoV-2, bat CoV, and a snake virus. However, this analysis was insufficient to reach a conclusive hypothesis, as several limita-
Table I. Characteristics of MERS-CoV, SARS-CoV and SARS-CoV-2.
Features MERS-CoV SARS-CoV SARS-CoV-2
Outbreak 2012, April 2002, November 2019, DecemberLocation of the first case Jeddah, Saudi Arabia Guangdong, China Wuhan, ChinaKey hosts Bat, camel Bat, palm civets,raccoon dogs Bat, pangolinActive cases confirmed 2519 (from 2012 until 8096 Over 243 million January 31, 2020) (as of October 26, 2021)Genome length (bp) 30,119 29,751 29,903Mortality 34.40% 10% (6.8-16.1%) 2-5%Days took to infect the first 1000 persons 903 130 48Incubation period (day) 5 to 6 2 to 7 7 to 14Basic reproduction number (R0) 1 2-4 1.4-5.5Receptor DPP4 ACE2 ACE2Mode of transmission Touching or consumption Believed to have Human-to-human of camel milk or meat. spread on close contact There is limited human-to-human from bats. transmission occurs when transmission despite close There is evidence there is close physical physical contact of human-to-human contact (mainly transmission through respiratory aerosols/ droplets). The transmissions may be possible through fecal-oral route and contaminated objects/ surfaces/fomites

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7166
tions were present in the study71. In any case, beta CoVs are less likely to infect reptiles by crossing over through mammals72. These findings made the natural reservoir of CoV a controversial topic, and a contingent of groups embrace the idea that different intermediate host species are yet to be discovered, other than bats73-75. The disease out-break related to SARS-CoV-2 demonstrates con-cealed virus reservoirs in animals that may spread into human populations occasionally76. The lower effective number of codons and the extreme co-don usage bias of SARS-CoV-2 in S, envelop, and matrix protein genes suggest higher gene expres-sion efficiency than that of SARS, bat SARS, or MERS-CoV, which is similar to Pangolin beta CoV77. In the human host, the SARS-CoV-2 dinu-cleotide pair, UpG and CpA dinucleotides, were highly preferred, and CpG dinucleotide was high-ly avoided. This strategy might imply evasion of the human immune system78. Multiple sequence alignments of the ACE2 receptor proteins of hu-mans with that of dogs, cats, tigers, minks, and other animals revealed a high homology and full conservation of the five AA residues, 353-KGD-FR-357, among the species, which may throw light on the possibility of transmission of SARS-CoV-2 from animals to humans78.
MERS-CoV is closely related to two bat CoV (HKU4, HKU5); it has been suggested that it may be isolated from bats, and dromedary camels probably act as intermediate host, as evidenced from serological studies79,80. In Qatar, the pres-ence of MERS-CoV RNA was reported in swabs obtained from dromedary camels that shared a correlation with two human cases of MERS81. A comprehensive evolutionary relationship analysis depicted the origin of MERS-CoV from bats due to the occurrence of recombination events within S and ORF1ab genes82,83. Recombination events were also reported in SARS-CoV as regions for putative recombination were detected via com-putational genomic studies84. The MERS-CoV strains isolated from humans and camels have been reported to share over 99% identity with variations located in the ORF3, ORF4b, and S genes85. SARS-CoV-2 shows 80% similarity with SARS-CoV and 51% with the MERS-CoV86. Most of the coding areas of SARS-CoV-2 indi-cate a similar genomic architecture to that of the bat-originating CoVs and SARS-CoV. The twelve coding regions predicted are; lab, 3, E, M, 7, 8, 9, 10B, N, S, 13, and 14. The proteins encoded by all the three CoVs are mostly similar in length87. However, there is a significant variation in the S
protein of SARS-CoV-2, which is longer in com-parison to the protein encoded in the bat CoVs, SARS-CoV, and MERS-CoV88.
SARS-CoV-2 shares many similarities in ar-chitecture and pathogenicity with SARS-CoV compared to MERS-CoV. Mathematical mod-els such as decision-tree experiments have also shown remarkable characteristics of an AA se-quence of SARS-CoV-2, which is different from MERS-CoV12. The CoVs use a similar S protein for binding to their respective host cells and the same cellular protease enzyme for the activation of the S protein89. The S protein in SARS-CoV-2 has a sequence similarity of about 77% with that of SARS-CoV, structural proteins are more than 90% similar to SARS-CoV, and 32.79% similar to MERS-CoV counterparts. The receptor-binding domain (S2) of SARS-CoV-2 has a sequence sim-ilarity of 74% with the S2 domain in SARS-CoV and an overall similarity of about 52% with that of SARS-CoV90. The E protein of SARS-CoV-2 is 96.00% similar to that of SARS-CoV and 36.00% similar to that of MERS-CoV. The M protein of SARS-CoV-2 is 89.59% similar to that of SARS-CoV and 39.27% similar to that of MERS-CoV. The SARS-CoV-2 N protein is 85.41% similar to that of SARS-CoV and 48.47% similar to that of MERS-CoV 91.
Accessory proteins are regarded as essential for in vitro replication of viral particles; however, some of these proteins are associated with viral pathogenesis92,93. The 3CLpro (nsp5) and RdRp (nsp12) proteins of SARS-CoV-2 are prime me-diators of replication and new virion production, and they share high sequence identity with SARS-CoV and MERS-CoV94. Recent reports95,96 have demonstrated that ORF8b and ORF3a of SARS-CoV catalyze the induction of proinflammatory cytokines and thus play a role in regulating che-motaxis in macrophages. ORF8b of SARS-CoV and MERS-CoV is also involved in suppressing the induction of interferon (IFN-I)97,98. Another study demonstrated that ORF8 of SARS-CoV-2 variant binds to major histocompatibility complex (MHC) and regulates its degradation in cell cul-ture, indicating that immune evasion may be me-diated by ORF8. However, SARS-CoV-2 ORF8 shows low homology to SARS-CoV ORF899. Generally, no homologous accessory proteins are found in CoV genera. However, some simi-lar kinds of proteins might be present in closely associated CoVs. For instance, SARS-CoV-2 and SARS-CoV show over 80% similarities in OR-F3a, 6, 7a, 7b, and 9b protein sequences.

Comparative review of three human coronaviruses
7167
Comparative Molecular Pathology of MERS-CoV, SARS-CoV, and SARS-CoV-2 Infection
SARS-CoV is considered a zoonotic virus that was transmitted to humans from birds pri-or to human-to-human transmission100. However, in humans, various risk factors including age, underlying metabolic disease like diabetes, and heart disease, lead to an increase in death risk101. SARS starts with viral infection in the respiratory tract of people of all ages via droplet transmis-sion of virus present in the mucus or saliva102. It was reported that viral loads of SARS-CoV de-creased with increased severity of the disease. On the contrary, a similar trend is still unclear for MERS-CoV103. Clinical symptoms associated with SARS-CoV infection include fever, chills, diarrhea, myalgia, and fatigue104. SARS-CoV en-ters into the human cell through the attachment of viral S glycoprotein (S protein) to the ACE2 receptor. ACE2 functions as a dominant host re-ceptor, and the presence of two co-receptors, DC-SIGN (CD209) and L-SIGN (CD209L), are also reported105,106. In dendritic cells, viral infection does not occur prior to DC-SIGN binding, but this binding may enhance SARS-CoV infection and dissemination substantially. On the other hand, L-SIGN is considered an alternative receptor that may bind with its spike protein and regulate cel-lular entry of SARS-CoV107. Changes occur in the S glycoprotein in the endosomal environment via the serine protease cathepsins B and L to assist in the union process108. The S glycoprotein is not just an essential structural protein of CoVs; it per-forms a vital role in the association of virus with the host cell. The S- protein is made up of two subunits: S1 and S2109. The S1 subunit contains the RBD, which is responsible for binding the virus to the host receptor, while the S2 subunit controls membrane fusion occurring during virus-host membrane interactions. These interactions lead to the penetration of the viral genome into the cy-toplasm of the host cell110. SARS-CoV-2 encodes a longer S protein compared to SARS-CoV and MERS-CoV, as identified by phylogenetic analy-sis20,76. The RBD of SARS-CoV, MERS-CoV, and SARS-CoV-2 binds to functional receptors pres-ent on the cellular surface, allowing penetration of the virus into host cells111. SARS-CoV and SARS-CoV-2 predominately utilize angiotensin-convert-ing enzyme 2 (ACE2) as a host receptor105,110,111. Additionally, viral entry by antibody-depen-dent enhancement (ADE) has been observed112. Through ADE, the B cell producing antibodies
may also expedite viral infection113. Surprising-ly, ACE2 exhibits stronger affinities for SARS-CoV-2 compared to SARS-CoV114. For instance, the interaction between host ACE2 and SARS-CoV-2 spike ectodomain displayed 10- to 20-fold higher binding affinity than that for SARS-CoV in a recent study115. Another study speculated that SARS-CoV-2 could use other cellular receptors and proteins to bind with host cell receptors such as integrins116. However, there is to date insuffi-cient evidence to corroborate this assumption. CD147-SP can be considered another entry portal of SARS-CoV-2117. In addition to attachment of S proteins to functional host receptors, priming of S proteins is necessary for invading the cellular machinery of the host118.
Apart from lung cells, the heart, kidney, liver, and tongue also express ACE2 receptors on their epithelial cells119,120. In fact, cilia could be the entry gate of the virus121. Surprisingly, after the S gly-coprotein attaches to ACE2, there is a significant cilia loss, squamous cell metaplasia, and elevated macrophage migration into the alveoli, causing notable damage to alveoli in the lungs. Addition-ally, SARS-CoV generates 7a and 3a proteins that lead to substantial programmed death of cells in the lungs, liver, and kidney122. Host translation elongation factor 1 (EF-1A) and serine protease 2 strongly bind to N protein of both SARS-CoV and MERS-CoV, and subsequently induce local or systemic inflammatory responses94. TH1 acti-vation also causes increased inflammation in the affected organs. MERS-CoV infection is more common in males than females123, and SARS-CoV and SARS-CoV-2 infection follow the same order of gender prevalence8. Clinical presentation of infection may range from being asymptom-atic to massive organ damage. Notably, MERS is closely associated with metabolic syndromes such as diabetes mellitus, obesity, and cardiovas-cular morbidities124. The developing metabolic syndrome in most cases alters the immunological function, exposing the infected person to further risk of more infections.
Many previous investigations reported that CoV infection leads to cytopathic effects, includ-ing cell lysis and apoptosis. Cellular fusion is caused by the virus and usually leads to syncytia formation. These processes are observed in the cell due to the mobilization of vesicles that form the replication complex and cause disruption of Golgi complexes at the time of viral replication94. Unlike in SARS-CoV, DPP4 CD26 is the MERS-CoV attachment site to lung and respiratory tract

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7168
epithelial cells125. Notably, MERS-CoV carries a particular RBD in its S glycogen that binds DDP4 on the host cells. DPP4 plays a significant role in altering glucose metabolism, activating the T cells, modulating cytotoxicity, and regulating apoptosis126. SARS-CoV-2 infects both the lower and upper respiratory systems and multiple other organs and systems, thus causing multiple patho-logical conditions, including neurological and gastrointestinal manifestations and kidney dam-age127-129. ACE2 receptors are abundant in oral mucosa, nasal secretory and ciliated cells, lower airways, lungs, cornea, ileum, and colon. Hence, patients suffer from collapsed lung and symptoms of diarrhea130,131. When spike D614 is replaced by mutant G614, S protein possesses greater stability and a potential to grow at a temperature of 37oC, compared to early SARS-CoV-2 isolates, which showed a preference for 33oC132. While SARS-CoV-2 is less pathogenic than MERS-CoV or SARS-CoV, its human-to-human transmission is faster1,4,133. Underlying illnesses (comorbidities) such as heart disease, diabetes, and hypertension have a close association with the severe patho-genesis of SARS-CoV-2 in affected patients134. These disorders reduce the generation of IFN and interleukin that leads to the downregulation of the host’s innate immunity via blockage of lym-phocyte and macrophage functions. In healthy people, ACE2 alters the renin-angiotensin system through angiotensin-II breakdown into angioten-sin-17 to prevent the development of acute lung failure135. Acute lung injury is directly related to a deficiency in ACE2 and an increase in Ang II136,137. Postmortem analysis of SARS-CoV-2 pa-tients has revealed pneumocyte hyperplasia and partial fibrosis leading to thickening and collapse of alveoli93,138
The sgRNAs are presumed to be translated into accessory and structural proteins of CoV in the cytoplasm. A recently concluded in vitro study indicated that the enzymatic function of the nsp14 exoribonuclease (ExoN) is crucial for replication of SARS-CoV-2 and MERS-CoV139. By enhanc-ing degradation and interfering with host mRNA translation, beta CoV nsp1 inhibits the expression of host genes and thus serves as a potent virulence factor140. MERS-CoV nsp1 inhibits mRNA trans-lation and induces mRNA degradation by selec-tively targeting nuclear mRNA translation and avoiding cytoplasmic viral mRNAs141. Current structural analysis and related studies have un-veiled that SARS-CoV-2 nsp1 inhibits ribosomal mRNA entrance142. The delta CoVs and gamma
CoVs cannot produce nsp1 due to lack of nsp1/nsp2 cleavage sites, though the same host shutoff is triggered by other mechanisms that have not been explored well.
Clinical and Immunological Features of MERS-CoV, SARS-CoV, and SARS-CoV-2 Infection
MERS is currently a common human corona-virus (hCoV) infection. MERS-CoV infection has lower transmissibility than the other two CoVs but causes severe symptoms, leading to a high-er case fatality rate40. Like SARS-CoV patients, patients with MERS-CoV usually show milder symptoms initially and later develop dyspnea and complications leading to respiratory failure, with most of the patients (63.4%) developing lethal pneumonia25. Organ function later deteriorates, leading to fatality within two weeks after infec-tion48. Prominent comorbidities associated with mortality among MERS-CoV patients are diabe-tes and renal failure, which result in poor health outcomes. To better understand the pathogenesis and immunological features of MERS-CoV, it is essential to undertand its comparative analysis with SARS-CoV infection1,4.
Unlike the SARS-CoV abortive mechanism of infection, MERS-CoV multiplies in lympho-cytes, dendritic cells, and macrophages52,54,55,143. Viral genomes, nucleoprotein expression, and vi-ral particles are detectable in virus-infected cells. Viral multiplication in macrophages and dendritic cells indicates that host cells are the source of vi-ral reservoirs thwarting host immunorecognition of the virus55. MERS-CoV has been reported to induce greater transcriptomic changes than those induced by SARS-CoV88. Cells carrying the virus facilitate systemic dissemination of the infection to lymph nodes. Naïve T cells interact with the vi-rus and trigger adaptive immune responses. This leads to the release of massive amounts of cyto-kines and chemokines. The diverse activation av-enues that trigger production of cytokines during MERS-CoV infection cause a distinct cytokine profile compared to SARS-CoV infection88,144.
The reason for productive replication is the high number of DDP4 receptors expressed in the dendritic and monocyte cells compared to the expression level of ACE2 receptors. This results in differential infection outcomes. MERS-CoV can infect cells from different human cell lines in ex-vivo studies36,145. DDP4 receptors are identi-fiable in endothelial and epithelial cells present in the prostate, liver, kidney, and intestines41,42. Dis-

