Comparative Immunophenotyping of Equine Multipotent Mesenchymal Stromal Cells: An Approach Toward a Standardized Definition Felicitas Paebst, 1,2 * Daniel Piehler, 3 Walter Brehm, 2 Sandra Heller, 1 Carmen Schroeck, 4 Attila T arnok, 5 Janina Burk 1,2 Abstract Horses are an approved large animal model for therapies of the musculoskeletal system. Especially for tendon disease where cell-based therapy is commonly used in equine patients, the translation of achieved results to human medicine would be a great accomplishment. Immunophenotyping of equine mesenchymal stromal cells (MSCs) remains the last obstacle to meet the criteria of the International Society for Cellular Therapy (ISCT) definition of human MSCs. Therefore, the surface antigen expression of CD 29, CD 44, CD 73, CD 90, CD 105, CD 14, CD 34, CD 45, CD 79a, and MHC II in equine MSCs from adipose tissue, bone marrow, umbilical cord blood, umbilical cord tissue, and tendon tissue was analyzed using flow cytometry. Isolated cells from the different sources and donors varied in their expression pattern of MSC-defining antigens. In particular, CD 90 and 105 showed most heterogeneity. However, cells from all samples were robustly positive for CD 29 and CD 44, while being mostly negative for CD 73 and the exclusion markers CD 14, CD 34, CD 45, CD 79a and MHC II. Fur- thermore, it was evident that enzymes used for cell detachment after in vitro-culture affected the detection of antigen expression. These results emphasize the need of stand- ardization of MSC isolation, culturing, and harvesting techniques. As the equine MSCs did not meet all criteria the ISCT defined for human MSCs, further investigations for a better characterization of the cell type should be conducted. V C 2014 International Society for Advancement of Cytometry Key terms horse; MSC; flow cytometry; surface antigen expression; detachment methods; animal model MESENCHYMAL stromal cells (MSCs) are a promising therapeutic option for a broad range of diseases. Encouraging results could be achieved in immunologic dis- orders like graft versus host disease (1,2) as well as in the treatment of inflammatory conditions (3) and degenerative diseases especially in orthopedics, like tendinopathy and osteoarthritis (4). Since the first isolation of MSCs from bone marrow (5), many different tissues were found to harbor MSCs as well, including adipose tissue (6,7), tendon tissue (8), umbilical cord blood (9), umbilical cord matrix (10,11), and peripheral blood (12,13). MSCs from all sources show a spindle shaped morphology but differ in growth characteristics, differentiation potential (14,15), and gene expression of che- mokine receptors (16). To find a common denomination, in 2006 the International Society for Cellular Therapy (ISCT) published a white paper (17). According to this, human multipotent MSCs (18) must meet the minimal criteria of being plastic adherent, showing the capacity of tri-lineage differentiation and express the surface markers CD 73, CD 90, 1 Translational Centre for Regenerative Medicine (TRM), University of Leipzig, Leipzig, Germany 2 Faculty of Veterinary Medicine, Large Animal Clinic for Surgery, University of Leipzig, Leipzig, Germany 3 Faculty of Veterinary Medicine, Institute of Immunology, University of Leipzig, Leipzig, Germany 4 Institute for Veterinary Anatomy, -Histol- ogy and -Embryology, Justus-Liebig-Uni- versity, Giessen, Germany 5 Faculty of Medicine, Department of Pedi- atric Cardiology, Heart Center GmbH, University of Leipzig, Leipzig, Germany Received 7 February 2014; Revised 1 April 2014; Accepted 1 May 2014 Grant sponsor: German Federal Ministry of Education and Research, Grant num- ber: BMBF 1315883; Grant sponsor: Aka- demie fuer Tiergesundheit (AFT) and Mehl-Muehlhens Foundation Additional Supporting Information may be found in the online version of this article. *Correspondence to: Felicitas Paebst, Large Animal Clinic for Surgery, An den Tierkliniken 21. E-mail: felicitas.- [email protected] Published online 3 June 2014 in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/cyto.22491 V C 2014 International Society for Advancement of Cytometry Cytometry Part A 85A: 678687, 2014 Original Article

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Comparative Immunophenotyping of Equine
Multipotent Mesenchymal Stromal Cells: An
Approach Toward a Standardized Definition
Felicitas Paebst,1,2* Daniel Piehler,3 Walter Brehm,2 Sandra Heller,1 Carmen Schroeck,4
Attila T�arnok,5 Janina Burk1,2
� AbstractHorses are an approved large animal model for therapies of the musculoskeletal system.Especially for tendon disease where cell-based therapy is commonly used in equinepatients, the translation of achieved results to human medicine would be a greataccomplishment. Immunophenotyping of equine mesenchymal stromal cells (MSCs)remains the last obstacle to meet the criteria of the International Society for CellularTherapy (ISCT) definition of human MSCs. Therefore, the surface antigen expressionof CD 29, CD 44, CD 73, CD 90, CD 105, CD 14, CD 34, CD 45, CD 79a, and MHC IIin equine MSCs from adipose tissue, bone marrow, umbilical cord blood, umbilicalcord tissue, and tendon tissue was analyzed using flow cytometry. Isolated cells fromthe different sources and donors varied in their expression pattern of MSC-definingantigens. In particular, CD 90 and 105 showed most heterogeneity. However, cells fromall samples were robustly positive for CD 29 and CD 44, while being mostly negativefor CD 73 and the exclusion markers CD 14, CD 34, CD 45, CD 79a and MHC II. Fur-thermore, it was evident that enzymes used for cell detachment after in vitro-cultureaffected the detection of antigen expression. These results emphasize the need of stand-ardization of MSC isolation, culturing, and harvesting techniques. As the equine MSCsdid not meet all criteria the ISCT defined for human MSCs, further investigations for abetter characterization of the cell type should be conducted. VC 2014 International Society
for Advancement of Cytometry
� Key termshorse; MSC; flow cytometry; surface antigen expression; detachment methods; animalmodel
MESENCHYMAL stromal cells (MSCs) are a promising therapeutic option for a
broad range of diseases. Encouraging results could be achieved in immunologic dis-
orders like graft versus host disease (1,2) as well as in the treatment of inflammatory
conditions (3) and degenerative diseases especially in orthopedics, like tendinopathy
and osteoarthritis (4).
Since the first isolation of MSCs from bone marrow (5), many different tissues
were found to harbor MSCs as well, including adipose tissue (6,7), tendon tissue (8),
umbilical cord blood (9), umbilical cord matrix (10,11), and peripheral blood
(12,13). MSCs from all sources show a spindle shaped morphology but differ in
growth characteristics, differentiation potential (14,15), and gene expression of che-
mokine receptors (16).
To find a common denomination, in 2006 the International Society for Cellular
Therapy (ISCT) published a white paper (17). According to this, human multipotent
MSCs (18) must meet the minimal criteria of being plastic adherent, showing the
capacity of tri-lineage differentiation and express the surface markers CD 73, CD 90,
1Translational Centre for RegenerativeMedicine (TRM), University of Leipzig,Leipzig, Germany
2Faculty of Veterinary Medicine, LargeAnimal Clinic for Surgery, University ofLeipzig, Leipzig, Germany
3Faculty of Veterinary Medicine, Instituteof Immunology, University of Leipzig,Leipzig, Germany
4Institute for Veterinary Anatomy, -Histol-ogy and -Embryology, Justus-Liebig-Uni-versity, Giessen, Germany
5Faculty of Medicine, Department of Pedi-atric Cardiology, Heart Center GmbH,University of Leipzig, Leipzig, Germany
Received 7 February 2014; Revised 1 April2014; Accepted 1 May 2014
Grant sponsor: German Federal Ministryof Education and Research, Grant num-ber: BMBF 1315883; Grant sponsor: Aka-demie fuer Tiergesundheit (AFT) andMehl-Muehlhens Foundation
Additional Supporting Information may befound in the online version of this article.
