Coleoid beaks from the Nusplingen Lithographic Limestone (Upper Kimmeridgian, SW Germany) CHRISTIAN KLUG, GU ¨ NTER SCHWEIGERT, GERD DIETL AND DIRK FUCHS Klug, C., Schweigert, G., Dietl, G. & Fuchs, D. 2005 09 12: Coleoid beaks from the Nusplingen Lithographic Limestone (Upper Kimmeridgian, SW Germany). Lethaia, Vol. 38, pp. 173–192. Oslo. ISSN 0024-1164. Over twenty specimens of coleoid beaks were recovered from the Nusplingen Litho- graphic Limestone (Upper Jurassic, Late Kimmeridgian, Beckeri Zone, Ulmense Subzone; SW Germany). Based on their morphology, four forms of lower beaks and three forms of upper beaks can be distinguished among the finds. Two gladii of Trachyteuthis hastiformis are associated with a complete beak and hence enable taxonomic assignment of two beak forms. In one of the two specimens, the beaks are still articulated. A third specimen shows another form of upper and lower beak on one slab, which is tentatively interpreted as the beak elements of Plesioteuthis prisca. The largest type of upper beak probably belongs to Leptotheuthis based on its scarcity and size. The remaining two forms are of uncertain taxonomic affiliation. The lower beak of Trachyteuthis resembles that of Recent Octopus and thus suggests a close phylogenetic relation to the Octopoda, as the lower beak of Vampyroteuthis has a different morphology. These originally partially chitinous structures were discovered in the Fossillagersta ¨tte of Nusplingen because the bituminous sediments still contain a significant portion of the original organic matter. This might be the case because of a low permeability of the respective strata. Otherwise, without the color contrast, these delicate structures would hardly be discernible against the carbonate matrix. & Beaks, Coleoidea, Germany, Late Jurassic, morphology, taphonomy, Trachy- teuthis. Christian Klug [[email protected]], Pala ¨ontologisches Institut und Museum, Karl Schmid-Str. 4, CH-8006 Zu ¨rich, Switzerland; Gu ¨nter Schweigert [schweigert.smns@na- turkundemuseum-bw.de], Gerd Dietl [[email protected]], Staatliches Museum fu ¨r Naturkunde, Rosenstein 1, D-70191 Stuttgart, Germany; Dirk Fuchs [drig@ zedat.fu-berlin.de], Institut fu ¨r Geowissenschaften, Freie Universita ¨t Berlin, Malteserstr. 74-100, D-12249 Berlin, Germany; 17th October 2004, revised 14th April 2005. In the course of excavations in the thin-bedded limestones of Late Jurassic age (Kimmeridgian, Beckeri Zone, Ulmense Subzone, hoelderi biohorizon; Schweigert et al. 1996; Schweigert 1998) at the protected Nusplingen Quarry (S of Balingen in SW Germany; Fig. 1) in 2002, two new well-preserved specimens of Trachyteuthis were recovered. Immedi- ately after having split the limestone slabs, it became clear that in both cases gladii were each directly associated with beaks of this coleoid. One of the beak elements closely resembles isolated beak elements that were collected earlier from the Nusplingen Litho- graphic Limestone but which could not be identified taxonomically. The fossiliferous site of the Nusplingen Litho- graphic Limestone near the village of Nusplingen was first recorded by Quenstedt (1843) from a small quarry exploited by a local farmer for paving tiles. In his early publications, Quenstedt (1843, 1855, 1857) already understood the significance of this site and predicted remarkable palaeontological discoveries. Subsequently, Oscar Fraas started to search for fossils intensively at this locality (Fraas 1849, 1854, 1855). From 1855 to 1869, the commercial exploitation of the Nusplingen Plattenkalk was disrupted and the number of finds decreased. The German fossil trader Stu ¨rtz resumed collecting and contributed valuable fossils to the collections of the Geologisch-Pala ¨ontologisches Institut und Museum in Tu ¨bingen and the Staatliche Museum fu ¨r Naturkunde in Stuttgart from 1896 to 1899 (Dietl & Schweigert 2001). Later excavations near Nusplingen were carried out by the University of Tu ¨bingen in 1929 and 1935 (Fahrion 1937) and continued in 1962. In 1993, the latest phase of systematic excavations, which still continue, was initiated by two of us (G. Dietl and G. Schweigert). Westphal (1992) published the first modern view of this exceptional Fossillagersta ¨tte. A bibliography of the older research activities of the Nusplingen Litho- graphic Limestone was compiled by Schweigert (1997); more recently, Dietl and Schweigert (1999b, 2001, 2004) provided comprehensive monographs and summaries on this site. Apart from representatives of the Belemnoidea, fossil remains of representatives of the Coleoidea are rare compared with those of Ammonoidea. This is DOI 10.1080/00241160510013303 # 2005 Taylor & Francis Group Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Coleoid beaks from the Nusplingen Lithographic Limestone(Upper Kimmeridgian, SW Germany)
CHRISTIAN KLUG, GUNTER SCHWEIGERT, GERD DIETL AND DIRK FUCHS
Klug, C., Schweigert, G., Dietl, G. & Fuchs, D. 2005 09 12: Coleoid beaks from theNusplingen Lithographic Limestone (Upper Kimmeridgian, SW Germany). Lethaia, Vol.38, pp. 173–192. Oslo. ISSN 0024-1164.
Over twenty specimens of coleoid beaks were recovered from the Nusplingen Litho-graphic Limestone (Upper Jurassic, Late Kimmeridgian, Beckeri Zone, Ulmense Subzone;SW Germany). Based on their morphology, four forms of lower beaks and three forms ofupper beaks can be distinguished among the finds. Two gladii of Trachyteuthis hastiformisare associated with a complete beak and hence enable taxonomic assignment of two beakforms. In one of the two specimens, the beaks are still articulated. A third specimen showsanother form of upper and lower beak on one slab, which is tentatively interpreted as thebeak elements of Plesioteuthis prisca. The largest type of upper beak probably belongs toLeptotheuthis based on its scarcity and size. The remaining two forms are of uncertaintaxonomic affiliation. The lower beak of Trachyteuthis resembles that of Recent Octopusand thus suggests a close phylogenetic relation to the Octopoda, as the lower beak ofVampyroteuthis has a different morphology. These originally partially chitinous structureswere discovered in the Fossillagerstatte of Nusplingen because the bituminous sedimentsstill contain a significant portion of the original organic matter. This might be the casebecause of a low permeability of the respective strata. Otherwise, without the colorcontrast, these delicate structures would hardly be discernible against the carbonatematrix. & Beaks, Coleoidea, Germany, Late Jurassic, morphology, taphonomy, Trachy-teuthis.
Christian Klug [[email protected]], Palaontologisches Institut und Museum, KarlSchmid-Str. 4, CH-8006 Zurich, Switzerland; Gunter Schweigert [[email protected]], Gerd Dietl [[email protected]], StaatlichesMuseum fur Naturkunde, Rosenstein 1, D-70191 Stuttgart, Germany; Dirk Fuchs [[email protected]], Institut fur Geowissenschaften, Freie Universitat Berlin, Malteserstr.74-100, D-12249 Berlin, Germany; 17th October 2004, revised 14th April 2005.
In the course of excavations in the thin-beddedlimestones of Late Jurassic age (Kimmeridgian,Beckeri Zone, Ulmense Subzone, hoelderi biohorizon;Schweigert et al. 1996; Schweigert 1998) at theprotected Nusplingen Quarry (S of Balingen in SWGermany; Fig. 1) in 2002, two new well-preservedspecimens of Trachyteuthis were recovered. Immedi-ately after having split the limestone slabs, it becameclear that in both cases gladii were each directlyassociated with beaks of this coleoid. One of the beakelements closely resembles isolated beak elements thatwere collected earlier from the Nusplingen Litho-graphic Limestone but which could not be identifiedtaxonomically.
The fossiliferous site of the Nusplingen Litho-graphic Limestone near the village of Nusplingen wasfirst recorded by Quenstedt (1843) from a smallquarry exploited by a local farmer for paving tiles. Inhis early publications, Quenstedt (1843, 1855, 1857)already understood the significance of this site andpredicted remarkable palaeontological discoveries.Subsequently, Oscar Fraas started to search for fossilsintensively at this locality (Fraas 1849, 1854, 1855).
From 1855 to 1869, the commercial exploitation of theNusplingen Plattenkalk was disrupted and the numberof finds decreased. The German fossil trader Sturtzresumed collecting and contributed valuable fossils tothe collections of the Geologisch-PalaontologischesInstitut und Museum in Tubingen and the StaatlicheMuseum fur Naturkunde in Stuttgart from 1896 to1899 (Dietl & Schweigert 2001). Later excavations nearNusplingen were carried out by the University ofTubingen in 1929 and 1935 (Fahrion 1937) andcontinued in 1962. In 1993, the latest phase ofsystematic excavations, which still continue, wasinitiated by two of us (G. Dietl and G. Schweigert).Westphal (1992) published the first modern view ofthis exceptional Fossillagerstatte. A bibliography of theolder research activities of the Nusplingen Litho-graphic Limestone was compiled by Schweigert(1997); more recently, Dietl and Schweigert (1999b,2001, 2004) provided comprehensive monographs andsummaries on this site.
Apart from representatives of the Belemnoidea,fossil remains of representatives of the Coleoidea arerare compared with those of Ammonoidea. This is
DOI 10.1080/00241160510013303 # 2005 Taylor & Francis Group Ltd

mainly because of the primary composition of thehard parts in Mesozoic coleoids, which presumablyconsisted of a mixture of chitinous material andaragonite. Remarkably, some exceptional Fossillager-statten (Seilacher 1970) yielded fossils of coleoids,which, in several cases, display most of the major softtissues, including tentacles, ink sac, eyes, mantlemuscles, intestines, blood vessels, and even gills (Kearet al. 1995). Examples include Mazon Creek (LatePennsylvanian; Schellenberg 2002), Monte SanGiorgio (Anisian; Rieber 1970, 1973), Raibl (Karnian;Bronn 1859; Mojsisovics 1882, 1902), Osteno (Sine-murian; Pinna 1972, 1985; Tang 2002a), Holzmaden(Toarcian; Reitner & Engeser 1982; Riegraf 1982, 1983;Reitner & Urlichs 1983; Riegraf & Hauff 1983; Riegrafet al. 1984, 1998; Etter & Tang 2002), Montlandon inLangres (Toarcian; Guerin-Franiatte & Gouspy 1993),La Voulte-sur-Rhone (Callovian; Frohlich et al. 1992;Wilby et al. 1995, 1996; Etter 2002; Haas 2002),Christian Malford (Callovian to Oxfordian; Donovan
1983; Martill 1987; Allison 1988b; Donovan & Crane1992; Kear et al. 1995; Tang 2002b), the Solnhofen/Eichstatt area (Tithonian; Naef 1922; Engeser &Reitner 1981, 1992; Bandel & Leich 1986; Allison1988a; Hewitt & Wignall 1988; Mehl 1990; Briggs et al.1993; Frickhinger 1994, 1999; Donovan 1995; Briggs &Wilby 1996; Engeser & Keupp 1997, 1999; Donovanet al. 2002), Sahel Alma (Santonian; Engeser & Reitner1986), and Nusplingen.
Fossil coleoid beaks, however, have been reported inonly a few articles (Bronn 1859; Mojsisovics 1882,1902; Lossel 1947; Rieber 1973; Pinna 1985; Dzik 1986;Kear et al. 1995; Harzhauser 1999; Haas 2002; Tanabeet al. in press, 2006). Well preserved coleoid beaks areexceedingly rare (Dzik 1986) although beak remainshave been recorded from several localities (Kear et al.1995). In recent years, the two excavation sites nearNusplingen (Nusplingen and Egesheim quarries)yielded flattened but otherwise well preserved materialof complete ammonoid and nautiloid beaks, many of
Fig. 1. Map with the location of the Nusplingen and Egesheim quarries, SW Germany, showing the distribution of land and sea during the LateKimmeridgian.
