Commonness and rarity of plants in a reserve network: just two faces of the same coin Sara Landi • Alessandro Chiarucci Received: 20 January 2014 / Accepted: 22 June 2014 / Published online: 17 July 2014 Ó Accademia Nazionale dei Lincei 2014 Abstract Occurrence of protected and rare species is regarded as a strong argument for establishing protected areas and monitoring biodiversity, but while protected species are clearly identified, some problems exist to define rare species. It is thus important to know whether common and unprotected native species are reliable indicators for protected and rare species. The aims of this paper were to: (a) analyse the distribution of rarity and commonness of species, by using different criteria and (b) test if groups of species with different conservation value (aliens, unpro- tected natives and protected natives) differ in terms of their rarity distribution, using the data collected in 604 plots sampled within 21 protected areas of the central Italy. Three different criteria were used to classify species as rare or common. Pearson correlation, least-squares regressions and Chi-square test were used to compare the species richness patterns or rare and common species as well as protected, unprotected native, and alien species. The number of species classified as common and rare widely differ according to the adopted criterion. The number of common and rare species were statistically correlated at both the plot and protected area scales, even if at the plot scale the predictive capacity was rather low. Protected species were significantly rarer than expected, while unprotected species were significantly more common than expected; alien species confirmed to be particularly rare in our study area, with some major alien species being totally absent in the recorded flora. The richness patterns of common and rare species defined according to different criteria have been found to be correlated one to the other, and both are well related to the richness of protected and alien species at both the plot and PA scales. Protected species were better related to common species, while alien species were better related to rare species. Despite rare species were numerically more than common species, and the richness pattern of total species was better predicted by common species than rare species. Common species con- firmed to be good indicators of species richness patterns and also of protected species. Keywords Alien species Á Biodiversity Á Common species Á Network Á Protected species Á Rare species 1 Introduction Common and rare species have fundamentally different ecology and understanding how species with different occurrence contribute to overall patterns of biodiversity (Jetz et al. 2004; Gaston et al. 2008) is an important challenge for modern ecology. Many authors reported that rare species, i.e. those with few individuals or occur- rences, are the majority in most ecosystems (e.g., Pielou 1969; Gaston 1994, 2011; Lennon et al. 2004; Gaston and Fuller 2008). Rarity can have different causes and it is thought to increase the risk of extinction, either through demographic stochasticity or because of the vulnerability to environmental changes of species occupying a restric- ted habitat. Despite the wide usage of the rarity concept in ecology and conservation biology, a consistent definition of rarity is still missing (Rabinowitz et al. 1986; Gaston 1994). Many measures have been used to define this phenomenon, as the breadth of geographic range size, degree of habitat S. Landi (&) Á A. Chiarucci BIOCONNET, BIOdiversity and CONservation NETwork, Department of Life Sciences, University of Siena, Via P.A. Mattioli 4, 53100 Siena, Italy e-mail: [email protected] 123 Rend. Fis. Acc. Lincei (2014) 25:369–380 DOI 10.1007/s12210-014-0313-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Commonness and rarity of plants in a reserve network: just twofaces of the same coin
Sara Landi • Alessandro Chiarucci
Received: 20 January 2014 / Accepted: 22 June 2014 / Published online: 17 July 2014
� Accademia Nazionale dei Lincei 2014
Abstract Occurrence of protected and rare species is
regarded as a strong argument for establishing protected
areas and monitoring biodiversity, but while protected
species are clearly identified, some problems exist to define
rare species. It is thus important to know whether common
and unprotected native species are reliable indicators for
protected and rare species. The aims of this paper were to:
(a) analyse the distribution of rarity and commonness of
species, by using different criteria and (b) test if groups of
species with different conservation value (aliens, unpro-
tected natives and protected natives) differ in terms of their
rarity distribution, using the data collected in 604 plots
sampled within 21 protected areas of the central Italy.
Three different criteria were used to classify species as rare
or common. Pearson correlation, least-squares regressions
and Chi-square test were used to compare the species
richness patterns or rare and common species as well as
protected, unprotected native, and alien species. The
number of species classified as common and rare widely
differ according to the adopted criterion. The number of
common and rare species were statistically correlated at
both the plot and protected area scales, even if at the plot
scale the predictive capacity was rather low. Protected
species were significantly rarer than expected, while
unprotected species were significantly more common than
expected; alien species confirmed to be particularly rare in
our study area, with some major alien species being totally
absent in the recorded flora. The richness patterns of
common and rare species defined according to different
criteria have been found to be correlated one to the other,
and both are well related to the richness of protected and
alien species at both the plot and PA scales. Protected
species were better related to common species, while alien
species were better related to rare species. Despite rare
species were numerically more than common species, and
the richness pattern of total species was better predicted by
common species than rare species. Common species con-
firmed to be good indicators of species richness patterns
and also of protected species.
Keywords Alien species � Biodiversity � Common
species � Network � Protected species � Rare species
1 Introduction
Common and rare species have fundamentally different
ecology and understanding how species with different
occurrence contribute to overall patterns of biodiversity
(Jetz et al. 2004; Gaston et al. 2008) is an important
challenge for modern ecology. Many authors reported that
rare species, i.e. those with few individuals or occur-
rences, are the majority in most ecosystems (e.g., Pielou
1969; Gaston 1994, 2011; Lennon et al. 2004; Gaston and
Fuller 2008). Rarity can have different causes and it is
thought to increase the risk of extinction, either through
demographic stochasticity or because of the vulnerability
to environmental changes of species occupying a restric-
ted habitat.
Despite the wide usage of the rarity concept in ecology
and conservation biology, a consistent definition of rarity is
still missing (Rabinowitz et al. 1986; Gaston 1994). Many
measures have been used to define this phenomenon, as the
breadth of geographic range size, degree of habitat
S. Landi (&) � A. Chiarucci
BIOCONNET, BIOdiversity and CONservation NETwork,
Department of Life Sciences, University of Siena, Via P.A.