*Correspondence to: Felicitas Paebst,Large Animal Clinic for Surgery, An denTierkliniken 21. E-mail: [email protected]
Published online 3 June 2014 in WileyOnline Library (wileyonlinelibrary.com)
DOI: 10.1002/cyto.22491
VC 2014 International Society forAdvancement of Cytometry
Cytometry Part A � 85A: 678�687, 2014
Original Article

and CD 105 while lacking the expression of CD 45, CD 34,
CD 14 or 11b, CD 79a or 19, and HLA-DR. Yet there is still
no consensus on a marker panel to define MSCs from other
species than human beings, especially from large model
animals.
Still, with the need for animal models that closely resem-
ble human diseases, the horse is considered to be a superior
large animal model for orthopedic diseases such as Achilles
tendinopathy or osteoarthritis (19–21). This is due to the
resemblance of equine joints and large tendons to their human
equivalents not only regarding anatomical aspects such as car-
tilage thickness (19) or tendon composition and biomechanical
properties (21), but also regarding their pathology. In contrast,
small laboratory animals lack comparability to humans due to
the immense differences in terms of mechanical loads on ten-
dons and cartilage (22). Using the horse as a model in clinical
and experimental studies, promising results have been obtained
with the therapeutical usage of MSCs for orthopedic disorders.
In particular, MSC treatment of equine tendinopathy yielded
lower re-injury rates (23–26) and an improved tendon struc-
ture (27–29). Therefore, the therapeutic results achieved in
equine athletes during the last decade could be valuable in
human medicine as well. However, the suitability of the equine
models for human diseases relies on a far more detailed analy-
sis of the equine MSCs used for therapeutical transfer.
On that account, it is necessary to better characterize
equine MSCs and to assess whether they meet the criteria
which the ISCT proposed for human MSCs. Until now, this
remains a challenge because there are only few monoclonal
antibodies (mAbs) available that show cross-reactivity with
equine epitopes (30–33). Moreover, it is known that there are
differences in cell surface marker expression of MSCs between
different species (34,35). Potentially due to these reasons, the
complete marker panel proposed for human MSCs was not
yet shown to be applicable to equine cells.
Therefore, we aimed to improve the transferability of
studies on MSC therapies in the equine model to human dis-
ease. The purpose of this study was to assess whether the com-
plete array of cell surface markers defined by the ISCT is
applicable to equine MSCs and to further investigate if equine
MSCs can be characterized by a uniform marker expression,
irrespective of their tissue origin.
MATERIALS AND METHODS
Experiment Overview
All monoclonal antibodies (mAb) used in the study were
first tested regarding their cross-reactivity with equine cells.
Positive surface markers (according to ISCT guidelines) were
evaluated on equine MSCs by confocal microscopy and flow
cytometry (FCM), and exclusion markers were tested on
equine peripheral blood mononuclear cells (PBMCs) by FCM.
Further, potential influences of cell detachment methods on
MSC marker expression were assessed using n 5 3 samples of
MSCs and PBMCs.
Based on the protocols that were then established, the
comparative characterization of equine MSCs derived from
five different sources was performed. For each cell source, n 5
6 samples were analyzed by FCM.
Isolation and Culture of MSCs
Six samples from healthy adult warmblood and Haflinger
horses (mean age 5.1 years) of each bone marrow (BM), adi-
pose tissue (AT), and tendon tissue (TT) were collected, as
well as six samples of umbilical cord blood (UCB) and umbili-
cal cord tissue (UCT) from healthy warmblood and thorough-
bred foals within the first hour after birth. Sample collection
had been approved by the local ethics committee (Landesdir-
ektion Leipzig, A 13/10). MSCs were isolated and their differ-
entiation potential was confirmed as described previously (36)
before they were stored at 2134�C until further analysis. The
cells were thawed at 37�C, washed twice and seeded at a den-
sity of 500,000/175 cm2 in cell culture flasks (BD FalconTM,
BD, Franklin Lakes, NJ) using low glucose concentration (1 g/
L) Dulbecco’s modified eagle medium (DMEM) (Life Tech-
nologies, Darmstadt, Germany), supplemented with 20%
heat-inactivated fetal bovine serum (FBS; Sigma–Aldrich,
Steinheim, Germany), 100 IU/mL penicillin, 0.1 mg/mL strep-
tomycin (PAA, Pasching, Austria), and 0.05 mg/mL gentamy-
cin (PAA). Cells were expanded until passage (P) 3 at 37�C,
5% CO2, and 95% humidity. Medium was changed twice a
week.
Isolation and Culture of PBMCs
Syringes were loaded with 15,000 IU heparin-sodium (B
Braun, Tuttlingen, Germany). The jugular vein was prepared
aseptically and a sample of 27 mL peripheral blood was taken
from each horse (mean age 8.3 years). Sample collection had
been approved by the local ethics committee (Landesdirektion
Leipzig, V 06/13). The blood was directly transferred to the
laboratory and PBMCs were isolated using Ficoll Histopa-
queTM (GE Healthcare, Freiburg, Germany) density centrifu-
gation according to the manufacturer’s instruction. Buffycoat
was removed, washed twice with phosphate buffered saline
solution (PBS; PAA), and the cells were immediately subjected
to mAb staining of surface markers. Cells were used as positive
controls for evaluating cross-reactivity of exclusion marker
mAbs and for the assessment of the influence of enzymes on
marker expression.
MSC Detachment
To assess potential influences of enzymes used for cell
detachment on marker expression, different approaches were
performed using MSCs and PBMCs. Cells were washed twice
with PBS without calcium and magnesium. Preliminary tests
were conducted using trypsin-EDTA (Life Technologies)
according to the manufacturer’s instructions. As the results
did not meet the demands, further experiments were per-
formed to compare enzymatic detachment with accutase
(Invitrogen, Darmstadt, Germany) to mechanic cell harvesting
(n 5 3). PBMCs and MSCs were incubated in accutase for 5
min at 37�C, 5% CO2 and 95% humidity. Untreated PBMCs
and MSCs that were detached mechanically were used as con-
trols. Mechanical detachment was carefully performed using
cell scrapers (TPP, Trasadingen, Switzerland) after cold PBS
Original Article
Cytometry Part A � 85A: 678�687, 2014 679
(4�C) containing 0.05 mM EDTA had been added to the MSC
cultures for 10 min. MSCs were subjected to staining for posi-
tive marker expression and PBMCs were stained for exclusion
markers as described below. Subsequently, cells were analyzed
by FCM.
As enzyme treatment impaired the detection of surface
antigens, mechanical cell detachment was performed for all
further investigations.
Flow Cytometry
Cells (1 3 106) were distributed into each well of 96-well
V bottom plates (Carl Roth, Karlsruhe, Germany) and washed
twice with 100 lL PBS. To enable identification of dead cells,
Fixable E-Fluor 780VR
(ebioscience, Heidelberg, Germany) at a
working dilution of 1:1,000 in PBS was used. Staining was per-
formed for 20 min on ice, protected from light. After washing,
cells were resuspended in 20 lL of 1:500 diluted and heat-
inactivated mouse or rat (for CD 44) serum (Serotec, Kidling-
ton, UK). Staining of CD 34/ CD 73 and MHC II additionally
required goat or rabbit serum (Sigma–Aldrich) (Table 1).
Incubation was again performed on ice in the dark for 15
min. Following another washing step in PBS, the staining was
performed combining the mAbs to the subsets of CD 90/34,
CD 14/45/105, CD 29/44/MHCII and single staining of CD 73
and 79a. Table 1 shows the antibodies used, their dilutions as
well as their isotype controls. After each antibody incubation
on ice and protected from light for 15 min, the cells were
washed once with FCM staining buffer (PBS, 3% FBS, 0.01%
sodium azide [Carl Roth]) and once with PBS. For CD 79a
staining, the FIX & PERMVR kit (Invitrogen) was used accord-
ing to the manufacturer’s instruction. Subsequently, these
samples were washed once with PBS and stained at room tem-
perature. Following all staining procedures, cells were fixed by
incubating them in 2% paraformaldehyde (PFA; Carl Roth) or
0.1% PFA for CD 79a for 15 min, washed with PBS, and
stored in FCM staining buffer until analysis.
