Comparative in vitro and early nursery performance of adventitious shoots from cryopreserved cotyledons and axillary shoots from epicotyls of the same zygotic embryo of control-pollinated Pinus radiata Cathy L. Hargreaves, Lynette J. Grace, Susan A. van der Maas, Mike I. Menzies, Satish Kumar, D. Grant Holden, Martin N. Foggo, Charlie B. Low, and Mike J. Dibley Abstract: This is the first published report comparing production and performance of adventitious shoots from cryopreserved cotyledons, with axillary shoots formed from epicotyls of the same zygotic embryo of radiata pine (Pinus radiata D. Don). Genotypes from 10 control-pollinated families of P. radiata in two treatments were compared for shoot initiation, in vitro growth, rooting, and early nursery performance. Plant growth in nursery beds was assessed by measuring height after 2 and 7 months. After 8 months in nursery beds, the physiological ages of genotypes were assessed before field planting. Genotype capture was higher from the cryopreserved cotyledons than from the epicotyls. This technique has the advantage of preserving juvenile material while field testing is done. Early shoot multiplication in both treatments was good. After four transfer cycles, epicotyl cultures showed improved elongation and higher mul- tiplication rates. After 6 months of in vitro growth, shoots from both treatments were given auxin pulses. Shoots of ad- ventitious origin were slower to root than epicotyl-derived shoots. Overall rooting rates were satisfactory. Plants of adventitious origin were shorter when planted into nursery beds and when transferred to field trials. Assessment of rel- ative physiological age indicated that all plants of adventitious origin showed some increase. Résumé : Ceci est le premier article à être publié sur la comparaison entre la production et la performance des pous- ses adventives obtenues à partir de cotylédons cryoconservés et celles des pousses axillaires formées à partir des épico- tyles du même embryon zygotique de pin de Monterey (Pinus radiata D. Don). Les génotypes de 10 familles de pin de Monterey obtenus par pollinisation dirigée ont été comparés dans deux traitements du point de vue de l’initiation des pousses, de la croissance in vitro, de l’enracinement et de la performance précoce en pépinière. La croissance des jeu- nes plants dans les planches a été évaluée par des mesures de la hauteur après 2 et 7 mois. Après 8 mois en pépinière, l’âge physiologique des génotypes a été évalué avant la plantation sur le terrain. La reproduction des génotypes était plus élevée à partir des cotylédons cryoconservés qu’à partir des épicotyles. Cette technique avait l’avantage de préser- ver le matériel juvénile pendant qu’ont lieu les tests sur le terrain. La multiplication précoce des pousses était bonne avec les deux traitements. Après quatre cycles de transfert, les cultures obtenues à partir d’épicotyles avaient une meil- leure élongation et un plus haut taux de multiplication. Après 6 mois de croissance in vitro, les pousses des deux trai- tements ont été stimulées avec des auxines. Les pousses d’origine adventive étaient plus lentes à s’enraciner que les pousses dérivées des épicotyles. Dans l’ensemble, le taux d’enracinement était satisfaisant. Les plants d’origine adven- tive étaient plus courts au moment d’être plantés en pépinière et au moment d’être transférés pour les essais sur le ter- rain. L’évaluation de l’âge physiologique relatif a indiqué que l’âge de tous les plants d’origine adventive avait connu une certaine augmentation. [Traduit par la Rédaction] Hargreaves et al. 2641 Can. J. For. Res. 35: 2629–2641 (2005) doi: 10.1139/X05-178 © 2005 NRC Canada 2629 Received 15 March 2005. Accepted 20 July 2005. Published on the NRC Research Press Web site at http://cjfr.nrc.ca on 11 November 2005. C.L. Hargreaves, 1 L.J. Grace, S.A. van der Maas, M.I. Menzies, S. Kumar, D.G. Holden, C.B. Low, and M.J. Dibley. New Zealand Forest Research Institute, Ensis, 49 Sala Street, Private Bag 3020, Rotorua, New Zealand. M.N. Foggo. 67 Tihi Road, Springfield, Rotorua, New Zealand. 1 Corresponding author (e-mail: [email protected]).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Comparative in vitro and early nursery performanceof adventitious shoots from cryopreservedcotyledons and axillary shoots from epicotyls ofthe same zygotic embryo of control-pollinatedPinus radiata
Cathy L. Hargreaves, Lynette J. Grace, Susan A. van der Maas, Mike I. Menzies,Satish Kumar, D. Grant Holden, Martin N. Foggo, Charlie B. Low, andMike J. Dibley
Abstract: This is the first published report comparing production and performance of adventitious shoots fromcryopreserved cotyledons, with axillary shoots formed from epicotyls of the same zygotic embryo of radiata pine(Pinus radiata D. Don). Genotypes from 10 control-pollinated families of P. radiata in two treatments were comparedfor shoot initiation, in vitro growth, rooting, and early nursery performance. Plant growth in nursery beds was assessedby measuring height after 2 and 7 months. After 8 months in nursery beds, the physiological ages of genotypes wereassessed before field planting. Genotype capture was higher from the cryopreserved cotyledons than from the epicotyls.This technique has the advantage of preserving juvenile material while field testing is done. Early shoot multiplicationin both treatments was good. After four transfer cycles, epicotyl cultures showed improved elongation and higher mul-tiplication rates. After 6 months of in vitro growth, shoots from both treatments were given auxin pulses. Shoots of ad-ventitious origin were slower to root than epicotyl-derived shoots. Overall rooting rates were satisfactory. Plants ofadventitious origin were shorter when planted into nursery beds and when transferred to field trials. Assessment of rel-ative physiological age indicated that all plants of adventitious origin showed some increase.
Résumé : Ceci est le premier article à être publié sur la comparaison entre la production et la performance des pous-ses adventives obtenues à partir de cotylédons cryoconservés et celles des pousses axillaires formées à partir des épico-tyles du même embryon zygotique de pin de Monterey (Pinus radiata D. Don). Les génotypes de 10 familles de pin deMonterey obtenus par pollinisation dirigée ont été comparés dans deux traitements du point de vue de l’initiation despousses, de la croissance in vitro, de l’enracinement et de la performance précoce en pépinière. La croissance des jeu-nes plants dans les planches a été évaluée par des mesures de la hauteur après 2 et 7 mois. Après 8 mois en pépinière,l’âge physiologique des génotypes a été évalué avant la plantation sur le terrain. La reproduction des génotypes étaitplus élevée à partir des cotylédons cryoconservés qu’à partir des épicotyles. Cette technique avait l’avantage de préser-ver le matériel juvénile pendant qu’ont lieu les tests sur le terrain. La multiplication précoce des pousses était bonneavec les deux traitements. Après quatre cycles de transfert, les cultures obtenues à partir d’épicotyles avaient une meil-leure élongation et un plus haut taux de multiplication. Après 6 mois de croissance in vitro, les pousses des deux trai-tements ont été stimulées avec des auxines. Les pousses d’origine adventive étaient plus lentes à s’enraciner que lespousses dérivées des épicotyles. Dans l’ensemble, le taux d’enracinement était satisfaisant. Les plants d’origine adven-tive étaient plus courts au moment d’être plantés en pépinière et au moment d’être transférés pour les essais sur le ter-rain. L’évaluation de l’âge physiologique relatif a indiqué que l’âge de tous les plants d’origine adventive avait connuune certaine augmentation.
[Traduit par la Rédaction] Hargreaves et al. 2641
Can. J. For. Res. 35: 2629–2641 (2005) doi: 10.1139/X05-178 © 2005 NRC Canada
2629
Received 15 March 2005. Accepted 20 July 2005. Published on the NRC Research Press Web site at http://cjfr.nrc.ca on11 November 2005.
C.L. Hargreaves,1 L.J. Grace, S.A. van der Maas, M.I. Menzies, S. Kumar, D.G. Holden, C.B. Low, and M.J. Dibley.New Zealand Forest Research Institute, Ensis, 49 Sala Street, Private Bag 3020, Rotorua, New Zealand.M.N. Foggo. 67 Tihi Road, Springfield, Rotorua, New Zealand.
1Corresponding author (e-mail: [email protected]).

