1 Collagen, cross-linking and advanced glycation endproducts in aging human skeletal muscle Authors: Jacob M. Haus, John A. Carrithers, Scott W. Trappe, and Todd A. Trappe Institutions: Human Performance Laboratory, Ball State University, Muncie, IN Running Title: Intramuscular connective tissue and aging Key Words: hydroxyproline, hydroxylysylpyridinoline, pentosidine, myosin, actin, sarcopenia Address for correspondence: Todd Trappe, Ph. D. Human Performance Laboratory Ball State University Muncie, IN 47306 Ph: 765 285-4456 Fax: 765 285-8596 e-mail: [email protected] Page 1 of 38 Articles in PresS. J Appl Physiol (September 27, 2007). doi:10.1152/japplphysiol.00670.2007 Copyright © 2007 by the American Physiological Society.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Collagen, cross-linking and advanced glycation endproducts in aging human skeletal muscle
Authors: Jacob M. Haus, John A. Carrithers, Scott W. Trappe, and Todd A. Trappe
Institutions: Human Performance Laboratory, Ball State University, Muncie, IN
Running Title: Intramuscular connective tissue and aging
Key Words: hydroxyproline, hydroxylysylpyridinoline, pentosidine, myosin, actin, sarcopenia
Address for correspondence: Todd Trappe, Ph. D. Human Performance Laboratory Ball State University Muncie, IN 47306 Ph: 765 285-4456 Fax: 765 285-8596 e-mail: [email protected]
Page 1 of 38Articles in PresS. J Appl Physiol (September 27, 2007). doi:10.1152/japplphysiol.00670.2007
Copyright © 2007 by the American Physiological Society.

2
ABSTRACT
We examined intramuscular endomysial collagen, cross-linking and advanced glycation
endproducts, as well as the general and contractile protein concentration of 20 young
(25±3y) and 22 old (78±6y, range: 70-93y) sedentary men and women to better
understand the underlying basis of changes in skeletal muscle mass and function that
occur with aging. The old individuals had an impaired ability (increased time) (p<0.05)
to climb stairs (80%), rise from a chair (56%), and walk (44%), as well as lower (p<0.05)
quadriceps muscle volume (-29%), muscle strength (-35%), muscle power (-48%), and
strength (-17%) and power (-33%) normalized to muscle size. Vastus lateralis muscle
biopsies revealed that intramuscular endomysial collagen (Young: 9.6±1.1, Old:
10.2±1.2 µg•mg muscle wet wt-1) and collagen cross-linking (hydroxylysylpyridinoline,
HP) (Young: 395±65, Old: 351±45 mmol HP•mol collagen-1) were unchanged (p<0.05)
with aging. The advanced glycation endproduct, pentosidine, was increased (p<0.05)
by ~200% (Young: 5.2±1.3, Old: 15.9±4.5 mmol pentosidine•mol collagen-1) with aging.
While myofibrillar protein concentration was lower (-5%, p<0.05), the concentration of
the main contractile proteins myosin and actin were unchanged (p>0.05) with aging.
These data suggest that the synthesis and degradation of proteins responsible for the
generation (myosin and actin) and transfer (collagen and pyridinoline cross-links) of
muscle force are tightly regulated in aging muscle. Glycation related cross-linking of
intramuscular connective tissue may contribute to altered muscle force transmission
and muscle function with healthy aging.
Page 2 of 38

3
INTRODUCTION
A critical component in the transfer of force from the contractile units of the
muscle out to the tendon and subsequent bone is the connective tissue scaffold that
surrounds individual muscle fibers, muscle bundles, and the whole muscle (20, 23, 37).
This skeletal muscle connective tissue network is primarily comprised of collagen fibers
and biochemical linkages within and between these fibers, which provide strength and
stability. Numerous data from animals have shown that collagen concentration of intact
skeletal muscle increases with aging (1, 12, 17, 24, 25, 34, 55), and a mature structural
characteristic of collagen known as pyridinium cross-linking has also been shown to be
increased in aged animal muscle (17, 18, 36, 40, 55). The pyridinium cross-links are
enzymatically added to collagen fibrils upon collagen maturity and replace an immature
cross-link molecule. These mature pyridinium cross-links provide strength to the
collagen molecule by linking the collagen fibrils in the characteristic one-third staggered
arrangement.
Separate from the enzymatically derived, mature cross-links are an additional
form of biochemical linkage found on connective tissue known as advanced glycation
endproducts (AGE’s). Accumulation of these compounds are the result of prolonged
exposure to monosaccharides, in which a spontaneous non-enzymatic bond is formed
between a reducing sugar and a protein residue. AGE’s have been linked to disease
and aging processes (2, 10, 35, 38, 41) and have been demonstrated to be strongly
correlated to increases in plasma glucose concentrations and chronological age.
Evidence from humans and animals demonstrates the formation of these compounds in
Page 3 of 38

4
vascular tissues and lens proteins with diabetes and has shown a progressive
accumulation in skin and cartilage with aging.
Skeletal muscle studied in aging animals demonstrates increases in collagen
concentration, HP cross-linking, and AGE’s, all of which have been associated with
increased muscle stiffness and reduced whole muscle function (1, 12, 17, 26, 52).
Increases in tissue stiffness have been suggested to play a role in reduced whole
muscle function with aging. However, almost no data exist from humans regarding
intramuscular connective tissue changes with aging (3) or the extent of collagen cross-
linking or the development of AGE’s in healthy human skeletal muscle (14, 45). In
aging humans, muscle strength and power is lost to a greater extent than the loss of
muscle mass (i.e., whole muscle specific force and specific power), suggesting that
factors other than the loss of muscle mass are contributing to the decline in whole
muscle function (47). We have shown that single muscle fiber strength and power
normalized to fiber size [i.e., myosin heavy chain (MHC) I and IIa myofiber specific force
and specific power] are not compromised with aging (47). These findings lead to the
possibility that the transfer of force and power from the single fiber out to the whole
muscle and subsequent tendon and bone may be impaired in aging skeletal muscle.
The intent of the current investigation was to expand on our previous findings of
the role of single muscle fiber function in the decline of whole muscle function in aging
through measurements of the intramuscular connective tissue network. Specifically, we
measured the concentration of collagen (endomysium), collagen cross-linking
(hydroxylysylpyridinoline, HP), and AGE’s (pentosidine) in skeletal muscle biopsy
samples taken from a well-characterized group of young and old sedentary women and
Page 4 of 38

5
men. We hypothesized that the concentrations of endomysial collagen, HP cross-links
and pentosidine would be elevated in old individuals compared to young. An additional
aim was to measure the concentration of the general muscle protein fractions (mixed,
sarcoplasmic, and myofibrillar) commonly studied in metabolic investigations of
sarcopenia (5, 19, 42, 50, 51, 54), as well as the concentration of the two main
contractile proteins that are responsible for force generation in skeletal muscle, myosin
and actin. These measurements extend our previous investigation from a subset of the
42 individuals studied for the present investigation (47).
Page 5 of 38

