Commensal worm traces and possible juvenile thalassinidean burrows associated with Ophiomorpha nodosa , Pleistocene, southern Brazil Jordi M. de Gibert a, * , Renata G. Netto b , Francisco M.W. Tognoli c , Marcelo E. Grangeiro b,F a Departament d’Estratigrafia, Paleontologia i Geocie `ncies Marines, Universitat de Barcelona. Martı ´ Franque `s s/n, 08028 Barcelona, Spain b Laborato ´rio da Histo ´ria da Vida e da Terra, PPGeo UNISINOS. Av. Unisinos, 950, 93022-000 Sa ˜o Leopoldo RS, Brazil c Curso de Po ´s Graduac ¸a ˜o em Geocie ˆncias, Universidade Estadual Paulista. Av. 24-A, 1515 Bela Vista, CEP 13506-900, Rio Claro SP, Brazil Received 21 September 2004; received in revised form 12 July 2005; accepted 19 July 2005 Abstract The Pleistocene Chuı ´ Formation at Oso ´ rio (Rio Grande do Sul, Brazil) consists of coastal marine and eolian sands, the former containing abundant and well-preserved Ophiomorpha nodosa burrow systems. Detailed ichnological study has revealed interesting features associated with them. Small-sized Ophiomorpha , here assigned to a new ichnospecies, O. puerilis , are interpreted as possible burrows of juvenile thalassinidean crustaceans probably belonging to the same species as the producers of larger O. nodosa. Additionally, helicoidal burrows with thick, concentrically laminated linings are associated with the walls of O. nodosa. They are assigned to the new ichnospecies Cylindrichnus helix , and they are interpreted as dwellings of commensal annelid worms. The association of these three ichnospecies constitutes a fossil example of the role of thalassinideans as ecosystem engineers able to modify their environment and to create new space and resources usable by other organisms. D 2005 Elsevier B.V. All rights reserved. Keywords: Ophiomorpha; Ontogeny; Paleoecology; Ecosystem engineering; Pleistocene; Brazil 1. Introduction Ophiomorpha is one of the best-known trace fossils for paleontologists and sedimentary geologists due to its abundance in Mesozoic and Cenozoic shallow and marginal marine deposits. The ichnogenus designates multiple-branching gallery systems of variable com- plexity characterized by having a thick pelletal lining. Early works revealed the striking similarity between 0031-0182/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2005.07.008 * Corresponding author. Fax: +34 934021340. E-mail addresses: [email protected] (J.M. de Gibert), [email protected] (R.G. Netto), [email protected] (F.M.W. Tognoli). F Deceased, June 2005. Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70– 84 www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology,
Commensal worm traces and possible juvenile thalassinidean
burrows associated with Ophiomorpha nodosa,
Pleistocene, southern Brazil
Jordi M. de Gibert a,*, Renata G. Netto b,
Francisco M.W. Tognoli c, Marcelo E. Grangeiro b,F
a Departament d’Estratigrafia, Paleontologia i Geociencies Marines, Universitat de Barcelona. Martı Franques s/n, 08028 Barcelona, Spainb Laboratorio da Historia da Vida e da Terra, PPGeo UNISINOS. Av. Unisinos, 950, 93022-000 Sao Leopoldo RS, Brazil
c Curso de Pos Graduacao em Geociencias, Universidade Estadual Paulista. Av. 24-A, 1515 Bela Vista, CEP 13506-900, Rio Claro SP, Brazil
Received 21 September 2004; received in revised form 12 July 2005; accepted 19 July 2005
Abstract
The Pleistocene Chuı Formation at Osorio (Rio Grande do Sul, Brazil) consists of coastal marine and eolian sands, the former
containing abundant and well-preserved Ophiomorpha nodosa burrow systems. Detailed ichnological study has revealed
interesting features associated with them. Small-sized Ophiomorpha, here assigned to a new ichnospecies, O. puerilis, are
interpreted as possible burrows of juvenile thalassinidean crustaceans probably belonging to the same species as the producers of
largerO. nodosa. Additionally, helicoidal burrows with thick, concentrically laminated linings are associated with the walls of O.
nodosa. They are assigned to the new ichnospecies Cylindrichnus helix, and they are interpreted as dwellings of commensal
annelid worms. The association of these three ichnospecies constitutes a fossil example of the role of thalassinideans as
ecosystem engineers able to modify their environment and to create new space and resources usable by other organisms.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Ophiomorpha; Ontogeny; Paleoecology; Ecosystem engineering; Pleistocene; Brazil
0031-0182/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.palaeo.2005.07.008
* Corresponding author. Fax: +34 934021340.
E-mail addresses: [email protected] (J.M. de Gibert),
[email protected] (R.G. Netto), [email protected]
(F.M.W. Tognoli).F Deceased, June 2005.
1. Introduction
Ophiomorpha is one of the best-known trace fossils
for paleontologists and sedimentary geologists due to
its abundance in Mesozoic and Cenozoic shallow and
marginal marine deposits. The ichnogenus designates
multiple-branching gallery systems of variable com-
plexity characterized by having a thick pelletal lining.
Early works revealed the striking similarity between
Palaeoecology 230 (2006) 70–84

Fig. 1. (A) Situation of Rio Grande do Sul State. (B) Simplified
geological map of Rio Grande do Sul State. (C) Geological map of
the Coastal Plain of Rio Grande do Sul (PCRS) showing the outcrop
distribution of the Barrier-Lagoon System III and the study area
(star). Modified of Tomazelli and Villwock, 2000.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 71
Ophiomorpha and modern burrows of thalassinidean
crustaceans (Weimer and Hoyt, 1964; Suguio and Mar-
tin, 1976; Frey et al., 1978). Ophiomorpha is a sub-
strate-controlled ichnogenus that occurs almost
exclusively in fine- to medium-grained sandy deposits
(Ekdale, 1992). Although very abundant in shallow
marine environments, some ichnospecies of Ophio-
morpha are known from deeper water settings (e.g.,
Tchoumachenko and Uchman, 2001). The extensive
literature regarding Ophiomorpha includes papers
focused on paleoenvironmental interpretation and ich-
nofabrics (e.g., Pollard et al., 1993; Anderson and
Droser, 1998), paleobiological and neoichnological
aspects (e.g., Frey et al., 1978; Miller and Curran,
2001) and evolutionary paleoecology (e.g., Bottjer et
al., 1988).
Despite being a very well known ichnogenus,
Ophiomorpha often occurs only as cross-sections
that do not allow in depth analysis of important
attributes, such as architecture or relation with other
traces. Some outcrops, however, offer the possibility
of such detailed studies, which reveal interesting fea-
tures of the paleobiology of the tracemaker. The
Pleistocene Chuı Formation in Brazil includes some
outcrops with those characteristics. Ophiomorpha
occurs here in well-sorted loose sands that allow care-
ful cleaning of the burrow systems. The study of
Ophiomorpha nodosa from the Chuı Formation,
besides providing the opportunity to analyze certain
aspects of its 3-dimensional architecture, has revealed
interesting new details about the paleoecology of this
trace fossil. Small-sized Ophiomorpha (O. puerilis
nov. isp.) and helicoidal Cylindrichnus (C. helix
nov. isp.) have been found in direct association with
Ophiomorpha nodosa. They are here interpreted as
burrows of juvenile thalassinidean crustaceans and
commensal worms, respectively. The objective of
this paper is to analyze these features in the light of
biological data on modern thalassinideans and their
burrows.
