Quaternary Science Reviews 24 (2005) 1287–1301 Co-existence of scimitar-toothed cats, lions and hominins in the European Pleistocene. Implications of the post-cranial anatomy of Homotherium latidens (Owen) for comparative palaeoecology Mauricio Anto´n a , Angel Galobart b , Alan Turner c, a Departamento de Paleobiologia, Museo Nacional de Ciencias Naturales, C. Jose Gutierrez Abascal 2, 28006 Madrid, Spain b Institut de Paleontologia Dr Miquel Crusafont, C. Escola Industrial 23, 08201 Sabadell, Spain c School of Biological and Earth Sciences, Liverpool John Moores University, Liverpool L3 3AF, UK Received 3 February 2004; accepted 24 September 2004 Abstract Human occupants of Europe shared food resources with a number of larger Carnivora, and their coexistence with two lion-sized felids, the lion and the scimitar-toothed machairodont Homotherium latidens, poses intriguing paleoecological problems. We investigate the ecology of Homotherium latidens using an exceptional sample of postcrania from the Spanish Early Pleistocene site of Incarcal, making comparisons with modern cats and with other machairodont species. Evidence of cursorial adaptations in Homotherium suggests a hunting technique different from modern cats or smilodontine sabre-tooths. Some, like reduction of the claws, would have limited the ability of individual homotheres to bring down large prey, implying group action. Homotherium would also have been disadvantaged in direct confrontation with Pleistocene lions by smaller body mass, reduced forepaw muscle strength, smaller claws and more fragile dentition. Its hunting technique would have worked best in more open habitats, but competition from lions would have forced it to seek moderate cover. Among factors that could de-stabilise coexistence of the two big cat species in Pleistocene Europe we invoke a decrease in environmental mosaicism associated with stepped climatic change over the last million years, and the increased importance of humans within the larger predator guild. r 2004 Elsevier Ltd. All rights reserved. 1. Introduction Whatever the antiquity of human occupation of Europe, it is clear that meat was most probably an important food source (Roebroeks, 2001). That interest in meat was shared by a guild of large Carnivora, and any understanding of earliest human presence must take account of the structure and activities of that guild (Turner, 1992). An obvious enigma among the guild members is the lion-sized sabre-toothed cat Homother- ium latidens, which was widespread as perhaps the dominant predator in a variety of Eurasian and African environments from the Pliocene until at least the Middle Pleistocene. It certainly coexisted with several species of hominins across its spatial and temporal range, from African Pliocene sites with Australopithecus species to later levels of Zhoukoudian cave, home to Asian Homo erectus, and the upper levels of Gran Dolina at Atapuerca with Homo heidelbergensis. In Europe, Homotherium latidens coexisted with the lion, Panthera leo, for an interval of at least 200 kyr after the arrival of the latter species on the continent some time during the Middle Pleistocene, a coexistence that raises interesting palaeoecological problems (Turner, 1995a; Turner and Anto´n, 1999). The recent discover of a mandible of Homotherium latidens dated to around 28,000 yr BP in the North Sea (Reumer et al., 2003) even suggests that either the animal survived in Eurasia until the last glaciation, thus lengthening considerably the period of co-existence with lions, or that it re-entered the ARTICLE IN PRESS 0277-3791/$ - see front matter r 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.quascirev.2004.09.008 Corresponding author. Tel.: +44 151 231 2182; fax: +44 151 207 3224. E-mail address: [email protected] (A. Turner).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
0277-3791/$ - se
doi:10.1016/j.qu