6
METHODS
Subjects
Twenty young (10 men, 10 women) and 22 old (10 men, 12 women) individuals
were included in this investigation (Table 1) following a physical exam, which included
blood and urine analyses, electrocardiogram (older subjects), and an interview to
document their life history of physical activity. Subjects were excluded if they had any
acute or chronic illness, cardiac, pulmonary, liver, or kidney abnormalities, uncontrolled
hypertension, insulin- or noninsulin-dependent diabetes, abnormal blood or urine
chemistries, arthritis, a history of neuromuscular problems, or if they smoked tobacco.
Women taking oral contraceptives or hormone replacement therapy were included. It
was our intent to only include life-long sedentary healthy older and younger individuals;
therefore, we excluded individuals that, by self-report, had ever completed any formal
exercise programs or physical activity outside of their activities of daily living. All
procedures, risks and benefits associated with the experimental testing were explained
to the subjects prior to signing a consent form adhering to the guidelines of the
Institutional Review Boards of the participating institutions. The study was conducted in
accordance with the Declaration of Helsinki.
Experimental Design
Subjects who qualified for the study completed six different experimental sessions
over a two to three week period, described in detail below. The first session involved
magnetic resonance imaging (MRI) for thigh muscle size. The second session involved
the measurement of body composition via whole body air-displacement plethysmography
Page 6 of 38

7
(Life Measurement Instruments, Concord, CA) and a muscle biopsy of the vastus
lateralis. The remaining four visits involved the measurement of functional ability and
muscle function. The first two of these four visits were used for familiarization to the
tests, while the third was used for data analysis (i.e., the data presented here) and the
fourth for the determination of the reliability of these measurements. Each of these four
visits was separated by at least two days. The muscle biopsy and the measurements of
muscle size and strength were completed on the dominant leg. Dominance was
determined by asking the subject which leg they would use to kick a ball.
Magnetic Resonance Imaging
Following one hour of supine rest to control for the influence of postural related
fluid shifts on muscle size (8), MRI were obtained for each subject as we have previously
described (48). Subjects were supine and their heels were fixed on a non-metallic
support to control joint and scan angle and to minimize compression of the legs against
each other and the MRI gurney. Imaging was completed in a 1.5T GE Signa scanner
(General Electric, Milwaukee, WI) to determine the volume and cross sectional area
(CSA) of the total quadriceps femoris, rectus femoris (RF), vastus lateralis (VL), vastus
intermedius (VI), and vastus medialis (VM). Following an initial scout scan, interleaved
transaxial images of 1 cm thick (TR/TE = 2000/9.0 ms, field of view 48 cm, 256 X 256
matrix) were taken from the top of the greater trochanter of the femur to the articular
surface of the tibia. MR images were transferred electronically from the scanner to a
personal computer (Macintosh Power PC) and analyzed with NIH Image software
(version 1.60) using manual planimetry. Analyses of the MR images began with the first
Page 7 of 38

8
proximal slice not containing gluteal muscle and continued distally to the last slice
containing RF. The average CSA (cm2) was taken as the average of all the analyzed
slices for an individual muscle and determined for the RF, VL, VI, and VM and summed
for the total quadriceps femoris. Muscle volume (cm3) was calculated by multiplying the
CSA of each individual muscle by the slice thickness (1 cm) for all analyzed images of
the RF, VL, VI, VM, and summed for the total quadriceps femoris. All measurements
were made by the same investigator.
Muscle Biopsy
Each subject underwent a muscle biopsy (9) of the vastus lateralis for the
measurement of intramuscular connective tissue, general and contractile proteins and
MHC distribution. Following the biopsy, excess blood, visible fat and connective tissue
were removed and portions of the muscle to be used for the aforementioned analyses
were immediately frozen and stored in liquid nitrogen (-190°C) until analysis. An
additional portion of the muscle was used to study the single muscle fiber contractile
characteristics of a subset (n=24) of the subjects and the results of those experiments
are presented elsewhere (47).
Intramuscular Connective Tissue Quantification
Collagen concentration. Skeletal muscle collagen concentration was determined
via measurement of the collagen specific amino acid, hydroxyproline (HYP). The
concentration of HYP was quantified by high-performance liquid chromatography and
fluorometric detection (1100 Series, Agilent Technologies, Wilmington, DE, USA) via the
Page 8 of 38

9
precolumn derivatization method described by Hutson et al. (21) with modifications for
human skeletal muscle. Muscle samples (~10 mg) were weighed at -35°C on a
precision microbalance (AD-2Z Autobalance, Perkin-Elmer, Wellesley, MA, USA) and
hydrolyzed in 1 ml of 6 M HCl at 110°C for 30 h. In addition, a sarcosine internal
standard (Sigma #S7672, St. Louis, MO) prepared in water was added to each vial.
The hydrolyzates were allowed to cool to room temperature and neutralized with 6 M
NaOH. Hydroxyproline standards (Sigma #56250) of 1, 10, 25, 50, 75, 100, and 125
µM were prepared along with the 2 mM sarcosine internal standard, and were used to
generate a standard curve.
Derivatization was performed according to the procedures described by Hutson
et al. (21) and separation was achieved through a XTerra RP 18, 5 µm, 250 mm x 4.6
mm column (Waters, Milford, MA, USA) using an isocratic mobile phase of 65% acetic
acid (3% glacial acetic acid, sodium acetate buffered to pH 4.3): 35% acetonitrile at a
1.0 ml•min-1 flow rate. Peaks were monitored at 260/316 nm (Ex/Em) with a gain of 8
and integrated with chromatography software (ChemStation, Agilent Technologies)
(Figure 1). HYP signals were normalized to the internal standard for each injection and
HYP concentration was determined from standard curves of HYP (correlation coefficient
of 0.983). All samples and standards were run in triplicate with a mean coefficient of
variation of 0.32% and 0.69% respectively. Collagen concentration was calculated from
the HYP concentration assuming collagen weighs 7.5 times the measured HYP weight
and the molecular weight of collagen is 300,000 (13, 14). Samples were normalized to
muscle wet weight and data are expressed as µg collagen•mg wet weight muscle-1.
Page 9 of 38

10
Collagen cross-link concentration. The extent of collagen cross-linking was
determined by measuring the amount of the pyridinium cross-link
hydroxylysylpyridinoline (HP) through HPLC as previously described (6) with
modifications for human skeletal muscle. Muscle samples (~10 mg) were weighed at -
35°C and immediately placed into screw cap vials containing 1 ml of 6 M HCl after which
the samples were hydrolyzed at 110°C for 20 h. A 500 µl aliquot of the hydrolyzate was
then added to a sample cocktail consisting of a 4:1:1 ratio of butanol: cellulose slurry:
acetic acid containing acetylated pyridinoline (Quidel Corp. #8006, San Diego, CA) as an
internal standard.
Samples for collagen cross-links were extracted through CF1 cellulose partition
chromatography as described previously (43). The cross-links were eluted with water
and evaporated to dryness in a Savant Speedvac sample concentrator. Samples were
reconstituted in 0.5% heptafluorobutyric acid (HFBA) in 10% ACN and then filtered
(0.22 µM nylon filter) through centrifuge spin columns (Costar Spin X) at 10,000 x g for
10 min to remove residual cellulose particles.
HP standards (Quidel Corp. # 8004) of 5, 10, 25, 50, 75 and 100 pmol were
prepared in 0.5% HFBA/10% ACN buffer with acetylated pyridinoline added as an
internal standard. Standards and samples were then injected into the HPLC (Agilent
Technologies) and eluted as previously described with modifications (6). The column
(XTerra RP 18, 5 µm, 250 mm x 4.6 mm, Waters) was equilibrated with 0.13% HFBA in
22% methanol and cross-links were eluted with a 1.0 ml • min-1 flow rate from 0 - 30
min. A washing step followed immediately using 0.1% HFBA in 75% ACN for 10 min.
Page 10 of 38

