Immunity Article Combined Deficiency of Proapoptotic Regulators Bim and Fas Results in the Early Onset of Systemic Autoimmunity Jack Hutcheson, 1,2 John C. Scatizzi, 1,3 Akbar M. Siddiqui, 1 G. Kenneth Haines III, 4 Tianfu Wu, 2 Quan-Zhen Li, 2 Laurie S. Davis, 2 Chandra Mohan, 2 and Harris Perlman 1, * 1 Department of Molecular Microbiology & Immunology, School of Medicine, Saint Louis University, Saint Louis, MO 63104, USA 2 Department of Internal Medicine-Rheumatology, Center for Immunology, University of Texas - Southwestern Medical Center, Dallas, TX 75390, USA 3 Department of Rheumatology, Allergy, and Immunology, School of Medicine, University of California, San Diego, San Diego, CA 92093, USA 4 Department of Pathology, School of Medicine, Yale University, New Haven, CT 06510, USA *Correspondence: [email protected] DOI 10.1016/j.immuni.2007.12.015 SUMMARY Alterations in the stoichiometric balance between members of Bcl-2 and Fas apoptotic pathway could lead to the pathogenesis of systemic lupus erythema- tosus (SLE). We showed that patients with SLE dis- played increased expression in antiapoptotic mem- bers of the Bcl-2 and Fas apoptotic pathways in isolated mononuclear cells. Further, mice (Bcl2l11 / Fas lpr/lpr ) lacking the Bcl-2 pro-apoptotic member, Bim (Bcl2l11 / ) and and with an lpr mutation in the gene encoding Fas (Fas lpr/lpr ) developed severe SLE-like disease by 16 weeks of age unlike Bcl2l11 / or Fas lpr/lpr mice. Bcl2l11 / Fas lpr/lpr antigen-presenting cells (APCs) were markedly acti- vated, and their numbers were increased in lymphoid tissues and in kidneys, yet numerous TUNEL-positive cells were observed in glomeruli of Bcl2l11 / Fas lpr/lpr mice. These data demonstrate that dysregu- lation of the Bcl-2 or Fas pathways can alter the func- tion of APCs, thereby leading to SLE pathogenesis. INTRODUCTION Systemic lupus erythematosus (SLE) is a multifactorial, multige- netic autoimmune disease that is primarily defined by increased incidence of autoantibodies that contribute to severe end-organ damage to the spleen, liver, joints, central nervous system, cardiovascular system, and kidneys (Munoz et al., 2005). Inflam- mation resulting from deposited immune complexes can lead to glomerulonephritis, dermatitis, serositis, and vasculitis (Liu and Mohan, 2006). Apoptotic cells might be a source of autoantigens (Cohen, 2006). Thus, a defect in apoptosis or a reduction in the clearance of apoptotic cells could promote SLE pathogenesis. In mammals, there are two pathways of apoptosis: extrinsic and intrinsic (Strasser et al., 2000). In the extrinsic apoptosis pathway, death ligands bind to their cognate death receptors on the cell surface, as in the case of Fas ligand (FasL) and Fas. The binding of FasL to Fas results in the recruitment of Fas-asso- ciated death domain (FADD) and procaspase-8 to the C terminus of the death receptor. Recruitment of multiple procaspase-8 molecules leads to autocatalysis of caspase-8 and subsequent activation of caspase-3 and caspase-7, which mark the begin- ning of the degradative phase of apoptosis. The intrinsic apopto- sis pathway is mediated by the Bcl-2 protein family. Members of this family are defined by their homology to the Bcl-2 protein on the basis of the presence of up to four Bcl-2 homology (BH) do- mains that are crucial for dimerization between family members (Strasser, 2005). The Bcl-2 family has both antiapoptotic and proapoptotic members. Antiapoptotic members such as Bcl-2, Bcl-x L , Mcl-1, A1, and Bcl-w contain three or all four of the BH domains, whereas the proapoptotic family members are subdi- vided into multi-BH domain (BH1-3) proteins such as Bax and Bak and BH3-only proteins such as Bim, Bid, Bad, Noxa, and Puma (Strasser, 2005; Strasser et al., 2000). The intrinsic apopto- tic pathway is largely dependent on a sequence of events occur- ring at or within the mitochondria. Both the extrinsic and intrinsic apoptosis pathways have been shown to play a role in the devel- opment of SLE that is strain and background dependent. Fas mutant mice (Fas lpr/lpr ) on the MRL background (MRL. Fas lpr/lpr ) develop systemic vasculitis, arthritis, early-onset splenomegaly, progressive lymphadenopathy, and glomerulone- phritis including mononuclear cell infiltration, crescent formation, and increased presence of autoantibodies in the kidney (Cohen and Eisenberg, 1991; Nagata and Suda, 1995), as well as 50% mortality by 6 months of age (Theofilopoulos and Dixon, 1985). However, on the C57BL/6 background, the development of disease is subdued, with reduced lymphoproliferation, later onset of autoantibody production, and few to no renal defects (Theofilopoulos and Dixon, 1985). Furthermore, MRL. Fas lpr/lpr and C57BL/6.Fas lpr/lpr mice display a large number of CD4 CD8 - CD3 + B220 + T lymphocytes in the periphery (Theofilo- poulos and Dixon, 1985; Vidal et al., 1998). Mice lacking Bim de- velop lymphadenopathy and splenomegaly, display increased amounts of circulating autoantibodies, and accumulate immuno- globin G (IgG)-secreting plasma cells resulting in hypergamma- globulinemia even on a C57BL/6 background (Bouillet et al., 1999; Hughes et al., 2006). Approximately half of Bcl2l11 / 206 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Immunity
Article
Combined Deficiency of Proapoptotic RegulatorsBim and Fas Results in the Early Onsetof Systemic AutoimmunityJack Hutcheson,1,2 John C. Scatizzi,1,3 Akbar M. Siddiqui,1 G. Kenneth Haines III,4 Tianfu Wu,2 Quan-Zhen Li,2
Laurie S. Davis,2 Chandra Mohan,2 and Harris Perlman1,*1Department of Molecular Microbiology & Immunology, School of Medicine, Saint Louis University, Saint Louis, MO 63104, USA2Department of Internal Medicine-Rheumatology, Center for Immunology, University of Texas - Southwestern Medical Center,Dallas, TX 75390, USA3Department of Rheumatology, Allergy, and Immunology, School of Medicine, University of California, San Diego,
San Diego, CA 92093, USA4Department of Pathology, School of Medicine, Yale University, New Haven, CT 06510, USA*Correspondence: [email protected]
DOI 10.1016/j.immuni.2007.12.015
SUMMARY
Alterations in the stoichiometric balance betweenmembers of Bcl-2 and Fas apoptotic pathway couldlead to the pathogenesis of systemic lupus erythema-tosus (SLE). We showed that patients with SLE dis-played increased expression in antiapoptotic mem-bers of the Bcl-2 and Fas apoptotic pathways inisolated mononuclear cells. Further, mice (Bcl2l11�/�
Faslpr/lpr) lacking the Bcl-2 pro-apoptotic member,Bim (Bcl2l11�/�) and and with an lpr mutation in thegene encoding Fas (Faslpr/lpr) developed severeSLE-like disease by 16 weeks of age unlikeBcl2l11�/� or Faslpr/lpr mice. Bcl2l11�/�Faslpr/lpr
antigen-presenting cells (APCs) were markedly acti-vated, and their numbers were increased in lymphoidtissues and in kidneys, yet numerous TUNEL-positivecells were observed in glomeruli of Bcl2l11�/�
Faslpr/lpr mice. These data demonstrate that dysregu-lation of the Bcl-2 or Fas pathways can alter the func-tion of APCs, thereby leading to SLE pathogenesis.
INTRODUCTION
Systemic lupus erythematosus (SLE) is a multifactorial, multige-
netic autoimmune disease that is primarily defined by increased
incidence of autoantibodies that contribute to severe end-organ
damage to the spleen, liver, joints, central nervous system,
cardiovascular system, and kidneys (Munoz et al., 2005). Inflam-
mation resulting from deposited immune complexes can lead to
glomerulonephritis, dermatitis, serositis, and vasculitis (Liu and
Mohan, 2006). Apoptotic cells might be a source of autoantigens
(Cohen, 2006). Thus, a defect in apoptosis or a reduction in the
clearance of apoptotic cells could promote SLE pathogenesis.
In mammals, there are two pathways of apoptosis: extrinsic
and intrinsic (Strasser et al., 2000). In the extrinsic apoptosis
pathway, death ligands bind to their cognate death receptors
on the cell surface, as in the case of Fas ligand (FasL) and Fas.
206 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.
The binding of FasL to Fas results in the recruitment of Fas-asso-
ciated death domain (FADD) and procaspase-8 to the C terminus
of the death receptor. Recruitment of multiple procaspase-8
molecules leads to autocatalysis of caspase-8 and subsequent
activation of caspase-3 and caspase-7, which mark the begin-
ning of the degradative phase of apoptosis. The intrinsic apopto-
sis pathway is mediated by the Bcl-2 protein family. Members of
this family are defined by their homology to the Bcl-2 protein on
the basis of the presence of up to four Bcl-2 homology (BH) do-
mains that are crucial for dimerization between family members
(Strasser, 2005). The Bcl-2 family has both antiapoptotic and
proapoptotic members. Antiapoptotic members such as Bcl-2,
Bcl-xL, Mcl-1, A1, and Bcl-w contain three or all four of the BH
domains, whereas the proapoptotic family members are subdi-
vided into multi-BH domain (BH1-3) proteins such as Bax and
Bak and BH3-only proteins such as Bim, Bid, Bad, Noxa, and
Puma (Strasser, 2005; Strasser et al., 2000). The intrinsic apopto-
tic pathway is largely dependent on a sequence of events occur-
ring at or within the mitochondria. Both the extrinsic and intrinsic
apoptosis pathways have been shown to play a role in the devel-
opment of SLE that is strain and background dependent.
Fas mutant mice (Faslpr/lpr) on the MRL background (MRL.
Faslpr/lpr) develop systemic vasculitis, arthritis, early-onset
splenomegaly, progressive lymphadenopathy, and glomerulone-
phritis including mononuclear cell infiltration, crescent formation,
and increased presence of autoantibodies in the kidney (Cohen
and Eisenberg, 1991; Nagata and Suda, 1995), as well as 50%
mortality by 6 months of age (Theofilopoulos and Dixon, 1985).
However, on the C57BL/6 background, the development of
disease is subdued, with reduced lymphoproliferation, later
onset of autoantibody production, and few to no renal defects
(Theofilopoulos and Dixon, 1985). Furthermore, MRL.