Comparative review of three human coronaviruses
7169
semination of the virus throughout the body was observed in patients with MERS-CoV infections, explaining the high incidence of systemic events like multi-organ failure and septic shock. Another important immunopathological feature is the an-tibody-dependent enhancement (ADE) confirmed in MERS-CoV146. The underlying mechanism is linked to the enhanced membrane fusion pro-cess. The interactions of antibodies and RBD of S protein tend to elevate proteolytic susceptibility, leading to conformational changes in the target host cells55. The binding Ab enhances virus entry via canonical receptor-dependent pathways.
Only three cytokines (IL-6, IP-10, and IFN-γ) display a marked increase in SARS-CoV, MERS-CoV, and SARS-CoV-235. SARS-CoV shows sig-nificant IFNantagonism, while the MERS-CoV has minor antagonist characteristics that lead to enhanced sensitivity to IFN-I antiviral respons-es147. Furthermore, due to differences in the vi-ral proteins among these human CoVs, SARS-CoV-2 is more sensitive to IFN-I-dependent antiviral response compared to other CoVs. In fact, the levels of IFN-I and IFN-III in patients with SARS-CoV-2 infections are reduced, un-like that in patients infected by SARS-CoV and other respiratory viruses144. MERS-CoV shares similar viral evasion strategies and IFN antago-nism with SARS-CoV. As a result, MERS-CoV tends to decrease upregulation of antiviral inter-feron-stimulated gene (ISG) responses through a novel approach, resulting in the induction of re-pressive histone modifications in the host cells. Similar histone modifications, which mediate several biological events such as gene regulation, were identified in patients with H5N1 flu infec-tion52. Inhibition of transcription factor binding is controlled by modifying the basal state of host chromatin, where genes are packed. This mechanism is linked to low ISG expression in IFN-administered MERS40. As in patients with SARS-CoV infection, levels of IFN-I in MERS-CoV infected patients are attenuated, and their rate of increase is slowed. The absence of IFN-I resulted in a lack of marked lung immunopa-thology in studies. It also improved the clinical outcomes compared to the delayed IFN-1 group, suggesting an atypical IFN-I effect linked to SARS infection36,37, but there were adverse out-comes linked to MERS-CoVinfection. In this respect, early IFN administration improved the protection of mice from severe infection, irre-spective of down-regulation of cytokine-related genes and ISG.
In seriously ill patients with MERS-CoV, the inability to activate Th1 cells reduces IFN-γ production, leading to activated natural killer (NK) cells and CD8+ T cells. This generates an uncontrolled immune response and attenuates viral clearance36,43,148,149. Extensive CD8+ T-cell responses were noted in critically ill patients during the acute phase, indicating no benefit as-sociated with hyper-activated T-cell responses150. The acute-phase T-cell response for SARS-CoV corresponds to that for MERS-CoV. The absence of T-cell response activation during the innate re-sponse provided sufficient enhancement of host survival and improved disease outcome. Addi-tionally, the adaptive immune response against MERS-CoV infection showed positive effects relative to those observed against SARS-CoV in-fection151.
Concentration of certain inflammatory cyto-kines and chemokines (CXCR3, SOCS5, IL-1β, IL-8, IL-15, IL-17, CCR2, IL-1α, IP-10, TNF-α, and IFN-γ) was reported to increase during MERS-CoV infection144,152. In terms of cytokines, MERS-CoV-associated IL-17 expression demon-strated significant upregulation compared to that associated with SARS-CoV. Secretion of IL-17 by CD4+ T cells can produce extensive pro-inflam-matory effects on host cells54,153. IL-17 expression is known to aggravate respiratory syncytial virus (RSV). In MERS-CoV, IL-17 expression tends to induce immune-mediated pathology, resulting in an elevated mortality rate143. Patients with MERS-CoV infection exhibit higher and more prolonged production of cytokines compared to those with SARS-CoV infection20.
The SARS-CoV-2 pandemic has resulted in devastating outcomes in the 21st century because of high transmissibility and viral-shedding prop-erties6,22. COVID-19 patients show clinical symp-toms that resemble flu during the onset of the dis-ease, including myalgia, fever, and dry cough128,154. Symptoms such as rhinorrhoea, pharyngalgia, al-veolar edema, amblygeustia, shortness of breath, dry mouth, nausea, and vomiting have also been recorded in a small number of COVID-19 pa-tients4,155-157. The laboratory findings obtained from patients infected by SARS-CoV, MERS-CoV, and SARS-CoV-2 are markedly similar; the most common abnormal findings include throm-bocytopenia and lymphocytopenia. Additionally, significant elevation in serum levels of alanine aminotransferase, lactate dehydrogenase, aspar-tate aminotransferase, and C-reactive protein have been recorded158-160. In severely affected pa-

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7170
tients, coagulation disorders, where D-dimer lev-el is elevated and prothrombin time is prolonged, are commonly observed161. Meanwhile, elevation in the creatine kinase and serum creatinine lev-el were reported in some patients, largely those infected with MERS-CoV158-160. Furthermore, a large number of COVID-19 cases with gastroin-testinal symptoms such as extreme diarrhea have been recorded in numerous laboratories, imply-ing that the virus is replicating in the digestive system and viral particles are shed via stool162,163. In addition, the sheer volume of ACE2 recep-tors in the bile duct relative to the alveolar cells contributes to the hypothesis that infection with SARS-CoV or SARS-CoV-2 causes serious liv-er damage164-166. SARS-CoV-2 infection has also been linked to neurological symptoms in several recent studies. Moreover, in some cases, SARS-CoV-2-RNA has been reported in cerebrospinal fluid167,168. The involvement of ACE2 receptors in the central nervous system (CNS) has been con-nected to neurological symptoms such as stroke, polyneuropathy, acute encephalitis, anosmia, age-usia169,170, and brain inflammation associated with COVID-19168,171. In previous studies, patients with COVID-19 were evaluated for symptoms such as anosmia and dysgeusia, and a high percentage (approximately 75%) revealed alteration in the senses of smell and taste172,173.
Cerebral arteriopathies and ischemic vascular stroke have been attributed as the direct effects of SARS-CoV-2 on ACE2 receptors of endothelial cells and as indirect effects of misdirected host immune response174. Neuro-opthalmic manifes-tations of COVID-19 syndrome are also increas-ingly recognized175,176. SARS-CoV-2 has recently been shown to infect different cells of the renal system, including tubular epithelial cells and podocytes, through direct tropism and indirect action by induction of cytokine storm and other mechanisms, resulting in a variety of renal ab-normalities, including acute kidney damage, and higher mortality177-180. Several studies have linked COVID-19 to impaired kidney function during the course of COVID-19 progression. In seriously ill patients with COVID-19, numerous renal dis-orders such as proteinuria, hematuria, and acute kidney failure (AKF) have been identified181-184. According to current observational evidence, AKF is one of the most important causes of ill-ness and death in SARS-CoV-2 patients, second only to ARDS185.
Secondary bacterial and fungal infections ob-served in patients infected with COVID-19 have
been implicated to further complicate the severity of the disease, constituting an important factor, especially for high-risk patients186,187. Even the microbiota, such as the bacterial microbiome, virome, and fungal microbiome, are found to af-fect the natural course of SARS-CoV-2 infection, along with comorbidities such as diabetes and hy-pertension188-190. Additional manifestations under-stated during the COVID-19 epidemic, including psychological illnesses such as depression, anxi-ety, and sleep disorders191 and skin disorders such as urticaria, rashes, erythema, and acro-ischemic lesions, also should be considered192. All the above COVID-19 manifestations are either the di-rect results of SARS-CoV-2 multiplication or of the indirect hyperinflammatory condition known as macrophage activation syndrome or cytokine storm. This syndrome leads to increased produc-tion of IL6, IL7, and TNF-alpha and inflammato-ry chemokines such as CCL2, CC13, and CXC10, as well as elevated amounts of serum ferritin, D-dimer, and chronic reactive protein; however, evidence for inflammasome activation is not pres-ent as IL1β production is not elevated193-195.
Chronic cough is common in long COVID (post-COVID syndrome) after SARS-CoV-2 in-fection and may be associated with the vagal sensory neurons and/or neuroinflammatory re-sponse. Mechanisms of post-COVID-19 chronic cough and optimal management are still unclear. New anti-inflammatory drugs or neuromodula-tors (gabapentin or opioids) could be considered for treatment; however, randomized studies are highly recommended to analyze the safety and efficacy of these potential treatments196. Gunst et al197 have performed a randomized trial to un-derstand the safety and efficacy of TMPRSS2 inhibitors in COVID-19 patients and found that TMPRSS2 inhibitors or camostat mesylate may be effective in the early phase of the disease and lowers the risk of disease progression; howev-er, this treatment is not effective for severely ill and hospitalized patients. According to another recent study, saliva and nasal swabs offered the best diagnostic performance and may be used as an alternative specimen collection method for di-agnosing SARS-CoV-2 infection198,199.
An overview of comparative clinical mani-festations in patients with MERS, SARS, and COVID-19 is presented in Table II.
Pathological studies on SARS-CoV-2 demon-strate increased infiltration into an infected per-son’s lung tissues154,206. Viral particles/macro-phages/inflammatory cells have been identified in