Cells were analyzed 8 h after staining using a FACS Can-
toIITM (BD) equipped with a blue 488 nm solid state and a
red 633 nm HeNe laser. Compensation was accomplished
with CompBeadsTM (BD). For each sample, preferably
200,000 and at least more than 10,000 events were recorded.
Further analysis was performed using FlowJo 10.0.6 (Tree
Star, Ashland, Oregon). Gating strategies focused on living
cells and exclusion of doublets (Fig. 1). Geometric mean fluo-
rescence intensity was used to define positive cells and percen-
tages were calculated.
Confocal Microscopy
Cells were stained as described in the previous section
and subjected to a cytospin (Shandon Cytopin 4, Thermo
Fisher Scientific, Waltham, MA) at 1,000 rpm for 3 min.
Cover slips were attached with CalbiochemVR
MowiolVR
(Merck
KGaA, Darmstadt, Germany) mixed with 1,4-Diazabicy-
clo[2.2.2]octane (DABCO, Sigma–Aldrich). Slides were stored
at 4�C protected from light. Analysis was performed by an
inverted confocal laser scanning microscope TCS SP5 (Leica
Microsystems GmbH, Wetzlar, Germany). To avoid a cross
talk in excitation of multiple stained compounds, a sequential
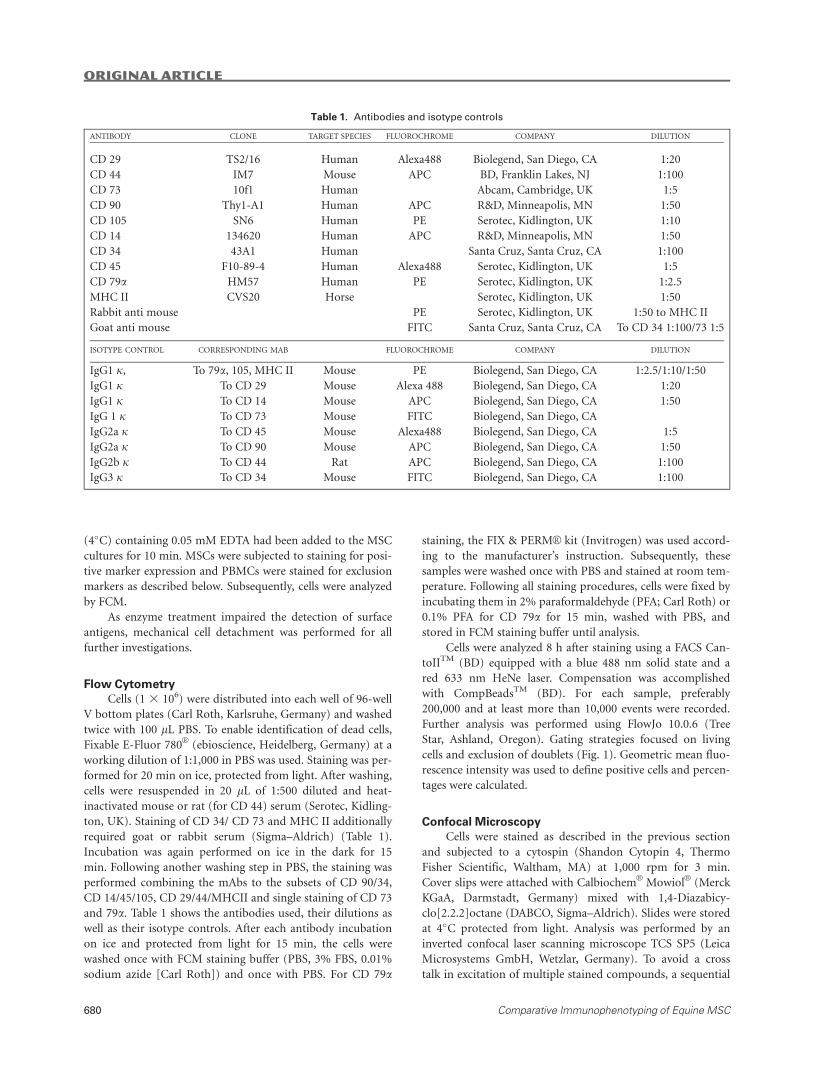
Table 1. Antibodies and isotype controls
ANTIBODY CLONE TARGET SPECIES FLUOROCHROME COMPANY DILUTION
CD 29 TS2/16 Human Alexa488 Biolegend, San Diego, CA 1:20
CD 44 IM7 Mouse APC BD, Franklin Lakes, NJ 1:100
CD 73 10f1 Human Abcam, Cambridge, UK 1:5
CD 90 Thy1-A1 Human APC R&D, Minneapolis, MN 1:50
CD 105 SN6 Human PE Serotec, Kidlington, UK 1:10
CD 14 134620 Human APC R&D, Minneapolis, MN 1:50
CD 34 43A1 Human Santa Cruz, Santa Cruz, CA 1:100
CD 45 F10-89-4 Human Alexa488 Serotec, Kidlington, UK 1:5
CD 79a HM57 Human PE Serotec, Kidlington, UK 1:2.5
MHC II CVS20 Horse Serotec, Kidlington, UK 1:50
Rabbit anti mouse PE Serotec, Kidlington, UK 1:50 to MHC II
Goat anti mouse FITC Santa Cruz, Santa Cruz, CA To CD 34 1:100/73 1:5
ISOTYPE CONTROL CORRESPONDING MAB FLUOROCHROME COMPANY DILUTION
IgG1 j, To 79a, 105, MHC II Mouse PE Biolegend, San Diego, CA 1:2.5/1:10/1:50
IgG1 j To CD 29 Mouse Alexa 488 Biolegend, San Diego, CA 1:20
IgG1 j To CD 14 Mouse APC Biolegend, San Diego, CA 1:50
IgG 1 j To CD 73 Mouse FITC Biolegend, San Diego, CA
IgG2a j To CD 45 Mouse Alexa488 Biolegend, San Diego, CA 1:5
IgG2a j To CD 90 Mouse APC Biolegend, San Diego, CA 1:50
IgG2b j To CD 44 Rat APC Biolegend, San Diego, CA 1:100
IgG3 j To CD 34 Mouse FITC Biolegend, San Diego, CA 1:100
Original Article
680 Comparative Immunophenotyping of Equine MSC

scanning mode was used. Images were acquired with photo
multipliers. Leica Application Suite Advanced Fluorescence
2.6.0 and Adobe Photoshop CS 8.0.1 were used for processing
the micrographs.
Statistical Analysis
Statistical analysis was performed using IBM SPSSVR
Statistics V22. Antigen expression was compared using a Krus-
kall–Wallis one-way analysis of variance (a 5 0.05). Subse-
quently, Mann–Whitney-U tests were performed (a 5 0.05).
Moreover, Spearman–Rho tests were applied to correlate
the positive markers (a 5 0.01 and a 5 0.05).
RESULTS
Cross-reactivity of Anti-human Antibodies on Equine
Cells
Monoclonal Abs for ISCT positive markers showed cross-
reactivity on mechanically detached adipose tissue derived
MSCs (Supporting Information Figs. S1 and S2). All mAbs for
exclusion markers were tested positively on PBMCs (Support-
ing Information Fig. S1).
Influence of the Cell Detachment Technique on
Surface Antigen Expression
Enzymatic cell detachment with trypsin–EDTA showed a
strong influence on surface marker expression. Among the
positive markers, only CD 29 and CD 44 could be detected
following this harvesting method (data not shown). The use
of accutase influenced the antigen expression less than trypsin
EDTA but still altered the expression of CD 29, CD 90, CD 45,
CD 79a, and MHC II (Fig. 2). Most prominently, accutase
treatment removed CD 79a in two out of three samples and
tremendously decreased CD 45 expression after 5 min of incu-
bation under standard conditions (Fig. 2).