Mattioli 4, 53100 Siena, Italy
e-mail: [email protected]
123
Rend. Fis. Acc. Lincei (2014) 25:369–380
DOI 10.1007/s12210-014-0313-1

specificity, local frequency, ephemerality, relative abun-
dance, occurrence, and population size (Hartley and Kunin
2003; Gaston 2008). Occurrence of rare and protected plant
species is regarded as a major argument for creating pro-
tected areas and defining their management plans. Some
attempts have also been done to design reserve networks by
using distributional data of selected indicator species
(Hopkinson et al. 2001; Lawler et al. 2003) and this is
likely to be a central theme in future conservation biology.
However, protected species are often chosen on the basis of
‘‘pragmatic’’ issues, such as the presence of one or some
flagship species (Simberloff 1998). While there are no
particular problems in identifying protected species, the
comprehension of which species are rare is still an unre-
solved issue. It is then fundamental to know whether
common and unprotected species are reliable indicator of
rare and protected species, since it is more convenient to
obtain occurrence data for the former than for the latter.
Rarity is a factor that should be taken into account in
conservation biology and, especially, in listing species
which are threatened or need specific management actions
(Hercos et al. 2012). On the other hand, if most species are
rare, it is difficult to include rare species into management
or conservation actions. Moreover, species that are rare in
one locality are not necessarily rare everywhere, because
their abundance may be depressed due to local conditions,
such as unsuitable habitats (Rabinowitz 1981), or even
random processes (Hercos et al. 2012). It is also necessary
to consider that management can have a major role in
controlling commonness and rarity patterns of species.
Therefore, when comparing groups of species with differ-
ent management importance, as it can be the case of alien
and protected species, it is necessary to consider whether
they are common or rare.
Despite common species can be less in number than the
rare species, there is evidence that their contribution to
species richness patterns is often greater than that of rare
species (Jetz and Rahbek 2002; Pearman and Weber 2007;
Mazaris et al. 2008). However, extremely common species
(e.g., those species which occur in all the sampled sites) are
scarcely informative on species richness patterns, since a
species which is present in each sample does not add
nothing to such a pattern or gradient, only uniformly ele-
vates its values of one unit. Nevertheless, analyses of
diversity patterns may be largely based on common rather
rare species, because the latter are swamped by variation
contributed by the former (Lennon et al. 2011). So, a clear
understanding of the contribution of common and rare
species to the richness patterns of groups of species with
different conservation values is fundamental.
The goal of this paper is to analyse the rarity and
commonness patterns of plant species in a regional network
of protected areas, by using an extensive data set on plant
species occurrences (Chiarucci et al. 2012). In particular,
we aim to: (a) analyse the distribution of rarity and com-
monness of species, by using different criteria to define
rarity and (b) test if groups of species with different con-
servation value (alien, native and protected species) differ
in terms of rarity distribution.
2 Methods
Information about the richness of common and rare spe-
cies were obtained from the occurrence data of 1,041
vascular plant species (hereafter referred as S) in 604
plots (each of 100 m2) recorded in 21 protected areas
(PAs) of the province of Siena (central Italy) by using a
stratified random design (Chiarucci et al. 2012). On the
basis of the check-list of alien species of Italy (Celesti-
Grapow et al. 2009) as well as international and regional
laws, each species was classified as: alien (A), unpro-
tected native (N), or protected native (P). The alien spe-
cies group included both archeophyte and neophyte
species, and they can be casual, naturalized or even
invasive (Pysek et al. 2004).
Each species was classified as common (C) or rare
(R) on the basis of three different criteria. In the first two
criteria, the classification of species as common or rare was
based on arbitrary levels of species occurrence: (1) 1 plot
and 1 PA as thresholds and (2) five plots and three PAs as
thresholds. With these criteria, the species recorded with
occurrence level equal or below the threshold levels were
classified as rare, while those with occurrence level above
the threshold were classified as common. Four lists of
common species (CPl1, CPl5 at plot scale, and CPA1,
CPA3 at PA scale) as well as four paired of rare species
(RPl1, RPl5 at plot scale, and RPA1, RPA3 at PA scale)
were obtained. The third criterion (3) used the quartile
method (Gaston 1994) to divide species into common and
rare species: in detail, the 25 % of most frequent species
were classified as common while the remaining 75 % of
species were classified as rare. This criterion resulted in
two lists of common species (CPlq at plot scale and CPAq
at PA scale) and rare species (RPlq at plot scale and RPAq
at PA scale).
Correlation analysis and ordinary linear regressions
were used to detect the existence of relations between pairs
of the species richness variables obtained by the above
classifications. A Chi-squared test was used to test if the
three groups of species with different conservation values
(A, N and P) differed in the distribution of commonness
and rarity, at both the plot and PA scales. The null
hypothesis to be tested is that alien, native, and protected
species do not differ in their patterns of rarity and
commonness.
370 Rend. Fis. Acc. Lincei (2014) 25:369–380
123

The contribution of common and rare species to the
overall richness patterns was assessed using sequential
correlation analysis (e.g., Vazquez and Gaston 2004;
Lennon et al. 2004, 2011; Kreft et al. 2006; Mazaris et al.
2008). For doing this, two rank orders of all the recorded
species (1,041) were built based on species occurrence in
each plot: from the most common to most rare, and from
the more rare to most common. Then, two series of 1,041
sub-assemblages (each made of 604 plots) were built by
sequentially including each species, from the most com-
mon to the most rare and vice versa. In detail, the following
procedure was used to build the series of sub-assemblages
with species ranked in order of commonness: the first sub-
assemblages included only the occurrences of the most
common species (i.e., the species with the highest occur-
rence value) in the 604 plots; then, the second sub-
assemblages were made by adding the second most com-
mon species to the first one. The same procedure was
repeated until all the 1,041 species, from the most common
to the most rare, were added to the sub-assemblages,
reaching a total of 1,041 sub-assemblages. The same pro-
cedure was repeated to build a series of sub-assemblages,
from the most rare to the most common species. Then, the
species richness values of each of the sub-assemblages in
the common to rare (C to R) and rare to common (R to C)
sequences were calculated and correlated with the species
richness of the full assemblages, by using the Pearson’s
product-moment correlation. The same procedure was used
at the PA scale (1,041 sub-assemblages, each of 21 PAs,
for both the C to R and R to C sequences). This resulted in
two series of correlation coefficients (one for the C to R
sequence and one for the R to C sequence), for both the
plot and PA scale. Then, these two sequences of correlation
coefficients were plotted against the number of species
included into the sub-assemblages. This allowed to
understand how the species richness of the full-assem-
blages can be predicted by the species richness values of
the sub-assemblages of the n most common or the n most
rare species.