Comparative review of three human coronaviruses
7171
the bronchoalveolar fluid (BALF) of COVID-19 patients103,207,208. SARS-CoV-2 also targets both types of pneumocytes (I and II), similar to SARS-CoV209. Monocyte-derived macrophages are pres-ent at high levels in the BALF, constituting 80% of the infiltrated cells observed in severely sick patients210. Various forms of activation of mono-cyte-originating macrophages have been noted211. ACE2 receptor expression on the surface revealed that entry-binding receptors for SARS-CoV-2 could be detected in alveolar macrophages, indi-cating the possibility of this path in the entry of SARS-CoV-2. From these findings, there is suf-ficient evidence that monocytes are essential in the cytokine storm and lung pathology103,212. The pro-inflammatory classically activated phenotype (M1) identified in wound-healing can activate the phenotype (M2) that leads to inflammatory tissue injuries and the development of fibrotic lesions in ARDS patients213,214. These results indicate fea-tures of SARS research findings that are common to those observed for SARS-CoV-2.
Studies211,214 evaluating chemokines and cy-tokines in SARS-CoV-2 infection would aid in clarifying the full cytokine profiles of patients with severe infection during the acute phase. This would help elucidate the pathogenic mechanisms that result in worse health outcomes. SARS-CoV-2 patients demonstrate increased concen-tration of pro-inflammatory cytokines, including IL1β, IL-2, IL-6, IL-8, IL-17, TNF-α, IP-10, MCP-1, GM-CSF, and G-CSF215, which may be attribut-ed to Th-1-cell responses216. Significant cytokine elevation was identified in patients with severe symptoms, and Th-1-cell and Th-2-cell-related
cytokines were detected simultaneously217. Pre-vious investigations confirmed that the increased level of certain pro-inflammatory cytokines (e.g., IFN-γ, IL1β, IL-6, IL-12) in serum positively correlated with severe lung damage and pulmo-nary inflammation in SARS-CoV patients218. T cell-related CD molecules and lymphocyte levels showed a negative correlation with changes in cy-tokines in patients with SARS-CoV-2 infection. Therefore, there is a potential correlation between cytokine storms and adaptive immunity. In pa-tients with mild symptoms, lymphocyte levels are usually normal during the convalescent phase and are undetectable later as the infection pro-gresses219,220. In acute stages in mild cases, lym-phocyte elevation was not linked to elevation in cytokines. This could be due to cellular immune response initiation that tends to accelerate viral clearance during the early phases, inhibiting cy-tokine production through innate immune activa-tion, thus alleviating disease severity221. In severe cases, cytokine hyperactivation during the acute phase of SARS-CoV-2 resulted in dysregulated systemic disease inflammation and deterioration, as shown by CRP, ferritin, D-dimers, and procal-citonin elevation217,220. Cytokine storms generate pathogenic effects instead of protective effects against SARS-CoV-2 infection of host cells222-225. Excessive cytokine and chemokine activation by macrophages has similar outcomes, as reported in macrophage activation syndrome (MAS) and hemophagocytic lymphohistiocytosis. In particu-lar, many proinflammatory cytokines, IL-1, IL-6, and TNF-α, are involved in COVID-19 pathogen-esis226. Single-cell RNA sequencing has revealed
Table II. Comparative analysis of clinical manifestations in patients with MERS, SARS, and COVID-19.
MERS-CoV SARS-CoV SARS-CoV-2Clinical symptoms/manifestations 8,157,200 8,157,160 8,201–205
Fever 63.5-83.5% 93-97.6% 67.7-90.8%Cough 51-70% 49-59% 45.5-62.9%Fatigue 21-35% 31.2% 23.3-38.1%Dyspnoea 51% 32% 21-40%Sputum 22-43% 16-27% 21.2-37.2%Sore throat ≤ 25% 11-25% 14.6-31.0%Headache 11-20% 30-46% 7.9-15.2%Gastrointestinal symptoms(like diarrhoea) ≤ 30% ≤ 32% 3-17%Bilateral pneumonia N.A. N.A. 58.2-81.0%Acute respiratory distress syndrome (ARDS) 20–30% 20% 18–30%Neurological manifestations such as stroke 17.4 % N.A. 17- 30%Acute kidney injury 41–50% 7% 3-20%
N.A.* (Not Available).
A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7172
potent interactions between immune and epithe-lial cells, with inflammatory macrophages that express multiple cytokines in samples with criti-cal COVID-19 conditions130. Amongst these, IL-8 and IL-6 are detected at elevated concentrations in individuals with critical or severe COVID-19. These high levels of both cytokines indicate lym-phocytopenia that predicts disease progression227.
During the initial stages of SARS-CoV-2 infec-tion, the lymphocyte count decreases in patients with severe symptoms associated with a dramat-ic decrease in NK cells, B cells, CD4+, CD8+ T, and CD3+ cells228. Mild cases of SARS-CoV-2 infection demonstrate a moderate increase in these lymphocytes220,221. These findings further reveal that lymphocytes can reach comparable
levels along with only slight variation in notice-able lymphocyte count214,221. Changes in adaptive immunity occur due to imbalanced activation of Th1/Th2, and alteration of T lymphocyte func-tion causes adverse effects on host cells, wors-ening the course of disease. Reduced function of CD8+ T cells serves as a predictor of the severity of SARS-CoV-2 infection208. Moreover, the pro-duction of endogenous IFNβ in the nasal mucosa of critical patients can be considered a prognostic tool of IFN therapy for managing COVID-19, as it predicts clinical outcomes229. The eosinophil level in COVID-19 patients was related to the reduced anticoagulant effect and therefore may be consid-ered for determining the prophylactic anticoagu-lant administration strategy230. The high eosino-
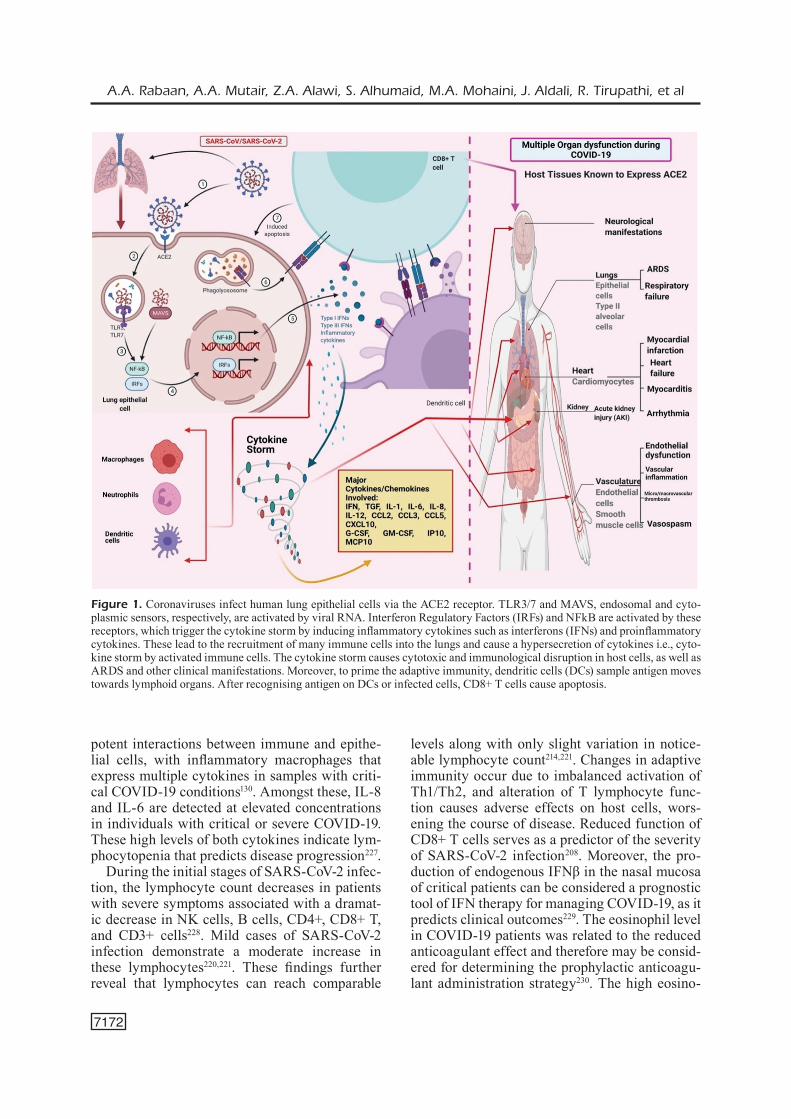
Figure 1. Coronaviruses infect human lung epithelial cells via the ACE2 receptor. TLR3/7 and MAVS, endosomal and cyto-plasmic sensors, respectively, are activated by viral RNA. Interferon Regulatory Factors (IRFs) and NFkB are activated by these receptors, which trigger the cytokine storm by inducing inflammatory cytokines such as interferons (IFNs) and proinflammatory cytokines. These lead to the recruitment of many immune cells into the lungs and cause a hypersecretion of cytokines i.e., cyto-kine storm by activated immune cells. The cytokine storm causes cytotoxic and immunological disruption in host cells, as well as ARDS and other clinical manifestations. Moreover, to prime the adaptive immunity, dendritic cells (DCs) sample antigen moves towards lymphoid organs. After recognising antigen on DCs or infected cells, CD8+ T cells cause apoptosis.

Comparative review of three human coronaviruses
7173
phil count is also associated with lower activity of anti-factor Xa230. Liu and coworkers have also suggested the importance of familial cluster (FC) and non-familial (NF) patients during the treat-ment of COVID-19 patients231.
An overview on pathology and pathobiology of SARS-CoV-2 is presented in Figure 1.
Immunocompetent infected individuals usual-ly present mild manifestations or become asymp-tomatic232. However, the immunocompromised, the elderly, and individuals with underlying con-ditions such as cardiovascular disease, diabetes, and cancer, develop severe symptoms and clinical disease134. A recent study linked renal, cardiovas-cular, and respiratory diseases with greater ICU admission and fatality rate in COVID-19 patients. Cancer and diabetes conditions also showed a strong correlation with severe disease outcomes in SARS-CoV-2 infected patients. Male patients and older people showed higher ICU admission and mortality. The risk of COVID-19 was dra-matically increased in older patients233. In another report, hypertension, cardiovascular disease, and diabetes were closely associated with severe out-comes in COVID-19-infected individuals across all age groups. In contrast to elderly patients, young patients exhibited a varied prevalence of cardiovascular comorbidities234. The De Ri-tis ratio has been associated with poor survival in SARS-COV-2 infected patients and is an im-portant prognostic factor. Moreover, the De Ritis ratio on admission was significantly associated with in-hospital mortality in COVID-19 patients. However, since the sample size in the clinical study was considerably smaller, additional inves-tigations are needed to validate the ability of this parameter to independently predict death in hos-pitalized patients235. Obesity is strongly associat-ed with immune cells, including MAIT and NK cells236-238. Hence, it may be considered a factor for increased risk of severe COVID-19. Popkin et al239 have recently conducted a meta-analysis and found 48% higher mortality in COVID-19 patients with obesity. In a recent cohort study, Gao et al240 found a linear increase in the risk of severity in patients with COVID-19 leading to hospitalization and death with an increase in body mass index. This excess risk was observed particularly in younger people and also in black individuals. According to another cross-sectional study, Lega et al241 found that COVID-19 patients with a severe psychiatric disorder (schizophre-nia and others) died at a younger age compared to those without any psychiatric disorder. Addi-
tionally, the vulnerability of COVID-19 patients with psychiatric disorders may reduce the chance of recovery. Pregnant women were affected more by COVID-19, particularly in the second wave of infections, which may be associated with the emergence of increasing numbers of pathogenic strains242. Moreover, people with physical disabil-ities are particularly at risk and need additional support from mental health services243. In a simi-lar report, authors have mentioned that older peo-ple with disabilities have been neglected during the COVID-19 pandemic244.
Conclusions
Three highly pathogenic coronaviruses (SARS-CoV, MERS-CoV, and SARS-CoV-2) have been reported in humans in the last two decades. Al-though these CoVs exhibit several similar features in their infection process, distinctive features and characteristics are observed in the immunopa-thology and clinical outcomes of each. Recurrent outbreaks of infectious and pathogenic strains of CoVs have posed a significant burden and danger to humankind, such as the current COVID-19 pandemic that has resulted in an unprecedented crisis with devastating social, health, and eco-nomic impacts worldwide. All three CoVs share immunological aspects that affect pathological characteristics. The viral agents undergo rep-lication in the host immune cells and set off an innate immune response that leads to induction of pro-inflammatory cells and cytokines. This cy-tokine storm can be life-threatening. Finally, the body responds by producing protective antibodies that clear the viral infection and also confer im-munity against future infection with the same vi-rus. In vitro studies have been particularly helpful for understanding the immunological and patho-logical aspects of the viruses and in conducting drug trials against these agents. However, there is a need to advance clinical research since these vi-ral agents can undergo further mutations and may give rise to viral species with enhanced patho-genicity in the future. Collaborative and intense efforts by scientists worldwide have resulted in advanced discoveries related to many aspects of SARS-CoV-2 and COVID-19. In addition, eluci-dating the immunopathology and clinical features of CoVs will help in developing better and more effective drugs, medicines, and vaccines to count-er the emergence and re-emergence of pathogenic CoVs. Prospective outcomes from clinical inves-

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7174
tigations of different vaccines and antiviral can-didates provide hope to end this pandemic soon. Continuous research efforts to better understand the pathogenesis, molecular biology, and immu-nological characteristics of SARS-CoV-2 and oth-er CoVs will help to stem the tide of the ongoing COVID-19 pandemic and formulate prevention plans for future pandemics.
Conflicts of interestAll authors declare that there exist no commercial or financial relationships that could, in any way, lead to a potential conflict of interest.
Author ContributionsAll the authors substantially contributed to the con-ception, compilation of data, checking and approving the final version of the manuscript, and agree to be ac-countable for its contents.
AcknowledgementsAll the authors acknowledge and thank their respec-tive Institutes and Universities.
FundingThis is a review article written by its authors and re-quired no substantial funding to be stated.
References
1) Shaw PD, Patel N, Patil S, Samuel R, Khanna P, Prajapati B, Sharun K, Tiwari R, Dhama K, Na-tesan S. Comparative evaluation of the origin, evolution, transmission, diagnosis, and vaccine development of three highly pathogenic human coronaviruses (Sars-cov, mers-cov and sars-cov-2). J Exp Biol Agric Sci 2020; 8: S103-S113.
2) Coleman CM, Frieman MB. Coronaviruses: Im-portant Emerging Human Pathogens. J Virol 2014; 88: 5209-5212.
3) WHO. WHO Director-General’s remarks at the media briefing on 2019-nCov. Who [Internet] 2020; 1-May-2020.
4) Rabaan AA, Al-Ahmed SH, Haque S, Sah R, Ti-wari R, Malik YS, Dhama K, Yatoo MI, Bonilla-Al-dana DK, Rodriguez-Morales AJ. SARS-CoV-2, SARS-CoV, and MERS-CoV: A comparative over-view. Infez Med 2020; 28: 174-184.
5) Rabaan AA, Al-Ahmed SH, Sah R, Alqumber MA, Haque S, Patel SK, Pathak M, Tiwari R, Yatoo MI,
Haq AU, Bilal M, Dhama K, Rodriguez-Morales AJ. MERS-CoV: epidemiology, molecular dynam-ics, therapeutics, and future challenges. Ann Clin Microbiol Antimicrob 2021; 20: 1-14.
6) Dhama K, Patel SK, Sharun K, Pathak M, Tiwari R, Yatoo MI, Malik YS, Sah R, Rabaan AA,Pan-war PK, Singh KP, Michalak I, Chaicumpa W, Martinez-Pulgarin DF, Bonilla-Aldana DK, Rodri-guez-Morales AJ.SARS-CoV-2 jumping the spe-cies barrier: Zoonotic lessons from SARS, MERS and recent advances to combat this pandemic virus. Travel Med Infect Dis 2020; 37: 101830.
7) Berger A, Drosten C, Doerr HW, Stürmer M, Preis-er W. Severe acute respiratory syndrome (SARS) - Paradigm of an emerging viral infection. J Clin Virol 2004; 29: 13-22.
8) Zhu Z, Lian X, Su X, Wu W, Marraro GA, Zeng Y. From SARS and MERS to COVID-19: A brief sum-mary and comparison of severe acute respiratory infections caused by three highly pathogenic hu-man coronaviruses. Respir Res 2020; 21: 1-14.
9) Peiris JSM, Chu CM, Cheng VCC, Chan KS, Hung IFN, Poon LLM, Law KI, Tang BSF, Hon TYW, Chan CS, Chan KH, Ng JSC, Zheng BJ, Ng WL, Lai RWM, GuanY, YuenKY.Clinical progression and viral load in a community outbreak of coro-navirus-associated SARS pneumonia: A prospec-tive study. Lancet 2003; 361: 1767-1772.
10) Prompetchara E, Ketloy C, Palaga T. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidem-ic. Asian Pacific J Allergy Immunol 2020; 38: 1-9.
11) Leung CHC, Gomersall CD. Middle East respiratory syndrome. Intensive Care Med 2014; 40:1015-1017.
12) Huh JE, Han S, Yoon T. Data mining of corona-virus: SARS-CoV-2, SARS-CoV and MERS-CoV. BMC Res Notes 2021; 14: 1-6.
13) Chong ZX, Liew WPP, Ong HK, Yong CY, Shit CS, Ho WY, Ng SYL, Yeap SK. Current diagnostic approaches to detect two important betacoronavi-ruses: Middle East respiratory syndrome corona-virus (MERS-CoV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Pathol - Res Pract 2021; 225: 153-565.
14) Weiss SR. Forty years with coronaviruses. J Exp Med 2020; 217: e20200537.
15) Suaifan GARY, Alkhawaja BA MA. RNA Coronavi-ruses’ Outbreaks: Recent Progress on the SARS-CoV-2 Pandemic Diagnostic Tests, Vaccination and Therapeutics. Mini Rev Med Chem 2021; 2021: 1-17.
16) Tiwari R, Dhama K, Sharun K, Iqbal Yatoo M, Malik YS, Singh R, Michalak I, Sah R, Bonilla-Aldana DK, Rodriguez-Morales AJ. COVID-19: animals, veteri-nary and zoonotic links. Vet Q 2020; 40: 169-182.
17) Dhama K, Khan S, Tiwari R, Sircar S, Bhat S, Ma-lik YS, Singh KP, Chaicumpa W, Bonilla-Aldana DK, Rodriguez-Morales AJ. Coronavirus disease 2019-COVID-19. Clin Microbiol Rev 2020; 33: 1-48.
18) Meo SA, Alhowikan AM, Khlaiwi TAL, Meo IM, Halepoto DM, Iqbal M, Usmani AM, Hajjar W, Ahmed N. Novel coronavirus 2019-nCoV: Preva-lence, biological and clinical characteristics com-parison with SARS-CoV and MERS-CoV. Eur Rev Med Pharmacol Sci 2020; 24: 2012-2019.