�Correspondfax: +44151 20
E-mail addr
Quaternary Science Reviews 24 (2005) 1287–1301
Co-existence of scimitar-toothed cats, lions and hominins in theEuropean Pleistocene. Implications of the post-cranial anatomy of
Homotherium latidens (Owen) for comparative palaeoecology
Mauricio Antona, Angel Galobartb, Alan Turnerc,�
aDepartamento de Paleobiologia, Museo Nacional de Ciencias Naturales, C. Jose Gutierrez Abascal 2, 28006 Madrid, SpainbInstitut de Paleontologia Dr Miquel Crusafont, C. Escola Industrial 23, 08201 Sabadell, Spain
cSchool of Biological and Earth Sciences, Liverpool John Moores University, Liverpool L3 3AF, UK
Received 3 February 2004; accepted 24 September 2004
Abstract
Human occupants of Europe shared food resources with a number of larger Carnivora, and their coexistence with two lion-sized
felids, the lion and the scimitar-toothed machairodont Homotherium latidens, poses intriguing paleoecological problems. We
investigate the ecology of Homotherium latidens using an exceptional sample of postcrania from the Spanish Early Pleistocene site of
Incarcal, making comparisons with modern cats and with other machairodont species. Evidence of cursorial adaptations in
Homotherium suggests a hunting technique different from modern cats or smilodontine sabre-tooths. Some, like reduction of the
claws, would have limited the ability of individual homotheres to bring down large prey, implying group action.Homotherium would
also have been disadvantaged in direct confrontation with Pleistocene lions by smaller body mass, reduced forepaw muscle strength,
smaller claws and more fragile dentition. Its hunting technique would have worked best in more open habitats, but competition from
lions would have forced it to seek moderate cover. Among factors that could de-stabilise coexistence of the two big cat species in
Pleistocene Europe we invoke a decrease in environmental mosaicism associated with stepped climatic change over the last million
years, and the increased importance of humans within the larger predator guild.
r 2004 Elsevier Ltd. All rights reserved.
1. Introduction
Whatever the antiquity of human occupation ofEurope, it is clear that meat was most probably animportant food source (Roebroeks, 2001). That interestin meat was shared by a guild of large Carnivora, andany understanding of earliest human presence must takeaccount of the structure and activities of that guild(Turner, 1992). An obvious enigma among the guildmembers is the lion-sized sabre-toothed cat Homother-
ium latidens, which was widespread as perhaps thedominant predator in a variety of Eurasian and Africanenvironments from the Pliocene until at least the Middle
e front matter r 2004 Elsevier Ltd. All rights reserved.
ascirev.2004.09.008
ing author. Tel.: +44151 231 2182;
7 3224.
ess: [email protected] (A. Turner).
Pleistocene. It certainly coexisted with several species ofhominins across its spatial and temporal range, fromAfrican Pliocene sites with Australopithecus species tolater levels of Zhoukoudian cave, home to Asian Homo
erectus, and the upper levels of Gran Dolina atAtapuerca with Homo heidelbergensis. In Europe,Homotherium latidens coexisted with the lion, Panthera
leo, for an interval of at least 200 kyr after the arrival ofthe latter species on the continent some time during theMiddle Pleistocene, a coexistence that raises interestingpalaeoecological problems (Turner, 1995a; Turner andAnton, 1999). The recent discover of a mandible ofHomotherium latidens dated to around 28,000 yr BP inthe North Sea (Reumer et al., 2003) even suggests thateither the animal survived in Eurasia until the lastglaciation, thus lengthening considerably the period ofco-existence with lions, or that it re-entered the

ARTICLE IN PRESS
Fig. 1. Schematic drawing of the skeleton of Homotherium latidens
with parts recovered at Incarcal shown in a darker gray. Scale bar
25 cm.
M. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011288
continent from North America (see discussion below onextinctions in Section 6.4). In either event it would haveco-existed with Neanderthals and fully modern humans.One assumption has been that Homotherium, like
other large machairodont cats, would tend to leave agreater amount of scavengeable food at a kill (Turner,1992). Assessing the ecological significance of Homo-
therium requires an understanding of its palaeobiology,and thus of its functional anatomy, but remains areusually rare and fragmentary with postcrania mostlyknown from a single semi-complete skeleton from thePliocene locality of Seneze in France (Ballesio, 1963).That specimen confirmed the anatomical distinctivenessof Homotherium as a gracile sabre-tooth with a skeletondiffering completely from the typical, hyper-robustsabre-tooths such as the well-known American genusSmilodon, or its likely ancestor Megantereon, asrecognised on the basis of partial remains fromHundsheim in Austria by Schaub (1925). Kurten(1952) coined the terms ‘‘scimitar-toothed cats’’ (suchas Homotherium) and ‘‘dirk-toothed cats’’ (such asSmilodon and Megantereon) for the two contrastingtypes of felid sabre-tooths defined on the basis of theshape of their upper canines and also set apart by theirdiffering postcranial anatomy. Martin (1980, 1989)defined the scimitar-tooths and the dirk-tooths as‘‘ecomorphs’’, meaning that both the type of uppercanines and the body proportions of each type were partof a set of adaptations to a particular lifestyle andpredatory strategy.Although the morphological distinctiveness of Homo-
therium is thus well established, the implications for itslocomotory and predatory behaviour are more difficultto define, especially since the hunting and killingtechniques of sabre-tooths as a whole are only nowbecoming clear (Turner and Anton, 1997, 1999; Antonand Galobart, 1999). Among earlier authors, Martin(1980) suggested that dirk-tooths were typical ambushhunters, while scimitar-tooths were more like pursuitpredators, an interpretation elaborated on by Rawn-Schatzinger (1992), who used a large sample of the LatePleistocene species Homotherium serum from Friesen-hahn Cave (Texas) to conclude that it would have been asprint runner like the modern cheetah. This agrees withthe ideas of Martin (1989, p. 544) who thought that H.
latidens had cheetah-like skeletal proportions. Morerecent studies by Anyonge (1996) and Lewis (1997)confirm the presence of cursorial adaptations in Homo-
therium, although neither considers the implications ofsuch adaptations in terms of prey procurement andhandling.In contrast, Ballesio (1963, pp. 118–119) had earlier
concluded after studying the Seneze specimen thatHomotherium latidens would be less capable of jumpingand sprinting than modern pantherine cats, arguing thatalthough the elongated limb-bones of Homotherium
suggest a similarity with the modern cheetah, the bodyproportions of these two cats were extremely different(1963, p. 112). Such contradictory interpretations do notseem to stem from real differences between the Amer-ican and Old-World species, but rather from differentauthors focusing their studies on different aspects ofanatomy. A comprehensive overview that unifies thesedisparate observations is required in order to obtain amore complete picture of Homotherium as a livingpredator.Since the publication of Ballesio’s monograph in
1963, virtually no new studies have considered the post-cranial anatomy of H. latidens in detail, althoughnumerous remains have been recovered. Moreover,some elements important for functional interpretations,such as the ungual phalanges, are virtually absent inSeneze (only one claw was found, and the digit to whichit corresponded is not known) and a single individualgives little idea of the range of morphological variationwithin a species. Fortunately, a rich assemblage from theEarly Pleistocene Spanish site of Incarcal offers anexceptional contribution to our knowledge of the post-cranial anatomy of this taxon since there are completeand well-preserved elements of the trunk and limbs ofseveral individuals (Fig. 1), including a substantialsample of ungual phalanges that allow us to assess themorphology and relative size of the claws in H. latidens
for the first time (Galobart, 1996; Anton and Galobart,1999).The aims of this paper are therefore to describe the
Incarcal sample in comparison with other publishedmaterial of H. latidens, and then to broaden ourcomparisons to other species of felid sabertooths andthe modern larger felids, with a special emphasis on thelion. From these comparisons and a detailed functionalinterpretation of the observed morphology we hope thata more comprehensive model of the predatory beha-viour and ecology of H. latidens will emerge, which willallow us to address broader ecological implications. Wethus hope to provide answers to a series of intriguing

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1289
questions posed by the coexistence of Homotherium, thelarge pantherines, other large Carnivora and homininsin the European fossil record: Would there, for example,be a direct competition between Homotherium and thelion in the places where they coexisted? How would sucha competition be affected by environmental factors?What can we learn from other examples of coexistenceof competing large Carnivora in the fossil record, inmodern environments or in historical times? Would thepresence of hominins and their entrance to the largepredator guild impinge upon Homotherium? To whatextent did Homotherium consume and defend its kills,and how important would the remains of those kills beas a resource for scavengers, including hominins? Wouldlions, hyaenids and hominins exert pressure on, or evenbecome kleptoparasites of, Homotherium? And, last butnot least, what combination of ecological factors couldbring about the extinction of Homotherium?