174 Christian Klug et al. LETHAIA 38 (2005)

them in situ (Schweigert 1998; Dietl & Schweigert1999a, 2001; Schweigert & Dietl 1999, 2001), as well ascoleoid gladii (Dietl et al. 1995, 1997). This is of specialinterest because the upper mandibles of ammonoidswere largely chitinous and lack mineralized portions.Thus, they are only preserved under special condi-tions, or else here their recognition was facilitated bythe preservation in the Nusplingen LithographicLimestone. At this site, the dark organic matter of thebeak elements is sometimes preserved and presents agood contrast to the light greyish limestone matrix.Consequently, it is not surprising that more than20 coleoid beak elements were excavated from thesestrata.
According to Allison (1988a), chitin requires amoderately early mineralization and a low rate ofdecay to become fossilized. During their experimentswith decaying squids, Kear et al. (1995, p. 115) foundthat most chitinous structures remained more orless intact for the duration of their observations (50weeks). With respect to coleoid beaks, they observedinitial disintegration of the less tanned and thinnerposterior parts of the beaks after ten weeks.
It is the aim of this study to document and interpretthe remains of coleoid beaks from the NusplingenLithographic Limestone and to provide reconstruc-tions of some of the beak elements and to discussthe phylogenetic and taphonomic framework of thefinds.
Material and measurements
Most of the studied specimens were collected by theStuttgart excavation team and are deposited in theStaatliches Museum fur Naturkunde in Stuttgart(SMNS). A few specimens were discovered in thecourse of the early excavations, some of which aredeposited in the Institut und Museum fur Geologieund Palaontologie of Tubingen University (GPIT). Inthe appendices, the gladii of Trachyteuthis hastiformis(Appendix 1) and Plesioteuthis prisca (Appendix 2) aswell as all beak remains are listed (Appendix 3).Parameters measured on the beak elements and thegladii are explained in Fig. 2.
Remarks on the systematic framework
In the last decade, several classifications of theColeoidea (Fig. 3) have been suggested (Engeser1990a, 1990b; Doyle et al. 1994; Young et al. 1998;Boletzky 1999; Haas 2002). With respect to thesister-group relationships between the Octobrachia(= Octopodiformes sensu Engeser 1990a or Young et al.1998, = Vampyropoda sensu Boletzky 1999) and theDecabrachia (= Decapodiformis sensu Young et al.1998, = Decabrachiomorpha sensu Haas 2002), orbetween the Octopoda (= Octobrachia sensu Boletzky1999) and the Vampyromorpha (= Pseudoctobrachia
Fig. 2. Parameters of the gladii and beak elements from the Nusplingen Lithographic limestone. &A. Gladius of Trachyteuthis with width (wgl)and length (lgl). &B. Lower beak of Trachyteuthis with lengths of the complete specimen (lb) and the internal lamella (lbil), as well as thewidths of both internal (lbil) and external lamella (lbel). &C. Lower beak of Trachyteuthis with height and width of the external lamella (lbel).&D. Upper beak of Trachyteuthis with height and length of the complete specimen (ub).
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 175

sensu Boletzky 1999), they are all consistent with eachother. It is not the aim of this paper, however, toclarify this terminology. All divisions listed above weremade on the basis of the shell and especially on thearm crown.
The differences between the taxonomic unitsmentioned above consist predominantly in theaffiliation of the so-called ‘fossil teuthids’ (Bandel &Leich 1986; Fig. 3). These include the majority ofMesozoic representatives of the Coleoidea except theBelemnoidea. Based on gladius characteristics and thetransformation of the second arm pair into retractilefilaments (still unknown from the fossil record),Engeser (1990a) placed these fossil forms togetherwith Vampyroteuthis in the Vampyromorpha.Although Doyle et al. (1994) accepted them as Octo-brachia, Engeser retained the Plesioteuthididae inthe Decabrachia. Young et al. (1998) also doubted theposition of all ‘fossil teuthids’ within the Octobrachiaand at least considered Plesioteuthis and Mastigophoraas Decabrachia. The question of whether Trachyteuthisis a ten- or an eight-armed cephalopod is more than150 years old and is still not satisfactorily answered.Donovan et al. (2002) described and figured one adultspecimen of Trachyteuthis (deposited in the SwedishMuseum of Natural History in Stockholm) with twopairs of fins preserved, which indicates a vampyro-morph affiliation. In contrast, Haas (2002) regardedthe ‘fossil teuthids’ as stem-lineage representatives ofthe Octobrachia (‘Trachyteuthimorpha’?).
Kluessendorf & Doyle (2000) described Pohlsepiamazonensis from the Carboniferous of Illinois (USA).They considered their find (one poorly preservedspecimen) as a representative of the Octopoda alreadylacking an internal shell (the structures they inter-preted as fins might well represent a trace of theinternal shell). If they are correct and this is not anartifact, phylogenetic principles require a separation ofOctopoda and Vampyromorpha before Carboniferoustimes. In that case, Jurassic ‘fossil teuthids’ cannotbe regarded as stem-lineage representatives of theOctobrachia. Consequently, the term ‘fossil teuthids’should be used as long as the placement of theseanimals is unsolved.
So far, the Nusplingen Lithographic Limestone hasyielded remains of three presumed octobrachiangenera: Trachyteuthis, Plesioteuthis, and Leptotheuthis.Trachyteuthis and Leptotheuthis belong to the suborderMesoteuthina, whereas Plesioteuthis is included in thesuborder Prototeuthina. In the following paragraphs,we attempt to identify the taxonomic affiliation of thenew finds of coleoid beaks.
Forms of coleoid beaks from theNusplingen Lithographic Limestone
The Fossillagerstatten between the villages ofNusplingen and Egesheim yielded the remains of morethan 20 beak elements, which have been assigned to
Fig. 3. Sister-group relationships within the Coleoidea involving the questionable placement of the ‘fossil teuthids’ (indicated by arrows). Notethe different terminology of: &A. Engeser (1990a) who positioned them in the Vampyromorpha; &B. Doyle et al. (1994) who left the positionof the ‘fossil teuthids’ open, like Young et al. (1998); according to Doyle, they are either a stemgroup of the Octobrachia or of the Decabrachia;&C. Young et al. (1998) who included the ‘fossil teuthids’ either in the Decapodiformes or in the Vampyromorpha; and &D. Haas (2002)who created a new parataxon comprising these ‘fossil teuthids’, the ‘Trachyteuthimorpha’.
176 Christian Klug et al. LETHAIA 38 (2005)
the Coleoidea based on their general morphology(Appendix 3; Figs 4–9). Among the specimens in-cluded in this study, seven forms of beaks can so far
be distinguished; forms (1) to (4) are lower beaks andforms (5) to (7) are upper beaks. The terminology isused according to Clarke (1986, fig. 4).
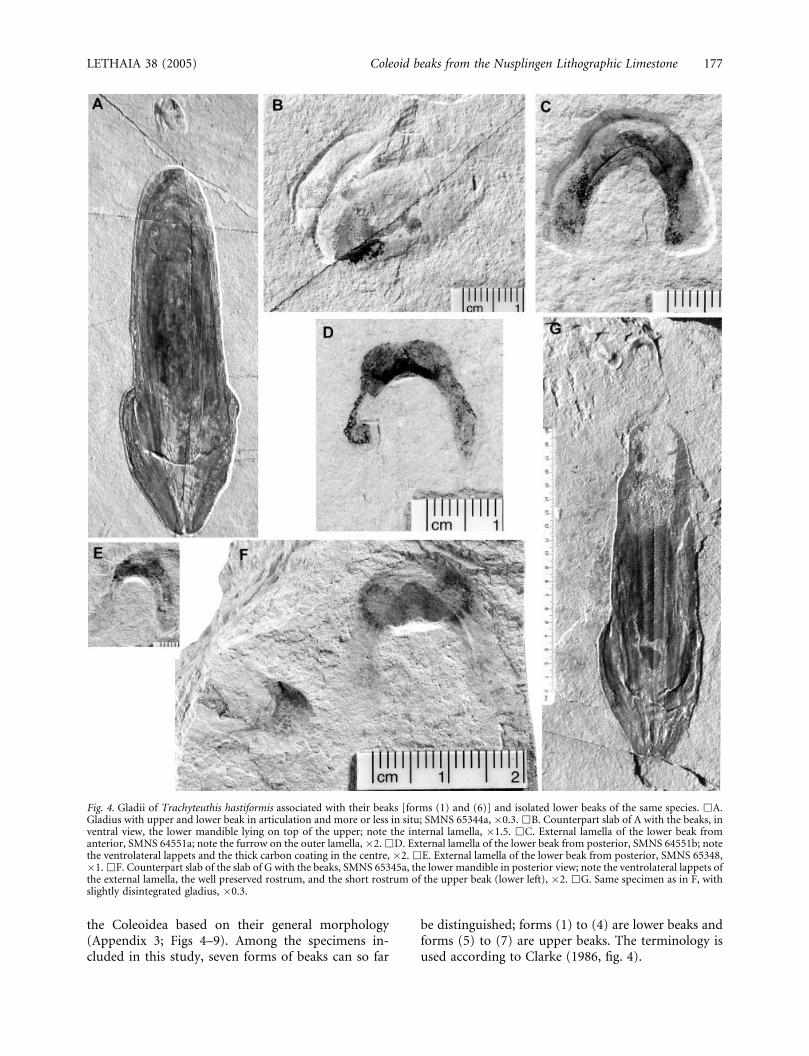
Fig. 4. Gladii of Trachyteuthis hastiformis associated with their beaks [forms (1) and (6)] and isolated lower beaks of the same species. &A.Gladius with upper and lower beak in articulation and more or less in situ; SMNS 65344a, �0.3. &B. Counterpart slab of A with the beaks, inventral view, the lower mandible lying on top of the upper; note the internal lamella, �1.5. &C. External lamella of the lower beak fromanterior, SMNS 64551a; note the furrow on the outer lamella, �2. &D. External lamella of the lower beak from posterior, SMNS 64551b; notethe ventrolateral lappets and the thick carbon coating in the centre, �2. &E. External lamella of the lower beak from posterior, SMNS 65348,�1. &F. Counterpart slab of the slab of G with the beaks, SMNS 65345a, the lower mandible in posterior view; note the ventrolateral lappets ofthe external lamella, the well preserved rostrum, and the short rostrum of the upper beak (lower left), �2. &G. Same specimen as in F, withslightly disintegrated gladius, �0.3.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 177
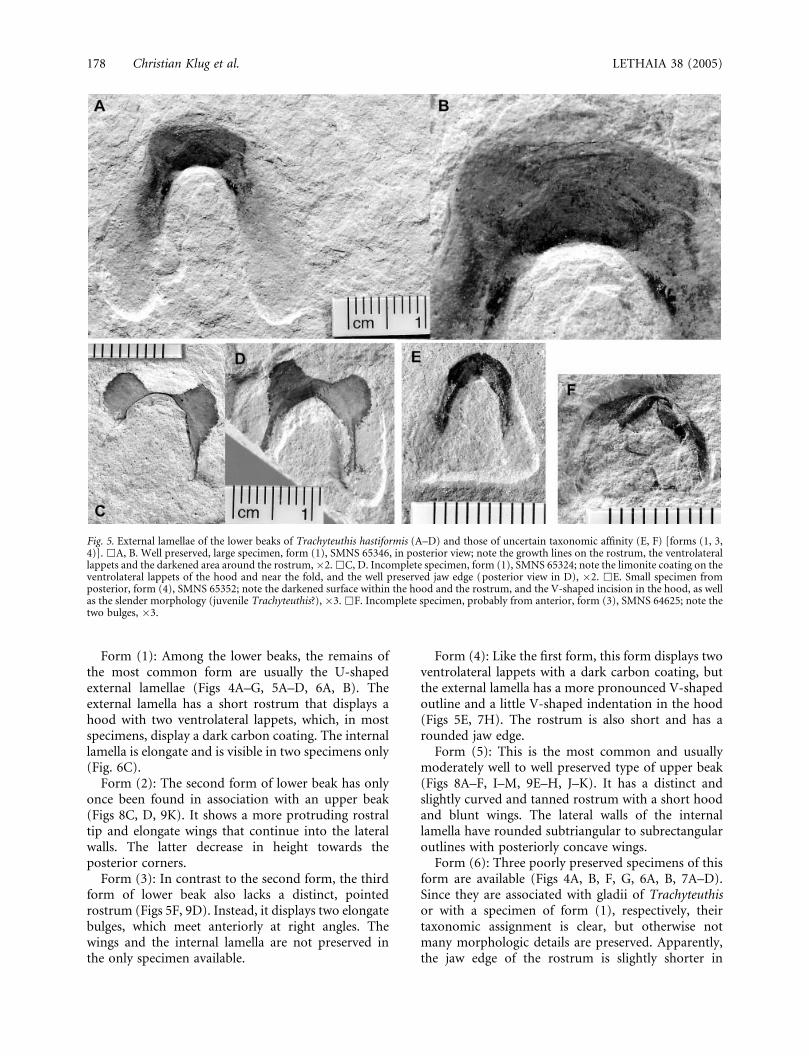
Form (1): Among the lower beaks, the remains ofthe most common form are usually the U-shapedexternal lamellae (Figs 4A–G, 5A–D, 6A, B). Theexternal lamella has a short rostrum that displays ahood with two ventrolateral lappets, which, in mostspecimens, display a dark carbon coating. The internallamella is elongate and is visible in two specimens only(Fig. 6C).
