162 Environmental Toxicology and Chemistry, Vol. 28, No. 1, pp. 162–172, 2009 2009 SETAC Printed in the USA 0730-7268/09 $12.00 .00 COMPARATIVE HEPATIC ACTIVITY OF XENOBIOTIC-METABOLIZINGENZYMES AND CONCENTRATIONS OF ORGANOHALOGENS AND THEIR HYDROXYLATED ANALOGUES IN CAPTIVE GREENLAND SLEDGE DOGS (CANIS FAMILIARIS) JONATHAN VERREAULT,†‡ FRANCE MAISONNEUVE,† RUNE DIETZ,§ CHRISTIAN SONNE,§ and ROBERT J. LETCHER*†‡ †Wildlife and Landscape Science Directorate, Science and Technology Branch, Environment Canada, National Wildlife Research Centre, Carleton University, Ottawa, Ontario K1A 0H3 ‡Carleton University, Department of Chemistry, Ottawa, Ontario K1S 5B6, Canada §University of Aarhus, National Environmental Research Institute, Department of Arctic Environment, DK-4000 Roskilde, Denmark ( Received 14 April 2008; Accepted 17 July 2008) Abstract—A captive study was performed with Greenland sledge dogs (Canis familiaris) fed a naturally organohalogen-contam- inated diet (Greenland minke whale [Balaenoptera acutorostrata] blubber; exposed group) or a control diet (pork fat; control group). The catalytic activity of major xenobiotic-metabolizing phase I and II hepatic microsomal enzymes was assessed. Relative to control dogs, ethoxyresorufin-O-deethylase (EROD) activity in exposed dogs was twofold higher ( p 0.001). Testosterone hydroxylation yielded 6- and 16-hydroxy (OH) testosterone and androstenedione, with higher rates of production (23–27%; p 0.03) in the exposed individuals. In the exposed dogs, epoxide hydrolase (EH) activity was 31% higher ( p 0.02) relative to the control dogs, whereas uridine diphosphoglucuronosyl transferase (UDPGT) activity was not different ( p 0.62). When the exposed and control dogs were combined, the summed () plasma concentrations of OH-polychlorinated biphenyl (PCB) congeners were predicted by plasma PCB concentrations and EROD activity ( p 0.04), whereas testosterone hydroxylase, EH, and UDPGT activities were not significant predictors of these concentrations. Consistent results were found for individual OH-PCB congeners and their theoretical precursor PCBs (e.g., 4-OH-CB-187 and CB-183, and 4-OH-CB-146 and CB-146) and for EROD activity. No association was found between OH–polybrominated diphenyl ether (PBDE) and PBDE plasma concentrations, or between potential precursor-metabolite pairs, and the enzyme activities. The present results suggest that liver microsomal EROD activity and plasma PCB concentrations have a greater (e.g., relative to EH activity) predictive power for the occurrence of plasma OH- PCB residues in sledge dogs. These results also suggest that plasma OH-PBDEs likely are not products of cytochrome P450– mediated transformation but, rather, are accumulated via the diet. Keywords—Biotransformation enzymes Sledge dog Arctic Organohalogens Hydroxylated metabolites INTRODUCTION The tissue-specific content, substrate specificity, and cata- lytic activity of xenobiotic-metabolizing enzymes are key char- acteristics influencing the toxicokinetics and fate of organo- halogen contaminants (OHCs). In mammals, the cytochrome P450 (CYP) monooxygenase isoforms (i.e., mainly subfami- lies 1–3) play a central role in the phase I, oxidative biotrans- formation of a broad variety of structurally diverse, chlorinated and brominated OHCs [1–3]. Two persistent and bioaccu- mulative OHC classes widely detected in mammalian wildlife, polychlorinated biphenyls (PCBs) and polybrominated diphe- nyl ether (PBDE) flame retardants, undergo oxidative metab- olism via catalytic mediation by specific CYP isoenzymes [1– 3]. The oxidative metabolism of PCBs and PBDEs can lead to the formation of potentially more toxic metabolites—namely, the hydroxylated (OH)-PCBs and OH-PBDEs [1–3]. The for- mation of OH-PCBs and OH-PBDEs has been demonstrated to occur either through direct insertion of a OH-group or via formation of an arene oxide intermediate and subsequent ep- oxide hydrolase (EH)–mediated ring opening (i.e., hydrolysis) with or without a 1,2-chlorine/bromine atom shift [1,3]. In * To whom correspondence may be addressed ([email protected]). Published on the Web 9/8/2008. addition to having a metabolic origin, however, certain OH- PBDE congeners containing OH-substitution on a phenyl ring carbon that is ortho to the ether linkage have been isolated in marine sponges, ascidians, and algae [4]. Recently, a number of these OH-PBDE congeners were confirmed to be naturally occurring, dietary-sourced compounds accumulated in the blubber of True’s beaked whales (Mesoplodon mirus) [4]. The presence of the OH-moiety on OH-PCBs and OH-PBDEs im- parts a greater electronic polarity relative to their PCB/PBDE analogues. This imparts increased water solubility and facil- itates subsequent excretion from the organism. The OH-con- taining compounds, however, can be further biotransformed by phase I and/or II enzyme catalysis, although their subse- quent metabolic transformation may be limited as a result of binding to circulating proteins (e.g., transthyretin). One major phase II reaction—glucuronidation—can conjugate glucuronic acid to the reactive OH-group of these molecules. This may occur via catalysis by uridine diphosphoglucuronosyl trans- ferase (UDPGT), leading to the formation of more hydrophilic and readily excretable end products in urine or bile [5]. The OH-PCBs and OH-PBDEs have been reported in a number of mammalian species (mainly in blood), especially those occupying top trophic positions in their respective ma- rine food webs [6,7], including Arctic species such as the polar bear (Ursus maritimus) [8,9]. In polar bears, for example, OH- PCBs were quantified in blood at concentrations several-fold

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

162
Environmental Toxicology and Chemistry, Vol. 28, No. 1, pp. 162–172, 2009� 2009 SETAC
Printed in the USA0730-7268/09 $12.00 � .00
COMPARATIVE HEPATIC ACTIVITY OF XENOBIOTIC-METABOLIZING ENZYMESAND CONCENTRATIONS OF ORGANOHALOGENS AND THEIR HYDROXYLATED
ANALOGUES IN CAPTIVE GREENLAND SLEDGE DOGS (CANIS FAMILIARIS)
JONATHAN VERREAULT,†‡ FRANCE MAISONNEUVE,† RUNE DIETZ,§ CHRISTIAN SONNE,§ andROBERT J. LETCHER*†‡
†Wildlife and Landscape Science Directorate, Science and Technology Branch, Environment Canada,National Wildlife Research Centre, Carleton University, Ottawa, Ontario K1A 0H3‡Carleton University, Department of Chemistry, Ottawa, Ontario K1S 5B6, Canada
§University of Aarhus, National Environmental Research Institute, Department of Arctic Environment, DK-4000 Roskilde, Denmark
(Received 14 April 2008; Accepted 17 July 2008)
Abstract—A captive study was performed with Greenland sledge dogs (Canis familiaris) fed a naturally organohalogen-contam-inated diet (Greenland minke whale [Balaenoptera acutorostrata] blubber; exposed group) or a control diet (pork fat; controlgroup). The catalytic activity of major xenobiotic-metabolizing phase I and II hepatic microsomal enzymes was assessed. Relativeto control dogs, ethoxyresorufin-O-deethylase (EROD) activity in exposed dogs was twofold higher ( p � 0.001). Testosteronehydroxylation yielded 6�- and 16�-hydroxy (OH) testosterone and androstenedione, with higher rates of production (23–27%; p� 0.03) in the exposed individuals. In the exposed dogs, epoxide hydrolase (EH) activity was 31% higher ( p � 0.02) relative tothe control dogs, whereas uridine diphosphoglucuronosyl transferase (UDPGT) activity was not different ( p � 0.62). When theexposed and control dogs were combined, the summed (�) plasma concentrations of OH-polychlorinated biphenyl (PCB) congenerswere predicted by plasma �PCB concentrations and EROD activity ( p � 0.04), whereas testosterone hydroxylase, EH, and UDPGTactivities were not significant predictors of these concentrations. Consistent results were found for individual OH-PCB congenersand their theoretical precursor PCBs (e.g., 4-OH-CB-187 and CB-183, and 4-OH-CB-146 and CB-146) and for EROD activity. Noassociation was found between �OH–polybrominated diphenyl ether (PBDE) and �PBDE plasma concentrations, or betweenpotential precursor-metabolite pairs, and the enzyme activities. The present results suggest that liver microsomal EROD activityand plasma PCB concentrations have a greater (e.g., relative to EH activity) predictive power for the occurrence of plasma OH-PCB residues in sledge dogs. These results also suggest that plasma OH-PBDEs likely are not products of cytochrome P450–mediated transformation but, rather, are accumulated via the diet.
Keywords—Biotransformation enzymes Sledge dog Arctic Organohalogens Hydroxylated metabolites
INTRODUCTION
The tissue-specific content, substrate specificity, and cata-lytic activity of xenobiotic-metabolizing enzymes are key char-acteristics influencing the toxicokinetics and fate of organo-halogen contaminants (OHCs). In mammals, the cytochromeP450 (CYP) monooxygenase isoforms (i.e., mainly subfami-lies 1–3) play a central role in the phase I, oxidative biotrans-formation of a broad variety of structurally diverse, chlorinatedand brominated OHCs [1–3]. Two persistent and bioaccu-mulative OHC classes widely detected in mammalian wildlife,polychlorinated biphenyls (PCBs) and polybrominated diphe-nyl ether (PBDE) flame retardants, undergo oxidative metab-olism via catalytic mediation by specific CYP isoenzymes [1–3].
The oxidative metabolism of PCBs and PBDEs can lead tothe formation of potentially more toxic metabolites—namely,the hydroxylated (OH)-PCBs and OH-PBDEs [1–3]. The for-mation of OH-PCBs and OH-PBDEs has been demonstratedto occur either through direct insertion of a OH-group or viaformation of an arene oxide intermediate and subsequent ep-oxide hydrolase (EH)–mediated ring opening (i.e., hydrolysis)with or without a 1,2-chlorine/bromine atom shift [1,3]. In
* To whom correspondence may be addressed([email protected]).