Mechanical cell detachment preserved all analyzed anti-
gens and was therefore chosen for the following comparative
assessment of antigen expression; however, it yielded low
viability.
MSC Marker Expression
The investigated MSCs showed a diverse marker expres-
sion pattern depending on the source of isolation (Table 2,
Figs. 3A and B). CD 29 and CD 44 were the only markers
which could consistently be detected on MSCs.
The highest percentage of CD 29-positive cells (65.42 6
14.87% SD) was identified in BM-MSCs, while the lowest per-
centage was found in UCB-MSCs (37.5 6 28.61% SD).
Regarding CD 44, the highest percentage of positive cells
was detected in AT-MSCs, with 97.18 6 1.57% SD, differing
significantly from MSCs derived from other sources (P <
0.05). BM and TT-derived MSCs also showed a consistent CD
44 expression pattern. However, MSCs recovered from UCB
and UCT showed a great variance in CD 44 expression, from a
complete lack up to 98.4% positive cells in UCB-MSC sam-
ples, and a range from 0.9% to 93.1% in UCT-MSC samples.
This is reflected by the respective standard deviations, which
exceed the average expression levels (46.08 6 46.1% SD in
UCB-MSCs and 32.2 6 42.6% SD in UCT-MSCs).
CD 73 was not expressed by most equine MSCs, with no
significant differences between the cell sources. Only few sam-
ples of UCT-, UCB-, and BM-MSC contained up to 12.7%
positive cells.
CD 90 was consistently found only on AT-MSCs, with
24.4 6 14.43% SD positive cells, which was significantly more
compared to MSCs from all other sources (P < 0.05). UCT-
MSCs showed no expression of this protein, which was signifi-
cant also compared to TT- and BM-MSCs (P < 0.05).
CD 105 was only expressed by AT-, TT-, and UCT-MSCs,
with the highest percentage of positive cells in AT-MSCs (40.2
6 21.28% SD), followed by TT-MSCs (28.38 6 20.33% SD)
(P < 0.05 compared to UCT-, UCB-, and BM-MSCs for both
AT- and TT-MSCs), and the lowest in UCT-MSCs (6.75 6
10.72% SD) (P < 0.05 compared to UCB-MSCs).
Furthermore, correlations between the expression of sur-
face marker were evident. CD 44 expression was positively
correlated with the expression of CD 90 and CD 105, respec-
tively (P < 0.01) and with the expression of CD 29 (P <
0.05). Moreover, the expression of CD 90 and CD 105 was
positively correlated (P < 0.01).
Exclusion markers were expressed in less than 2% of cells
in most samples. Nevertheless, significantly more CD 14-
positive cells, although still less than 2%, were identified in
AT-MSC samples than in UCT- or BM-MSC samples (P <
0.05). Furthermore, CD 14 and CD 45 were expressed highly
in one UCB-MSC sample (97.1% and 23.8%). Analysis of CD
79a-positive cells showed outliers in another UCB-MSC sam-
ple and in one BM-MSC sample (16.1% and 7.8%).
Figure 1. Gating strategies. The figures show the gating strategies in a representative sample of MSCs derived from tendon tissue. Con-
tour plots in (A) and (D) display the area of forward scatter (FSC-A) versus the area of the side scatter (SSC-A). From all recorded events
(A), living cells were selected based on Live/Dead staining (B). Doublets were excluded on the area versus width forward scatter (FSC-A
vs. FSC-W) (C). Only live and single cells MSCs were used for further analysis (D).
Original Article
Cytometry Part A � 85A: 678�687, 2014 681
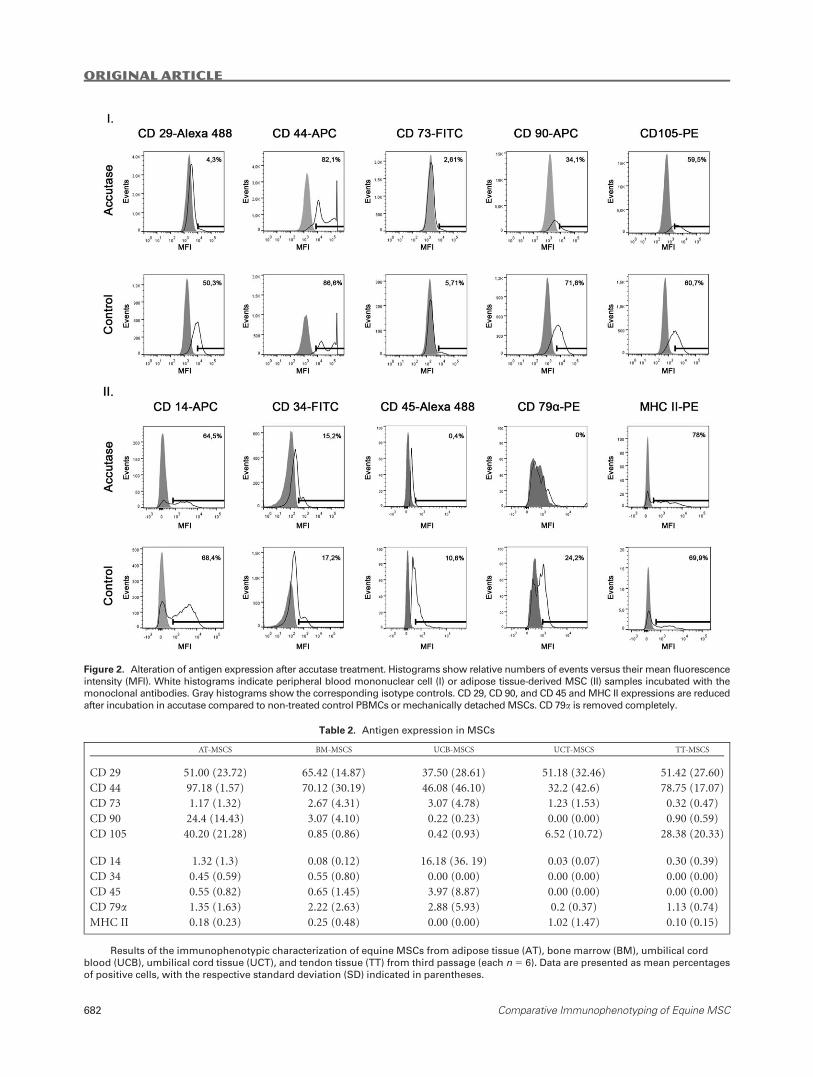
Figure 2. Alteration of antigen expression after accutase treatment. Histograms show relative numbers of events versus their mean fluorescence
intensity (MFI). White histograms indicate peripheral blood mononuclear cell (I) or adipose tissue-derived MSC (II) samples incubated with the
monoclonal antibodies. Gray histograms show the corresponding isotype controls. CD 29, CD 90, and CD 45 and MHC II expressions are reduced
after incubation in accutase compared to non-treated control PBMCs or mechanically detached MSCs. CD 79a is removed completely.