Comparative review of three human coronaviruses
7175
19) Gorbalenya AE, Baker SC, Baric RS, de Groot RJ, Drosten C, Gulyaeva AA, Haagmans BL, Lauber C, Leontovich AM, Neuman BW, Penzar D,Perl-man S, Poon LLM, Samborskiy DV , Sidorov IA, Sola I, Ziebuhr J.The species Severe acute respi-ratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Mi-crobiol 2020; 5: 536-544.
20) Noor R. A comparative review of pathogene-sis and host innate immunity evasion strategies among the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), severe acute respi-ratory syndrome coronavirus (SARS-CoV) and the Middle East respiratory syndrome coro. Arch Microbiol 2021; 203: 1943-1951.
21) Chan JFW, Lau SKP, To KKW, Cheng VCC, Woo PCY, Yue KY. Middle East Respiratory syndrome coronavirus: Another zoonotic betacoronavirus causing SARS-like disease. Clin Microbiol Rev 2015; 28: 465-522.
22) WHO. WHO COVID-19 Dashboard - Up to date data on pandemic. WHO Heal Emerg Dasboard 2021.
23) To KKW, Tsang OTY, Leung WS, Tam AR, Wu TC, Lung DC, Yip CCY, Cai JP, Chan JMC, Chik TSH, Lau DPL, Choi, CYC, Chen LL, Chan WM, Chan K-H, Daniel J, Chin-Ki A, Poon RWS, Luo CT, Cheng VCC, Chan JFW, Yuen KY. Temporal pro-files of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: an observational co-hort study. Lancet Infect Dis 2020; 20: 565-574.
24) Lam TTY, Shum MHH, Zhu HC, Tong YG, Ni XB, Liao YS, Wei W, Cheung WYM, Li WJ, Li LF, Ji-ang BG, Wei W, Yuan TT, Zheng K, Cui XM, Li J, Pei GQ, Qiang X, Cheung WY, Cao W. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 2020; 583: 282-285.
25) Ioannidis JPA. The infection fatality rate of COVID-19 inferred from seroprevalence data. medRxiv 2020; 2020.05.13.20101253.
26) Abdallat MM, Abroug F, Al Dhahry SHS, Alha-jri MM, Al-Hakeem R, Al Hosani FI, Al Qasrawi SMA, Al-Romaihi HE, Assiri A, Baillie JK,Ben Embarek PK, Ben Salah A, Blümel B, Briese T, Buchholz U, Cognat SBF, Defang GN, De La Roc-que S, Donatelli I, Drosten C, Drury PA, Eremin SR,Ferguson NM, Fontanet A, Formenty PBH,-Fouchier RAM, Gao CQ, Garcia E, Gerber SI, Guery B, Haagmans BL, Haddadin AJ, Hardiman MC, Hensley LE, Hugonnet SAL, Hui DSC, Isla N, Karesh WB, Koopmans M, Kuehne A, Lipkin WI, Mafi AR, Malik M, Manuguerra JC, Memish Z, Mounts AW, Mumford E, Opoka L, Osterhaus A, John Oxenford C, Pang J, Pebody R, Peiris JSM, Jay Plotkin B, Poumerol G, Reusken C, Rezza G, Roth CE, Shindo N, Shumate AM, Siwula M, Slim A, Smallwood C, van der Werf S, Van Kerkhove MD, Zambon M. State of knowledge and data gaps of middle east respiratory syndrome coro-navirus (MERS-CoV) in humans. PLoS Curr 2013; 5: 1-36.
27) Rabaan AA, Al-Ahmed SH, Sah R, Tiwari R, Ya-too MI, Patel SK, Pathak M, Malik YS, Dhama K, Singh KP, Bonilla-Aldana K, Haque S, Marti-
nez-Pulgarin DF, Rodriguez-Morales AJ, Leblebi-cioglu H. SARS-CoV-2/COVID-19 and advances in developing potential therapeutics and vaccines to counter this emerging pandemic. Ann Clin Mi-crobiol Antimicrob 2020; 19: 1-37.
28) Sharun K, Tiwari R, Iqbal Yatoo M, Patel SK, Na-tesan S, Dhama J, Malik YS, Harapan H, Singh RK, Dhama K. Antibody-based immunotherapeu-tics and use of convalescent plasma to counter COVID-19: advances and prospects. Expert Opin Biol Ther 2020; 20: 1033-1046.
29) Iqbal Yatoo M, Hamid Z, Parray OR, Wani AH, Ul Haq A, Saxena A, Patel SK, Pathak M, Tiwari R, Malik YS, Sah R, Rabaan AA, Rodriguez Morales AJ, Dhama K. COVID-19 - Recent advancements in identifying novel vaccine candidates and cur-rent status of upcoming SARS-CoV-2 vaccines. Hum Vaccines Immunother 2020; 16: 2891-2904.
30) Iqbal Yatoo M, Hamid Z, Rather I, Nazir QUA, Bhat RA, Ul Haq A, Magray SN, Haq Z, Sah R, Tiwari R, Natesan S, Bilal M, Harapan H, Dha-ma K. Immunotherapies and immunomodulatory approaches in clinical trials - a mini review. Hum Vaccines Immunother 2021; 17: 1897-1909.
31) WHO Covid-19. Draft landscape of COVID-19 candidate vaccines. WHO 2020; 3.
32) Dallocchio RN, Dessì A, De Vito A, Delogu G, Ser-ra PA, Madeddu G. Early combination treatment with existing HIV antivirals: An effective treatment for COVID-19? Eur Rev Med Pharmacol Sci 2021; 25: 2435-2448.
33) Wang D, Hu B, Hu C, Zhu F, Liu X, Zhang J, Wang B, Xiang H, Cheng Z, Xiong Y, Zhao Y, Li Y, Wang X, Peng, Z. Clinical Characteristics of 138 Hospi-talized Patients with 2019 Novel Coronavirus-In-fected Pneumonia in Wuhan, China. JAMA - J Am Med Assoc 2020; 323: 1061-1069.
34) Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, He JX, Liu L, Shan H, Lei CL, Hui DSC,Du B, Li LJ, Zeng G, Yuen KY , Chen RC , Tang CL , Wang T, Chen PY, Xiang J, Li SY, Wang JL, Liang ZJ, Peng YX, Wei L, Liu Y, Hu YH, Peng P, Wang JM, Liu JY, Chen Z, Li G, Zheng ZJ, Qiu SQ, Luo J, Ye CJ, Zhu SY, Zhong NS.China Medical Treatment Ex-pert Group for Covid-19. Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med 2020; 382: 1708-1720.
35) Wu JT, Leung K, Leung GM. Nowcasting and Forecasting the Potential Domestic and Interna-tional Spread of the 2019-nCoV Outbreak Origi-nating in Wuhan, China: A Modeling Study. Lan-cet 2020; 395: 689-697.
36) Xiao K, Zhai J, Feng Y, Zhou N, Zhang X, Zou JJ, Li N, Guo Y, Li X, Shen X, Zhang Z, Shu F, Huang W, Li Y, Zhang Z, Chen RA, Wu YJ, Peng SM, Huang M, Xie WJ, Cai QH, Hou FH, Chen W, Xiao L, Shen Y.Isolation of SARS-CoV-2-relat-ed coronavirus from Malayan pangolins. Nature 2020; 583: 286-289.
37) Ruan Q, Yang K, Wang W, Jiang L, Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med 2020; 46: 846-848.
38) Wu Z, McGoogan JM. Characteristics of and Im-portant Lessons from the Coronavirus Disease

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7176
2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases from the Chinese Center for Disease Control and Prevention. JAMA - J Am Med Assoc 2020;323:1239-1242.
39) Wu A, Peng Y, Huang B, Ding X, Wang X, Niu P, Meng J, Zhu Z, Zhang Z, Wang J, Sheng J, Quan L, Xia Z, Tan W, Cheng G, Jiang T.Genome Com-position and Divergence of the Novel Coronavirus (2019-nCoV) Originating in China. Cell Host Mi-crobe 2020; 27: 325-328.
40) Hamming I, Timens W, Bulthuis MLC, Lely AT, Na-vis GJ, van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS corona-virus. A first step in understanding SARS patho-genesis. J Pathol 2004; 203: 631-637.
41) Raj VS, Mou H, Smits SL, Dekkers DH, Müller MA, Dijkman R, Muth D, Demmers JA, Zaki A, Fouchi-er RA, Thiel V. Drosten C, Rottier PJM, Osterhaus ADME, Bosch BJ, Haagmans BL.Dipeptidyl pepti-dase 4 is a functional receptor for the emerging hu-man coronavirus-EMC. Nature 2013; 495: 251-254.
42) Lu R, Zhao X, Li J, Niu P, Yang B, Wu H, Wang W, Song H, Huang B, Zhu N,Bi Y, Ma X, Zhan F, Wang L, Hu T, Zhou H, Hu Z, Zhou W, Zhao L, Chen J, Meng Y, Wang J, Lin Y, Yuan J, Xie Z, Ma J, Liu WJ, Wang D, Xu W, Holmes EC, Gao GF, Wu G, Chen W, Shi W, Tan W.Genomic charac-terisation and epidemiology of 2019 novel coro-navirus: implications for virus origins and receptor binding. Lancet 2020; 395: 565-574.
43) Zhou P, Yang X Lou, Wang XG, Hu B, Zhang L, Zhang W, Si HR, Zhu Y, Li B, Huang CL,Chen HD, Chen J, Luo Y, Guo H, Jiang RD, Liu MQ, Chen Y, Shen XR, Wang X, Zheng XS, Zhao K, Chen QJ, Deng F, Liu LL, Yan B, Zhan FX, Wang YY, Xiao GF, Shi ZL.A pneumonia outbreak associat-ed with a new coronavirus of probable bat origin. Nature 2020; 579: 270-273.
44) Ozono S, Zhang Y, Ode H, Sano K, Tan TS, Imai K, Miyoshi K, Kishigami S, Ueno T, Iwatani Y,Su-zuki T, Tokunaga K.SARS-CoV-2 D614G spike mutation increases entry efficiency with enhanced ACE2-binding affinity. Nat Commun 2021; 12: 1-9.
45) Wu F, Zhao S, Yu B, Chen YM, Wang W, Song ZG, Hu Y, Tao ZW, Tian JH, Pei YY,Yuan ML, Zhang YL, Dai FH, Liu Y, Wang QM, Zheng JJ, Xu L, Holmes EC, Zhang YZ.A new coronavirus as-sociated with human respiratory disease in China. Nature 2020;579: 265-269.
46) Walls AC, Park Y-J, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, Function, and Antigenic-ity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020; 181: 281-292.
47) Wrapp D, Wang N, Corbett KS, Goldsmith JA, Hsieh CL, Abiona O, Graham BS, McLellan JS. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. bioRxiv 2020; 10.1101/2020.02.11.944462.
48) Benvenuto D, Giovanetti M, Ciccozzi A, Spoto S, Angeletti S, Ciccozzi M. The 2019-new coronavi-rus epidemic: Evidence for virus evolution. J Med Virol 2020; 92: 455-459.
49) Epidemiology Working Group for NCIP Epidemic Response, Chinese Center for Disease Control
and Prevention. The epidemiological characteris-tics of an outbreak of 2019 novel coronavirus dis-eases (COVID-19) in China. Zhonghua Liu Xing Bing Xue Za Zhi 2020; 41: 145-151.
50) D’Cruz RJ, Currier AW, Sampson VB. Laboratory Testing Methods for Novel Severe Acute Respi-ratory Syndrome-Coronavirus-2 (SARS-CoV-2). Front Cell Dev Biol 2020; 8: 468.
51) Wang WK, Chen SY, Liu IJ, Kao CL, Chen HL, Chiang BL, Wang JT, Sheng WH, Hsueh PR, Yang CF, Yang PC, Chang SC. Temporal relation-ship of viral load, ribavirin, interleukin (IL)-6, IL-8, and clinical progression in patients with severe acute respiratory syndrome. Clin Infect Dis 2004; 39: 1071-1075.
52) Chen W, Xu Z, Mu J, Yang L, Gan H, Mu F, Fan B, He B, Huang S, You B, Yang Y,TangX, Qiu L, Qiu Y, WenJ, FangJ, Wang J. Antibody response and viraemia during the course of severe acute respi-ratory syndrome (SARS)-associated coronavirus infection. J Med Microbiol 2004; 53: 435-438.
53) Choi HM, Moon SY, Yang HI, Kim KS. Under-standing viral infection mechanisms and patient symptoms for the development of covid-19 thera-peutics. Int J Mol Sci 2021; 22: 1-25.
54) Hung IFN, Cheng VCC, Wu AKL, Tang BSF, Chan KH, Chu CM, Wong MML, Hui WT, Poon LLM, Tse DMW, ChanKS, WooPCY, LauSKP, PeirisJSM, YuenKY.Viral loads in clinical specimens and SARS manifestations. Emerg Infect Dis 2004; 10: 1550-1557.
55) Chu CM, Poon LLM, Cheng VCC, Chan KS, Hung IFN, Wong MML, Chan KH, Leung WS, Tang BSF, Chan VL,NgWL, SimTC, NgPW, LawKI, TseDMW, PeirisJSM, YuenKY. Initial viral load and the out-comes of SARS. Cmaj 2004; 171: 1349-1352.
56) Fajnzylber J, Regan J, Coxen K, Corry H, Wong C, Rosenthal A, Worrall D, Giguel F, Piechoc-ka-Trocha A, Atyeo C, Fischinger S, Chan A, Flaherty KT, Hall K, Dougan M, Ryan ET, Gilles-pie E, Chishti R, Li Y, Jilg N, Hanidziar D, Baron RM, Baden L, Tsibris AM, Armstrong KA, Kuritz-kes DR, Alter G, Walker BD, Yu X, Li JZ, Betty Abayneh BA, Allen P, Antille D, Balazs A, Bals J, Barbash M, Bartsch Y, Boucau J, Boyce S, Bra-ley J, Branch K, Broderick K, Carney J, Cheva-lier J, Choudhary MC, Chowdhury N, Daley G, Davidson S, Desjardins M, Donahue L, Drew D, Einkauf K, Elizabeth S, Elliman A, Etemad B, Fal-lon J, Fedirko L, Finn K, Flannery J, Forde P, Gar-cia-Broncano P, Gettings E, Golan D, Goodman K, Griffin A, Grimmel S, Grinke K, Hartana CA, Healy M, Heller H, Henault D, Holland G, Jiang C, Jordan H, Kaplonek P, Karlson EW, Karpell M, Kayitesi C, Lam EC, LaValle V, Lefteri K, Lian X, Lichterfeld M, Lingwood D, Liu H, Liu J, Lopez K, Lu Y, Luthern S, Ly NL, MacGowan M, Magispoc K, Marchewka J, Martino B, McNamara R, Michell A, Millstrom I, Miranda N, Nambu C, Nelson S, Noone M, Novack L, O’Callaghan C, Ommerborn C, Osborn M, Pacheco LC, Phan N, Pillai S, Porto FA, Rassadkina Y, Reissis A, Ruzicka F, Seiger K, Selleck K, Sessa L, Sharpe A, Sharr C, Shin S, Singh N, Slaughenhaupt S, Sheppard KS, Sun W, Sun X, Suschana EL, Talabi O, Ticheli H, Weiss