2. Materials and methods
We examined a sample of post-cranial bones ofHomotherium latidens from the Spanish site of Incarcalcomparing their morphology with that of modernpantherine species including skeletons of lion (Panthera
leo), tiger (Panthera tigris), leopard (Panthera pardus),jaguar (Panthera onca), and cheetah (Acinonyx jubatus) inthe Museu de Zoologia de Barcelona and one lionskeleton from the Museo Nacional de Ciencias Naturalesof Madrid (Spain). We also used for reference publisheddescriptions of Homotherium, Smilodon, and other sabre-toothed taxa. For morphological comparisons we havefollowed the traditional approach of comparative anat-omy, expressing the observed differences as qualitativeobservations. This allows us to incorporate the observa-tions of previous authors who described the relevantmaterial (Merriam and Stock, 1932; Meade, 1961;Ballesio, 1963; Rawn-Schatzinger, 1992).We used descriptions of muscular anatomy and
muscle insertions in extant carnivores as a guide forour reconstructions of muscles in Homotherium; for adiscussion of methods see Anton (2003) and Turner andAnton (1997, 2004). Principal sources were the descrip-tions of Barone (1967, 1989), Evans and Christensen(1976), and Spoor and Badoux (1986), and, unlessotherwise stated, our observations on attachments ofparticular muscles are based on these references. We areaware of the caution expressed by Bryant and Seymour(1990) about the reliability of muscle reconstructions,but, as they state, the Carnivora are a good group towork with in this regard because a high percentage ofmuscles leave discrete marks on the bones where theyattach.Following the methodological approach of Bryant
and Russell (1992) and Witmer (1995) we used the
phylogenetic relationships between the extinct saber-toothed felids and extant carnivores to infer unpreservedanatomical features. For a general phylogenetic context(see Anton and Galobart, 1999, Fig. 1a), we havefollowed the hypothesis of Werdelin (1996), whichaddresses the relationships among the Carnivora at thefamily level. The phylogeny of felids recently publishedby Turner and Anton (1997) is adopted here (Anton andGalobart, 1999, Fig. 1b).
3. The Incarcal site
The Incarcal site complex is located in the lacustrinebasin of Banyoles-Besalu (Northeastern Spain). A totalof nine karst cavities filled with fossiliferous sediments inPliocene limestones are currently known (Ros et al.,2003). Three of these fillings (Incarcal I, II, and V) haveyielded the majority of the faunal remains, while theothers have a much lower fossiliferous richness. Thefaunal association of the site is typical for the lowerBiharian (Lower Pleistocene) of Western Europe (Ga-lobart et al., 1990, 1996; Colomer et al., 1990; Galobart,1996; Anton and Galobart, 1999).Anatomical connections are not very common, and
the Homotherium remains in particular are mostlyunassociated. Taphonomic analyses (Galobart, 1996,2003) show these deposits to be unlike any other sitemodel (lacustrine, palustrine, carnivore traps, hyaenadens), but similarities were found with the Miocenekarst fillings of Thomas Farm, Florida (Pratt, 1990).
4. Observations and comparisons
4.1. Thoracic and lumbar vertebrae
The thoracic vertebrae of the Incarcal Homotherium
display no remarkable differences from those ofpantherine cats, but the centra of the lumbars arerelatively shorter (Fig. 2). The anterozygapophises inanterior view are more ‘‘enveloping’’ than in moderncheetah and lion, but less so than in jaguar. The shape ofthe transverse processes, with a wide insertion in thevertebral body extending laterally in the first half andwith only moderate ventral inclination, resembles thejaguar more than any other modern big cat in oursample. The neural spines have an anterio-posteriorlylong and wide summit. Among the modern cats, onlythe jaguar approximates Homotherium in these features.