Introduction
A critical requirement for clonal forestry using radiatapine, Pinus radiata D. Don, is a cost-effective propagationsystem that is successful for most genotypes and gives largenumbers of uniform plants per clone that show minimalphysiological aging in field testing (Menzies and Aimers-Halliday 1997). Several clonal-propagation approaches havebeen tested on P. radiata. These can be summarized, in orderof increasing cost, as nursery cuttings; tissue-culture ap-proaches that include axillary- and adventitious-shoot pro-duction; and somatic embryogenesis. Methods using cuttingsinvolve the manipulation of seedlings in stool beds/hedges tostimulate axillary-shoot formation from fascicles. Multipli-cation rates as high as 50–80 times per seedling within20 months of sowing have been reported (Faulds and Dibley1991). Cuttings from these hedges can in turn be set to pro-duce further stool beds so that over a 5-year period a largenumber of cuttings can be produced from a single genotype.In tissue culture, axillary shoot multiplication methods areinitiated by the germination of control-pollinated seed in vi-tro. The isolates can then be multiplied rapidly by tissue cul-ture and this results in the generation of plant numbers inexcess of several thousand after 1 year in vitro (C.L.Hargreaves, unpublished data). Adventitious-shoot methodsoften use pregermination cotyledons from zygotic embryosthat are given a cytokinin (benzylaminopurine) treatment toproduce shoots. These are then used for axillary-shoot multi-plication and can produce even higher plant numbers fromsingle genotypes in 1 year (Aitken-Christie and Jones 1987).Either of these two in vitro methods can be combined withtechniques using cuttings to provide large numbers of stoolbeds, thus reducing the time and costs associated with in vi-tro methods and further increasing the plant numbers avail-able from single genotypes.
The most serious limitation to these three methodologiesis that the source material has a limited life-span, owing tothe constraints of physiological aging or maturation. By thetime clonal material has been proven in field testing for 6–8 years or more, the stool beds/shoot cultures are no longerproducing juvenile material. Some arrest of this physiologi-cal aging has been achieved by both the hedging process ofstool-bed production (Menzies et al. 1985; Menzies andAimers-Halliday 1997) and the cool storage of in vitrogrown shoots (Aitken-Christie and Singh 1987; Horgan et al.1997). Cryogenic methods of storage that arrest aging areunder development for axillary-shoot meristems. However,this method is currently limited by low genotype capture af-ter freezing and subsequent slow growth of recovered meri-stems (C.L. Hargreaves, unpublished data).
Somatic embryogenesis was originally seen as the futureway of conifer clonal forestry for commercial-forest opera-tors because the technique results in tissue that can be easilycryopreserved (thus ensuring juvenility while field testingtakes place) and has the potential to produce millions ofplants quickly (Durzan and Gupta 1988; Attree and Fowke1991). However, a major drawback to the technology is lowgenotype capture and the large labour cost of in vitro proce-dures before genotypes are at the stage where less technicalmethods of amplification can be used (Timmis 1998; Park etal. 1998).
An alternative method that may offer some of the advan-tages of somatic embryogenesis without the drawbacks is theadventitious production of shoots from cryopreserved em-bryo cotyledons (Hargreaves et al. 2004). The cryopreserva-tion of cotyledons, from which a subset has been removed toproduce plants for initial field trials, ensures that the un-used/stored cotyledons retain juvenile growth characteristics.Earlier results with P. radiata showed that cryopreservationof cotyledons did not reduce shoot production, root forma-tion on shoots, or plant growth in the first 2 years in the fieldcompared with non-cryopreserved controls (Hargreaves et al.2004). Genotype capture from mature zygotic embryos (andtherefore the dominant embryo) is high and the in vitro cul-ture period short. Media formulations include few plant-growth regulators, and less skilled observation through de-velopmental stages is required than with somatic embryo-genesis.
However, the use of adventitious-bud formation has otherconsiderations. Although genotype capture is high, there isconcern that shoots produced adventitiously from cotyledonsmay perform differently from shoots that arise from theepicotyl without the use of any growth regulators. Researchwith P. radiata (K. Horgan, A. Joe, D. Skudder, W. Hackett,and A. Clausen, unpublished data) and loblolly pine, Pinustaeda L. (Frampton et al. 1998), has indicated slower initialgrowth rates in the field and early maturation in propagulesof adventitious origin of some genotypes. However, even ifthe performance of some genotypes is adversely affected bytheir adventitious origin, many more genotypes would reachthe field from breeding programs than from somatic-embryogensis systems. There may still be sufficient geno-types that do perform well in the field to enable the methodto be used for clonal forestry, and probably at significantlyless cost than if somatic embryogenesis was employed.
In the present investigation, the performance of adventi-tious shoots derived from cryopreserved cotyledons wascompared with that of axillary shoots derived from the samegenotypes, thus testing the applicability of the two propaga-tion methodologies for clonal forestry. Genotype capture,shoot production, root initiation, physiological aging, andearly nursery performance were recorded.
Materials and methods
Control-pollinated seed from mature cones of 10 crossesof P. radiata collected in 2000 from the Amberley seed or-chard (North Canterbury, South Island, New Zealand) wereused. The seed originated from second- or third-generationstands with 80% probability of their being Ana Nuevo and20% probability of being Monterey. The families selectedwere the result of controlled pollinations between six treesused as females and seven different trees used as males, withseveral trees being used more than once (as the parent of thesame sex). Seeds were sterilized in November 2002 (South-ern Hemisphere summer) as described by Hargreaves et al.(2004).
Embryos (20 from each of the 10 families) were dissectedand left for 3 h on a modified Quoirin and Lepoivre medium(LP) (Quoirin and Lepoivre 1977, modified by Aitken-Christie et al. 1988) until the cotyledons had opened a little.Any yellowish or malformed embryos were deemed unsound
© 2005 NRC Canada
2630 Can. J. For. Res. Vol. 35, 2005
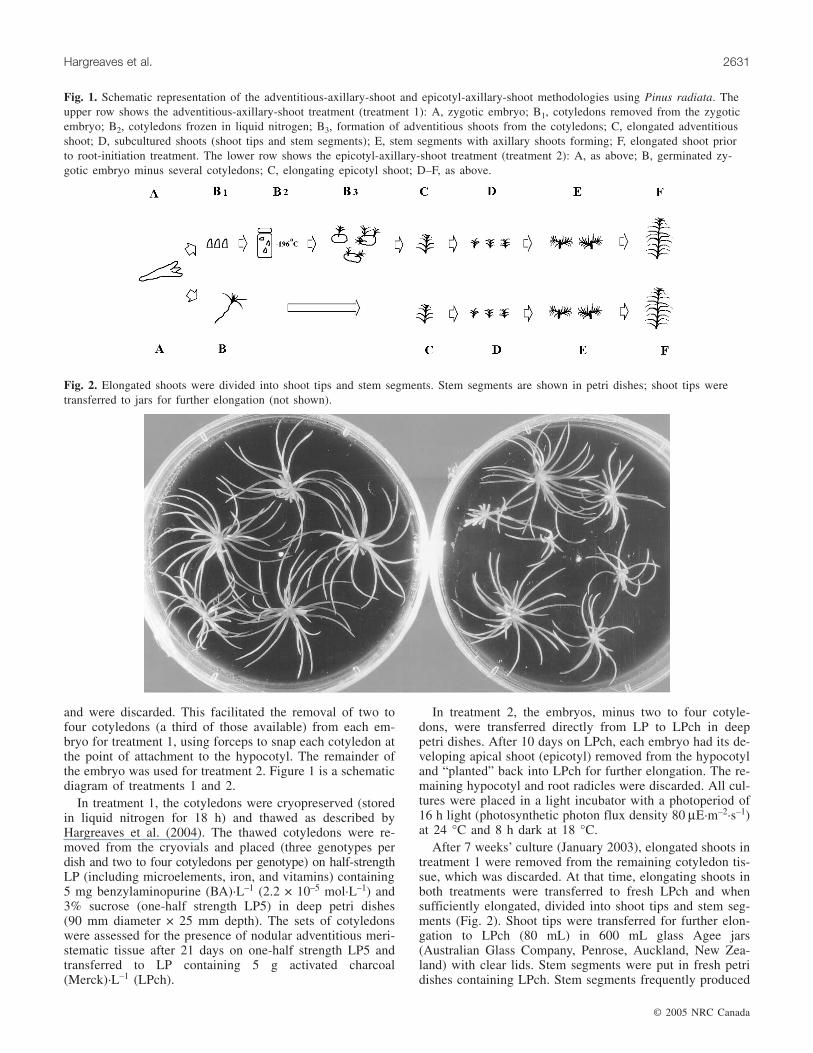
and were discarded. This facilitated the removal of two tofour cotyledons (a third of those available) from each em-bryo for treatment 1, using forceps to snap each cotyledon atthe point of attachment to the hypocotyl. The remainder ofthe embryo was used for treatment 2. Figure 1 is a schematicdiagram of treatments 1 and 2.