Comparative review of three human coronaviruses
7177
ST, Wilson V, Zhu A. SARS-CoV-2 viral load is associated with increased disease severity and mortality. Nat Commun 2020; 11: 1-9.
57) Ganesh B, Rajakumar T, Malathi M, Manikandan N, Nagaraj J, Santhakumar A, Elangovan A, Ma-lik YS. Epidemiology and pathobiology of SARS-CoV-2 (COVID-19) in comparison with SARS, MERS: An updated overview of current knowl-edge and future perspectives. Clin Epidemiol Glob Heal 2021; 10: 100694.
58) GISAID Global Initiative on Sharing All Influenza Data. Phylogeny of SARS-like betacoronaviruses in-cluding novel coronavirus (nCoV) 2020; 22: 30494.
59) Malik YS, Sircar S, Bhat S, Sharun K, Dhama K, Dadar M, Tiwari R, Chaicumpa W. Emerging novel coronavirus (2019-nCoV)—current scenario, evo-lutionary perspective based on genome analysis and recent developments. Vet Q 2020; 40: 68-76.
60) Mohapatra RK, Pintilie L, Kandi V, Sarangi AK, Das D, Sahu R, Perekhoda L. The recent chal-lenges of highly contagious COVID-19, causing respiratory infections: Symptoms, diagnosis, transmission, possible vaccines, animal models, and immunotherapy. Chem Biol Drug Des 2020; 96: 1187-1208.
61) Zhou H, Chen X, Hu T, Li J, Song H, Liu Y, Wang P, Liu D, Yang J, Holmes E, HughesA, BiY, ShiW. A novel bat coronavirus reveals natural insertions at the S1/S2 cleavage site of the Spike protein and a possible recombinant origin of HCoV-19. bioRx-iv2020; 10.1101/2020.03.02.974139.
62) Zheng J. SARS-coV-2: An emerging coronavirus that causes a global threat. Int J Biol Sci 2020; 16: 1678-1685.
63) Kaur N, Singh R, Dar Z, Bijarnia RK, Dhingra N, Kaur T. Genetic comparison among various coro-navirus strains for the identification of potential vaccine targets of SARS-CoV2. Infect Genet Evol 2021; 89: 104490.
64) Kumar V. Emerging Human Coronavirus Infec-tions (SARS, MERS, and COVID-19): Where They Are Leading Us. Int Rev Immunol 2021; 40: 5-53.
65) Malik YA. Properties of coronavirus and SARS-CoV-2. Malays J Pathol 2020; 42: 3-11.
66) Urra JM, Cabrera CM, Porras L, Ródenas I. Se-lective CD8 cell reduction by SARS-CoV-2 is as-sociated with a worse prognosis and systemic inflammation in COVID-19 patients. Clin Immunol 2020; 217: 108486.
67) Zhang T, Wu Q, Zhang Z. Probable Pangolin Ori-gin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr Biol 2020; 30: 1346-1351.
68) Yuan S, Jiang SC, Li ZL. Analysis of Possible In-termediate Hosts of the New Coronavirus SARS-CoV-2. Front Vet Sci 2020; 7: 379.
69) Brierley L, Fowler A. Predicting the animal hosts of coronaviruses from compositional biases of spike protein and whole genome sequences through ma-chine learning. PLoS Pathog 2021; 17: e1009149.
70) Ji W, Wang W, Zhao X, Zai J, Li X. Cross-species transmission of the newly identified coronavirus 2019-nCoV. J Med Virol 2020; 92: 433-440.
71) Qian J, Feng Y, Li J. Comments on “Cross-spe-cies transmission of the newly identified coronavi-rus 2019-nCoV.” J Med Virol 2020; 92: 1437-1439.
72) Liu Z, Xiao X, Wei X, Li J, Yang J, Tan H, Zhu J, Zhang Q, Wu J, Liu L. Composition and divergence of coronavirus spike proteins and host ACE2 re-ceptors predict potential intermediate hosts of SARS-CoV-2. J Med Virol 2020; 92: 595-601.
73) Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF. The proximal origin of SARS-CoV-2. Nat Med 2020; 26: 450-452.
74) Sun P, Qie S, Liu Z, Ren J, Li K, Xi J. Clinical char-acteristics of hospitalized patients with SARS-CoV-2 infection: A single arm meta-analysis. J Med Virol 2020; 92: 612-617.
75) Zhang YZ, Holmes EC. A Genomic Perspective on the Origin and Emergence of SARS-CoV-2. Cell 2020; 181: 223-227.
76) Ramanathan K, Antognini D, Combes A, Paden M, Zakhary B, Ogino M, Maclaren G, Brodie D. Ge-nomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet 2020; 2020: 19-21.
77) Kandeel M, Ibrahim A, Fayez M, Al-Nazawi M. From SARS and MERS CoVs to SARS-CoV-2: Moving toward more biased codon usage in viral structural and nonstructural genes. J Med Virol 2020; 92: 660-666.
78) Roy A, Guo F, Singh B, Gupta S, Paul K, Chen X, Sharma NR, Jaishee N, Irwin DM, Shen Y. Base Composition and Host Adaptation of the SARS-CoV-2: Insight From the Codon Usage Perspec-tive. Front Microbiol 2021; 12: 548275.
79) van Boheemen S, de Graaf M, Lauber C, Beste-broer TM, Raj VS, Zaki AM, Osterhaus ADME, Haagmans BL, Gorbalenya AE, Snijder EJ,Fouch-ierRAM.Genomic characterization of a newly dis-covered coronavirus associated with acute respi-ratory distress syndrome in humans. MBio 2012; 3: e00473-12.
80) Meyer B, Müller MA, Corman VM, Reusken CBEM, Ritz D, Godeke GJ, Lattwein E, Kallies S, Siemens A, van Beek J,DrexlerJF, MuthD, BoschBJ, WerneryU, KoopmansMPG, WerneryR, Drosten C.Antibodies against MERS coronavirus in dromedaries, United Arab Emirates, 2003 and 2013. Emerg Infect Dis 2014; 20: 552-559.
81) Stalin Raj V, Farag EABA, Reusken CBEM, Lam-ers MM, Pas SD, Voermans J, Smits SL, Oster-haus ADME, Al-Mawlawi N, Al-Romaihi HE,Alha-jriMM, El-SayedAM, MohranKA, GhobashyH, Al-hajriF, Al-ThaniM, Al-MarriSA, El-MaghrabyMM, KoopmansMPG, HaagmansBL.Isolation of MERS coronavirus from dromedary camel, Qatar, 2014. Emerg Infect Dis 2014; 20: 1339-1342.
82) Dudas G, Rambaut A. MERS-CoV recombination: implications about the reservoir and potential for adaptation. Virus Evol 2016; 2: vev023.
83) Wang Y, Liu D, Shi W, Lu R, Wang W, Zhao Y, Deng Y, Zhou W, Ren H, Wu J, Wang Y, Wu G, GaoGF, TanaW. Origin and possible genetic re-combination of the middle east respiratory syn-drome coronavirus from the first imported case in china: Phylogenetics and coalescence analysis. MBio 2015; 6: e01280-15.
84) Kenney SP, Helmy YA, Fawzy M, Elaswad A, So-bieh A, Shehata AA. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics,

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7178
Epidemiology, Diagnosis, Treatment, and Control. J Clin Med 2020; 9: 2-29.
85) Chu DKW, Hui KPY, Perera RAPM, Miguel E, Niemeyer D, Zhao J, Channappanavar R, Du-das G, Oladipo JO, Traoré A, Fassi-Fihri O, Ali A, Demissié GF, Muth D, Chan MCW, Nicholls JM, Meyerholz DK, Kuranga SA, Mamo G, Zhou Z, So RTY, Hemida MG, Webby RJ, Roger F, Rambaut A, Poon LLM, Perlman S, Drosten C, Chevalier V, Peiris M. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc Natl Acad Sci U S A 2018; 115: 3144-3149.
86) Hu B, Guo H, Zhou P, Shi ZL. Characteristics of SARS-CoV-2 and COVID-19. Nat Rev Microbiol 2021; 19: 141-154.
87) Sahin AR, Erdogan A, Agaoglu PM, Dineri Y, Ca-kirci AY, Senel ME, Okyay RA, Tasdogan AM. 2019 Novel Coronavirus (COVID-19) Outbreak: A Review of the Current Literature. Eurasian J Med Oncol 2020; 4: 1-7.
88) Krishnamoorthy P, Raj AS, Roy S, Kumar NS, Kumar H. Comparative transcriptome analysis of SARS-CoV, MERS-CoV, and SARS-CoV-2 to identify potential pathways for drug repurposing. Comput Biol Med 2021; 128: 104123.
89) De Albuquerque LP, Siqueira Patriota LL de, Gon-zatto V, Pontual EV, Guedes Paiva PM, Napoleão TH. Coronavirus Spike (S) Protein: A Brief Review on Structure-Function Relationship, Host Recep-tors, and Role in Cell Infection. Adv Res 2020; 28: 116-124.
90) Pandey P, Rane JS, Chatterjee A, Kumar A, Khan R, Prakash A, Ray S. Targeting SARS-CoV-2 spike protein of COVID-19 with naturally occurring phytochemicals: an in silico study for drug devel-opment. J Biomol Struct Dyn 2020; 2020: 1-11.
91) Kaur N, Singh R, Dar Z, Bijarnia RK, Dhingra N, Kaur T. Genetic comparison among various coro-navirus strains for the identification of potential vaccine targets of SARS-CoV2. Infect Genet Evol 2020; 89: 104490.
92) Liu DX, Fung TS, Chong KKL, Shukla A, Hilgen-feld R. Accessory proteins of SARS-CoV and oth-er coronaviruses. Antiviral Res 2014; 109: 97-109.
93) Fung TS, Liu DX. Similarities and Dissimilarities of COVID-19 and Other Coronavirus Diseases. Annu Rev Microbiol 2021; 75: 1-18.
94) Naqvi AAT, Fatima K, Mohammad T, Fatima U, Singh IK, Singh A, Atif SM, Hariprasad G, Hasan GM, Hassan MI. Insights into SARS-CoV-2 ge-nome, structure, evolution, pathogenesis and ther-apies: Structural genomics approach. Biochim Bio-phys Acta - Mol Basis Dis 2020; 1866: 165878.
95) Shi CS, Nabar NR, Huang NN, Kehrl JH. SARS-Coronavirus Open Reading Frame-8b trig-gers intracellular stress pathways and activates NLRP3 inflammasomes. Cell Death Discov 2019; 5: 1-12.
96) Yue Y, Nabar NR, Shi CS, Kamenyeva O, Xiao X, Hwang IY, Wang M, Kehrl JH. SARS-Corona-virus Open Reading Frame-3a drives multimodal necrotic cell death. Cell Death Dis 2018; 9: 1-15.
97) Wong HH, Fung TS, Fang S, Huang M, Le MT, Liu DX. Accessory proteins 8b and 8ab of severe acute respiratory syndrome coronavirus suppress
the interferon signaling pathway by mediating ubiquitin-dependent rapid degradation of inter-feron regulatory factor 3. Virology 2018; 515: 165-175.
98) Wong LYR, Ye ZW, Lui PY, Zheng X, Yuan S, Zhu L, Fung SY, Yuen KS, Siu KL, Yeung ML, Cai Z, Woo PCY, Yuen KY, Chan CP, Jin DY. Middle East Respiratory Syndrome Coronavirus ORF8b Ac-cessory Protein Suppresses Type I IFN Expres-sion by Impeding HSP70-Dependent Activation of IRF3 Kinase IKKε. J Immunol 2020; 205: 1564-1579.
99) V’kovski P, Kratzel A, Steiner S, Stalder H, Thiel V. Coronavirus biology and replication: implica-tions for SARS-CoV-2. Nat Rev Microbiol 2021; 19: 155-170.
100) Millán-Oñate J, Rodriguez-Morales AJ, Cama-cho-Moreno G, Mendoza-Ramírez H,Rodrí-guez-Sabogal IA, Álvarez-Moreno C. A new emerg-ing zoonotic virus of concern : the 2019 novel Coro-navirus (SARS CoV-2). Infection 2020; 24: 187-192.
101) Cappuccio FP, Siani A. Covid-19 and cardiovas-cular risk: Susceptibility to infection to SARS-CoV-2, severity and prognosis of Covid-19 and blockade of the renin-angiotensin-aldosterone system. An evidence-based viewpoint. Nutr Me-tab Cardiovasc Dis 2020; 30: 1227-1235.
102) L’Huillier AG, Torriani G, Pigny F, Kaiser L, Eck-erle I. Shedding of infectious SARS-CoV-2 in symptomatic neonates, children and adolescents. medRxiv 2020; 10.1101/2020.04.27.20076778.
103) de Wit E, van Doremalen N, Falzarano D, VJ M. SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol 2020; 14: 23-34.
104) Pal M, Berhanu G, Desalegn C, Kandi V. Se-vere Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2): An Update. Cureus 2020; 12: e7423.
105) Li W, Moore MJ, Vasllieva N, Sui J, Wong SK, Berne MA, Somasundaran M, Sullivan JL, Lu-zuriaga K, Greeneugh TC, Choe H, FarzanM. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003; 426: 450-454.
106) Gu J, Korteweg C. Pathology and pathogenesis of severe acute respiratory syndrome. Am J Pathol 2007; 170: 1136-1147.
107) Chen J, Subbarao K. The immunobiology of SARS. Annu Rev Immunol 2007; 25: 443-472.
108) Zhou T, Tsybovsky Y, Gorman J, Rapp M, Cerut-ti G, Chuang GY, Katsamba PS, Sampson JM, Schön A, Bimela J, Boyington JC, Nazzari A, Olia AS, Shi W, Sastry M, Stephens T, Stuckey J, Teng IT, Wang P, Wang S, Zhang B, Friesner RA, Ho DD, Mascola JR, Shapiro L, Kwong PD. Cryo-EM Structures of SARS-CoV-2 Spike without and with ACE2 Reveal a pH-Dependent Switch to Mediate Endosomal Positioning of Receptor-Binding Do-mains. Cell Host Microbe 2020; 28: 867-879.
109) Schoeman D, Fielding BC. Coronavirus envelope protein: Current knowledge. Virol J 2019; 16: 1-22.
110) Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, Guo L, Guo R, Chen T, Hu J, Xiang Z, Mu Z, ChenX, Chen J, Hu K, Jin Q, Wang J, Qian Z. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry

Comparative review of three human coronaviruses
7179
and its immune cross-reactivity with SARS-CoV. Nat Commun 2020; 11: 1-12.
111) Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, MüllerMA, Dro-stenC, PöhlmannS.SARS-CoV-2 Cell Entry De-pends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020; 181: 271-280.
112) Wan Y, Shang J, Sun S, Tai W, Chen J, Geng Q, He L, Chen Y, Wu J, Shi Z, ZhouY, DuL, LiF.Mo-lecular Mechanism for Antibody-Dependent En-hancement of Coronavirus Entry. J Virol 2019; 94: e02015-19.
113) Tay MZ, Poh CM, Rénia L, MacAry PA, Ng LFP. The trinity of COVID-19: immunity, inflammation and in-tervention. Nat Rev Immunol 2020; 20: 363-374.
114) Wrapp D, Wang N, Corbett KS, Goldsmith JA, Hsieh CL, Abiona O, Graham BS, McLellan JS. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020; 367: 1260-1263.
115) Chu H, Chan JFW, Yuen TTT, Shuai H, Yuan S, Wang Y, Hu B, Yip CCY, Tsang JOL, Huang X, Chai Y, Yang D, Hou Y, Chik KKH, Zhang X, Fung AYF, Tsoi HW, Cai JP, Chan WM, Ip JD, Chu AWH, Zhou J, Lung DC, Kok KH, To KKW, Tsang OTY, Chan KH, Yuen KY. Comparative tropism, replication kinetics, and cell damage profiling of SARS-CoV-2 and SARS-CoV with implications for clinical manifestations, transmissibility, and laboratory studies of COVID-19: an observational study. The Lancet Microbe 2020; 1: e14-e23.
116) Sigrist CJ, Bridge A, Le Mercier P. A potential role for integrins in host cell entry by SARS-CoV-2. Antiviral Res 2020; 177: 104759.
117) Wang K, Chen W, Zhang Z, Deng Y, Lian JQ, Du P, Wei D, Zhang Y, Sun XX, Gong L,Yang X, He L, Zhang L, Yang Z, Geng JJ, Chen R, Zhang H, Wang B, Zhu YM, Nan G, Jiang JL, Li L, Wu J, Lin P, Huang W, Xie L, Zheng ZH, Zhang K, Miao JL, Cui HY, Huang M, Zhang J, Fu L, Yang XM, Zhao Z, Sun S, Gu H, Wang Z, Wang CF, Lu Y, Liu YY, Wang QY, Bian H, Zhu P, Chen ZN. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct Target Ther 2020; 5: 1-10.
118) Cui J, Li F, Shi ZL. Origin and evolution of patho-genic coronaviruses. Nat Rev Microbiol 2019; 17: 181-192.
119) Xu H, Zhong L, Deng J, Peng J, Dan H, Zeng X, Li T, Chen Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int J Oral Sci 2020; 12: 1-5.
120) Sun K, Gu L, Ma L, Duan Y. Atlas of ACE2 gene expression reveals novel insights into transmis-sion of SARS-CoV-2. Heliyon 2021; 7: e05850.
121) Buqaileh R, Saternos H, Ley S, Aranda A, Fore-ro K AW. Can cilia provide an entry gateway for SARS-CoV-2 to human ciliated cells? Physiol Ge-nomics 2021; 53: 249-258.
122) Fani M, Teimoori A, Ghafari S. Comparison of the COVID-2019 (SARS-CoV-2) pathogenesis with SARS-CoV and MERS-CoV infections. Future Vi-rol 2020; 15: 317-323.
123) Khan S, El Morabet R, Khan RA, Bindajam A, Alqadhi S, Alsubih M, Khan NA. Where we
missed? Middle East Respiratory Syndrome (MERS-CoV) epidemiology in Saudi Arabia; 2012-2019. Sci Total Environ 2020; 747: 141369.
124) Navand, Soltani S, Moghadami M, Hosseini P, Nasimzadeh S, Zandi M. Diabetes and corona-virus infections (SARS-CoV, MERS-CoV, and SARS-CoV-2). J Acute Dis 2020; 9: 244-247.
125) Li Y, Zhang Z, Yang L, Lian X, Xie Y, Li S, Xin S, Cao P, Lu J. The MERS-CoV Receptor DPP4 as a Candidate Binding Target of the SARS-CoV-2 Spike. iScience 2020; 23: 101160.
126) Deacon CF. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treat-ment of Type 2 Diabetes. Front Endocrinol (Laus-anne) 2019; 10: 80-99.
127) Somsen GA, van Rijn C, Kooij S, Bem RA, Bonn D. Small droplet aerosols in poorly ventilated spaces and SARS-CoV-2 transmission. Lancet Respir Med 2020; 8: 658-659.
128) Dhama K, Patel SK, Pathak M, Yatoo MI, Tiwari R, Malik YS, Singh R, Sah R, Rabaan AA, Bonil-la-Aldana DK R-MA. An update on SARS-CoV-2/COVID-19 with particular reference to its clinical pathology, pathogenesis, immunopathology and mitigation strategies. Travel Med Infect Dis 2020; 37: 101755.
129) Tripathi S, Pandey MK, Malik YS, Bilal M, Dha-ma K, Chaicumpa W, Chandra R. Novel corona-virus (Sars-cov-2): Molecular biology, pathogen-esis, pathobiology and advances in treatment of covid-19 patients- an update. J Exp Biol Agric Sci 2020; 8: 683-708.
130) Chua RL, Lukassen S, Trump S, Hennig BP, Wen-disch D, Pott F, Debnath O, Thürmann L, Kurth F, Völker MT, Kazmierski J, Timmermann B, Twardziok S, Schneider S, Machleidt F, Müller-Re-detzky H, Maier M, Krannich A, Schmidt S, Balzer F, Liebig J, Loske J, Suttorp N, Eils J, Ishaque N, Liebert UG, von Kalle C, Hocke A, Witzenrath M, Goffinet C, Drosten C, Laudi S, Lehmann I, Conrad C, Sander LE, Eils R.COVID-19 severi-ty correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat Biotechnol 2020; 38: 970-979.
131) Tsai SF, Lu KY, Chuang HM, Liu CA. Surviving the Rookie Virus, Severe Acute Respiratory Syn-drome Coronavirus 2 (SARS-CoV2): The Immu-nopathology of a SARS-CoV2 Infection. Cell Transplant 2021; 30: 096368972199376.
132) Laporte M, Stevaert A, Raeymaekers V, van Ber-waer R, Martens K, Pöhlmann S, Naesens L. The SARS-CoV-2 and other human coronavirus spike proteins are fine-tuned towards temperature and proteases of the human airways. PLoS Pathog 2021; 17: e1009500.
133) Halaji M, Farahani A, Ranjbar R, Heiat M, Dehkordi FS. Emerging coronaviruses: First SARS, second MERS and third SARS-COV-2. epidemiological updates of COVID-19. Infez Med 2020; 28: 6-17.
134) Arumugam VA, Thangavelu S, Fathah Z, Ravindran P, Sanjeev AMA, Babu S, Meyyazhagan A, Yatoo MI, Sharun K, Tiwari R, PandeyMK, SahR, ChandraR, DhamaK.COVID-19 and the world with co-morbidi-ties of heart disease, hypertension and diabetes. J Pure Appl Microbiol 2020; 14: 1623-1638.

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7180
135) Corvol P. The renin-angiotensin-aldosterone sys-tem and its suppression. Ann Pediatr (Paris) 1982; 29: 657-658.
136) Oz M, Lorke DE. Multifunctional angiotensin con-verting enzyme 2, the SARS-CoV-2 entry recep-tor, and critical appraisal of its role in acute lung injury. Biomed Pharmacother 2021; 136: 111193.
137) Cheng H, Wang Y, Wang GQ. Organ-protective effect of angiotensin-converting enzyme 2 and its effect on the prognosis of COVID-19. J Med Virol 2020; 92: 726-730.
138) Schaller T, Hirschbühl K, Burkhardt K, Braun G, Trepel M, Märkl B, Claus R. Postmortem Exam-ination of Patients with COVID-19. JAMA - J Am Med Assoc 2020; 323: 2518-2520.
139) Ogando NS, Zevenhoven-Dobbe JC, van der Meer Y, Bredenbeek PJ, Posthuma CC, Snijder EJ. The Enzymatic Activity of the nsp14 Exoribo-nuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2. J Virol 2020; 94: e01246-20.
140) Narayanan K, Huang C, Lokugamage K, Kamitani W, Ikegami T, Tseng CT, Makino S. Severe acute respiratory syndrome coronavirus nsp1 suppresses host gene expression, including that of type I inter-feron, in infected cells. J Virol 2008; 82: 4471-4479.
141) Lokugamage KG, Narayanan K, Nakagawa K, Terasaki K, Ramirez SI, Tseng CTK, Makino S. Middle East Respiratory Syndrome Coronavirus nsp1 Inhibits Host Gene Expression by Selective-ly Targeting mRNAs Transcribed in the Nucleus while Sparing mRNAs of Cytoplasmic Origin. J Virol 2015; 89: 10970-10981.
142) Thoms M, Buschauer R, Ameismeier M, Koepke L, Denk T, Hirschenberger M, Kratzat H, Hayn M, MacKens-Kiani T, Cheng J, Straub JH, Stür-zel CM, Fröhlich T, Berninghausen O, Becker T, Kirchhoff F, Sparrer KMJ, Beckmann R. Structural basis for translational shutdown and immune eva-sion by the Nsp1 protein of SARS-CoV-2. Science 2020; 369: 1249-1256.
143) Snijder EJ, van der Meer Y, Zevenhoven-Dobbe J, Onderwater JJM, van der Meulen J, Koerten HK, Mommaas AM. Ultrastructure and Origin of Mem-brane Vesicles Associated with the Severe Acute Respiratory Syndrome Coronavirus Replication Complex. J Virol 2006; 80: 5927-5940.
144) Hemmat N, Asadzadeh Z, Ahangar NK, Alemo-hammad H, Najafzadeh B, Derakhshani A, Bagh-banzadeh A, Baghi HB, Javadrashid D, Najafi S, Ar Gouilh M, Baradaran B.The roles of signal-ing pathways in SARS-CoV-2 infection; lessons learned from SARS-CoV and MERS-CoV. Arch Virol 2021; 166: 675-696.
145) Granados A, Peci A, McGeer A, Gubbay JB. Influ-enza and rhinovirus viral load and disease severi-ty in upper respiratory tract infections. J Clin Virol 2017; 86: 14-19.
146) Jiang X, Rayner S, Luo MH. Does SARS-CoV-2 has a longer incubation period than SARS and MERS? J Med Virol 2020; 92: 476-478.
147) Wen J, Cheng Y, Ling R, Dai Y, Huang B, Huang W, Zhang S, Jiang Y. Antibody-dependent en-hancement of coronavirus. Int J Infect Dis 2020; 100: 483-489.
148) The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19) in China. Zhonghua Liu Xing Bing Xue Za Zhi 2020; 41: 145-151.
149) Miorin L, Kehrer T, Sanchez-Aparicio MT, Zhang K, Cohen P, Patel RS, Cupic A, Makio T, Mei M, Moreno E, Danziger O, White KM, Rathnasinghe R, Uccellini M, Gao S, Aydillo T, Mena I, Yin X, Martin-Sancho L, Krogan NJ, Chanda SK, Schot-saert M, Wozniak RW, Ren Y, Rosenberg BR, Fontoura BMA, García-Sastre A.SARS-CoV-2 Orf6 hijacks Nup98 to block STAT nuclear import and antagonize interferon signaling. Proc Natl Acad Sci U S A 2020; 117: 28344-28354.
150) Sinderewicz E, Czelejewska W, Jezier-ska-Wozniak K, Staszkiewicz-Chodor J, Maksy-mowicz W. Immune response to covid-19: Can we benefit from the sars-cov and mers-cov pandemic experience? Pathogens 2020; 9: 739.
151) Thieme CJ, Anft M, Paniskaki K, Blazquez-Navar-ro A, Doevelaar A, Seibert FS, Hoelzer B, Konik MJ, Berger MM, Brenner T, Tempfer C, Watzl C, Meister TL, Pfaender S, Steinmann E, Dolff S, Ditt-mer U, Westhoff TH, Witzke O, Stervbo U, Roch T, Babel N. Robust T Cell Response Toward Spike, Membrane, and Nucleocapsid SARS-CoV-2 Pro-teins Is Not Associated with Recovery in Critical COVID-19 Patients. Cell Reports Med 2020; 1: 100092.
152) Mahallawi WH, Khabour OF, Zhang Q, Makhdoum HM, Suliman BA. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine 2018; 104: 8-13.
153) Kumar S, Nyodu R, Maurya VK, Saxena SK. Host Immune Response and Immunobiology of Human SARS-CoV-2 Infection. 2020; 2020: 43-53.
154) Bouvet M, Debarnot C, Imbert I, Selisko B, Snijder EJ, Canard B, Decroly E. In vitro reconstitution of sars-coronavirus mRNA cap methylation. PLoS Pathog 2010; 6: 1-13.
155) Chen L, Zhao J, Peng J, Li X, Deng X, Geng Z, Shen Z, Guo F, Zhang Q, Jin Y,WangL, WangS.Detection of 2019-nCoV in Saliva and Character-ization of Oral Symptoms in COVID-19 Patients. Cell Prolif 2020; 53: e12923.
156) Rakib A, Sami SA, Mimi NJ, Chowdhury MM, Eva TA, Nainu F, Paul A, Shahriar A, Tareq AM, Emon NU, Chakraborty S, Shil S, Mily SJ, Hadda TB, Al-malki FA, Emran TB. Immunoinformatics-guided design of an epitope-based vaccine against severe acute respiratory syndrome coronavirus 2 spike glycoprotein. Comput Biol Med 2020; 124: 103967.
157) Pormohammad A, Ghorbani S, Khatami A, Farzi R, Baradaran B, Turner DL, Turner RJ, Mansour-nia MA, Kyriacou DN, Bahr NC, IdrovoJ-P.Com-parison of Confirmed COVID-19 with SARS and MERS Cases - Clinical Characteristics, Laborato-ry Findings, Radiographic Signs and Outcomes: A Systematic Review and Meta-Analysis. Rev Med Virol 2020; 30: e2112.
158) Gao Y, Li T, Han M, Li X, Wu D, Xu Y, Zhu Y, Liu Y, Wang X, Wang L. Diagnostic utility of clinical lab-oratory data determinations for patients with the severe COVID-19. J Med Virol 2020; 92: 791-796.

