ORIGINAL PAPER Combined treatment of carfilzomib and z-VAD-fmk inhibits skeletal proteolysis and apoptosis and ameliorates cancer cachexia Qiang Wang • Chunhong Li • Xudong Peng • Qingjie Kang • Dawei Deng • Liuping Zhang • Yueyong Zheng • Chaoyi Wang • Zhongpeng Qiao • Dunwei Guo • Song You • Hua Tang Received: 3 January 2015 / Accepted: 13 February 2015 Ó Springer Science+Business Media New York 2015 Abstract The purpose of the study was to evaluate the therapeutic benefit of treatments with carfilzomib (CFZ) and z-VAD-fmk in a mouse model of cancer-induced cachexia. The model of cancer-associated cachexia was generated by injecting murine C26 adenocarcinoma cells into BALB/C mice. CFZ and z-VAD-fmk were adminis- tered individually or in combination at 5 and 12 days after inoculation. Changes in body weight, gastrocnemius mus- cle mass, tumor burden, spontaneous activity, survival, and metabolic profiles were noted. Also evaluated were the circulatory levels of renin and angiotensin II, and levels of apoptotic, proteolytic, and renin-angiotensin system-asso- ciated markers and transcription factor 2 (ATF2) in gas- trocnemius muscle. The CFZ and z-VAD-fmk treatments were associated with less muscle wasting, reduced tumor burden, modulated metabolism, higher levels of glucose, albumin, and total proteins, and lower levels of triglyceride fatty acids, more spontaneous physical activity, and longer survival in C26-inoculated mice compared with PBS- treated cachectic mice. CFZ and z-VAD-fmk treatments resulted in higher levels of caspase-3 and BAX and lower level of BCL-XL in gastrocnemius muscles and altered the level of proteins in the renin-angiotensin system. The combined treatment administered 5 days after C26 inoculation was more effective than other regimens. Combined treatment with CFZ and z-VAD-fmk early in the development of cachexia was associated with signs of less proteolysis and apoptosis and less severe cachexia in a mouse model of cancer-induced cachexia. Keywords Carfilzomib z-VAD-fmk pATF2 Proteolysis Apoptosis Introduction Cancer cachexia is a multifactorial syndrome characterized by ongoing loss of skeletal muscle mass that is not fully responsive to conventional treatments such as nutritional support [1]. In cancer patients, especially patients with advanced disease, the incidence of cachexia is high, rang- ing from 50 to 80 % [2, 3]. Cachexia not only reduces the quality of life of cancer patients and their caregivers, but also obstructs patients’ response to treatment and reduces their ability to tolerate treatment-associated toxicity [3–5]. Over 30 % of cancer-related deaths can be attributed to cachexia [2]. To identify potential therapies for cachexia and develop effective treatments, it is important to obtain a better understanding of the underlying mechanism of this detrimental syndrome. The most prominent feature of cachexia is skeletal muscle wasting, a process during which excessive prote- olysis of myofibrils occurs. The role of the ubiquitin–pro- teasome system (UPS) in skeletal muscle proteolysis has been well established [6, 7]. In addition, the presence of DNA fragments in skeletal muscle during muscle wasting suggests that an apoptotic pathway is activated in this catabolic state. Caspase-3, one of the key players in apoptosis, was activated at the initial step to break down the myofibril, providing protein substrates for UPS [8–10]. Caspase-3 Q. Wang C. Li X. Peng Q. Kang D. Deng L. Zhang Y. Zheng C. Wang Z. Qiao D. Guo S. You H. Tang (&) Department of General Surgery, The First Affiliated Hospital of Chongqing Medical University, YouYi Road, Yuzhong District, Chongqing 400016, China e-mail: [email protected] 123 Med Oncol (2015) 32:100 DOI 10.1007/s12032-015-0538-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Combined treatment of carfilzomib and z-VAD-fmk inhibitsskeletal proteolysis and apoptosis and ameliorates cancer cachexia
Qiang Wang • Chunhong Li • Xudong Peng • Qingjie Kang • Dawei Deng •
Liuping Zhang • Yueyong Zheng • Chaoyi Wang • Zhongpeng Qiao •
Dunwei Guo • Song You • Hua Tang
Received: 3 January 2015 / Accepted: 13 February 2015
� Springer Science+Business Media New York 2015
Abstract The purpose of the study was to evaluate the
therapeutic benefit of treatments with carfilzomib (CFZ)
and z-VAD-fmk in a mouse model of cancer-induced
cachexia. The model of cancer-associated cachexia was
generated by injecting murine C26 adenocarcinoma cells
into BALB/C mice. CFZ and z-VAD-fmk were adminis-
tered individually or in combination at 5 and 12 days after
inoculation. Changes in body weight, gastrocnemius mus-
cle mass, tumor burden, spontaneous activity, survival, and
metabolic profiles were noted. Also evaluated were the
circulatory levels of renin and angiotensin II, and levels of
apoptotic, proteolytic, and renin-angiotensin system-asso-
ciated markers and transcription factor 2 (ATF2) in gas-
trocnemius muscle. The CFZ and z-VAD-fmk treatments
were associated with less muscle wasting, reduced tumor
burden, modulated metabolism, higher levels of glucose,
albumin, and total proteins, and lower levels of triglyceride
fatty acids, more spontaneous physical activity, and longer
survival in C26-inoculated mice compared with PBS-
treated cachectic mice. CFZ and z-VAD-fmk treatments
resulted in higher levels of caspase-3 and BAX and lower
level of BCL-XL in gastrocnemius muscles and altered the
level of proteins in the renin-angiotensin system. The
combined treatment administered 5 days after C26
inoculation was more effective than other regimens.
Combined treatment with CFZ and z-VAD-fmk early in the
development of cachexia was associated with signs of less
proteolysis and apoptosis and less severe cachexia in a
mouse model of cancer-induced cachexia.
Keywords Carfilzomib � z-VAD-fmk � pATF2 �Proteolysis � Apoptosis
Introduction
Cancer cachexia is a multifactorial syndrome characterized
by ongoing loss of skeletal muscle mass that is not fully
responsive to conventional treatments such as nutritional
support [1]. In cancer patients, especially patients with
advanced disease, the incidence of cachexia is high, rang-
ing from 50 to 80 % [2, 3]. Cachexia not only reduces the
quality of life of cancer patients and their caregivers, but
also obstructs patients’ response to treatment and reduces
their ability to tolerate treatment-associated toxicity [3–5].
Over 30 % of cancer-related deaths can be attributed to
cachexia [2]. To identify potential therapies for cachexia
and develop effective treatments, it is important to obtain a
better understanding of the underlying mechanism of this
detrimental syndrome.