In treatment 1, the cotyledons were cryopreserved (storedin liquid nitrogen for 18 h) and thawed as described byHargreaves et al. (2004). The thawed cotyledons were re-moved from the cryovials and placed (three genotypes perdish and two to four cotyledons per genotype) on half-strengthLP (including microelements, iron, and vitamins) containing5 mg benzylaminopurine (BA)·L–1 (2.2 × 10–5 mol·L–1) and3% sucrose (one-half strength LP5) in deep petri dishes(90 mm diameter × 25 mm depth). The sets of cotyledonswere assessed for the presence of nodular adventitious meri-stematic tissue after 21 days on one-half strength LP5 andtransferred to LP containing 5 g activated charcoal(Merck)·L–1 (LPch).
In treatment 2, the embryos, minus two to four cotyle-dons, were transferred directly from LP to LPch in deeppetri dishes. After 10 days on LPch, each embryo had its de-veloping apical shoot (epicotyl) removed from the hypocotyland “planted” back into LPch for further elongation. The re-maining hypocotyl and root radicles were discarded. All cul-tures were placed in a light incubator with a photoperiod of16 h light (photosynthetic photon flux density 80 µE·m–2·s–1)at 24 °C and 8 h dark at 18 °C.
After 7 weeks’ culture (January 2003), elongated shoots intreatment 1 were removed from the remaining cotyledon tis-sue, which was discarded. At that time, elongating shoots inboth treatments were transferred to fresh LPch and whensufficiently elongated, divided into shoot tips and stem seg-ments (Fig. 2). Shoot tips were transferred for further elon-gation to LPch (80 mL) in 600 mL glass Agee jars(Australian Glass Company, Penrose, Auckland, New Zea-land) with clear lids. Stem segments were put in fresh petridishes containing LPch. Stem segments frequently produced
© 2005 NRC Canada
Hargreaves et al. 2631
Fig. 1. Schematic representation of the adventitious-axillary-shoot and epicotyl-axillary-shoot methodologies using Pinus radiata. Theupper row shows the adventitious-axillary-shoot treatment (treatment 1): A, zygotic embryo; B1, cotyledons removed from the zygoticembryo; B2, cotyledons frozen in liquid nitrogen; B3, formation of adventitious shoots from the cotyledons; C, elongated adventitiousshoot; D, subcultured shoots (shoot tips and stem segments); E, stem segments with axillary shoots forming; F, elongated shoot priorto root-initiation treatment. The lower row shows the epicotyl-axillary-shoot treatment (treatment 2): A, as above; B, germinated zy-gotic embryo minus several cotyledons; C, elongating epicotyl shoot; D–F, as above.
Fig. 2. Elongated shoots were divided into shoot tips and stem segments. Stem segments are shown in petri dishes; shoot tips weretransferred to jars for further elongation (not shown).
side shoots that further enhanced multiplication (Fig. 3). Forclarity, shoots that arose initially from cryopreserved cotyle-dons are described as adventitious-axillary, and shoots de-rived from the developing epicotyl are termed epicotyl-axillary.
After 10 weeks’ culture (late January), genotypes in bothtreatments that had one or more vigorous explants, and usu-ally shoot tips and stem segments, were selected. Shoottransfers were continued in both treatments at 4- to 6-weekintervals for a further 18 weeks. From initiation through allsubsequent subculturing stages, any shoots that appeared“wet” or hyperhydric, indicating reduced cuticular wax,were discarded (Aitken-Christie et al. 1985; Debergh et al.1992). At approximately 28 weeks (May 2003) from thestart of the experiment, a total of 54 genotypes from 9 of the10 control pollinations with a minimum of 25 shoot tips pertreatment per genotype were selected. These shoot tips hadelongated to approximately 3 cm on LPch in 600 mL Ageejars. After 6 weeks on LPch, shoots were given a furthertransfer to LPch. This facilitated shoot setting in late Julyand thus planting out in the nursery beds on the preferreddate.
At 34 weeks from the beginning of the experiment (July2003), all shoots in both treatments were given a fresh basecut to remove any callus and an in vitro auxin treatment for12–14 days on a Greshoff and Doy medium (as modified bySommer et al. 1975 and Horgan and Holland 1989) contain-ing 2% sucrose, 0.5 mg napthalene acetic acid·L–1, and1.0 mg indole butyric acid·L–1. Following auxin treatment,shoots were individually transferred (set) into trays ofpeat:perlite:sand (1:1:1) on 2–4 August 2003. High humiditywas maintained for 1 week with a combination of ventedplastic lids and hand-misting twice daily. Set shoots wereheld under light and temperature conditions similar to thosedescribed earlier for in vitro growth. After 1 week, shootswere gradually hardened off to ambient humidity conditions(60% ± 10%). Four weeks after setting, shoots were assessed
for early root formation (August 2003) and potted into HikoV90 (plastic trays) with the same potting mix as used forsetting, with the addition of 5 g Osmocote·L–1 (Fig. 4). Theplants were transferred to the greenhouse (September 2003)and after 8 weeks placed in a nursery shade house (late Oc-tober 2003) until planted out in the nursery bed (15 cm ×15 cm spacing) in early December 2003.
Growth was assessed by measuring plant height in Febru-ary and July 2004 (Fig. 5). The physiological age of plantswas assessed prior to field planting in August 2004, basedon characteristics of the apical bud and primary and second-ary needles (Menzies et al. 2000).
The data were analysed using PROC GLM (SAS InstituteInc. 1989) for analysis of variance (ANOVA) using themodel y = x + family + genotypes (families) + treatment +treatment × families + error. For all analysis models, fami-lies, genotypes within families, treatment, and interactionterms were considered random effects. The model treated alleffects as random, as this is the most conservative test,which should not restrict the significance of effects to justthose treatments or families used in this experiment. Tukey’smultiple range test option was used to calculate significantdifferences between family and treatment means.
Results
There were small differences in the numbers of sound em-bryos among the 10 crosses selected for this experiment (Ta-ble 1), with cross 9 having the lowest (13), cross 10 havingthe highest (20), and an average of 17 per cross (Table 1).Axillary-shoot formation from the epicotyl was more vari-able than adventitious-shoot formation, with a range of30%–94% compared with 50%–100% across the 10 fami-lies, with some of the differences significant (Tables 1 and2). There was a trend towards higher capture of genotypeswith the adventitious-shoot treatment (80%) than with theaxillary-shoot treatment (59%). For the purposes of this ex-
© 2005 NRC Canada
2632 Can. J. For. Res. Vol. 35, 2005
Fig. 3. The stem segments shown in Fig. 2 after 4 weeks’ growth. Side shoots have formed at the needle stem axils from axillarymeristems present in both treatment 1 and treatment 2.

periment, it was important to have viable shoots in bothtreatments for subsequent comparative work. In most cases,if a genotype had produced viable shoots in the axillary-shoot treatment, it would also have shoots represented in theadventitious-shoot treatment (Table 1). However, not all ofthe genotypes that formed adventitious shoots had also pro-duced epicotyl-axillary shoots.
Shoot-multiplication rates were deduced by dividing thesubsequent total number of explants (February 2003, April
2003, May 2003) by the total number of explants in January2003 (Table 3). The number of adventitious shoots (treat-ment 1) was higher initially because more meristems (formedadventitiously over the surface of the cotyledons) wereavailable than the initial apical meristem of the epicotyl.This is illustrated by the ranges in January 2003: 3.6–8.00for treatment 1 compared with 1.89–2.67 for treatment 2(Table 3). Subsequent amplification rates differed betweenthe two treatments over the next three transfers. In February
© 2005 NRC Canada
Hargreaves et al. 2633
Fig. 4. (a) Family 1, genotype 13 plants in treatment 1 (adventitious-axillary-shoot treatment; left-hand side) and treatment 2 (epicotyl-axillary-shoot treatment; right-hand side) in the greenhouse 8 weeks after planting out (September 2003). (b) Family 1, genotype 15 intreatment 1 (adventitious-axillary-shoot treatment; left-hand side) and treatment 2 (epicotyl-axillary-shoot treatment; right-hand side) inthe greenhouse 8 weeks after planting out (September 2003).