Then, a frequency index that weighs each species
according to its occurrence at plot and PA scale was
computed. The proportion of plots and PAs occupied by a
given species (p) was used to calculate the index p(1-p).
This index represents a measure of the spatial information
provided by each species (Lennon et al. 2004), with species
with intermediate occurrence providing more information
than extremely rare or extremely common ones. The index
p(1-p) was compared across the different groups of spe-
cies (A, N, and P, as well as common and rare species), to
compare the information contribution they provide. The
existence of statistically significant differences between the
p(1-p) index between common and rare species, at both
plot (ICPl1 vs. IRPl1, ICPl5 vs. IRPl5, and ICPlq vs.
IRPlq) and PA scale (ICPA1 vs. IRPA1, ICPA3 vs. IRPA3,
and ICPAq vs. IRPAq) were tested by the Mann–Whitney
test for independent samples. The non parametric Kruskal–
Wallis test was adopted to test for statistically significant
differences in the p(1-p) index among the three groups of
species with different conservation interest (A, N, and P).
All analyses were performed with R version 3.0.2 (R Core
Team 2013), in particular ‘‘vegan’’ package (Oksanen et al.
2013).
3 Results
None of the species was recorded in all the sampled plots,
nor in all the PAs. The species with the highest frequency
at the plot scale was Hedera helix (298 plots, 49.3 % of the
total), and the species with the highest frequency at the PA
scale were Hedera helix, Quercus cerris, and Brachypo-
dium rupestre (19 PAs, 90.5 % of the total). The species
classified as protected were 95, being 9.1 % of the total
richness (mean per plot = 1.3; mean per PA = 12.0),
while 48 species were classified as alien (4.6 %; mean per
plot = 0.5; mean per PA = 6.2) and 898 as unprotected
natives (86.3 %; mean per plot = 29.7; mean per
PA = 208.2).
The classification of species common and rare showed
markedly different results, according to the different
adopted criteria (Table 1), with the number of species
classified as common, at the plot and PA scales, decreasing
from the first to the third approach.
The number of common species classified as protected
was significantly lower than expected, whereas the number
of rare species classified as protected was significantly
higher than expected (Table 2). This pattern was observed
at the both plot and PA scales, for the two first
Table 1 Number of species classified as common (CPl1, CPl5, CPlq)
or rare (RPl1, RPl5, RPlq) at plot scale, and as common (CPA1,
CPA3, CPAq) or rare (RPA1, RPA3, RPAq) at PA scale, according to
the three different criteria used
Pl1 Pl5 Plq
CPl1 RPl1 CPl5 RPl5 CPlq RPlq
Mean 31.1 0.4 29.6 1.9 25.4 6.1
Range 0–118 0–6 0–107 0–15 0–90 0–33
Tot 808 233 511 530 267 774
PA1 PA3 PAq
CPA1 RPA1 CPA3 RPA3 CPAq RPAq
Mean 212.1 14.3 181.9 44.5 131.4 95.0
Range 23–526 0–45 17–419 1–151 8–258 2–312
Tot 741 300 484 557 267 774
Rend. Fis. Acc. Lincei (2014) 25:369–380 371
123

classification criteria. The results of the quartile criterion
were slightly different and showed that, at both the plot and
PA scales, both the number of alien and protected species
classified as common were significantly less than expected
(Table 2).
The species richness values of rare species were statis-
tically related to those of common species, at both the plot
and PA scales (Fig. 1), even if at the plot scale the pre-
dictive capacity was rather low (Fig. 1, upper panels). At
the plot scale (Fig. 1), the capacity to predict the number of
rare species by using the number of common species
increased from the more restrictive criterion to the less
restrictive ones. A much higher predictive capacity is
observed at the PA scale for all the different adopted cri-
teria (Fig. 1). Given the autocorrelated nature of the data
(total species richness is made by summing rare and
common species), the number of common and rare species
were also significant predictors of total species richness, at
both plot and PA scales.
The richness of protected species was statistically
predictable by using the richness of rare and protected
species (Fig. 2). At the plot scale, the predictive capacity
of protected species by rare species was rather low, while
it was higher using common species. At the PA scale the
predictive capacity of protected species by common and
rare species was similar and much higher, but the
covariance in species-area relationships certainly con-
tributed to this.