11
Fluorescence of HP was monitored at 295/395 nm (Ex/Em) and integrated with
chromatography software (ChemStation, Agilent Technologies) (Figure 1).
Standard and sample signals were normalized to the internal standard signal and
sample HP recovery was then calculated as the percentage of sample internal standard
signal relative to the mean internal standard signal obtained from the known HP
standard curve. Mean recovery of skeletal muscle HP was ~ 80% after CF1 purification.
The concentration of skeletal muscle HP was determined from the standard curve and
adjusted for recovery. Cross-link concentration of each tissue sample was expressed
as mmol of HP per mol collagen assuming the molecular weight of collagen is 300,000
(7).
Advanced glycation endproduct (AGE) concentration. The AGE, pentosidine, was
measured in human skeletal muscle through HPLC and fluorometric detection (Agilent
Technologies). Muscle samples (~10 mg) were weighed at -35°C and immediately
placed into screw cap vials containing 1 ml of 6 M HCl. Samples were then hydrolyzed
at 110°C for 20 h. Following hydrolysis, samples were evaporated to dryness in a
Savant Speedvac sample concentrator followed by reconstitution with 0.5% HFBA/10%
ACN buffer containing 300 pmol of pyridoxine internal standard (Sigma #P5669). After
reconstitution the samples were filtered (0.22 µM nylon filter) through centrifuge spin
columns (Costar Spin X) at 10,000 x g for 10 min. Recovery of internal standard after
spin column filtration was tested and found to be 100%. Pentosidine standards of 0.05,
0.1, 0.25, 0.5, 0.75, 1.0, 2.5, and 5 pmol were prepared from a known concentration of
purified pentosidine obtained from the International Malliard Reaction Society (Vincent
Page 11 of 38

12
Monnier, CWRU, Cleveland, OH) in the same buffer as the samples (0.5%HFBA/10%
ACN), which included 300 pmol of pyridoxine.
The column (XTerra RP 18, 5 µm, 250 mm x 4.6 mm, Waters) was equilibrated
with 0.05% HFBA in water and the standards and samples were subsequently injected
into the HPLC (Agilent Technologies). Pyridoxine was eluted at ~ 10 min with an
inverse gradient of HFBA (0.05% to 0%; 0-20 min) and methanol (0% - 2.5%; 0-20 min)
at 1.0 ml • min-1 flow rate. Pentosidine was eluted at ~ 22 min with 2.5% methanol from
20-30 min. The elution gradients were followed by a washing step of 100% acetonitrile.
Fluorescence was monitored at 295/395 at a gain of 12 from 0-17 min for detection of
pyridoxine and 328/378 at a gain of 15 from 17-25 min for the detection of pentosidine
(Figure 1). Peaks were integrated with chromatography software (ChemStation, Agilent
Technologies) and standard and sample signals were reduced with and without
normalization to pyridoxine internal standard. Normalization had no effect on the
concentrations of pentosidine compared to raw signal when calculated from the
standard curve. Skeletal muscle pentosidine concentration was expressed as mmol per
mol collagen, assuming the molecular weight of collagen is 300,000 (7).
General Protein Quantification
Mixed, sarcoplasmic, and myofibrillar muscle protein concentration. For each
subject, a piece of muscle weighing ~10 mg was divided and weighed on a precision
microbalance (Cahn 35, Orion Research, Beverly, MA) at -35°C. Each sample was
homogenized in 40 volumes of cold homogenizing buffer (250 mM sucrose, 100 mM
potassium chloride, 20 mM imidazole, and 5 mM EDTA; pH 6.8) in a ground-glass
Page 12 of 38

13
homogenizer (Radnoti Glass Technology, Monrovia, CA) (11). Samples were then
centrifuged at 20,000 x g for 30 min at 4°C. The supernatant was taken as the
sarcoplasmic protein fraction and the pellet was resuspended in 40 volumes of cold
homogenizing buffer and taken as the myofibrillar protein fraction (22, 44). Aliquots of
the homogenate (total mixed protein), sarcoplasmic, and myofibrillar protein fractions
were measured for protein concentration using the bicinchoninic acid (BCA) assay
(Sigma) with bovine serum albumin used as the protein standard. The amount of protein
in each of the three fractions was normalized to muscle wet weight and expressed as µg
protein•mg wet weight muscle-1.
As collagen is contained in the mixed protein and myofibrillar protein (i.e. collagen
pellets following centrifugation) (3, 32, 33), follow-up BCA protein assays with pure
collagen (Sigma #C9791) were completed to confirm that collagen contributes to the
overall absorbance of the muscle measurements (Figure 2). These measurements were
completed because it has been suggested that collagen does not contribute to some
assays of mixed protein in skeletal muscle (39, 53).
Contractile Protein Quantification
Myosin and actin concentration. Myosin (myosin heavy chain, MHC) and actin
concentrations were determined by quantitative gel electrophoresis as previously
described (22, 49). Aliquots of the myofibrillar protein fraction were diluted with sodium
dodecyl sulphate (SDS) buffer (2% SDS, 125 mM Tris HCl (pH 6.8), 12.5% glycerol, 5%
2-mercaptoethanol, 0.005% bromophenol blue) and heated at 60°C for 4 min.
Myofibrillar protein (800 ng) was separated by SDS-polyacrylamide gel electrophoresis
Page 13 of 38

14
(PAGE) (28). MHC was resolved with a 4% stacking gel and 10% separating gel. Actin
was resolved with a 4% stacking gel and a 6-12% gradient separating gel that were
allowed to polymerize overnight. For MHC and actin, electrophoresis was performed at
a constant current of 20 mA per gel in the stacking gel, and 25 mA per gel in the
separating gel with a Tris-glycine electrode buffer at 4°C (Hoeffer SE 600, Amersham
Pharmacia Biotech, Piscataway, NJ).
The separating gels were silver stained (15), digitally photographed (ChemImager
5500, Alpha Innotech, San Leandro, CA), and densitometry was completed using NIH
Image software (Ver. 1.60) (47). Each gel was loaded with five standards of either MHC
or actin (Sigma #’s M7659, A2522), a molecular weight standard (Pierce, Rockford, IL),
and subject samples. All standards and samples were loaded in duplicate and each gel
contained samples from young and old men and women. An average of the duplicate
densities was taken to represent each standard and sample. All measurements were
made in blinded fashion by the same investigator. Unknown sample amounts of MHC
and actin were determined from regression analysis of the standard curves on each gel.
Correlation coefficients were 0.98-1.00 for both MHC and actin. The amount of myosin
or actin was normalized to muscle wet weight and expressed as µg•mg wet weight
muscle-1.
MHC Isoform Distribution
MHC isoform composition of each of the myofibrillar protein samples quantified for
myosin and actin protein concentrations was determined in triplicate from a SDS buffer-
diluted aliquot of the myofibrillar protein fraction heated to 100°C for 5 min. MHC
Page 14 of 38