Published on the Web 9/8/2008.
addition to having a metabolic origin, however, certain OH-PBDE congeners containing OH-substitution on a phenyl ringcarbon that is ortho to the ether linkage have been isolated inmarine sponges, ascidians, and algae [4]. Recently, a numberof these OH-PBDE congeners were confirmed to be naturallyoccurring, dietary-sourced compounds accumulated in theblubber of True’s beaked whales (Mesoplodon mirus) [4]. Thepresence of the OH-moiety on OH-PCBs and OH-PBDEs im-parts a greater electronic polarity relative to their PCB/PBDEanalogues. This imparts increased water solubility and facil-itates subsequent excretion from the organism. The OH-con-taining compounds, however, can be further biotransformedby phase I and/or II enzyme catalysis, although their subse-quent metabolic transformation may be limited as a result ofbinding to circulating proteins (e.g., transthyretin). One majorphase II reaction—glucuronidation—can conjugate glucuronicacid to the reactive OH-group of these molecules. This mayoccur via catalysis by uridine diphosphoglucuronosyl trans-ferase (UDPGT), leading to the formation of more hydrophilicand readily excretable end products in urine or bile [5].
The OH-PCBs and OH-PBDEs have been reported in anumber of mammalian species (mainly in blood), especiallythose occupying top trophic positions in their respective ma-rine food webs [6,7], including Arctic species such as the polarbear (Ursus maritimus) [8,9]. In polar bears, for example, OH-PCBs were quantified in blood at concentrations several-fold

Enzyme activity and organohalogens in sledge dogs Environ. Toxicol. Chem. 28, 2009 163
Table 1. Sample size (sex- and cohort-categorized) and mean (� standard error) age, body mass, plasma lipid content, and plasma organohalogen(OHC) concentrations in control (CON) and exposed (EXP) adult sledge dogs (i.e., parents [P] and their first-generation adult male [M] and
female [F] pups [F1])a
CON diet(pork fat)
CON sledge dogs(plasma)
EXP diet(Greenland minke
whale blubber)EXP sledge dogs
(plasma)
Sample size 3 (pooled samples) P: 7 F 3 (pooled samples) P: 7 FF1: 2 F and 3 M F1: 1 F and 2 M
Age (d) —b 455 � 33 —b 469 � 38Body mass (kg) — 23.4 � 0.9 — 24.1 � 0.8Lipid content (%) 96.8 � 1.7 0.76 � 0.03 67.5 � 2.4 0.67 � 0.05
Concentrations (ng/g wet wt.)�PBDEc,* 2.1 � 0.4 0.08 � 0.02 41.9 � 0.7 0.26 � 0.04�PCBd,* ND 1.01 � 0.27 1,147 � 58.8 11.9 � 1.35�OH-PBDEe,* ND 0.51 � 0.17 3.08 � 0.99f 1.46 � 0.31�OH-PCBg,* ND 48.1 � 8.5 ND 224 � 13.4
a Lipid content and OHC concentrations also are listed for the CON and EXP diets (pork fat and Greenland minke whale blubber, respectively).Partial data as reported in Verreault et al. [15,16]. Significant differences ( p � 0.05) between the CON and EXP sledge dog groups are indicatedwith an asterisk. ND � not detected.
b The age and body mass of the pork (fat) and minke whale (blubber) are unknown.c �PBDE � sum of 13 polybrominated diphenyl ether (PBDE) congeners: BDEs 17, 28, 47, 49, 66, 85, 99, 100, 138, 153, 154, 183, and 209.d �PCB � sum of 60 polychlorinated biphenyl (PCB) congeners: CBs 16/32, 17, 18, 22, 31/28, 33/20, 42, 44, 47/48, 49, 52, 56/60, 64/41, 66,
70/76, 74, 85, 87, 92, 95, 97, 99, 101/90, 105, 110, 114, 118, 128, 130, 137, 138, 141, 146, 149, 151, 153, 156, 157, 158, 170/190, 171, 172,174, 176, 177, 178, 179, 180, 183, 187, 189, 194, 195, 196/203, 199, 200, 202, 206, 207, and 208.
e �OH-PBDE � sum of 14 hydroxylated (OH) PBDE congeners: 6�-OH-BDE-17, 4�-OH-BDE-17, 4-OH-BDE-42, 6-OH-BDE-47, 3-OH-BDE-47, 5-OH-BDE-47, 6�-OH-BDE-49, 4�-OH-BDE-49, 2�-OH-BDE-68, 6-OH-BDE-85, 6-OH-BDE-90, 6-OH-BDE-99, 2-OH-BDE-123, and 6-OH-BDE-137.
f Comprised exclusively of the congener 6-OH-BDE-47.g �OH-PCB � sum of 32 OH-PCB congeners: 4�-OH-CB-79, 4-OH-CB-97, 4�-OH-CB-101, 4-OH-CB-134, 4-OH-CB-107/4�-OH-CB-108, 2�-
OH-CB-114, 3-OH-CB-118, 4�-OH-CB-127, 4�-OH-CB-120, 4�-OH-CB-130, 3�-OH-CB-138, 4-OH-CB-146, 4�-OH-CB-159, 4-OH-CB-162, 4-OH-CB-163, 4�-OH-CB-172, 4�-OH-CB-177, 4-OH-CB-178, 3�-OH-CB-180, 3�-OH-CB-182, 3�-OH-CB-183, 3�-OH-CB-184, 4-OH-CB-187,4-OH-CB-193, 4�-OH-CB-198, 4�-OH-CB-199, 4�-OH-CB-200, 4�-OH-CB-201, 4�-OH-CB-202, 4,4�-di-OH-CB-202, 3�-OH-CB-203, and 4�-OH-CB-208.
higher relative to those in liver, adipose, and brain tissue andat concentrations far greater than those of their theoreticalprecursor PCB congeners. This demonstrates the unique ca-pacity of these carnivorous, top Arctic predators to biotrans-form PCBs and/or retain OH-PCBs in blood. For comparison,in the same polar bears, the OH-PBDE congeners monitoredin blood, liver, brain, and adipose tissue samples have beendetected at concentrations substantially lower than those oftheir PBDE analogues and considerably lower than those ofOH-PCBs.
Recently, in a semifield study, captive sledge dogs (Canisfamiliaris), also known as sleigh dogs or sled dogs, fed anaturally organohalogen-contaminated diet consisting of theblubber of a Greenland minke whale (Balaenoptera acuto-rostrata) have been examined as a potential top carnivoresurrogate Canoidea species for the Greenland polar bear inthe study of exposure-associated effects [10–14] and fate andtoxicokinetics of OHCs [15,16]. Verreault et al. [15], however,reported differences in the plasma congener composition andconcentrations of PCBs and PBDEs, as well as those of theirOH-containing analogues, between sledge dogs and polar bearsfrom Greenland. These differences in the fate and toxicoki-netics of PCBs and PBDEs were hypothesized to be attributed,in part, to species-specific disparities in phase I and II xeno-biotic-metabolizing enzyme content, catalytic activity, andspecificity for congeners of these persistent and ubiquitouschemical classes. Hence, this could influence in turn OH-PCBand OH-PBDE formation and retention in their circulatorysystem. Studies with polar bears from the Canadian Arctichave reported high CYP1A isoenzyme induction based on liverprotein content (determined immunoquantitatively) and mi-crosomal ethoxyresorufin-O-deethylase (EROD) activity [17–
19]. Regardless, there exist major gaps in our knowledge per-taining to the link between PCB and PBDE concentrations,OH-PCB and OH-PBDE metabolite formation and retention,and liver microsomal enzyme activities in top Arctic carnivoresof the Canoidea superfamily, including the polar bear, Arcticfox (Alopex lagopus), and Arctic wolf (Canis lupus arctos).
The goal of the present study was to examine, using cat-alytic enzyme assays, the comparative activity of major phaseI and II xenobiotic-metabolizing enzymes (CYP-like, EH, andUGPGT) in liver microsomes of captive adult female sledgedogs and their first-generation male and female adult pups fedeither Greenland minke whale blubber or a control diet con-sisting of pork fat. Furthermore, in these sledge dog cohorts,the correlative associations between catalytic enzyme activitiesand blood (plasma) concentrations of PCBs and PBDEs andtheir OH-PCB and OH-PBDE analogues were examined. Over-all, the present study investigated the inferred role of thesexenobiotic-metabolizing enzyme systems on the fate and tox-icokinetics of PCBs and PBDEs in connection with apparentOH-PCB and OH-PBDE metabolites in the captive sledge dog,with possible extrapolation to other members of the Canoideasuperfamily, such as the polar bear, Arctic fox, and Arcticwolf.
MATERIALS AND METHODS
Sample collection
Two-month-old, female sledge dogs (P cohort) were ob-tained in the community of Aasiaat (Disco Bay, West Green-land) and assigned to two experimental groups: An exposed(EXP) group, and a control (CON) group (Table 1). The CONand EXP groups were composed of paired sisters, with one

164 Environ. Toxicol. Chem. 28, 2009 J. Verreault et al.
sister in each group, to minimize genetic differences betweengroups. Over the nearly two-year study duration (90–94 weeksin 2003–2005), one and two of the EXP and CON P cohortdogs, respectively, were mated by one single male, and theirfirst-generation male and female pups (F1 cohort) were inte-grated in the feeding experiment (last 48–49 weeks of thestudy). The CON and EXP pups were all sexually mature atthe end of the study. The EXP P and F1 cohorts were fed withnaturally OHC-contaminated minke whale blubber [20,21] har-vested as part of the Greenlandic native subsistence hunt,whereas the CON cohorts were given commercially availablepork fat (Blume, Randers, Denmark). The daily blubber/fatfeeding regime was adjusted each week based on the individ-ual’s body mass (average ration, 112 g/d) and was supple-mented with standardized Royal Canin (Randers, Denmark)Energy 4300 and 4800 diets (http://www.royalcanin.com/) tocover the essential nutritional needs of dogs living in coldregions. Details regarding the diet composition (e.g., fatty ac-ids, vitamins, and microelements) can be found in Sonne etal. [22] (http://www.ncbi.nlm.nih.gov/pubmed/18342937).The dogs were exercised on a daily basis, maintained in con-trolled and healthy body condition, and otherwise kept sepa-rated in chains to control the food intake. The dogs also weresubjected to various treatments during the study period (i.e.,vaccination and immune challenge [intradermal test]) as de-scribed elsewhere [10] and were humanely killed on comple-tion of the experiment by physical means (shooting). Blood(plasma) was collected from each individual and kept at �20Cor lower until chemically analyzed. Liver samples from thesame lobe were excised within 5 min after death, perfused toremove blood, rinsed in a NaCl/distilled H2O solution, andstored immediately in liquid nitrogen until microsome prep-aration. The sledge dog experimental design was performedunder a license granted by the Official Chief Veterinarian fromthe Home Rule Government in Greenland (Community ofNuuk, 2003).