Faslpr/lpr and C57BL/6.Faslpr/lpr mice display a large number of
CD4�CD8-CD3+B220+ T lymphocytes in the periphery (Theofilo-
poulos and Dixon, 1985; Vidal et al., 1998). Mice lacking Bim de-
velop lymphadenopathy and splenomegaly, display increased
amounts of circulating autoantibodies, and accumulate immuno-
globin G (IgG)-secreting plasma cells resulting in hypergamma-
globulinemia even on a C57BL/6 background (Bouillet et al.,
1999; Hughes et al., 2006). Approximately half of Bcl2l11�/�

Immunity
Loss of Bim and Fas Leads to SLE
mice on a mixed C57BL/6:SV129 background develop glomeru-
lonephritis, characterized by increased serum concentrations of
urea, glomerular hypercellularity, increased mesangial matrix,
and deposits of immune complexes (Bouillet et al., 1999). It has
also been shown that Bim plays an important role in the mainte-
nance of anergic B cells and that the loss of Bim can lead to the
survival of autoreactive B cells that would normally have been
eliminated by anergy (Oliver et al., 2006). In addition, Bim is
required for negative selection of T cells (Bunin et al., 2005;
Villunger et al., 2004). These data suggest that both the intrinsic
and extrinsic apoptosis pathways play important roles in the
development of SLE-like disease. However, contribution of both
pathways to the development of SLE-like disease is unknown.
Here, we examined mononuclear cells from patients with SLE
and showed that they have an antiapoptotic signature. To under-
stand the significance in the skewing the ratio of antiapoptotic
to proapoptotic members in SLE development, we generated
mice deficient for both the extrinsic death receptor Fas and the
proapoptotic Bcl-2 family member Bim (Bcl2l11�/�Faslpr/lpr).
Bcl2l11�/�Faslpr/lpr mice displayed enhanced and accelerated
splenomegaly and lymphadenopathy, including an increase in
the number of hematopoietic cells. Furthermore, Bcl2l11�/�
Faslpr/lpr mice showed severe damage to the glomeruli of the
kidneys that was associated with an increase in circulating and
deposited autoantibodies. Bcl2l11�/�Faslpr/lpr mice had an in-
crease in interglomerular proliferation and displayed an en-
hanced number of macrophages surrounding the glomeruli
and an increased number of TUNEL+ cells within the glomeruli.
Taken together, these data suggest that deficiency in Bim and
Fas is sufficient to induce a full blown lupus-like disease even
on a resistant background. Thus, therapies that target both
Bim and Fas might be more effective at controlling or inducing
remission of SLE.
RESULTS
To date, the critical factors involved in the apoptotic pathways
and their influence on the development of SLE are unknown.
Further, the interpretation of studies in mice examining the direct
role of proapoptotic genes in SLE are complicated by the strain
and background of the mice. To identify the potential apoptotic
genes involved in SLE, we performed a microarray analysis on
SLE patients and healthy controls. Previous studies have
suggested that type I interferon is critical for SLE pathogenesis
(Bennett et al., 2003); as such, all patients with SLE displayed
an increase in expression of MX1, an interferon-inducible gene.
We observed that expression of genes encoding antiapoptotic
members of the Bcl-2 family including BCL2A1 (1.8-fold; p <
0.04) and MCL1 (1.5-fold; p < 0.02) were enhanced in myeloid
cells from a subset of SLE patients as compared to healthy con-
trols (Figure 1A). There was no difference in the amounts of
expression in BCL2A1 and MCL1 from isolated T and B lympho-
cyte populations. Additionally, the expression of CFLAR, the
gene encoding FLIP, a naturally occurring inhibitor of the Fas
death pathway, was markedly elevated in myeloid cells (1.9-
fold; p < 0.007), T lymphocytes (1.4-fold; p < 0.04), and B lym-
phocytes (2.1-fold; p < 0.04) from subset of SLE patients as
compared to healthy controls (Figures 1A–1C). Taken together,
these data demonstrate a correlation between the increase in
expression of MX1 with the expression of CFLAR and MCL1 in
B cells (CFLAR: r = 0.52, p < 0.01; MCL1: r = 0.43, p < 0.04)
and myeloid cells (r = 0.52, p < 0.01; r = 0.68, p < 0.0006), and
BCL2A1 (r = 0.53, p < 0.01) in myeloid cells and the SLE pheno-
type. Given the previously established relationship between MX1
expression and disease severity (Bennett et al., 2003), these data
further suggest a connection between increased antiapoptotic
gene expression and SLE severity.
Bcl2l11�/�Faslpr/lpr Mice Display Severe Splenomegalyand LymphadenopathyTo understand the significance of the differential increase in the
expression of genes that are involved in the Bcl-2 and Fas path-
ways, we focused on mice lacking components of the Bcl-2 and
Fas pathways, specifically Bim and Fas. Previous studies with
mice lacking the antiapoptotic Bcl-2 proteins have been ham-
pered by early lethality and mice that overexpress antiapoptotic
Bcl-2 family members in a cell-type-specific manner have either
failed to display or show a mild autoimmune disease phenotype
(Hughes et al., 2006). To date, only mice lacking Bim have been
shown to develop autoimmune disease on a mixed background
(Bouillet et al., 1999). Further, unlike the vast majority of the BH3-
only proteins, Bim interacts with and sequesters all antiapoptotic
proteins (Chen et al., 2005).
Similar to mice lacking antiapoptotic Bcl-2 members, mice lack-
ing Flip die in utero, and overexpression of Flip in lymphocytes has
yielded conflicting results, none of which led to an autoimmune-
disease phenotype. Thus, thevastmajorityof studies have focused
on Fas. A previous study showed that Faslpr/lpr mice expressing
a human BCL2-Ig minigene display modest splenomegaly and
profound lymphadenopathy (Reap et al., 1995). Because recent
Figure 1. Apoptosis-Related Gene Expression Profiles of SLE and
Healthy Control PBMC Subsets
CFLAR, MCL1, and BCL2A1 gene expression in sorted CD33+ myeloid cells (A)
and CD4+ T cell (B) and CD19+ B cell (C) subsets from healthy controls (HC)
and SLE patients. Red, black, and green indicate that the gene expression is
greater than, equal to, or less than the mean amount of expression of all sam-
ples. Myxovirus resistance-1 (MX1) has been shown previously to correlate
with SLE disease activity (Bennett et al., 2003).
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 207
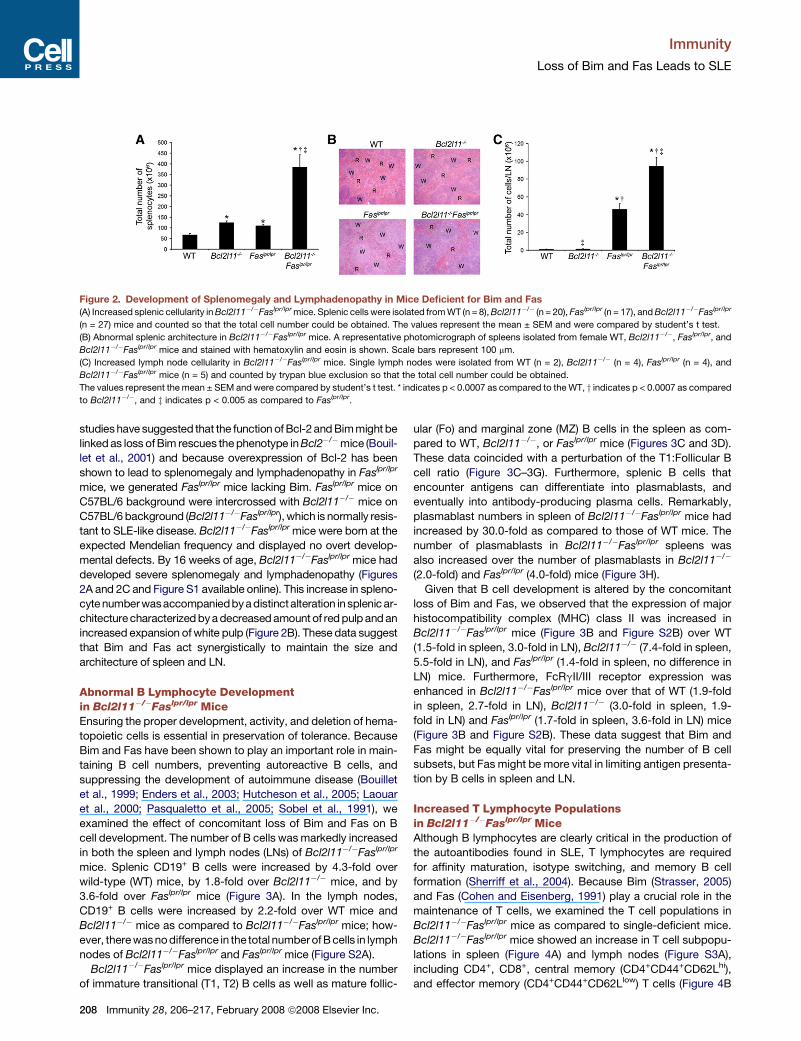
Immunity
Loss of Bim and Fas Leads to SLE
Figure 2. Development of Splenomegaly and Lymphadenopathy in Mice Deficient for Bim and Fas(A) Increased splenic cellularity in Bcl2l11�/�Faslpr/lpr mice. Splenic cells were isolated from WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17), and Bcl2l11�/�Faslpr/lpr
(n = 27) mice and counted so that the total cell number could be obtained. The values represent the mean ± SEM and were compared by student’s t test.
(B) Abnormal splenic architecture in Bcl2l11�/�Faslpr/lpr mice. A representative photomicrograph of spleens isolated from female WT, Bcl2l11�/�, Faslpr/lpr, and
Bcl2l11�/�Faslpr/lpr mice and stained with hematoxylin and eosin is shown. Scale bars represent 100 mm.
(C) Increased lymph node cellularity in Bcl2l11�/�Faslpr/lpr mice. Single lymph nodes were isolated from WT (n = 2), Bcl2l11�/� (n = 4), Faslpr/lpr (n = 4), and
Bcl2l11�/�Faslpr/lpr mice (n = 5) and counted by trypan blue exclusion so that the total cell number could be obtained.
The values represent the mean ± SEM and were compared by student’s t test. * indicates p < 0.0007 as compared to the WT, y indicates p < 0.0007 as compared
to Bcl2l11�/�, and z indicates p < 0.005 as compared to Faslpr/lpr.
studies have suggested that the function of Bcl-2 and Bim might be
linked as loss of Bim rescues the phenotype in Bcl2�/�mice (Bouil-
let et al., 2001) and because overexpression of Bcl-2 has been
shown to lead to splenomegaly and lymphadenopathy in Faslpr/lpr
mice, we generated Faslpr/lpr mice lacking Bim. Faslpr/lpr mice on
C57BL/6 background were intercrossed with Bcl2l11�/� mice on
C57BL/6 background (Bcl2l11�/�Faslpr/lpr), which is normally resis-
tant to SLE-like disease. Bcl2l11�/�Faslpr/lpr mice were born at the
expected Mendelian frequency and displayed no overt develop-
mental defects. By 16 weeks of age, Bcl2l11�/�Faslpr/lpr mice had
developed severe splenomegaly and lymphadenopathy (Figures
2A and 2C and Figure S1 available online). This increase in spleno-
cyte numberwasaccompaniedbya distinctalteration in splenic ar-
chitecture characterized by a decreased amount of red pulp and an
increased expansion of white pulp (Figure 2B). These data suggest
that Bim and Fas act synergistically to maintain the size and
architecture of spleen and LN.