Form (2): The second form of lower beak has onlyonce been found in association with an upper beak(Figs 8C, D, 9K). It shows a more protruding rostraltip and elongate wings that continue into the lateralwalls. The latter decrease in height towards theposterior corners.
Form (3): In contrast to the second form, the thirdform of lower beak also lacks a distinct, pointedrostrum (Figs 5F, 9D). Instead, it displays two elongatebulges, which meet anteriorly at right angles. Thewings and the internal lamella are not preserved inthe only specimen available.
Form (4): Like the first form, this form displays twoventrolateral lappets with a dark carbon coating, butthe external lamella has a more pronounced V-shapedoutline and a little V-shaped indentation in the hood(Figs 5E, 7H). The rostrum is also short and has arounded jaw edge.
Form (5): This is the most common and usuallymoderately well to well preserved type of upper beak(Figs 8A–F, I–M, 9E–H, J–K). It has a distinct andslightly curved and tanned rostrum with a short hoodand blunt wings. The lateral walls of the internallamella have rounded subtriangular to subrectangularoutlines with posteriorly concave wings.
Form (6): Three poorly preserved specimens of thisform are available (Figs 4A, B, F, G, 6A, B, 7A–D).Since they are associated with gladii of Trachyteuthisor with a specimen of form (1), respectively, theirtaxonomic assignment is clear, but otherwise notmany morphologic details are preserved. Apparently,the jaw edge of the rostrum is slightly shorter in
Fig. 5. External lamellae of the lower beaks of Trachyteuthis hastiformis (A–D) and those of uncertain taxonomic affinity (E, F) [forms (1, 3,4)]. &A, B. Well preserved, large specimen, form (1), SMNS 65346, in posterior view; note the growth lines on the rostrum, the ventrolaterallappets and the darkened area around the rostrum, �2. &C, D. Incomplete specimen, form (1), SMNS 65324; note the limonite coating on theventrolateral lappets of the hood and near the fold, and the well preserved jaw edge (posterior view in D), �2. &E. Small specimen fromposterior, form (4), SMNS 65352; note the darkened surface within the hood and the rostrum, and the V-shaped incision in the hood, as wellas the slender morphology (juvenile Trachyteuthis?), �3. &F. Incomplete specimen, probably from anterior, form (3), SMNS 64625; note thetwo bulges, �3.
178 Christian Klug et al. LETHAIA 38 (2005)

contrast to that of form (5) or (7). It thus fits into therounded rostrum of the lower beak of form (1).
Form (7): The last form is represented by only oneisolated specimen (Figs 8G, H, 9I). It is abraded butotherwise shows fine details. The anterior duplicatureis narrow and high and both the hood and the rostrumare long. It has a larger height/length ratio than theforms (5) and (6).
The beak of Trachyteuthis hastiformis
With the discovery of two specimens of Trachyteuthisthat are preserved with their gladii as well as withupper and lower beaks (SMNS 65344a; Figs 4A, B, F,G, 6A, B, 7A–D), the taxonomic affiliation of earlierfinds of isolated beaks from the same localities becamelikely. Of the 24 gladii of this species (Appendix 1),only two are preserved with the beak elements. Onthe other hand, nine isolated lower beaks of form (1)and one pair of beaks [forms (1) and (6)] can nowbe assigned to this species with some reservation(Figs 4–7). The lower beak of Plesioteuthis and Lepto-theuthis could also be of a morphology similar to thatof Trachyteuthis. Nevertheless, it is reasonable toassign this form of lower beak to the ‘Trachyteuthi-morpha’ (sensu Haas 2002), since the Nusplingen
Lithographic Limestone has so far yielded noother coleoid gladii. We have tentatively assigned these13 beaks to Trachyteuthis, since the second mostcommon beak form [(5), eight specimens] couldbelong to the latter genus, according to the numberof gladii of Plesioteuthis discovered so far (sevenspecimens).
The lower beaks, here assigned to Trachyteuthis,predominantly display remains of the external lamella,except for two specimens (SMNS 65344a, 65349;Figs 4A, B, 6, 7A, B). In its compacted and thus flat-tened state, the outer lamella has a U-shaped outlinewith a hood consisting of two dorsolateral lappets. Thecarbon coating is usually the thickest in two sub-circular spots situated within the lappets. In mostspecimens, these two lappets are interconnected by anarrower band of a carbon coating, which is of equalthickness in many specimens (e.g. SMNS 65346,65324) and even thicker in one other specimen (SMNS64551b; Figs 4D, 7G). In some specimens, two darkcarbonaceous bands, which originate at these lappets,run dorsally on the wing folds. In most remains ofthese lower beaks, a subtle three-dimensional relief ispreserved (Figs 5A, B, 7F). This relief mainly consistsof a depression, located centrally on the inside of theU-shape. It is smoothly rounded and, even in the
Fig. 6. &A, B. Upper and lower beaks of Trachyteuthis hastiformis (SMNS 65478), probably from one specimen; part and counterpart slab,forms (1) and (6), �1.5. &C. Lower beak of Trachyteuthis hastiformis (SMNS 65349) from dorsal, with abraded internal lamella pointing tothe lower left and incomplete remains of the anterior duplicature stretched out towards the upper right; note the darkened rostrum and hood,form (1), �2.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 179

largest and best preserved specimens, very shallow.This led us to conclude that the rostral tip was rathermore rounded than pointed, as in the lower beaks of
Recent Octopus. The wings of the external lamella are10.6 to 26 mm high and the external lamella is, in itsflattened state, 10.5 to 27 mm wide.
Fig. 7. Outline drawings of the remains of Trachyteuthis hastiformis (except H.) from the Late Kimmeridgian Plattenkalk of Nusplingen; areasshaded in light grey indicate a thickened carbon coating and in G., the darker grey indicates the increased thickness of the carbon layer. &A.Articulated upper and lower beak, form (1) and (6), SMNS 65344a, �2. &B. Same specimen as in A, gladius with articulated upper and lowerbeak in situ, SMNS 65344a, �0.2. &C. Associated upper and lower beak, form (1) and (6), SMNS 65345, �2. &D. Same specimen as in C,partly disintegrated gladius with upper and lower beak, SMNS 65345, �0.2. &E. Lower beak, form (1), SMNS 65351a, �2. &F. Large lowerbeak, form (1), SMNS 65346, with growth lines near the rostral tip and on the hood, �2. &G. Lower beak, form (1), SMNS 64551b, �2. &H.Lower beak of unknown taxonomic affiliation, form (4), SMNS 65352, �3.
180 Christian Klug et al. LETHAIA 38 (2005)
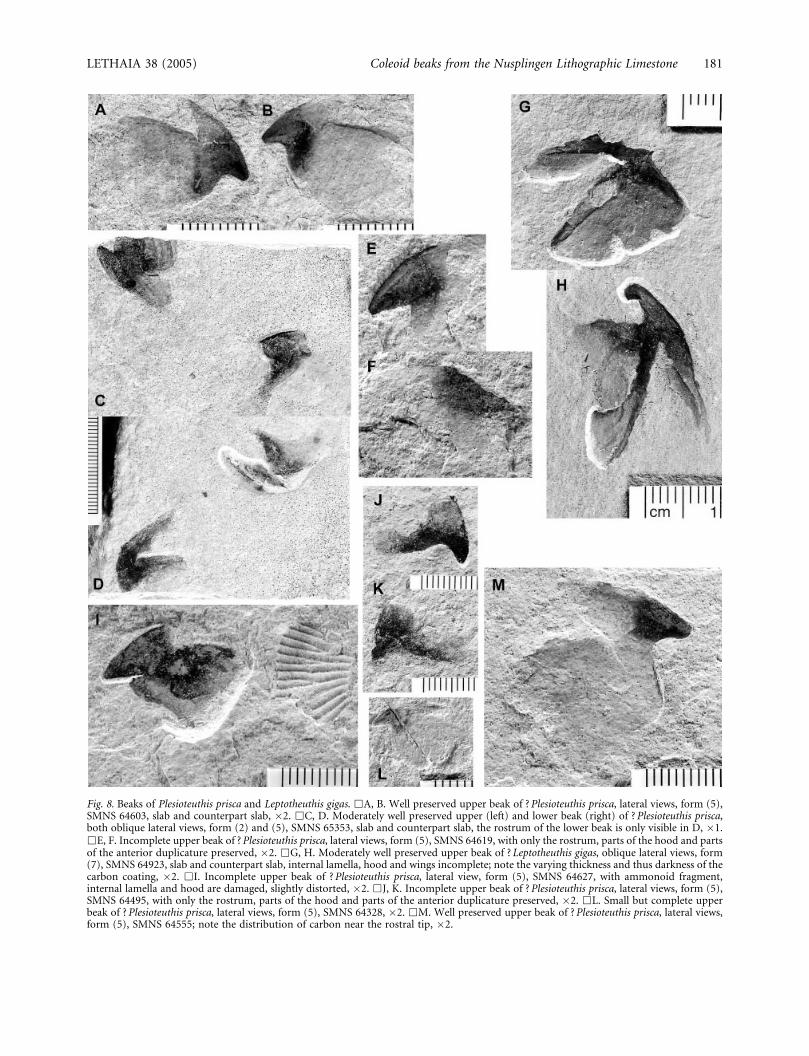
Fig. 8. Beaks of Plesioteuthis prisca and Leptotheuthis gigas. &A, B. Well preserved upper beak of ? Plesioteuthis prisca, lateral views, form (5),SMNS 64603, slab and counterpart slab, �2. &C, D. Moderately well preserved upper (left) and lower beak (right) of ? Plesioteuthis prisca,both oblique lateral views, form (2) and (5), SMNS 65353, slab and counterpart slab, the rostrum of the lower beak is only visible in D, �1.&E, F. Incomplete upper beak of ? Plesioteuthis prisca, lateral views, form (5), SMNS 64619, with only the rostrum, parts of the hood and partsof the anterior duplicature preserved, �2. &G, H. Moderately well preserved upper beak of ? Leptotheuthis gigas, oblique lateral views, form(7), SMNS 64923, slab and counterpart slab, internal lamella, hood and wings incomplete; note the varying thickness and thus darkness of thecarbon coating, �2. &I. Incomplete upper beak of ? Plesioteuthis prisca, lateral view, form (5), SMNS 64627, with ammonoid fragment,internal lamella and hood are damaged, slightly distorted, �2. &J, K. Incomplete upper beak of ? Plesioteuthis prisca, lateral views, form (5),SMNS 64495, with only the rostrum, parts of the hood and parts of the anterior duplicature preserved, �2. &L. Small but complete upperbeak of ? Plesioteuthis prisca, lateral views, form (5), SMNS 64328, �2. &M. Well preserved upper beak of ? Plesioteuthis prisca, lateral views,form (5), SMNS 64555; note the distribution of carbon near the rostral tip, �2.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 181
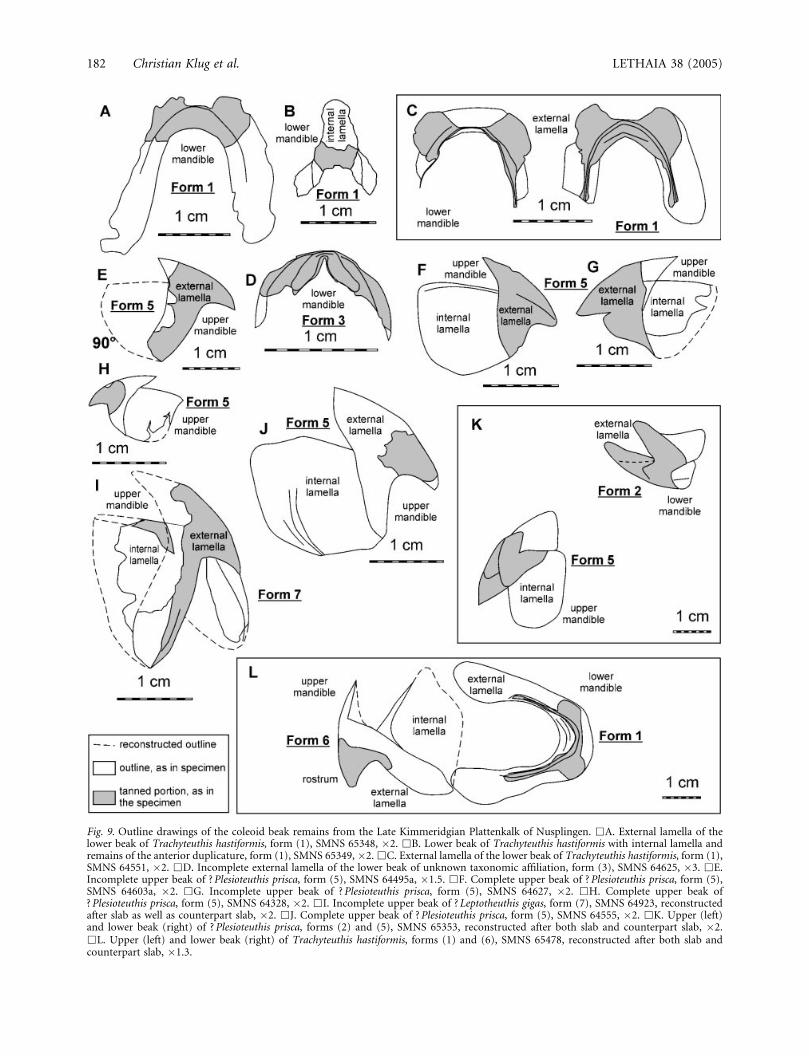
Fig. 9. Outline drawings of the coleoid beak remains from the Late Kimmeridgian Plattenkalk of Nusplingen. &A. External lamella of thelower beak of Trachyteuthis hastiformis, form (1), SMNS 65348, �2. &B. Lower beak of Trachyteuthis hastiformis with internal lamella andremains of the anterior duplicature, form (1), SMNS 65349, �2. &C. External lamella of the lower beak of Trachyteuthis hastiformis, form (1),SMNS 64551, �2. &D. Incomplete external lamella of the lower beak of unknown taxonomic affiliation, form (3), SMNS 64625, �3. &E.Incomplete upper beak of ?Plesioteuthis prisca, form (5), SMNS 64495a, �1.5. &F. Complete upper beak of ? Plesioteuthis prisca, form (5),SMNS 64603a, �2. &G. Incomplete upper beak of ? Plesioteuthis prisca, form (5), SMNS 64627, �2. &H. Complete upper beak of? Plesioteuthis prisca, form (5), SMNS 64328, �2. &I. Incomplete upper beak of ? Leptotheuthis gigas, form (7), SMNS 64923, reconstructedafter slab as well as counterpart slab, �2. &J. Complete upper beak of ? Plesioteuthis prisca, form (5), SMNS 64555, �2. &K. Upper (left)and lower beak (right) of ? Plesioteuthis prisca, forms (2) and (5), SMNS 65353, reconstructed after both slab and counterpart slab, �2.&L. Upper (left) and lower beak (right) of Trachyteuthis hastiformis, forms (1) and (6), SMNS 65478, reconstructed after both slab andcounterpart slab, �1.3.