Liver microsome preparation and total proteindetermination
Liver microsomes were prepared by differential ultracen-trifugation according to procedures described elsewhere [23].Briefly, liver tissues (9–11 g) were thawed on ice and minced,after which an aliquot of approximately 1.0 g was added to 4ml of cold phosphate buffer (0.1 M KH2PO4 and 0.1 MNa2HPO4, pH 7.4; Fisher Scientific, Ottawa, ON, Canada) andhomogenized with an Ultra-Turrex. The homogenate was cen-trifuged at 9,000 g (15 min, 4C), and 2 ml of the supernatant(S-9) were centrifuged at 100,000 g (60 min, 4C). The pelletwas resuspended in 2 ml of cold phosphate buffer and storedin liquid nitrogen until determination of catalytic assay activ-ity. The total microsomal protein concentration was deter-mined in triplicates using the spectrophotometric method de-scribed by Kennedy and Jones [23] using bovine serum al-bumin (Sigma-Aldrich, Oakville, ON, Canada) as the standard(0.0–500 g/ml). Fluorescence was measured using a multi-well plate reader (Cytofluor model 2350; Millipore, Etobicoke,ON, Canada).
Ethoxyresorufin-O-deethylase activity
Hepatic microsomal EROD activity (analyzed in triplicates)was determined according to methods described previously byKennedy and Jones [23]. The only modification to this methodwas that the incubation time was set to 3 min (37C, pH 8.0),
because EROD activity was found to be linear for up to 7 to10 min in the lower activity range (�150 pmol/min/mg pro-tein) and for up to 5 to 7 min in the higher activity range(300–400 pmol/min/mg protein). The percentages of variationbetween the triplicate samples were all less than 7.8%. A com-mercially available reference material consisting of an adultfemale beagle dog liver microsome pool (n � 3 dogs; BDBiosciences, Woburn, MA, USA) was assayed along with eachset of sledge dog samples to monitor for precision and re-peatability (coefficient of variation [CV], 3.5%; n � 15).
Testosterone hydroxylase activity
The determination of testosterone hydroxylase (TH) activ-ity in liver microsomes was based on procedures describedelsewhere [24,25]. The 2�-, 6�-, 6�-, 7�-, 11�-, 15�-, 16�-,and 16�-OH-testosterone, androstenedione, and testosteronestandards were obtained from Steraloids (Wilton, NH, USA).The stock solutions were prepared in methanol (high-perfor-mance liquid chromatograph [HPLC] grade). Briefly, the re-action mixture (analyzed in duplicates), which included 0.75mg of liver microsomal protein (final concentration, 1 mg/ml),100 M testosterone, and 20 l (5 g) of the internal standard6�-OH-testosterone (not detected in sledge dog liver micro-somes) in 750 l of 0.1 M Tris buffer (pH 7.4), was prein-cubated for 5 min (37C). The reaction was initiated by theaddition of 100 l of the reduced form of nicotinamide adeninedinucleotide phosphate (final concentration, 1 mM; Sigma-Aldrich) and terminated after 30 min by the addition of 750l of ice-cold methanol. The microsomal proteins were re-moved from the suspension by centrifugation (5,500 g, 10min). A 1-ml aliquot of the supernatant was filtered (meshsize, 0.2 m), and 50 l were injected directly onto a HPLC.The HPLC system consisted of a Waters Alliance 2795 sep-aration module (Milford, MA, USA) equipped with a tem-perature-controlled column holder and autosampler and a Wa-ters 2996 photodiode-array (PDA) detector set at 241 nm. Theanalytical column was a Hypersil BDS (C18; length, 150 mm;inner diameter, 4.6; film thickness, 3 m; Thermo Fisher Sci-entific, Waltham, MA, USA). The column was maintained at40C and the autosampler chamber at 25C. The flow rate wasconstant at 0.8 ml/min. The TH activity was determined basedon the peak areas of the OH-testosterone metabolites and an-drostenedione. The instrumental detection limits of 6�-,16�-, and 16�-OH-testosterone and androstenedione were 16,8, 8, and 9 pmol, respectively, per 50-l injection. Recoveryof the 6�-OH-testosterone internal standards was 95% or great-er for all samples. The percentages of variation between theduplicate samples were all less than 9.5%. The CV of THactivity in the reference female beagle dog liver microsomepool (BD Biosciences) ranged between 0.9 and 6.0% for6�-, 16�-, and 16�-OH-testosterone and androstenedione (n� 5 for each product).
Epoxide hydrolase activity
The EH activity in liver microsomes was determined basedon the methods described by Eaton and Stapleton [26]. Briefly,the reaction mixture (analyzed in duplicates), comprised of100 g of liver microsomal protein in 500 l of 0.1 M Trisbuffer (pH 7.4), was preincubated for 5 min (37C), and thereaction was initiated by the addition of 25 l of 4,5-ben-zo[a]pyrene (BaP) epoxide (0.35 mg/ml; catalog no. L0622;National Cancer Institute [NCI] Chemical Carcinogen Refer-ence Standard Repository, Midwest Research Institute, Kansas

Enzyme activity and organohalogens in sledge dogs Environ. Toxicol. Chem. 28, 2009 165
City, MO, USA). The reaction was terminated after 10 minby the addition of 500 l of ice-cold acetonitrile. The micro-somal proteins were removed from the suspension by centri-fugation (5,500 g, 5 min). The supernatant was filtered (meshsize, 0.2 m), and 50 l were injected directly onto a HPLC.The HPLC system consisted of a Waters Alliance 2795 sep-aration module equipped with a temperature-controlled columnholder and autosampler, a Waters 2996 PDA ultraviolet/visiblewavelength detector, and a PerkinElmer (Waltham, MA, USA)LC240 fluorescence detector. The PDA ultraviolet/visiblewavelength detector was set at 260 nm, and the fluorescencedetector was set at an excitation wavelength of 241 nm andan emission wavelength of 389 nm. Because of its high sen-sitivity, the fluorescence detector was used to confirm the iden-tity of the peaks, whereas the determination of EH activitywas done using the ultraviolet/visible peak area response (ab-sorption wavelength, 260 nm) of the 4,5-BaP dihydrodiol (cat-alog no. L0113; NCI Chemical Carcinogen Reference StandardRepository, Midwest Research Institute). The analytical col-umn (Ultracarb 5 octadecylsilane; length, 100 mm; inner di-ameter, 4.6 mm; film thickness, 0.25 mm; Phenomenex, Tor-rance, CA, USA) was maintained at 30C and the autosamplerchamber at 25C. The flow rate was constant at 1.0 ml/min.The instrumental detection limit of 4,5-BaP dihydrodiol was4 pmol per 50-l injection. The percentages of variation be-tween the duplicate samples were all 9.0% or less. The CV ofEH activity in the reference female beagle dog liver microsomepool (BD Biosciences) was 3.5% (n � 3).
Uridine diphosphate glucuronyltransferase activity
The UDPGT activity (analyzed in triplicates) in liver mi-crosomes was determined based on procedures described pre-viously by Collier et al. [27]. In brief, 96-well microplatescontaining 30 l of diluted liver microsomes (final concen-tration, 30–90 g protein/well), 105 l of 4-methylumbelli-ferone (4-MU; final concentration: 1000 M; Fluka catalogno. 69580; Sigma-Aldrich) in 0.1 M Tris buffer (pH 7.4) con-taining 5 mM MgCl2 (Sigma-Aldrich) and 0.05% bovine serumalbumin (Sigma-Aldrich) were preincubated for 5 min (37C).The reaction was initiated by the addition of 15 l of uridine5�-diphosphoglucuronic acid (final concentration, 2 mM; cat-alog no. 197U6751; Sigma-Aldrich). The final volume was150 l/well. The plates were read on a SpectraMax Geminimicroplate spectrofluorometer (Molecular Devices, Sunnyvale,CA, USA) set at an excitation wavelength of 360 nm and anemission wavelength of 455 nm. The lag time was 5 min, andthe kinetic activity was measured over a 10-min period at 37C.The maximum reaction rate (Vmax) used for the determinationof UDPGT activity was calculated between 5 and 15 min (r2
� 0.90). The instrumental detection limit of 4-MU was 10M. The percentages of variation between the triplicate sam-ples were all 15.0% or less. The CV of UDPGT activity inthe reference female beagle dog liver microsome pool (BDBiosciences) was 6.0% (n � 7).
Chemical analysis
The analytical procedures, including sample lipid deter-mination, extraction, partitioning, and cleanup, as well as in-strumental quantification, quality-assurance/quality-controlparameters, and method limits of quantification for the deter-mination of PCBs, PBDEs, OH-PCBs, and OH-PBDEs in plas-ma of sledge dogs, have been comprehensively described byVerreault et al. [15]. A complete list of the PCB, PBDE, OH-
PCB, and OH-PBDE congeners determined in the presentstudy can be found in Table 1. The PCB and PBDE congenersare numbered according to the system proposed by the Inter-national Union for Pure and Applied Chemistry, whereas theOH-PCBs and OH-PBDEs are named based on the nomen-clature revised by Maervoet et al. [28].
Data treatment
The relationships among liver microsomal phase I and IIenzyme activities, plasma concentrations (wet-wt basis) ofPCBs, PBDEs, OH-PCBs, and OH-PBDEs, and potential con-founding factors (age, body mass, and plasma lipid percent-ages) for the combined sledge dog cohorts were investigatedusing principal component analysis (PCA) on the correlationmatrix. The PCA was conducted using either the sums (i.e.,�PCB and �OH-PCB, or �PBDE and �OH-PBDE) of all thecongeners determined (Table 1) or specific congener pairs con-sisting of individual PCBs or PBDEs and their theoretical OH-metabolites as reported elsewhere [1–3]. For these individualcongener analyses, however, only the congeners detected in60% or more of the plasma samples were used. For thesecongeners, the samples with concentrations of a given con-gener below the method limit of quantification were arbitrarilyassigned a randomly generated value between zero and thecompound-specific method limit of quantification (detailed inVerreault et al. [15]). The random values that were obtainedcomplied with the criterion of normal distribution and, thus,did not introduce biased variance in the dataset. The principalcomponents (PCs) with eigenvalues above one were consid-ered to account for a significant contribution to the total var-iance according to the latent root criterion [29]. Furthermore,to investigate the strength and significance of these PCA re-lationships, general linear models (GLMs; e.g., analysis ofvariance and Fisher post-hoc test) were employed. The cor-relations between two variables were expressed using the Pear-son coefficient (r). The enzyme activities, plasma OHC con-centrations, age, body mass, and lipid percentages were log10-transformed before the analyses to achieve or approach thenormal distribution. The statistical package used was Statis-tica� (StatSoft, Tulsa, OK, USA), and the level of significancewas set at 0.05.