Abnormal B Lymphocyte Developmentin Bcl2l11�/�Faslpr/lpr MiceEnsuring the proper development, activity, and deletion of hema-
topoietic cells is essential in preservation of tolerance. Because
Bim and Fas have been shown to play an important role in main-
taining B cell numbers, preventing autoreactive B cells, and
suppressing the development of autoimmune disease (Bouillet
et al., 1999; Enders et al., 2003; Hutcheson et al., 2005; Laouar
et al., 2000; Pasqualetto et al., 2005; Sobel et al., 1991), we
examined the effect of concomitant loss of Bim and Fas on B
cell development. The number of B cells was markedly increased
in both the spleen and lymph nodes (LNs) of Bcl2l11�/�Faslpr/lpr
mice. Splenic CD19+ B cells were increased by 4.3-fold over
wild-type (WT) mice, by 1.8-fold over Bcl2l11�/� mice, and by
3.6-fold over Faslpr/lpr mice (Figure 3A). In the lymph nodes,
CD19+ B cells were increased by 2.2-fold over WT mice and
Bcl2l11�/� mice as compared to Bcl2l11�/�Faslpr/lpr mice; how-
ever, there wasno difference in the total number of B cells in lymph
nodes of Bcl2l11�/�Faslpr/lpr and Faslpr/lpr mice (Figure S2A).
Bcl2l11�/�Faslpr/lpr mice displayed an increase in the number
of immature transitional (T1, T2) B cells as well as mature follic-
208 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.
ular (Fo) and marginal zone (MZ) B cells in the spleen as com-
pared to WT, Bcl2l11�/�, or Faslpr/lpr mice (Figures 3C and 3D).
These data coincided with a perturbation of the T1:Follicular B
cell ratio (Figure 3C–3G). Furthermore, splenic B cells that
encounter antigens can differentiate into plasmablasts, and
eventually into antibody-producing plasma cells. Remarkably,
plasmablast numbers in spleen of Bcl2l11�/�Faslpr/lpr mice had
increased by 30.0-fold as compared to those of WT mice. The
number of plasmablasts in Bcl2l11�/�Faslpr/lpr spleens was
also increased over the number of plasmablasts in Bcl2l11�/�
(2.0-fold) and Faslpr/lpr (4.0-fold) mice (Figure 3H).
Given that B cell development is altered by the concomitant
loss of Bim and Fas, we observed that the expression of major
histocompatibility complex (MHC) class II was increased in
Bcl2l11�/�Faslpr/lpr mice (Figure 3B and Figure S2B) over WT
(1.5-fold in spleen, 3.0-fold in LN), Bcl2l11�/� (7.4-fold in spleen,
5.5-fold in LN), and Faslpr/lpr (1.4-fold in spleen, no difference in
LN) mice. Furthermore, FcRgII/III receptor expression was
enhanced in Bcl2l11�/�Faslpr/lpr mice over that of WT (1.9-fold
in spleen, 2.7-fold in LN), Bcl2l11�/� (3.0-fold in spleen, 1.9-
fold in LN) and Faslpr/lpr (1.7-fold in spleen, 3.6-fold in LN) mice
(Figure 3B and Figure S2B). These data suggest that Bim and
Fas might be equally vital for preserving the number of B cell
subsets, but Fas might be more vital in limiting antigen presenta-
tion by B cells in spleen and LN.
Increased T Lymphocyte Populationsin Bcl2l11�/�Faslpr/lpr MiceAlthough B lymphocytes are clearly critical in the production of
the autoantibodies found in SLE, T lymphocytes are required
for affinity maturation, isotype switching, and memory B cell
formation (Sherriff et al., 2004). Because Bim (Strasser, 2005)
and Fas (Cohen and Eisenberg, 1991) play a crucial role in the
maintenance of T cells, we examined the T cell populations in
Bcl2l11�/�Faslpr/lpr mice as compared to single-deficient mice.
Bcl2l11�/�Faslpr/lpr mice showed an increase in T cell subpopu-
lations in spleen (Figure 4A) and lymph nodes (Figure S3A),
including CD4+, CD8+, central memory (CD4+CD44+CD62Lhi),
and effector memory (CD4+CD44+CD62Llow) T cells (Figure 4B

Immunity
Loss of Bim and Fas Leads to SLE
and Figure S3B). However, Bcl2l11�/�Faslpr/lpr mice displayed
a reduced amount of naive T cells (CD4+CD44lowCD62Lhi) as
compared to Bcl2l11�/� and WT mice, whereas there was no dif-
ference in the number of these cells as compared to Faslpr/lpr
mice (Figure 4B and Figure S3B). Taken together, the increase
in both central and effector memory T cells and the decrease
in naive T cells in Bcl2l11�/�Faslpr/lpr mice suggest an overall in-
crease in activation of Bcl2l11�/�Faslpr/lpr T cells. Despite the
marked increase in splenocytes in Bcl2l11�/�Faslpr/lpr mice (Fig-
ure 2), there was no difference in the number of T regulatory
(Treg) cells (CD4+CD25+FoxP3+) as compared to WT,
Bcl2l11�/�, or Faslpr/lpr mice (Figure S4A). Bcl2l11�/�Faslpr/lpr
mice also displayed a defect in the deletion and removal of po-
tentially self-reactive T cells, as indicated by the increase in the
number of CD3+B220+ DN (CD4�CD8�) T cells in Bcl2l11�/�
Faslpr/lpr mice (Figure S4B), and these cells have been previously
described in Faslpr/lpr mice (Cohen and Eisenberg, 1991). These
data suggest that Bim and Fas synergistically regulate the acti-
vation, maturation, and removal of T lymphocytes.
Loss of Bim and Fas Leads to Accumulation of ActivatedMacrophages in SpleenAlthough studies on SLE often focus on T and B lymphocytes,
antigen-presenting cells such as macrophages might play vital
roles in the pathogenesis of the disease. This population might
be of particular interest in the Bcl2l11�/�Faslpr/lpr system given
that both Bim and Fas have previously been shown to limit circu-
lating monocyte numbers as well as macrophage activation
(Brown et al., 2004; Hutcheson et al., 2005; Scatizzi et al.,
2006). There was an increase in the number of splenic and lymph
node macrophages in Bcl2l11�/�Faslpr/lpr mice (Figure 4C and
Figure S5A). Not only were there more macrophages in the
spleen of Bcl2l11�/�Faslpr/lpr mice, but these macrophages dis-
played an enhanced activation phenotype, including increased
expression of CD40, CD86, MHC class II, and FcgRII/III
(Figure 4D and Figure S5B). In contrast, the dendritic cell
population displayed only a marginal increase in markers of
activation in Bcl2l11�/�Faslpr/lpr mice (Figure S6A) as compared
to single-deficient mice. Further, there was only an increase in
numbers of dendritic cells in spleen (Figure 4C) but not in lymph
nodes as compared to Faslpr/lpr mice (Figure S6B). These data
suggest that Bim and Fas synergize to prevent abnormal macro-
phage accumulation and activation.
Increased Circulating Autoantibodiesin Bcl2l11�/�Faslpr/lpr MiceIn SLE, the presence of autoantibodies against self-antigens is
a hallmark of the disease. Because Bcl2l11�/�Faslpr/lpr mice
display several characteristics consistent with SLE, we exam-
ined serum for several common circulating autoantibodies in
Bcl2l11�/�Faslpr/lpr mice. Bcl2l11�/�Faslpr/lpr mice displayed
an increase over WT mice in total anti-IgM, total anti-IgG,
anti-dsDNA IgM (dsDNA: double-stranded DNA), anti-dsDNA
IgG, anti-ssDNA IgM (ssDNA: single-stranded DNA), anti-
ssDNA IgG, anti-histone, and anti-nuclear antibodies (Figures
5A–5E). Furthermore, the amounts of total anti-IgM, anti-dsDNA
IgM, anti-dsDNA IgG, anti-ssDNA IgM, anti-ssDNA IgG, anti-
histone, and anti-nuclear antibodies were increased over the
amounts in Bcl2l11�/� mice, and there was also an elevation
in the circulating amounts of total anti-IgG, total anti-IgM,
anti-dsDNA IgM, anti-ssDNA IgM, and anti-nuclear antibodies
over those amounts found in Faslpr/lpr mice as compared to
amounts in Bcl2l11�/�Faslpr/lpr mice (Figures 5A–5E). Although
Bcl2l11�/�Faslpr/lpr mice displayed increases in anti-DNA anti-
body amounts, these data show that the loss of Fas contributes
to the increase in class-switched IgG antibodies predominantly
over the loss of Bim. These potentially represent the autoanti-
bodies most responsible for disease pathogenesis (Figure 5).