2. Geological setting
The Chuı Formation crops out in the Coastal Plain
of Rio Grande do Sul (Planicie Costeira do Rio
Grande do Sul, PCRS) in southern Brazil. This plain
extends for about 33,000 km2 along the eastern part of
the state of Rio Grande do Sul, parallel to the present
shoreline (Fig. 1). The PCRS was formed during the
Quaternary by the progradation of sediments deriving
from the western highlands. The proximal part of the
plain consists of alluvial fans fed by the Precambrian
Sul-riograndense Shield and by Paleozoic and Meso-
zoic sedimentary rocks and the volcanic plateau of
Serra Geral. The distal region is composed of four
lagoon-barrier depositional systems (Villwock et al.,
1986; Villwock and Tomazelli, 1995; Tomazelli and
Villwock, 2000). They are known as Lagoon-Barrier
Systems I to IV, and they were formed during high-
stand sea levels related to glacio-eustatic cycles during
the Quaternary. The oldest systems are located to the
west, while the youngest are situated to the east as a
result of the progradation of the plain. Systems I to III
are Pleistocene, and System IV is Holocene (Toma-
zelli and Villwock, 2000).
The Chuı Formation extends all along the coast of
Rio Grande do Sul and it belongs to System III (Figs.
1 and 2). Detailed sedimentologic study of this system
was accomplished by Tomazelli et al. (1982) and
Tomazelli (1985). From base to top, the barrier con-
sists of shallow marine, beach and eolian deposits
composed of quartzose, fine-grained, well-sorted

Fig. 2. Stratigraphic chart of the PCRS. Modified of Tomazelli and
Villwock, 2000.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8472
sand. The lagoonal facies consist of silty and muddy
fine sands with carbonate and ferruginous concretions.
Radiometric dating of fossils in the marine sands
(Martin et al., 1982) and thermoluminescence dating
of eolian sands (Poupeau et al., 1985) coincide in
assigning an age of about 120 ky to System III.
Hence, it corresponds to the last transgressive peak
of the Pleistocene, known as the Cananeia Transgres-
sion (Suguio and Martin, 1978).
3. The Osorio outcrops
The outcrops studied for this project are located in
two adjacent quarries, Jazida Gomes and Transareia,
situated in the surroundings of Osorio in the north-
eastern part of Rio Grande do Sul (Fig. 1). The
sedimentology of these outcrops was previously stu-
died by Tomazelli et al. (1982).
Several sections were made to establish the strati-
graphy of the Chuı Formation at Osorio. A represen-
tative section (Gomes Quarry) is shown in Fig. 3. Two
main units are differentiated: a lower sandy marine
unit and an upper sandy eolian unit. They correspond
to facies B and A, respectively, of Tomazelli et al.
(1982). These authors described an additional under-
lying unit consisting of silty–muddy sands that was
only temporarily exposed.
The marine unit consists of well-sorted fine-
grained mainly quartzose sands with an observed
maximum thickness of 5.5 m. They exhibit horizontal
lamination, and low-angle, herringbone, and planar
cross-stratification, besides symmetrical ripples at
the uppermost part. Trace fossils (Fig. 4) are ubiqui-
tous in both quarries, the most obvious being exten-
sive Ophiomorpha nodosa systems occurring mainly
in the lower half of the unit. Very abundant small
Macaronichnus isp. (no more than 2 mm in diameter)
and rare Diplocraterion parallelum occur in the same
part of the unit. The upper half hosts abundant Rosse-
lia socialis. Ichnologic and sedimentologic data indi-
cate that this unit was deposited in a very shallow
subtidal setting.
The upper eolian unit comprises about 5 m of fine-
grained sands showing planar cross-lamination inter-
calated with several few-centimeter-thick horizons
having higher mud content. A strongly pedogenized
horizon consisting of reddish fine- to medium-grained
sandstones is found at the top of the unit. Insect
burrows are very abundant, including Krausichnus,
?Vondrichnus and wasp cells (cf. Celliforma) (Fig. 5)
(Grangeiro et al., 2003). Isolated meniscate burrows
(cf. Taenidium) are probably related to ?Vondrichnus
burrow systems. These traces originate in the paleo-
soil horizon (and some probably in the muddier inter-
calations as well) and penetrate into the eolian sands.
Root traces, otherwise, are rare or poorly preserved,
suggesting vegetation dominated by grasses and
bushes.
4. Ophiomorpha systems and associated structures
4.1. Ophiomorpha nodosa burrow systems
Ophiomorpha burrow systems in the Chuı For-
mation constitute complex labyrinths with horizontal
to vertical elements and multiple branching. Quali-
tative observations show that horizontal and subhor-
izontal elements are important as constituent of
mazes connected to less-branched vertical shafts
(Fig. 6B). Vertical elements are rectilinear, while
the rest are rectilinear to gently curved. The branch-
ing style of the mazes is mostly Y-shaped but also
T-shaped (Fig. 6A, C).
The burrows are circular in cross-section. They
exhibit a strongly developed pelletal lining consti-
tuted by mud with isolated quartz sand grains. The
lining is externally knobby but internally smooth. It
is constituted by ovoidal to spherical pellets 0.5 to 1
cm in diameter. The pellets are not aligned or clus-
tered, but they are homogeneously distributed around
the burrow without displaying any particular arrange-

Fig. 3. Representative section (Gomes quarry) of the Chuı Formation in the study area showing its main sedimentological and ichnological
characteristics.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 73
ment (Fig. 6D). The internal diameter of the burrows
ranges between 2 and 5 cm, being the average 3.2
cm and the mode 3 cm. The thickness of the lining is
very variable between 0.25 and 1.5 cm with an
average of 0.9 cm and a mode of 1 cm. Arrangement
and shape of pellets in the lining are diagnostic
features to differentiate among Ophiomorpha ichnos-
pecies (Frey et al., 1978; Bromley and Ekdale,
1998). The characteristics described above allow
assignment of the burrow systems from Chuı to the
ichnospecies O. nodosa.
Other elements have been found that allow com-
pletion of the 3-dimensional configuration of the O.
nodosa systems. Chambers are sometimes found
connected to the end of some galleries. These cul-
de-sac chambers are ovoidal in shape (Fig. 6E), with
a length of about 15 cm and an external diameter of
about 8 cm. They exhibit the same type of pelleted
lining as other elements of the burrow system.
Additionally, some elements with very distinct
characteristics have been recognized as well. They
are vertical cylindrical structures with an external
smooth wall and an internal narrow tube (Fig. 7).
The lining is very thick (8–14 mm) compared with
the diameter of the central tube (about 5 mm) (Fig.
7C, D). One specimen has been observed that shows
evidence of slight lateral shifting of the shaft (Fig.
7B–D). The maximum vertical length observed was
17.5 cm. These elements are connected to Ophio-
morpha burrows and are part of the same burrow

Fig. 4. Shallow marine trace fossils from the Chuı Formation. (A) Ophiomorpha nodosa. (B) Small Macaronichnus isp. and O. nodosa (right).
(C) Rosselia socialis. (D) Diplocraterion parallelum (right) and O. nodosa (left).
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8474
system (Fig. 7A). They are comparable with the
constricted burrow entrances of thalassinidean bur-
rows figured by Frey et al. (1978) and Pollard et al.
(1993).