2003 there was little difference in the mean number ofexplants per treatment (1.71 in treatment 1 and 1.70 in treat-ment 2). In April 2003 the mean number of explants pershoot had increased to 4.95 in treatment 1 and 6.19 in treat-ment 2, although the difference was not significant (Tables 2and 3). In May 2003 the difference was also significant (P <0.001), with means of 8.70 explants per shoot in treatment 1and 14.77 in treatment 2 (Tables 2 and 3). The shoots intreatment 1 did not elongate as quickly as those in treatment
2. The phenotypes of shoots in treatments 1 and 2, growingunder identical conditions, could not be distinguished by eye.
Following the shoot transfers in May 2003, a maximum ofseven genotypes per family that had a minimum of 25 shoottips were randomly selected in both treatments. This reducedthe total number of genotypes from 74 (Table 3) to 54 (Ta-ble 4). The total number of shoots transferred to Greshoffand Doy auxin medium per family that formed roots after4 weeks is given in Table 4. There were significant differ-
© 2005 NRC Canada
2634 Can. J. For. Res. Vol. 35, 2005
Fig. 5. (a) Family 2, genotype 8 after 7 months in the nursery bed (July 2004) plants in treatment 2 (epicotyl-axillary treatment; left-hand side) and treatment 1 (adventitious-axillary treatment; right-hand side) after 7 months in the nursery bed (July 2004). (b) Family3, genotype 9 plants in treatment 2 (epicotyl-axillary treatment; left-hand side) and treatment 1 (adventitious-axillary treatment; right-hand side) after 7 months in the nursery bed (July 2004).

ences in early rooting between families in both treatments,ranging from 49% to 100% in treatment 1 and from 49% to99% in treatment 2 (Table 4). Early-rooting results in treat-ment 2 were better than those in treatment 1 for seven of thenine families tested. At transfer into rooting medium, shootsappeared similar and care was taken to use equally elon-gated shoot tips ex vitro. But it was not always possible tomatch family pairs shoot for shoot.
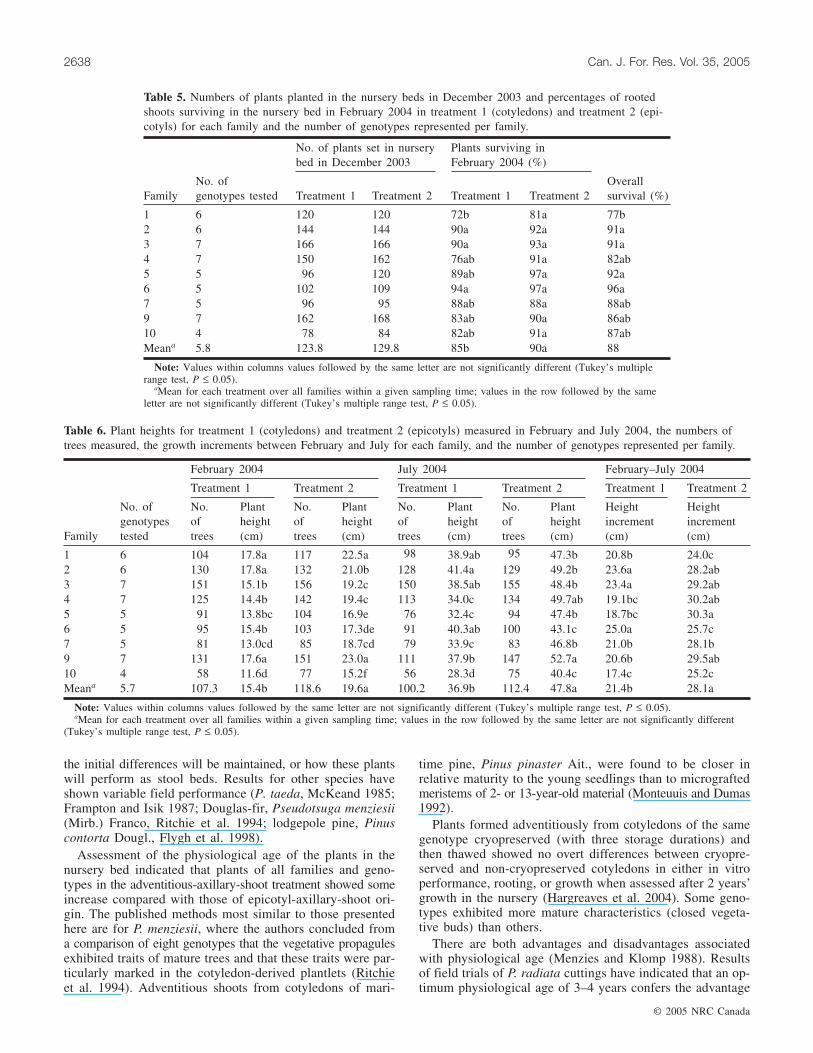
After growing on in the greenhouse and subsequent hard-ening off in shade-house conditions, rooted plants were plantedout into the nursery bed (December 2003). Two genotypeslisted in Table 4 data were not included in the nursery trialsbecause of the aberrant phenotype of family 2, genotype 17in both treatments and poor rooting of family 5, genotype 17in both treatments. In February 2004, plants were assessedfor survival (Table 5). In treatment 2, eight of nine familiesshowed better plant survival. There were between-family dif-ferences ranging from 67% to 90% survival in treatment-1plants compared with 78%–95% in treatment-2 plants (Ta-ble 5) and these differences were significant between fami-lies in treatment 1 (Table 2).
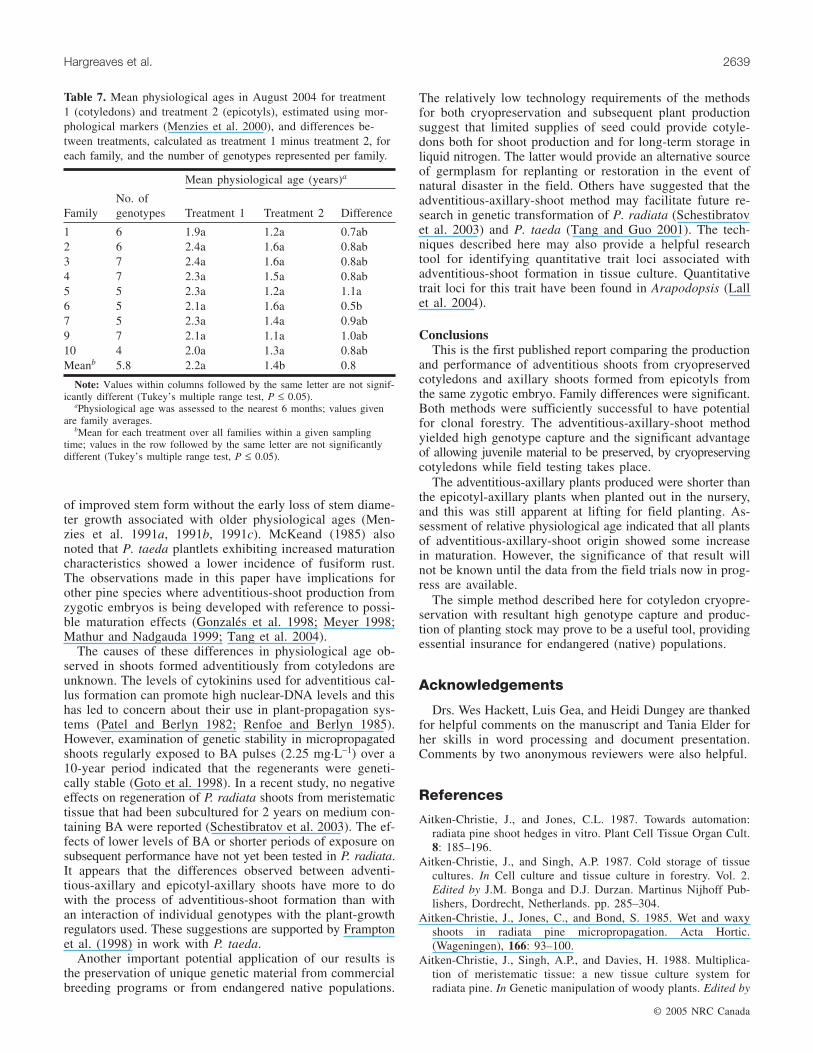
Tree heights (Fig. 5) were measured in the nursery in Feb-ruary and July 2004. The heights and increments measuredbetween these dates are given in Table 6. In February, treesin treatment 1 (mean 15.4 cm) were significantly smaller(P < 0.001; Table 2) than those in treatment 2 (mean 19.6 cm)and there were significant family effects (P < 0.001; Ta-ble 2). In July the treatment means were also significantlydifferent (P < 0.001; Table 2) but there was no family effect.The treatment difference in height increment between thetwo dates was significant (P < 0.001; Table 2), with meansof 21.4 cm for treatment 1 and 28.1 cm for treatment 2, butthere was no family effect (Table 2). When percent heightdifference between the two treatments was calculated, treat-
ment-1 plants were 78% of the height of treatment-2 plantsin February and 77% in July. The difference did not there-fore appear to be increasing.