The most prominent feature of cachexia is skeletal
muscle wasting, a process during which excessive prote-
olysis of myofibrils occurs. The role of the ubiquitin–pro-
teasome system (UPS) in skeletal muscle proteolysis has
been well established [6, 7]. In addition, the presence of
DNA fragments in skeletal muscle during muscle wasting
suggests that an apoptotic pathway is activated in this
catabolic state.
Caspase-3, one of the key players in apoptosis, was
activated at the initial step to break down the myofibril,
providing protein substrates for UPS [8–10]. Caspase-3
Q. Wang � C. Li � X. Peng � Q. Kang � D. Deng � L. Zhang �Y. Zheng � C. Wang � Z. Qiao � D. Guo � S. You � H. Tang (&)
Department of General Surgery, The First Affiliated Hospital of
Chongqing Medical University, YouYi Road, Yuzhong District,
Chongqing 400016, China
e-mail: [email protected]
123
Med Oncol (2015) 32:100
DOI 10.1007/s12032-015-0538-6

also cleaves specific 19S proteasome subunits in skeletal
muscle to stimulate proteasome activity [11]. This suggests
that both the UPS and apoptotic pathways are important in
mediating the wasting of skeletal muscle in cachexia.
Interestingly, angiotensin II (ANGII), in the renin-an-
giotensin system (RAS), can activate the UPS and several
proteins in the proteolytic pathway, including caspase 3,
MuRFl, and MAFbx [12]. ANGII also regulates the activity
of activating transcription factor 2 (ATF2), a transcription
factor for renin [13]. Murphy et al. [14] found that ANGII
can induce the expression of MuRFl and MAFbx; an in-
hibitor of the RAS hormone system, specifically an in-
hibitor of angiotensin converting enzyme (ACE), which
mediates conversion of AngI to AngII, enhanced skeletal
muscle function in mice with mild or severe cachexia.
Their results support the contribution of the RAS in
cachexia.
Given the significant contribution of UPS and caspase-3
in skeletal muscle wasting, we hypothesized that inhibiting
UPS and caspase-3 activities may confer therapeutic ben-
efits to patients with cancer-associated cachexia. CFZ is an
epoxyketone-based irreversible 20S proteasome inhibitor
[14] that selectively inhibits the chymotrypsin-like activity
of the proteasome and induces cell death in multiple types
of cancers [15, 16]. Currently, CFZ is approved as a sec-
ond-line treatment for patients with multiple myeloma
[17]. z-VAD-fmk is a widely used broad-spectrum caspase
inhibitor that can protect muscle from compression-in-
duced damage and restores muscle function [18] and im-
pairs angiogenesis, which may inhibit tumor growth [19].
In the current study, we tested various treatments with
Carfilzomib (CFZ) and z-VAD-fmk at different stage of
cachexia in a mouse model of cancer-associated cachexia.
To further understand the molecular mechanism underlying
the therapeutic benefit of these treatments, we also char-
acterized the expression of various apoptotic, proteolytic,
and RAS-associated markers.
Methods and materials
Reagents and cell lines
Anti-ATF2 (ab47476) and anti-pATF2 (ab13106) were
purchased from Abcam (USA). Anti-B cell lymphoma
(BCL)-xL (sc-7195), anti-BCL2-associated X protein
(BAX; sc-493), anti-caspase 3 (sc-7148), and anti-b-actin
(sc-130656) were obtained from Santa Cruz Biotechnology
(CA, USA). Secondary goat anti-rabbit IgG conjugated
with horseradish peroxidase (BA1054) was purchased from
BOSTER (Wuhan, China). The western blot analysis kit
was from Beyotime (China). Real-time PCR reagents and
primers came from TaKaRa (Dalian, China). CFZ (S2853)
and z-VAD-fmk (S7023) were purchased from Selleck
(USA) and resolved with dimethyl sulfoxide. Commer-
cially available murine colon 26 adenocarcinoma cell line
C26 cells were a gift from the Department of Pathology,
Chongqing University of Medical Sciences.
Animal studies
All animal experimental procedures were reviewed and
approved by the Institutional Animal Care and Use Com-
mittee of Chongqing University of Medical Sciences.
Male BALB/C mice (6–8 weeks old; 20–24 g) were
obtained from the Experimental Animal Center of
Chongqing University of Medical Sciences, China. The
animals were housed in a controlled environment with
specific temperature (22 ± 1 �C) and humidity
(55 ± 5 %) settings and a 12-h light–dark cycle; all ani-
mals were fed ad libitum. To acclimate to local conditions,
the animals were housed in the facility for at least 7 days
before the experiments began.
To induce cancer cachexia, C26 cells growing in ex-
ponential phase were harvested with trypsin and injected
subcutaneously into an axilla of the mouse. A total of 175
animals received C26 cell injections with 1 9 106 cells per
site; 10 animals received PBS injection instead of C26 cells
to serve as healthy controls.
The tumor-bearing animals were divided into 7 groups
of 25 animals each, according to the treatment and the time
when the treatment started: CFZ (2 mg/kg, twice a week)
and z-VAD-fmk (1.5 mg/kg, daily), alone (designated as
‘‘C’’ or ‘‘Z,’’ respectively) or in combination (designated as
‘‘U’’). Each of these treatments was administered either
5 days after cell inoculation (preventive), when the tumor
nodules were palpable, or 12 days after cell inoculation
(post-cachexia), when the mice presented signs of
cachexia. In addition, a group of tumor-bearing mice re-
ceived sterile phosphate-buffered saline (PBS) to serve as
the cachexia control (CC); another group of mice received
subcutaneous injection of PBS, instead of C26, were the
healthy controls (HC).
On day 19, 10 animals from each group were killed for
bioanalytical studies; the remaining mice (15 mice from
each group) were used for longitudinal studies of the
spontaneous physical activity and survival.
We recorded spontaneous physical activity, condition of
the fur, and body weight of the animals on a daily basis.
The spontaneous physical activity was monitored by an
infrared monitoring system (WV-CF314LCH, Panasonic,
Japan). In addition, tumor growth was measured by a
caliper, and the tumor volume was determined using the
following formula: tumor volume V (cm3) = (a 9 b2)/2,
where a represents the length in millimeter and b the width
in millimeter.
100 Page 2 of 10 Med Oncol (2015) 32:100
123

On day 19, retro-orbital blood samples were collected
from 10 mice in each group. Serum was obtained by
clotting of the blood at room temperature for 1 h followed
by centrifugation at 4000 rpm/min for 10 min. The serum
samples were stored at -20 �C before analyses. After
blood collection, the animals were euthanized by cervical
dislocation; tumors and gastrocnemius muscles from the
left leg were dissected and weighed. Muscle specimens
were divided into two portions, with one snap-frozen at
-80 �C for biochemical analyses and the other fixed in
10 % buffered formalin for histochemical studies.