Table 2 Chi-square test, showing the observed values of alien (A),
protected native (P) and unprotected native (N) species classified as
common (CPl1, CPl5, CPlq) or rare (RPl1, RPl5, RPlq) at plot scale,
and as common (CPA1, CPA3, CPAq) or rare (RPA1, RPA3, RPAq)
at PA scale
Pl1 Pl5 Plq
CPl1 RPl1 CPl5 RPl5 CPlq RPlq
A 27 21 10 38 4 ; 44
P 40 ; 55 : 25 ; 70 : 9 ; 86
N 726 172 476 422 254 644
v2 = 86.97 v2 = 40.56 v2 = 23.85
PA1 PA3 PAq
CPA1 RPA1 CPA3 RPA3 CPAq RPAq
A 24 24 11 37 5 ; 43
P 45 ; 50 : 22 ; 73 : 10 ; 85
N 672 226 451 447 252 646
v2 = 42.60 v2 = 36.54 v2 = 19.98
The bold values indicate statistically significant values, and if the
observed number is significantly higher (:) or lower (;) than expected
Fig. 1 Regression graphs showing the predictive power of rare
species (RPl1, RPl5, RPlq, RPA1, RPA3, RPAq) by common species
(CPl1, CPl5, CPlq, CPA1, CPA3, CPAq), according to the different
criteria used for defining rare species (see ‘‘Methods’’ for details), at
both the plot (Pl) and the PA scales. Pl1: RPl1 = 0.02 CPl1-0.160,
R2 = 0.16; Pl5: RPl5 = 0.07 CPl5-0.23, R2 = 0.26; Plq:
RPlq = 0.27 CPlq-0.79, R2 = 0.432; PA 1: RPA1 = 0.09 CPA1-
3.99, R2 = 0.787; PR3: RPA3 = 0.34 CPA3-17.12, R2 = 0.847;
PAq: RPAq = 1.07 CPAq-45.88, R2 = 0.843. All values were
statistically significant p \ 0.001
372 Rend. Fis. Acc. Lincei (2014) 25:369–380
123

The richness of alien species was statistically predict-
able by using the richness of rare and protected species
(Fig. 3), but with a different pattern than protected species.
Contrary to the protected species, the richness of alien
species was better predicted by rare species than by com-
mon species at the plot scale. At the PA scale, the
Fig. 2 Regression graphs showing (in the first and third lines) the
predictive power of protected species (P) by rare species (RPl1, RPl5,
RPlq, RPA1, RPA3, RPAq), according to the different criteria used for
defining rare species (see ‘‘Methods’’ for details), at both the plot and the
PA scales. Pl1: P = 0.33 RPl1 ? 0.14, R2 = 0.037; Pl5: P = 0.13
RPl5 ? 1.01, R2 = 0.059; Plq: P = 0.08 RPlq ? 0.77, R2 = 0.124;
PA1: P = 0.66 RPA1 ? 2.62, R2 = 0.875; PA3: P = 0.21
RPA3 ? 2.57, R2 = 0.852; PAq: P = 0.10 RPAq ? 2.02, R2 =
0.848. All values were statistically significant p \ 0.001. Regression
graphs showing (in the second and fourth lines) the predictive power of
protected species (P) by common species (CPl1,CPl5, CPlq, CPA1,
CPA3, CPAq), according to the different criteria used for defining
common species (see ‘‘Methods’’ for details), at both the plot and the PA
scales. Pl1: P = 0.04 CPl1 ? 0.12, R2 = 0.250; Pl5: P = 0.04
CPl5 ? 0.10, R2 = 0.255; Plq: P = 0.05 CPlq ? 0.04, R2 = 0.259;
PA1: P = 0.06 CPA1-1.42, R2 = 0.862; PA3: P = 0.08 CPA3-
2.17, R2 = 0.852; PAq: P = 0.12 CPAq-4.01, R2 = 0.838. All values
were statistically significant p \ 0.001
Rend. Fis. Acc. Lincei (2014) 25:369–380 373
123

Fig. 3 Regression graphs showing (in the first and third lines) the
predictive power of alien species (A) by rare species (RPl1, RPl5,
RPlq, RPA1, RPA3, RPAq), according to the different criteria used
for defining rare species (see ‘‘Methods’’ for details), at both the plot
(Pl) and the PA scales. Pl1: A = 0.35 RPl1 ? 0.38, R2 = 0.112; Pl5:
A = 0.1366 RPl5 ? 0.25, R2 = 0.159; Plq: A = 0.18 RPlq ? 0.05,
R2 = 0.146; PA1: A = 0.39 RPA1 ? 0.69, R2 = 0.715; PA3:
A = 0.14 RPA3 ? 0.17, R2 = 0.826, PAq: A = 0.07 RPAq-0.25,
R2 = 0.839. Regression graphs showing (in the second and fourth
lines) the predictive power of alien species (A) by common species
(CPl1,CPl5, CPlq, CPA1, CPA3, CPAq), according to the different
criteria used for defining common species (see ‘‘Methods’’ for
details), at both the plot (Pl) and the PA scales. Pl1:
A = 0.01 CPl1 ? 0.19, R2 = 0.051; Pl5: A = 0.01 CPl5 ? 0.23,
R2 = 0.039; Plq: A = 0.01 CPlq ? 0.29, R2 = 0.022; PA1:
A = 0.04 CPA1-2.12, R2 = 0.781; PA3: A = 0.05 CPA3-2.42,
R2 = 0.743; PAq: A = 0.07 CPAq-0.25, R2 = 0.839. All values
were statistically significant p \ 0.001
374 Rend. Fis. Acc. Lincei (2014) 25:369–380
123
predictive capacity of alien species by common and rare
species at PA scale was similar and much higher, but even
in this case the covariance in the species-area relationships
certainly contributed to this.