4.2. Small Ophiomorpha
Adjacent to the large burrows, very small Ophio-
morpha locally occur (Fig. 8A). They have external
diameters between 1.6 and 3.3 mm (average 2.4
mm). The configuration of these small Ophiomor-
Fig. 5. Insect burrows from the continental part of the Chuı Formation.
pha is rather simple, consisting of a rectilinear or
gently curved tube, most times parallel to the large
adjacent Ophiomorpha, and sometimes ending in a
slightly enlarged chamber (Fig. 8D). The lining of
these burrows consists of cylindrical round-ended
pellets, about 1 mm in length and 0.5 mm in
diameter. The pellets consist of mud with isolated
quartz grains. They constitute a single-layer lining
(Fig. 8E) and display variable orientations that are
parallel, oblique or perpendicular to the burrow axis
(Fig. 8B, C).
(A) Krausichnus isp. (B) ?Vondrichnus isp. (C) Possible bee nest.
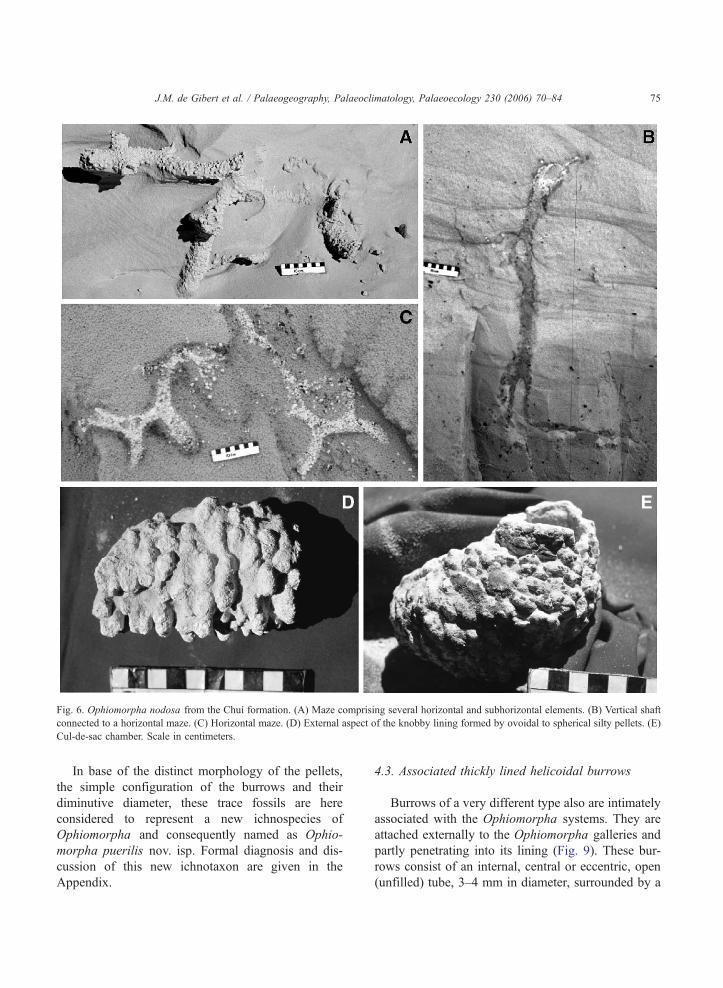
Fig. 6. Ophiomorpha nodosa from the Chuı formation. (A) Maze comprising several horizontal and subhorizontal elements. (B) Vertical shaft
connected to a horizontal maze. (C) Horizontal maze. (D) External aspect of the knobby lining formed by ovoidal to spherical silty pellets. (E)
Cul-de-sac chamber. Scale in centimeters.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 75
In base of the distinct morphology of the pellets,
the simple configuration of the burrows and their
diminutive diameter, these trace fossils are here
considered to represent a new ichnospecies of
Ophiomorpha and consequently named as Ophio-
morpha puerilis nov. isp. Formal diagnosis and dis-
cussion of this new ichnotaxon are given in the
Appendix.
4.3. Associated thickly lined helicoidal burrows
Burrows of a very different type also are intimately
associated with the Ophiomorpha systems. They are
attached externally to the Ophiomorpha galleries and
partly penetrating into its lining (Fig. 9). These bur-
rows consist of an internal, central or eccentric, open
(unfilled) tube, 3–4 mm in diameter, surrounded by a
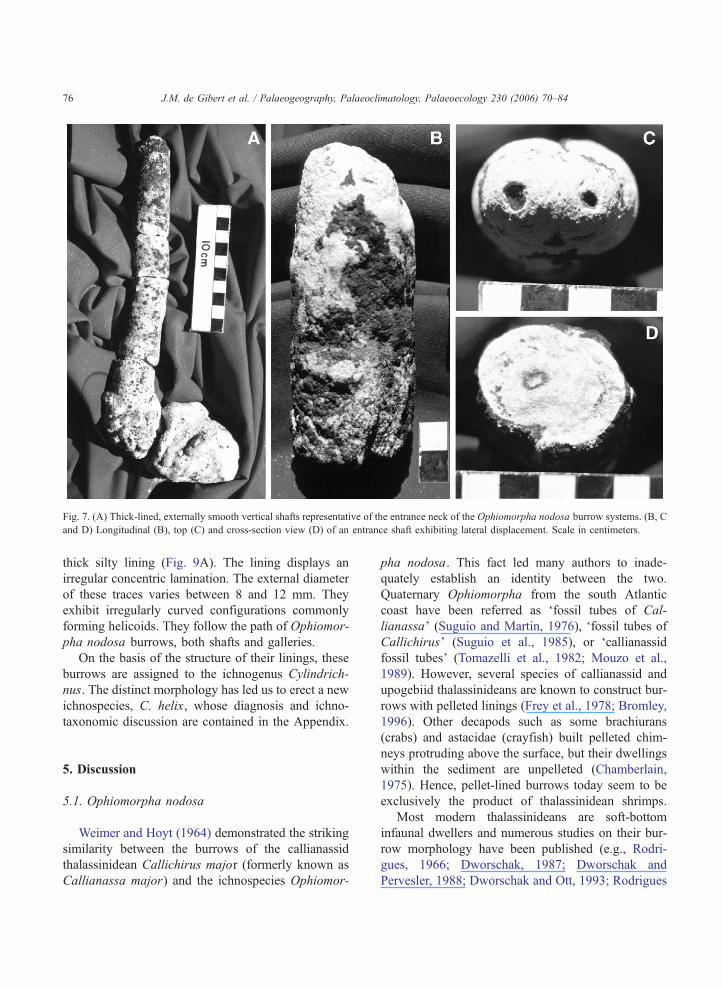
Fig. 7. (A) Thick-lined, externally smooth vertical shafts representative of the entrance neck of the Ophiomorpha nodosa burrow systems. (B, C
and D) Longitudinal (B), top (C) and cross-section view (D) of an entrance shaft exhibiting lateral displacement. Scale in centimeters.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8476
thick silty lining (Fig. 9A). The lining displays an
irregular concentric lamination. The external diameter
of these traces varies between 8 and 12 mm. They
exhibit irregularly curved configurations commonly
forming helicoids. They follow the path of Ophiomor-
pha nodosa burrows, both shafts and galleries.
On the basis of the structure of their linings, these
burrows are assigned to the ichnogenus Cylindrich-
nus. The distinct morphology has led us to erect a new
ichnospecies, C. helix, whose diagnosis and ichno-
taxonomic discussion are contained in the Appendix.
5. Discussion
5.1. Ophiomorpha nodosa
Weimer and Hoyt (1964) demonstrated the striking
similarity between the burrows of the callianassid
thalassinidean Callichirus major (formerly known as
Callianassa major) and the ichnospecies Ophiomor-
pha nodosa. This fact led many authors to inade-
quately establish an identity between the two.