Histology examination
Five-micron serial sections of paraffin-preserved gastroc-
nemius muscles were stained with hematoxylin and eosin.
The images were acquired under an inverted light micro-
scope (2009) and analyzed with Image J (http://rsb.info.
nih.gov/ij/). Approximately 200 muscle fibers from each
muscle sample were analyzed for cross-sectional area
quantification.
Bioanalytical assays
Serum glucose, triglyceride, total protein, and albumin
were measured using a Beckman counter (USA). Rennin
(sc-22752) and AngII (sc-9040) (santacruze, USA) levels
in serum were determined using sandwich enzyme-linked
immunosorbent assay.
Real-time reverse transcription PCR (qRT-PCR)
analysis
Total RNA was isolated from frozen gastrocnemius muscle
samples with TRIzol reagent (TaKaRa, China) and reverse
transcribed to cDNA in accordance with the manufactur-
er’s instructions. RT-PCR was performed using the SYBR
Green quantitative reverse transcription PCR System
(KAPA Biosystems, USA). Each qRT-PCR reaction
(10 lL) included 1 lL of diluted cDNA, 0.6 lL of primers,
5 lL of KAPA SYBR fast qPCR master mix, and double-
distilled water.
Table 1 contains a list of all the primers used in the real-
time PCR analysis. Real-time-PCR was performed using an
ABI Prism 7500 sequence detection system (Applied
Biosystems), under the following reaction conditions:
95 �C for 1 min, 40 cycles of a three-step reaction, de-
naturation at 95 �C for 5 s, annealing at 58 �C for 15 s, and
extension at 72 �C for 15 s.
Melting curve analyses were performed with the Bio-
Rad CFX Manager v1.6 (Bio-Rad) to ensure amplification
specificity. The amount of the target transcripts were nor-
malized to glyceraldehyde 3-phosphate dehydrogenase
(GAPDH), and the relative fold changes in expression were
calculated using the comparative cycle threshold (Ct)
method.
Western blot analysis
Nuclear and total proteins were extracted from gastrocne-
mius muscles with RIPA buffer (50 mM Tris pH 7.4,
150 mM NaCl, 1 % Triton X-100, 1 % sodium deoxy-
cholate, 0.1 % sodium dodecyl sulfate), and multiple pro-
tease inhibitors (including sodium orthovanadate, sodium
fluoride, ethylenediaminetetraacetic acid, leupeptin), and
quantified with a bicinchoninic acid assay. Protein lysates
were subsequently resolved with 10 % sodium dodecyl
sulfate–PAGE and transferred onto a polyvinylidene di-
fluoride membrane (Beyotime Institute of Biotechnology,
China). The membranes was blocked with 5 % skim milk
at 37 �C for 1 h and then incubated with specific primary
antibodies overnight at 4 �C. Thereafter, the membranes
were washed with Tris-buffered saline with Tween 20 and
incubated with secondary goat anti-rabbit IgG-horseradish
peroxidase for 1 h at room temperature. Proteins of interest
were visualized using a UVP BioImaging system (Super
bright scientific instrument co., LTD, New York, America)
and analyzed with LabWorks software (Gene co., LTD.,
Shanghai, China).
Statistical analyses
SPSS 17.0 was used for statistical analyses, and all data are
presented as mean ± standard deviation. One-way
ANOVA was used to compare more than two groups;
Newman–Keuls analyses were employed for pairwise
Table 1 Primers for real-time PCR
Gene Primer sequence
ATF2 50-GCCCTTCCTCTCCTCAACCA-30
50-AGTCCTAACCAATCCACTGCCA-30
Caspase3 50-GGACTGATGAGGAGATGGCTTG-30
50-AGGGACTGGATGAACCACGAC-30
BAX 50-GGATGCGTCCACCAAGAAGC-30
50-AAAGTAGAAGAGGGCAACCACG-30
BCL-XL 50-GGTAGTGAATGAACTCTTTCGGGA-30
50-CATCTCCTTGTCTACGCTTTCCAC-30
MAFbx 50-GAAGAGAGCAGTATGGGGTCAC-30
50-CTTGAGGGGAAAGRGAGACG-30
MuRFl 50-GGAACACGAAGACGAGAAAATC-30
50-TGGCTATTCTCCTTGGTCACTC-30
GAPDH 50-GGTGAAGGTCGGTGTGAACG-30
50-CTCGCTCCTGGAAGATGGTG-30
Med Oncol (2015) 32:100 Page 3 of 10 100
123
comparisons. Survival data were analyzed using a log-rank
test. P \ 0.05 was considered statistically significant.
Results
Combined treatments of CFZ and z-VAD-fmk
ameliorate weight loss and reduce tumor burden
in the C26 tumor-bearing mice
Mice bearing subcutaneous C26 tumors are commonly
used as a rodent model of cancer-associated cachexia [20],
since these mice undergo lethal wasting as the tumors
grow. To evaluate the effect of CFZ and z-VAD-fmk on
cancer-associated cachexia, we administered the two drugs
to the C26-inoculated mice at 5 and 12 days after
inoculation and evaluated changes in body weight, gas-
trocnemius muscle mass, and tumor growth at 5-, 12-, and
19-day post-inoculation.