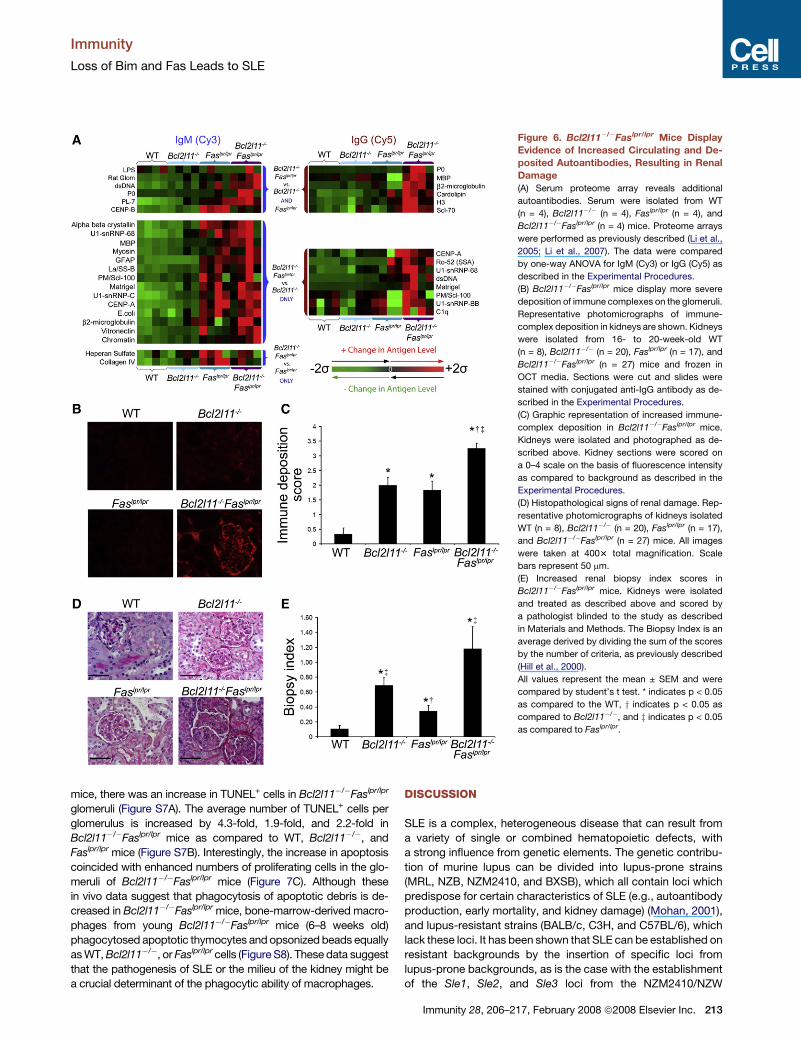
A proteome array (Li et al., 2005; Li et al., 2007), which
examines over 85 different autoantigens, revealed that
Bcl2l11�/�Faslpr/lpr serum had an increase in IgM autoantibodies
specific for collagen I, ribosomal phosphoprotein P0 (P0), trans-
fer RNA (tRNA) synthetase antigen PL-7 (PL-7), dsDNA, and rat
glomerular sonicate and IgG autoantibodies specific for Scl-70
(DNA topoisomerase I), P0, histone H3 (H3), myelin basic protein
(MBP), cardiolipin, and b2-microglobulin as compared to sin-
gle-deficient or WT mice (Figure 6A). These data indicate an
increased amount of anti-nuclear, anti-cytoplasmic, and anti-
glomerular autoantibodies in Bcl2l11�/�Faslpr/lpr mice. Further-
more, Bcl2l11�/�Faslpr/lpr mice had a markedly increased
amount of deposited IgG antibody, mainly localized at the
glomerular basement membrane, as compared to WT,
Bcl2l11�/�, and Faslpr/lpr mice (Figure 6B). When quantitated,
Bcl2l11�/�Faslpr/lpr mice displayed a 9.8-fold, 1.6-fold, and 1.8-
fold increase in fluorescence intensity as compared to WT,
Bcl2l11�/�, and Faslpr/lpr mice, respectively (Figure 6C). The
autoantigen array (Figure 6A) and immunohistology (Figures 6B
and 6C) data indicate an increased amount of anti-glomerular
antibodies in Bcl2l11�/�Faslpr/lpr mice as compared to WT,
Bcl2l11�/�, and Faslpr/lpr mice, and this could, in part, explain
the increased severity of disease in these mice.
Increased Evidence of Renal Pathologyin Bcl2l11�/�Faslpr/lpr MiceKidneys from Bcl2l11�/�Faslpr/lpr mice displayed increased glo-
merular size as compared to that of WT, Bcl2l11�/�, or Faslpr/lpr
mice. Furthermore, the glomeruli from Bcl2l11�/�Faslpr/lpr mice
showedhistologicalevidenceof renaldamage, including increased
mesangial expansion, basement membrane thickening, increased
interstitial infiltration, and loss of open capillary loops (Figure 6D).
Histological scoring of kidney destruction (Hill et al., 2000) revealed
an 11.0-fold increase in biopsy index over WT mice, a 1.7-fold
increase over Bcl2l11�/� mice, and a 3.4-fold increase over
Faslpr/lpr miceascompared toBcl2l11�/�Faslpr/lpr (Figure 6E).There
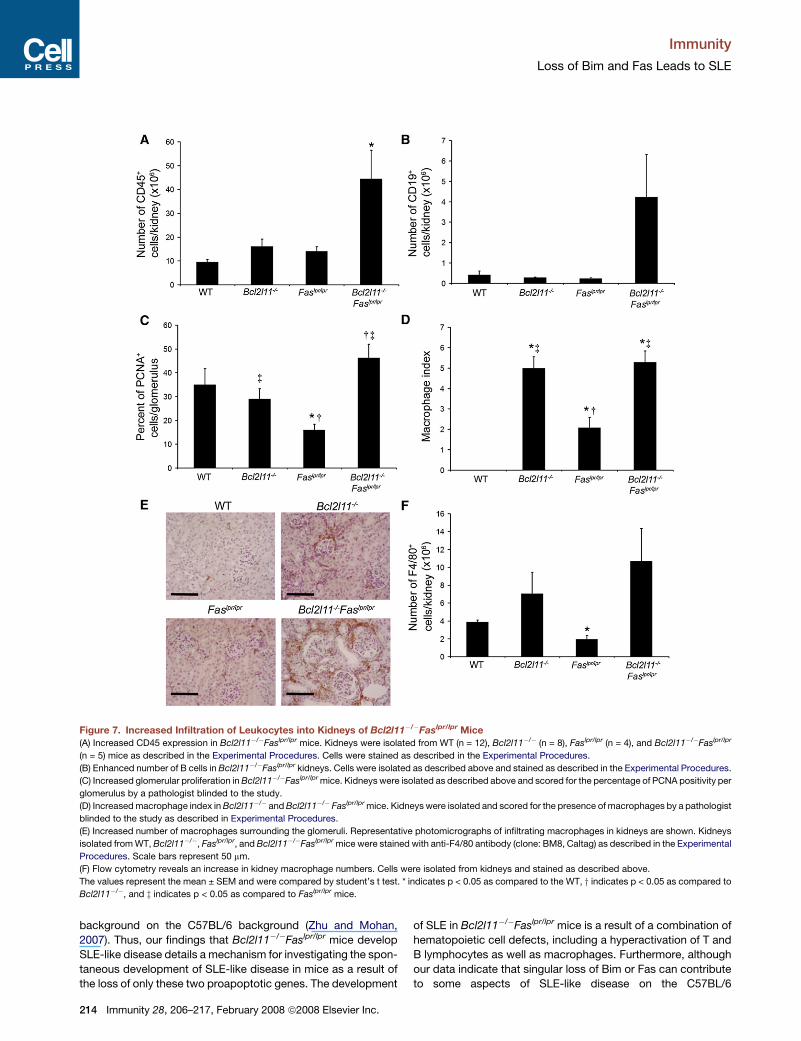
was a marked increase in the number of CD45+ leukocytes in kid-
neys isolated from Bcl2l11�/�Faslpr/lpr mice as compared to WT
(4.6-fold), Bcl2l11�/� (2.8-fold), or Faslpr/lpr mice (3.2-fold)
(Figure 7A). As a result, Bcl2l11�/�Faslpr/lpr mice displayed
a 10.2-fold increase in renal B cells (CD19+) as compared to WT
mice and an 14.8-fold and 18.4-fold increase over Bcl2l11�/�
and Faslpr/lpr mice (Figure 7B). Furthermore, there was an increase
in glomerular proliferation in Bcl2l11�/�Faslpr/lpr mice as displayed
by increased proliferating cell nuclear antigen (PCNA) staining
within the glomeruli (Figure 7C).
Because B cells only account for a portion of the cells infil-
trating into the kidneys of Bcl2l11�/�Faslpr/lpr mice, we ana-
lyzed the presence of myeloid cells. Both Bcl2l11�/� and
Bcl2l11�/�Faslpr/lpr mice exhibited an increase in the number
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 209
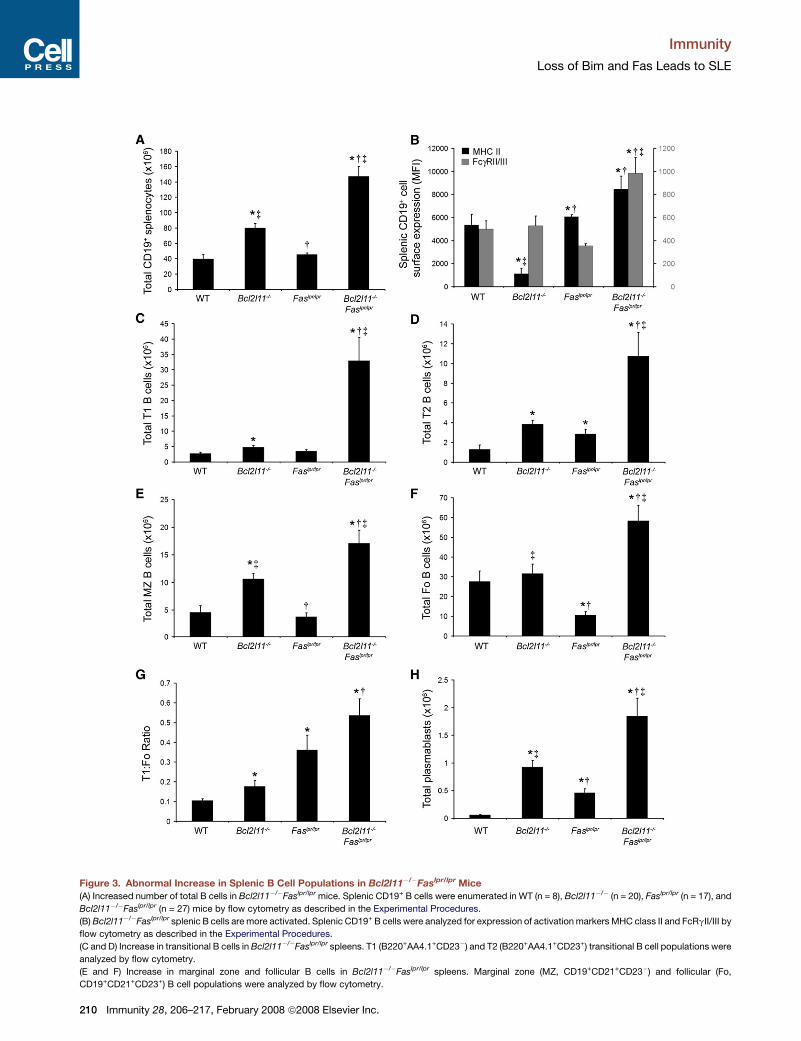
Immunity
Loss of Bim and Fas Leads to SLE
Figure 3. Abnormal Increase in Splenic B Cell Populations in Bcl2l11�/�Faslpr/lpr Mice
(A) Increased number of total B cells in Bcl2l11�/�Faslpr/lpr mice. Splenic CD19+ B cells were enumerated in WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17), and
Bcl2l11�/�Faslpr/lpr (n = 27) mice by flow cytometry as described in the Experimental Procedures.