4.2. Humerus
The greater tuberosity is higher relative to the humeralhead than in pantherines (Fig. 3A and B), resemblingthe cheetah. Insertion areas for the supraspinatus
ARTICLE IN PRESS
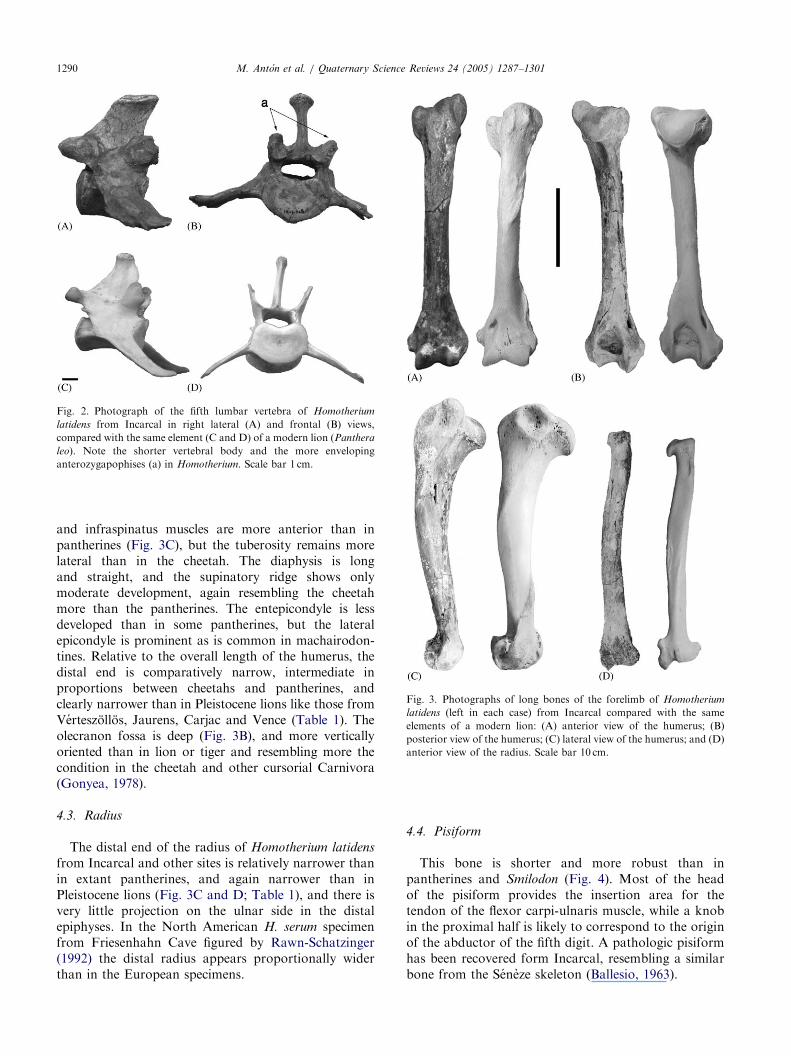
Fig. 2. Photograph of the fifth lumbar vertebra of Homotherium
latidens from Incarcal in right lateral (A) and frontal (B) views,
compared with the same element (C and D) of a modern lion (Panthera
leo). Note the shorter vertebral body and the more enveloping
anterozygapophises (a) in Homotherium. Scale bar 1 cm.
Fig. 3. Photographs of long bones of the forelimb of Homotherium
latidens (left in each case) from Incarcal compared with the same
elements of a modern lion: (A) anterior view of the humerus; (B)
posterior view of the humerus; (C) lateral view of the humerus; and (D)
anterior view of the radius. Scale bar 10 cm.
M. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011290
and infraspinatus muscles are more anterior than inpantherines (Fig. 3C), but the tuberosity remains morelateral than in the cheetah. The diaphysis is longand straight, and the supinatory ridge shows onlymoderate development, again resembling the cheetahmore than the pantherines. The entepicondyle is lessdeveloped than in some pantherines, but the lateralepicondyle is prominent as is common in machairodon-tines. Relative to the overall length of the humerus, thedistal end is comparatively narrow, intermediate inproportions between cheetahs and pantherines, andclearly narrower than in Pleistocene lions like those fromVerteszollos, Jaurens, Carjac and Vence (Table 1). Theolecranon fossa is deep (Fig. 3B), and more verticallyoriented than in lion or tiger and resembling more thecondition in the cheetah and other cursorial Carnivora(Gonyea, 1978).