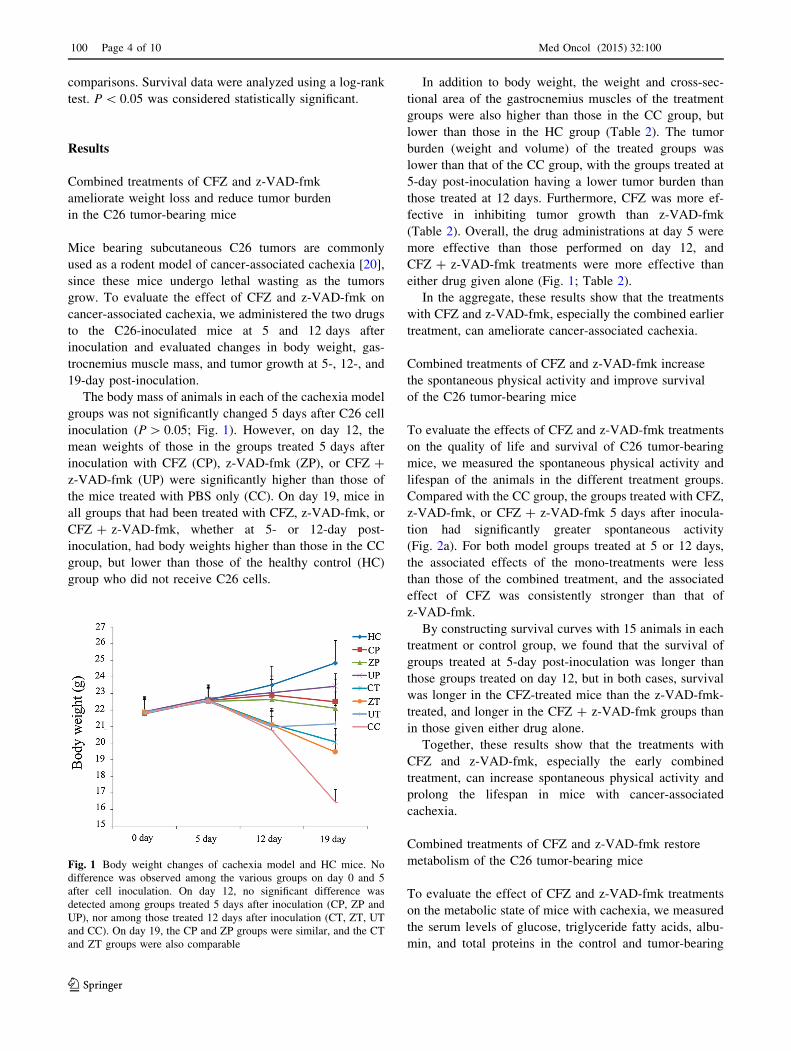
The body mass of animals in each of the cachexia model
groups was not significantly changed 5 days after C26 cell
inoculation (P [ 0.05; Fig. 1). However, on day 12, the
mean weights of those in the groups treated 5 days after
inoculation with CFZ (CP), z-VAD-fmk (ZP), or CFZ ?
z-VAD-fmk (UP) were significantly higher than those of
the mice treated with PBS only (CC). On day 19, mice in
all groups that had been treated with CFZ, z-VAD-fmk, or
CFZ ? z-VAD-fmk, whether at 5- or 12-day post-
inoculation, had body weights higher than those in the CC
group, but lower than those of the healthy control (HC)
group who did not receive C26 cells.
In addition to body weight, the weight and cross-sec-
tional area of the gastrocnemius muscles of the treatment
groups were also higher than those in the CC group, but
lower than those in the HC group (Table 2). The tumor
burden (weight and volume) of the treated groups was
lower than that of the CC group, with the groups treated at
5-day post-inoculation having a lower tumor burden than
those treated at 12 days. Furthermore, CFZ was more ef-
fective in inhibiting tumor growth than z-VAD-fmk
(Table 2). Overall, the drug administrations at day 5 were
more effective than those performed on day 12, and
CFZ ? z-VAD-fmk treatments were more effective than
either drug given alone (Fig. 1; Table 2).
In the aggregate, these results show that the treatments
with CFZ and z-VAD-fmk, especially the combined earlier
treatment, can ameliorate cancer-associated cachexia.
Combined treatments of CFZ and z-VAD-fmk increase
the spontaneous physical activity and improve survival
of the C26 tumor-bearing mice
To evaluate the effects of CFZ and z-VAD-fmk treatments
on the quality of life and survival of C26 tumor-bearing
mice, we measured the spontaneous physical activity and
lifespan of the animals in the different treatment groups.
Compared with the CC group, the groups treated with CFZ,
z-VAD-fmk, or CFZ ? z-VAD-fmk 5 days after inocula-
tion had significantly greater spontaneous activity
(Fig. 2a). For both model groups treated at 5 or 12 days,
the associated effects of the mono-treatments were less
than those of the combined treatment, and the associated
effect of CFZ was consistently stronger than that of
z-VAD-fmk.
By constructing survival curves with 15 animals in each
treatment or control group, we found that the survival of
groups treated at 5-day post-inoculation was longer than
those groups treated on day 12, but in both cases, survival
was longer in the CFZ-treated mice than the z-VAD-fmk-
treated, and longer in the CFZ ? z-VAD-fmk groups than
in those given either drug alone.
Together, these results show that the treatments with
CFZ and z-VAD-fmk, especially the early combined
treatment, can increase spontaneous physical activity and
prolong the lifespan in mice with cancer-associated
cachexia.
Combined treatments of CFZ and z-VAD-fmk restore
metabolism of the C26 tumor-bearing mice
To evaluate the effect of CFZ and z-VAD-fmk treatments
on the metabolic state of mice with cachexia, we measured
the serum levels of glucose, triglyceride fatty acids, albu-
min, and total proteins in the control and tumor-bearing
Fig. 1 Body weight changes of cachexia model and HC mice. No
difference was observed among the various groups on day 0 and 5
after cell inoculation. On day 12, no significant difference was
detected among groups treated 5 days after inoculation (CP, ZP and
UP), nor among those treated 12 days after inoculation (CT, ZT, UT
and CC). On day 19, the CP and ZP groups were similar, and the CT
and ZT groups were also comparable
100 Page 4 of 10 Med Oncol (2015) 32:100
123

groups 19 days after C26 cell inoculation. Compared with
the HC group, all the tumor-bearing mice had lower levels
of glucose, albumin, and total proteins, and higher levels of
triglyceride fatty acids (Table 3). The CFZ and z-VAD-
fmk treatments administered either 5- or 12-day post-
inoculation were associated with higher levels of glucose,
albumin, and total proteins and lower levels of triglyceride
fatty acid, compared with the group given PBS. Consistent
with our above observation in the physiological examina-
tion, survival analysis, and tumor measurement, the pre-
ventive setting was more effective than the post-cachexia
setting; the effect of CFZ treatment was consistently
stronger than that of z-VAD-fmk; the combined treatment
was more effective than the mono-treatments in both set-
tings (Fig. 2a). Together, these results demonstrated that
the treatments with CFA and z-VAD-fmk, especially the
combined treatment in the prevention setting, can reverse
the catabolic state in animals with cachexia.