15
isoforms were separated with SDS-PAGE with a 3.5% stacking gel and 5% separating
gel. Electrophoresis was performed at 150 V for ~15 h with a Tris-glycine electrode
buffer at 4°C (Hoeffer SE 600, Amersham Pharmacia Biotech, Piscataway, NJ). The
separating gels were silver stained (15), digitally photographed (ChemImager 5500,
Alpha Innotech, San Leandro, CA), and densitometry was completed to determine the
percent contribution of each isoform of the total (100%). The average density of each
isoform from the three lanes loaded was taken as the MHC distribution for that sample.
Functional Ability
The ability to perform activities of daily living was determined with three
functional tests: walk, chair stand, and stair climb. Each of the functional tests was
completed three times during each session, with the average of the three tests
recorded. Timing of the functional tests was accomplished using retroreflective sensors
(Banner Engineering Corp., Minneapolis, MN) interfaced to a digital timer and display
(Veeder-Root, Danaher Controls, Gurnee, IL). The sensors were placed such that the
forehead (chair stand or walk tests) or foot (stair climb test) would break the light beams
to start and stop timing.
Walk test. Subjects walked (i.e., one foot in contact with the ground at all times)
a distance of 6.1 m as quickly as possible. Subjects started three steps behind the first
photo eye and continued through the second photo eye. The coefficient of variation for
the walk test was 3.8 ± 0.5% with no group, age, or gender differences.
Page 15 of 38

16
Chair stand test. Subjects stood from a seated position in a metal chair as fast
as possible without support (i.e., via chair or thighs). The coefficient of variation for the
chair stand was 6.9 ± 0.9% with no group, age, or gender differences.
Stair climb test. Subjects started two steps before the first stair and climbed a
flight of 10 stairs (3.3m vertical distance) as quickly as possible without skipping a stair.
The coefficient of variation for the stair climb was 3.6 ± 0.5% with no group, age, or
gender differences.
Muscle Function
Following the functional ability measurements, subjects completed light
stretching of the legs and a five-minute warm-up on a bicycle ergometer (Monark
Exercise AB, Sweden). Muscle function testing was performed on a Cybex Norm
dynamometer (Lumenex, Ronkonkoma, NY) and associated equipment fabricated by
the investigators. Measurements were made of in-vivo maximal shortening velocity
(Vmax), as well as static and dynamic muscle strength and power of the quadriceps
femoris muscle. Adequate rest periods separated all of the tests within each testing
session.
Maximal shortening velocity. Vmax was determined by having each subject
extend their lower leg from a metal bar placed behind their heel to establish a starting
position at 90° of knee flexion through a 90° arc and into a heavily padded bar as fast as
possible. Fiber optic sensors (Banner Engineering Corp., Minneapolis, MN) interfaced
with a computer timer (LabVIEW, National Instruments, Inc., Austin, TX) were
positioned at 60° and 30° of knee flexion to measure the time to move through the 30°.
Page 16 of 38

17
Subjects were allowed three warm-up attempts followed by three timed leg extensions,
with the fastest time of the three recorded. The coefficient of variation was 3.5 ±0.5%
with no group, age or gender differences.
Static and dynamic force and power. Following completion of the Vmax
measurements, maximal isometric (Po), and concentric muscle forces were determined.
Po was measured at 60° of knee flexion and subjects were allowed one warm-up trial
followed by two maximal contractions, with the greater value taken as Po. A one-minute
rest period separated the two maximal contractions. Coefficient of variation for the
measurement of Po averaged 6.2%, with no age, group, or gender difference.
Maximal isokinetic concentric force was determined at 60, 120, 180, 240, 300,
360, 420, 480, and 500°•sec-1. At each velocity the subjects were allowed three warm-
up attempts immediately followed by three maximal concentric contractions, with the
highest force attained for each velocity used for analysis. A two-minute rest period was
given between velocities. Some subjects were not able to produce force at the higher
velocities and/or their measured Vmax was lower than some of the higher measurement
velocities. Coefficient of variation for all concentric velocities, 60 to 500°•sec-1,
averaged 5.3, 3.9, 4.2, 5.5, 5.5, 6.4, 5.8, 7.6, 8.2%, respectively, with no age, group, or
gender difference.
Peak power was calculated for each subject by curve fitting the data of power
(force x velocity) vs. the force produced at each velocity represented as a percentage of
maximal isometric force (%Po). Data to develop the power curve was used if the
measurement velocity was below the measured Vmax for that subject and force
production at two consecutive velocities differed by greater than 4 Nm.
Page 17 of 38

18
Statistics
A two-way (age and gender) analysis of variance (ANOVA) was used to compare all
variables. No significant interactions were detected; therefore, post-hoc comparisons
were not completed and the data were collapsed and presented as young and old.
Significance was accepted at p<0.05. Data are presented as mean ± S.E.
Page 18 of 38