RESULTS
Hepatic microsomal enzyme activity
Comparable phase I and II enzyme activities were measuredbetween the P and F1 sledge dog cohorts ( p � 0.12) withineach of the EXP and CON dietary groups. Concurrently, nodifference in enzyme activity ( p � 0.37) was found betweenmales and females within the CON F1 cohort, which was theonly cohort for which a male–female comparison could beperformed. Therefore, the P and F1 dogs (including both sexes)were merged into a single cohort for each of the EXP andCON groups for comparisons of enzyme activities (Fig. 1).
Ethoxyresorufin-O-deethylase activity
The EROD activity was nearly twofold higher ( p � 0.001)in the EXP sledge dogs (mean � standard error [SE], 265 �36 pmol/min/mg protein) compared to the CON sledge dogs(mean � SE, 138 � 16 pmol/min/mg protein) (Fig. 1). Amongthe phase I and II catalytic rates determined, EROD yieldedthe largest difference in activity between these two dietarygroups. The EROD activity in the liver microsome pool of
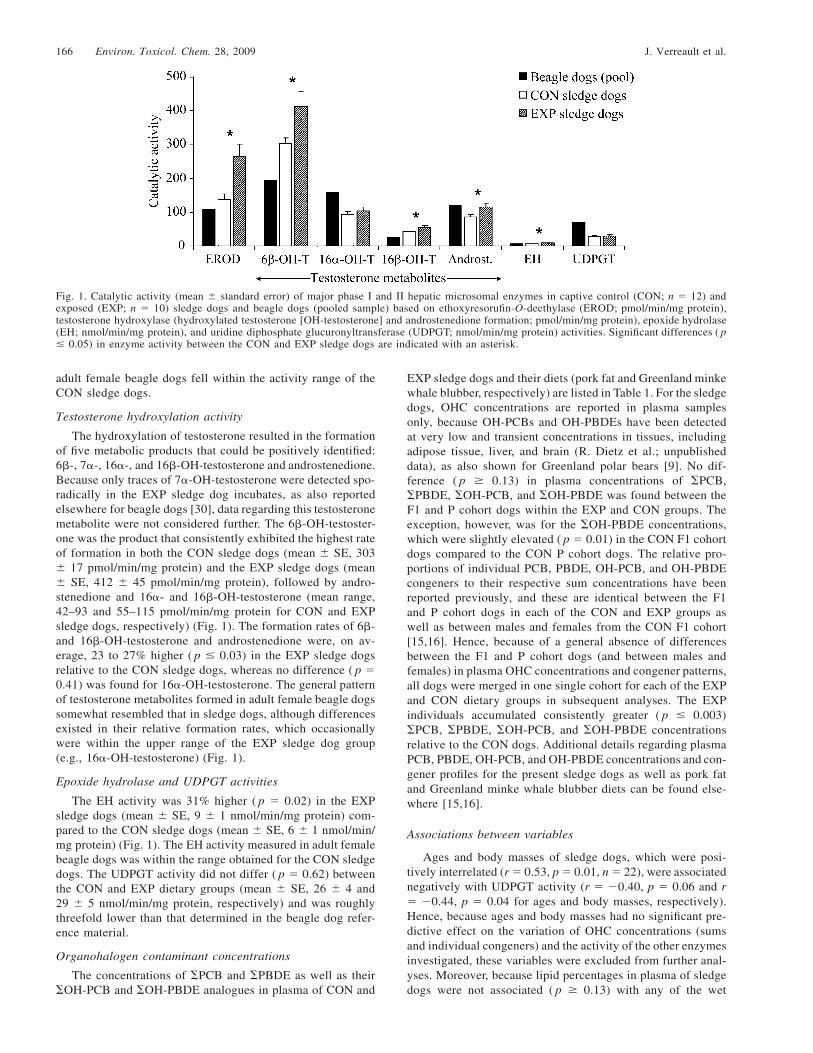
166 Environ. Toxicol. Chem. 28, 2009 J. Verreault et al.
Fig. 1. Catalytic activity (mean � standard error) of major phase I and II hepatic microsomal enzymes in captive control (CON; n � 12) andexposed (EXP; n � 10) sledge dogs and beagle dogs (pooled sample) based on ethoxyresorufin-O-deethylase (EROD; pmol/min/mg protein),testosterone hydroxylase (hydroxylated testosterone [OH-testosterone] and androstenedione formation; pmol/min/mg protein), epoxide hydrolase(EH; nmol/min/mg protein), and uridine diphosphate glucuronyltransferase (UDPGT; nmol/min/mg protein) activities. Significant differences ( p� 0.05) in enzyme activity between the CON and EXP sledge dogs are indicated with an asterisk.
adult female beagle dogs fell within the activity range of theCON sledge dogs.
Testosterone hydroxylation activity
The hydroxylation of testosterone resulted in the formationof five metabolic products that could be positively identified:6�-, 7�-, 16�-, and 16�-OH-testosterone and androstenedione.Because only traces of 7�-OH-testosterone were detected spo-radically in the EXP sledge dog incubates, as also reportedelsewhere for beagle dogs [30], data regarding this testosteronemetabolite were not considered further. The 6�-OH-testoster-one was the product that consistently exhibited the highest rateof formation in both the CON sledge dogs (mean � SE, 303� 17 pmol/min/mg protein) and the EXP sledge dogs (mean� SE, 412 � 45 pmol/min/mg protein), followed by andro-stenedione and 16�- and 16�-OH-testosterone (mean range,42–93 and 55–115 pmol/min/mg protein for CON and EXPsledge dogs, respectively) (Fig. 1). The formation rates of 6�-and 16�-OH-testosterone and androstenedione were, on av-erage, 23 to 27% higher ( p � 0.03) in the EXP sledge dogsrelative to the CON sledge dogs, whereas no difference ( p �0.41) was found for 16�-OH-testosterone. The general patternof testosterone metabolites formed in adult female beagle dogssomewhat resembled that in sledge dogs, although differencesexisted in their relative formation rates, which occasionallywere within the upper range of the EXP sledge dog group(e.g., 16�-OH-testosterone) (Fig. 1).
Epoxide hydrolase and UDPGT activities
The EH activity was 31% higher ( p � 0.02) in the EXPsledge dogs (mean � SE, 9 � 1 nmol/min/mg protein) com-pared to the CON sledge dogs (mean � SE, 6 � 1 nmol/min/mg protein) (Fig. 1). The EH activity measured in adult femalebeagle dogs was within the range obtained for the CON sledgedogs. The UDPGT activity did not differ ( p � 0.62) betweenthe CON and EXP dietary groups (mean � SE, 26 � 4 and29 � 5 nmol/min/mg protein, respectively) and was roughlythreefold lower than that determined in the beagle dog refer-ence material.
Organohalogen contaminant concentrations
The concentrations of �PCB and �PBDE as well as their�OH-PCB and �OH-PBDE analogues in plasma of CON and
EXP sledge dogs and their diets (pork fat and Greenland minkewhale blubber, respectively) are listed in Table 1. For the sledgedogs, OHC concentrations are reported in plasma samplesonly, because OH-PCBs and OH-PBDEs have been detectedat very low and transient concentrations in tissues, includingadipose tissue, liver, and brain (R. Dietz et al.; unpublisheddata), as also shown for Greenland polar bears [9]. No dif-ference ( p � 0.13) in plasma concentrations of �PCB,�PBDE, �OH-PCB, and �OH-PBDE was found between theF1 and P cohort dogs within the EXP and CON groups. Theexception, however, was for the �OH-PBDE concentrations,which were slightly elevated ( p � 0.01) in the CON F1 cohortdogs compared to the CON P cohort dogs. The relative pro-portions of individual PCB, PBDE, OH-PCB, and OH-PBDEcongeners to their respective sum concentrations have beenreported previously, and these are identical between the F1and P cohort dogs in each of the CON and EXP groups aswell as between males and females from the CON F1 cohort[15,16]. Hence, because of a general absence of differencesbetween the F1 and P cohort dogs (and between males andfemales) in plasma OHC concentrations and congener patterns,all dogs were merged in one single cohort for each of the EXPand CON dietary groups in subsequent analyses. The EXPindividuals accumulated consistently greater ( p � 0.003)�PCB, �PBDE, �OH-PCB, and �OH-PBDE concentrationsrelative to the CON dogs. Additional details regarding plasmaPCB, PBDE, OH-PCB, and OH-PBDE concentrations and con-gener profiles for the present sledge dogs as well as pork fatand Greenland minke whale blubber diets can be found else-where [15,16].