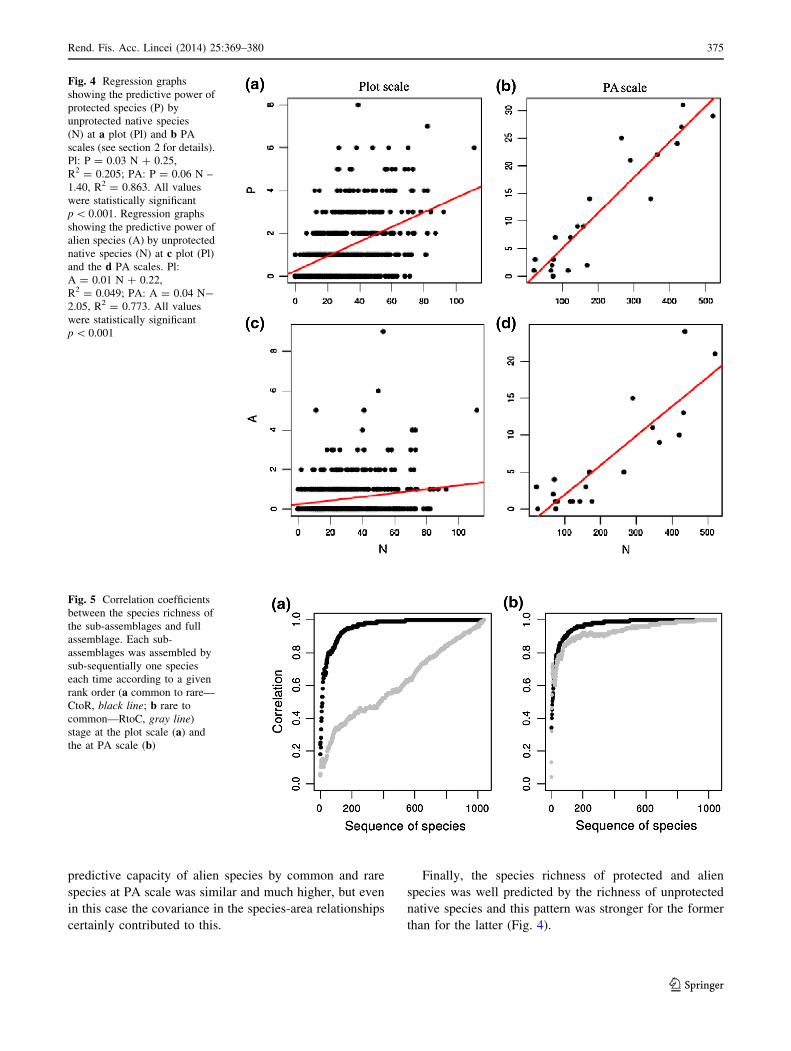
Finally, the species richness of protected and alien
species was well predicted by the richness of unprotected
native species and this pattern was stronger for the former
than for the latter (Fig. 4).
Fig. 4 Regression graphs
showing the predictive power of
protected species (P) by
unprotected native species
(N) at a plot (Pl) and b PA
scales (see section 2 for details).
Pl: P = 0.03 N ? 0.25,
R2 = 0.205; PA: P = 0.06 N –
1.40, R2 = 0.863. All values
were statistically significant
p \ 0.001. Regression graphs
showing the predictive power of
alien species (A) by unprotected
native species (N) at c plot (Pl)
and the d PA scales. Pl:
A = 0.01 N ? 0.22,
R2 = 0.049; PA: A = 0.04 N-
2.05, R2 = 0.773. All values
were statistically significant
p \ 0.001
Fig. 5 Correlation coefficients
between the species richness of
the sub-assemblages and full
assemblage. Each sub-
assemblages was assembled by
sub-sequentially one species
each time according to a given
rank order (a common to rare—
CtoR, black line; b rare to
common—RtoC, gray line)
stage at the plot scale (a) and
the at PA scale (b)
Rend. Fis. Acc. Lincei (2014) 25:369–380 375
123

The correlation coefficients between the species richness
of each of the sub-assemblages and the species richness of
the full assemblages increased with different patterns along
the C to R and R to C sequences (Fig. 5). Common species
contributed more to species richness patterns than rare
species, creating most of spatial structure in richness pat-
terns. At the plot scale, the correlation coefficients for the C
to R sequence were much higher than those achieved in the
R to C sequence (Fig. 5a). Thus, common species gave a
much closer approximation to the overall species richness
patterns than rare species; only 6.3 % of common species
were needed in the C to R to achieve a correlation coeffi-
cient of 0.8 (p \ 0.001, Fig. 5a), and 11.2 % to reach a
correlation coefficient of 0.9 (p \ 0.001, Fig. 5a). On the
other hand, the correlation coefficient of R to C sequence
increased slowly following an almost linear curve and it
was necessary to include 71.6 % of rare species into the
sub-assemblages to achieve a correlation coefficient of 0.8
(p \ 0.001, Fig. 5a), and 85.5 % to reach a correlation
coefficient of 0.9 (p \ 0.001, Fig. 5a). A less clear
difference between the C to R and sequences was observed
at the PA scale: 4.3 and 8.4 % of the common species were
required to reach correlation coefficient of 0.8 (p \ 0.001,
Fig. 5b) and 0.9 (p \ 0.001, Fig. 5b) in C to R sequence,
while 6.8 and 17.6 % of rare species were needed to
achieve correlation coefficient of 0.8 (p \ 0.001, Fig. 5b)
and 0.9 (p \ 0.001, Fig. 5b) in the R to C sequence.
Common and rare species differed in the values of the
information content about variability of species richness, as
shown by the values of the p(1-p) index (Fig. 6). The non
parametric Mann–Whitney test revealed statistically signifi-
cant differences between common and rare species at both the
plot scale and PA scale (df = 1; p \ 0.001). Even the groups
of species with different conservation values (A, N, and P)
provided statistically significant different values of the
information content p(1-p) (Fig. 7). In particular, the Krus-
kal–Wallis test revealed statistically significant differences at
both the plot scale and at PA scale (df = 2, p \ 0.001), with
the unprotected native species having higher values of the
index than protected and alien species (Fig. 7).
Fig. 6 Box-plot of values of the p(1-p) index between common and rare species at both plot (ICPl,1 IRPl1, ICPl5, IRPl5, ICPlq, IRPlq) and PA
scale (ICPA1, IRPA1, ICPA3, IRPA3, ICPAq, IRPAq)
376 Rend. Fis. Acc. Lincei (2014) 25:369–380
123

4 Discussion
4.1 Commonness and rarity
A growing evidence suggests that common rather than rare
species shape the overall distribution patterns of species
richness (Vazquez and Gaston 2004; Gaston and Fuller
2007; Gaston 2008; Perez-Quesada and Brazeiro 2012).