19
RESULTS
Intramuscular Connective Tissue
Collagen concentration (Young: 9.6±1.1, Old: 10.2±1.2 µg•mg muscle wet wt-1)
and pyridinoline cross-links (Young: 395±65, Old: 351±45 mmol HP•mol collagen-1)
were unchanged (p>0.05) with aging, while AGE’s were increased (p<0.05) by ~200%
(Young: 5.2±1.3, Old: 15.9±4.5 mmol pentosidine•mol collagen-1) with aging (Figure 3).
General and Contractile Proteins
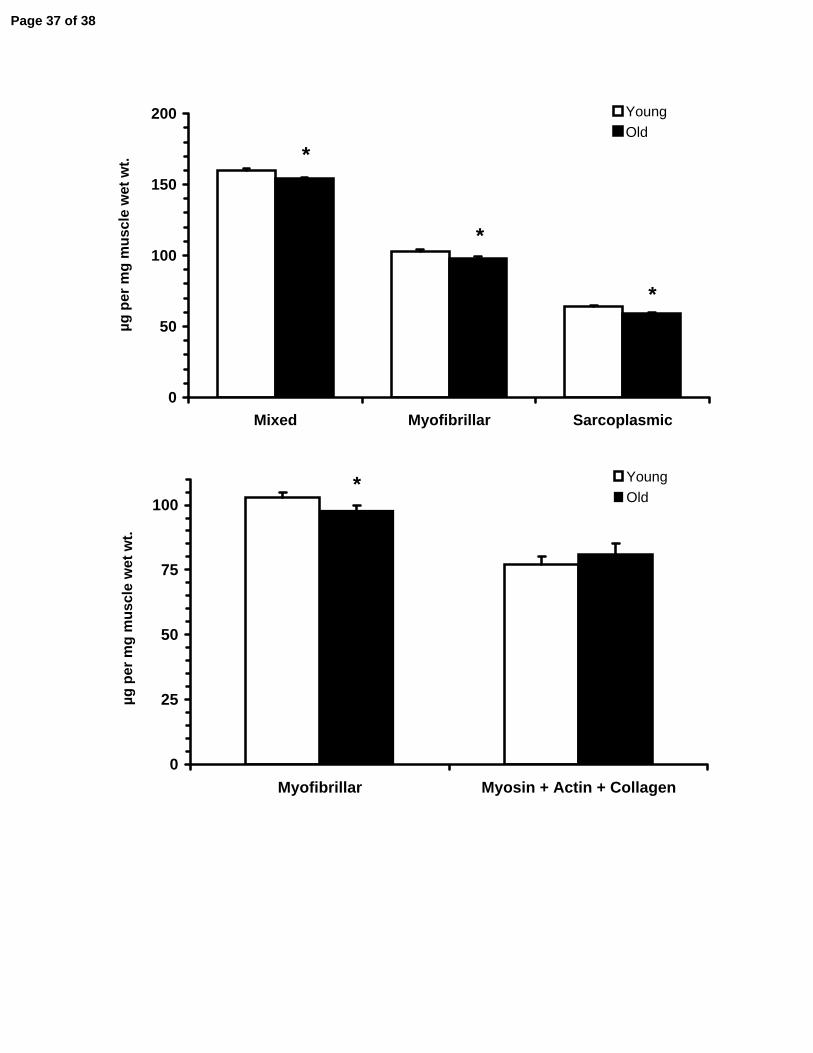
All three general skeletal muscle protein pools were reduced (p<0.05) with aging
(Figure 4). Mixed protein concentration was reduced by 4% (Young: 160±3, Old: 154±1
µg•mg muscle wet wt-1), myofibrillar by 5% (Young: 103±2, Old: 98±2 µg•mg muscle wet
wt-1), and sarcoplasmic by 8% (Young: 64±1, Old: 59±1 µg•mg muscle wet wt-1). The
two main contractile proteins myosin (Young: 46±3, Old: 54±4 µg•mg muscle wet wt-1)
and actin (Young: 21±1, Old: 17±1 µg•mg muscle wet wt-1) were not changed (p>0.05)
with aging. The proportion of MHC I in the muscle increased (P<0.05) with aging, while
MHC IIa and IIx were reduced (p<0.05) (Table 2).
Whole Muscle Characteristics
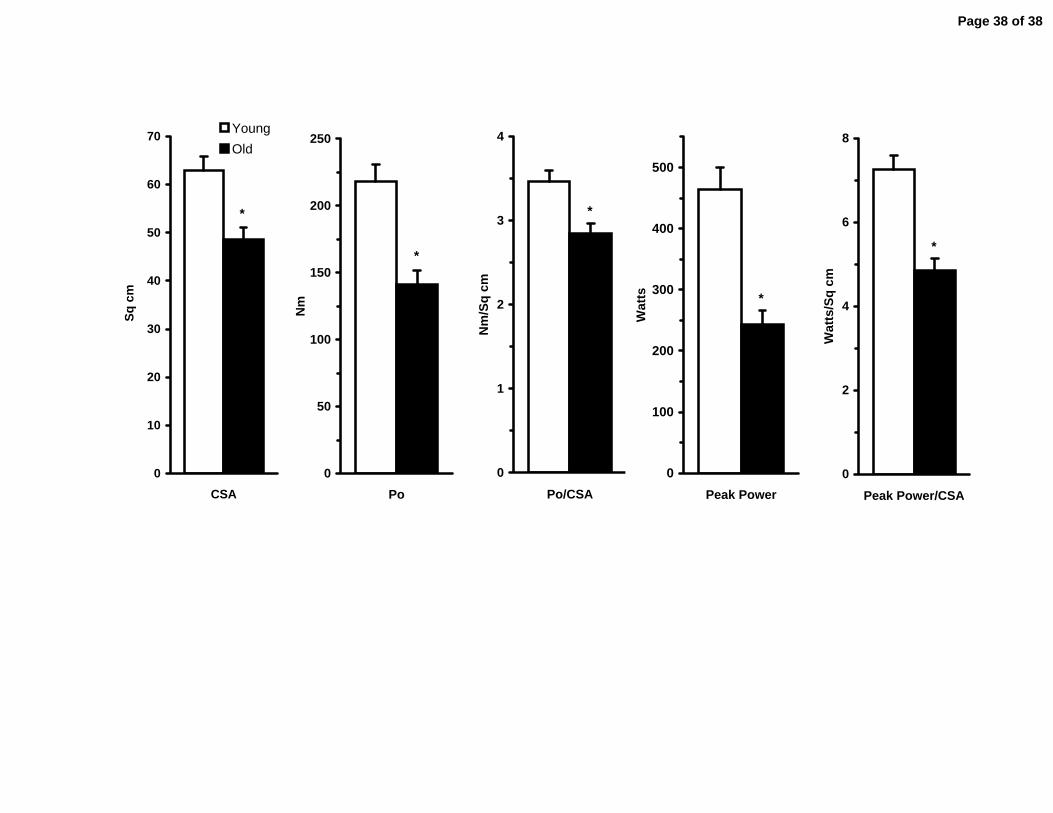
Quadriceps muscle volume was reduced (p<0.05) by 29% with aging (Table 2).
Quadriceps muscle CSA followed this same trend and was reduced (p<0.05) by 23%
(Young: 62.9±2.9, Old: 48.7±2.3 cm2) (Figure 5). Maximal isometric force (Po) of the
quadriceps was reduced (p<0.05) by 35% (Young: 218±13, Old: 141±11 Nm) and Po
normalized to muscle size was reduced (p<0.05) by 17% (Young: 3.46±0.14, Old:-
Page 19 of 38

20
285±0.11 Nm/cm2) (Figure 5). Similarly, peak power of the quadriceps was reduced
(p<0.05) by 48% (Young: 464±36, Old: 243±23 W) and peak power normalized to
muscle CSA was reduced (p<0.05) by 33% (Young: 7.25±0.33, Old: 4.85±0.29 W/cm2)
(Figure 5). Stair climb, chair rise, and walk time were increased by 45 to 80%, and stair
climbing power was reduced by 40% (Table 2) with aging. Quadriceps Vmax was
reduced by 23% (Table 2).
Page 20 of 38

21
DISCUSSION
The results of the present study show that endomysial collagen concentration
and enzymatically mediated collagen cross-linking are tightly regulated with aging as
evidenced by the similar concentrations in young and old individuals. However, non-
enzymatically regulated advanced glycation end-product cross-linking is significantly
increased in muscle from healthy sedentary elderly individuals. These results suggest
that the formation of AGE’s over the life span of an individual may contribute to
increased muscle connective tissue protein stiffness and thus contribute to impaired
muscle function in the elderly.
Contrary to our hypotheses, aging had no effect on the intramuscular collagen
concentration. In addition, there was no sex-specific influence on intramuscular
collagen levels. Several recent studies have shown the collagen fraction of skeletal
muscle is much more dynamic that previously thought (3, 32, 33), and the synthesis rate
of intramuscular collagen is elevated in older men (3). Based on these findings and the
collagen concentration data from the current study, collagen breakdown would also
have to be elevated to a similar degree to prevent the accretion of collagen within the
muscle. The discrepancy between the current study and previous animal findings may
be related to the specific portions of the muscle connective tissue studied. Data
obtained from the muscle biopsy technique mainly reflect the endomysial collagen
layers and not the peri- or epimysial layers of connective tissue that have a different
composition of collagen types and roles in the transfer of mechanical forces (20, 23,
37). This is contrary to the animal data which utilizes excised whole muscles for the
analysis of connective tissue. Thus, the potential that biochemical changes are
Page 21 of 38