Comparative review of three human coronaviruses
7181
159) Al-Tawfiq JA, Hinedi K, Abbasi S, Babiker M, Sun-ji A, Eltigani M. Hematologic, hepatic, and renal function changes in hospitalized patients with Middle East respiratory syndrome coronavirus. Int J Lab Hematol 2017; 39: 272-278.
160) Wang JT, Sheng WH, Fang CT, Chen YC, Wang JL, Yu CJ, Chang SC, Yang PC. Clinical Manifes-tations, Laboratory Findings, and Treatment Out-comes of SARS Patients. Emerg Infect Dis 2004; 10: 818-824.
161) Giannis D, Ziogas IA, Gianni P. Coagulation disor-ders in coronavirus infected patients: COVID-19, SARS-CoV-1, MERS-CoV and lessons from the past. J Clin Virol 2020; 127.
162) Ma C, Cong Y, Zhang H. COVID-19 and the Di-gestive System. Am J Gastroenterol 2020; 115: 1003-1006.
163) Zhou Z, Zhao N, Shu Y, Han S, Chen B, Shu X. Ef-fect of Gastrointestinal Symptoms in Patients With COVID-19. Gastroenterology 2020; 158: 2294-2297.
164) Loganathan S, Kuppusamy M, Wankhar W, Gurugubelli KR, Mahadevappa VH, Lepcha L, Choudhary AK. Angiotensin-converting enzyme 2 (ACE2): COVID-19 gate way to multiple organ fail-ure syndromes. Respir Physiol Neurobiol 2021; 283: 103548.
165) Chai X, Hu L, Zhang Y, Han W, Lu Z, Ke A, Zhou J, Shi G, Fang N, Fan J, CaiJ, FanJ, LanF. Specif-ic ACE2 expression in cholangiocytes may cause liver damage after 2019-nCoV infection. bioRxiv 2020; 10.1101/2020.02.03.931766.
166) Lei HY, Ding YH, Nie K, Dong YM, Xu JH, Yang ML, Liu MQ, Wei L, Nasser MI, Xu LY, ZhuP, ZhaoMY. Potential effects of SARS-CoV-2 on the gastrointestinal tract and liver. Biomed Pharma-cother 2021; 133: 111064.
167) Puelles VG, Lütgehetmann M, Lindenmeyer MT, Sperhake JP, Wong MN, Allweiss L, Chilla S, Heinemann A, Wanner N, Liu S, Braun F, Lu S, Pfefferle S, Schröder AS, Edler C, Gross O, Glat-zel M, Wichmann D, Wiech T, Kluge S, Pueschel K, Aepfelbacher M, Huber TB. Multiorgan and Renal Tropism of SARS-CoV-2. N Engl J Med 2020; 383: 590-592.
168) Montalvan V, Lee J, Bueso T, De Toledo J, Rivas K. Neurological manifestations of COVID-19 and other coronavirus infections: A systematic review. Clin Neurol Neurosurg 2020; 194: 105921.
169) Al-Washahi M, Al-Abri R. Loss of smell and taste are newly emerging symptoms in COVID-19 pa-tients necessitating more insights into their diag-nostic evaluation. Oman Med J 2021; 36: e257.
170) De Vito A, Geremia N, Fiore V, Princic E, Babud-ieri S, Madeddu G. Clinical features, laboratory findings and predictors of death in hospitalized patients with COVID-19 in Sardinia, Italy. Eur Rev Med Pharmacol Sci 2020; 24: 7861-7868.
171) Neumann B, Schmidbauer ML, Dimitriadis K, Otto S, Knier B, Niesen WD, Hosp JA, Günther A, Lindemann S, Nagy G,SteinbergT, LinkerRA, HemmerB, BöselJ.Cerebrospinal fluid findings in COVID-19 patients with neurological symptoms. J Neurol Sci 2020; 418: 117090.
172) Vaira LA, Deiana G, Lechien JR, De Vito A, Cos-su A, Dettori M, Del Rio A, Saussez S, Madeddu
G, Babudieri S, Fois AG, Clementina C, Hopkins C, De Riu G, Piana AF. Correlations Between Ol-factory Psychophysical Scores and SARS-CoV-2 Viral Load in COVID-19 Patients. Laryngoscope 2021; 42: 1560-1590.
173) Vaira LA, Hopkins C, Salzano G, Petrocelli M, Me-lis A, Cucurullo M, Ferrari M, Gagliardini L, Pipolo C, Deiana G, Fiore V, De Vito A, Turra N, Canu S, Maglio A, Serra A, Bussu F, Madeddu G, Babudieri S, Giuseppe Fois A, Pirina P, Salzano FA, De Riu P, Biglioli F, De Riu G. Olfactory and gustatory function impairment in COVID-19 patients: Italian objective multicenter-study. Head Neck 2020; 42: 1560-1569.
174) Hosseini MJ, Halaji M, Nejad JH, Ranjbar R. Cen-tral nervous system vasculopathy associated with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2): a novel case report from Iran. J Neurovirol 2021; 27: 507-509.
175) Vasanthapuram VH, Badakere A. Internucle-ar ophthalmoplegia as a presenting feature in a COVID-19-positive patient. BMJ Case Rep 2021; 14: e241873.
176) Conrady CD, Faia LJ, Gregg KS, Rao RC. Coro-navirus-19-Associated Retinopathy. Ocul Immu-nol Inflamm 2021; 2021: 1-2.
177) Gabarre P, Dumas G, Dupont T, Darmon M, Azou-lay E, Zafrani L. Acute kidney injury in critically ill patients with COVID-19. Intensive Care Med 2020; 46: 1339-1348.
178) Henry BM, Lippi G. Chronic kidney disease is as-sociated with severe coronavirus disease 2019 (COVID-19) infection. Int Urol Nephrol 2020; 52: 1193-1194.
179) Patel SK, Singh R, Rana J, Tiwari R, Natesan S, Harapan H, Arteaga-Livias K, Bonilla-Aldana DK, Rodríguez-Morales AJ, Dhama K. The kidney and COVID-19 patients - Important considerations. Travel Med Infect Dis 2020; 37: 101831.
180) Ronco C, Reis T. Kidney involvement in COVID-19 and rationale for extracorporeal therapies. Nat Rev Nephrol 2020; 16: 308-310.
181) Gäckler A, Rohn H, Witzke O. Acute renal failure in COVID-19. Nephrologe 2021; 16:66-70.
182) Hardenberg JHB, Stockmann H, Eckardt KU, Schmidt-Ott KM. COVID-19 and acute kidney injury in the inten-sive care unit. Nephrologe 2021; 16: 20-25.
183) Ng JH, Hirsch JS, Hazzan A, Wanchoo R, Shah HH, Malieckal DA, Ross DW, Sharma P, Sakhi-ya V, Fishbane S, Jhaveri KD, Abate M, Andrade HP, Barnett RL, Bellucci A, Bhaskaran MC, Co-rona AG, Flores Chang BS, Finger M, Gitman M, Halinski C, Hasan S, Hazzan AD, Hong S, KhaninY, KuanA, Madireddy V, Malieckal D, Muz-ib A, Nair G, Nair VV, Ng JH, Parikh R, Sachdeva M, Schwarz R, Singhal PC, Uppal NN. Outcomes Among Patients Hospitalized With COVID-19 and Acute Kidney Injury. Am J Kidney Dis 2021; 77: 204-215.e1.
184) Martinez-Rojas MA, Vega-Vega O, Bobadilla XNA. Is the kidney a target of SARS-CoV-2? Am J Physiol - Ren Physiol 2020; 318: F1454-F1462.
185) Chong WH, Saha BK. Relationship Between Se-vere Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) and the Etiology of Acute Kidney Injury (AKI). Am J Med Sci 2021; 361: 287-296.

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7182
186) Mitaka H, Kuno T, Takagi H PP. Incidence and Mortality of COVID-19-associated Pulmonary As-pergillosis: A Systematic Review and Meta-analy-sis. Mycoses 2021; 2021: 1-13.
187) Silva DL, Lima CM, Magalhães VCR, Baltazar LM, Peres NTA, Caligiorne RB, Moura AS, Fereguetti T, Martins JC, Rabelo LF, AbrahãoJS, LyonAC, JohannS, SantosDA. Fungal and bacterial coin-fections increase mortality of severely ill COVID-19 patients. J Hosp Infect 2021; 113: 145-154.
188) Zuo T, Liu Q, Zhang F, Yeoh YK, Wan Y, Zhan H, Lui GCY, Chen Z, Li AYL, Cheung CP, Chen N, Lv W, Ng RWY, Tso EYK, Fung KSC, Chan V, Ling L, Joynt G, Hui DSC, Chan FKL, Chan PKS,Ng SC. Temporal landscape of human gut RNA and DNA virome in SARS-CoV-2 infection and severity. Mi-crobiome 2021; 9: 1-16.
189) Zuo T, Zhang F, Lui GCY, Yeoh YK, Li AYL, Zhan H, Wan Y, Chung ACK, Cheung CP, Chen N, Lai CKC, Chen Z, Tso EYK, Fung KSC, Chan V, Ling L, Joynt G, Hui DSC, Chan FKL,Chan PKS, Ng SC. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastro-enterology 2020; 159: 944-955.
190) Zuo T, Zhan H, Zhang F, Liu Q, Tso EYK, Lui GCY, Chen N, Li A, Lu W, Chan FKL, Chan PKS, Ng SC. Alterations in Fecal Fungal Microbiome of Patients With COVID-19 During Time of Hospitalization until Discharge. Gastroenterology 2020; 159: 1302-1310.
191) Elhadi M, Alsoufi A, Msherghi A, Alshareea E, Ashini A, Nagib T, Abuzid N, Abodabos S, Alrifai H, Gresea E, Yahya W, Ashour D, Abomengal S, Qarqab N, Albibas A, Anaiba M, Idheiraj H, Abra-heem H, Fayyad M, Alkilani Y, Alsuwiyah S, El-ghezewi A, Zaid A. Psychological Health, Sleep Quality, Behavior, and Internet Use Among Peo-ple During the COVID-19 Pandemic: A Cross-Sec-tional Study. Front Psychiatry 2021; 12: 632496.
192) Cazzato G, Foti C, Colagrande A, Cimmino A, Scarcella S, Cicco G, Sablone S, Arezzo F, Romi-ta P, Lettini T,RestaL, IngravalloG.Skin Manifes-tation of SARS-CoV-2: The Italian Experience. J Clin Med 2021; 10: 1566.
193) Rabaan AA, Al-Ahmed SH, Muhammad J, Khan A, Sule AA, Tirupathi R, Mutair AA, Alhumaid S, Al-Omari A, Dhawan M, Tiwari R, Sharun K, Mo-hapatra RK, Mitra S, Bilal M, Alyami SA, Emran TB, Moni MA, Dhama K. Role of Inflammatory Cytokines in COVID-19 Patients: A Review on Molecular Mechanisms, Immune Functions, Im-munopathology and Immunomodulatory Drugs to Counter Cytokine Storm. Vaccines 2021; 9: 436.
194) Merad M, Martin JC. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nat Rev Immunol 2020; 20: 355-362.
195) Obando-Pereda G. Can molecular mimicry ex-plain the cytokine storm of SARS-CoV-2?: an in silico approach. J Med Virol 2021; 93: 5350-5357.
196) Song W-J, Hui CKM, Hull JH, Birring SS, Mc-Garvey L, Mazzone SB, Chung KF. Confronting COVID-19-associated cough and the post-COVID syndrome: role of viral neurotropism, neuroinflam-mation, and neuroimmune responses. Lancet Re-spir Med 2021; 9: 533-544.
197) Gunst JD, Staerke NB, Pahus MH, Kristensen LH, Bodilsen J, Lohse N, Dalgaard LS, Brønnum D, Fröbert O, Hønge B, Johansen IS, Monrad I, Erikstrup C, Rosendal R, Vilstrup E, Mariager T, Bove DG, Offersen R, Shakar S, Cajander S, Jør-gensen NP, Sritharan SS, Breining P, Jespersen S, Mortensen KL, Jensen ML, Kolte L, Frattari GS, Larsen CS, Storgaard M, Nielsen LP, Tolstrup M, Sædder EA, Østergaard LJ, Ngo HTT, Jensen MH, Højen JF, Kjolby M, Søgaard OS. Efficacy of the TMPRSS2 inhibitor camostat mesilate in pa-tients hospitalized with Covid-19-a double-blind randomized controlled trial. EClinicalMedicine 2021; 35: 100849.
198) Tan SH, Allicock O, Armstrong-Hough M, Wyllie AL. Saliva as a gold-standard sample for SARS-CoV-2 detection. Lancet Respir Med 2021; 9: 562-564.
199) Tsang NNY, So HC, Ng KY, Cowling BJ, Leung GM, Ip DKM. Diagnostic performance of different sampling approaches for SARS-CoV-2 RT-PCR testing: a systematic review and meta-analysis. Lancet Infect Dis 2021; 21: 1-13.
200) Kim JE, Heo JH, Kim HO, Song SH, Park SS, Park TH, Ahn JY, Kim MK, Choi JP. Neurological compli-cations during treatment of middle east respiratory syndrome. J Clin Neurol 2017; 13: 227-233.
201) Lai AL, Millet JK, Daniel S, Freed JH, Whittaker GR. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: a systematic review and meta-analysis. Int J Infect Dis 2020; 94: 91-95.
202) Khamis AH, Jaber M, Azar A, AlQahtani F, Bisha-wi K, Shanably A. Clinical and laboratory findings of COVID-19: A systematic review and meta-anal-ysis. J Formos Med Assoc 2020; 2020: 1-18.
203) Lee KW, Yusof Khan AHK, Ching SM, Chia PK, Loh WC, Abdul Rashid AM, Baharin J, Inche Mat LN, Wan Sulaiman WA, Devaraj NK, Sivaratnam D, Basri H, Hoo FK. Stroke and Novel Coronavirus Infection in Humans: A Systematic Review and Meta-Analysis. Front Neurol 2020; 11: 579070.
204) Correia AO, Feitosa PWG, Moreira JL de S, Nogueira SÁR, Fonseca RB, Nobre MEP. Neu-rological manifestations of COVID-19 and other coronaviruses: A systematic review. Neurol Psy-chiatry Brain Res 2020; 37: 27-32.
205) Borges do Nascimento IJ, Cacic N, Abdulazeem HM, von Groote TC, Jayarajah U, Weerasekara I, Esfahani MA, Civile VT, Marusic A, Jeroncic A,Carvas Junior N, Pericic TP, Zakarija-Grkov-ic I, Meirelles Guimarães SM, Luigi BragazziN, Bjorklund M, Sofi-Mahmudi A, Altujjar M, Tian M, ArcaniDMC, O’Mathúna DP, Marcolino MS. Novel Coronavirus Infection (COVID-19) in Humans: A Scoping Review and Meta-Analysis. J Clin Med 2020; 9: 941.
206) Hu W, Yen YT, Singh S, Kao CL, Wu-Hsieh BA. SARS-CoV regulates immune function-related gene expression in human monocytic cells. Viral Immunol 2012; 25: 277-288.
207) Connor RF, Roper RL. Unique SARS-CoV protein nsp1: bioinformatics, biochemistry and potential ef-fects on virulence. Trends Microbiol 2007; 15: 51-53.
208) Cheng VCC, Lau SKP, Woo PCY, Yuen KY. Se-vere Acute Respiratory Syndrome Coronavirus as