Treatments of CFZ and z-VAD-fmk alter RAS in C26
tumor-bearing mice
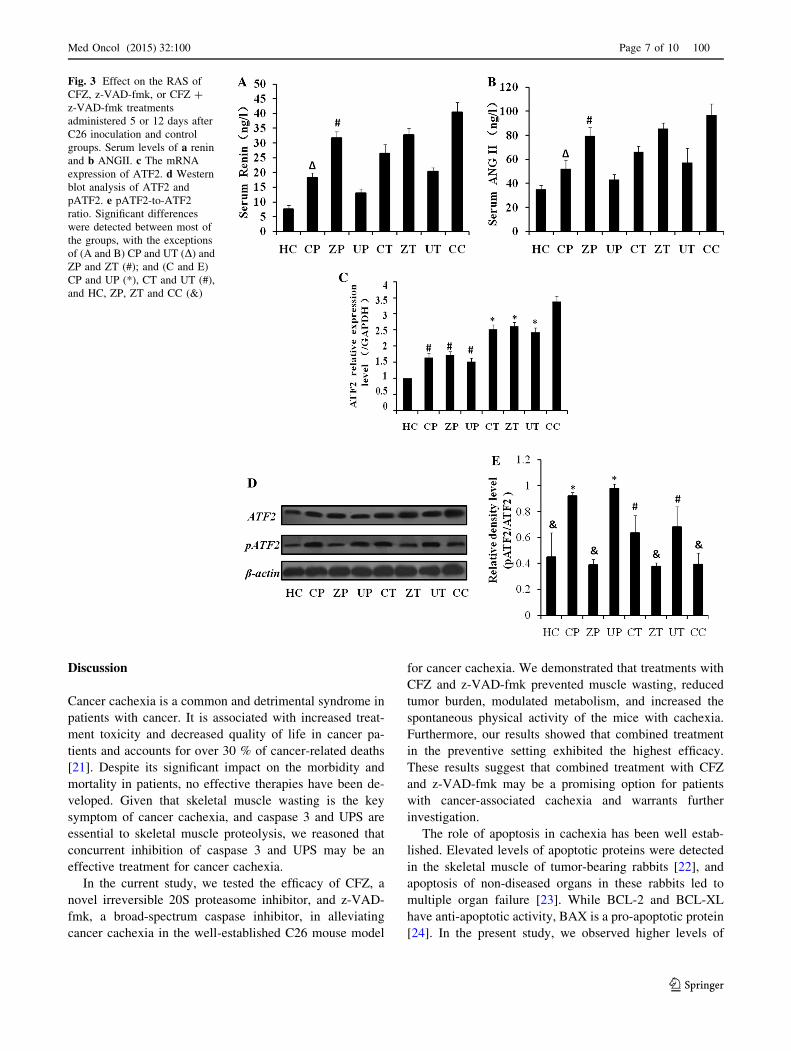
Previous studies [14] have shown that activation of RAS is
associated with cachexia, and ANGII can activate proteins
involved in proteolysis. To test whether treatments with
CFZ and z-VAD-fmk affect RAS activity in mice with
cachexia, we measured the levels of renin and ANGII in
the different experimental groups. Compared with the HC
group, the tumor-bearing animals had much higher levels
of renin and ANGII. Treatments with CFZ and z-VAD-fmk
were associated with lower levels of renin and ANGII. The
renin and ANGII levels of the CFZ-treated groups were
lower than those of the z-VAD-fmk-treated group, and
lower still in the CFZ ? z-VAD-fmk group than those
receiving either drug alone. The renin and ANGII levels of
the group treated with CFZ 5 days after inoculation were
lower than those of the CFZ group treated at 12 days, but
no difference was observed between the groups receiving
z-VAD-fmk treatments at the two times (Fig. 3a, b).
To explore the molecular mechanism underlying CFZ and
z-VAD-fmk-associated alteration in RAS, we measured the
levels of ATF2 in the gastrocnemius muscles of animals in
the different groups. Results from both qRT-PCR and
western blot analyses showed that ATF2 levels were higher
in the tumor-bearing animals than in the healthy controls
(Fig. 3c, d; Table 4). ATF2 levels in the groups treated with
CFZ, z-VAD-fmk, or CFZ ? z-VAD-fmk, at either 5 or
12 days after inoculation, were lower than in the CC group
(treated with PBS). There was no difference in ATF2 levels
between corresponding groups treated at 5 and 12 days.
Regarding the levels of pATF2, the active form of ATF2,
these were dramatically higher in the CFZ-treated groups
(CFZ-only and CFZ ? z-VAD-fmk) than in the z-VAD-fmkTa
ble
2T
he
effe
cto
fC
FZ
and
z-V
AD
-fm
ktr
eatm
ents
on
bo
dy
wei
gh
t,g
astr
ocn
emiu
sm
usc
le,
and
tum
or
bu
rden
Tre
ated
5-d
ayp
ost
-in
ocu
lati
on
Tre
ated
12
-day
po
st-i
no
cula
tio
n
HC
CC
CP
ZP
UP
CT
ZT
UT
Bo
dy
Wei
gh
t,g
24
.85
±1
.36
16
.44
±0
.78
22
.50
±1
.36
12
2.1
1±
0.9
91
23
.45
±0
.74
20
.08
±0
.81
21
9.4
8±
1.0
12
21
.16
±1
.11
Gas
tro
cnem
ius
Wt,
mg
13
7.6
±1
3.3
74
.6±
6.0
11
5.8
±1
3.2
39
8.5
±6
.04
12
6.5
±1
0.6
10
2.9
±9
.84
85
.2±
10
.31
14
.3±
14
.13
XS
,u
m2
2.4
8±
0.2
41
.22
±0
.14
2.0
0±
0.2
85
1.6
3±
0.1
76
2.2
4±
0.2
31
.69
±0
.19
61
.38
±0
.12
1.8
8±
0.2
95
Tu
mo
rW
t,g
–4
.87
±0
.39
2.2
6±
0.3
53
.43
±0
.27
71
.91
±0
.33
3.3
8±
0.4
07
4.2
5±
0.4
32
.67
±0
.27
Vo
l,cm
3–
5.4
7±
0.4
82
.62
±0
.40
3.9
2±
0.3
28
2.2
1±
0.4
43
.90
±0
.47
84
.82
±0
.44
3.1
2±
0.4
5
All
the
mea
sure
men
tsw
ere
tak
eno
nd
ay1
9p
ost
-tu
mo
rce
llin
ocu
lati
on
.G
rou
ps
wit
hth
esa
me
sup
ersc
rip
ted
nu
mb
ers
wer
en
ot
sig
nifi
can
tly
dif
fere
nt;
the
dif
fere
nce
sb
etw
een
the
oth
erg
rou
p
pai
rsw
ere
stat
isti
call
ysi
gn
ifica
nt
CC
cach
exia
con
tro
l,C
PC
FZ
pre
ven
tio
n,
CT
CF
Ztr
eatm
ent,
HC
hea
lth
yco
ntr
ol,
UP
CF
Z?
z-V
AD
-fm
kp
rev
enti
on
,U
TC
FZ
?z-
VA
D-f
mk
trea
tmen
t,X
Scr
oss
sect
ion
,Z
Pz-
VA
D-f
mk
pre
ven
tio
n,
ZT
z-V
AD
-fm
ktr
eatm
ent
Med Oncol (2015) 32:100 Page 5 of 10 100
123

groups, with correspondingly higher ratios of pATF2 to
ATF2 (Fig. 3d, e). Since high ATF2 activity can inhibit renin
transcription, this result suggested a possible mechanism by
which CFZ inhibits RAS activity.