22
occurring in the outer connective tissue layers with aging cannot be excluded. Finally, it
should also be considered that while intramuscular collagen concentration did not
change with aging, it is possible that the type or isoforms of collagen changed. Aging
animal muscle has been shown to alter the collagen isoforms, either by increasing Type
IV collagen (27) or decreasing Type III collagen (16). In these cases, the collagen may
be changing morphology to accommodate a functional need.
To our knowledge, the current investigation is the first to examine the effects of
aging on human skeletal muscle collagen cross-linking. Limited reports exist from post-
mortem analysis of human skeletal muscle that shows the primary mature intramuscular
pyridinium cross-link is HP and lysylpyridinoline (LP) is found only in trace amounts (14,
45). As a result our focus in healthy adult skeletal muscle was limited to that of the
abundant, mature HP species. The existing literature of collagen cross-linking in aging
animals has focused mainly on HP and demonstrates that aging results in a significant
increase in HP with concomitant increases in muscle stiffness (17, 18). The lack of
change seen in aging human muscle may indicate that the normal turnover of skeletal
muscle collagen is robust enough to also turnover mature endomysial connective tissue
cross-links, and these cross-links may not contribute to the reduced muscle function
that occurs with aging.
In contrast to HP collagen cross-linking, the formation of AGE cross-links is not
enzymatically regulated but dictated by the presence of a reducing sugar, the
appropriate protein side chain and oxygen (4). The differences seen in AGE
concentration between young and old in the current investigation are most likely related
to the temporal component of aging. That is, the more time that protein residues have
Page 22 of 38

23
to come into contact with glucose, there is a greater chance that AGE’s will form. It
should be noted that all the subjects demonstrated normoglycemia and were absent of
overt disease, thus the increased AGE’s noted in the old are reflective of factors other
than altered glucose metabolism, such as that seen in diabetic individuals. The
accumulation of AGE’s in collagenous tissues has been shown to negatively affect
function such as stiffening of the blood vessel walls and kidney structures (4, 35). It is
likely that the 200% increase in pentosidine seen in the older individuals of the current
study influenced tissue stiffness and the passive viscoelastic properties of the muscle
and thus contributed to declines in muscle function. In this context, it should be noted
that pentosidine is commonly used as a surrogate for the many other AGE’s cross-links
(4), and the large increase seen herein is reflective of the accumulation of all possible
AGE’s. The accumulation of AGE’s in the skeletal muscle of the aging women and men
in the current study may be due to the lack of a regular and robust tissue turnover
stimulus such as exercise (19, 31, 33). These concepts warrant further study of AGE
formation in aging human skeletal muscle and the influence of both acute and chronic
exercise.
As the concentrations of mixed, myofibrillar, and sarcoplasmic proteins found in
skeletal muscle are a result of the net turnover (i.e., the sum of protein synthesis and
degradation) (5, 46, 50, 51, 54) and were reduced in the old men and women, there
appears to be a net imbalance between these two processes in aging skeletal muscle.
In addition, the reduction in the myofibrillar protein concentration, in light of the
maintenance of myosin, actin and collagen concentrations (Figure 5), suggests that
other proteins in the myofibrillar apparatus (i.e., titin, nebulin, c-protein, m-protein (37))
Page 23 of 38

24
are disproportionately lost with aging. Collectively, the current and previous (47) data
collected on the individuals in this study suggest a primary contributor to sarcopenia is
the loss of MHC II fiber number (30) (Table 2). To this end, MHC IIa muscle fibers
produce 5-6 times the normalized power as a MHC I fiber (~9 vs. ~1.5 watts•liter-1) (47).
Thus, an aging individual could afford to lose 5-6 MHC I fibers for every one MHC IIa
fiber lost. Having a higher proportion of MHC I would lend itself to a slower contracting
whole muscle, which is supported by the 23% slower in vivo maximal contraction
velocity in the older subjects. Of course, we cannot rule out the role of a change in the
neural control of muscle with aging (29).
This study and the data reported here are the first to comprehensively examine
the intramuscular connective tissue network in aging men and women. These data
suggest that despite large changes in muscle mass, the concentrations of the two main
contractile proteins, myosin and actin, the protein responsible for the transfer of force
out to the whole muscle, collagen, and the enzymatically regulated cross-linking of
collagen are tightly regulated in aging human skeletal muscle. It does appear that non-
enzymatic addition of advanced glycation endproducts to the intramuscular connective
tissue network may play a role in the reduction of muscle and physical function with
aging.
Page 24 of 38

25
REFERENCES
1. Alnaqeeb MA, Al Zaid NS, and Goldspink G. Connective tissue changes and physical properties of developing and ageing skeletal muscle. J Anat 139 ( Pt 4): 677-689, 1984.
2. Alt N, Carson JA, Alderson NL, Wang Y, Nagai R, Henle T, Thorpe SR, and Baynes JW. Chemical modification of muscle protein in diabetes. Arch Biochem Biophys 425: 200-206, 2004.
3. Babraj JA, Cuthbertson DJ, Smith K, Langberg H, Miller B, Krogsgaard MR, Kjaer M, and Rennie MJ. Collagen synthesis in human musculoskeletal tissues and skin. Am J Physiol Endocrinol Metab 289: E864-869, 2005.
4. Bailey AJ. Molecular mechanisms of ageing in connective tissues. Mech Ageing Dev 122: 735-755, 2001.
5. Balagopal P, Rooyackers OE, Adey DB, Ades PA, and Nair KS. Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans. The American journal of physiology 273: E790-800, 1997.
6. Bank RA, Beekman B, Verzijl N, de Roos JA, Sakkee AN, and TeKoppele JM. Sensitive fluorimetric quantitation of pyridinium and pentosidine crosslinks in biological samples in a single high-performance liquid chromatographic run. JChromatogr B Biomed Sci Appl 703: 37-44, 1997.
7. Bank RA, TeKoppele JM, Oostingh G, Hazleman BL, and Riley GP.Lysylhydroxylation and non-reducible crosslinking of human supraspinatus tendon collagen: changes with age and in chronic rotator cuff tendinitis. Ann Rheum Dis 58: 35-41, 1999.
8. Berg HE, Tedner B, and Tesch PA. Changes in lower limb muscle cross-sectional area and tissue fluid volume after transition from standing to supine. Acta Physiol Scand 148: 379-385, 1993.
9. Bergstrom J. Muscle electrolytes in man. Scandinavian Journal of Clinical Laboratory Investigation 14: 7-110, 1962.
10. Brown SM, Smith DM, Alt N, Thorpe SR, and Baynes JW. Tissue-specific variation in glycation of proteins in diabetes: evidence for a functional role of amadoriase enzymes. Ann N Y Acad Sci 1043: 817-823, 2005.
11. Chi M-Y, Hintz CS, Coyle EF, Martin WH, Ivy JL, Nemeth PH, Holloszy JO, and Lowry OH. Effects of detraining on enzymes of energy metabolism in individual human muscle fibers. American Journal of Physiology 244: C276-C287, 1983.
Page 25 of 38