Quaternary Ophiomorpha from the south Atlantic
coast have been referred as dfossil tubes of Cal-
lianassaT (Suguio and Martin, 1976), dfossil tubes ofCallichirusT (Suguio et al., 1985), or dcallianassidfossil tubesT (Tomazelli et al., 1982; Mouzo et al.,
1989). However, several species of callianassid and
upogebiid thalassinideans are known to construct bur-
rows with pelleted linings (Frey et al., 1978; Bromley,
1996). Other decapods such as some brachiurans
(crabs) and astacidae (crayfish) built pelleted chim-
neys protruding above the surface, but their dwellings
within the sediment are unpelleted (Chamberlain,
1975). Hence, pellet-lined burrows today seem to be
exclusively the product of thalassinidean shrimps.
Most modern thalassinideans are soft-bottom
infaunal dwellers and numerous studies on their bur-
row morphology have been published (e.g., Rodri-
gues, 1966; Dworschak, 1987; Dworschak and
Pervesler, 1988; Dworschak and Ott, 1993; Rodrigues
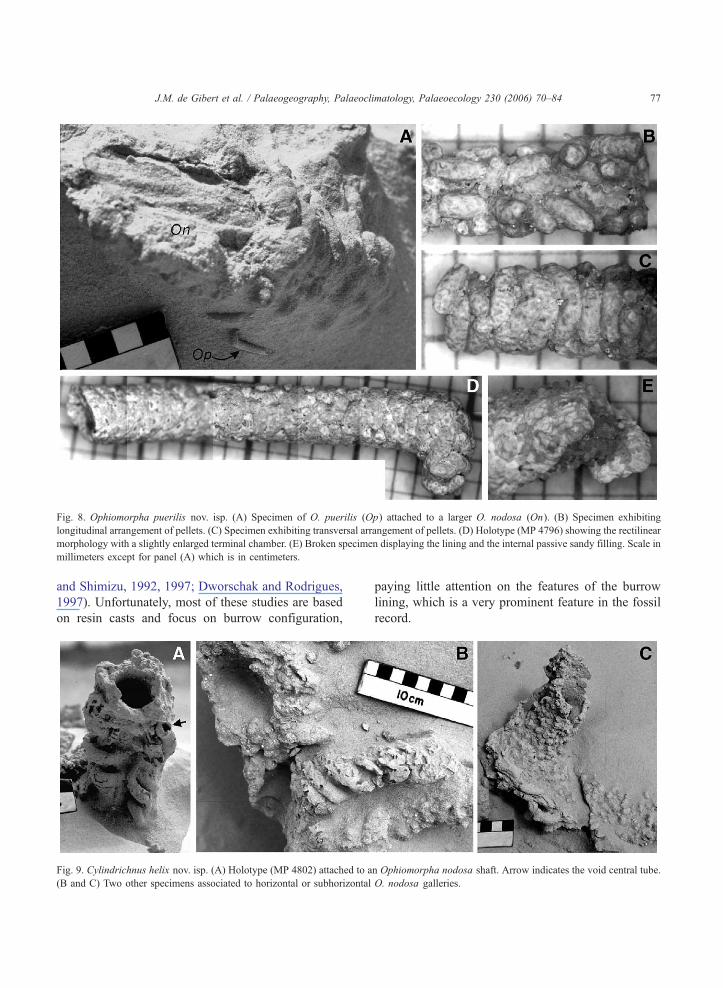
Fig. 8. Ophiomorpha puerilis nov. isp. (A) Specimen of O. puerilis (Op) attached to a larger O. nodosa (On). (B) Specimen exhibiting
longitudinal arrangement of pellets. (C) Specimen exhibiting transversal arrangement of pellets. (D) Holotype (MP 4796) showing the rectilinear
morphology with a slightly enlarged terminal chamber. (E) Broken specimen displaying the lining and the internal passive sandy filling. Scale in
millimeters except for panel (A) which is in centimeters.
J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 77
and Shimizu, 1992, 1997; Dworschak and Rodrigues,
1997). Unfortunately, most of these studies are based
on resin casts and focus on burrow configuration,
Fig. 9. Cylindrichnus helix nov. isp. (A) Holotype (MP 4802) attached to a
(B and C) Two other specimens associated to horizontal or subhorizontal
paying little attention on the features of the burrow
lining, which is a very prominent feature in the fossil
record.
n Ophiomorpha nodosa shaft. Arrow indicates the void central tube.
O. nodosa galleries.

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8478
Griffis and Suchanek (1991) studied the burrows of
44 modern species of thalassinideans and observed
that they construct species-specific burrows. These
authors proposed a simple model to classify their
architecture and trophic mode. Although this model
has been criticized (Nickell and Atkinson, 1995) for
being too simplistic, it provides useful basis on which
to analyze the Ophiomorpha from Chuı Formation at
Osorio.
Griffis and Suchanek (1991) classified thalassini-
dean burrows in six types. Type 4 is the one that
compares best with Chuı Ophiomorpha. The burrow
systems grouped within this type are dprimarily reti-
culate branches extending horizontally from a long
vertical shaft.T They have a smooth, hard burrow wall
and distinctively narrow shafts at the burrow opening.
All these features are found in the Ophiomorpha from
Osorio. The five species that the authors listed as
producers of Type 4 burrows belong to the family
Callianassidae: Callichirus major, Callianassa guas-
sutinga, C. jamaicense, C. louisianensis and Glyp-
turus jousseaumei. The first three species are known
from the modern Brazilian coast (Rodrigues, 1966;
Rodrigues and Shimizu, 1997), although they are not
present in the shores of Rio Grande do Sul, where
thalassinideans are today represented by the species
Sergio mirim.
The trophic significance of Type 4 burrows is not
clear as Griffis and Suchanek (1991) recognized. The
deep branching pattern suggests some sort of sedi-
ment processing (deposit feeding), but the hard walls
point to rather permanent burrows favoring an inter-
pretation of sea-grass harvesting (although no sea-
grass was found on them) or suspension feeding
behavior of the dweller. Pending new data, the men-
tioned authors interpreted these burrow systems as
being constructed by suspension feeders that may
complete their diet with some sort of wall grazing.
Considering all above, we interpret that the trace-
maker of the Chuı Ophiomorpha was a thalassini-
dean shrimp, probably a callianassid, with a
complex feeding behavior based mainly in sus-
pended material.
5.2. Ophiomorpha puerilis
The small Ophiomorpha puerilis and the larger
O. nodosa are both pellet-lined open burrows, and
so they share the same essential constructional fea-
tures. However, they exhibit two main differences.
The first is the shape of the pellets, subcylindrical
in the first, and ovoidal to spherical in the second.
The second difference is the unbranched and simple
configuration of the small burrows. These differ-
ences may have resulted from different burrow
architects but they may also be due to ontogenetic
changes within a single species. Both ichnospecies
occur not only in the same facies but also in direct
association with one another. Although burrow con-
nections between them have not been observed, the
occurrence of the small burrows on the surround-
ings of the larger ones suggests that connections
existed.
The pellets found in O. puerilis are similar in
morphology to the fecal pellets of some polychaetes
(Schafer, 1972; Bayuk and Radwanski, 1979). Never-
theless, annelid excrements are rather find as aggre-
gates, trails or burrow fills and not known as
forming part of a constructed lining. Pelleted linings
seem to be an exclusive feature of thalassinidean
burrows (see discussion above). Thus, although a
polychaete origin for O. puerilis cannot be comple-
tely ruled out, we believe that it is most likely that
this ichnospecies was produced by thalassinideans.