Table 2. Antigen expression in MSCs
AT-MSCS BM-MSCS UCB-MSCS UCT-MSCS TT-MSCS
CD 29 51.00 (23.72) 65.42 (14.87) 37.50 (28.61) 51.18 (32.46) 51.42 (27.60)
CD 44 97.18 (1.57) 70.12 (30.19) 46.08 (46.10) 32.2 (42.6) 78.75 (17.07)
CD 73 1.17 (1.32) 2.67 (4.31) 3.07 (4.78) 1.23 (1.53) 0.32 (0.47)
CD 90 24.4 (14.43) 3.07 (4.10) 0.22 (0.23) 0.00 (0.00) 0.90 (0.59)
CD 105 40.20 (21.28) 0.85 (0.86) 0.42 (0.93) 6.52 (10.72) 28.38 (20.33)
CD 14 1.32 (1.3) 0.08 (0.12) 16.18 (36. 19) 0.03 (0.07) 0.30 (0.39)
CD 34 0.45 (0.59) 0.55 (0.80) 0.00 (0.00) 0.00 (0.00) 0.00 (0.00)
CD 45 0.55 (0.82) 0.65 (1.45) 3.97 (8.87) 0.00 (0.00) 0.00 (0.00)
CD 79a 1.35 (1.63) 2.22 (2.63) 2.88 (5.93) 0.2 (0.37) 1.13 (0.74)
MHC II 0.18 (0.23) 0.25 (0.48) 0.00 (0.00) 1.02 (1.47) 0.10 (0.15)
Results of the immunophenotypic characterization of equine MSCs from adipose tissue (AT), bone marrow (BM), umbilical cord
blood (UCB), umbilical cord tissue (UCT), and tendon tissue (TT) from third passage (each n 5 6). Data are presented as mean percentages
of positive cells, with the respective standard deviation (SD) indicated in parentheses.
Original Article
682 Comparative Immunophenotyping of Equine MSC

DISCUSSION
This is the first study assessing the complete ISCT marker
set in equine MSCs, the difficulties concerning the lack of
cross-reactive mAbs being overcome (31,33,37). Aiming to
obtain comprehensive results, equine MSCs derived from five
different potential sources were evaluated in the study, includ-
ing equine tendon-derived MSCs which had never been sub-
jected to immunophenotyping before.
The results showed that equine MSCs from different
sources exhibit significant differences in their surface antigen
expression. Unfortunately, none of the cell sources could reli-
ably meet the ISCT criteria of human MSC definition. More-
over, cells from different donors often displayed a
heterogenous expression pattern even within the same cell
source, indicating individual differences between donors. In
former studies, conflicting results on mean variation of anti-
gen expression were reported (31,33,38,39). Although in BM-
(31) and AT-MSCs (38), large standard deviations of mean
were reported, other research groups could identify distinct
populations (33,39).
In this study, CD 29 and CD 44 were the only surface
antigens robustly positive in cells derived from all sources.
This finding was consistently reported in different studies and
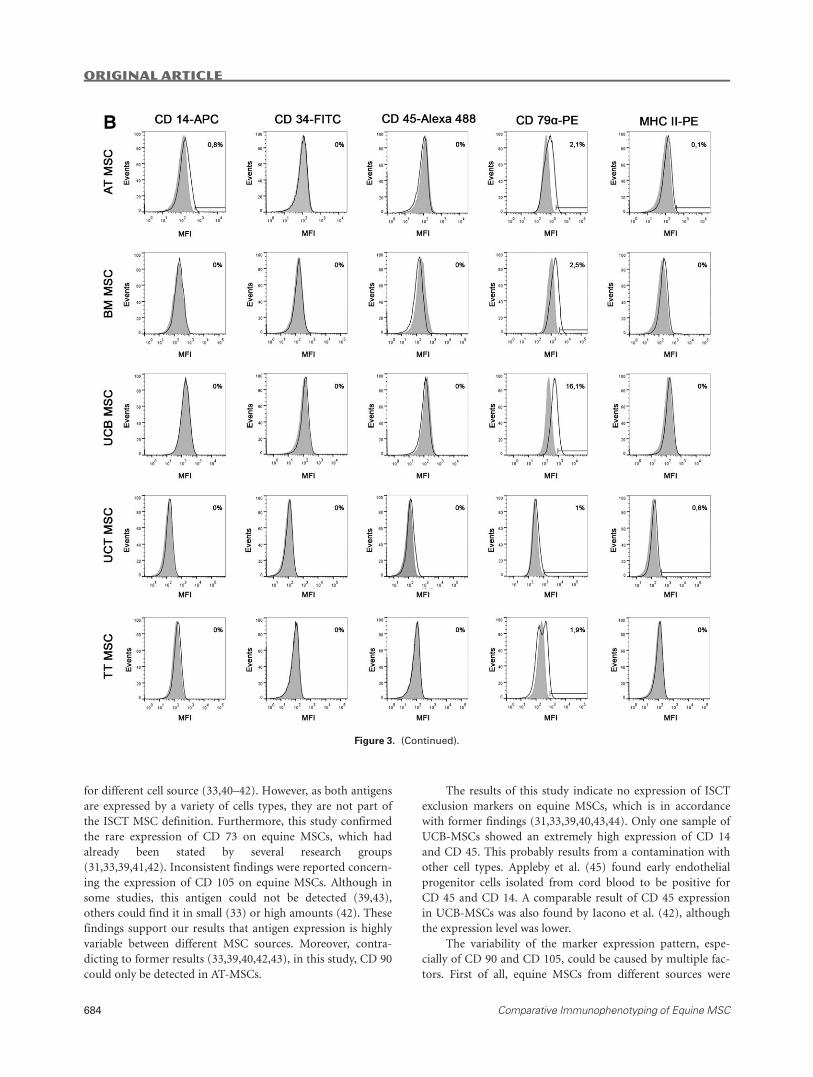
Figure 3. (A) Positive marker expression in MSCs. (B) Exclusion marker expression in MSCs. The figures illustrate the expression of posi-
tive (A) and exclusion (B) marker antigens in representative samples of each cell source. Histograms display relative numbers of events
versus their mean fluorescence intensity (MFI). White histograms indicate samples incubated with monoclonal antibodies, while gray his-
tograms show the corresponding isotype controls. Measuring bars indicate cut off values and percentages of positive cells. AT: adipose
tissue; BM: bone marrow; UCB: umbilical cord blood; UCT: umbilical cord tissue; TT: tendon tissue; MSC: mesenchymal stromal cell.
Original Article
Cytometry Part A � 85A: 678�687, 2014 683
for different cell source (33,40–42). However, as both antigens
are expressed by a variety of cells types, they are not part of
the ISCT MSC definition. Furthermore, this study confirmed
the rare expression of CD 73 on equine MSCs, which had
already been stated by several research groups
(31,33,39,41,42). Inconsistent findings were reported concern-
ing the expression of CD 105 on equine MSCs. Although in
some studies, this antigen could not be detected (39,43),
others could find it in small (33) or high amounts (42). These
findings support our results that antigen expression is highly
variable between different MSC sources. Moreover, contra-
dicting to former results (33,39,40,42,43), in this study, CD 90
could only be detected in AT-MSCs.
The results of this study indicate no expression of ISCT
exclusion markers on equine MSCs, which is in accordance
with former findings (31,33,39,40,43,44). Only one sample of
UCB-MSCs showed an extremely high expression of CD 14
and CD 45. This probably results from a contamination with
other cell types. Appleby et al. (45) found early endothelial
progenitor cells isolated from cord blood to be positive for
CD 45 and CD 14. A comparable result of CD 45 expression
in UCB-MSCs was also found by Iacono et al. (42), although
the expression level was lower.
The variability of the marker expression pattern, espe-
cially of CD 90 and CD 105, could be caused by multiple fac-
tors. First of all, equine MSCs from different sources were
Figure 3. (Continued).
Original Article
684 Comparative Immunophenotyping of Equine MSC

shown to display differences in growth kinetics (36,46), differ-
entiation potential (36,47), and gene expression (36), thus dif-
ferences in surface marker expression could be expected as
well. Furthermore, differences in the culture medium could
influence the cell characteristics (48,49). In this study, to ena-
ble obtaining sufficient cell numbers from all MSC sources,
DMEM supplemented with 20% FBS was used, instead of
10% which is more commonly used (31,33,39). Increased FBS
content could enhance cell growth and influences other cell
features (48). Moreover, the timeframe of plastic-adherent
culture is known to affect the surface marker expression of
MSCs (38,40). Radcliffe et al. found that in BM-derived
MSCs, the expression of CD 90 increased over time and
showed a consistent expression at day 14 (40). BM-MSCs
were also shown to lose CD 14 at the same time point and to
acquire a homogenous marker profile (38) although contra-
dicting the other findings, a consistent expression of CD 90
from P0 on was found in this latter study. However, in this
study, only P3 cells were used for immunophenotyping and
still did not show uniform marker profiles.