4.3. Radius
The distal end of the radius of Homotherium latidens
from Incarcal and other sites is relatively narrower thanin extant pantherines, and again narrower than inPleistocene lions (Fig. 3C and D; Table 1), and there isvery little projection on the ulnar side in the distalepiphyses. In the North American H. serum specimenfrom Friesenhahn Cave figured by Rawn-Schatzinger(1992) the distal radius appears proportionally widerthan in the European specimens.
4.4. Pisiform
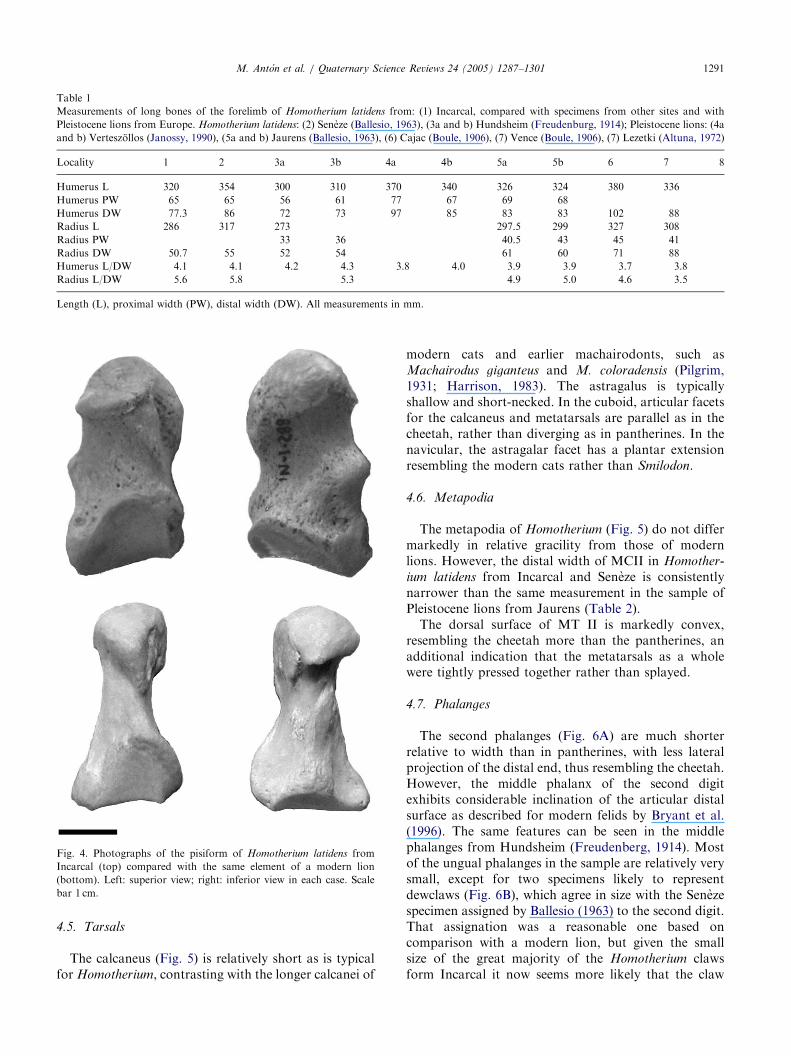
This bone is shorter and more robust than inpantherines and Smilodon (Fig. 4). Most of the headof the pisiform provides the insertion area for thetendon of the flexor carpi-ulnaris muscle, while a knobin the proximal half is likely to correspond to the originof the abductor of the fifth digit. A pathologic pisiformhas been recovered form Incarcal, resembling a similarbone from the Seneze skeleton (Ballesio, 1963).