Treatments of CFZ and z-VAD-fmk inhibit apoptosis
and the ubiquitin-proteolysis pathways
To characterize the effect of CFZ and z-VAD-fmk on
apoptosis and the UPS, we measured the expression of
several well-characterized genes involved in the pathways,
including caspase 3, BAX, and BCL-XL for apoptosis, and
MuRF1 and MAFbx for the ubiquitin degradation pathway.
Meanwhile, we measured the levels of caspase 3, BAX,
and BCL-XL proteins. Results from both qRT-PCR and
western blot analyses revealed that treatments with CFZ
and z-VAD-fmk were associated with an inhibition of the
expression of caspase 3 and BAX and induced the ex-
pression of BCL-XL, resulting in a decrease in the BAX-
to-BCL-XL ratio (Fig. 4a–e). The CFZ ? z-VAD-fmk
treatments (at 5 and 12 days) had a greater effect than the
corresponding mono-treatments; treatments administered
5 days after inoculation were more effective than those
administered at 12 days; no differences were observed
between the CFZ and z-VAD-fmk groups. Similarly, both
CFZ and z-VAD-fmk treatments inhibited the expression
of MuRF1 and MAFbxm, two genes involved in ubiquitin-
mediated protein degradation pathway. The inhibitory ef-
fect of CFZ was stronger than that of z-VAD-fmk (Fig. 4e,
f). These results suggested that CFZ and z-VAD-fmk may
ameliorate cachexia via inhibiting apoptosis and ubiquitin-
medicated protein degradation pathway.
Fig. 2 The spontaneous physical activity and overall survival of the
C26 tumor-bearing mice treated with CFZ, z-VAD-fmk, or CFZ ? z-
VAD-fmk at 5- or 12-day post-inoculation and control mice. a The
spontaneous physical activities of animals in different treatment
groups. The P values between most of the group pairs were \0.001,
except for P \ 0.05 between CP and UT (*) and between ZP and CT
(D). b Kaplan–Meier survival curves of animals in different treatment
groups. The P values between most of the group pairs were \0.001,
except for P \ 0.05 between CP and UT (#) and between ZP and CT
(&)
Table 3 Metabolic parameters of the experimental groups at 19 days
Treated 5-day post-inoculation Treated 12-day post-inoculation
HC CC CP ZP UP CT ZT UT
Serum
glucose,
mmol/L
5.99 ± 0.26 3.06 ± 0.18 4.71 ± 0.171 3.95 ± 0.182 4.94 ± 0.55 4.17 ± 0.172 3.48 ± 0.14 4.53 ± 0.171
Serum
TFA,
mmol/L
1.16 ± 0.15 4.99 ± 0.32 2.93 ± 0.181 3.60 ± 0.222 2.26 ± 0.20 3.75 ± 0.282 4.29 ± 0.31 3.13 ± 0.171
Albumin,
g/L
19.84 ± 0.47 13.23 ± 0.28 16.45 ± 0.291 14.96 ± 0.322 18.43 ± 0.35 15.26 ± 0.412 14.15 ± 0.19 16.29 ± 0.431
Total
protein,
g/L
59.17 ± 0.90 49.21 ± 1.21 56.61 ± 1.081 54.07 ± 1.222 57.83 ± 1.18 55.13 ± 0.942,3 52.28 ± 1.29 55.91 ± 1.381,3
The difference between most of the groups was statistically significant (P \ 0.05), with the exception of the groups with the same superscripted
numbers. TFA triglyceride fatty acid
100 Page 6 of 10 Med Oncol (2015) 32:100
123
Discussion
Cancer cachexia is a common and detrimental syndrome in
patients with cancer. It is associated with increased treat-
ment toxicity and decreased quality of life in cancer pa-
tients and accounts for over 30 % of cancer-related deaths
[21]. Despite its significant impact on the morbidity and
mortality in patients, no effective therapies have been de-
veloped. Given that skeletal muscle wasting is the key
symptom of cancer cachexia, and caspase 3 and UPS are
essential to skeletal muscle proteolysis, we reasoned that
concurrent inhibition of caspase 3 and UPS may be an
effective treatment for cancer cachexia.
In the current study, we tested the efficacy of CFZ, a
novel irreversible 20S proteasome inhibitor, and z-VAD-
fmk, a broad-spectrum caspase inhibitor, in alleviating
cancer cachexia in the well-established C26 mouse model
for cancer cachexia. We demonstrated that treatments with
CFZ and z-VAD-fmk prevented muscle wasting, reduced
tumor burden, modulated metabolism, and increased the
spontaneous physical activity of the mice with cachexia.
Furthermore, our results showed that combined treatment
in the preventive setting exhibited the highest efficacy.
These results suggest that combined treatment with CFZ
and z-VAD-fmk may be a promising option for patients
with cancer-associated cachexia and warrants further
investigation.
The role of apoptosis in cachexia has been well estab-
lished. Elevated levels of apoptotic proteins were detected
in the skeletal muscle of tumor-bearing rabbits [22], and
apoptosis of non-diseased organs in these rabbits led to
multiple organ failure [23]. While BCL-2 and BCL-XL
have anti-apoptotic activity, BAX is a pro-apoptotic protein
[24]. In the present study, we observed higher levels of
Fig. 3 Effect on the RAS of
CFZ, z-VAD-fmk, or CFZ ?
z-VAD-fmk treatments
administered 5 or 12 days after
C26 inoculation and control
groups. Serum levels of a renin
and b ANGII. c The mRNA
expression of ATF2. d Western
blot analysis of ATF2 and
pATF2. e pATF2-to-ATF2
ratio. Significant differences
were detected between most of
the groups, with the exceptions
of (A and B) CP and UT (D) and
ZP and ZT (#); and (C and E)
CP and UP (*), CT and UT (#),
and HC, ZP, ZT and CC (&)
Med Oncol (2015) 32:100 Page 7 of 10 100
123
BAX and caspase 3 and lower levels of BCL-XL in mice
with cachexia, and we also found that CFZ and z-VAD-
fmk treatments were associated with lower levels of cas-
pase 3 and BAX and higher levels of BCL-XL, leading to a
lower BAX-to-BCL-XL ratio than in PBS-treated cachexia
mice. These observations suggest that these treatments
inhibited skeletal muscle apoptosis. Since caspase 3 also
serves as the initial activator in proteolysis, the lower
levels of caspase 3 also provide an explanation for the
lesser amount of muscle proteolysis observed in cachexia
mice treated with CFZ and z-VAD-fmk.