(B) Bcl2l11�/�Faslpr/lpr splenic B cells are more activated. Splenic CD19+ B cells were analyzed for expression of activation markers MHC class II and FcRgII/III by
flow cytometry as described in the Experimental Procedures.
(C and D) Increase in transitional B cells in Bcl2l11�/�Faslpr/lpr spleens. T1 (B220+AA4.1+CD23�) and T2 (B220+AA4.1+CD23+) transitional B cell populations were
analyzed by flow cytometry.
(E and F) Increase in marginal zone and follicular B cells in Bcl2l11�/�Faslpr/lpr spleens. Marginal zone (MZ, CD19+CD21+CD23�) and follicular (Fo,
CD19+CD21+CD23+) B cell populations were analyzed by flow cytometry.
210 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.
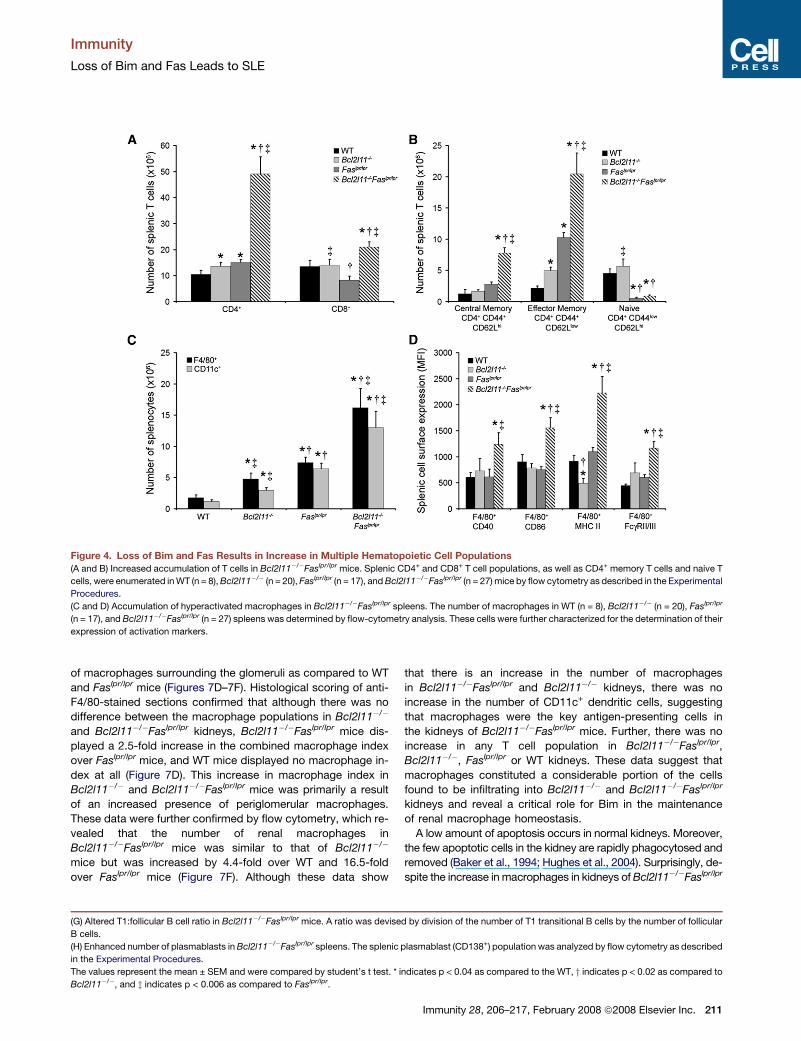
Immunity
Loss of Bim and Fas Leads to SLE
Figure 4. Loss of Bim and Fas Results in Increase in Multiple Hematopoietic Cell Populations
(A and B) Increased accumulation of T cells in Bcl2l11�/�Faslpr/lpr mice. Splenic CD4+ and CD8+ T cell populations, as well as CD4+ memory T cells and naive T
cells, were enumerated in WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17), and Bcl2l11�/�Faslpr/lpr (n = 27) mice by flow cytometry as described in the Experimental
Procedures.
(C and D) Accumulation of hyperactivated macrophages in Bcl2l11�/�Faslpr/lpr spleens. The number of macrophages in WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr
(n = 17), and Bcl2l11�/�Faslpr/lpr (n = 27) spleens was determined by flow-cytometry analysis. These cells were further characterized for the determination of their
expression of activation markers.
of macrophages surrounding the glomeruli as compared to WT
and Faslpr/lpr mice (Figures 7D–7F). Histological scoring of anti-
F4/80-stained sections confirmed that although there was no
difference between the macrophage populations in Bcl2l11�/�
and Bcl2l11�/�Faslpr/lpr kidneys, Bcl2l11�/�Faslpr/lpr mice dis-
played a 2.5-fold increase in the combined macrophage index
over Faslpr/lpr mice, and WT mice displayed no macrophage in-
dex at all (Figure 7D). This increase in macrophage index in
Bcl2l11�/� and Bcl2l11�/�Faslpr/lpr mice was primarily a result
of an increased presence of periglomerular macrophages.
These data were further confirmed by flow cytometry, which re-
vealed that the number of renal macrophages in
Bcl2l11�/�Faslpr/lpr mice was similar to that of Bcl2l11�/�
mice but was increased by 4.4-fold over WT and 16.5-fold
over Faslpr/lpr mice (Figure 7F). Although these data show
that there is an increase in the number of macrophages
in Bcl2l11�/�Faslpr/lpr and Bcl2l11�/� kidneys, there was no
increase in the number of CD11c+ dendritic cells, suggesting
that macrophages were the key antigen-presenting cells in
the kidneys of Bcl2l11�/�Faslpr/lpr mice. Further, there was no
increase in any T cell population in Bcl2l11�/�Faslpr/lpr,
Bcl2l11�/�, Faslpr/lpr or WT kidneys. These data suggest that
macrophages constituted a considerable portion of the cells
found to be infiltrating into Bcl2l11�/� and Bcl2l11�/�Faslpr/lpr
kidneys and reveal a critical role for Bim in the maintenance
of renal macrophage homeostasis.
A low amount of apoptosis occurs in normal kidneys. Moreover,
the few apoptotic cells in the kidney are rapidly phagocytosed and
removed (Baker et al., 1994; Hughes et al., 2004). Surprisingly, de-
spite the increase in macrophages in kidneys of Bcl2l11�/�Faslpr/lpr
(G) Altered T1:follicular B cell ratio in Bcl2l11�/�Faslpr/lpr mice. A ratio was devised by division of the number of T1 transitional B cells by the number of follicular
B cells.
(H) Enhanced number of plasmablasts in Bcl2l11�/�Faslpr/lpr spleens. The splenic plasmablast (CD138+) population was analyzed by flow cytometry as described
in the Experimental Procedures.
The values represent the mean ± SEM and were compared by student’s t test. * indicates p < 0.04 as compared to the WT, y indicates p < 0.02 as compared to
Bcl2l11�/�, and z indicates p < 0.006 as compared to Faslpr/lpr.
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 211
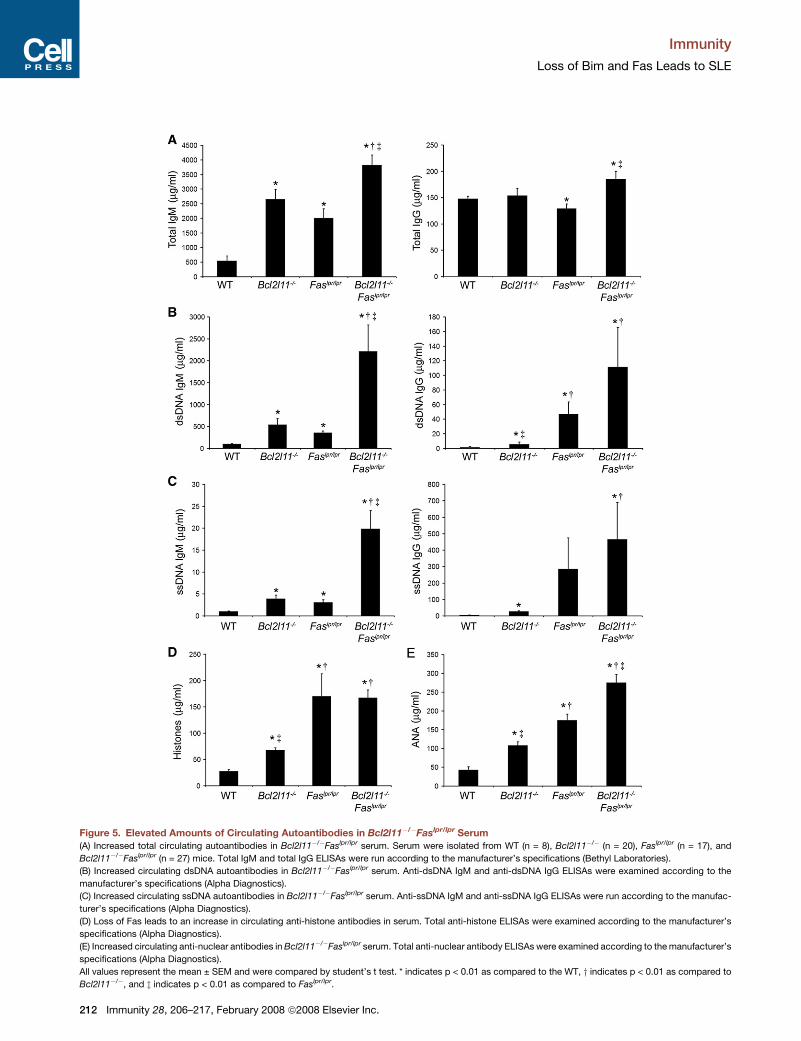
Immunity
Loss of Bim and Fas Leads to SLE
Figure 5. Elevated Amounts of Circulating Autoantibodies in Bcl2l11�/�Faslpr/lpr Serum
(A) Increased total circulating autoantibodies in Bcl2l11�/�Faslpr/lpr serum. Serum were isolated from WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17), and
Bcl2l11�/�Faslpr/lpr (n = 27) mice. Total IgM and total IgG ELISAs were run according to the manufacturer’s specifications (Bethyl Laboratories).
(B) Increased circulating dsDNA autoantibodies in Bcl2l11�/�Faslpr/lpr serum. Anti-dsDNA IgM and anti-dsDNA IgG ELISAs were examined according to the
manufacturer’s specifications (Alpha Diagnostics).
(C) Increased circulating ssDNA autoantibodies in Bcl2l11�/�Faslpr/lpr serum. Anti-ssDNA IgM and anti-ssDNA IgG ELISAs were run according to the manufac-
turer’s specifications (Alpha Diagnostics).
(D) Loss of Fas leads to an increase in circulating anti-histone antibodies in serum. Total anti-histone ELISAs were examined according to the manufacturer’s
specifications (Alpha Diagnostics).