Prior to field planting, an assessment of physiological agewas made in August 2004 (Table 7). If “day 1” is consideredto be the start of in vitro culture from the isolated cryo-preserved cotyledons in treatment 1 and zygotic embryos intreatment 2, the chronological age of the plants could beconveniently considered to date from in December 2002.Thus, in August 2004, the age of the plants was about20 months (1.7 years). Based on their morphological fea-tures, the adventitious plants (treatment 1) appeared to beolder (at 2.2 years) than their chronological age and theaxillary plants (treatment 2) younger (at 1.4 years) (Table 7).This difference in age was significant (P < 0.001; Table 2),but there was no significant difference between families (Ta-ble 2).
Discussion
This is the first report of research comparing the productionand performance of adventitious shoots from cryopreservedcotyledons with the production and performance of axillaryshoots formed from epicotyls of the same zygotic embryo,and hence of the same genotype, of P. radiata. The experi-ments tested seed from 10 control-pollinated families and atotal of 170 genotypes. All results showed significant familyand interactive effects (Table 2). These were specific for theparticular commercial genotypes used. In any application ofthese techniques to clonal forestry using other seed, familydifferences will need to be assessed. Both methods of shootand plant production demonstrated here have been used incommercial forestry (Nairn 1992).
© 2005 NRC Canada
Hargreaves et al. 2635
Genotypes producing viableshoots (%)
FamilyNo. of seeds witha sound embryoa Treatment 1 Treatment 2
Genotypes representedin both treatments (%)
1 16 56b 44bc 442 18 83ab 67abc 613 16 100a 94a 944 18 100a 67abc 675 17 88ab 71abc 716 18 56b 67abc 447 18 89ab 89a 788 16 75ab 31c 259 13 100a 69abc 6910 20 50b 30c 25Meanb 17 80a 59b 58
Note: Within columns, values followed by the same letter are not significantly different (Tukey’smultiple range test, P ≤ 0.05).
aYellowish or malformed embryos were deemed unsound and were discarded.bMean for each treatment over all families within a given sampling time; values in the row followed
by the same letter are not significantly different (Tukey’s multiple range test, P ≤ 0.05).
Table 1. Numbers of Pinus radiata seeds with a sound embryo (from an initial samplesize of 20), percentages of genotypes producing viable shoots from cotyledons (treatment1) and epicotyls (treatment 2) 5 weeks after initiation, and percentages of genotypes repre-sented in both treatments, for 10 families.
In similar work with P. taeda, the adventitious shoots usedwere not of the same genetic origin as the seedlings (McKeand1985; Frampton and Isik 1987), rooted hypocotyls, or excisedembryos (Frampton and Isik 1987) used for comparison.
Genotype capture following initiation of adventitious shootsfrom cryopreserved cotyledons of control-pollinated seed re-ported here was comparable to that in earlier work usingopen-pollinated seed (Hargreaves et al. 1997, 1999, 2004).Viable shoots were reported for 79%–83% of 72 genotypestested by Hargreaves et al. (2004), which included a non-cryopreserved cotyledon control. In the work presented here,82% of 170 genotypes from 10 different control-pollinatedfamilies formed viable shoots.
Results from other studies comparing the production ofadventitious-axillary shoots with the epicotyl-axillary methodusing control-pollinated families of P. radiata appeared toshow much poorer genotype capture per family overall, butespecially for adventitious-axillary-shoot methods, where therewas an overall mean of 6% of genotypes for all familiestested (Nairn 1992). In the results presented here, based onviable shoot numbers, 80% of tested genotypes from the 10families did form shoots. In our experiment we used 74(44%) paired genotypes of the 170 that were screened and
all of these had in excess of 48 shoots after only 4 months inculture.
Using epicotyl-axillary-shoot methods, Nairn (1992) re-ported an overall mean of 28% of genotypes producing morethan 48 shoots after 12 months for 16 families tested, and63% for 1 family. Our results showed that an overall averageof 59% of genotypes formed viable shoots using the epicotyl-axillary-shoot methods, with 7 of the 10 families having inexcess of 63% genotype capture. Shoot numbers from theepicotyl-axillary-shoot method had not exceeded 48 after4 months, although after 12 months most would have at least2–3 times this number of shoots. The lower genotype cap-ture reported by Nairn (1992) may have been a result of vit-rification (or shoots with poor cuticular-wax formation), citedas a problem in both adventitious-axillary-shoot and epicotyl-axillary-shoot propagation methods (Nairn 1992; Nairn et al.1995). In contrast, vitrification did not influence our currentresults, or previous results reported by Hargreaves et al.(1997, 1999, 2004), because vitrified or wet shoots were re-jected at subculturing (Aitken-Christie et al. 1985; Deberghet al. 1992).
Early-rooting percentages 4 weeks after setting were high forboth treatments, with an average of 74% for the adventitious-
© 2005 NRC Canada
2636 Can. J. For. Res. Vol. 35, 2005
Trait Source Mean square df F P
Viable shoot formation Family 1.25 9 5.56 <0.01Treatment 2.42 1 10.80 <0.01Family × treatment 0.23 9 1.31 ns
Shoots formed by January 2003 Family 6.39 8 0.83 nsTreatment 367.15 1 48.81 <0.001Family × treatment 7.74 8 3.37 <0.01
Shoots formed by February 2003 per shootexplant present in January
Family
Treatment
0.47
0.01
8
1
0.90
0.01
ns
nsFamily × treatment 0.52 8 1.39 ns
Shoots formed by April 2003 per shoot explantpresent in January
Family
Treatment
3.45
47.81
8
1
0.35
4.97
ns
nsFamily × treatment 9.81 8 2.11 <0.05
Shoots formed by May 2003 Family 53.17 8 2.12 nsTreatment 1269.89 1 50.76 <0.001Family × treatment 25.11 8 1.11 ns
Shoots tips forming roots Family 3009.45 8 7.83 <0.01Treatment 1071.60 1 2.72 nsFamily × treatment 384.57 8 0.56 ns
Plant survival in the nursery in February 2004 Family 367.09 8 5.00 <0.05Treatment 547.20 1 7.33 <0.05Family × treatment 73.42 8 0.82 ns
Height of all trees in February 2004 Family 1191.88 8 12.79 <0.001Treatment 8794.03 1 96.76 <0.001Family × treatment 93.19 8 1.45 ns
Height of all trees in July 2004 Family 2228.73 8 2.29 nsTreatment 54516.00 1 58.46 <0.001Family × treatment 975.08 8 4.43 <0.001
Height increment in February–July 2004 Family 495.03 8 0.78 nsTreatment 20340.00 1 33.41 <0.001Family × treatment 638.55 8 8.70 <0.001
Physiological age in August 2004 Family 0.38 8 1.49 nsTreatment 16.44 1 210.29 <0.001Family × treatment 0.08 8 2.13 ns
Table 2. Summary of ANOVA results.
axillary shoots and 82% for the epicotyl-axillary shoots. Theserooting percentages were higher than the 69% reported forthe adventitious shoots from cryopreserved cotyledons 13–18 weeks after setting (Hargreaves et al. 2004). The shootsin the current experiment were set in July (winter) ratherthan October–November (spring) and consequently were notexposed to the higher ambient temperatures late spring canproduce in New Zealand (Hargreaves et al. 2004). Nairn(1992) reported ex vitro rooting of 5%–85% across geno-types and a mean of 45%. However, it was not stated whetherthese shoots were initiated adventitiously or were of epicotyl-axillary origin, or for how many weeks the shoots were setbefore the rooting assessment was made.