RAS has an important role in cachexia, and ANGII
levels were found to be higher in patients with cachexia
[9, 10, 25]. ANGII can promote the E3 ligases MuRFl and
MAFbx, inducing proteolysis of skeletal muscle [26].
ANGII can also activate caspase 3 [25]. Increased levels
of MuRF-1 and MAFbx via ANGII have been suggested
as the underlying mechanism for ANG II-induced skeletal
muscle atrophy in rodents [12]. In the present study, we
found that mice treated with CFZ and z-VAD-fmk had
lower levels of ANGII, MuRF1, and MAFbx. Therefore, it
is possible that the effect of CFZ and z-VAD-fmk is
mediated in part through the downregulation of ANGII
levels, which subsequently lowers levels of MuRF-1 and
MAFbx.
In the RAS, renin is the rate-limiting step for the con-
version of angiotensinogen to ANGI. Therefore, inhibiting
renin expression can restrict levels of ANGI and subse-
quently that of ANGII. In the current study, we found that
the combined treatment of CFZ and z-VAD-fmk led to
lower renin levels. This provides an explanation for the
lower ANGII levels found in these mice. The cAMP re-
sponse element (enhCRE) in the distal enhancer regulatory
region of the renin gene has been reported to be important
in controlling renin transcription. enhCRE is also a binding
target of ATF2: Once activated by phosphorylation, ATF2
binds to enhCRE to replace the CRE-binding protein
(CREB), causing suppression in renin expression [13, 27].
In the present study, we observed higher pATF2 levels in
the CFZ-treated animals but not the other groups. The high
pATF2 in CFZ-treated animals could be the result of the
inhibition of UPS by CFZ. The higher pATF2 levels lead
to a higher pATF2-to-ATF2 ratio, providing a plausible
explanation for the decrease in renin expression in CFZ-
treated mice with cachexia.
It is worth noting that z-VAD-fmk treatment was not
associated with higher pATF2 levels. z-VAD-fmk is an
apoptosis inhibitor and is not expected to increase pATF2
levels by downregulating proteasome activity. The fact that
z-VAD-fmk treatment caused no pATF2 upregulation
suggests that downregulation of RAS by pATF2 is specific
to CFZ treatment, and other mechanisms are involved in
z-VAD-fmk-mediated cachexia rescue.Ta
ble
4P
rote
inle
vel
so
fA
TF
2,
pA
TF
2,
casp
ase
3,
BA
X,
and
BC
L-X
Lin
the
gas
tro
cnem
ius
mu
scle
so
fm
ice
Tre
ated
5-d
ayp
ost
-in
ocu
lati
on
Tre
ated
12
-day
po
st-i
no
cula
tio
n
HC
CC
CP
ZP
UP
CT
ZT
UT
AT
F2
0.4
3±
0.0
90
.97
±0
.11
0.6
3±
0.0
81
0.6
9±
0.0
81
0.6
5±
0.0
51
0.8
5±
0.0
52
0.8
4±
0.0
52
0.8
2±
0.0
52
pA
TF
20
.19
±0
.06
30
.39
±0
.06
60
.58
±0
.08
40
.27
±0
.06
3,5
0.6
3±
0.0
54
0.5
4±
0.0
84
0.3
2±
0.0
35,6
0.5
4±
0.0
74
Cas
pas
e3
0.3
7±
0.0
40
.97
±0
.11
0.6
6±
0.0
77
0.6
3±
0.0
57
0.5
1±
0.0
90
.83
±0
.04
80
.78
±0
.05
80
.65
±0
.09
7
BA
X0
.23
±0
.03
0.7
0±
0.0
40
.43
±0
.02
90
.41
±0
.02
90
.32
±0
.06
0.6
0±
0.0
910
0.5
7±
0.0
610
0.4
6±
0.0
69
BC
L-X
L0
.60
±0
.07
11
0.3
8±
0.0
50
.73
±0
.03
12
0.6
7±
0.0
411,1
20
.84
±0
.04
0.5
5±
0.0
611,1
30
.50
±0
.03
13
0.6
7±
0.0
711,1
2
No
rmal
ized
tob
-act
in.
Th
ed
iffe
ren
ces
bet
wee
nm
ost
of
the
gro
up
sw
ere
sig
nifi
can
t(P
\0
.05
),w
ith
the
exce
pti
on
of
the
gro
up
sw
ith
the
sam
esu
per
scri
pte
dn
um
ber
s
100 Page 8 of 10 Med Oncol (2015) 32:100
123

Fig. 4 Treatments with CFZ,
z-VAD-fmk, and CFZ ?
z-VAD-fmk altered the
expression of genes involved in
apoptosis and the UPS in
gastrocnemius muscles. (A to C)
The mRNA expression levels of
a caspase 3, b BCL-XL, and
c Bax. d The ratio of Bax to
BCL-XL. e Caspase 3, BCL-
XL, and BAX proteins. The
mRNA expressions of
(f) MuRF1 and (g) MAFbx.
Significant differences were
detected between most of the
groups with the exceptions of
those whose labels are the same
in the graph. For example, there
is no statistically significant
difference between HC, ZP, and
UT (labeled with symbol ‘‘$’’)
and no difference between CT
and ZT (labeled with ‘‘¥’’) in
panel A
Med Oncol (2015) 32:100 Page 9 of 10 100
123

In the aggregate, we found that treatments with CFZ and
z-VAD-fmk were effective in preventing weight loss,
muscle atrophy, and tumor growth and increased the
quality of life and survival in mice with cancer-associated
cachexia. In particular, we found that the early combined
treatment of these drugs was more effective than the other
treatment options. Our molecular exploration suggests that
CFZ and z-VAD-fmk treatments can inhibit apoptosis and
the proteolysis pathways in skeletal muscles. Furthermore,
the RAS system may be involved in downregulating pro-
teolytic proteins in the UPS, and regulation of ATF2 ac-
tivity may be involved in regulating RAS in cachexia
model mice treated with CFZ. These results warrant further
evaluation of the viability of administering combined CFZ
and z-VAD-fmk early in the treatment of cancer patients
with cachexia.
Acknowledgments This work was supported by the Scientific Re-
search Project of Chongqing Health Bureau, China (Grant number:
2011-2-101). Hua Tang and Qiang Wang designed and executed the
study. Chunhong Li cultured the cells used in the study. Qiang Wang
is responsible for all the statistical analysis and manuscript writing.
All the authors participated in the development of the animal model
and the revision of the manuscript.
Conflict of interest The authors declare no conflict of interest.