26
12. Ducomps C, Mauriege P, Darche B, Combes S, Lebas F, and Doutreloux JP.Effects of jump training on passive mechanical stress and stiffness in rabbit skeletal muscle: role of collagen. Acta Physiol Scand 178: 215-224, 2003.
13. Eyre DR, Koob TJ, and Van Ness KP. Quantitation of hydroxypyridinium crosslinks in collagen by high-performance liquid chromatography. Anal Biochem 137: 380-388, 1984.
14. Gineyts E, Cloos PA, Borel O, Grimaud L, Delmas PD, and Garnero P.Racemization and isomerization of type I collagen C-telopeptides in human bone and soft tissues: assessment of tissue turnover. Biochem J 345 Pt 3: 481-485, 2000.
15. Giulian G, R. M, and Greaser M. Improved methodology for analysis and quantification of proteins on one-dimensional silver-stained slab gels. Analytical Biochemistry 129: 277-287, 1983.
16. Goldspink G, Fernandes K, Williams PE, and Wells DJ. Age-related changes in collagen gene expression in the muscles of mdx dystrophic and normal mice. Neuromuscul Disord 4: 183-191, 1994.
17. Gosselin LE, Adams C, Cotter TA, McCormick RJ, and Thomas DP. Effect of exercise training on passive stiffness in locomotor skeletal muscle: role of extracellular matrix. J Appl Physiol 85: 1011-1016, 1998.
18. Gosselin LE, Martinez DA, Vailas AC, and Sieck GC. Passive length-force properties of senescent diaphragm: relationship with collagen characteristics. J Appl Physiol 76: 2680-2685, 1994.
19. Hasten DL, Pak-Loduca J, Obert KA, and Yarasheski KE. Resistance exercise acutely increases MHC and mixed muscle protein synthesis rates in 78-84 and 23-32 yr olds. Am J Physiol Endocrinol Metab 278: E620-626, 2000.
20. Huijing PA. Muscle as a collagen fiber reinforced composite: a review of force transmission in muscle and whole limb. J Biomech 32: 329-345, 1999.
21. Hutson PR, Crawford ME, and Sorkness RL. Liquid chromatographic determination of hydroxyproline in tissue samples. J Chromatogr B Analyt Technol Biomed Life Sci 791: 427-430, 2003.
22. Ingalls CP, Warren GL, and Armstrong RB. Dissociation of force production from MHC and actin contents in muscles injured by eccentric contractions. J Muscle Res Cell Motil 19: 215-224, 1998.
23. Kjaer M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol Rev 84: 649-698, 2004.
Page 26 of 38

27
24. Kovanen V. Effects of ageing and physical training on rat skeletal muscle. An experimental study on the properties of collagen, laminin, and fibre types in muscles serving different functions. Acta Physiol Scand Suppl 577: 1-56, 1989.
25. Kovanen V, and Suominen H. Effects of age and life-long endurance training on the passive mechanical properties of rat skeletal muscle. Compr Gerontol [A] 2: 18-23, 1988.
26. Kovanen V, Suominen H, and Heikkinen E. Mechanical properties of fast and slow skeletal muscle with special reference to collagen and endurance training. JBiomech 17: 725-735, 1984.
27. Kovanen V, Suominen H, Risteli J, and Risteli L. Type IV collagen and laminin in slow and fast skeletal muscle in rats--effects of age and life-time endurance training. Coll Relat Res 8: 145-153, 1988.
28. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685., 1970.
29. Lambert CP, and Evans WJ. Adaptations to aerobic and resistance exercise in the elderly. Reviews in endocrine & metabolic disorders 6: 137-143, 2005.
30. Lexell J, Henriksson-Larsen K, Winblad B, and Sjostrom M. Distribution of different fiber types in human skeletal muscles: effects of aging studied in whole muscle cross sections. Muscle Nerve 6: 588-595, 1983.
31. Louis M, Poortmans JR, Francaux M, Berre J, Boisseau N, Brassine E, Cuthbertson DJ, Smith K, Babraj JA, Waddell T, and Rennie MJ. No effect of creatine supplementation on human myofibrillar and sarcoplasmic protein synthesis after resistance exercise. Am J Physiol Endocrinol Metab 285: E1089-1094, 2003.
32. Miller BF, Hansen M, Olesen JL, Flyvbjerg A, Schwarz P, Babraj JA, Smith K, Rennie MJ, and Kjaer M. No effect of menstrual cycle on myofibrillar and connective tissue protein synthesis in contracting skeletal muscle. Am J Physiol Endocrinol Metab 290: E163-E168, 2006.
33. Miller BF, Olesen JL, Hansen M, Dossing S, Crameri RM, Welling RJ, Langberg H, Flyvbjerg A, Kjaer M, Babraj JA, Smith K, and Rennie MJ. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J Physiol 567: 1021-1033, 2005.
34. Mohan S, and Radha E. Age-related changes in rat muscle collagen. Gerontology 26: 61-67, 1980.
35. Monnier VM, Mustata GT, Biemel KL, Reihl O, Lederer MO, Zhenyu D, and Sell DR. Cross-linking of the extracellular matrix by the maillard reaction in aging and diabetes: an update on "a puzzle nearing resolution". Ann N Y Acad Sci 1043: 533-544, 2005.
Page 27 of 38

28
36. Palokangas H, Kovanen V, Duncan A, and Robins SP. Age-related changes in the concentration of hydroxypyridinium crosslinks in functionally different skeletal muscles. Matrix 12: 291-296, 1992.
37. Patel TJ, and Lieber RL. Force transmission in skeletal muscle: from actomyosin to external tendons. Exerc Sport Sci Rev 25: 321-363, 1997.
38. Reddy GK. Cross-linking in collagen by nonenzymatic glycation increases the matrix stiffness in rabbit achilles tendon. Exp Diabesity Res 5: 143-153, 2004.
39. Rizzi C, Mazzoleni F, Sandri M, Rossini K, Brunson A, Rossi M, Cantani C, Cantani M, and Carraro U. Molecular markers of muscle plasticity, damage, regeneration, and repair. In: Muscle Damage, edited by Salmons S. New York: Oxford University Press, Inc., 1997, p. 107-127.
40. Rodrigues CJ, Rodrigues Junior AJ, and Bohm GM. Effects of aging on muscle fibers and collagen content of the diaphragm: a comparison with the rectus abdominis muscle. Gerontology 42: 218-228, 1996.
41. Sajithlal GB, Chithra P, and Chandrakasan G. Advanced glycation end products induce crosslinking of collagen in vitro. Biochim Biophys Acta 1407: 215-224, 1998.
42. Short KR, Vittone JL, Bigelow ML, Proctor DN, and Nair KS. Age and aerobic exercise training effects on whole body and muscle protein metabolism. Am J Physiol Endocrinol Metab 286: E92-101, 2004.
43. Skinner SJ. Rapid method for the purification of the elastin cross-links, desmosine and isodesmosine. J Chromatogr 229: 200-204, 1982.
44. Solaro RJ, Pang DC, and Briggs FN. The purification of cardiac myofibrils with Triton X-100. Biochimica Biophysica Acta 245: 259-262, 1971.
45. Takahashi M, Hoshino H, Kushida K, and Inoue T. Direct measurement of crosslinks, pyridinoline, deoxypyridinoline, and pentosidine, in the hydrolysate of tissues using high-performance liquid chromatography. Anal Biochem 232: 158-162, 1995.
46. Toth MJ, Matthews DE, Tracy RP, and Previs MJ. Age-related differences in skeletal muscle protein synthesis: relation to markers of immune activation. Am J Physiol Endocrinol Metab 288: E883-891, 2005.
47. Trappe S, Gallagher P, Harber M, Carrithers J, Fluckey J, and Trappe T.Single muscle fibre contractile properties in young and old men and women. J Physiol 552: 47-58, 2003.
48. Trappe TA, Lindquist DM, and Carrithers JA. Muscle-specific atrophy of the quadriceps femoris with aging. Journal of Applied Physiology 90: 2070-2074., 2001.
Page 28 of 38