McKeand and Allen (1984) have reported root-system ab-normalities in tissue-cultured P. taeda. Our adventitious-axillary
shoots tended to have fewer roots than the epicotyl-axillaryshoots, and at planting out into the nursery bed in Decem-ber, there were more vigorously rooted shoots from theepicotyl-axillary-shoot treatments to choose from. This mayhave compounded the differences in subsequent growth andmaturation characteristics observed in the two treatments.Modifications to rooting treatments will be investigated toimprove the rooting of adventitious shoots, which may sig-nificantly improve early growth.
The adventitious-axillary plants were significantly smallerthan the epicotyl-axillary plants 2 months after planting outin the nursery, irrespective of family or genotype. Percentheight differences between the treatments remained similaroverall at 77% (February) and 78% (July). It is too early topredict the field performance of the two stock types, whether
© 2005 NRC Canada
Hargreaves et al. 2637
Initial no. of explants Mean no. of explants from each shoot explant present in January
January 2003 February 2003 April 2003 May 2003
FamilyNo. ofgenotypesa Treatment 1 Treatment 2 Treatment 1 Treatment 2 Treatment 1 Treatment 2 Treatment 1 Treatment 2
1 6 4.17b 2.67a 1.66a 1.42a 4.45a 5.92a 8.02a 13.69a2 9 5.33ab 2.11ab 2.00a 1.96a 4.94a 7.54a 10.70a 19.80a3 9 6.56ab 2.00ab 1.44a 2.22a 4.49a 7.06a 7.84a 16.72a4 9 6.11ab 1.89b 1.92a 1.67a 5.67a 6.39a 8.95a 16.22a5 9 8.00a 2.22ab 1.35a 1.67a 4.45a 5.32a 8.09a 12.85a6 9 5.56ab 2.44ab 1.57a 1.50a 3.56a 7.20a 8.27a 15.89a7 9 4.44b 2.22ab 1.86a 1.67a 5.19a 5.09a 7.26a 10.80a9 9 5.11ab 2.33ab 1.70a 1.50a 6.02a 5.22a 9.79a 12.04a10 5 3.60b 2.00ab 1.92a 1.70a 6.09a 5.60a 9.58a 14.04aMeanb 8.2 5.58a 2.20b 1.71a 1.70a 4.95a 6.19a 8.70b 14.77a
Note: Values within columns followed by the same letter are not significantly different (Tukey’s multiple range test, P ≤ 0.05).aGenotypes from sound embryos; yellowish or malformed embryos were deemed unsound and were discarded.bMean for each treatment over all families within a given sampling time; values in the row followed by the same letter are not significantly different
(Tukey’s multiple range test, P ≤ 0.05).
Table 3. Initial mean numbers of explants present in January 2003 and mean numbers of explants (shoot tips plus stem segments) pro-duced from each shoot present in January at subsequent time intervals from cotyledons (treatment 1) and epicotyls (treatment 2) foreach family, and the number of genotypes represented per family.
No. of shoot tips setin July
Shoot tips rooted inAugust (%)
FamilyNo. ofgenotypes tested Treatment 1 Treatment 2 Treatment 1 Treatment 2
Overall meanrooting (%)
1 6 176 179 100a 98a 99a2 7 210 214 84a 97a 91ab3 7 214 220 93a 99a 96ab4 7 209 214 65a 91ab 79ab5 6 213 185 51a 75ab 68ab6 5 152 144 78a 81ab 80ab7 5 141 147 63a 46b 54b9 7 213 219 83a 89ab 86ab10 4 108 118 49a 59ab 54bMeana 6 181.8 182.2 77a 84a 79
Note: Values within columns values followed by the same letter are not significantly different (Tukey’s multiplerange test, P ≤ 0.05).
aMean for each treatment over all families within a given sampling time; values in the row followed by the sameletter are not significantly different (Tukey’s multiple range test, P ≤ 0.05).
Table 4. Numbers of shoot tips set in July 2003 and percentages of shoot tips showing root elongationin August 2003 in treatment 1 (cotyledons) and treatment 2 (epicotyls) and overall mean percent root-ing for each family, and the number of genotypes represented per family.
the initial differences will be maintained, or how these plantswill perform as stool beds. Results for other species haveshown variable field performance (P. taeda, McKeand 1985;Frampton and Isik 1987; Douglas-fir, Pseudotsuga menziesii(Mirb.) Franco, Ritchie et al. 1994; lodgepole pine, Pinuscontorta Dougl., Flygh et al. 1998).
Assessment of the physiological age of the plants in thenursery bed indicated that plants of all families and geno-types in the adventitious-axillary-shoot treatment showed someincrease compared with those of epicotyl-axillary-shoot ori-gin. The published methods most similar to those presentedhere are for P. menziesii, where the authors concluded froma comparison of eight genotypes that the vegetative propagulesexhibited traits of mature trees and that these traits were par-ticularly marked in the cotyledon-derived plantlets (Ritchieet al. 1994). Adventitious shoots from cotyledons of mari-
time pine, Pinus pinaster Ait., were found to be closer inrelative maturity to the young seedlings than to micrograftedmeristems of 2- or 13-year-old material (Monteuuis and Dumas1992).
Plants formed adventitiously from cotyledons of the samegenotype cryopreserved (with three storage durations) andthen thawed showed no overt differences between cryopre-served and non-cryopreserved cotyledons in either in vitroperformance, rooting, or growth when assessed after 2 years’growth in the nursery (Hargreaves et al. 2004). Some geno-types exhibited more mature characteristics (closed vegeta-tive buds) than others.
There are both advantages and disadvantages associatedwith physiological age (Menzies and Klomp 1988). Resultsof field trials of P. radiata cuttings have indicated that an op-timum physiological age of 3–4 years confers the advantage
© 2005 NRC Canada
2638 Can. J. For. Res. Vol. 35, 2005
No. of plants set in nurserybed in December 2003
Plants surviving inFebruary 2004 (%)
FamilyNo. ofgenotypes tested Treatment 1 Treatment 2 Treatment 1 Treatment 2
Overallsurvival (%)
1 6 120 120 72b 81a 77b2 6 144 144 90a 92a 91a3 7 166 166 90a 93a 91a4 7 150 162 76ab 91a 82ab5 5 96 120 89ab 97a 92a6 5 102 109 94a 97a 96a7 5 96 95 88ab 88a 88ab9 7 162 168 83ab 90a 86ab10 4 78 84 82ab 91a 87abMeana 5.8 123.8 129.8 85b 90a 88
Note: Values within columns values followed by the same letter are not significantly different (Tukey’s multiplerange test, P ≤ 0.05).
aMean for each treatment over all families within a given sampling time; values in the row followed by the sameletter are not significantly different (Tukey’s multiple range test, P ≤ 0.05).
Table 5. Numbers of plants planted in the nursery beds in December 2003 and percentages of rootedshoots surviving in the nursery bed in February 2004 in treatment 1 (cotyledons) and treatment 2 (epi-cotyls) for each family and the number of genotypes represented per family.
February 2004 July 2004 February–July 2004
Treatment 1 Treatment 2 Treatment 1 Treatment 2 Treatment 1 Treatment 2
Family
No. ofgenotypestested
No.oftrees
Plantheight(cm)
No.oftrees
Plantheight(cm)
No.oftrees
Plantheight(cm)
No.oftrees
Plantheight(cm)
Heightincrement(cm)
Heightincrement(cm)
1 6 104 17.8a 117 22.5a 98 38.9ab 95 47.3b 20.8b 24.0c2 6 130 17.8a 132 21.0b 128 41.4a 129 49.2b 23.6a 28.2ab3 7 151 15.1b 156 19.2c 150 38.5ab 155 48.4b 23.4a 29.2ab4 7 125 14.4b 142 19.4c 113 34.0c 134 49.7ab 19.1bc 30.2ab5 5 91 13.8bc 104 16.9e 76 32.4c 94 47.4b 18.7bc 30.3a6 5 95 15.4b 103 17.3de 91 40.3ab 100 43.1c 25.0a 25.7c7 5 81 13.0cd 85 18.7cd 79 33.9c 83 46.8b 21.0b 28.1b9 7 131 17.6a 151 23.0a 111 37.9b 147 52.7a 20.6b 29.5ab10 4 58 11.6d 77 15.2f 56 28.3d 75 40.4c 17.4c 25.2cMeana 5.7 107.3 15.4b 118.6 19.6a 100.2 36.9b 112.4 47.8a 21.4b 28.1a
Note: Values within columns values followed by the same letter are not significantly different (Tukey’s multiple range test, P ≤ 0.05).aMean for each treatment over all families within a given sampling time; values in the row followed by the same letter are not significantly different
(Tukey’s multiple range test, P ≤ 0.05).