The tracemakers could have been juveniles of the O.
nodosa producers or a completely different species
of small-sized crustaceans. We favor the first hypoth-
esis given the direct association between the two
types of burrows and the information available on
the burrowing behavior of modern and ancient tha-
lassinideans (discussed below). Hence, we consider
that juveniles and adults of a single species were the
tracemakers of smaller and larger Ophiomorpha,
respectively.
Nevertheless, the distribution of sizes of Ophio-
morpha (puerilis and nodosa) from the Chuı Forma-
tion is not continuous. There is an important gap
between small (1.6–3.3 mm in diameter) and large
burrows (2–5 cm in diameter). Intermediate forms (8–
10 mm in diameter) are not absent but very rare.
Tamaki and Ingole (1993) and Berkenbusch and Row-
den (1998) recorded the existence of bimodality on
populations of Callianassa japonica in Japan and
Callianassa filholi in New Zealand, respectively.
The two size modes corresponded to juvenile and
adult animals and resulted from the existence of a

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 79
period of recruitment during the year. However, in the
fossil record, we should expect this situation to have
been modified by the effect of time averaging. Hence,
the bimodality of the fossil Ophiomorpha from Chuı
may have been the result of a high mortality of
juveniles or of the abandonment of juvenile dwellings
to occupy the parental burrows.
Modern burrows of juvenile thalassinideans asso-
ciated with their adults have been reported by Forbes
(1973; Callianassa kraussi), Frey and Howard
(1975; Upogebia affinis), Swinbanks (1981; Callia-
nassa californiensis) and Tamaki and Ingole (1993;
Callianassa japonica). In the case of U. affinis, two
possible scenarios were suggested for this situation
(Frey and Howard, 1975): (a) larvae development
within the parent burrow or (b) settlement of plank-
tonic larvae within burrows. The first would imply
some sort of parental care in the sense it is described
by Clutton-Brock (1991). Thiel (1999) demonstrated
how permanence in parent burrows increases the
survival chances of juveniles of two amphipod crus-
tacean species. In the second scenario, the presence
of juveniles within burrows would simply be a sec-
ondary consequence of settlement in an area with
open holes leading into the burrows.
Most thalassinideans have planktonic larvae, but
Forbes (1973) showed that the larvae of C. kraussi
were not capable of swimming, and so he interpreted
that they stayed in the parental burrows until they
were capable of burrowing at the juvenile stage.
Thus, the burrows of the juveniles protruded as
extensions from their parental burrow. In Forbes’
(1973) and Frey and Howard’s (1975) papers, juve-
nile burrows were described as originating in adult
burrow chambers that could be interpreted as brood
chambers. The small Ophiomorpha from Osorio
occur isolated and not associated with adult burrow
enlargements. That supports the interpretation of set-
tlement of planktonic larvae within adult burrows.
Several examples of juvenile–adult association are
known in the fossil record of thalassinidean or other
crustacean burrows. From the Cretaceous of Utah,
Howard (1966) described associations of very small
and dnormal sizeT Thalassinoides, and Frey and
Howard (1975) recorded Ophiomorpha burrows
locally dencrustedT by clusters of tiny thalassinoid
burrows. Curran (1976) and Curran and Frey (1977)
interpreted bulbous enlargements in Ophiomorpha
burrows with small radiating tubules as dpossiblecallianassid brood structuresT in the Pleistocene of
North Carolina. Verde and Martınez (2004) recorded
similar structures in the Miocene of Uruguay con-
nected to Thalassinoides and Ophiomorpha burrows.
These authors erected a new ichnogenus and ichnos-
pecies, Maiakarichnus currani to name these struc-
tures. Gibert (1996) and Gibert et al. (1999)
interpreted the association of small and large Sinu-
sichnus sinuosus as connected juvenile and adult
crustacean burrows. Curran (1985) described small
Ophiomorpha shafts (5–7.5 mm in diameter) from
the Cretaceous Englishtown Formation of Delaware
(U.S.A.). These burrows were found in the same
facies as normal size O. nodosa systems although,
in contrast to the case of O. puerilis, not directly
connected.
Studies indicate the burrowing capabilities of
modern juvenile thalassinideans and demonstrate
that their dwellings may merge with adult burrow
systems. The fossil examples demonstrate that this
situation can be tracked back to the Cretaceous.
Hence, the example of the Pleistocene Ophiomorpha
from the Chuı Formation is an additional contribu-
tion to understanding the burrowing capabilities of
juvenile thalassinidean. Nevertheless, other hypoth-
esis (polychaete or different crustacean species origin
for O. puerilis) cannot be totally rejected.
5.3. Cylindrichnus helix
These trace fossils always occur adjacent to large
Ophiomorpha burrows and are partly excavated into
their lining. They have never been found in isola-
tion. This fact suggests that the association is not
casual.
This case is different from that of Ophiomorpha
puerilis. The thickly lined helicoidal burrows (Cylin-
drichnus helix) exhibit very different constructional
features and configuration from that of Ophiomorpha,
which makes it very unlikely that both have been
produced by the same species. C. helix was probably
produced by a worm, as it shows a concentric lining
comparable to that of the burrow of the terebellid
polychaete Amphitrite ornata (Aller and Yingst,
1978) and a helicoidal shape such as the traces pro-
duced by the capitellid Notomastus latericeus (Rein-
eck et al., 1967; Bromley, 1996). The thick lining with

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8480
irregular concentric layers suggests continuous re-
excavation of the burrow, probably by grazing on
the burrow walls.
The restriction of occurrence of C. helix to the
surroundings of Ophiomorpha linings suggests that
their tracemakers had a preference for this location.
Studies of the lining and surroundings of the burrows
of three different species of Callianassa (C. kraussi in
Branch and Pringle, 1987; C. trilobata in Dobbs and
Guckert, 1988; C. tyrrhena in Dworschak, 2000)
reveal that burrow linings are zones with high bacter-
ial biomass content that decrease rapidly with
increased distance from the burrow. Dworschak
(2000) pointed out that bacterial content in the inner
part of the lining of the burrow was one order of
magnitude higher than that on the sediment surface.
This dbacterial enhancementT can be explained by two
causes (Branch and Pringle, 1987): (a) irrigation of the
burrows allowing oxygenation deep within the sedi-
ment and (b) modification of the organic content of
the sediment by shrimp activity that may incorporate
this when constructing the lining.
Considering these possibilities, it seems likely that
the producer of C. helix inhabited the environs of the
Ophiomorpha burrows in order to benefit from (a)
access to oxygenated waters, (b) a high bacterial
content to feed on, and (c) protection by being
deep in the sediment far from epibenthic and shallow
endobenthic predators. By this interpretation, C.
helix may well represent an example of a fossilized
commensal relationship between its tracemaker and
the thalassinidean producing the Ophiomorpha.
Curran (1985) recorded a similar association of
worm-produced burrows and crustacean burrow net-
works in the Cretaceous Englishtown Formation. He
described two different types of traces associated to
Ophiomorpha nodosa. One of them consisted of sin-
uous, sometimes irregularly branching burrows exca-
vated within the lining of Ophiomorpha. The other
comprised polygonal-patterned burrow networks
sometimes found wrapping around or anchoring in
Ophiomorpha shafts. Curran (1985) interpreted the
first as a probable fossil example of commensalism.
Both types, despite being morphologically distinctive
from C. helix, may record a similar ecologic relation-
ship between a commensal worm benefiting from
constructing its dwelling adjacent to open burrow
systems of crustaceans.
6. Conclusions: thalassinideans as ecosystem
engineers
The association of Ophiomorpha nodosa, O.
puerilis and Cylindrichnus helix in the Pleistocene
Chuı Formation constitutes a fossil example of how
thalassinidean crustaceans modify their environment
favoring the occupation of new ecological niches.