29
49. Tsika RW, Herrick RE, and Baldwin KM. Time course adaptations in rat skeletal muscle isomyosins during compensatory growth and regression. Journal of Applied Physiology 63: 2111-2121, 1987.
50. Volpi E, Sheffield-Moore M, Rasmussen BB, and Wolfe RR. Basal muscle amino acid kinetics and protein synthesis in healthy young and older men. Jama 286: 1206-1212, 2001.
51. Welle S, Thornton C, Statt M, and McHenry B. Postprandial myofibrillar and whole body protein synthesis in young and old human subjects. Am J Physiol 267: E599-604, 1994.
52. Willems ME, Miller GR, and Stauber WT. Force deficits after stretches of activated rat muscle-tendon complex with reduced collagen cross-linking. Eur J Appl Physiol 85: 405-411, 2001.
53. Wilson CM. Studies and critique of Amido Black 10B, Coomassie Blue R, and Fast Green FCF as stains for proteins after polyacrylamide gel electrophoresis. Anal Biochem 96: 263-278, 1979.
54. Yarasheski KE, Pak-Loduca J, Hasten DL, Obert KA, Brown MB, and Sinacore DR. Resistance exercise training increases mixed muscle protein synthesis rate in frail women and men >/=76 yr old. Am J Physiol 277: E118-125, 1999.
55. Zimmerman SD, McCormick RJ, Vadlamudi RK, and Thomas DP. Age and training alter collagen characteristics in fast- and slow-twitch rat limb muscle. J Appl Physiol 75: 1670-1674, 1993.
Page 29 of 38

30
ACKNOWLEDGEMENTS
The authors would like to thank the subjects for their participation. This work was
supported by NIH grants R21 AG15833 (TT), K01 AG00831 (TT), M01 RR-14288, and
R01 AG20532 (TT).
Page 30 of 38

31
FIGURE LEGENDS
Figure 1. Representative chromatographs for each of the connective tissue assays performed in human skeletal muscle. Elution profiles for collagen and collagen cross-linking were similar between young and old, however aging samples displayed increases in advanced glycation endproducts. HYP: hydroxyproline, PRO: proline, HP: hydroxylysylpyridinoline, PE: pentosidine, IS: internal standard. Figure 2. Standard curve generated from pure collagen using the bicinchoninic acid (BCA) protein assay; y=0.002x + 0.0135. Pure collagen (Sigma C9791) was prepared in 0.1 M acetic acid and absorbance was measured at 540 nm confirming the contribution of collagen to the myofibrillar protein quantification with the BCA assay. Figure 3. Connective tissue characteristics of young and old human skeletal muscle. A) Intramuscular collagen concentration determined from hydroxyproline. B) Intramuscular collagen cross-linking described by the concentration of hydroxylysylpyridinoline (HP). C) Intramuscular advanced glycation endproducts described by the concentration of pentosidine. * p<0.05 from Young. Figure 4. Top: Change in the concentration of the main skeletal muscle pools (mixed, myofibrillar and sarcoplasmic) of the vastus lateralis of young and old individuals. Bottom: Primary components of the assayed myofibrillar fraction (myosin, actin and collagen) are no different between young and old despite decreases in the myofibrillar fraction with aging. *p<0.05 from Young. Figure 5. Comparison between young and old skeletal muscle functional characteristics. CSA: cross-sectional area, Po: Peak isometric force, Po/CSA: Peak isometric force normalized to muscle CSA, Peak Power/CSA: Peak power normalized to muscle CSA. *p<0.05 from Young.
Page 31 of 38

Table 1. Subject Characterstics.
* P<0.05 from Young. The Young group contained 10 men and 10 women. The Old group contained 10 men and 12 women.
Variable Young (n=20)
Old (n=22)
Age, y 25 ± 1 (22-30) 78 ± 1 (70-93) *
Height, cm 172 ± 2 167 ± 2 *
Weight, kg 70.6 ± 3.2 70.9 ± 2.6
Body fat, % 25 ± 2 32 ± 2 *
Page 32 of 38

Table 2. Functional Ability and Muscle Characterstics.
* P<0.05 from Young.
Variable Young(n=20)
Old(n=22) %∆
Stair ClimbTime, sPower, W
2.57 ± 0.11550 ± 36
4.62 ± 0.33 *334 ± 32 *
+80-40
Chair rise time, s 0.59 ± 0.02 0.92 ± 0.07 * +56
Walk time, s 2.52 ± 0.09 3.65 ± 0.20 * +45
Quadriceps volume, cm3 881 ± 56 622 ± 35 * -29
Quadriceps Vmax, deg/s 660 ± 16 509 ± 11 * -23
Vastus Lateralis Fiber Type% MHC I% MHC IIa% MHC IIx
27 ± 254 ± 219 ± 1
42 ± 3 *45 ± 2 *14 ± 2 *
+55-17-26
Page 33 of 38

Page 34 of 38

R2 = 0.9964
0.00
0.05
0.10
0.15
0.20
0.25
0 200 400 600 800 1000Collagen Concentration (μg/ml)
Abs
orba
nce
at 5
40 n
m
Page 35 of 38

0
5
10
15
20
Young Old
μg c
olla
gen
per
mg
mus
cle
wet
wei
ght
A
0
125
250
375
500
Young Old
mm
ol H
P pe
r mol
col
lage
n
B
0
6
12
18
24
Young Old
mm
ol p
ento
sidi
nepe
r mol
col
lage
n
*C
Page 36 of 38
0
50
100
150
200
Mixed Myofibrillar Sarcoplasmic
µg p
er m
g m
uscl
e w
et w
t.YoungOld
*
*
*
*
0
25
50
75
100
Myofibrillar Myosin + Actin + Collagen
µg p
er m
g m
uscl
e w
et w
t.
YoungOld
*
Page 37 of 38
0
10
20
30
40
50
60
70
CSA
Sq c
mYoungOld
*
0
50
100
150
200
250
Po
Nm
*
0
1
2
3
4
Po/CSAN
m/S
q cm
*
0
100
200
300
400
500
Peak Power
Wat
ts *
0
2
4
6
8
Peak Power/CSA
Wat
ts/S
q cm
*
Page 38 of 38
Related Documents