Table 6. Plant heights for treatment 1 (cotyledons) and treatment 2 (epicotyls) measured in February and July 2004, the numbers oftrees measured, the growth increments between February and July for each family, and the number of genotypes represented per family.
of improved stem form without the early loss of stem diame-ter growth associated with older physiological ages (Men-zies et al. 1991a, 1991b, 1991c). McKeand (1985) alsonoted that P. taeda plantlets exhibiting increased maturationcharacteristics showed a lower incidence of fusiform rust.The observations made in this paper have implications forother pine species where adventitious-shoot production fromzygotic embryos is being developed with reference to possi-ble maturation effects (Gonzalés et al. 1998; Meyer 1998;Mathur and Nadgauda 1999; Tang et al. 2004).
The causes of these differences in physiological age ob-served in shoots formed adventitiously from cotyledons areunknown. The levels of cytokinins used for adventitious cal-lus formation can promote high nuclear-DNA levels and thishas led to concern about their use in plant-propagation sys-tems (Patel and Berlyn 1982; Renfoe and Berlyn 1985).However, examination of genetic stability in micropropagatedshoots regularly exposed to BA pulses (2.25 mg·L–1) over a10-year period indicated that the regenerants were geneti-cally stable (Goto et al. 1998). In a recent study, no negativeeffects on regeneration of P. radiata shoots from meristematictissue that had been subcultured for 2 years on medium con-taining BA were reported (Schestibratov et al. 2003). The ef-fects of lower levels of BA or shorter periods of exposure onsubsequent performance have not yet been tested in P. radiata.It appears that the differences observed between adventi-tious-axillary and epicotyl-axillary shoots have more to dowith the process of adventitious-shoot formation than withan interaction of individual genotypes with the plant-growthregulators used. These suggestions are supported by Framptonet al. (1998) in work with P. taeda.
Another important potential application of our results isthe preservation of unique genetic material from commercialbreeding programs or from endangered native populations.
The relatively low technology requirements of the methodsfor both cryopreservation and subsequent plant productionsuggest that limited supplies of seed could provide cotyle-dons both for shoot production and for long-term storage inliquid nitrogen. The latter would provide an alternative sourceof germplasm for replanting or restoration in the event ofnatural disaster in the field. Others have suggested that theadventitious-axillary-shoot method may facilitate future re-search in genetic transformation of P. radiata (Schestibratovet al. 2003) and P. taeda (Tang and Guo 2001). The tech-niques described here may also provide a helpful researchtool for identifying quantitative trait loci associated withadventitious-shoot formation in tissue culture. Quantitativetrait loci for this trait have been found in Arapodopsis (Lallet al. 2004).
ConclusionsThis is the first published report comparing the production
and performance of adventitious shoots from cryopreservedcotyledons and axillary shoots formed from epicotyls fromthe same zygotic embryo. Family differences were significant.Both methods were sufficiently successful to have potentialfor clonal forestry. The adventitious-axillary-shoot methodyielded high genotype capture and the significant advantageof allowing juvenile material to be preserved, by cryopreservingcotyledons while field testing takes place.
The adventitious-axillary plants produced were shorter thanthe epicotyl-axillary plants when planted out in the nursery,and this was still apparent at lifting for field planting. As-sessment of relative physiological age indicated that all plantsof adventitious-axillary-shoot origin showed some increasein maturation. However, the significance of that result willnot be known until the data from the field trials now in prog-ress are available.
The simple method described here for cotyledon cryopre-servation with resultant high genotype capture and produc-tion of planting stock may prove to be a useful tool, providingessential insurance for endangered (native) populations.
Acknowledgements
Drs. Wes Hackett, Luis Gea, and Heidi Dungey are thankedfor helpful comments on the manuscript and Tania Elder forher skills in word processing and document presentation.Comments by two anonymous reviewers were also helpful.
References
Aitken-Christie, J., and Jones, C.L. 1987. Towards automation:radiata pine shoot hedges in vitro. Plant Cell Tissue Organ Cult.8: 185–196.
Aitken-Christie, J., and Singh, A.P. 1987. Cold storage of tissuecultures. In Cell culture and tissue culture in forestry. Vol. 2.Edited by J.M. Bonga and D.J. Durzan. Martinus Nijhoff Pub-lishers, Dordrecht, Netherlands. pp. 285–304.
Aitken-Christie, J., Jones, C., and Bond, S. 1985. Wet and waxyshoots in radiata pine micropropagation. Acta Hortic.(Wageningen), 166: 93–100.
Aitken-Christie, J., Singh, A.P., and Davies, H. 1988. Multiplica-tion of meristematic tissue: a new tissue culture system forradiata pine. In Genetic manipulation of woody plants. Edited by
© 2005 NRC Canada
Hargreaves et al. 2639
Mean physiological age (years)a
FamilyNo. ofgenotypes Treatment 1 Treatment 2 Difference
1 6 1.9a 1.2a 0.7ab2 6 2.4a 1.6a 0.8ab3 7 2.4a 1.6a 0.8ab4 7 2.3a 1.5a 0.8ab5 5 2.3a 1.2a 1.1a6 5 2.1a 1.6a 0.5b7 5 2.3a 1.4a 0.9ab9 7 2.1a 1.1a 1.0ab10 4 2.0a 1.3a 0.8abMeanb 5.8 2.2a 1.4b 0.8
Note: Values within columns followed by the same letter are not signif-icantly different (Tukey’s multiple range test, P ≤ 0.05).
aPhysiological age was assessed to the nearest 6 months; values givenare family averages.
bMean for each treatment over all families within a given samplingtime; values in the row followed by the same letter are not significantlydifferent (Tukey’s multiple range test, P ≤ 0.05).
Table 7. Mean physiological ages in August 2004 for treatment1 (cotyledons) and treatment 2 (epicotyls), estimated using mor-phological markers (Menzies et al. 2000), and differences be-tween treatments, calculated as treatment 1 minus treatment 2, foreach family, and the number of genotypes represented per family.

J.W. Hanover and D.E. Keathley. Plenum Publishing Corp.,New York. pp. 413–432.
Attree, S.M., and Fowke, L.C. 1991. Micropropagation through so-matic embryogenesis in conifers. In Biotechnology in agricul-ture and forestry. 17. High-tech and micropropagation 1. Editedby Y.P.S. Bajaj. Springer-Verlag, New York. pp. 53–70.
Debergh, P., Aitken-Christie, J., Cohen, D., Grout, B., von Arnold,S., Zimmerman, R., and Ziv, M. 1992. Reconsideration of theterm ‘vitrification’ as used in micropropagation. Plant Cell Tis-sue Organ Cult. 30: 135–140.
Durzan, D.J., and Gupta, P.K. 1988. Somatic embryogenesis andpolyembryogenesis in conifers. Adv. Biotechnol. Process. 9: 53–81.
Faulds, T., and Dibley, M.J. 1991. Potential propagation methodsfor clonal forestry. In Proceedings of the FRI/NZFP Forests Ltd.Clonal Forestry Workshop. Edited by J.T. Miller. New ZealandMinistry of Forestry, Rotorua. FRI Bull. 160. pp. 67–68.
Flygh, G., Grönroos, R., Högberg, K.-A., and von Arnold, S. 1998.Development and growth of plantlets of Pinus contorta regener-ated from adventitious buds. Scand. J. For. Res. 13: 331–339.
Frampton, L.J., Jr., and Isik, K. 1987. Comparison of field growthamong loblolly pine seedlings and three plant types produced invitro. TAPPI J. 70(7): 119–123.
Frampton, L.J., Jr., Amerson, H.V., and Leach, G.N. 1998. Tissueculture method affects ex vitro growth and development ofloblolly pine. New For. 16: 125–138.
Gonzalés, M.V., Rey, M., Tavazza, R., La Malfa, S., Cuozzo, L.,and Ancora, G. 1998. In vitro adventitious shoot formation oncotyledons of Pinus pinea. Hort. Sci. 33(4): 749–750.
Goto, S., Thakur, R.C., and Ishii, K. 1998. Determination of ge-netic stability in long-term micropropagated shoots of Pinusthunbergii Parl. using RAPD markers. Plant Cell Rep. 18: 193–197.