Modern thalassinideans and other crustaceans play
a very important ecological role, particularly in shal-
low marine environments, as a result of their intense
burrowing activity and high-density populations.
They can be considered as true dphysical ecosystemengineersT in the sense of Jones et al. (1994, 1997).
They define ecosystem engineering as the physical
modification, maintenance or creation of habitats by
directly or indirectly controlling the availability of
resources to other organisms.
Curran and Martin (2003) have pointed out the
importance of modern callianassids as ecosystem
engineers in marginal marine environments. They
recorded how the burrowing activity of Glypturus
acanthochirus is responsible for the creation of a
highly mounded topography in intertidal carbonates
in the Bahamas. The mounds are stabilized by micro-
bial mats resulting in a modified substrate suitable for
colonization by other burrowing crustaceans. The
trace fossils from the Chuı Formation are an example
of this effect in deeper tiers. In this case, the activity
of the producers of O. nodosa created new living
space by increasing the area of the water–substrate
interface. Additionally, O. nodosa networks allowed
oxygenated seawater to circulate deep within the sedi-
ment and organic matter to be incorporated in the
burrow wall. Both factors favored microbial activity
in the surroundings of the burrows. This situation
allowed the producers of C. helix to exploit these
resources (oxygen and food) in a deep-tier niche
achieving protection from epifaunal and shallow
infaunal predators. On the other hand, planktonic
larvae of crustaceans could have entered the burrow
networks and constructed their juvenile burrows (O.
puerilis). In the case we considered O. puerilis as
produced by a different species than larger O. nodosa
(a hypothesis that cannot be completely ruled out, as
indicated above), the former would have to be inter-
preted in similar terms than C. helix, that is as the
work of a commensal organism.

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 81
Hence, the construction of the O. nodosa burrow
systems indirectly resulted in the creation of new
space and resources suitable for larval recruitment
and/or commensal occupation. These two effects
have been recognized in rock units back to the Cre-
taceous, revealing the ecological role of thalassini-
deans (and other crustaceans as well) as ecosystem
engineers.
Acknowledgements
The authors wish to thank Marcelo Zagonel de
Oliveira for his assistance in the field. Tony Ekdale
and Richard Bromley contributed with their sugges-
tions to improve the manuscript. This paper is a con-
tribution to the projects 31.00.006/01-0 and 31.00.002/
04-4 of UNISINOS, 524415/1996-0 and 474345/03-3
of CNPq (Brazilian Scientific and Technological
Development Council), BTE 200-0584 of the Spanish
Government and GRC 2001/SGR/00077 of the Gen-
eralitat de Catalunya.
One of the authors, Marcelo E. Grangeiro, passed
away when this paper was close to be definitively
accepted. The rest of the authors wish to dedicate this
work to his memory.
Appendix A. Systematic Ichnology
A.1. Ophiomorpha puerilis nov. isp. Fig. 8
Diagnosis. Small (external diameter less than 4
mm), rectilinear Ophiomorpha, in some cases ending
in a slightly enlarged chamber, with a lining formed of
a single layer of cylindrical, rod-shaped pellets with
rounded ends.
Derivation of name. From the Latin dpuerilisTmeaning dyouthful,T djuvenile.T
Holotype. MP 4796 housed in the Museu de
Paleontologia da UNISINOS, Sao Leopoldo, Rio
Grande do Sul, Brazil.
Paratypes. MP 4794, MP 4798, MP 4799, MP
4804 housed in the same collection as the holotype.
Remarks. The diagnostic feature of the ichno-
genus Ophiomorpha is the presence of a distinct
lining made of agglutinated pelletoidal sediment
(Frey et al., 1978). This feature is present in O.
puerilis despite of its unusual small size for the
ichnogenus. Fursich (1973) and Schlirf (2000) sug-
gested that Ophiomorpha had to be considered a
junior synonym of Spongeliomorpha. Nevertheless,
we retain Ophiomorpha as a valid name because of
the obvious etho-constructional significance of the
pelleted lining.
Ichnospecies of Ophiomorpha are defined on
basis of the characteristics of their linings, particu-
larly the shape and distribution of the pellets (e.g.,
Frey et al., 1978; Bromley and Ekdale, 1998; Uch-
man, 2001). Thus, pellets are regularly ovoidal in O.
nodosa, O. boornensis and O. annulata but they are
differently arranged; uniformly distributed in the
first, grouped in pairs in O. boornensis, and orga-
nized in transverse rows in O. annulata. O. irregu-
laire bears irregular conical pellets, while O. rudis
exhibits a rather irregular distribution of the pelleted
lining. The elongated round-ended morphology of
the pellets in O. puerilis is very distinctive and not
found in any other ichnospecies of Ophiomorpha.
Additionally, its small size (one order of magnitude
smaller than the rest of ichnospecies) and its simple
configuration are also diagnostic features of this
ichnospecies.
Several other ichnogenera have been described as
constituted by pellets similar in size and morphology
to those seen in O. puerilis. Eiserhardt et al. (2001)
revised some of them to conclude that they can be
reduced to two main types: pellet-filled burrows
(Alcyonidiopsis) and epigenic pellet trails (Tomacu-
lum). Nevertheless, Uchman (1999) considered Toma-
culum as a junior synonym of Alcyonidiopsis as well.
Katto (1974) Sakoites yukioia from the Miocene of
Japan, which also superficially resembles O. puerilis,
is also described as a pellet-filled burrow and, so it
should probably be re-assigned to Alcyonidiopsis.
Tibikoia is described as forming irregular aggregates
(Bayuk and Radwanski, 1979). In any case, none of
those ichnotaxa are described as pellet-lined burrows
such as O. puerilis.
A.2. Cylindrichnus helix nov. isp. Fig. 9
Diagnosis. Cylindrichnus exhibiting irregular sinu-
soidal to helicoidal configuration.
Derivation of name. From the helicoidal burrow
morphology.

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8482
Holotype. MP 4802 housed in the Museu de
Paleontologia da UNISINOS, Sao Leopoldo, Rio
Grande do Sul, Brazil.
Paratype. MP 4803 housed in the same collection
than the holotype.
Remarks. The ichnogenus Cylindrichnus was
erected by Howard (1966) to designate burrows with
a thick, concentrically laminated lining. Despite the
common subsequent use of the ichnotaxon, Goldring
(1996) and Goldring et al. (2002) pointed out its
dubious validity. These authors demonstrated that
some concentrically laminated burrows, assignable
to Cylindrichnus, were passively filled (draught fill-
ing) while others had actually constructed linings. The
validity of the ichnogenus is pending of revision of
type (or near-type) material from the Cretaceous of
Utah. Meanwhile, we have decided to conservatively
include the new ichnospecies erected here, helix,
within the ichnogenus because it bears a distinct con-
centrically laminated lining, which is commonly con-
sidered as its diagnostic feature.
The sinuous to helicoidal pattern of the material
studied herein is not known from previous records of
Cylindrichnus (see Goldring, 1996 for revision) and
hence, justifies the erection of a new ichnospecies.
Other helicoidal burrows are obligatorily vertical
(Gyrolithes) or horizontal (Helicodromites, Helico-
lithus) and lack the thick concentric lining. The axes
of the helices in C. helix are parallel to the associated
Ophiomorpha shafts (vertical) and tunnels (inclined to
horizontal), and so, orientation is not diagnostic.
References
Aller, R.C., Yingst, J.Y., 1978. Biogeochemistry of tube-dwellings:
a study of the sedentary polychaete Amphitrite ornata (Leidy).