This is not intuitive, since local and regional assemblages
are normally composed by numerous rare species and few
common ones (Colwell and Coddington 1994; Gotelli and
Colwell 2001). Despite some differences in the operational
division of species into common and rare, according to
their occurrences into the plots or protected areas, this
study showed that a very high proportion of species are rare
but the common ones are most responsible for richness
patterns, as already reported by other studies (Jiguet and
Julliard 2006; Pearman and Weber 2007). This study also
confirmed the rule that in a regional survey most of the
recorded species are rare and few are common (Pielou
1969; Gaston 1994): this pattern is known as the ‘‘Law of
infrequency’’ (Raunkiaer 1934; Palmer 1995) and it has
negative implications for biodiversity surveying and
recording, since any of such programs will inevitably miss
rare species (Gaston 1994; Chiarucci et al. 2011). The
recording of rare species also depend on the adopted
sampling approach, as well as on the abundance or
occurrence data which are collected (Gotelli and Colwell
2001). Specific sampling designs for recording rare species
are almost impossible, for both practical and biological
reasons (Elzinga et al. 2001). So, rare species can only be
defined as those with abundance or occurrence values
below arbitrary thresholds in a given sample or data col-
lection. There are also additional caveats which affect
rarity measures in observational studies, as those repre-
sented by bias in sampling design (Chiarucci 2007) and
statistical artefacts (Palmer et al. 2008).
Biologists frequently lump various groups of species
under the term rare, partly because a consistent vocabulary
for the types of rarity is still missing, or not globally
accepted, and this may obscure important features of a
diverse group (Rabinowitz et al. 1986; Gaston 1994). In
addition to definition arguments, it is really difficult to
recognise common and rare species even because changes
in the rarity thresholds affect the definition of rare species
and the relationships they are involved. This is also con-
firmed by the present findings, which demonstrated
important changes in grouping rare species on the basis of
the adopted criteria. Despite the division of species into
common and rare is dependent on the selected criterion,
almost all data sets are intrinsically incomplete and they
are certainly more biased for rare species than for common
species (Guisan et al. 2006; Kreft et al. 2006; Diekmann
et al. 2007; Chiarucci et al. 2008; Hedgren and Weslien
2008). This makes inference on rare species more difficult
than that on common species.
Fig. 7 Box-plot of values of the
p(1-p) index between alien,
native, and protected species at
both plot (IPlA, IPlN, IPlP) and
PA scale (IPAA, IPAN, IPAP)
Rend. Fis. Acc. Lincei (2014) 25:369–380 377
123

Since common species contribute more than rare species
to the species richness patterns, their occurrence is a good
proxy for biodiversity and this can be used because com-
mon species are easier to record than rare species. Com-
mon species can also be used as important indicators of the
overall habitat quality, since they often constitute struc-
turally important components of habitats (McGeoch 2007).
From a practical point of view, focusing the survey on
common and easily recognized species could improve the
understanding of species richness patterns, and their
determinants, with relatively low costs (Vazquez and
Gaston 2004). The greater contribution of common species
to the patterns of species richness can profitably be used to
investigate where common species are absent, rather than
where rare species are present, to determine patterns of
species richness on the basis of easily detectable species
(Lennon et al. 2004). In addition, richness of common
species is well correlated to environmental variables, as it
was reported for birds in South Africa (Jetz and Rahbek
2002) and Britain (Evans et al. 2005), tropical fishes (Mora
and Robertson 2005), neotropical palms (Kreft et al. 2006)
and birds in South America (Rahbek et al. 2007). Given the
relatively large amount of information available for com-
mon species (e.g., life-history and physiological traits),
explanations for the richness patterns may be more pre-
cisely sought than have hitherto been possible.
In this and other studies (e.g., Lennon et al. 2004;
Mazaris et al. 2008), the information contents provided by
the common species were higher than that provided by the
rare species, since the former gave a closer approximation
to overall patterns of species richness than the latter. Then,
given that the richness of common species results to be
more correlated with overall species richness and more
informative, common species could be considered as better
indicators of that overall richness. Pearman and Weber
(2007), using data on butterflies, birds, and vascular plants,
confirmed that sampling common species alone can well
describe the spatial patterns of total species richness. This
is apparently contrary to the long-prevailing assumptions
that rare species would make better indicators (Gaston
2008).
This study also confirmed the importance of the scale in
controlling the patterns of commonness and rarity. At small
grain, as the plot scale, the relations between the richness
of common and rare species are significant but often with
low predictive value. On the other hand, at larger grains, as
it was the case of the PA scale, the relationships are much
higher. The covariance due to the species–area relation-
ships is likely to be the determinant of the relations
observed at larger scale in this study (see Chiarucci et al.
2012 for an analysis on the same data set).
In summary, it is suggested that common species can be
used as adequate indicators of biodiversity patterns, at
various scales, allowing better understanding of the deter-
minants of species richness patterns. This study showed
that, despite the intensity of relationship changes at plot
and PA scale, the general pattern does not change. Mazaris
et al. (2008) stated that this pattern is independent of the
size of sampling unit. Then, the richness of common spe-
cies can be considered the best predictor of overall species
richness at different spatial scales.
4.2 Protected and alien species
Often the focus of conservation planning and biodiversity
monitoring is limited to groups of species with special
features or management interest, as in the case of protected
(e.g., Regnery et al. 2013; Hochkirch et al. 2013) or alien
(e.g., Mace and Baillie 2007) species. Protected and alien
species are highly heterogeneous groups and can include
species with various degrees of rarity, in dependence of
different mechanisms controlling their distribution and
abundance. The richness patterns of common and rare
species defined according to different criteria have been
found to be correlated one to the other, and both well
related to the richness of protected and alien species.
However, protected and alien species showed different
patterns: the richness of protected species was better rela-
ted to the richness of common species while the richness of
alien species was better related to the richness of rare
species. This is a peculiar pattern, despite some potential
links to the sampling design adopted for collecting the data.
The probabilistic design adopted in this study is certainly
more suited for recording the occurrence of common rather
than rare species. A stratified sampling is an objective,
well-defined, and easy method, free from personal bias
(Chiarucci 2007), and it is suitable in long-term monitoring
programs and for statistical analysis, but it is not effective
for recording those species which have strict habitat
requirement, small range size or population density
(Diekmann et al. 2007), as it can be the case for protected
or alien species.