In addition, as was demonstrated in this study, the cell
harvesting technique is of tremendous importance for the
detection of surface markers. To our knowledge, all other
reports are based on enzymatic detachment either with tryp-
sin or accumax/accutase prior to the mAb staining. However,
in none of these studies, the influence of the enzyme on anti-
gen expression was tested on proper positive controls for the
exclusion markers. So far, there is only limited knowledge
about the effects of enzymes on the stability of surface antigen
expression. Panchision et al. found that papain most aggres-
sively reduced expression of surface proteins on murine neu-
ronal and human tumor cells, whereas accutase seemed to
preserve them but also showed lowest detachment efficiency
(50). In this study, accutase and trypsin EDTA both affected
the expression of surface markers. Accutase was less aggressive,
but still reduced the expression of CD 45 and completely abol-
ished the staining of CD 79a. Therefore, cells were detached
mechanically in the further course of the study, which in con-
trast to previous studies could explain small amounts of CD
79a found on MSCs from different sources. Although
mechanical cell detachment is an uncommon technique,
probably due to the increase of dead cells in the sample, we
conclude from our results that it is very valuable for immuno-
phenotyping to prevent false negative results.
The study revealed distinct differences in the surface anti-
gen expression between equine and human MSCs. Therefore,
one should consider the known differences in features of
MSCs from different species. As in equine MSCs, CD 29 and
CD 44 expressions are found consistently in canine
(34,35,51,52), ovine (53), and porcine (54) MSCs. Corre-
sponding to the equine antigen expression profile as well, CD
73 could not be detected in canine, rabbit, or sheep MSCs
(35,55,56), while rat tendon derived, feline and bovine neona-
tal cell sources and caprine bone marrow-derived MSCs
expressed this antigen (57–60). CD 73 protein expression
could also be detected in bone marrow derived MSCs from
mice but only in moderate amounts (61). MSCs isolated from
different tissues of pigs robustly express CD 90 (54,62–64)
and CD 105 (54). CD 73 investigation in pig is hampered by a
lack of cross-reactive mAbs (63).
Immunophenotyping of MSCs from large animals is still
a challenge and a common definition of the cell type for the
respective species is lacking while the characterization of these
cells in humans and small laboratory animals is not hampered
by such difficulties. However, to our knowledge, there is only
one study on human MSCs in which the complete ISCT
marker set was actually used (65). Considering the results of
this study, which demonstrate significant differences between
MSCs from different sources and a high variability of marker
expression in MSCs from different donors, it could be debated
if this is solely due to the fact that equine cells were used.
Nevertheless, these results give evidence that standard isola-
tion procedures do not lead to uniform cell populations,
which is likely to apply to all species. Furthermore, as none of
the markers included in the ISCT MSC definition is specific
for MSCs itself, a broad array of antibodies needs to be inves-
tigated for binding to the cells of interest, especially in clinical
trials, to avoid false positive results in the identification of
MSCs.
Aiming to identify some putative markers which could
facilitate the isolation of more homogeneous equine MSC
populations, the correlation of surface antigen expression was
also assessed in this study. CD 44, although very unspecific
and expressed by many different cell types, seems to be a reli-
able target, as expression of this surface marker was correlated
with each positive marker except for CD 73. Correspondingly,
de Schauwer et al. had also found a strong correlation between
CD 29 and CD 44-positive cells as well as MHC II negative
cells (33). Consequently, further experiments should be per-
formed to investigate if cell sorting of CD 44 positive cells
prior to cell culture could lead to uniform populations with
increased percentages of cells positive for the required ISCT
markers.
In conclusion, equine MSCs, although being plastic-
adherent and capable of tri-lineage differentiation, could not
meet the minimal criteria which are demanded for human
MSCs. Moreover, plastic adherence as a selection method for
equine MSCs does not lead to uniform populations. Sorting
of CD 441 cells prior to plastic adherence isolation could lead
to more homogenous populations. Nevertheless, cell charac-
terization based on the defined surface markers should be
reconsidered for equine MSCs. However, if the MSC defini-
tion of the ISCT is to be applied, we recommend using the
complete marker panel, as none of the markers is specific for
MSCs. Furthermore, there is a need to standardize cell culture
and harvesting techniques to achieve comparable results in
the future.
ACKNOWLEDGMENTS
The authors acknowledge Prof. Dr. A. Bader and Prof.
Dr. P. Seibel (both Center for Biotechnology and Biomedicine,
University of Leipzig) for kindly providing the laboratory
facilities. Furthermore, the authors would like to thank Prof.
Original Article
Cytometry Part A � 85A: 678�687, 2014 685

Dr. G. Alber (Institute of Immunology, Faculty of Veterinary
Medicine, University of Leipzig) for his valuable support.
Finally, the authors acknowledge the Orthopedic and Stem
Cell Research (OSC-R) team (Large Animal Clinic for Surgery,
University of Leipzig), especially C. Gittel and Dr. I. Ribitsch,
for the isolation of MSCs. The work presented in this paper
was made possible by funding from the German Federal
Ministry of Education and Research (BMBF 1315883).
LITERATURE CITED
1. Muroi K, Miyamura K, Ohashi K, Murata M, Eto T, Kobayashi N, Taniguchi S,Imamura M, Ando K, Kato S, et al. Unrelated allogeneic bone marrow-derived mes-enchymal stem cells for steroid-refractory acute graft-versus-host disease: A phase I/II study. Int J Hematol 2013;2:206–213.
2. Resnick IB, Barkats C, Shapira MY, Stepensky P, Bloom AI, Shimoni A, Mankuta D,Varda-Bloom N, Rheingold L, Yeshurun M, et al. Treatment of severe steroid resist-ant acute GVHD with mesenchymal stromal cells (MSC). Am J Blood Res 2013;3:225–238.
3. Higashimoto M, Sakai Y, Takamura M, Usui S, Nasti A, Yoshida K, Seki A, KomuraT, Honda M, Wada T, et al. Adipose tissue derived stromal stem cell therapy inmurine ConA-derived hepatitis is dependent on myeloid-lineage and CD4(1) T-cellsuppression. Eur J Immunol 2013;43:2956–2968.
4. Orozco L, Munar A, Soler R, Alberca M, Soler F, Huguet M, Sent�ıs J, S�anchez A,Garc�ıa-Sancho J. Treatment of knee osteoarthritis with autologous mesenchymalstem cells: A pilot study. Transplantation 2013;12:1535–1541.
5. Friedenstein AJ, Gorskaja JF, Kulagina NN. Fibroblast precursors in normal and irra-diated mouse hematopoietic organs. Exp Hematol 1976;5:267–274.
6. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP,Hedrick MH. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng 2001;2:211–228.
7. Zuk PA, Zhu M, Ashjian P, de Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, FraserJK, Benhaim P, Hedrick MH. Human adipose tissue is a source of multipotent stemcells. Mol Biol Cell 2002;12:4279–4295.
8. Bi Y, Ehirchiou D, Kilts TM, Inkson CA, Embree MC, Sonoyama W, Li L, Leet AI,Seo B-M, Zhang L, et al. Identification of tendon stem/progenitor cells and the roleof the extracellular matrix in their niche. Nat Med 2007;10:1219–1227.
9. K€ogler G, Sensken S, Airey JA, Trapp T, M€uschen M, Feldhahn N, Liedtke S, SorgRV, Fischer J, Rosenbaum C, et al. A new human somatic stem cell from placentalcord blood with intrinsic pluripotent differentiation potential. J Exp Med 2004;2:123–135.
10. Takechi K, Kuwabara Y, Mizuno M. Ultrastructural and immunohistochemical stud-ies of Wharton’s jelly umbilical cord cells. Placenta 1993;2:235–245.