Associations between variables
Ages and body masses of sledge dogs, which were posi-tively interrelated (r � 0.53, p � 0.01, n � 22), were associatednegatively with UDPGT activity (r � �0.40, p � 0.06 and r� �0.44, p � 0.04 for ages and body masses, respectively).Hence, because ages and body masses had no significant pre-dictive effect on the variation of OHC concentrations (sumsand individual congeners) and the activity of the other enzymesinvestigated, these variables were excluded from further anal-yses. Moreover, because lipid percentages in plasma of sledgedogs were not associated ( p � 0.13) with any of the wet
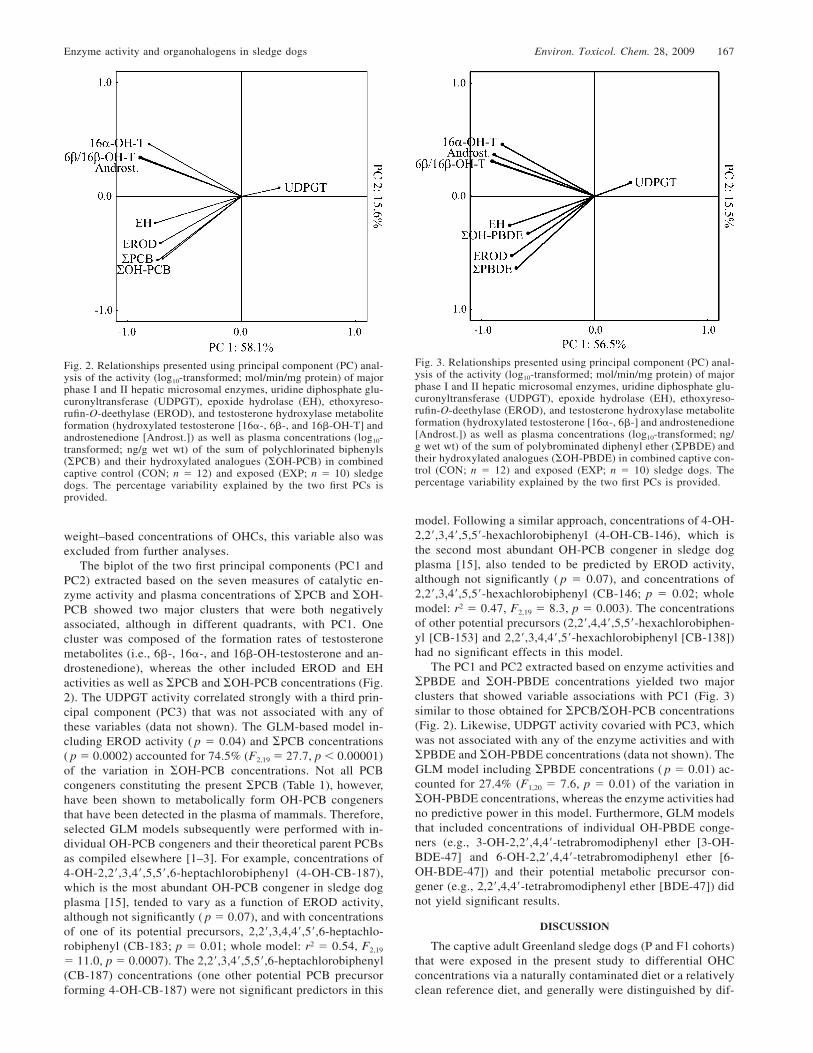
Enzyme activity and organohalogens in sledge dogs Environ. Toxicol. Chem. 28, 2009 167
Fig. 2. Relationships presented using principal component (PC) anal-ysis of the activity (log10-transformed; mol/min/mg protein) of majorphase I and II hepatic microsomal enzymes, uridine diphosphate glu-curonyltransferase (UDPGT), epoxide hydrolase (EH), ethoxyreso-rufin-O-deethylase (EROD), and testosterone hydroxylase metaboliteformation (hydroxylated testosterone [16�-, 6�-, and 16�-OH-T] andandrostenedione [Androst.]) as well as plasma concentrations (log10-transformed; ng/g wet wt) of the sum of polychlorinated biphenyls(�PCB) and their hydroxylated analogues (�OH-PCB) in combinedcaptive control (CON; n � 12) and exposed (EXP; n � 10) sledgedogs. The percentage variability explained by the two first PCs isprovided.
Fig. 3. Relationships presented using principal component (PC) anal-ysis of the activity (log10-transformed; mol/min/mg protein) of majorphase I and II hepatic microsomal enzymes, uridine diphosphate glu-curonyltransferase (UDPGT), epoxide hydrolase (EH), ethoxyreso-rufin-O-deethylase (EROD), and testosterone hydroxylase metaboliteformation (hydroxylated testosterone [16�-, 6�-] and androstenedione[Androst.]) as well as plasma concentrations (log10-transformed; ng/g wet wt) of the sum of polybrominated diphenyl ether (�PBDE) andtheir hydroxylated analogues (�OH-PBDE) in combined captive con-trol (CON; n � 12) and exposed (EXP; n � 10) sledge dogs. Thepercentage variability explained by the two first PCs is provided.
weight–based concentrations of OHCs, this variable also wasexcluded from further analyses.
The biplot of the two first principal components (PC1 andPC2) extracted based on the seven measures of catalytic en-zyme activity and plasma concentrations of �PCB and �OH-PCB showed two major clusters that were both negativelyassociated, although in different quadrants, with PC1. Onecluster was composed of the formation rates of testosteronemetabolites (i.e., 6�-, 16�-, and 16�-OH-testosterone and an-drostenedione), whereas the other included EROD and EHactivities as well as �PCB and �OH-PCB concentrations (Fig.2). The UDPGT activity correlated strongly with a third prin-cipal component (PC3) that was not associated with any ofthese variables (data not shown). The GLM-based model in-cluding EROD activity ( p � 0.04) and �PCB concentrations( p � 0.0002) accounted for 74.5% (F2,19 � 27.7, p � 0.00001)of the variation in �OH-PCB concentrations. Not all PCBcongeners constituting the present �PCB (Table 1), however,have been shown to metabolically form OH-PCB congenersthat have been detected in the plasma of mammals. Therefore,selected GLM models subsequently were performed with in-dividual OH-PCB congeners and their theoretical parent PCBsas compiled elsewhere [1–3]. For example, concentrations of4-OH-2,2�,3,4�,5,5�,6-heptachlorobiphenyl (4-OH-CB-187),which is the most abundant OH-PCB congener in sledge dogplasma [15], tended to vary as a function of EROD activity,although not significantly ( p � 0.07), and with concentrationsof one of its potential precursors, 2,2�,3,4,4�,5�,6-heptachlo-robiphenyl (CB-183; p � 0.01; whole model: r2 � 0.54, F2,19
� 11.0, p � 0.0007). The 2,2�,3,4�,5,5�,6-heptachlorobiphenyl(CB-187) concentrations (one other potential PCB precursorforming 4-OH-CB-187) were not significant predictors in this
model. Following a similar approach, concentrations of 4-OH-2,2�,3,4�,5,5�-hexachlorobiphenyl (4-OH-CB-146), which isthe second most abundant OH-PCB congener in sledge dogplasma [15], also tended to be predicted by EROD activity,although not significantly ( p � 0.07), and concentrations of2,2�,3,4�,5,5�-hexachlorobiphenyl (CB-146; p � 0.02; wholemodel: r2 � 0.47, F2,19 � 8.3, p � 0.003). The concentrationsof other potential precursors (2,2�,4,4�,5,5�-hexachlorobiphen-yl [CB-153] and 2,2�,3,4,4�,5�-hexachlorobiphenyl [CB-138])had no significant effects in this model.
The PC1 and PC2 extracted based on enzyme activities and�PBDE and �OH-PBDE concentrations yielded two majorclusters that showed variable associations with PC1 (Fig. 3)similar to those obtained for �PCB/�OH-PCB concentrations(Fig. 2). Likewise, UDPGT activity covaried with PC3, whichwas not associated with any of the enzyme activities and with�PBDE and �OH-PBDE concentrations (data not shown). TheGLM model including �PBDE concentrations ( p � 0.01) ac-counted for 27.4% (F1,20 � 7.6, p � 0.01) of the variation in�OH-PBDE concentrations, whereas the enzyme activities hadno predictive power in this model. Furthermore, GLM modelsthat included concentrations of individual OH-PBDE conge-ners (e.g., 3-OH-2,2�,4,4�-tetrabromodiphenyl ether [3-OH-BDE-47] and 6-OH-2,2�,4,4�-tetrabromodiphenyl ether [6-OH-BDE-47]) and their potential metabolic precursor con-gener (e.g., 2,2�,4,4�-tetrabromodiphenyl ether [BDE-47]) didnot yield significant results.
DISCUSSION
The captive adult Greenland sledge dogs (P and F1 cohorts)that were exposed in the present study to differential OHCconcentrations via a naturally contaminated diet or a relativelyclean reference diet, and generally were distinguished by dif-

168 Environ. Toxicol. Chem. 28, 2009 J. Verreault et al.
ferent hepatic microsomal phase I enzyme activities (CYP-likeand EH). In contrast, no difference in catalytic rates was foundfor the phase II, UDPGT-mediated enzyme activity. The dis-tinct variable between the CON and EXP sledge dogs was thediet, in which the Greenland minke whale blubber containedmuch greater levels of a complex suite of OHCs relative tothose in the pork fat diet (Table 1). Numerous OHC compoundsthat were measured in the present study are inducers of CYPenzyme expression in mammals, including dogs (see followingsections). This strongly suggests that dietary OHC exposuremediated the elevation of CYP-like and EH activities in sledgedogs.
Enzyme induction and substrate specificity
The CYP-like and EH activities in sledge dogs did not varyas a function of age or body mass and were identical betweensexes. A modest, significant, age-dependent (and body mass–dependent, because these two variables covaried) variation,however, was observed for UDPGT activity. Pharmacokineticstudies in dogs frequently have shown drug-specific charac-teristics in the hepatic microsomal protein induction, expres-sion, and activity of CYP isoenzymes between individuals ofdifferent ages [31] and between males and females [32]. Forthe present sledge dog cohorts, however, our assessment re-garding the effect of these confounding variables (i.e., age,body mass, and sex) was based on a very small sample size;hence, no conclusion could be drawn on the basis of theseresults.
Hitherto, nine constitutive CYP isoenzymes within the sub-families 1A, 2B–E, and 3A have been isolated and sequencedfrom the liver of canines (mainly beagle dogs): CYP1A1/2[33], CYP2B11 [34], CYP2C21/41 [33,35], CYP2D15 [36],CYP2E1 [37], and CYP3A12/26 [38,39]. Metabolic degra-dation of a large number of xenobiotic substrates has beenassociated with specific dog CYP isoenzymes and their hepaticmicrosomal activities. Pharmacokinetic studies using dogshave involved pharmacologically active compounds (drugs)primarily and OHCs of environmental concern (almost exclu-sively PCBs) to a much less extent. For example, dog livermRNA of CYP1A1/2 and CYP2C21 was first purified fromPCB-treated beagle dogs [33]. In another study, mongrel dogsexposed through the diet to a PCB mixture (Aroclor 1248) hadhigher liver microsomal CYP1A1 content based on immu-noblots and higher EROD activity, whereas no induction ofCYP2B-like activity based on pentoxyresorufin-O-deethylaseactivity was found [40]. Beagle dog CYP2B11 also has beenreported to have high metabolic activity toward the major PCBcongener detected in biota, CB-153 [34,41–43], which wasmore efficiently metabolized and excreted (via feces and urine)in beagle dogs compared to other noncanine mammalian spe-cies [44]. At present, the specificity and inductive capacity oftechnical PBDE mixtures or individual PBDE congeners onphase I and II enzymes has yet to be investigated in dogs.Nonetheless, studies in other mammalian species (e.g., ratsand mice) have found low hepatic microsomal CYP1A1/2-likeinduction for a few environmentally relevant PBDE congeners(e.g., BDE-47, 2,2�,4,4�,5-pentabromodiphenyl ether [BDE-99], 2,2�,4,4�,6-pentabromodiphenyl ether [BDE-100], 2,2�,4,4�,5,5�-hexabromodiphenyl ether [BDE-153], and 2,2�,4,4�,5�,6-hexabromodiphenyl ether [BDE-154]) (for review, seeHakk and Letcher [1]).