In fact, both rare and alien species revealed to be rarer
than the unprotected native, and this makes more difficult
to collect good data about their abundance or distribution
and this is likely to be a major limitation for management
and monitoring purposes.
Regarding protected species, the results of this study
suggest that higher the richness of common species higher
is the richness of protected species, confirming the
importance of protecting sites with high species richness
and using common species as the best currency for biodi-
versity estimates. Common species largely determine the
species richness patterns and these are also reflected by the
patterns of protected species richness. The coincidence
between common and protected species can be a sampling
378 Rend. Fis. Acc. Lincei (2014) 25:369–380
123

artefact (Diekmann et al. 2008; Palmer et al. 2008) due to
differences in sampling design and intensity. However,
while the relation and the larger scale (whole protected
area) could be biased because of the covariance in species–
area relationships, the relation at the plot scale is certainly
worth of understanding since this was a well standardised
sampling.
Alien species are group made by a variety of different
species, with extremely different strategies and abundance
patterns; they can be invasive and colonize most of the
habitats, thus being really common, or limited to a few
places where they have been introduced. Species as
Robinia pseudoacacia, Arundo donax, and Ailanthus
altissima are considered to be the major invasive neophyte
species in Tuscany (Arrigoni and Viegi 2011), but they
were almost absent in the data recorded by this survey.
Robinia pseudoacacia is invasive neophyte and has been
recorded in 8 PAs and 17 plots, confirming its abundance.
On the other hand, Arundo donax and Ailanthus altissima
have not been recorded in the present survey, confirming
the low level of alien invasion of this area but also the
low probability of including such alien species in a
probabilistic data collection. Thus, alien species are con-
firmed to be rare in this area, as it was indicated by the
results of the p(1-p) index but also by the lack of some
major alien species in the recorded data. The scarce level
of alien species occurrence is easy to understand for this
specific data, given their focus on a network of protected
areas, which are relatively well preserved (Chiarucci et al.
2012).
Alien species usually occupy more nutrient-rich, human-
modified habitats, and show tighter habitat similarity as a
group, compared with the more variable native species
(Dawson et al. 2011). The difference in the level of habitat
similarity within alien and native species groups may
reflect introduction sources and habitat disturbance events
(Chrobock et al. 2011). Different results, with potentially
many common alien species, could have been found in
more disturbed areas, but there is no much data available
on the distribution of rarity among alien species in com-
parison to native species.
Acknowledgments We thank everyone who contributed to data
collection within the MoBiSIC project, the University of Siena and
the Province of Siena for their support. Moreover, we would like to
thank the Editor and two anonymous referees for valuable comments
and suggestions on a previpus version of the manuscript.
References
Arrigoni PV, Viegi L (2011) La flora vascolare esotica spontaneizzata
della Toscana. Regione Toscana, Firenze
Celesti-Grapow L, Alessandrini A, Arrigoni PV, Banfi E, Bernardo L,
Bovio M, Brundu G, Cagiotti MR, Camarda I, Carli E, Conti F,
Fascetti S, Galasso G, Gubellini L, La Valva V, Lucchese F,
Marchiori S, Mazzola P, Peccenini S, Poldini L, Pretto F, Prosser
F, Siniscalco C, Villani MC, Viegi L, Wilhalm T, Blasi C (2009)
Inventory of the non-native flora of Italy. Plant Biosyst
143:386–430
Chiarucci A (2007) To sample or not to sample? That is the question
for the vegetation scientist. Folia Geobot 42:209–216
Chiarucci A, Bacaro G, Rocchini D (2008) Quantifying plant species
diversity in a Natura 2000 network: old ideas and new proposals.
Biol Conserv 141:2608–2618
Chiarucci A, Bacaro G, Scheiner SM (2011) Old and new challenges
in using species diversity for assessing biodiversity. Phil Trans R
Soc B 366:2426–2437
Chiarucci A, Bacaro G, Filibeck G, Landi S, Maccherini S, Scoppola
A (2012) Scale dependence of plant species richness in a
network of protected areas. Biodivers Conserv 21:503–516
Chrobock T, Kempel A, Fischer M, Kleunen M (2011) Introduction
bias: cultivated alien plant species germinate faster and more
abundantly than native species in Switzerland. Basic Appl Ecol
12:244–250
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity
through extrapolation. Philos Tran R Soc London B 345:101–118
Dawson W, Fischer M, Kleunen M (2011) The maximum relative
growth rate of common UK plant species is positively associated
with their global invasiveness. Global Ecol Biogeogr 20:299–306
Diekmann M, Kuhne A, Isermann M (2007) Random vs non-random
sampling: effects on species abundance, species richness and
vegetation-environment relationships. Folia Geobot 42:179–190
Diekmann M, Dupre C, Kolb A, Metzing D (2008) Forest vascular plants
as indicators of plant species richness: a data analysis of a flora atlas
from northwestern Germany. Plant Biosyst 142:584–593
Elzinga CL, Salzer DW, Willoughby JW, Gibbs JP (2001) Monitoring
plant and animal population. Blackwell Science, Malden
Evans KL, Greenwood JJD, Gaston KJ (2005) Relative contribution
of abundant and rare species to species–energy relationships.