11. Wang H-S, Hung S-C, Peng S-T, Huang C-C, Wei H-M, Guo Y-J, Fu Y-S, Lai M-C,Chen C-C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilicalcord. Stem Cells (Dayton, Ohio) 2004;7:1330–1337.
12. Fern�andez M, Simon V, Herrera G, Cao C, Del Favero H, Minguell JJ. Detection ofstromal cells in peripheral blood progenitor cell collections from breast cancerpatients. Bone Marrow Transplant 1997;4:265–271.
13. Zvaifler NJ, Marinova-Mutafchieva L, Adams G, Edwards CJ, Moss J, Burger JA,Maini RN. Mesenchymal precursor cells in the blood of normal individuals. ArthritisRes 2000;6:477–488.
14. Baksh D, Yao R, Tuan RS. Comparison of proliferative and multilineage differentia-tion potential of human mesenchymal stem cells derived from umbilical cord andbone marrow. Stem Cells (Dayton, Ohio) 2007;6:1384–1392.
15. Ishige I, Nagamura-Inoue T, Honda MJ, Harnprasopwat R, Kido M, Sugimoto M,Nakauchi H, Tojo A. Comparison of mesenchymal stem cells derived from arterial,venous, and Wharton’s jelly explants of human umbilical cord. Int J Hematol 2009;2:261–269.
16. Balasubramanian S, Venugopal P, Sundarraj S, Zakaria Z, Majumdar AS, Ta M.Comparison of chemokine and receptor gene expression between Wharton’s jellyand bone marrow-derived mesenchymal stromal cells. Cytotherapy 2012;1:26–33.
17. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, DeansR, Keating A, Prockop D, Horwitz E. Minimal criteria for defining multipotent mes-enchymal stromal cells. The International Society for Cellular Therapy positionstatement. Cytotherapy 2006;4:315–317.
18. Horwitz E M, Le Blanc K, Dominici M, Mueller I, Slaper-Cortenbach I, Marini FC,Deans RJ, Krause DS, Keating A. Clarification of the nomenclature for MSC: TheInternational Society for Cellular Therapy position statement. Cytotherapy 2005;5:393–395.
19. Ahern BJ, Parvizi J, Boston R, Schaer TP. Preclinical animal models in single site car-tilage defect testing: A systematic review. Osteoarthritis Cartilage 2009;6:705–713.
20. McLean M. Spotlight on: Dr. Lynne Oliver, Office of New Animal Drug Evaluation.Available at: http://www.fda.gov/AnimalVeterinary/NewsEvents/FDAVeterinarian-Newsletter/ucm210093.htm 2011; (20/12/2011).
21. Patterson-Kane JC, Becker DL, Rich T. The pathogenesis of tendon microdamage inathletes: The horse as a natural model for basic cellular research. J Comp Pathol2012;2–3:227–247.
22. Lui PPY, Maffulli N, Rolf C, Smith RKW. What are the validated animal models fortendinopathy? Scand J Med Sci Sports 2011;1:3–17.
23. Smith RKW, Korda M, Blunn GW, Goodship AE. Isolation and implantation ofautologous equine mesenchymal stem cells from bone marrow into the superficialdigital flexor tendon as a potential novel treatment. Equine Vet J 2003;1:99–102.
24. Dowling BA, Dart AJ, Hodgson DR, Smith RK. Superficial digital flexor tendonitisin the hors. Equine Vet J 2000;5:369–378.
25. Dyson SJ. Medical management of superficial digital flexor tendonitis: A comparativestudy in 219 horses (1992-2000). Equine Vet J 2004;5:415–419.
26. Godwin EE, Young NJ, Dudhia J, Beamish IC, Smith RKW. Implantation of bonemarrow-derived mesenchymal stem cells demonstrates improved outcome in horseswith overstrain injury of the superficial digital flexor tendon. Equine Vet J 2012;1:25–32.
27. Nixon AJ, Dahlgren LA, Haupt JL, Yeager AE, Ward DL. Effect of adipose-derivednucleated cell fractions on tendon repair in horses with collagenase-induced tendini-tis. Am J Vet Res 2008;7:928–937.
28. Schnabel LV, Lynch ME, van der Meulen MCH, Yeager AE, Kornatowski MA, NixonAJ. Mesenchymal stem cells and insulin-like growth factor-I gene-enhanced mesen-chymal stem cells improve structural aspects of healing in equine flexor digitorumsuperficialis tendons. J Orthop Res 2009;10:1392–1398.
29. Smith RKW, Werling NJ, Dakin SG, Alam R, Goodship AE, Dudhia J. Beneficialeffects of autologous bone marrow-derived mesenchymal stem cells in naturallyoccurring tendinopathy. PLoS One 2013;9:e75697.
30. Ibrahim S, Saunders K, Kydd JH, Lunn DP, Steinbach F. Screening of anti-humanleukocyte monoclonal antibodies for reactivity with equine leukocytes. Vet ImmunolImmunopathol 2007;1–2:63–80.
31. Braun J, Hack A, Weis-Klemm M, Conrad S, Treml S, Kohler K, Walliser U, SkutellaT, Aicher WK. Evaluation of the osteogenic and chondrogenic differentiation capaci-ties of equine adipose tissue-derived mesenchymal stem cells. Am J Vet Res 2010;10:1228–1236.
32. de Schauwer C, Meyer E, van de Walle G, Soom AV. Markers of stemness inequine mesenchymal stem cells: A plea for uniformity. Theriogenology 2011;75:1431–1443.
33. de Schauwer C, Piepers S, van de Walle GR, Demeyere K, Hoogewijs MK, GovaereJLJ, Braeckmans K, van Soom A, Meyer E. In search for cross-reactivity to immuno-phenotype equine mesenchymal stromal cells by multicolor flow cytometry. Cytome-try Part A 2012;81A:312–323.
34. Seo M-S, Jeong Y-H, Park J-R, Park S-B, Rho K-H, Kim H-S, Yu K-R, Lee S-H, JungJ-W, Lee Y-S, et al. Isolation and characterization of canine umbilical cord blood-derived mesenchymal stem cells. J Vet Sci 2009;3:181–187.
35. Vieira NM, Brandalise V, Zucconi E, Secco M, Strauss BE, Zatz M. Isolation, charac-terization, and differentiation potential of canine adipose-derived stem cells. CellTransplant 2010;3:279–289.
36. Burk J, Ribitsch I, Gittel C, Juelke H, Kasper C, Staszyk C, Brehm W. Growth and dif-ferentiation characteristics of equine mesenchymal stromal cells derived from differ-ent sources. Vet J (London, England: 1997) 2013;1:98–106.
37. de Schauwer C, Meyer E, van de Walle GR, van Soom A. Markers of stemness inequine mesenchymal stem cells: A plea for uniformity. Theriogenology 2011;8:1431–1443.
38. de Mattos Carvalho A, Alves ALG, Golim MA, Moroz A, Hussni CA, de OliveiraPGG, Deffune E. Isolation and immunophenotypic characterization of mesenchymalstem cells derived from equine species adipose tissue. Vet Immunol Immunopathol2009;2–4:303–306.
39. Ranera B, Lyahyai J, Romero A, V�azquez FJ, Remacha AR, Bernal ML, Zaragoza P,Rodellar C, Mart�ın-Burriel I. Immunophenotype and gene expression profiles of cellsurface markers of mesenchymal stem cells derived from equine bone marrow andadipose tissue. Vet Immunol Immunopathol 2011;1–2:147–154.
40. Radcliffe CH, Flaminio MJBF, Fortier LA. Temporal analysis of equine bone marrowaspirate during establishment of putative mesenchymal progenitor cell populations.Stem Cells Dev 2010;2:269–282.