182 Christian Klug et al. LETHAIA 38 (2005)

The best preserved and also largest specimen clearlyshows the gentle bulging of the rostrum, which iscovered by faint growth lines (SMNS 65346; Figs 5A,B, 7F). Like all other external lamellae of this kind, itdisplays a fine fold, which runs subparallel to the jawedge and the wing fold, fading out towards the dorsalend of the wings. Above the jaw angle, this foldapproaches the shoulder.
The internal lamella of the lower beak shows anelongate triangular outline with a rounded posteriorend. In both specimens, however, the outline is veryirregular, which prohibits conclusions on the details ofits morphology. Consequently, the posterior end ofthe internal lamella was tentatively reconstructedaccording to the shape of this part in Recent Octopus.In the specimens SMNS 65344 and 65349, the remainsof the internal lamellae are 9 and 29 mm long.
The beak of Plesioteuthis prisca
Quenstedt (1857) recorded a single specimen of acoleoid beak, which he tentatively assigned to Plesio-teuthis. Naef (1922, fig. 42) followed this interpreta-tion in his reconstruction of Plesioteuthis prisca. Thepoorly preserved original specimen, housed in the IGPTubingen collection, was re-figured in a sketch byRiegraf & Schmitt-Riegraf (1995, pl. 3, fig. 1). Sincethere are no further specimens of Plesioteuthis gladiiwith in situ beaks available, the assignment of isolatedbeak elements to this genus is still preliminary anduncertain. Comparison is based largely on the differ-ences in morphology of the upper beaks associatedwith Trachyteuthis which are, however, poorlypreserved.
Nevertheless, the upper beak of form (5) probablydid not belong to Trachyteuthis according to thedifferent morphology of those upper beaks associatedwith the two gladii (SMNS 65344, 65345). Further-more, seven specimens each of the gladii of Plesio-teuthis as well as of the beak form (5) were alreadyfound.
Additionally, we have studied one pair of associatedbeaks, here interpreted as one lower [form (2)] andone upper beak [form (5)] of one individual (Figs 8C,D, 9K). Apparently the part interpreted as lower beak[form (2)] displays a pointed rostrum, in contrast tothe specimens assigned to Trachyteuthis and to RecentOctopus. Furthermore, most isolated upper beaks ofform (5) have a moderate size; the only larger speci-men SMNS 64923 [form (7)] also displays a differentmorphology (Figs 8G, H, 9I). Hence, it was interpretedas belonging to the largest species so far recoveredfrom the Nusplingen Lithographic Limestone, Lepto-theuthis gigas.
The only lower beak [form (2)] which might belongto Plesioteuthis is the one mentioned above, which wasfound together with an upper beak [SMNS 65353;form (5)]. The height of the internal lamella decreasesposteriorward simultaneously with the thickness of itsfaint carbonaceous coating. Both wings of the externallamella are stretched out in an oblique anteriordirection. They are darkened by a carbonaceouscoating which is thicker than that of the internallamella. This increased thickness of the coating canbest be explained by the anterior duplicature incombination with the stronger chitinization. In thispart of the beak, the chitinous material had to takemore mechanical power when in operation thanthe more dorsal and more posterior portions. Therostrum and the hood are visible only on one ofthe two slabs, but it is clear that the rostrum is quitedistinct and the hood extended posteriorly overapproximately one fifth of the length of the internallamella.
The upper beak [form (5)] also displays a distinctrostrum, which is slightly bent ventrally (Figs 8A–F,I–M, 9E–H, J, K). The rostrum is darkened by carbonin all specimens but the dark colour fades out ventrallyand posteriorly to the end of the hood. In all speci-mens, the anterior duplicature does not extend all theway to the most ventral edge of the lateral wings. Theinternal lamella decreases in height posteriorly.
The beak of Leptotheuthis gigas
So far, only two large specimens of Leptotheuthis gigaswere recovered (Fig. 10). The only complete indi-vidual, housed in IGP Tubingen, was discoveredduring an early excavation (Dietl & Schweigert 2001,fig. 74). The specimen shows the pairs of small fins,the head with the eyes, the funnel tube, and severalof the arms including the cirri. Additionally, faintremains of the beak are visible. Since it was poorlyprepared, many fine details of the specimen are lost. Itis only possible to discern the rough dimensions butotherwise the specimen yielded no information on themorphology of its beaks.
One isolated upper beak [form (7)] was tentativelyassigned to Leptotheuthis because of its large size andunique morphology (Figs 8G, H, 9I). This specimen isquite well preserved, although fragments of the beakare distributed on both the part and the counterpartlimestone slabs. In this case, the thin carbon film iswell preserved and indicates the location of the ante-rior duplicature and the stronger rostral chitinousmaterial, which probably also reflects the outline of theportions having been tanned prior to the cephalopod’sdeath; the beak measures 27 mm in height. Because ithad settled at an oblique angle on one side, it is
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 183

longitudinally distorted. Furthermore, the posteriormargin of the internal lamella is missing and that ofthe external lamella is abraded. Since the chitinouspart thins out posteriorly in both lamellae, and theedge has the irregular aspect of a fracture, the finerparts of this specimen were probably mechanicallydamaged during transport or else might have beenbiologically decomposed (Kear et al. 1995). Theanterior fold is rather slender and so is the hood. Theedge of the rostrum is obscured by one of the lateralwings and its outline ill-defined.
Problematic specimens
Two specimens preclude firm taxonomic assignment[forms (3) and (4)]. One of them (SMNS 65352;Figs 5E, 7H) closely resembles the external lamella ofthe lower beak of Trachyteuthis in overall outline and
in the rough distribution of the thickened carboncoating [form (4)]. It differs from Trachyteuthis in itsnarrower and more V-shaped outline as well as in thepresence of a V-shaped indentation on its ventral edge.Additionally, the distribution of the carbonaceousmatter is not identical. Nevertheless, it might as wellbelong to a younger specimen of Trachyteuthis andcould be preserved in a different mode.
The second specimen has an entirely different shape[form (3); Figs 5F, 9D]. Given its overall morphology,it appears likely that it is a crescent-shaped externallamella with only little remains of the lateral wings andnone of the internal lamella. Adjacent to the ques-tionable rostral tip, two elongate bulges are visible.The bulges measure 5 mm in length, are arranged atan angle of approximately 90� and are fused dorsallywith a narrow fold. They resemble the teeth of deca-pod lower beaks, apart from the fact that the bulges are
Fig. 10. Leptotheuthis gigas, GPIT 17633. &A. Complete specimen, �0.2. &B. Detail of the head with faint remains of the beak [form (7)?];note the cirri on the arms, the buccal mass and the eye, �0.5.
184 Christian Klug et al. LETHAIA 38 (2005)

more or less fused and there is no rostrum on theirdorsal side. Besides a narrow anterior fold, the rest ofthe external lamella revealed no further morphologicdetails.
Taphonomy
All remains of coleoid beaks discussed in this articleare more or less flattened. The maximum thickness ofthe specimens from the Nusplingen and Egesheimquarries is about 2 mm. Assuming a maximumprimary vertical extension of the beaks of 20 mmabove the sediment surface, this would imply a tenfoldreduction in the thickness of the specimens. Thisdistortion hampers the identification of some partsand thus also the reconstruction in general. It mostlikely derives from compaction due to expulsion ofwater from the original soft sediment.
In order to clarify the elemental composition ofthese fossils, an EDAX analysis was performed on amandible fragment of Plesioteuthis. It clearly showedthat the dark coatings mainly consist of carbon withoxygen and small amounts of calcium, potassium,silicon, aluminum, and sulfur. Phosphorus is appar-ently completely absent in the beak material, althoughphosphatized soft-tissues are preserved in variousarthropods (e.g. Antrimpos and Mesolimulus) andcephalopods (Leptotheuthis) from the same strata andlocality. In the case of the coleoid mandibles, eitheronly carbon remains from the original organic matteror it was replaced by clay minerals.