Journal of Marine Research 36, 201–254.
Anderson, B.G., Droser, M.L., 1998. Ichnofabrics and geometric
configurations of Ophiomorpha within a sequence stratigraphic
framework: an example from the Upper Cretaceous US western
interior. Sedimentology 45, 379–396.
Bayuk, W., Radwanski, A., 1979. Polychaete-attributable faecal
pellets, Tibikoia sanctacrucensis ichnosp. n., from the Korytnica
Clays (middle Miocene; Holy Cross Mountains, Central Poland.
Acta Geologica Polonica 29, 339–344.
Berkenbusch, K., Rowden, A.A., 1998. Population dynamics of the
burrowing ghost shrimp Callianassa filholi on an intertidal
sandflat in New Zealand (Decapoda: Thalassinidea). Ophelia
49, 55–69.
Bottjer, D.J., Droser, M.L., Jablonski, D., 1988. Palaeoenviron-
mental trends in the history of trace fossils. Nature 333,
252–255.
Branch, G.M., Pringle, A., 1987. The impact of the sand prawn
Callianassa kraussi Stebbing on sediment turnover and on
bacteria, meiofauna, and benthic microflora. Journal of Experi-
mental Marine Biology and Ecology 107, 219–235.
Bromley, R.G., 1996. Trace fossils, Biology and Taphonomy, 2a ed.
Chapman & Hall, London.
Bromley, R.G., Ekdale, A.A., 1998. Ophiomorpha irregulaire
(trace fossil); redescription from the Cretaceous of the Book
Cliffs and Wasatch Plateau, Utah. Journal of Paleontology 72,
773–778.
Chamberlain, C.K., 1975. Recent lebensspuren in nonmarine aqua-
tic environments. In: Frey, R.W. (Ed.), The Study of Trace
Fossils. Springer-Verlag, New York, pp. 431–458.
Clutton-Brock, T.H., 1991. Evolution of Parental Care. Princeton
University Press, Princeton.
Curran, H.A., 1976. A trace fossil brood structure of probable
callianassid origin. Journal of Paleontology 50, 249–259.
Curran, H.A., 1985. The trace fossil assemblage of a Cretaceous
nearshore environment: Englishtown Formation of Delaware,
U.S.A. In: Curran, H.A. (Ed.), Biogenic Structures: Their Use
in Interpreting Depositional Environments, Special Publication-
Society of Economic Paleontologists and Mineralogists, vol. 35,
pp. 261–276.
Curran, H.A., Frey, R.W., 1977. Pleistocene trace fossils from North
Carolina (U.S.A.) and their Holocene analogues. In: Crimes,
T.P., Harper, J.C. (Eds.), Trace Fossils, vol. 2. Seel House Press,
Liverpool, pp. 139–162.
Curran, H.A., Martin, A.J., 2003. Complex decapod burrows and
ecological relationships in modern and Pleistocene intertidal
carbonate environments, San Salvador Island, Bahamas. Palaeo-
geography, Palaeoclimatology, Palaeoecology 192, 229–245.
Dobbs, F.C., Guckert, J.B., 1988. Callianassa trilobata (Crustacea:
Thalassinidea) influences abundance of meiofauna and biomass,
composition, and physiologic state of microbial communities
within its burrow. Marine Ecology Progress Series 45, 69–79.
Dworschak, P.C., 1987. Feeding behaviour of Upogebia pusilla and
Callianassa tyrrhena (Crustacea, Decapoda, Thalassinidea).
Investigaciones Pesqueras 51 (Suppl. 1), 421–429.
Dworschak, P.C., 2000. The burrows of Callianassa tyrrhena
(Petagna 1792) (Decapoda: Thalassinidea). Marine Ecology
22, 155–166.
Dworschak, P.C., Ott, J.A., 1993. Decapod burrow in mangrove-
channel and back-reef environments at the Atlantic Barrier Reef,
Belize. Ichnos 2, 277–290.
Dworschak, P.C., Pervesler, P., 1988. Burrows of Callianassa bou-
vieri Nobile 1904 from Safaga (Egypt, Red Sea) with some
remarks on the biology of the species. Senckenbergiana Mar-
itima 20, 1–17.
Dworschak, P.C., Rodrigues, S. de A., 1997. A modern analogue for
the trace fossil Gyrolithes: burrows of the thalassinidean shrimp
Axianassa australis. Lethaia 30, 41–52.
Eiserhardt, K.H., Koch, L., Eiserhardt, W.L., 2001. Revision des
ichnotaxon Tomaculum Groom, 1902. Neues Jahrbuch fur Geo-
logie und Palaontologie Abhandlungen 221, 325–358.

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–84 83
Ekdale, A.A., 1992. Muckcraking and mudslinging: the joys of
deposit-feeding. In: Maples, C.G., West, R.W. (Eds.), Trace
Fossils, Short Courses in Paleontology, vol. 5. Paleontological
Society, Knoxville, pp. 145–171.
Forbes, A.T., 1973. An unusual abbreviated larval life in the estuar-
ine burrowing prawn Callianassa kraussi (Crustacea:Decapo-
da:Thalassinidea). Marine Biology 22, 361–365.
Frey, R.W., Howard, J.D., 1975. Endobenthic adaptations of juve-
nile thalassinidean shrimp. Bulletin of the Geological Society of
Denmark 24, 283–297.
Frey, R.W., Howard, J.D., Pryor, W.A., 1978. Ophiomorpha: its
morphologic, taxonomic and environmental significance. Palaeo-
geography, Palaeoclimatology, Palaeoecology 23, 199–229.
Fursich, F.T., 1973. A revision of the trace fossils Spongeliomor-
pha, Ophiomorpha and Thalassinoides. Neues Jahrbuch fur
Geologie und Palaontologie, Monatshefte, 719–735.
Gibert, J.M. de, 1996. A new decapod burrow system from the NW
Mediterranean Pliocene. Revista Espanola de Paleontologıa 11,
251–254.
Gibert, J.M. de, Jeong, K., Martinell, J., 1999. Ethologic and
ontogenic significance of the trace fossil Sinusichnus sinuosus,
Pliocene, northwestern Mediterranean. Lethaia 32, 31–40.
Grangeiro, M.E., Netto, R.G., Genise, J., Gibert, J.M. de, Tognoli,
F.M.W., 2003. Pleistocene insect trace fossils from coastal plain
of Rio Grande do Sul State, Brazil. 3rd Latinoamerican Sedi-
mentological Congress, Abstracts. p. 174.
Goldring, R., 1996. The sedimentological significance of concen-
trically laminated burrows from the Lower Cretaceous Ca-ben-
tonites, Oxfordshire. Journal of the Geological Society, London
153, 255–263.
Goldring, R., Gruszczynski, M., Gatt, P.A., 2002. A bow-form
burrow and its sedimentological and paleoecological signifi-
cance. Palaios 17, 622–630.
Griffis, R.B., Suchanek, T.H., 1991. A model of burrow architecture
and trophic modes in thalassinidean shrimp (Decapoda: Thalas-
sinidea). Marine Ecology Progress Series 79, 171–183.
Howard, J.D., 1966. Characteristic trace fossils in Upper Cretaceous
sandstones of the Book Cliffs and Wasatch Plateau. Bulletin-
Utah Geological and Mineralogical Survey 80, 35–33.
Jones, C.V., Lawton, J.H., Shachak, M., 1994. Organisms as eco-
system engineers. Oikos 69, 373–386.
Jones, C.V., Lawton, J.H., Shachak, M., 1997. Positive and negative
effects of organisms as physical ecosystem engineers. Ecology
78, 1946–1957.