Hargreaves, C.L., Smith, D.R., Foggo, M.N., and Gordon, M.E.1997. Cryopreservation of zygotic embryos of Pinus radiata andsubsequent plant regeneration. In Proceedings of IUFRO ‘97:Genetics of Radiata Pine, 1–5 December 1997, Rotorua, NewZealand. Edited by R.D. Burdon and J.M. Moore. New ZealandForest Research Institute, Rotorua. FRI Bull. 203. pp. 281–284.
Hargreaves, C.L., Foggo, M.N., Smith, D.R., and Gordon, M.E.1999. Development of protocols for the cryopreservation of zy-gotic embryos of Pinus radiata and subsequent plant regenera-tion. N.Z. J. For. Sci. 29: 54–63.
Hargreaves, C., Grace, L., van der Mass, S., Reeves, C., Holden,G., Menzies, M., Kumar, S., and Foggo, M. 2004. Cryopreserva-tion of Pinus radiata zygotic embryo cotyledons: effect of stor-age duration on adventitious shoot formation and plant growthafter 2 years in the field. Can. J. For. Res. 34: 600–608.
Horgan, K., and Holland, L. 1989. Rooting micropropagated shootsfrom mature radiata pine. Can. J. For. Res. 19: 1309–1315.
Horgan, K., Skudder, D., and Holden, G. 1997. Clonal storage andrejuvenation. In Proceedings of IUFRO ‘97: Genetics of RadiataPine, 1–5 December 1997, Rotorua, New Zealand. Edited byR.D. Burdon and J.M. Moore. New Zealand Forest Research In-stitute, Rotorua. FRI Bull. 203. pp. 273–280.
Lall, S., Nettleton, D., DeCook, R., Che, P., and Howell, S.H.2004. Quantitative trait loci associated with adventitious shootformation in tissue culture and the program of shoot develop-ment in Arabidopsis. Genetics, 167: 1883–1892.
Mathur, G., and Nadgauda, R. 1999. In vitro plantlet regenerationfrom mature zygotic embryos of Pinus wallichiana A.B. Jacks.Plant Cell Rep. 19: 74–80.
McKeand, S.E. 1985. Expression of mature characteristics by tis-sue culture plantlets derived from embryos of loblolly pine. J.Am. Soc. Hort. Sci. 110: 619–623.
McKeand, S.E., and Allen, H.L. 1984. Nutritional and root devel-opment factors affecting growth of tissue culture plantlets ofloblolly pine. Physiol. Plant. 61: 523–528.
Menzies, M.I., and Aimers-Halliday, J. 1997. Propagation optionsfor clonal forestry with Pinus radiata. In Proceedings of IUFRO‘97: Genetics of Radiata Pine, 1–5 December 1997, Rotorua,New Zealand. Edited by R.D. Burdon and J.M. Moore. NewZealand Forest Research Institute, Rotorua. FRI Bull. 203.pp. 256–263.
Menzies, M.I., and Klomp, B.K. 1988. Effects of parent age ongrowth and form of cuttings and comparison with seedlings. InWorkshop on Growing Pinus radiata from Cuttings, Rotorua, 5–7 May 1986. Edited by M.I. Menzies, J.P. Aimer, and L.J.Whitehouse. New Zealand Ministry of Forestry, Rotorua. FRIBull. 135 pp. 18–40.
Menzies, M.I., Faulds, T., Dibley, M.J., and Aitken-Christie, J.1985. Vegetative propagation of radiata pine in New Zealand. InProceedings of the International Symposium on Nursery Man-agement Practices for the Southern Pines, 4–9 August 1985,Montgomery, Ala. Edited by D.B. South. Department of Re-search Information, Auburn University, Auburn, Ala. pp. 167–190.
Menzies, M.I., Klomp, B.K., and Holden, D.G. 1991a. Optimalphysiological age of propagules for uses in clonal forestry. InProceedings of FRI/NZFP Forests Ltd. Clonal Forestry Work-shop. Edited by J.T. Miller. New Zealand Ministry of Forestry,Rotorua. FRI Bull. 160. pp. 142–145.
Menzies, M., Klomp, B., and Holden, G. 1991b. Promising futurefor radiata pine cuttings. In What’s new in forest researchNo. 212. New Zealand Forest Research Institute, Rotorua.
Menzies, M.I., Klomp, B.K., Holden, D.G., and Hong, S.O. 1991c.The effects of initial spacing on growth and crop selection ofradiata pine seedlings and cuttings. In Efficiency of stand estab-lishment operations. Edited by M.I. Menzies, G.E. Parrott,and L.J. Whitehouse. New Zealand Ministry of Forestry,Rotorua. FRI Bull. 156. pp. 152–164.
Menzies, M.I., Dibley, M.J., Faulds, T., Aimers-Halliday, J., andHolden, D.G. 2000. Research note: morphological markers ofphysiological age for Pinus radiata. N.Z. J. For. Sci. 30: 359–364.
Meyer, H.J. 1998. In vitro formation of adventitious buds on ma-ture embryos of Pinus elliottii Engelm. × P. caribaea Morelethybrids. 1998. S. Afr. J. Bot. 64(3): 220–225.
Monteuuis, O., and Dumas, E. 1992. Morphological features as in-dicators of maturity in acclimatized Pinus pinaster from differ-ent in vitro origins. Can. J. For. Res. 22: 1417–1421.
Nairn, B.J. 1992. Commercial micropropagation of radiata pine. InMicropropagation of woody plants. Edited by M.R. Ahuja.Kluwer, Dordrecht, Netherlands. For. Sci. 41: 383–394.
Nairn, B.J., Furneaux, R.H., and Stevenson, T.S. 1995. Identifica-tion of an agar constituent responsible for hydric control inmicropropagation of radiata pine. Plant Cell Tissue Organ Cult.43: 1–11.
Park, Y.S., Barrett, J.D., and Bonga, J.M. 1998. Application of so-matic embryogenesis in high-value clonal forestry: deployment,genetic control and stability of cryopreserved clones. In VitroCell. & Dev. Biol. Plant, 34: 231–239.
Patel, K.R., and Berlyn, G.P. 1982. Genetic instability of multiplebuds of Pinus coulteri regenerated from tissue culture. Can. J.For. Res. 12: 93–101.
Quoirin, M., and Lepoivre, P. 1977. Études des milieux adaptésaux cultures in vitro de Prunus. Acta Hortic. 78: 437–442.
Renfoe, M.H., and Berlyn, G.P. 1985. Variation in nuclear DNAcontent in Pinus taeda L. tissue culture of diploid origin. J.Plant Physiol. 121: 131–139.
© 2005 NRC Canada
2640 Can. J. For. Res. Vol. 35, 2005

Ritchie, G.A., Duke, S.D., and Timmis, R. 1994. Maturation inDouglas-fir: II. Maturation characteristics of genetically matchedDouglas-fir seedlings, rooted cuttings and tissue culture plantletsduring and after 5 years of field growth. Tree Physiol. 14: 1261–1275.
SAS Institute Inc. 1989. SAS/STAT® users’s guide. Version 6. 4thed. SAS Institute Inc., Cary, N.C.
Schestibratov, K.A., Mikhailov, R.V., and Dolgov, S.V. 2003. Plantletregeneration from subculturable nodular callus of Pinus radiata.Plant Cell Tissue Organ Cult. 72: 139–146.
Sommer, H.E., Brown, C.L., and Kormanik, P.P. 1975. Differentia-tion of plantlets in long leaf pine (Pinus palustris Mill.) tissueculture in vitro. Bot. Gaz. 136: 196–200.
Tang, W., and Guo, Z. 2001. In vitro propagation of loblolly pinevia direct somatic organogenesis from mature cotyledons andhypocotyls. Plant Growth Regul. 33: 25–31.
Tang, W., Harris, L.C., Outhavong, V., and Newton, R.J. 2004. Theeffect of different plant growth regulators on adventitious shootformation from Virginia pine (Pinus virginiana) zygotic embryoexplants. Plant Cell Organ Cult. 78: 237–240.
Timmis, R. 1998. Bioprocessing for tree production in the forestryindustry: conifer somatic embryogenesis. Biotechnol. Prog. 14:156–166.
© 2005 NRC Canada
Hargreaves et al. 2641
Related Documents