All isolated specimens of the upper beaks weredeposited lying on one side (except for SMNS 64923which was deposited obliquely; Figs 8G, H, 9I), whichprobably happened because both length and heightclearly exceeded the width of the upper beaks. Thus,this mode of embedding can best be explained by theirlaterally compressed morphology. The upper beaksprobably sank down through the water with theirrounded dorsum or the rostrum facing downwards.Since these two orientations would not have beenstable when a specimen was lying on the sediment, thebeaks fell onto one side after the rostrum or the hoodfirst touched the sediment surface.
It is more difficult to explain the orientation of mostof the lower beaks. Apparently, their high and broadexternal lamella offered enough stability, so that theyrested on it when they were embedded. For the lowerbeak of Germanonautilus, a Triassic nautiloid,Mundlos (1973) showed that the stable position in anenvironment with bottom currents is that with thedorsum facing downwards. In this case, the heavilycalcified conchorhynch served as an anchor, supportedby the two flexible wings of chitinous material.
A slightly differing biostratinomy has been recon-structed for the upper beak of Germanonautilus. Inthis nautiloid, the anterior fold of the lateral wings ofchitinous material served as an anchor, supported bythe posterior end of the internal lamella (Klug 2001).During compaction, the anterior parts of the wingswere stretched out anteriorly. A similar preservationwas found in one specimen of Trachyteuthis (SMNS65349; Figs 6, 9B), i.e. the internal lamella is flattenedand the wings of the external lamella are extendedanteriorly. All but one other specimen (SMNS 65344a;Figs 4A, B, 7A, B) lack the internal lamella. Theseprobably rested on the anterior side of the externallamella with an upright internal lamella. In that case,the internal lamella would have been preserved in adifferent layer or might have been eroded due to thelonger exposure to seawater. In some cases, the innerlamella might still be preserved in the counterpart slabor it was destroyed during preparation.
The fact that coleoid beaks of the quality of theNusplingen localities have not yet been discovered atSolnhofen can be explained by the presence of a higheramount of organic matter in the sediment in theformer (Bantel et al. 1999; Dietl & Schweigert 2001),which is evident from the darker color of some hori-zons of the Nusplingen Lithographic Limestone. Mostof the fossil beak elements are preserved as a thincarbon coating. It is darkest in the originally tannedareas, predominantly in the anterior part of theexternal lamellae, especially near the rostral tip (e.g.Figs 4D, F, 5, 7). At the rostral tip, the chitinousmaterial probably had to be strongest and therefore, itspreservation potential was the highest. This is cor-roborated by some remains of upper beaks that do notdisplay much more than the rostral tip and the ante-rior parts of the external lamella (Figs 8E, F, J, K).These interpretations also coincide with the results ofthe experiments by Kear et al. (1995).
Some of the lower beaks also display a secondarycoating of limonite. This mineralization probablyhappened later in diagenesis. In specimen SMNS65324 (Figs 5C, D), the limonitization is largelyrestricted to the extent of the primarily tanned portionof the external lamella.
Possible phylogenetic implications
Since fossil coleoid beaks are rare, they have not yetbeen included in phylogenetic analyses that comprisefossil taxa. Consequently, these new specimens ofcoleoid beaks (some of which are preserved in situ)encouraged us to discuss the rather obscure phylo-genetic relations of coleoids and the indications fromthe fossil beaks.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 185
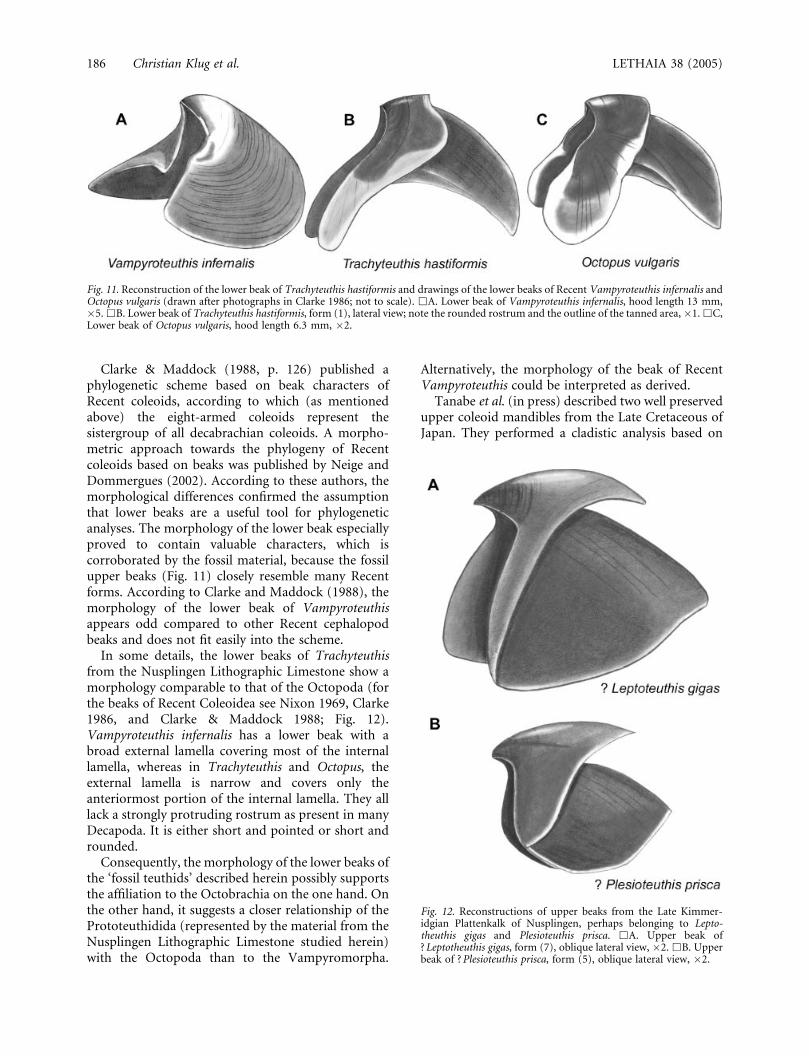
Clarke & Maddock (1988, p. 126) published aphylogenetic scheme based on beak characters ofRecent coleoids, according to which (as mentionedabove) the eight-armed coleoids represent thesistergroup of all decabrachian coleoids. A morpho-metric approach towards the phylogeny of Recentcoleoids based on beaks was published by Neige andDommergues (2002). According to these authors, themorphological differences confirmed the assumptionthat lower beaks are a useful tool for phylogeneticanalyses. The morphology of the lower beak especiallyproved to contain valuable characters, which iscorroborated by the fossil material, because the fossilupper beaks (Fig. 11) closely resemble many Recentforms. According to Clarke and Maddock (1988), themorphology of the lower beak of Vampyroteuthisappears odd compared to other Recent cephalopodbeaks and does not fit easily into the scheme.
In some details, the lower beaks of Trachyteuthisfrom the Nusplingen Lithographic Limestone show amorphology comparable to that of the Octopoda (forthe beaks of Recent Coleoidea see Nixon 1969, Clarke1986, and Clarke & Maddock 1988; Fig. 12).Vampyroteuthis infernalis has a lower beak with abroad external lamella covering most of the internallamella, whereas in Trachyteuthis and Octopus, theexternal lamella is narrow and covers only theanteriormost portion of the internal lamella. They alllack a strongly protruding rostrum as present in manyDecapoda. It is either short and pointed or short androunded.
Consequently, the morphology of the lower beaks ofthe ‘fossil teuthids’ described herein possibly supportsthe affiliation to the Octobrachia on the one hand. Onthe other hand, it suggests a closer relationship of thePrototeuthidida (represented by the material from theNusplingen Lithographic Limestone studied herein)with the Octopoda than to the Vampyromorpha.
Alternatively, the morphology of the beak of RecentVampyroteuthis could be interpreted as derived.
Tanabe et al. (in press) described two well preservedupper coleoid mandibles from the Late Cretaceous ofJapan. They performed a cladistic analysis based on
Fig. 11. Reconstruction of the lower beak of Trachyteuthis hastiformis and drawings of the lower beaks of Recent Vampyroteuthis infernalis andOctopus vulgaris (drawn after photographs in Clarke 1986; not to scale). &A. Lower beak of Vampyroteuthis infernalis, hood length 13 mm,�5. &B. Lower beak of Trachyteuthis hastiformis, form (1), lateral view; note the rounded rostrum and the outline of the tanned area, �1. &C,Lower beak of Octopus vulgaris, hood length 6.3 mm, �2.
Fig. 12. Reconstructions of upper beaks from the Late Kimmer-idgian Plattenkalk of Nusplingen, perhaps belonging to Lepto-theuthis gigas and Plesioteuthis prisca. &A. Upper beak of? Leptotheuthis gigas, form (7), oblique lateral view, �2. &B. Upperbeak of ? Plesioteuthis prisca, form (5), oblique lateral view, �2.
186 Christian Klug et al. LETHAIA 38 (2005)

the upper mandibles of 22 Recent and this fossilspecies, i.e. 23 species. In this analysis, they used sixcharacters (A: rostrum shape, B: jaw angle, C: shouldershape, D: shape of posterior margin of wing, E: shapeof posterior margin of lateral wall, F: presence orabsence of a calcareous tip). In the resulting clado-gram, the upper mandible of this new taxon fromJapan has a position within the Oegopsina.
The material presented in our study is insufficientlypreserved to decide on the precise state of some ofthe characters used by Tanabe et al. (in press).Considering the jaw angle, most of the upper man-dibles excavated in the Nusplingen quarry yield valuesaround 90� or slightly below. This is interpreted asindicating a jaw angle of less than 90�, but this could aswell be an artifact of syn-compactional deformation.Additionally, the posterior margin of the lateral wall ofthe internal lamella is never perfectly preserved. This isthe reason, why a posterior concavity of the posteriormargin of the lateral wall is visible only in the speci-men SMNS 64328 (Figs 8L, 9H). Presuming that therostrum angle is obtuse (<90�) and that the posteriormargin of the lateral wall is medially concave, thecharacter states of the Nusplingen specimens would be
identical with those of the material from the JapaneseCretaceous, thus indicating a close relationship torecent decapod coleoids (Oegopsina). It still has to bekept in mind, that: (1) the identification of some ofthese character states are rather uncertain, (2) thematerial is strongly distorted, and (3) this cladisticanalysis was based exclusively on upper beaks.
According to W. Riegraf (personal communication,2003), the absence of tentacles among the ‘fossilteuthids’ can be explained by the rapid disintegrationof these organs after the animal’s death. He hypo-thesizes that many ‘fossil teuthids’ actually had tenarms. Additionally, he suggests that the upper beakswhich we here assign to Plesioteuthis with some reser-vation show similarities to the upper beaks of Recentdecapods (compare also Mangold & Fioroni 1966).Nevertheless, the reconstruction of Trachyteuthis(Fig. 13) is based on the assumption of a closerelationship to Vampyroteuthis.
The coleoid beak certainly does not represent themost important character and, additionally, futurefinds of the beaks of other fossil vampyromorphs mayreveal a different morphology and thus will enable totest phylogenetic models such as those of Engeser
Fig. 13. &A. A reconstruction of Trachyteuthis, presuming a close relationship to Vampyroteuthis, based on the presence of four fins and cirriand the (so far) lacking tentacles. Most of the morphology is taken from the reconstruction by Bandel & Leich (1986), but here it is shown withsmall fins according to the specimens published by Donovan (1995) and Donovan et al. (2002), �0.25. &B. For comparison, a drawing ofVampyroteuthis (after a photograph by R. E. Young, 1998 from the tree of life web page, http://tolweb.org/) is shown, �0.7.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 187

(1990a) or Haas (2002). Since coleoid beaks areexceedingly rare and poorly understood, this materialis still insufficient to help establish a coherent phylo-genetic scheme for the Coleoidea.
Conclusions
The beak forms (1) and (6) can be assigned toTrachyteuthis hastiformis with some certainty sincethey were found in association with the characteristicgladii, roughly in the original place of the buccal mass.The affiliation of isolated lower beaks of the same form(1) is corroborated by the facts that both these lowerbeaks and the gladii of Trachyteuthis are similarlycommon in the Nusplingen Lithographic Limestone.Additionally, the mode of preservation, the moderatesize and the morphology of the external lamellasupport this hypothesis. The external lamella of thislower beak is U-shaped and exhibits a smoothlycurved and only slightly protruding rostrum. In onespecimen, the beaks are still articulated with oneanother (Figs 4A, B, 7A, B, 11B).