Katto, J., 1974. A pellet-cord from the Cenozoic (Miocene) of
Higashi-Muro-gun, Wakayama Prefecture, southwest Japan.
Research Reports Kochi University 23, Natural Sciences 4, 1–7.
Martin, L., Bittencourt, A.C.S.P., Villas-Boas, G.S., 1982. Primeira
ocorrencia de corais pleistocenicos da costa brasileira: datacao do
maximo da penultima transgressao. Ciencia da Terra 1, 16–17.
Miller, M.F., Curran, H.A., 2001. Behavioral plasticity of modern
and Cenozoic burrowing thalassinidean shrimp. Palaeogeogra-
phy, Palaeoclimatology, Palaeoecology 166, 219–236.
Mouzo, F.R., Farinati, E.A., Esposito, G.J., 1989. Tubos fosiles de
callianassidos en la playa de Pehuen-Co, Provincia de Buenos
Aires. Primeras Jornadas Geologicas Bonaerenses, Actas, Tan-
dil, pp. 263–274.
Nickell, L.A., Atkinson, J.A., 1995. Functional morphology of
burrows and trophic modes of three thalassinidean shrimp
species, and a new approach to the classification of thalassi-
nidean burrow morphology. Marine Ecology. Progress Series
128, 181–197.
Pollard, J.D., Goldring, R., Buck, S.G., 1993. Ichnofabrics con-
taining Ophiomorpha: significance in shallow-water facies
interpretation. Journal of the Geological Society, London 150,
149–164.
Poupeau, G., Rivera, A., Solıan Jr., E., Vasconcelos, M.B.A., Souza,
J.H., 1985. Datacao por termoluminescencia de depositos are-
nosos costeiros do Rio Grande do Sul: resultados e implicacoes.
28 Simposio Sul-Brasileiro de Geologia, Florianopolis. Socie-
dade Brasileira de Geologia, Anais, p. 403.
Reineck, H.E., Gutmann, W.F., Hertweck, G., 1967. Das Schlick-
gebiet suslich Helgoland als Beispiel rezenter Schelfablagerun-
gen. Senckenbergiana Maritima 3, 185–201.
Rodrigues, S.A., 1966. Estudos sobre Callianassa. Sistematica,
Biologia e Anatomia. PhD thesis, Faculdade de Filosofia, Cien-
cias e Letras, Universidade de Sao Paulo, Brazil.
Rodrigues, S. de A., Shimizu, R.M., 1992. Description of a new
Axianassa (Crustacea: Decapoda: Thalassinidea) from Brazil,
and its larval stage. Proceedings of the Biological Society of
Washington 105, 317–323.
Rodrigues, S. de A., Shimizu, R.M., 1997. Autoecologia de
Callichirus major (Say, 1818) (Crustacea: Decapoda: Thalas-
sinidea). In: Absalao, R.S., Esteves, A.M. (Eds.), Ecologia de
Praias Arenosas do Litoral Brasileiro. Instituto de Biologia-
Universidade Federal do Rio de Janeiro, Rio de Janeiro,
pp. 155–170.
Schafer, W., 1972. Ecology and Palaeoecology of Marine Environ-
ments. Oliver & Boyd, Edinburgh.
Schlirf, M., 2000. Upper Jurassic trace fossils from the Boulonnais
(northern France). Geologica et Palaeontologica 34, 145–213.
Suguio, K., Martin, L., 1976. Presenca de tubos fosseis de
bCallianassaQ nas formacoes quaternarias do litoral paulista e
sua utilizacao na reconstrucao paleoambiental. Boletim do IG-
USP 7, 17–26.
Suguio, K., Martin, L., 1978. Quaternary marine formations of the
states of Sao Paulo and southern Rio de Janeiro. International
Symposium on Coastal evolution in the Quaternary, IGCP
Project 61, Special Publication 1, Sao Paulo.
Suguio, K., Martin, L., Bittencourt, A.C.S.P., Dominguez, J.M.L.,
Flexor, J.M., Azevedo, A.E.G., 1985. Fluctuacoes do nıvel
relativo do mar durante o Quaternario superior ao longo do
litoral brasileiro e suas implicacoes na sedimentacao costeira.
Revista Brasileira de Geociencias 15, 273–286.
Swinbanks, D.D., 1981. Sedimentology photo. Journal of Sedimen-
tary Petrology 51, 1146.
Tamaki, A., Ingole, B., 1993. Distribution of juvenile and adult
ghost shrimps, Callianassa japonica Ortmann (Thalassinidea),
on an intertidal sand flat: intraspecific facilitation as a possible
pattern-generating factor. Journal of Crustacean Biology 13,
175–183.
Tchoumachenko, P., Uchman, A., 2001. The oldest deep-sea Ophio-
morpha and Scolicia and associated trace fossils from the Upper
Jurassic–Lower Cretaceous deep-water turbidite deposits of SW

J.M. de Gibert et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 230 (2006) 70–8484
Bulgaria. Palaeogeography, Palaeoclimatology, Palaeoecology
169, 85–99.
Thiel, M., 1999. Extended parental care in marine amphipods: II.
Maternal protection of juveniles from predation. Journal of
Experimental Marine Biology and Ecology 234, 235–253.
Tomazelli, L.J., 1985. Contribuicao ao conhecimento das facies
de ambiente praial a partir de elementos do Pleistoceno
Costeiro do Rio Grande do Sul. 28 Simposio Sul-Brasileiro
de Geologia, Florianopolis. Sociedade Brasileira de Geologia,
Anais, pp. 325–338.
Tomazelli, L.J., Villwock, J.A., 2000. O Cenozoico no Rio Grande
do Sul: Geologia da Planıcie Costeira. In: Holz, M., De Ros,
L.F. (Eds.), Geologia do Rio Grande do Sul. Edicao CIGO/
UFRGS, Porto Alegre, pp. 375–406.
Tomazelli, L.J., Villwock, J.A., Loss, E.L., Denhardt, E.A., 1982.
Caracterizacao de um deposito praial pleistocenico na Provıncia
Costeira do Rio Grande do Sul. 328 Congresso Brasileiro de
Geologia, Salvador, vol. 4. Sociedade Brasileira de Geologia,
Anais, pp. 1514–1523.
Uchman, A., 1999. Ichnology of the Rhenodanubian Flysch (Lower
Cretaceous–Eocene) in Austria and Germany. Beringeria 25,
67–173.
Uchman, A., 2001. Eocene flysch trace fossils from the Hecho
Group of the Pyrenees, northern Spain. Beringeria 28, 3–41.
Verde, M., Martınez, S., 2004. A new ichnogenus for crustacean
trace fossils from the upper Miocene Camacho Formation of
Uruguay. Palaeontology 47, 39–49.
Villwock, J.A., Tomazelli, L.J., 1995. Geologia Costeira do Rio
Grande do Sul. Notas Tecnicas 8, 1–45.
Villwock, J.A., Tomazelli, L.J., Loss, E.L., Dehnhardt, E.A., Horn,
N.O., Bachi, F.A., Dehnhardt, B.A., 1986. Geology of the Rio
Grande do Sul Coastal Province. In: Rabassa, J. (Ed.), Quatern-
ary of South America and Antarctic Peninsula. A.A. Balkema,
Rotterdam, pp. 79–97.
Weimer, R.J., Hoyt, J.H., 1964. Burrows of Callianassa major Say,
geologic indicators of littoral and shallow neritic environments.
Journal of Paleontology 38, 761–767.
Related Documents