(E) Increased circulating anti-nuclear antibodies in Bcl2l11�/�Faslpr/lpr serum. Total anti-nuclear antibody ELISAs were examined according to the manufacturer’s
specifications (Alpha Diagnostics).
All values represent the mean ± SEM and were compared by student’s t test. * indicates p < 0.01 as compared to the WT, y indicates p < 0.01 as compared to
Bcl2l11�/�, and z indicates p < 0.01 as compared to Faslpr/lpr.
212 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.
Immunity
Loss of Bim and Fas Leads to SLE
Figure 6. Bcl2l11�/�Faslpr/lpr Mice Display
Evidence of Increased Circulating and De-
posited Autoantibodies, Resulting in Renal
Damage
(A) Serum proteome array reveals additional
autoantibodies. Serum were isolated from WT
(n = 4), Bcl2l11�/� (n = 4), Faslpr/lpr (n = 4), and
Bcl2l11�/�Faslpr/lpr (n = 4) mice. Proteome arrays
were performed as previously described (Li et al.,
2005; Li et al., 2007). The data were compared
by one-way ANOVA for IgM (Cy3) or IgG (Cy5) as
described in the Experimental Procedures.
(B) Bcl2l11�/�Faslpr/lpr mice display more severe
deposition of immune complexes on the glomeruli.
Representative photomicrographs of immune-
complex deposition in kidneys are shown. Kidneys
were isolated from 16- to 20-week-old WT
(n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17), and
Bcl2l11�/�Faslpr/lpr (n = 27) mice and frozen in
OCT media. Sections were cut and slides were
stained with conjugated anti-IgG antibody as de-
scribed in the Experimental Procedures.
(C) Graphic representation of increased immune-
complex deposition in Bcl2l11�/�Faslpr/lpr mice.
Kidneys were isolated and photographed as de-
scribed above. Kidney sections were scored on
a 0–4 scale on the basis of fluorescence intensity
as compared to background as described in the
Experimental Procedures.
(D) Histopathological signs of renal damage. Rep-
resentative photomicrographs of kidneys isolated
WT (n = 8), Bcl2l11�/� (n = 20), Faslpr/lpr (n = 17),
and Bcl2l11�/�Faslpr/lpr (n = 27) mice. All images
were taken at 4003 total magnification. Scale
bars represent 50 mm.
(E) Increased renal biopsy index scores in
Bcl2l11�/�Faslpr/lpr mice. Kidneys were isolated
and treated as described above and scored by
a pathologist blinded to the study as described
in Materials and Methods. The Biopsy Index is an
average derived by dividing the sum of the scores
by the number of criteria, as previously described
(Hill et al., 2000).
All values represent the mean ± SEM and were
compared by student’s t test. * indicates p < 0.05
as compared to the WT, y indicates p < 0.05 as
compared to Bcl2l11�/�, and z indicates p < 0.05
as compared to Faslpr/lpr.
mice, there was an increase in TUNEL+ cells in Bcl2l11�/�Faslpr/lpr
glomeruli (Figure S7A). The average number of TUNEL+ cells per
glomerulus is increased by 4.3-fold, 1.9-fold, and 2.2-fold in
Bcl2l11�/�Faslpr/lpr mice as compared to WT, Bcl2l11�/�, and
Faslpr/lpr mice (Figure S7B). Interestingly, the increase in apoptosis
coincided with enhanced numbers of proliferating cells in the glo-
meruli of Bcl2l11�/�Faslpr/lpr mice (Figure 7C). Although these
in vivo data suggest that phagocytosis of apoptotic debris is de-
creased in Bcl2l11�/�Faslpr/lpr mice, bone-marrow-derived macro-
phages from young Bcl2l11�/�Faslpr/lpr mice (6–8 weeks old)
phagocytosed apoptotic thymocytes and opsonized beads equally
as WT, Bcl2l11�/�, or Faslpr/lpr cells (Figure S8). These data suggest
that the pathogenesis of SLE or the milieu of the kidney might be
a crucial determinant of the phagocytic ability of macrophages.
DISCUSSION
SLE is a complex, heterogeneous disease that can result from
a variety of single or combined hematopoietic defects, with
a strong influence from genetic elements. The genetic contribu-
tion of murine lupus can be divided into lupus-prone strains
(MRL, NZB, NZM2410, and BXSB), which all contain loci which
predispose for certain characteristics of SLE (e.g., autoantibody
production, early mortality, and kidney damage) (Mohan, 2001),
and lupus-resistant strains (BALB/c, C3H, and C57BL/6), which
lack these loci. It has been shown that SLE can be established on
resistant backgrounds by the insertion of specific loci from
lupus-prone backgrounds, as is the case with the establishment
of the Sle1, Sle2, and Sle3 loci from the NZM2410/NZW
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 213
Immunity
Loss of Bim and Fas Leads to SLE
background on the C57BL/6 background (Zhu and Mohan,
2007). Thus, our findings that Bcl2l11�/�Faslpr/lpr mice develop
SLE-like disease details a mechanism for investigating the spon-
taneous development of SLE-like disease in mice as a result of
the loss of only these two proapoptotic genes. The development
of SLE in Bcl2l11�/�Faslpr/lpr mice is a result of a combination of
hematopoietic cell defects, including a hyperactivation of T and
B lymphocytes as well as macrophages. Furthermore, although
our data indicate that singular loss of Bim or Fas can contribute
to some aspects of SLE-like disease on the C57BL/6
Figure 7. Increased Infiltration of Leukocytes into Kidneys of Bcl2l11�/�Faslpr/lpr Mice(A) Increased CD45 expression in Bcl2l11�/�Faslpr/lpr mice. Kidneys were isolated from WT (n = 12), Bcl2l11�/� (n = 8), Faslpr/lpr (n = 4), and Bcl2l11�/�Faslpr/lpr
(n = 5) mice as described in the Experimental Procedures. Cells were stained as described in the Experimental Procedures.
(B) Enhanced number of B cells in Bcl2l11�/�Faslpr/lpr kidneys. Cells were isolated as described above and stained as described in the Experimental Procedures.
(C) Increased glomerular proliferation in Bcl2l11�/�Faslpr/lpr mice. Kidneys were isolated as described above and scored for the percentage of PCNA positivity per
glomerulus by a pathologist blinded to the study.
(D) Increased macrophage index in Bcl2l11�/� and Bcl2l11�/� Faslpr/lpr mice. Kidneys were isolated and scored for the presence of macrophages by a pathologist
blinded to the study as described in Experimental Procedures.
(E) Increased number of macrophages surrounding the glomeruli. Representative photomicrographs of infiltrating macrophages in kidneys are shown. Kidneys
isolated from WT, Bcl2l11�/�, Faslpr/lpr, and Bcl2l11�/�Faslpr/lpr mice were stained with anti-F4/80 antibody (clone: BM8, Caltag) as described in the Experimental
Procedures. Scale bars represent 50 mm.
(F) Flow cytometry reveals an increase in kidney macrophage numbers. Cells were isolated from kidneys and stained as described above.
The values represent the mean ± SEM and were compared by student’s t test. * indicates p < 0.05 as compared to the WT, y indicates p < 0.05 as compared to
Bcl2l11�/�, and z indicates p < 0.05 as compared to Faslpr/lpr mice.
214 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.

Immunity
Loss of Bim and Fas Leads to SLE
background, they also specifically implicate Fas as a mediator of
B cell class switching and antigen presentation and Bim as a po-
tent regulator of renal macrophage infiltration, suggesting a syn-
ergistic role for the intrinsic and extrinsic apoptosis pathways in
the development of murine SLE and providing a model of SLE on
the C57BL/6 background that requires only 4 months to develop.
Although it has been suggested that Fas does not play a role in
the BCR-induced deletion of transitional B cells (King and Mon-
roe, 2000), the increase in T1 B cells in Bcl2l11�/�Faslpr/lpr mice
as compared to Bcl2l11�/�mice suggests that the loss of Fas en-
hances the build up of T1 B cells in Bcl2l11�/�Faslpr/lpr mice. A
lack of apoptosis in T1 B cells has been implicated in the develop-
ment of lupus in the New Zealand black (NZB) model, and it has
been suggested that this is an effect of increased Bcl-2 expres-
sion (Roy et al., 2005). Because Bcl-2 is an antiapoptotic protein
and a binding partner of Bim, loss of Bim could result in higher
amounts or higher concentrations of Bcl-2. An increase in the ra-
tio of immature transitional T1 to mature follicular B cells suggests
that Bcl2l11�/�Faslpr/lpr mice have a defect in the deletion of B
cells, and potentially a defect in B cell anergy, as has been previ-
ously reported in Bcl2l11�/� mice (Oliver et al., 2006). Further-
more, a loss of tolerance in early B cell development has been
associated with the development of SLE in humans (Yurasov
et al., 2005). Given that SLE is mediated by autoantibodies, an in-
crease in plasmablasts, as in Bcl2l11�/�Faslpr/lpr mice, would
allow for greater production of autoantibodies and increase the
severity of SLE. This increase in plasmablasts and the activated
B cell phenotype would explain the high titer of autoantibodies
found in Bcl2l11�/�Faslpr/lpr mice.
Although B lymphocytes are critical in the production of the
autoantibodies found in SLE, T lymphocytes are required for
affinity maturation, isotype switching, and memory B cell forma-
tion (Sherriff et al., 2004). The increase in both central and effec-
tor memory T cells suggests an enhanced state of activation in
Bcl2l11�/�Faslpr/lpr mice that correlates with the hyperactivated
B cell phenotype observed in both spleen and lymph nodes. It
has recently been shown that the numbers of T regulatory cells
are reduced in mouse models of SLE as well as in human
patients (Hsu et al., 2006); correspondingly, we show that T reg-
ulatory cell numbers in Bcl2l11�/�Faslpr/lpr mice were not differ-
ent than those in WT, Bcl2l11�/�, or Faslpr/lpr mice. Accordingly,
the titers of IL-17 in Bcl2l11�/�Faslpr/lpr serum were not increased
(J.H. and H.P., unpublished data). Bcl2l11�/�Faslpr/lpr mice also
display a defect in the deletion and removal of potentially self-
reactive T cells, as indicated by the increase in the number of
CD3+B220+ DN T cells in Bcl2l11�/�Faslpr/lpr mice.
SLE is characterized by circulating autoantibodies, especially
autoantibodies specific for anti-nuclear antigens (Tan et al.,
1966). In Bcl2l11�/� and Faslpr/lpr mice (Figures 5A–5E), there is
a consistent increase in both total IgM and total IgG autoantibodies,
as well as other antibodies, as determined by enzyme-linked
immunosorbent assay (ELISA). Bcl2l11�/�Faslpr/lpr mice also
displayed an increase in the expression of autoantibodies found
in patients with lupus as compared to WT, Bcl2l11�/�, or Faslpr/lpr
mice, as determined by a serum proteome array. Furthermore,
Bcl2l11�/�Faslpr/lpr mice displayed a higher amount of autoanti-
bodiesspecific for rat glomerular sonicate, further suggesting an in-
crease in glomerular autoantibodies in Bcl2l11�/�Faslpr/lpr mice.