Comparative review of three human coronaviruses
7183
an Agent of Emerging and Reemerging Infection. Clin Microbiol Rev 2007; 20: 660-694.
209) Chu H, Chan JF, Wang Y, Yuen TT, Chai Y, Hou Y, Shuai H, Yang D, Hu B, Huang X, Zhang X, Cai JP, Zhou J, Yuan S, Kok KH, To KK, Chan IH, Zhang AJ, Sit KY, Au WK, Yuen KY. Compara-tive replication and immune activation profiles of SARS-CoV-2 and SARS-CoV in human lungs: an ex vivo study with implications for the pathogen-esis of COVID-19. Clin Infect Dis 2020; 71: 1400-1409.
210) Siddique F, Abbas RZ, Mansoor MK, Alghamdi ES, Saeed M, Ayaz MM, Rahman M, Mahmood MS, Iqbal A, Manzoor M, Abbas A, JavaidA, HussainI.An Insight Into COVID-19: A 21st Century Disaster and Its Relation to Immunocompetence and Food Antioxidants. Front Vet Sci 2021; 7: 586637.
211) Channappanavar R, Perlman S. Pathogenic hu-man coronavirus infections: causes and conse-quences of cytokine storm and immunopathology. Semin Immunopathol 2017; 39: 529-539.
212) Law HKW, Chung YC, Hoi YN, Sin FS, Yuk OC, Luk W, Nicholls JM, Peiris JSM, Lau YL. Chemo-kine up-regulation in SARS-coronavirus-infected, monocyte-derived human dendritic cells. Blood 2005; 106: 2366-2374.
213) Zheng J, Wang Y, Li K, Meyerholz DK, Allamargot C, Perlman S. Severe Acute Respiratory Syndrome Coronavirus 2-Induced Immune Activation and Death of Monocyte-Derived Human Macrophages and Dendritic Cells. J Infect Dis 2021; 223: 785-795.
214) Cheung CY, Poon LLM, Ng IHY, Luk W, Sia SF, Wu MHS, Chan KH, Yuen KY, Gordon S, Guan Y, Peiris JSM. Cytokine Responses in Severe Acute Respiratory Syndrome Coronavirus-Infect-ed Macrophages In Vitro: Possible Relevance to Pathogenesis. J Virol 2005; 79: 7819-7826.
215) Lee P, Kim DJ. Newly emerging human corona-viruses: Animal models and vaccine research for SARS, MERS, and COVID-19. Immune Netw 2020; 20: 1-25.
216) Tetro JA. Is COVID-19 receiving ADE from other coronaviruses? Microbes Infect 2020; 22: 72-73.
217) Broxmeyer HE, Sherry B, Cooper S, Lu L, Maze R, Beckmann MP, Cerami A, Ralph P. Compar-ative analysis of the human macrophage inflam-matory protein family of cytokines (chemokines) on proliferation of human myeloid progenitor cells. Interacting effects involving suppression, syner-gistic suppression, and blocking of suppression. J Immunol 1993; 150: 3448-3458.
218) Wong CK, Lam CW, Wu AK, Ip WK, Lee NL, Chan IH, Lit LC, Hui DS, Chan MH, Chung SS, Sung JJ. Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin Exp Immunol 2004; 136: 95-103.
219) Herold S, Steinmueller M, Von Wulffen W, Ca-karova L, Pinto R, Pleschka S, Mack M, Kuziel WA, Corazza N, Brunner T, Seeger W, Lohmey-er J. Lung epithelial apoptosis in influenza virus pneumonia: The role of macrophage-expressed TNF-related apoptosis-inducing ligand. J Exp Med 2008; 205: 3065-3077.
220) Boonnak K, Vogel L, Feldmann F, Feldmann H, Legge KL, Subbarao K. Lymphopenia Associated
with Highly Virulent H5N1 Virus Infection Due to Plasmacytoid Dendritic Cell-Mediated Apoptosis of T Cells. J Immunol 2014; 192: 5906-5912.
221) Cameron MJ, Ran L, Xu L, Danesh A, Berme-jo-Martin JF, Cameron CM, Muller MP, Gold WL, Richardson SE, Poutanen SM,Willey BM, DeVries ME, Fang Y, Seneviratne C, Bosinger SE, Persad D, Wilkinson P, Greller LD, Somogyi R, Humar A, Keshavjee S, Louie M, Loeb MB, BruntonJ, Mc-Geer AJ, Kelvin DJ. Interferon-Mediated Immuno-pathological Events Are Associated with Atypical Innate and Adaptive Immune Responses in Pa-tients with Severe Acute Respiratory Syndrome. J Virol 2007; 81: 8692-8706.
222) Croce L, Gangemi D, Ancona G, Liboà F, Bendotti G, Minelli L, Chiovato L. The cytokine storm and thyroid hormone changes in COVID-19. J Endo-crinol Invest 2021; 44: 891-904.
223) Hirawat R, Saifi MA, Godugu C. Targeting inflam-matory cytokine storm to fight against COVID-19 associated severe complications. Life Sci 2021; 267: 118923.
224) Keam S, Megawati D, Patel SK, Tiwari R, Dhama K, Harapan H. Immunopathology and immuno-therapeutic strategies in severe acute respiratory syndrome coronavirus 2 infection. Rev Med Virol 2020; 30: e2123.
225) Yongzhi X. COVID-19-associated cytokine storm syndrome and diagnostic principles: an old and new Issue. Emerg Microbes Infect 2021; 10: 266-276.
226) Cao X. COVID-19: immunopathology and its im-plications for therapy. Nat Rev Immunol 2020; 20: 269-270.
227) Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z, Jia X, Wu M, Shi B, Xu S, Chen J, Wang W, Chen B, Jiang L, Yu S, Lu J, Wang J, Xu M, Yuan Z, Zhang Q, Zhang X, Zhao G, Chen S, Lu H. Viral and host factors related to the clinical outcome of COVID-19. Nature 2020; 583: 437-440.
228) Ruytinx P, Proost P, Van Damme J, Struyf S. Chemo-kine-induced macrophage polarization in inflamma-tory conditions. Front Immunol 2018; 9: 1930.
229) Menezes SM, Braz M, Llorens-Rico V, Wauters J, Van Weyenbergh J. Endogenous IFNβ expression predicts outcome in critical patients with COVID-19. The Lancet Microbe 2021; 2: e235-e236.
230) Ari S, Can V, Demir ÖF, Ari H, Ağca FV, Melek M, Çamci S, Dikiş ÖŞ, Huysal K, Türk T. Elevated eo-sinophil count is related with lower anti-factor Xa activity in COVID-19 patients. J Hematop 2020; 13: 249-258.
231) Liu S, Yuan H, Zhang B, Li W, You J, Liu J, Zhong Q, Zhang L, Chen L, Li S, ZouY, ZhangS. Com-parison of Clinical Features and CT Temporal Changes Between Familial Clusters and Non-fa-milial Patients With COVID-19 Pneumonia. Front Med 2021; 8: 1-9.
232) Surendar J, Frohberger SJ, Karunakaran I, Schmitt V, Stamminger W, Neumann AL, Wilhelm C, Hoerauf A, Hübner MP. Adiponectin limits IFN-γand IL-17 producing CD4 T cells in obesity by restraining cell intrinsic glycolysis. Front Immunol 2019; 10: 2555.
233) Laires PA, Dias S, Gama A, Moniz M, Pedro AR, Soares P, Aguiar P, Nunes C. The association

A.A. Rabaan, A.A. Mutair, Z.A. Alawi, S. Alhumaid, M.A. Mohaini, J. Aldali, R. Tirupathi, et al
7184
between chronic disease and serious COVID-19 outcomes and its influence on risk perception: Survey study and database analysis. JMIR Public Heal Surveill 2021; 7: e22794.
234) Bae SA, Kim SR, Kim MN, Shim WJ, Park SM. Im-pact of cardiovascular disease and risk factors on fatal outcomes in patients with COVID-19 accord-ing to age: A systematic review and meta-analy-sis. Heart 2021; 107: 373-380.
235) Zinellu A, Arru F, De Vito A, Sassu A, Valdes G, Scano V, Zinellu E, Perra R, Madeddu G, Carru C,PirinaP, MangoniAA, BabudieriS, FoisAG.The De Ritis ratio as prognostic biomarker of in-hos-pital mortality in COVID-19 patients. Eur J Clin Invest 2021; 51: e13427.
236) McCarthy C, O’Donnell CP, Kelly NEW, O’Shea D, Hogan AE. COVID-19 severity and obesity: are MAIT cells a factor? Lancet Respir Med 2021; 9: 445-447.
237) O’Shea D, Hogan AE. Dysregulation of natural kill-er cells in obesity. Cancers (Basel) 2019; 11: 573.
238) Brien AO, Kedia-Mehta N, Tobin L, Veerapen N, Besra GS, Shea DO, Hogan AE. Targeting mi-tochondrial dysfunction in MAIT cells limits IL-17 production in obesity. Cell Mol Immunol 2020; 17: 1193-1195.
239) Popkin BM, Du S, Green WD, Beck MA, Algaith T, Herbst CH, Alsukait RF, Alluhidan M, Ala-zemi N, Shekar M. Individuals with obesity and
COVID-19: A global perspective on the epide-miology and biological relationships. Obes Rev 2020; 21: e13128.
240) Gao M, Piernas C, Astbury NM, Hippisley-Cox J, O’Rahilly S, Aveyard P, Jebb SA. Associations between body-mass index and COVID-19 severi-ty in 6·9 million people in England: a prospective, community-based, cohort study. Lancet Diabetes Endocrinol 2021; 9: 350-359.
241) Lega I, Nisticò L, Palmieri L, Caroppo E, Noce CL, Donfrancesco C, Vanacore N, Scattoni ML, Picar-di A, Gigantesco A, Brusaferro S. Psychiatric disorders among hospitalized patients deceased with COVID-19 in Italy. EClinicalMedicine 2021; 35: 100854.
242) Kadiwar S, Smith JJ, Ledot S, Johnson M, Bianchi P, Singh N, Montanaro C, Gatzoulis M, Shah N, Ukor EF. Were pregnant women more affected by COVID-19 in the second wave of the pandemic? Lancet 2021; 397: 1539-1540.
243) Steptoe A, Di Gessa G. Mental health and social interactions of older people with physical disabil-ities in England during the COVID-19 pandemic : a longitudinal cohort study. Lancet Public Heal 2021; 6: e365-e373.
244) Kuper H, Shakespeare T. Comment Are old-er people with disabilities neglected in the COVID-19 pandemic ? Lancet Public Heal 2021; 6: e347-e348.
Related Documents