Forms (2) and (5) probably are related becauseone specimen of each form was found very close tothe other. These forms were tentatively assigned toPlesioteuthis prisca, because form (5) is the commonestupper beak (Figs 8, 12B) but appears to differ fromform (6) (Trachyteuthis), which has not yet beenfound in isolated specimens. The lower beak ofPlesioteuthis had a more pointed rostrum.
Because of its large size and relative scarcity, form(7) is tentatively assigned to the largest and rarestcoleoid species so far found in the Nusplingen Litho-graphic Limestone, Leptotheuthis gigas. This upperbeak is higher than the others and it differs from thatof Plesioteuthis and Trachyteuthis in the more elongatethe wing of external lamella (Figs 8G, H, 9I, 12A). Thelower beaks of form (3) and (4) cannot yet be assignedto any one of these three taxa. Future discoveries ofadditional complete specimens or gladii associatedwith beaks might shed more light on the taxonomicaffiliation of these forms.
The comparison of the lower beaks showed thatforms (1) and (4) somewhat resemble the outerlamella of Recent Octopus. In contrast, the externallamella of the lower beak of Vampyroteuthis is muchbroader and covers the internal lamella (Fig. 11). Onthe one hand, the affiliation of Trachyteuthis and thePrototeuthidida in general to the Octobrachia seemssupported. On the other hand, some of the fossilupper beaks display similarities to decapod beaks.Whether or not the phylogenetic relationship ofrepresentatives of the ‘fossil teuthids’ is closer tothe Vampyromorpha, to the Octopoda or to the
Decapoda, needs to be clarified in future studies. Thenew data on these coleoid beaks, however, may thenhelp in clarifying these poorly understood relation-ships.
The presence of a higher proportion of organiccomponents in the sediments of the NusplingenLithographic limestone probably enabled the preser-vation of these rarely preserved structures thatprimarily consisted of chitinous material. Because oftheir proportions, the lower beaks of Trachyteuthis andthe two lower beaks of uncertain affiliation must havesettled on the external lamella on the sediment andthus are only rarely preserved with the internallamella. The upper beaks of Plesioteuthis and ?Lepto-theuthis were high and long but narrow and thereforewere embedded lying on one flank.
Acknowledgements. – Markus Rieter and Martin Kapitzke (Stutt-gart) both did an excellent job on the preparation of the specimens.Their careful work is gratefully acknowledged. Rolf Hugger(Albstadt) recovered many important specimens used in this study.We thank Alexander Fels (Stuttgart) who performed an EDAX-analysis of the beak material for us. Desmond T. Donovan(London), Susan Turner (Kenmore, Queensland) and WolfgangRiegraf (Munster) kindly proofread the manuscript and offeredsubstantial help in improving its structure and content as well aslanguage. Sigurd von Boletzky (Banyuls-sur-mer) and James W.Haggart (Vancouver) mindfully reviewed the manuscript andcontributed some valuable ideas. Additionally, we thank MalcolmR. Clarke (Plymouth, UK) for sharing his valuable opinion on someaspects of these cephalopod beaks. Kazushige Tanabe (Tokyo)generously provided access to unpublished data on Cretaceouscoleoid beaks from Japan. We are indebted to Wolfgang Gerber(Tubingen) who produced most of the photographs.
References
Allison, P.A. 1988a: Konservat-Lagerstatten: cause and classifica-tion. Paleobiology 14, 331–344.
Allison, P.A. 1988b: Phosphatised soft-bodied squids from theJurassic Oxford Clay. Lethaia 21, 403–410.
Bandel, K. & Leich, H. 1986: Jurassic Vampyromorpha(dibranchiate cephalopods). Neues Jahrbuch fur Geologie undPalaontologie Monatshefte 1986 (3), 129–148.
Bantel, G., Schweigert, G., Nose, M. & Schulz, H.-M. 1999:Mikrofazies, Mikro- und Nannofossilien aus dem NusplingerPlattenkalk (Ober-Kimmeridgium, Schwabische Alb). Stuttgar-ter Beitrage zur Naturkunde, Serie B 279, pp 55.
Boletzky, S. 1999: Breve mise au point sur la classification descephalopodes actuels. Bulletin de la Societe zoologique de France124 (3), 271–278.
Briggs, D.E.G., Kear, A.J., Martill, D.M. & Wilby, P.R. 1993:Phosphatization of soft-tissue in experiments and fossils. Journalof the Geological Society, London 150, 1035–1038.
Briggs, D.E.G. & Wilby, P.R. 1996: The role of the calciumcarbonate-calcium phosphate switch in the mineralization ofsoft-bodied fossils. Journal of the Geological Society, London 153,665–668.
Bronn, H.G. 1859: Nachtrag uber die Triasfauna von Raibl. NeuesJahrbuch fur Mineralogie, Geognosie, Geologie und Petre-faktenkunde 1859, 39–45.
Clarke, M.R. 1986: A Handbook for the Identification of CephalopodBeaks. Oxford University Press, Oxford, xiv þ 273 pp.
Clarke, M.R. & Maddock, L. 1988: Beaks of living coleoid Cephalo-poda. In Clarke, M.R. & Trueman, R. (eds): Paleontology andNeontology of Cephalopods. 123–131. Academic Press, San Diego.
188 Christian Klug et al. LETHAIA 38 (2005)

Dietl, G., Dietl, O., Kapitzke, M., Rieter, M., Schweigert, G., Ilg, A.& Hugger, R. 1997: Der Nusplinger Plattenkalk (Weißer Jura) –Grabungskampagne 1996. Jahreshefte der Gesellschaft furNaturkunde in Wurttemberg 153, 185–203.
Dietl, G., Kapitzke, M. & Rieter, M. 1995: Neue Grabungen imNusplinger Plattenkalk (Weißer Jura zeta) der SchwabischenAlb. Fossilien 1995, 170–174.
Dietl, G. & Schweigert, G. 1999a: Ein Nautilus mit in situ liegen-dem, vollstandigem Kieferapparat aus dem Nusplinger Platten-kalk (Oberjura, SW-Deutschland). Neues Jahrbuch fur Geologieund Palaontologie Abhandlungen 211, 75–87.
Dietl, G. & Schweigert, G. 1999b: Der Nusplinger Plattenkalk undseine Fossilien (Weißer Jura, Ober-Kimmeridgium) (ExkursionM am 10. April 1999). Jahresberichte und Mitteilungen desoberrheinischen geologischen Vereins, neue Folge 81, 157–171.
Dietl, G. & Schweigert, G. 2001: Im Reich der Meerengel. 144 pp. F.Pfeil, Munchen.
Dietl, G. & Schweigert, G. 2004: The Nusplingen LithographicLimestone – a ‘fossil lagerstaette’ of Late Kimmeridgian agefrom the Swabian Alb (Germany). Rivista Italiana di Paleonto-logia e Stratigrafia 110, 303–309.
Donovan, D.T. 1983: Mastigophora Owen 1856, a little knowngenus of Jurassic coleoids. Neues Jahrbuch fur Geologie undPalaontologie Abhandlungen 165, 484–495.
Donovan, D.T. 1995: A specimen of Trachyteuthis (Coleoidea) withfins from the Upper Jurassic of Solnhofen (Bavaria). StuttgarterBeitrage zur Naturkunde, Serie B 235, 8 pp.
Donovan, D.T. & Crane, M. 1992: The type material of the Jurassiccephalopod Belemnotheutis. Palaeontology 35, 273–296.
Donovan, D.T., Doguzhaeva, L.A. & Mutvei, H. 2002: Trachyteuthis(Upper Jurassic): two pairs of fins and their phylogeneticsignificance. Berliner palaobiologische Abhandlungen 1, 43–46.
Doyle, P., Donovan, D.T. & Nixon, M. 1994: Phylogeny andsystematics of the Coleoidea. University of Kansas PaleontologicalContributions, New Series 5, 15 pp.
Dzik, J. 1986: Uncalcified cephalopod jaws from the MiddleJurassic of Poland. Neues Jahrbuch fur Geologie und PalaontologieMonatshefte 1986, 405–417.
Engeser, T. 1990a: Phylogeny of the fossil coleoid Cephalopoda(Mollusca). In Schudack, M.E. (ed.): Beitrage zur Palaontologie.Berliner geowissenschaftliche Abhandlungen, Reihe A: Geologieund Palaontologie 124, 123–191.
Engeser, T. 1990b: Major events in cephalopod evolution. InTaylor, P.D. & Larwood, G.P. (eds): Major Evolutionary Radia-tions. The Systematics Association Special Volume 42, 119–138.
Engeser, T. & Keupp, H. 1997: Zwei neue Gattungen und eine neueArt von vampyromorphen Tintenfischen (Coleoidea, Cephalo-poda) aus dem Untertithonium von Eichstatt. Archaeopteryx 15,47–58.
Engeser, T. & Keupp, H. 1999: Zwei neue vampyromorpheTintenfische (Coleoidea, Cephalopoda) aus dem oberjur-assischen Solnhofener Plattenkalk von Eichstatt. Archaeopteryx17, 21–32.
Engeser, T. & Reitner, J. 1981: Beitrage zur Systematik vonphragmokontragenden Coleoiden aus dem Untertithonium(Malm zeta, ‘Solnhofener Plattenkalk’) von Solnhofen undEichstatt (Bayern). Neues Jahrbuch fur Geologie und PalaontologieMonatshefte 1981 (9), 527–545.
Engeser, T. & Reitner, J. 1986: Coleoidenreste aus der Oberkreidedes Libanon im Staatlichen Museum fur Naturkunde in Stutt-gart. Stuttgarter Beitrage zur Naturkunde, Serie B 124, 17 pp.
Engeser, T. & Reitner, J. 1992: Ein neues Exemplar von Belemno-teuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda)aus dem Solnhofener Plattenkalk (Untertithonium) vonWintershof, Bayern. Archaeopteryx 10, 13–17.
Etter, W. 2002: La Voulte-sur-Rhone: exquisite cephalopodpreservation. In Bottjer, D.J., Etter, W., Hagadorn, J.W. & Tang,C.M. (eds): Exceptional Fossil Preservation. 293–305. ColumbiaUniversity Press, New York.
Etter, W. & Tang, C.M. 2002: Posidonia Shale: Germany’s Jurassicmarine park. In Bottjer, D.J., Etter, W., Hagadorn, J.W. & Tang,C. (eds): Exceptional Fossil Preservation. 265–291. ColumbiaUniversity Press, New York.
Fahrion, H. 1937: Ein Beitrag zur Entstehung der NusplingerKalkschiefer auf Grund neuer Fossilfunde. Jahresberichte undMitteilungen des oberrheinischen geologischen Vereins, Neue Folge26, 60–65.
Fraas, O. 1849: Beobachtungen uber die Gestaltung des oberenweissen Jura in Schwaben und Franken. 26. Versammlungdeutscher Naturforscher und Arzte (18–24 Sept. 1849), 66–67.
Fraas, O. 1854: Nusplingen – ein Seitenstuck zu Solnhofen.Correspondenzblatt des zoologisch-mineralogischen VereinsRegensburg 8, 71–76.
Fraas, O. 1855: Beitrage zum obersten weissen Jura in Schwaben.Jahreshefte des Vereins fur vaterlandische Naturkunde in Wurt-temberg 11, 76–107.
Frickhinger, K.A. 1994: Die Fossilien von Solnhofen. 336 pp. Gold-schneck, Weinstadt.
Frickhinger, K.A. 1999: Die Fossilien von Solnhofen, 2. 190 pp.Goldschneck, Weinstadt.
Frohlich, F., Mayrat, A., Riou, B. & Secretan, S. 1992: Structuresretiniennes phosphatees dans l’oeil geant de Dollocaris, uncrustace fossile. Annales de Paleontologie 78, 193–204.
Guerin-Franiatte, S. & Gouspy, C. 1993: Decouverte de Cephalo-podes Teuthides (Coleoidea) dans le Lias superieur deHaute-Marne, France. In Elmi, S., Mangold, C. & Almeras, Y.(eds): 3eme symposium international sur les Cephalopodes actuelset fossiles; symposium F. Roman Geobios, Memoire Special 15,181–189.