Biol Lett 1:87–90
Gaston KJ (1994) Rarity. Chapman Hall, London
Gaston KJ (2008) Biodiversity and extinction: the importance of
being common. Prog Phys Geogr 32:73–79
Gaston KJ (2011) Common ecology. Bioscience 61:354–362
Gaston KJ, Fuller RA (2007) Biodiversity and extinction: losing the
common and the widespread. Prog Phys Geogr 31:213–225
Gaston KJ, Fuller RA (2008) Commonness, population depletion and
conservation biology. Trends Ecol Evol 23:14–19
Gaston KJ, Chown SL, Evans KL (2008) Ecogeographical rules:
elements of a synthesis. J Biogeogr 35:483–500
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures
and pitfalls in the measurement and comparison of species
richness. Ecol Lett 4:379–391
Guisan A, Broennimann O, Engler R, Vust M, Yoccoz NG, Lehmann
A, Zimmermann NE (2006) Using niche-based models to
improve the sampling of rare species. Conserv Biol 20:501–511
Hartley S, Kunin WE (2003) Scale dependency of rarity, extinction
risk and conservation priority. Conserv Biol 17:1559–1570
Hedgren O, Weslien J (2008) Detecting rare species with random or
subjective sampling: a case study of red-listed saproxylic beetles
in Boreal Sweden. Conserv Biol 22:212–215
Hercos AP, Sobansky M, Queiroz HL, Magurran AE (2012) Local
and regional rarity in a diverse tropical fish assemblage. Proc R
Soc B 280:2012–2076
Hochkirch A, Schmitt T, Beninde J, Hiery M, Kinitz T, Kirschey J,
Matenaar D, Rohde K, Stoefen A, Wagner N, Zink A, Lotters S,
Veith M, Proelss A (2013) Europe needs a new vision for a
Natura 2020 network. Conserv Lett 6:462–467
Hopkinson P, Travis JMJ, Evans J, Gregory R, Telfer MG, Williams
PH (2001) Flexibility and the use of indicator taxa in the
Rend. Fis. Acc. Lincei (2014) 25:369–380 379
123

selection of sites for nature reserves. Biodiv Conserv
10:271–285
Jetz W, Rahbek C (2002) Geographic range size and determinants of
avian species richness. Science 297:1548–1551
Jetz W, Rahbek C, Colwell RK (2004) The coincidence of rarity and
richness and the potential signature of history in centers of
endemism. Ecol Lett 7:1180–1191
Jiguet F, Julliard R (2006) Inferences from common species
communities for selecting conservation areas. Biodivers Conserv
15:799–815
Kreft H, Sommer JH, Barthlott W (2006) The significance of
geographic range size for spatial diversity patterns in Neotrop-
ical palms. Ecography 29:21–30
Lawler JJ, White D, Sifneos JC, Master LL (2003) Rare species and
the use of indicatorgroups for conservation planning. Conserv
Biol 17:875–882
Lennon JJ, Koleff P, Greenwood JJD, Gaston KJ (2004) Contribution
of rarity and commonness to patterns of species richness. Ecol
Lett 7:81–87
Lennon JJ, Beale CM, Reid CL, Kent M, Pakeman RJ (2011) Are
richness patterns of common and rare species equally well
explained by environmental variables? Ecography 34:529–539
Mace GM, Baillie JEM (2007) The 2010 Biodiversity indicators:
challenges for science and policy. Conserv Biol 21:1406–1413
Mazaris A, Tzanopoulos J, Kallimanis A, Matsinos YG, Sgardelis SP,
Pantis JD (2008) The contribution of common and rare species to
plant species richness patterns: the effect of habitat type and size
of sampling unit. Biodivers Conserv 17:3567–3577
McGeoch MA (2007) Insects and bioindication: theory and progress.
In: Stewart AJA, New TR, Lewis OT (eds) Insect conservation
biology, Commonwealth Agricultural Bureau (CABI Interna-
tional).Wallingford: pp 144–74
Mora C, Robertson DR (2005) Causes of latitudinal gradients in
species richness: a test with fishes of the Tropical Eastern
Pacific. Ecology 86:1771–1782
Oksanen J, Guillaume Blanchet F, Kindt R, Legendre P, Minchin PR,
O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H
(2013) vegan: community ecology package. R package version
2.0–9. http://CRAN.R-project.org/package=vegan
Palmer MW (1995) How should one count species. Nat Area J
15:124–135
Palmer MW, McGlinn D, Fridley JD (2008) Artifacts and artifictions
in biodiversity research. Folia Geobot 43:245–257
Pearman PB, Weber D (2007) Common species determine richness
patterns in biodiversity indicator taxa. Biol Conserv
138:109–119
Perez-Quesada A, Brazeiro A (2012) Contribution of rarity and
commonness to patterns of species richness in biogeographic
transitions regions: woody plants of Uruguay. Austral Ecol
38:639–645
Pielou EC (1969) An introduction to mathematical ecology. Wiley,
New York
Pysek P, Richardson DM, Rejmanek M, Webster GL, Williamson M,
Kirschner J (2004) Alien plants in checklist and floras: towards
better communication between taxonomists and ecologists.
Taxon 53:131–143
R Core Team (2013) R: a language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna,
Austria. http://www.R-project.org/
Rabinowitz D (1981) Seven forms of rarity. In: Synge H (ed)
Biological aspects of rare plant conservation. Wiley, Chichester,
pp 205–217
Rabinowitz D, Cairns S, Dillon T (1986) Seven forms of rarity and
their frequency in the flora of the British Isles. In: Soule ME (ed)
Conservation biology: the science of scarcity and diversity.
Sinauer Associates, Sunderland, pp 182–204
Rahbek C, Gotelli NJ, Colwell RK, Entsminger GL, Rangel TFLVB,
Graves GR (2007) Predicting continental-scale patterns of bird
species richness with spatially explicit models. Proc R Soc Lond
B 274:165–174
Raunkiaer C (1934) The life forms of plants and statistical plant
geography. Clarendon Press, Oxford. Soc London B 274:165–174
Regnery B, Couvet D, Kerbiriou C (2013) Offsets and conservation of
the species of the EU habitats and birds directives. Conserv Biol
27:1335–1343
Simberloff D (1998) Flagships, umbrellas and keystones: is single-
species management passe in the landscape era? Biol Conserv
83:247–257
Vazquez LB, Gaston KJ (2004) Rarity, commonness, and patterns of
species richness: the mammals of Mexico. Global Ecol Biogeogr
13:535–542
380 Rend. Fis. Acc. Lincei (2014) 25:369–380
123
Related Documents