References
1. Fearon K, Strasser F, Anker SD, Bosaeus I, Bruera E, Fainsinger
RL, Jatoi A, Loprinzi C, MacDonald N, Mantovani G, Davis M,
Muscaritoli M, Ottery F, Radbruch L, Ravasco P, Walsh D,
Wilcock A, Kaasa S, Baracos VE. Definition and classification of
cancer cachexia: an international consensus. Lancet Oncol.
2011;12:489–95.
2. von Haehling S, Anker SD. Cachexia as major underestimated
unmet medical need: facts and numbers. Int J Cardiol.
2012;161:121–3.
3. Kumar NB, Kazi A, Smith T, Crocker T, Yu D, Reich RR, Reddy
K, Hastings S, Exterman M, Balducci L, Dalton K, Bepler G.
Cancer cachexia: traditional therapies and novel molecular
mechanism-based approaches to treatment. Curr Treat Options
Oncol. 2010;11:107–17.
4. Fearon K, Arends J, Baracos V. Understanding the mechanisms
and treatment options in cancer cachexia. Nat Rev Clin Oncol.
2013;10:90–9.
5. Ross PJ, Ashley S, Norton A, Priest K, Waters JS, Eisen T, Smith
IE, O’Brien ME. Do patients with weight loss have a worse
outcome when undergoing chemotherapy for lung cancers? Br J
Cancer. 2004;90:1905–11.
6. Wing SS, Lecker SH, Jagoe RT. Proteolysis in illness-associated
skeletal muscle atrophy: from pathways to networks. Crit Rev
Clin Lab Sci. 2011;48:49–70.
7. Johns N, Stephens NA, Fearon KCH. Muscle wasting in cancer.
Int J Biochem Cell Biol. 2013;45:2215–29.
8. Argiles JM, Lopez-Soriano FJ, Busquets S. Apoptosis signalling
is essential and precedes protein degradation in wasting skeletal
muscle during catabolic conditions. Int J Biochem Cell Biol.
2008;40:1674–8.
9. Du J, Wang X, Miereles C, Bailey JL, Debigare R, Zheng B,
Price SR, Mitch WE. Activation of caspase-3 is an initial step
triggering accelerated muscle proteolysis in catabolic conditions.
J Clin Invest. 2004;113:115–23.
10. Rajan VR, Mitch WE. Muscle wasting in chronic kidney disease:
the role of the ubiquitin proteasome system and its clinical im-
pact. Pediatr Nephrol. 2008;23:527–35.
11. Wang XH, Zhang L, Mitch WE, LeDoux JM, Hu J, Du J. Cas-
pase-3 cleaves specific 19 S proteasome subunits in skeletal
muscle stimulating proteasome activity. J Biol Chem. 2010;
285:21249–57.
12. Sukhanov S, Semprun-Prieto L, Yoshida T, Michael Tabony A,
Higashi Y, Galvez S, Delafontaine P. Angiotensin II, oxidative
stress and skeletal muscle wasting. Am J Med Sci. 2011;342:143–7.
13. Desch M, Hackmayer G, Todorov VT. Identification of ATF2 as
a transcriptional regulator of renin gene. Biol Chem. 2012;393:
93–100.
14. Kuhn DJ, Orlowski RZ, Bjorklund CC. Second generation pro-
teasome inhibitors: carfilzomib and immunoproteasome-specific
inhibitors (IPSIs). Curr Cancer Drug Targets. 2011;11:285–95.
15. Mansour MA, Aljoufi MA, Al-Hosaini K, Al-Rikabi AC, Nagi
MN. Possible role of selective, irreversible, proteasome inhibitor
(carfilzomib) in the treatment of rat hepatocellular carcinoma.
Chem Biol Interact. 2014;215:17–24.
16. Parlati F, Lee SJ, Aujay M, Suzuki E, Levitsky K, Lorens JB,
Micklem DR, Ruurs P, Sylvain C, Lu Y, Shenk KD, Bennett MK.
Carfilzomib can induce tumor cell death through selective inhi-
bition of the chymotrypsin-like activity of the proteasome. Blood.
2009;114:3439–47.
17. Jakubowiak AJ. Evolution of carfilzomib dose and schedule in
patients with multiple myeloma: A historical overview. Cancer
Treat Rev. 2014;40:781–90.
18. Teng BT, Tam EW, Benzie IF, Siu PM. Protective effect of
caspase inhibition on compression-induced muscle damage.
J Physiol. 2011;589:3349–69.
19. Segura I, Serrano A, De Buitrago GG, Gonzalez MA, Abad JL,
Claveria C, Gomez L, Bernad A, Martinez AC, Riese HH. In-
hibition of programmed cell death impairs in vitro vascular-like
structure formation and reduces in vivo angiogenesis. FASEB J.
2002;16:833–41.
20. Tanaka Y, Eda H, Tanaka T, Udagawa T, Ishikawa T, Horii I,
Ishitsuka H, Kataoka T, Taguchi T. Experimental cancer cachexia
induced by transplantable colon 26 adenocarcinoma in mice.
Cancer Res. 1990;50:2290–5.
21. Fearon KC. Cancer cachexia: developing multimodal therapy for
a multidimensional problem. Eur J Cancer. 2008;44:1124–32.
22. Ishiko O, Sumi T, Yoshida H, Hyun Y, Ogita S. Angiogenesis in
the adipose tissue of tumor-bearing rabbits treated by cyclic
plasma perfusion. Int J Oncol. 2001;19:785–90.
23. Fukuda T, Sumi T, Nobeyama H, Yoshida H, Matsumoto Y,
Yasui T, Honda K, Ishiko O. Multiple organ failure of tumor-
bearing rabbits in cancer cachexia is caused by apoptosis of
normal organ cells. Int J Oncol. 2009;34:61–7.
24. Czabotar PE, Lessene G, Strasser A, Adams JM. Control of
apoptosis by the BCL-2 protein family: implications for phy-
siology and therapy. Nat Rev Mol Cell Biol. 2014;15:49–63.
25. Rajan V, Mitch WE. Ubiquitin, proteasomes and proteolytic
mechanisms activated by kidney disease. Biochim Biophys Acta.
2008;1782:795–9.
26. Zhang L, Du J, Hu Z, Han G, Delafontaine P, Garcia G, Mitch
WE. IL-6 and serum amyloid A synergy mediates angiotensin II-
induced muscle wasting. J Am Soc Nephrol. 2009;20:604–12.
27. Bhoumik A, Ronai ZE. ATF2: a transcription factor that elicits
oncogenic or tumor suppressor activities. Cell Cycle. 2008;7:
2341–5.
100 Page 10 of 10 Med Oncol (2015) 32:100
123
Related Documents