Ethoxyresorufin-O-deethylase activity
The EROD activity was twofold higher in adult female (Pcohort) EXP sledge dogs relative to that in the analogous CONcohort, which was associated with 12-fold higher plasma�PCB concentrations (from 1.1 to 12.9 ng/g wet wt) reachedover a time frame of 90 to 94 weeks of feeding. By comparison,in the study by Korytko et al. [40], in which a 5-ppm dose ofPCB mixture (Aroclor 1248) was administered daily via thediet to adult female mongrel dogs during a 20-week period,liver microsomal EROD activity was roughly threefold higherin the exposed individuals relative to the controls, and thiswas associated with a fourfold increase in serum �PCB con-centrations (from 0.3 to 1.2 ng/g wet wt). This may suggestthat CYP1A1/2-like enzymes in mongrel dogs are more in-ducible by aryl hydrocarbon–receptor agonists (e.g., dioxins/furans and dioxin-like PCBs) relative to the present sledgedogs, which were exposed to a substantially more complexsuite of potential CYP1A1/2-like inducers (Table 1) [15,16].These data should be interpreted with caution, however, be-cause the concentrations of the most potent aryl hydrocarbon–receptor inducers either were not determined (e.g., dioxins/furans and coplanar PCBs) or were below the limit of detectionin plasma of sledge dogs (e.g., mono-ortho-2,3,3�,4,4�-pen-tachlorobiphenyl [CB-105], 2,3�4,4�,5-pentachlorobiphenyl[CB-118], 2,3,3�,4,4�,5-hexachlorobiphenyl [CB-156], 2,3,3�,4,4�,5�-hexachlorobiphenyl [CB-157], and 2,3,3�,4,4�,5,5�-heptachlorobiphenyl [CB-189]). Nonetheless, these congenershave been shown in several mammalian species (e.g., polarbears) to vary with the concentrations of other nonplanar andmore bulky PCB congeners [19]. Regardless, the present find-ings also may be suggestive that sledge dogs are less efficientmetabolizers of PCBs relative to mongrel dogs because of, forexample, potential dysfunctionality of certain liver microsomalCYP isoforms (i.e., genetic polymorphism) and/or their rela-tive content. This was postulated by Hay-Kraus et al. [45] toexplain a threefold lower propofol (an anesthetic agent) hy-droxylase activity that was observed in liver microsomes ofgreyhound dogs relative to beagle dogs [46], which was con-firmed to be a CYP2B11-mediated reaction. At present, geneticpolymorphism of CYP isoenzymes has been demonstratedamong pure and mixed-breed beagle dogs for five CYP sub-families: CYP1A2, CYP2C41, CYP2D15, CYP2E1, andCYP3A12 [47].
Testosterone hydroxylation activity
The relative pattern of testosterone metabolite formation(i.e., four mono-OH-testosterone and androstenedione) in CONand EXP sledge dog liver microsomes was consistent with thatreported elsewhere for beagle dog liver microsomes (Fig. 1)[48,49]. The hydroxylation of testosterone in dogs has beenshown to be selectively catalyzed by certain hepatic CYP iso-enzymes, leading to the formation of mono-OH-testosteroneproducts at specific carbons of testosterone. More specifically,16�-OH-testosterone and, to a lesser extent, 16�-OH-testos-terone formation in dog liver microsome incubates have beenused as markers of CYP2B11 [32,48,50] and CYP2C21 in-duction [33,50]. Metabolism of testosterone also has been re-ported to yield androstenedione as one of the major productsformed in dog liver microsomes [49], which likely involves aCYP2B/2C-mediated transformation, as demonstrated withrats [51]. For the present CON and EXP sledge dogs, CYP2B/2C-like enzymes appeared to be moderately elevated based onthe relatively low formation rates of 16�- and 16�-OH-tes-

Enzyme activity and organohalogens in sledge dogs Environ. Toxicol. Chem. 28, 2009 169
tosterone as well as androstenedione. Moreover, 6�-OH-tes-tosterone in dogs has been shown to be generated largely byCYP3A12 mediation [32,50,52]. For example, treatment withrifampicin (a bactericidal antibiotic drug) has been docu-mented in beagle dogs to induce CYP3A12-mediated forma-tion of 16�-OH-testosterone in addition to 6�-OH-testosterone[32]. Because 6�-OH-testosterone was the major testosteronemetabolite formed in the sledge dogs of the present study, andbecause 6�- and 16�-OH-testosterone, both of which weresignificantly different between the CON and EXP groups, aresuggested markers of CYP3A12 induction, CYP3A-like en-zymes may be comparatively more inducible in sledge dogs.In fact, CYP3A isoenzymes are some of the most versatileCYP isoforms in mammals, exhibiting very low substrate spec-ificity and being induced by a large number of more bulky,globular exogenous molecules [53].
Epoxide hydrolase activity
Epoxide hydrolase was the phase I enzyme that showed thesecond-highest difference in catalytic activity (after EROD)between the CON and EXP sledge dog groups. In mammalianspecies, at least five EH isoforms have been identified [54],of which one hepatic microsomal form, resembling that of therat, has been purified and characterized in a phenobarbital-dosed male beagle dog [55]. It has been shown that the mostcommon environmental chemical class metabolized by EH areepoxide derivatives of the polycyclic aromatic hydrocarbons(e.g., benzo[a]pyrene 4,5-oxide [54]). Although data are lack-ing for dogs, PCB dosage has been reported to induce micro-somal EH activity in rodents (e.g., in rats [56] and mice [57]).Nonetheless, studies of beagle dogs administered phenobar-bital orally have demonstrated that dogs possess a unique abil-ity to metabolically form arene oxide intermediates from PCBcongeners lacking two adjacent, chlorine-unsubstituted carbonatoms [41,58]. Although no direct link to EH expression oractivity was established in these beagle dog studies, it wasconfirmed using liver microsomes that the metabolism of CB-153 and 2,2�,4,4�,6,6�-hexachlorobiphenyl (CB-155) occursvia 2,3- and 3,4-arene oxide intermediates, respectively, lead-ing to the formation of several OH-PCB congeners. The au-thentic standards of these OH-PCB congeners were not avail-able for the present study, but a similar mechanism can bepostulated to occur in sledge dogs. At present, the EH-medi-ated metabolism of PBDE epoxide derivatives has yet to bereported for canines [1].
Uridine diphosphoglucuronosyl transferase activity
To date, at least two canine UDPGT isoforms, UGT1A6and UGT2B31, have been cloned and characterized in liver ofbeagle dogs [59,60]. Among the potential substrates identifiedfor UDPGT-mediated reactions (e.g., carboxylic acids, phe-nols, opioids, and steroids), dog liver microsomal UGT1A6was reported to predominantly catalyze the glucuronidation ofa single class of compounds, the simple planar phenolic com-pounds [59]. Moreover, Soars et al. [60] demonstrated that dogliver microsomal UGT2B31 can catalyze a broader range ofsubstrates, including phenolic compounds but also selectedopioids and carboxylic acid-containing drugs. In the presentsledge dogs, catalytic UDPGT activity was comparable be-tween the CON and EXP individuals, showing a marginal butnonsignificant difference between these two dietary groups. Inline with these findings, studies of other chemically exposedmammals (mainly rodents) also have reported subtle and, oc-
casionally, non-dose-related changes in the induced expressionand catalytic activity of liver microsomal UDPGT, as shown,for example, in rats and mice exposed to PCBs and PBDEs[61–63]. Hence, the limited dietary treatment–related changesin liver microsomal UDPGT activity in sledge dogs suggestthat the activity of this phase II enzyme is a poor predictor/biomarker of OHC-mediated induction for the present suite ofPCB and PBDE congeners. It also may be postulated that theOH-PCBs and OH-PBDEs identified thus far in plasma ofsledge dogs, which consist mainly of nonplanar and bulkyphenolic congeners (Table 1), are poor substrates or inducersfor liver microsomal UDPGT isoforms relative to the simpleplanar phenolics [59,60]. Moreover, the cocktail of OHCs andother unknown and potential UDPGT inducers in the Green-land minke whale blubber or pork fat diet of sledge dogs iscomplex, but the present results indicate that this type of ex-posure is not differential with respect to the UDPGT catalyticactivity.
OH-PCB and OH-PBDE concentrations and enzymeactivities
The hepatic microsomal activity of EROD (CYP1A1/2-like) and plasma �PCB concentrations were significant pre-dictors for the variation of plasma �OH-PCB concentrationsin the combined CON and EXP sledge dogs. Despite apparentvariable associations between plasma �PCB and �OH-PCBconcentrations and EH activity (Fig. 2), GLM-based analysisyielded nonsignificant results. Consistent and significant as-sociations were found between major OH-PCB congenersidentified thus far in sledge dog plasma and their theoreticalprecursor PCB congeners and EROD activity. Nonetheless,because most of the PCB and OH-PCB congeners presentlydetected in plasma of sledge dogs strongly covaried withineach class, the discriminatory effect of specific congeners onthe basis of the magnitude and significance of their associationswith EROD activity could hardly be achieved for the presentdataset. Although this remains to be verified for the sledgedogs of the present study, these tendencies may provide anindication that hepatic microsomal CYP1A1/2-like isoen-zymes mediate in concert with EH during the multistep con-version of PCBs to their OH-containing forms retained in plas-ma (Fig. 4), as, for example, has been reported in rats [64].Overall, the present results may suggest that among the cur-rently determined catalytic assays, the hepatic microsomalEROD activity is a predictive indicator for the occurrence ofplasma OH-PCB residues along with those of PCBs in captiveadult Greenland sledge dogs. This also reinforces that OH-PCBs were entirely metabolically derived from dietary-accu-mulated PCBs in the CON and EXP sledge dogs, because OH-PCBs were notably absent in the Greenland minke whale blub-ber and pork fat diets (Table 1). Furthermore, because onlythe �PBDE concentrations significantly, but weakly, predictedthe �OH-PBDE variation in EXP sledge dog plasma, this sup-ports the idea that Greenland minke whale blubber was theunique accumulative source of the present plasma OH-PBDEs,as opposed to their having an origin via metabolic formationfrom precursor PBDEs. The present lack of association be-tween EROD activity and plasma PBDE concentrations maybe explained by the lower specificity and inductive propensityof these compounds to CYP1A1/2-like enzymes relative todioxin-like PCBs and other planar aromatics [1]. Alternatively,the present results also may indicate that environmentally rel-evant PBDEs possibly interfere with EROD activity in vitro,

170 Environ. Toxicol. Chem. 28, 2009 J. Verreault et al.
Fig. 4. Simplified metabolic scheme for polychlorinated biphenyls (PCBs) exemplified for 2,2�,4,5,5�-pentachlorobiphenyl (CB-101) showing themajor phase I (cytochrome P450 [CYP] 1/2/3 subfamilies and epoxide hydrolase [EH]) and phase II enzymes (uridine diphosphate glucuron-yltransferase [UDPGT], glutathione [GSH], and glutathione-S-transferase [GST]) involved. The major PCB metabolites formed are the methyl-sulfonated (MeSO2) and hydroxylated (OH) PCBs.
as reported, for example, in primary cultured carp (Cyprinuscarpio) hepatocytes [65].