Haas, W. 2002: The evolutionary history of the eight-armedColeoidea. Abhandlungen der geologischen Bundesanstalt Wien57, 341–351.
Harzhauser, M. 1999: Filling a gap – beaks and hooks ofCenozoic coleoids (Cephalopoda). Annalen des Naturhistori-schen Museums in Wien 101A, 123–135.
Hewitt, R.A. & Wignall, P.B. 1988: Structure and phylogeneticsignificance of Trachyteuthis (Coleoidea) from the KimmeridgeClay of England. Proceedings of the Yorkshire Geological Society47, 149–153.
Kear, A.J., Briggs, D.E.G. & Donovan, D.T. 1995: Decay andfossilization of non-mineralized tissue in coleoid cephalopods.Palaeontology 38, 105–131.
Kluessendorf, J. & Doyle, P. 2000: Pohlsepia mazonensis, an early‘octopus’ from the Carboniferous of Illinois, USA. Palaeontology43, 919–926.
Klug, C. 2001: Functional morphology and taphonomy of nautiloidbeaks from the Middle Triassic of Southwest Germany. ActaPalaeontologica Polonica 46, 43–68.
Lossel, P. 1947: A propos des machoires de cephalopodes fossilesdites ‘en bec de perroquet’. Compte Rendu Sommaire des Seancesde la Societe Geologique de France 14, 280–282.
Mangold, K.M. & Fioroni, P. 1966: Morphologie et biometrie desmandibules de quelques cephalopodes mediterraneens. Vie etMilieu (A) 17 (3-A), 1139–1196.
Martill, D.M. 1987: A taphonomic and diagenetic case study of apartially articulated ichthyosaur. Palaeontology 30, 543–556.
Mehl, J. 1990: Fossilerhaltung von Kiemen bei Plesioteuthisprisca (Ruppell 1829) (Vampyromorpha, Cephalopoda) ausuntertithonen Plattenkalken der Altmuhlalb. Archaeopteryx8, 77–91.
Mojsisovics, E.V. 1882: Die Cephalopoden der mediterranenTriasprovinz. Abhandlungen der kaiserlich-koniglichen Geo-logischen Reichsanstalt 10, 322 pp.
Mojsisovics, E.V. 1902: Das Gebirge um Hallstatt. 1. Abtheilung.Die Cephalopoden der Hallstatter Kalke. Abhandlungen derkaiserlich-koniglichen Geologischen Reichsanstalt 1, 1. Lfg. 1873)82 pp. pls 1–32; 2. Lfg. (1875) 83–174, pls 33–70; 3. Lfg. (1902)175–356, pls 1–23. Lechner, Wien.
Mundlos, R. 1973: Ist Conchorhynchus ein Ceratiten-Kiefer?Palaontologische Zeitschrift 47, 156–162.
Naef, A. 1922: Die fossilen Tintenfische. 322 pp. Gustav Fischer,Jena.
Neige, P. & Dommergues, J.-L. 2002: Disparity of beaks andstatoliths of some coleoids: a morphometric approach todepict shape differentiation. Abhandlungen der geologischenBundesanstalt Wien 57, 393–399.
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 189

Nixon, M. 1969: Growth of the beak and radula of Octopus vulgaris.Journal of Zoology 159, 363–379.
Pinna, G. 1972: Rinvenimento di un raro cefalopode coleoideo nelgiacimento sinemuriano di Osteno in Lombardia. Atti dellaSocieta Italiana di Scienze Naturali e del Museo Civico di StoriaNaturale de Milano 113, 141–149.
Pinna, G. 1985: Exceptional preservation in the Jurassic of Osteno.Philosophical Transactions of the Royal Society of London 311B,170–180.
Quenstedt, F.A. 1843: Das Flozgebirge Wurtembergs. Mit besondererRucksicht auf den Jura. 558 pp. Laupp, Tubingen.
Quenstedt, F.A. 1855: 1. Petrefaktensammler Schwabens. 2. UberPterodactylus suevicus im lithographischen Schiefer Wurttembergs.52 pp. Laupp, Tubingen.
Quenstedt, F.A. 1856–1857: Der Jura. 842 pp. Laupp, Tubingen.Reitner, J. & Engeser, T. 1982: Zwei neue Coleoidea-Arten aus
dem Posidonienschiefer (Untertoarcium) aus der Gegend vonHolzmaden (Baden-Wurttemberg). Stuttgarter Beitrage zurNaturkunde, Serie B 84, 19 pp.
Reitner, J. & Urlichs, M. 1983: Echte Weichteilbelemniten aus demUntertoarcium (Posidonienschiefer) Sudwestdeutschlands.Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen165, 450–465.
Rieber, H. 1970: Phragmoteuthis? ticinensis n. sp., ein Coleoidea-Rest aus der Grenzbitumenzone (mittlere Trias) des MonteSan Giorgio (Kt. Tessin, Schweiz). Palaontologische Zeitschrift 44,32–40.
Rieber, H. 1973: Cephalopoden aus der Grenzbitumenzone(Mittlere Trias) des Monte San Giorgio (Kanton Tessin,Schweiz). Schweizerische Palaontologische Abhandlungen 93, 96pp.
Riegraf, W. 1982: New Coleoidea from the Lower Jurassic ofSouthwest Germany. Neues Jahrbuch fur Geologie und Palaonto-logie Monatshefte 1982 (2), 91–97.
Riegraf, W. 1983: Uber ‘Sueviteuthis’ zellensis Reitner & Engeser,1982 (Coleoidea) aus dem Untertoarcium von Sudwest-deutschland. Neues Jahrbuch fur Geologie und PalaontologieAbhandlungen 165, 502–507.
Riegraf, W. & Hauff, R. 1983: Belemniten mit Weichkorper,Fangarmen und Gladius aus dem Untertoarcium (Posido-nienschiefer) und Unteraalenium (Opalinuston) Sudwest-deutschlands. Neues Jahrbuch fur Geologie und PalaontologieAbhandlungen 165, 466–483.
Riegraf, W., Janssen, N.M.M. & Schmitt-Riegraf, C. 1998: Cephalo-poda dibranchiata fossiles (Coleoidea) II. In Westphal, F. (ed.):Fossilium Catalogus; I, Animalia. 135, 512 pp. BackhuysPublishers, Leiden.
Riegraf, W. & Schmitt-Riegraf, C. 1995: Mandibula fossilesammonitorum et nautilorum (Rhyncholithi et rhynchoteuthes,excl. aptychi et anaptychi). In Westphal, F. (ed.): FossiliumCatalogus. I. Animalia. 134, 219 pp. Kugler, Amsterdam &New York.
Riegraf, W., Werner, G. & Lorcher, F. 1984: Der Posidonienschiefer.Biostratigraphie, Fauna und Fazies des sudwestdeutschen Unter-toarciums (Lias epsilon). 195 pp. F. Enke, Stuttgart.
Schellenberg 2002: Mazon Creek: preservation in LatePaleozoic deltaic and marginal marine environments. In Bottjer,D.J., Etter, W., Hagadorn, J.W. & Tang, C.M. (eds): ExceptionalFossil Preservation. 185–204. Columbia University Press,New York.
Schweigert, G. 1997: Bibliographie des Nusplinger Plattenkalks(Oberjura, Schwabische Alb). Profil 11, 341–349; Stuttgart.
Schweigert, G. 1998: Die Ammonitenfauna des NusplingerPlattenkalks (Ober-Kimmeridgium, Beckeri-Zone, Ulmense-Subzone, Schwabische Alb). Stuttgarter Beitrage zur Naturkunde,Serie B 267, 61 pp.
Schweigert, G. & Dietl, G. 1999: Zur Erhaltung und Einbettung vonAmmoniten im Nusplinger Plattenkalk (Oberjura, Sudwest-deutschland). Stuttgarter Beitrage zur Naturkunde, Serie B 272,31 pp.
Schweigert, G. & Dietl, G. 2001: Die Kieferelemente von Physo-doceras (Ammonitina, Aspidoceratidae) im Nusplinger Platten-kalk (Oberjura, Schwabische Alb). Berliner geowissenschaftlicheAbhandlungen E36, 131–143.
Schweigert, G., Krishna, J., Pandey, B. & Pathak, D.B. 1996: A newapproach to the correlation of the Upper Kimmeridgian BeckeriZone across the Tethyan sea. Neues Jahrbuch fur Geologie undPalaontologie Abhandlungen 202, 345–373.
Seilacher, A. 1970: Begriff und Bedeutung der Fossil-Lagerstatten.Neues Jahrbuch fur Geologie und Palaontologie Monatshefte 1970,34–39.
Tanabe, K., Hikida, Y. & Iba, Y. in press: Two Coleoid jaws fromthe Upper Cretaceous of Hokkaido, Japan. Journal of Paleon-tology 79, 1, ca. 15 pp.
Tang, C.M. 2002a: Osteno: Jurassic Preservation on the cellularlevel. In Bottjer, D.J., Etter, W., Hagadorn, J.W. & Tang, C.M.(eds): Exceptional Fossil Preservation. 251–264. ColumbiaUniversity Press, New York.
Tang, C.M. 2002b: Oxford clay: England’s Jurassic marine park. InBottjer, D.J., Etter, W., Hagadorn, J.W. & Tang, C.M. (eds)Exceptional Fossil Preservation. 307–325. Columbia UniversityPress, New York.
Westphal, F. 1992: Nusplingen. Entstehung einer Fossillagerstatteim Oberjura-Plattenkalk. Kaupia 1, 63–70.
Wilby, M.A., Briggs, D.E.G. & Riou, B. 1995: La Voulte-sur-Rhone (Callovian): fossilized soft-tissues from a ‘toxicsludge’. The Palaeontology Newsletter (the PalaeontologicalAssociation) 28, 25.
Wilby, M.A., Briggs, D.E.G. & Riou, B. 1996: Mineralization ofsoft-bodied invertebrates in a Jurassic metalliferous deposit.Geology 24, 847–850.
Young, R.E., Vecchione, M. & Donovan, D.T. 1998: The evolutionof cephalopods and their present biodiversity and ecology. SouthAfrican Journal of Marine Science 20, 393–420.