The overall profile generated by these proteome array data is
more similar between Bcl2l11�/�Faslpr/lpr and Faslpr/lpr mice as
compared to Bcl2l11�/� or WT mice. These data suggest that the
combined hematopoietic defects in Bcl2l11�/�Faslpr/lpr mice con-
tribute to the loss of tolerance and the development of SLE-like
characteristics. In addition to the increased titer of circulating IgM
and IgG antibodies, Bcl2l11�/�Faslpr/lpr mice also have an increase
in the amount of IgG deposited in kidneys, which might be a conse-
quenceof increasedBcell presence in thekidney.The depositionof
immune complexes at the basement membrane of the kidney is
consistent with increased renal destruction in Bcl2l11�/�Faslpr/lpr
mice. Collectively, these data demonstrate that the deficiency in
Bim and Fas alone induce a rapid and severe lupus-like phenotype
on a normally autoimmune-resistant background.
EXPERIMENTAL PROCEDURES
Mice
Bcl2l11�/� (C57BL/6) mice were a kind gift from A. Strasser (Walter and Eliza
Hall Institute, Melbourne, Australia) (Bouillet et al., 1999). Congenic control
(C57BL/6, WT) and Fas mutant Faslpr/lpr (B6.MRL-Faslpr/J) mice were pur-
chased from the Jackson Laboratory (Bar Harbor, ME). Bcl2l11�/�Faslpr/lpr
mice on the C57BL/6 background were established at Saint Louis University
by the crossing of Bcl2l11�/� and Faslpr/lpr mice. Mice used for this study
were 16- to 20-week-old females maintained in a stress-free environment.
The Animal Care and Use Committee at Saint Louis University approved all
experiments on mice.
Study Participants
The study protocol was approved by the Internal Review Board of the Univer-
sity of Texas Southwestern Medical Center (UTSW). Fourteen SLE patients
and matched healthy controls were recruited from the UTSW registry. All
patients fulfilled at least four of the 11 American College of Rheumatology
classification criteria for SLE. Detailed clinical characteristics of study subjects
along with their medications were collected for each patient as described (Han
et al., 2005). Disease activity at time of blood acquisition was determined with
the Systemic Lupus Erythematosus Disease Activity Index (SLEDAI) score
(Bombardier et al., 1992), and patients were further classified according to
disease severity where SLEDAI less than 6 was considered mild and SLEDAI
more than 6 was moderate to severe.
Cell Acquisition and Sorting
Peripheral blood was collected in heparinized tubes and processed. Peripheral
blood mononuclear cells (PBMCs) were isolated with density-gradient centri-
fugation over Ficoll-Hypaque, and cells were labeled with fluorochrome-
conjugated antibodies including CD33, CD3, CD4, and CD19 (BD Biosciences,
PharMingen) and sorted with a 6+ color analyzer and sorter MoFlo Flow
Cytometer (DakoCytomation) into individual subpopulations of CD19+ B cells,
CD33+ myeloid cells, and CD4+ T cells. Not all subsets were collected for all
samples. Purity of the individual populations after sorting was verified to be
greater than 95%.
RNA Extraction and Chip Hybridization
RNA was extracted with Trizol (Invitrogen) and further purified by RNeasy
columns (QIAGEN). RNA integrity and yield was confirmed by Bioanalyzer (Agi-
lent Technologies). Complementary RNA (cRNA) probes were generated (Am-
bion), and cRNA was hybridized to Affymetrix HG-U133A GeneChips with ad-
herence to stringent quality control criteria for chip hybridization (Expression
Analysis Technical Manual, Affymetrix) for all samples.
Data Processing
Genechip Operating Software v1.1 by Affymetrix was employed for the gener-
ation of expression values,and data was imported into GeneSpring v. 7.3 soft-
ware (Agilent Technologies). The data were normalized, filtered for flags and
expression, and subjected to statistical analysis (one-way analysis of variance
[ANOVA]). Hierarchical clustering was performed with Cluster and data was
visualized with TreeView (EisenSoftware).
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 215

Immunity
Loss of Bim and Fas Leads to SLE
Histopathologic Studies
Kidneys were isolated and prepared for histological analysis by being cut in
half laterally and fixed in 10% formalin. Specimens were then embedded in
paraffin, sectioned (3 mm), and stained with hematoxylin and eosin (H&E) or pe-
riodic acid-Schiff (PAS) stain. All sections were scored by a pathologist blinded
to the study (G.K.H.) on the basis of a modification of a previously reported
scoring system (Hill et al., 2000). Paraffin embedded sections were incubated
with Dako target retrieval solution prior to the staining for F4/80 as previously
described (Hutcheson et al., 2005). Anti-F4/80-stained kidneys were scored by
a pathologist blinded to the study for the presence of macrophages in the
glomeruli, periglomerular space, cortex, medulla, and lymphoid infiltrate.
Each region was scored on a scale from 0 to 3. An overall macrophage index
was determined by addition of the values from each region and division by the
number of regions (macrophage index = total score/5). Kidneys were also
embedded in optimal cutting temperature (OCT) embedding media, snap
frozen, sectioned (3 mm), and stained for IgG. The sections were incubated
with rhodamine-red-conjugated donkey anti-mouse IgG (Jackson ImmunoRe-
search Laboratories, West Grove, PA) at 37�C for 30 min in a moist box.
Images taken at 4003 were scored from 0 to 4 on the basis of fluorescence
intensity as compared to the background for the sample. For apoptotic-
body analysis, paraffin-embedded kidney sections were stained for TUNEL
(Roche Biochemical, Indianapolis, IN, in situ death detection kit). Three differ-
ent areas per kidney of TUNEL+ cells were identified at 4003 power. TUNEL
scores were obtained by calculation of an average number of TUNEL+ cells/
glomerulus in five glomeruli. Spleens were isolated and fixed in 10% parafor-
maldehyde. Fixed specimens were embedded in paraffin and sectioned.
Sections were stained with H&E, and photomicrographs were taken at 2003
magnification for the determination of splenic architecture. All photographs
were taken on a Nikon (Tokyo, Japan) microscope equipped with the Nikon
digital camera DMX1200.
Measurement of Autoantibody Titers
Peripheral blood was isolated from mice by cardiac stick and allowed to clot,
and serum was separated out by centrifugation. Serum was screened for total
IgM or IgG (Bethyl Laboratories, Montgomery, TX) and for autoantibodies to
dsDNA (IgM and IgG), ssDNA (IgM and IgG), histones (total Ig), and anti-
nuclear antibodies (ANAs, total Ig) with ELISA, as described by the manufac-
turer (Alpha Diagnostics International, San Antonio, TX).
Immunophenotyping
Spleen, cervical lymph nodes, and kidneys were isolated from 16- to 20-week-
old mice and prepared as previously described (Hutcheson et al., 2005). Cells
were then stained for 30 min with fluorochrome-conjugated antibodies specific
to CD3, CD4, CD8, CD11b, CD11c, CD19, CD21, CD23, CD25, CD40, CD44,
CD45, CD62L, CD69, CD86, CD138 (Syndecan-1), Gr-1, NK1.1, TCRb, AA4.1
(BD Biosciences, PharMingen), CD45R (B220), F4/80 (BM8) (Invitrogen, Cal-
tag, Carlsbad, CA), or FoxP3 (eBioscience, San Diego, CA). After the staining,
cells were washed and fixed as previously described (Hutcheson et al., 2005).
For the performance of intracellular FoxP3 staining, after fixation, cells were
permeabilized with 0.3% saponin for 15 min prior to antibody staining, and
then antibody was added for 30 min. Cells were then washed with 0.1% sapo-
nin. Data were acquired and analyzed as previously described (Hutcheson and
Perlman, 2007).
Glomerular Proteome Array
Serum from WT, Bcl2l11�/�, Faslpr/lpr, and Bcl2l11�/�Faslpr/lpr (all n = 4) mice
were analyzed by a proteome array (Li et al., 2005; Li et al., 2007). The raw
data were normalized to multiple technical controls on each of the 16 arrays.
Antigen spots that showed no fluorescence were assigned a signal intensity
of 0 and designated as missing values in the subsequent statistical analysis,
which was conducted with a one-way ANOVA separately on the IgM (Cy3)
and IgG (Cy5) datasets. Pair-wise comparisons were conducted as follows:
Bcl2l11�/�Faslpr/lpr versus WT, Bcl2l11�/�Faslpr/lpr versus Bcl2l11�/�,
Bcl2l11�/�Faslpr/lpr versus Faslpr/lpr Bcl2l11�/� versus WT, and Faslpr/lpr versus
WT. The comparisons of interest were Bcl2l11�/�Faslpr/lpr versus Bcl2l11�/�
and Bcl2l11�/�Faslpr/lpr versus Faslpr/lpr. Significant differences in autoantibody
profile were identified with p % 0.05 for the two comparisons of interest. The
significant autoantibody profile differences were then placed into three groups:
216 Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc.
only changed in Bcl2l11�/�Faslpr/lpr�/� versus Bcl2l11�/�, only changed in
Bcl2l11�/�Faslpr/lpr versus Faslpr/lpr, and changed in both comparisons.
Statistical Analysis
Results were expressed as the mean ± standard error of the mean (SEM).
Differences between groups were analyzed with Student’s t tests unless
otherwise noted. Correlations were determined by Spearman rank order.
Supplemental Data
Eight figures are available at http://www.immunity.com/cgi/content/full/28/2/
206/DC1/.
ACKNOWLEDGMENTS
We would like to thank J. Eslick and S. Koehm for technical assistance in pre-
paring and collecting flow-cytometry data for this project, as well as I. Rifkin
and M. Rauchman for their help in interpreting our preliminary data. Our thanks
also go to M. Green, L. Morrison, G. Chinnadurai, and T. Ferguson for providing
advice in the preparation of this work. The authors thank A.W. Becker for as-
sistance with patient sample collection and A. Mobley of the UTSW flow-cy-
tometry core for cell sorting. We thank K.H. Dao and B.K. Han for patients’ clin-
ical evaluations. We thank D.R. Karp and E.K. Wakeland and the members of
the Simmons Arthritis Research Center for their support. This project is sup-
ported from a predoctoral American Heart Association grant to J.H.
(0710060Z), from a predoctoral American Heart Association grant to J.C.S.
(0515499Z), and from National Institutes of Health (NIH) (AR050250,
AI067590) and the Saint Louis University Autoimmune Disease fund to H.P.