41. Pascucci L, Curina G, Mercati F, Marini C, Dall’Aglio C, Paternesi B, Ceccarelli P.Flow cytometric characterization of culture expanded multipotent mesenchymalstromal cells (MSCs) from horse adipose tissue: Towards the definition of minimalstemness criteria. Vet Immunol Immunopathol 2011;3–4:499–506.
42. Iacono E, Brunori L, Pirrone A, Pagliaro PP, Ricci F, Tazzari PL, Merlo B. Isolation,characterization and differentiation of mesenchymal stem cells from amniotic fluid,umbilical cord blood and Wharton’s jelly in the horse. Reproduction (Cambridge,England) 2012;4:455–468.
43. Corradetti B, Lange-Consiglio A, Barucca M, Cremonesi F, Bizzaro D. Size-sievedsubpopulations of mesenchymal stem cells from intervascular and perivascularequine umbilical cord matrix. Cell Prolif 2011;4:330–342.
44. Hoynowski SM, Fry MM, Gardner BM, Leming MT, Tucker JR, Black L, Sand T,Mitchell KE. Characterization and differentiation of equine umbilical cord-derivedmatrix cells. Biochem Biophys Res Commun 2007;2:347–353.
45. Appleby SL, Cockshell MP, Pippal JB, Thompson EJ, Barrett JM, Tooley K, Sen S,Sun WY, Grose R, Nicholson I, et al. Characterization of a distinct population of cir-culating human non-adherent endothelial forming cells and their recruitment viaintercellular adhesion molecule-3. PLoS One 2012;11:e46996.
46. Vidal MA, Walker NJ, Napoli E, Borjesson DL. Evaluation of senescence in mesen-chymal stem cells isolated from equine bone marrow, #adipose |tissue, and umbilicalcord tissue. Stem Cells Dev 2012;2:273–283.
47. Toupadakis CA, Wong A, Genetos DC, Cheung WK, Borjesson DL, Ferraro GL,Galuppo LD, Leach JK, Owens SD, Yellowley CE. Comparison of the osteogenicpotential of equine mesenchymal stem cells from bone marrow, #adipose|tissue, umbilical cord blood, and umbilical cord tissue. Am J Vet Res 2010;10:1237–1245.
Original Article
686 Comparative Immunophenotyping of Equine MSC

48. Adamzyk C, Emonds T, Falkenstein J, Tolba R, Jahnen-Dechent W, Lethaus B, NeussS. Different culture media affect proliferation, surface epitope expression, and differ-entiation of ovine MSC. Stem Cells Int 2013:387324.
49. Hagmann S, Moradi B, Frank S, Dreher T, K€ammerer PW, Richter W, Gotterbarm T.Different culture media affect growth characteristics, surface marker distributionand chondrogenic differentiation of human bone marrow-derived mesenchymalstromal cells. BMC Musculoskelet Disord 2013;14:223.
50. Panchision DM, Chen H-L, Pistollato F, Papini D, Ni H-T, Hawley TS. Optimizedflow cytometric analysis of central nervous system tissue reveals novel functionalrelationships among cells expressing CD133, CD15, and CD24. Stem Cells (Dayton,Ohio) 2007;6:1560–1570.
51. Takemitsu H, Zhao D, Yamamoto I, Harada Y, Michishita M, Arai T. Comparison ofbone marrow and adipose tissue-derived canine mesenchymal stem cells. BMC VetRes 2012;8:150.
52. Choi S-A, Choi H-S, Kim KJ, Lee D-S, Lee JH, Park JY, Kim EY, Li X, Oh H-Y, LeeD-S, et al. Isolation of canine mesenchymal stem cells from amniotic fluid and differ-entiation into hepatocyte-like cells. In vitro cellular & developmental biology. Ani-mal 2013;1:42–51.
53. Boos AM, Loew JS, Deschler G, Arkudas A, Bleiziffer O, Gulle H, Dragu A, KneserU, Horch RE, Beier JP. Directly auto-transplanted mesenchymal stem cells inducebone formation in a ceramic bone substitute in an ectopic sheep model. J Cell MolMed 2011;6:1364–1378.
54. Vacanti V, Kong E, Suzuki G, Sato K, Canty JM, Lee T. Phenotypic changes of adultporcine mesenchymal stem cells induced by prolonged passaging in culture. J CellPhysiol 2005;2:194–201.
55. Dissanayaka WL, Zhu X, Zhang C, Jin L. Characterization of dental pulp stem cellsisolated from canine premolars. J Endod 2011;8:1074–1080.
56. Mart�ınez-Lorenzo MJ, Royo-Ca~nas M, Alegre-Aguar�on E, Desportes P, Castiella T,Garc�ıa-Alvarez F, Larrad L. Phenotype and chondrogenic differentiation of mesen-chymal cells from adipose tissue of different species. J Orthop Res 2009;11:1499–1507.
57. Tan Q, Lui PP, Rui YF, Wong YM. Comparison of potentials of stem cells isolatedfrom tendon and bone marrow for musculoskeletal tissue engineering. Tissue EngPart A 2012;7–8:840–851.
58. Iacono E, Cunto M, Zambelli D, Ricci F, Tazzari PL, Merlo B. Could fetal fluid andmembranes be an alternative source for mesenchymal stem cells (MSCs) in the felinespecies. A preliminary study. Vet Res Commun 2012;2:107–118.
59. Cardoso TC, Ferrari HF, Garcia AF, Novais JB, Silva-Frade C, Ferrarezi MC, AndradeAL, Gameiro R. Isolation and characterization of Wharton’s jelly-derived multipo-tent mesenchymal stromal cells obtained from bovine umbilical cord and maintainedin a defined serum-free three-dimensional system. BMC Biotechnol 2012:18.
60. Murphy JM, Fink DJ, Hunziker EB, Barry FP. Stem cell therapy in a caprine model ofosteoarthritis. Arthritis Rheum 2003;12:3464–3474.
61. Rostovskaya M, Anastassiadis K. Differential expression of surface markers in mousebone marrow mesenchymal stromal cell subpopulations with distinct lineage com-mitment. PLoS One 2012;12:e51221.
62. Hsiao FSH, Lian WS, Lin SP, Lin CJ, Lin YS, Cheng ECH, Liu CW, Cheng CC,Cheng PH, Ding ST, et al. Toward an ideal animal model to trace donor cell fatesafter stem cell therapy: Production of stably labeled multipotent mesenchymal stemcells from bone marrow of transgenic pigs harboring enhanced green fluorescenceprotein gene. J Anim Sci 2011;11:3460–3472.
63. Noort WA, Oerlemans MIFJ, Rozemuller H, Feyen D, Jaksani S, Stecher D, NaaijkensB, Martens AC, B€uhring HJ, Doevendans PA, et al. Human versus porcine mesenchy-mal stromal cells: Phenotype, #differentiation |potential, immunomodulation andcardiac improvement after transplantation. J Cell Mol Med 2012;8:1827–1839.
64. Zhu X-Y, Urbieta-Caceres V, Krier JD, Textor SC, Lerman A, Lerman LO. Mesenchy-mal stem cells and endothelial progenitor cells decrease renal injury in experimentalswine renal artery stenosis through different mechanisms. Stem Cells (Dayton,Ohio) 2013;1:117–125.
65. Jin HJ, Bae YK, Kim M, Kwon S-J, Jeon HB, Choi SJ, Kim SW, Yang YS, Oh W,Chang JW. Comparative analysis of human mesenchymal stem cells from bone mar-row, adipose tissue, and umbilical cord blood as sources of cell therapy. Int J Mol Sci2013;9:17986–18001.
Original Article
Cytometry Part A � 85A: 678�687, 2014 687
Related Documents




![RESEARCH Open Access Human multipotent stromal cells ...mesenchymal lineage cell types including bone, cartilage, adipose tissue, muscle and tendon [4]. MSCs have been isolated from](https://static.cupdf.com/doc/110x72/5e6b65b920f9b208741edf9a/research-open-access-human-multipotent-stromal-cells-mesenchymal-lineage-cell.jpg)