190 Christian Klug et al. LETHAIA 38 (2005)
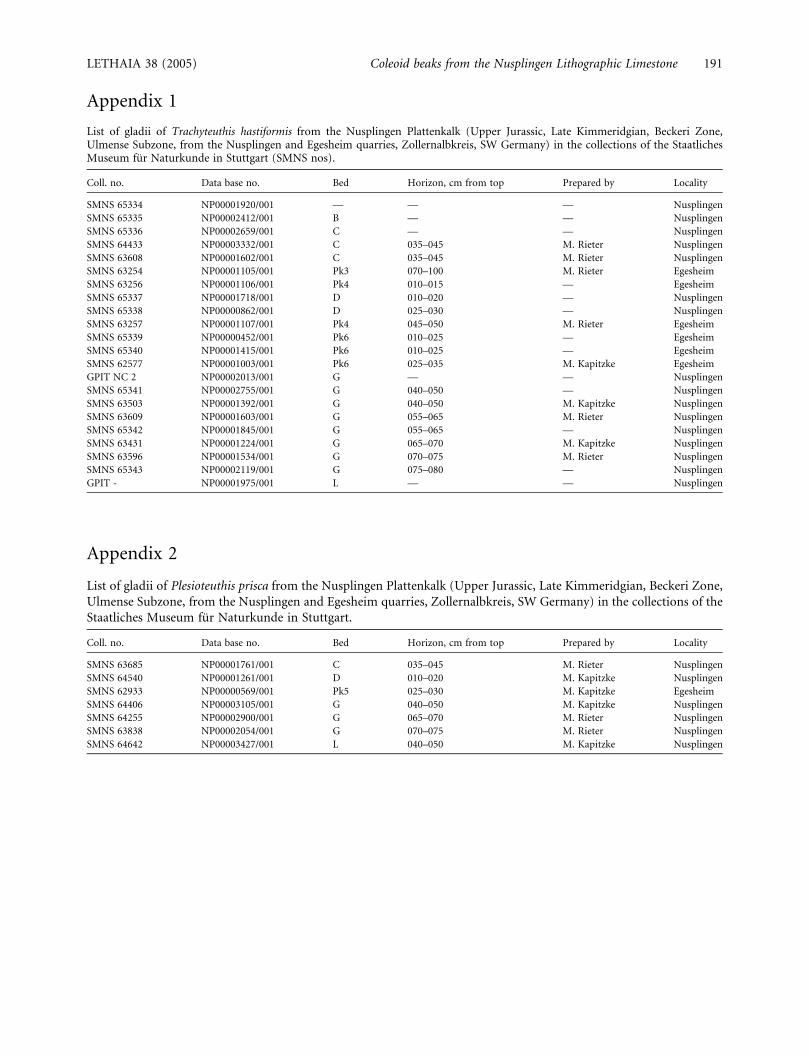
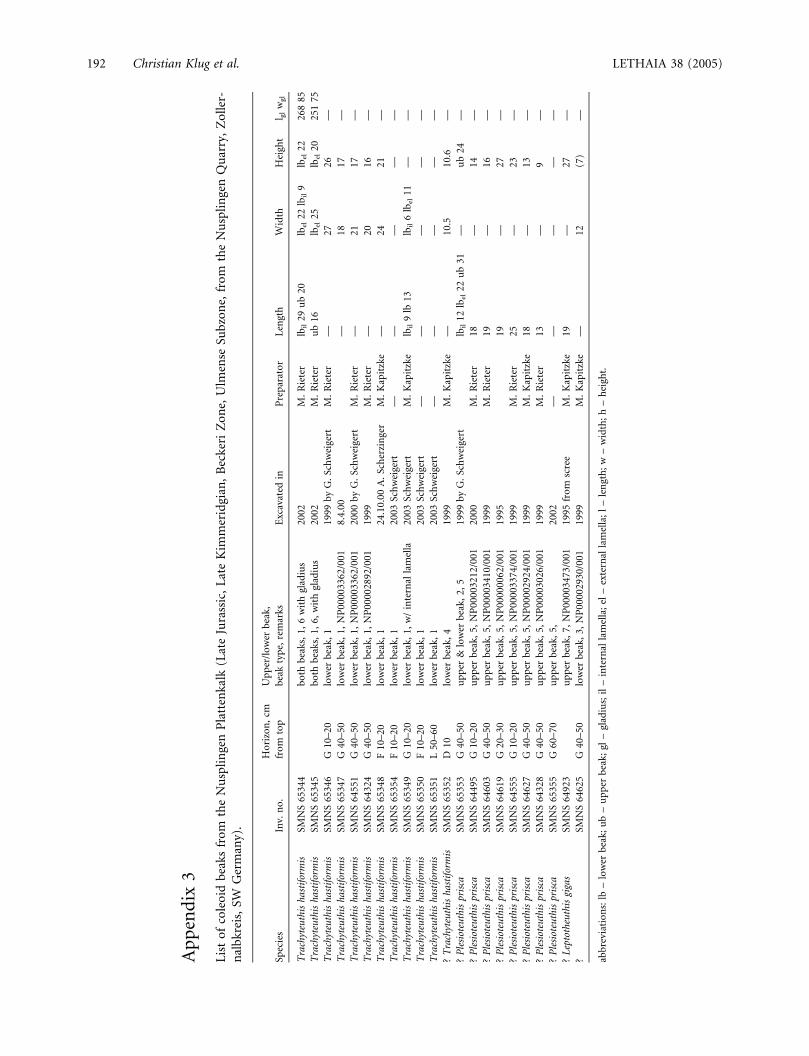
Appendix 1
List of gladii of Trachyteuthis hastiformis from the Nusplingen Plattenkalk (Upper Jurassic, Late Kimmeridgian, Beckeri Zone,Ulmense Subzone, from the Nusplingen and Egesheim quarries, Zollernalbkreis, SW Germany) in the collections of the StaatlichesMuseum fur Naturkunde in Stuttgart (SMNS nos).
Coll. no. Data base no. Bed Horizon, cm from top Prepared by Locality
SMNS 65334 NP00001920/001 — — — Nusplingen
SMNS 65335 NP00002412/001 B — — Nusplingen
SMNS 65336 NP00002659/001 C — — Nusplingen
SMNS 64433 NP00003332/001 C 035–045 M. Rieter Nusplingen
SMNS 63608 NP00001602/001 C 035–045 M. Rieter Nusplingen
SMNS 63254 NP00001105/001 Pk3 070–100 M. Rieter Egesheim
SMNS 63256 NP00001106/001 Pk4 010–015 — Egesheim
SMNS 65337 NP00001718/001 D 010–020 — Nusplingen
SMNS 65338 NP00000862/001 D 025–030 — Nusplingen
SMNS 63257 NP00001107/001 Pk4 045–050 M. Rieter Egesheim
SMNS 65339 NP00000452/001 Pk6 010–025 — Egesheim
SMNS 65340 NP00001415/001 Pk6 010–025 — Egesheim
SMNS 62577 NP00001003/001 Pk6 025–035 M. Kapitzke Egesheim
GPIT NC 2 NP00002013/001 G — — Nusplingen
SMNS 65341 NP00002755/001 G 040–050 — Nusplingen
SMNS 63503 NP00001392/001 G 040–050 M. Kapitzke Nusplingen
SMNS 63609 NP00001603/001 G 055–065 M. Rieter Nusplingen
SMNS 65342 NP00001845/001 G 055–065 — Nusplingen
SMNS 63431 NP00001224/001 G 065–070 M. Kapitzke Nusplingen
SMNS 63596 NP00001534/001 G 070–075 M. Rieter Nusplingen
SMNS 65343 NP00002119/001 G 075–080 — Nusplingen
GPIT - NP00001975/001 L — — Nusplingen
Appendix 2
List of gladii of Plesioteuthis prisca from the Nusplingen Plattenkalk (Upper Jurassic, Late Kimmeridgian, Beckeri Zone,
Ulmense Subzone, from the Nusplingen and Egesheim quarries, Zollernalbkreis, SW Germany) in the collections of the
Staatliches Museum fur Naturkunde in Stuttgart.
Coll. no. Data base no. Bed Horizon, cm from top Prepared by Locality
SMNS 63685 NP00001761/001 C 035–045 M. Rieter Nusplingen
SMNS 64540 NP00001261/001 D 010–020 M. Kapitzke Nusplingen
SMNS 62933 NP00000569/001 Pk5 025–030 M. Kapitzke Egesheim
SMNS 64406 NP00003105/001 G 040–050 M. Kapitzke Nusplingen
SMNS 64255 NP00002900/001 G 065–070 M. Rieter Nusplingen
SMNS 63838 NP00002054/001 G 070–075 M. Rieter Nusplingen
SMNS 64642 NP00003427/001 L 040–050 M. Kapitzke Nusplingen
LETHAIA 38 (2005) Coleoid beaks from the Nusplingen Lithographic Limestone 191
Ap
pen
dix
3
Lis
to
fco
leo
idb
eak
sfr
om
the
Nu
spli
nge
nP
latt
enk
alk
(Lat
eJu
rass
ic,
Lat
eK
imm
erid
gian
,B
eck
eri
Zo
ne,
Ulm
ense
Su
bzo
ne,
fro
mth
eN
usp
lin
gen
Qu
arry
,Z
oll
er-
nal
bk
reis
,S
WG
erm
any)
.
Sp
ecie
sIn
v.n
o.
Ho
rizo
n,
cm
fro
mto
p
Up
per
/lo
wer
bea
k,
bea
kty
pe,
rem
ark
sE
xcav
ated
inP
rep
arat
or
Len
gth
Wid
thH
eigh
tl g
lw
gl
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
4b
oth
bea
ks,
1,
6w
ith
glad
ius
20
02
M.
Rie
ter
lbil
29
ub
20
lbel
22
lbil
9lb
el2
22
68
85
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
5b
oth
bea
ks,
1,
6,
wit
hgl
adiu
s2
00
2M
.R
iete
ru
b1
6lb
el2
5lb
el2
02
51
75
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
6G
10
–2
0lo
wer
bea
k,
11
99
9b
yG
.S
chw
eige
rtM
.R
iete
r—
27
26
—
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
7G
40
–5
0lo
wer
bea
k,
1,
NP
00
003
36
2/0
01
8.4
.00
—1
81
7—
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
64
55
1G
40
–5
0lo
wer
bea
k,
1,
NP
00
003
36
2/0
01
20
00
by
G.
Sch
wei
gert
M.
Rie
ter
—2
11
7—
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
64
32
4G
40
–5
0lo
wer
bea
k,
1,
NP
00
002
89
2/0
01
19
99
M.
Rie
ter
—2
01
6—
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
8F
10
–2
0lo
wer
bea
k,
12
4.1
0.0
0A
.S
cher
zin
ger
M.
Kap
itzk
e—
24
21
—
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
35
4F
10
–2
0lo
wer
bea
k,
12
00
3S
chw
eige
rt—
——
——
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
34
9G
10
–2
0lo
wer
bea
k,
1,
w/
inte
rnal
lam
ella
20
03
Sch
wei
gert
M.
Kap
itzk
elb
il9
lb1
3lb
il6
lbel
11
——
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
35
0F
10
–2
0lo
wer
bea
k,
12
00
3S
chw
eige
rt—
——
——
Tra
chyt
euth
ish
ast
ifor
mis
SM
NS
65
35
1L
50
–6
0lo
wer
bea
k,
12
00
3S
chw
eige
rt—
——
——
?T
rach
yteu
this
ha
stif
orm
isS
MN
S6
53
52
D1
0lo
wer
bea
k,
41
99
9M
.K
apit
zke
—1
0.5
10
.6—
?P
lesi
oteu
this
pri
sca
SM
NS
65
35
3G
40
–5
0u
pp
er&
low
erb
eak
,2
,5
19
99
by
G.
Sch
wei
gert
lbil
12
lbel
22
ub
31
—u
b2
4—
?P
lesi
oteu
this
pri
sca
SM
NS
64
49
5G
10
–2
0u
pp
erb
eak
,5
,N
P0
000
32
12
/00
12
00
0M
.R
iete
r1
8—
14
—
?P
lesi
oteu
this
pri
sca
SM
NS
64
60
3G
40
–5
0u
pp
erb
eak
,5
,N
P0
000
34
10
/00
11
99
9M
.R
iete
r1
9—
16
—
?P
lesi
oteu
this
pri
sca
SM
NS
64
61
9G
20
–3
0u
pp
erb
eak
,5
,N
P0
000
00
62
/00
11
99
51
9—
27
—
?P
lesi
oteu
this
pri
sca
SM
NS
64
55
5G
10
–2
0u
pp
erb
eak
,5
,N
P0
000
33
74
/00
11
99
9M
.R
iete
r2
5—
23
—
?P
lesi
oteu
this
pri
sca
SM
NS
64
62
7G
40
–5
0u
pp
erb
eak
,5
,N
P0
000
29
24
/00
11
99
9M
.K
apit
zke
18
—1
3—
?P
lesi
oteu
this
pri
sca
SM
NS
64
32
8G
40
–5
0u
pp
erb
eak
,5
,N
P0
000
30
26
/00
11
99
9M
.R
iete
r1
3—
9—
?P
lesi
oteu
this
pri
sca
SM
NS
65
35
5G
60
–7
0u
pp
erb
eak
,5
,2
00
2—
——
——
?L
epto
theu
this
giga
sS
MN
S6
49
23
up
per
bea
k,
7,
NP
00
003
47
3/0
01
19
95
fro
msc
ree
M.
Kap
itzk
e1
9—
27
—
?S
MN
S6
46
25
G4
0–
50
low
erb
eak
,3
,N
P0
000
29
30
/00
11
99
9M
.K
apit
zke
—1
2(7
)—
abb
revi
atio
ns:
lb–
low
erb
eak
;u
b–
up
per
bea
k;
gl–
glad
ius;
il–
inte
rnal
lam
ella
;el
–ex
tern
alla
mel
la;
l–
len
gth
;w
–w
idth
;h
–h
eigh
t.
192 Christian Klug et al. LETHAIA 38 (2005)
Related Documents