The reasons for the nonsignificant associations betweenplasma concentrations of OH-PCBs and PCBs, or OH-PBDEsand PBDEs, and TH activity (CYP2B/2C/3A-like) are unclear,because in the present study, most of the measured compoundsin sledge dog plasma were good theoretical substrates forCYP2B/2C/3A-like enzymes. This lack of relationship, how-ever, could be attributed to the choice of assay, because tes-tosterone may not be the substrate exhibiting the best speci-ficity for these CYP isoforms in sledge dogs. Furthermore, thenotable absence of a relationship between UDPGT activity andplasma OH-PCB and OH-PBDE concentrations could be aresult of the multiple mechanisms that can compete with theUDPGT-mediated reaction (Fig. 4). Among other pathways,these include excretion, protection (i.e., via protein binding),and secondary oxidation (i.e., CYP-mediated formation of di-OH congeners) of OH-PCBs and OH-PBDEs as well as po-tential interactions with other conjugative pathways mediatedby the glutathione and glutathione-S-transferase enzymes. Al-ternatively, this could be a consequence of the minor and,perhaps, negligible contribution of simple planar phenolicsincluding OH-PCB and OH-PBDE congeners in sledge dogplasma, although these are yet to be identified in this species.
Future studies should aim to define the implications of mul-tiple isoforms and their substrate specificity and activity forthe present suite of liver microsomal phase I and II enzymesas well as other biotransformation enzymes that have receivedless attention in dogs, such as glutathione-S-transferase andglutathione. Complementary investigations also should ex-amine in greater depth the effect of sex and age on the met-abolic enzyme content and catalytic activity in sledge dogs.Such studies will provide a better understanding about someof the factors influencing the fate and toxicokinetics of OHCsas well as potential bioactivation via metabolite formation insledge dogs and, perhaps, in other top Arctic carnivore Can-oidea species as well, such as the polar bear, Arctic fox, andArctic wolf.
Acknowledgement—The main financial support for the present studywas a Discovery Grant from the Natural Science and Engineering
Research Council (NSERC) of Canada (to R.J. Letcher), the Envi-ronment Canada Wildlife Toxicology and Disease Program (to R.J.Letcher), and the Danish Cooperation for Environment in the Arcticand the Lundbeck Foundation (to R. Dietz and C. Sonne). We grate-fully acknowledge Thomas Dau Rasmussen, Anne Silverbau, Mikaeland Jane Rasmussen, Jørn Breinholt, and Jens Kjeldsen for their tech-nical assistance in the field as well as Steen Andersen and SiggaJoensen for sample collection. We also are very grateful to SoheilaShahmiri for assistance with the chemical analyses.
REFERENCES
1. Hakk H, Letcher RJ. 2003. Metabolism in the toxicokinetics andfate of brominated flame retardants—A review. Environ Int 29:801–828.
2. Kawano M, Hasegawa J, Enomoto T, Onishi H, Nishio Y, MatsudaM, Wakimoto T. 2005. Hydroxylated polychlorinated biphenyls(OH-PCBs): Recent advances in wildlife contamination study.Environ Sci 12:315–324.
3. Letcher RJ, Klasson-Wehler E, Bergman A. 2000. Methyl sulfoneand hydroxylated metabolites of polychlorinated biphenyls. InPaasivita J, ed, The Handbook of Environmental Chemistry—New Types of Persistent Halogenated Compounds, Vol 3, PartK. Springer-Verlag, Heidelberg, Germany, pp 315–359.
4. Teuten EL, Xu L, Reddy CM. 2005. Two abundant bioaccumu-lated halogenated compounds are natural products. Science 307:917–920.
5. Tampal N, Lehmler HJ, Espandiari P, Malmberg T, Robertson LW.2002. Glucuronidation of hydroxylated polychlorinated biphenyls(PCBs). Chem Res Toxicol 15:1259–1266.
6. Houde M, Pacepavicius G, Wells RS, Fair PA, Letcher RJ, AlaeeM, Bossart GD, Hohn AA, Sweeney J, Solomon KR, Muir DCG.2006. Polychlorinated biphenyls and hydroxylated polychlori-nated biphenyls in plasma of bottlenose dolphins (Tursiops trun-catus) from the western Atlantic and the Gulf of Mexico. EnvironSci Technol 40:5860–5866.
7. McKinney MA, Martineau D, Dallaire AD, Beland P, De GuiseS, LeBeuf M, Letcher RJ. 2006. Organohalogen contaminantsand metabolites in the liver of beluga whales (Delphinapterusleucas) from two Canadian populations. Environ Toxicol Chem25:1246–1257.
8. Verreault J, Gabrielsen GW, Chu S, Muir DCG, Andersen M,Hamaed A, Letcher RJ. 2005. Flame retardants and methoxylatedand hydroxylated polybrominated diphenyl ethers in two Nor-wegian Arctic top predators: Glaucous gulls and polar bears. En-viron Sci Technol 39:6021–6028.
9. Gebbink WA, Sonne C, Dietz R, Kirkegaard M, Born EW, MuirDCG, Letcher RJ. 2008. Target tissue selectivity and burdens ofdiverse classes of brominated and chlorinated contaminants inpolar bears (Ursus maritimus) from East Greenland. Environ SciTechnol 152:621–629.

Enzyme activity and organohalogens in sledge dogs Environ. Toxicol. Chem. 28, 2009 171
10. Sonne C, Dietz R, Larsen HJ, Loft KE, Kirkegaard M, LetcherRJ, Shahmiri S, Moller P. 2006. Impairment of cellular immunityin west Greenland sledge dogs (Canis familiaris) dietary exposedto polluted minke whale (Balaenoptera acutorostrata) blubber.Environ Sci Technol 40:2056–2062.
11. Sonne C, Leifsson PS, Dietz R, Kirkegaard M, Møller P, JensenAJ, Letcher RJ, Shahmiri S. 2007. Renal lesions in Greenlandsledge dogs (Canis familiaris) exposed to a natural dietary cock-tail of persistent organic pollutants. Toxicol Environ Chem 89:563–576.
12. Sonne C, Fonfara S, Dietz R, Kirkegaard M, Letcher RJ, ShahmiriS, Andersen S, Moller P. 2007. Multiple cytokine and acute-phaseprotein gene transcription in West Greenland sledge dogs (Canisfamiliaris) dietary exposed to organic environmental pollutants.Arch Environ Contam Toxicol 53:110–118.
13. Sonne C, Dietz R, Kirkegaard M, Letcher RJ, Shahmiri S, An-dersen S, Moller P, Olsen AK, Jensen AL. 2008. Effects of or-ganohalogen pollutants on hematological and urine clinical-chem-ical parameters in Greenland sledge dogs (Canis familiaris). Eco-toxicol Environ Saf 69:381–390.
14. Sonne C, Leifsson PS, Dietz R, Kirkegaard M, Møller P, JensenAL, Letcher RJ. 2008. Greenland sledge dogs (Canis familiaris)develop liver lesions when exposed to a chronic and dietary lowdose of an environmental organohalogen cocktail. Environ Res106:72–80.
15. Verreault J, Dietz R, Sonne C, Gebbink WA, Shahmiri S, LetcherRJ. 2008. Comparative fate of organohalogen contaminants intwo top carnivores in Greenland: Captive sledge dogs and wildpolar bears. Comp Biochem Physiol C Toxicol Pharmacol 147:306–315.
16. Verreault J, Sonne C, Dietz R, Letcher RJ. 2008. Dietary, age,and trans-generational effects on the fate of organohalogen con-taminants in captive sledge dogs in Greenland. Environ Int (inpress).
17. Bandiera SM, Torok SM, Lin S, Ramsay MA, Norstrom RJ. 1995.Catalytic and immunologic characterization of hepatic and lungcytochromes P450 in the polar bear. Biochem Pharmacol 49:1135–1146.
18. Bandiera SM, Torok SM, Letcher RJ, Norstrom RJ. 1997. Im-munoquantitation of cytochromes P4501A and P4502B and com-parison with chlorinated hydrocarbon levels in archived polar bearliver samples. Chemosphere 34:1469–1479.
19. Letcher RJ, Norstrom RJ, Lin S, Ramsay MA, Bandiera SM.1996. Immunoquantitation and microsomal monooxygenase ac-tivities of hepatic cytochromes P4501A and P4502B and chlo-rinated hydrocarbon contaminant levels in polar bear (Ursus mar-itimus). Toxicol Appl Pharmacol 137:127–140.
20. Hobbs KE, Muir DCG, Born EW, Dietz R, Haug T, Metcalfe T,Metcalfe C, Øien N. 2003. Levels and patterns of persistent or-ganochlorines in minke whale (Balaenoptera acutorostrata) fromthe North Atlantic and European Arctic. Environ Pollut 121:239–252.
21. Kleivane L, Skaare JU. 1998. Organochlorine contaminants innortheast Atlantic minke whales (Balaenoptera acutorostrata).Environ Pollut 101:231–239.
22. Sonne C, Riget FF, Jensen JEB, Hyldstrup L, Teilmann J, DietzR, Kirkegaard M, Andersen S, Letcher RJ, Jakobsen J. 2008.Does the nutrition profile of vitamins, fatty acids, and microel-ements counteract the negative impact from organohalogen pol-lutants on bone mineral density in Greenland sledge dogs (Canisfamiliaris)? Environ Int 34:811–820.
23. Kennedy SW, Jones SP. 1994. Simultaneous measurement of cy-tochrome P4501A catalytic activity and total protein concentra-tion with a fluorescence plate reader. Anal Biochem 222:217–223.
24. Li H, Letcher RJ. 2002. A high-performance-liquid-chromatog-raphy–based method for the determination of hydroxylated tes-tosterone metabolites formed in vitro in liver microsomes fromgray seal (Halichoerus grypus). J Chromatogr Sci 40:397–402.
25. Whalley PM, Bakes D, Grime K, Weaver RJ. 2001. Rapid high-performance liquid chromatographic method for the separation ofhydroxylated testosterone metabolites. J Chromatogr B BiomedSci Appl 760:281–288.