L.D. was supported in part by AR45293 and the Simmons Arthritis Research
Center. This work was also supported by NIH grant U54 AI057160 to the Mid-
west Regional Center of Excellence for Biodefense and Emerging Infectious
Diseases Research.
Received: October 25, 2007
Revised: December 11, 2007
Accepted: December 14, 2007
Published online: February 14, 2008
REFERENCES
Baker, A.J., Mooney, A., Hughes, J., Lombardi, D., Johnson, R.J., and Savill, J.
(1994). Mesangial cell apoptosis: The major mechanism for resolution of
glomerular hypercellularity in experimental mesangial proliferative nephritis.
J. Clin. Invest. 94, 2105–2116.
Bennett, L., Palucka, A.K., Arce, E., Cantrell, V., Borvak, J., Banchereau, J.,
and Pascual, V. (2003). Interferon and granulopoiesis signatures in systemic
lupus erythematosus blood. J. Exp. Med. 197, 711–723.
Bombardier, C., Gladman, D.D., Urowitz, M.B., Caron, D., and Chang, C.H.
(1992). Derivation of the SLEDAI. A disease activity index for lupus patients.
The Committee on Prognosis Studies in SLE. Arthritis Rheum. 35, 630–640.
Bouillet, P., Metcalf, D., Huang, D.C., Tarlinton, D.M., Kay, T.W., Kontgen, F.,
Adams, J.M., and Strasser, A. (1999). Proapoptotic Bcl-2 relative Bim required
for certain apoptotic responses, leukocyte homeostasis, and to preclude
autoimmunity. Science 286, 1735–1738.
Bouillet, P., Cory, S., Zhang, L.C., Strasser, A., and Adams, J.M. (2001).
Degenerative disorders caused by Bcl-2 deficiency prevented by loss of its
BH3-only antagonist Bim. Dev. Cell 1, 645–653.
Brown, N.J., Hutcheson, J., Bickel, E., Scatizzi, J.C., Albee, L.D., Haines, G.K.,
3rd, Eslick, J., Bradley, K., Taricone, E., and Perlman, H. (2004). Fas death
receptor signaling represses monocyte numbers and macrophage activation
in vivo. J. Immunol. 173, 7584–7593.
Bunin, A., Khwaja, F.W., and Kersh, G.J. (2005). Regulation of Bim by TCR
signals in CD4/CD8 double-positive thymocytes. J. Immunol. 175, 1532–1539.
Chen, L., Willis, S.N., Wei, A., Smith, B.J., Fletcher, J.I., Hinds, M.G., Colman,
P.M., Day, C.L., Adams, J.M., and Huang, D.C. (2005). Differential targeting of
prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary
apoptotic function. Mol. Cell 17, 393–403.

Immunity
Loss of Bim and Fas Leads to SLE
Cohen, P.L. (2006). Apoptotic cell death and lupus. Springer Semin. Immuno-
pathol. 28, 145–152.
Cohen, P.L., and Eisenberg, R.A. (1991). Lpr and gld: Single gene models of
systemic autoimmunity and lymphoproliferative disease. Annu. Rev. Immunol.
9, 243–269.
Enders, A., Bouillet, P., Puthalakath, H., Xu, Y., Tarlinton, D.M., and Strasser,
A. (2003). Loss of the pro-apoptotic BH3-only Bcl-2 family member Bim
inhibits BCR stimulation-induced apoptosis and deletion of autoreactive B
cells. J. Exp. Med. 198, 1119–1126.
Han, B.K., White, A.M., Dao, K.H., Karp, D.R., Wakelan, E.K., and Davis, L.S.
(2005). Increased prevalence of activated CD70+CD4+ T cells in the periphery
of patients with systemic lupus erythematosus. Lupus 14, 598–606.
Hill, G.S., Delahousse, M., Nochy, D., Tomkiewicz, E., Remy, P., Mignon, F.,
and Mery, J.P. (2000). A new morphologic index for the evaluation of renal
biopsies in lupus nephritis. Kidney Int. 58, 1160–1173.
Hsu, W.T., Suen, J.L., and Chiang, B.L. (2006). The role of CD4CD25 T cells in
autoantibody production in murine lupus. Clin. Exp. Immunol. 145, 513–519.
Hughes, J., Cailhier, J.F., Watson, S., and Savill, J.S. (2004). Apoptosis in
glomerulonephritis. Rheum. Dis. Clin. North Am. 30, 655–676.
Hughes, P., Bouillet, P., and Strasser, A. (2006). Role of Bim and other Bcl-2
family members in autoimmune and degenerative diseases. Curr. Dir.
Autoimmun. 9, 74–94.
Hutcheson, J., and Perlman, H. (2007). Loss of Bim results in abnormal
accumulation of mature CD4(-)CD8(-)CD44(-)CD25(-) thymocytes. Immunobi-
ology 212, 629–636.
Hutcheson, J., Scatizzi, J.C., Bickel, E., Brown, N.J., Bouillet, P., Strasser, A.,
and Perlman, H. (2005). Combined loss of proapoptotic genes Bak or Bax with
Bim synergizes to cause defects in hematopoiesis and in thymocyte apopto-
sis. J. Exp. Med. 201, 1949–1960.
King, L.B., and Monroe, J.G. (2000). Immunobiology of the immature B cell:
Plasticity in the B-cell antigen receptor-induced response fine tunes negative
selection. Immunol. Rev. 176, 86–104.
Laouar, Y., Vasseur, F., Moreau, G., Garcia, C., Pasqualetto, V., Wache, A.C.,
and Ezine, S. (2000). Selective involvement of the Fas (CD95)/Fas ligand
pathway in bone marrow B cell progenitors. Eur. J. Immunol. 30, 1402–1409.
Li, Q.Z., Xie, C., Wu, T., Mackay, M., Aranow, C., Putterman, C., and Mohan, C.
(2005). Identification of autoantibody clusters that best predict lupus disease
activity using glomerular proteome arrays. J. Clin. Invest. 115, 3428–3439.
Li, Q.Z., Zhou, J., Wandstrat, A.E., Carr-Johnson, F., Branch, V., Karp, D.R.,
Mohan, C., Wakeland, E.K., and Olsen, N.J. (2007). Protein array autoantibody
profiles for insights into systemic lupus erythematosus and incomplete lupus
syndromes. Clin. Exp. Immunol. 147, 60–70.
Liu, K., and Mohan, C. (2006). What do mouse models teach us about human
SLE? Clin. Immunol. 119, 123–130.
Mohan, C. (2001). Murine lupus genetics: Lessons learned. Curr. Opin.
Rheumatol. 13, 352–360.
Munoz, L.E., Gaipl, U.S., Franz, S., Sheriff, A., Voll, R.E., Kalden, J.R., and
Herrmann, M. (2005). SLE–a disease of clearance deficiency? Rheumatology
(Oxford) 44, 1101–1107.
Nagata, S., and Suda, T. (1995). Fas and Fas ligand: lpr and gld mutations.
Immunol. Today 16, 39–43.
Oliver, P.M., Vass, T., Kappler, J., and Marrack, P. (2006). Loss of the proapop-
totic protein, Bim, breaks B cell anergy. J. Exp. Med. 203, 731–741.
Pasqualetto, V., Vasseur, F., Zavala, F., Schneider, E., and Ezine, S. (2005). Fas
receptor signaling is requisite for B cell differentiation. J. Leukoc. Biol. 78,
1106–1117.
Reap, E.A., Felix, N.J., Wolthusen, P.A., Kotzin, B.L., Cohen, P.L., and
Eisenberg, R.A. (1995). bcl-2 transgenic Lpr mice show profound enhance-
ment of lymphadenopathy. J. Immunol. 155, 5455–5462.
Roy, V., Chang, N.H., Cai, Y., Bonventi, G., and Wither, J. (2005). Aberrant IgM
signaling promotes survival of transitional T1 B cells and prevents tolerance in-
duction in lupus-prone New Zealand black mice. J. Immunol. 175, 7363–7371.
Scatizzi, J.C., Bickel, E., Hutcheson, J., Haines, G.K., 3rd, and Perlman, H.
(2006). Bim deficiency leads to exacerbation and prolongation of joint inflam-
mation in experimental arthritis. Arthritis Rheum. 54, 3182–3193.
Sherriff, A., Gaipl, U.S., Voll, R.E., Kalden, J.R., and Hermann, M. (2004).
Apoptosis and systemic lupus erythmatosus. Rheum. Dis. Clin. North Am.
30, 505–527.
Sobel, E.S., Katagiri, T., Katagiri, K., Morris, S.C., Cohen, P.L., and Eisenberg,
R.A. (1991). An intrinsic B cell defect is required for the production of autoan-
tibodies in the lpr model of murine systemic autoimmunity. J. Exp. Med. 173,
1441–1449.
Strasser, A. (2005). The role of BH3-only proteins in the immune system. Nat.
Rev. Immunol. 5, 189–200.
Strasser, A., O’Connor, L., and Dixit, V.M. (2000). Apoptosis signaling. Annu.
Rev. Biochem. 69, 217–245.
Tan, E.M., Schur, P.H., Carr, R.I., and Kunkel, H.G. (1966). Deoxybonucleic
acid (DNA) and antibodies to DNA in the serum of patients with systemic lupus
erythematosus. J. Clin. Invest. 45, 1732–1740.
Theofilopoulos, A.N., and Dixon, F.J. (1985). Murine models of systemic lupus
erythematosus. Adv. Immunol. 37, 269–390.
Vidal, S., Kono, D.H., and Theofilopoulos, A.N. (1998). Loci predisposing to
autoimmunity in MRL-Fas lpr and C57BL/6-Faslpr mice. J. Clin. Invest. 101,
696–702.
Villunger, A., Marsden, V.S., Zhan, Y., Erlacher, M., Lew, A.M., Bouillet, P.,
Berzins, S., Godfrey, D.I., Heath, W.R., and Strasser, A. (2004). Negative selec-
tion of semimature CD4+8-HSA+ thymocytes requires the BH3-only protein
Bim but is independent of death receptor signaling. Proc. Natl. Acad. Sci.
USA 101, 7052–7057.
Yurasov, S., Wardemann, H., Hammersen, J., Tsuiji, M., Meffre, E., Pascual,
V., and Nussenzweig, M.C. (2005). Defective B cell tolerance checkpoints in
systemic lupus erythematosus. J. Exp. Med. 201, 703–711.
Zhu, J., and Mohan, C. (2007). SLE 1, 2, 3.genetic dissection of lupus. Adv.
Exp. Med. Biol. 601, 85–95.
Accession Numbers
The raw data reported for the microarray analyses in this paper have been de-
posited in the Gene Expression Omnibus database (www.ncbi.nlm.nih.gov/
geo/) with the accession number GSE10325.
Immunity 28, 206–217, February 2008 ª2008 Elsevier Inc. 217
Related Documents