26. Eaton DL, Stapleton PL. 1989. Simultaneous determination ofcytosolic glutathione-S-transferase and microsomal epoxide hy-drolase activity toward benzo[a]pyrene-4,5-oxide by high-per-formance liquid chromatography. Anal Biochem 178:153–158.
27. Collier AC, Tingle MD, Keelan JA, Paxton JW, Mitchell MD.
2000. A highly sensitive fluorescent microplate method for thedetermination of UDP-glucuronosyl transferase activity in tissuesand placental cell lines. Drug Metab Dispos 28:1184–1186.
28. Maervoet J, Covaci A, Schepens P, Sandau CD, Letcher RJ. 2004.A reassessment of the nomenclature of polychlorinated biphenyl(PCB) metabolites. Environ Health Perspect 112:291–294.
29. Hair JF, Anderson RE, Tatham RL, Black WC. 1998. MultivariateData Analysis, 5th ed. Prentice Hall, Upper Saddle River, NJ,USA.
30. Jayyosi Z, Muc M, Erick J, Thomas PE, Kelley M. 1996. Catalyticand immunochemical characterization of cytochrome P450 iso-zyme induction in dog liver. Fundam Appl Toxicol 31:95–102.
31. Tanaka E, Narisawa C, Nakamura H, Sawa Y, Etoh H, TadanoK, Horie T, Ohkawa H, Misawa S. 1998. Changes in the enzymaticactivities of beagle liver during maturation as assessed both invitro and in vivo. Xenobiotica 28:795–802.
32. Nishibe Y, Wakabayashi M, Harauchi T, Ohno K. 1998. Char-acterization of cytochrome P450 (CYP3A12) induction by rifam-picin in dog liver. Xenobiotica 28:549–557.
33. Uchida T, Komori M, Kitada M, Kamataki T. 1990. Isolation ofcDNAs coding for three different forms of liver microsomal cy-tochrome P450 from polychlorinated biphenyl–treated beagledogs. Mol Pharmacol 38:644–651.
34. Graves PE, Elhag GA, Ciaccio PJ, Bourque DP, Halpert JR. 1990.cDNA and deduced amino acid sequences of a dog hepatic cy-tochrome P450IIB responsible for the metabolism of 2,2�,4,4�,5,5�-hexachlorobiphenyl. Arch Biochem Biophys 281:106–115.
35. Blaisdell J, Goldstein JA, Bai SA. 1998. Isolation of a new caninecytochrome P450 CDNA from the cytochrome P450 2C subfamily(CYP2C41) and evidence for polymorphic differences in its ex-pression. Drug Metab Dispos 26:278–283.
36. Sakamoto K, Kirita S, Baba T, Nakamura Y, Yamazoe Y, KatoR, Takanaka A, Matsubara T. 1995. A new cytochrome P450 formbelonging to the CYP2D in dog liver microsomes: Purification,cDNA cloning, and enzyme characterization. Arch Biochem Bio-phys 319:372–382.
37. Lankford SM, Bai SA, Goldstein JA. 2000. Cloning of caninecytochrome P450 2E1 cDNA: Identification and characterizationof two variant alleles. Drug Metab Dispos 28:981–986.
38. Ciaccio PJ, Graves PE, Bourque DP, Glinsmann-Gibson B, Hal-pert JR. 1991. cDNA and deduced amino acid sequences of adog liver cytochrome P450 of the IIIA gene subfamily. BiochimBiophys Acta 1088:319–322.
39. Fraser DJ, Feyereisen R, Harlow GR, Halpert JR. 1997. Isolation,heterologous expression, and functional characterization of a nov-el cytochrome P4503A enzyme from a canine liver cDNA library.J Pharmacol Exp Ther 283:1425–1432.
40. Korytko PJ, Casey AC, Bush B, Quimby FW. 1999. Induction ofhepatic cytochromes P450 in dogs exposed to a chronic low doseof polychlorinated biphenyls. Toxicol Sci 47:52–61.
41. Ariyoshi N, Koga N, Oguri K, Yoshimura H. 1992. Metabolismof 2,4,5,2�,4�,5�-hexachlorobiphenyl with liver microsomes ofphenobarbital-treated dog: The possible formation of PCB 2,3-arene oxide intermediate. Xenobiotica 22:1275–1290.
42. Duignan DB, Sipes IG, Leonard TB, Halpert JR. 1987. Purifi-cation and characterization of the dog hepatic cytochrome P450isozyme responsible for the metabolism of 2,2�,4,4�,5,5�-hexa-chlorobiphenyl. Arch Biochem Biophys 255:290–303.
43. Duignan DB, Sipes IG, Ciaccio PJ, Halpert JR. 1988. The me-tabolism of xenobiotics and endogenous compounds by the con-stitutive dog liver cytochrome P450 PBD-2. Arch Biochem Bio-phys 267:294–304.
44. Sipes IG, Slocumb ML, Perry DF, Carter DE. 1982. 2,4,5,2�,4�,5�-Hexachlorobiphenyl: Distribution, metabolism, and excretion inthe dog and the monkey. Toxicol Appl Pharmacol 65:264–272.
45. Hay-Kraus BL, Greenblatt DJ, Venkatakrishnan K, Court MH.2000. Evidence for propofol hydroxylation by cytochromeP4502B11 in canine liver microsomes: Breed and gender differ-ences. Xenobiotica 30:575–588.
46. Court MH, Hay-Kraus BL, Hill DW, Kind AJ, Greenblatt DJ.1999. Propofol hydroxylation by dog liver microsomes: Assaydevelopment and dog breed differences. Drug Metab Dispos 27:1293–1299.
47. Kamimura H. 2006. Genetic polymorphism of cytochrome P450sin beagles: Possible influence of CYP1A2 deficiency on toxico-logical evaluations. Arch Toxicol 80:732–738.
48. Ohmori S, Taniguchi T, Rikihisa T, Kanakubo Y, Kitada M. 1993.

172 Environ. Toxicol. Chem. 28, 2009 J. Verreault et al.
Species differences of testosterone 16-hydroxylases in liver mi-crosomes of guinea pig, rat and dog. Xenobiotica 23:419–426.
49. Williams TM, Kind AJ, Hill DW. 2000. Drug metabolism: Invitro biotransformation of anabolic steroids in canines. J VetPharmacol Ther 23:57–66.
50. Shou M, Norcross R, Sandig G, Lu P, Li Y, Lin Y, Mei Q, Rod-rigues AD, Rushmore TH. 2003. Substrate specificity and kineticproperties of seven heterologously expressed dog cytochromesP450. Drug Metab Dispos 31:1161–1169.
51. Ryan DE, Levin W. 1990. Purification and characterization ofhepatic microsomal cytochrome P450. Pharmacol Ther 45:153–239.
52. Ciaccio PJ, Halpert JR. 1989. Characterization of a phenobarbital-inducible dog liver cytochrome P450 structurally related to ratand human enzymes of the P450IIIA (steroid-inducible) genesubfamily. Arch Biochem Biophys 271:284–299.
53. Liu YT, Hao HP, Liu CX, Wang GJ, Xie HG. 2007. Drugs asCYP3A probes, inducers, and inhibitors. Drug Metab Rev 39:699–721.
54. Fretland AJ, Omiecinski CJ. 2000. Epoxide hydrolases: Bio-chemistry and molecular biology. Chem Biol Interact 129:41–59.
55. Ariyoshi N, Tanaka M, Ishii Y, Oguri K. 1994. Purification andcharacterization of dog liver microsomal epoxide hydrolase. JBiochem (Tokyo) 115:985–990.
56. Dragnev KH, Beebe LE, Jones CR, Fox SD, Thomas PE, NimsRW, Lubet RA. 1994. Subchronic dietary exposure to Aroclor1254 in rats: Accumulation of PCBs in liver, blood, and adiposetissue and its relationship to induction of various hepatic drug-metabolizing enzymes. Toxicol Appl Pharmacol 125:111–122.
57. Meijer J, DePierre JW. 1987. Hepatic levels of cytosolic, micro-somal, and ‘‘mitochondrial’’ epoxide hydrolases and other drug-metabolizing enzymes after treatment of mice with various xe-nobiotics and endogenous compounds. Chem Biol Interact 62:249–269.
58. Ariyoshi N, Yoshimura H, Oguri K. 1993. Identification of invitro metabolites of 2,4,6,2�,4�,6�-hexachlorobiphenyl from phe-nobarbital-treated dog liver microsomes. Biol Pharm Bull 16:852–857.
59. Soars MG, Smith DJ, Riley RJ, Burchell B. 2001. Cloning andcharacterization of a canine UDP-glucuronosyltransferase. ArchBiochem Biophys 391:218–224.
60. Soars MG, Fettes M, O’Sullivan AC, Riley RJ, Ethell BT, Bur-chell B. 2003. Cloning and characterization of the first drug-metabolizing canine UDP-glucuronosyltransferase of the 2B sub-family. Biochem Pharmacol 65:1251–1259.
61. Fisher JW, Campbell J, Muralidhara S, Bruckner JV, Ferguson D,Mumtaz M, Harmon B, Hedge JM, Crofton KM, Kim H, Al-mekinder TL. 2006. Effect of PCB 126 on hepatic metabolismof thyroxine and perturbations in the hypothalamic-pituitary-thy-roid axis in the rat. Toxicol Sci 90:87–95.
62. Hallgren S, Sinjari T, Hakansson H, Darnerud PO. 2001. Effectsof polybrominated diphenyl ethers (PBDEs) and polychlorinatedbiphenyls (PCBs) on thyroid hormone and vitamin A levels inrats and mice. Arch Toxicol 75:200–208.
63. Tseng LH, Li MH, Tsai SS, Lee CW, Pan MH, Yao WJ, Hsu PC.2008. Developmental exposure to decabromodiphenyl ether(PBDE 209): Effects on thyroid hormone and hepatic enzymeactivity in male mouse offspring. Chemosphere 70:640–647.
64. Taura KI, Yamada H, Hagino Y, Ishii Y, Mori MA, Oguri K.2000. Interaction between cytochrome P450 and other drug-me-tabolizing enzymes: Evidence for an association of CYP1A1 withmicrosomal epoxide hydrolase and UDP-glucuronosyltransferase.Biochem Biophys Res Commun 273:1048–1052.
65. Kuiper RV, Bergman A, Vos JG, van den Berg M. 2004. Somepolybrominated diphenyl ether (PBDE) flame retardants withwide environmental distribution inhibit TCDD-induced ERODactivity in primary cultured carp (Cyprinus carpio) hepatocytes.Aquat Toxicol 68:129–139.
Related Documents