ARTICLE IN PRESS
Table 1
Measurements of long bones of the forelimb of Homotherium latidens from: (1) Incarcal, compared with specimens from other sites and with
Pleistocene lions from Europe. Homotherium latidens: (2) Seneze (Ballesio, 1963), (3a and b) Hundsheim (Freudenburg, 1914); Pleistocene lions: (4a
and b) Verteszollos (Janossy, 1990), (5a and b) Jaurens (Ballesio, 1963), (6) Cajac (Boule, 1906), (7) Vence (Boule, 1906), (7) Lezetki (Altuna, 1972)
Locality 1 2 3a 3b 4a 4b 5a 5b 6 7 8
Humerus L 320 354 300 310 370 340 326 324 380 336
Humerus PW 65 65 56 61 77 67 69 68
Humerus DW 77.3 86 72 73 97 85 83 83 102 88
Radius L 286 317 273 297.5 299 327 308
Radius PW 33 36 40.5 43 45 41
Radius DW 50.7 55 52 54 61 60 71 88
Humerus L/DW 4.1 4.1 4.2 4.3 3.8 4.0 3.9 3.9 3.7 3.8
Radius L/DW 5.6 5.8 5.3 4.9 5.0 4.6 3.5
Length (L), proximal width (PW), distal width (DW). All measurements in mm.
Fig. 4. Photographs of the pisiform of Homotherium latidens from
Incarcal (top) compared with the same element of a modern lion
(bottom). Left: superior view; right: inferior view in each case. Scale
bar 1 cm.
M. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1291
4.5. Tarsals
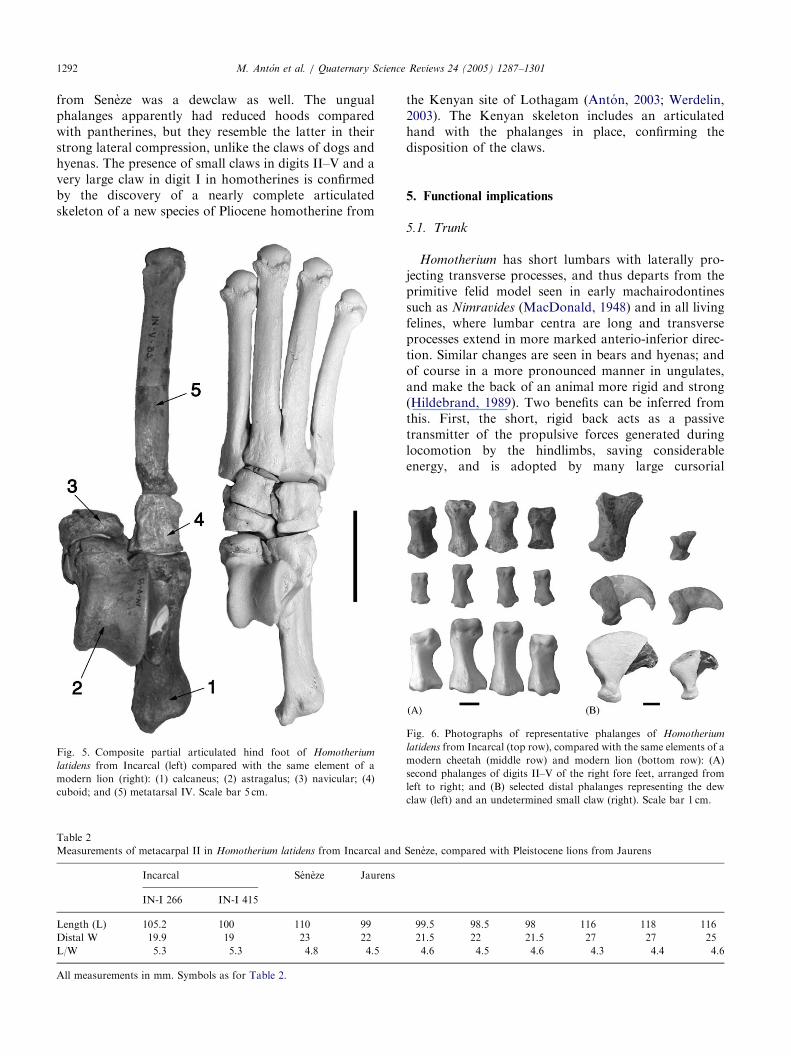
The calcaneus (Fig. 5) is relatively short as is typicalforHomotherium, contrasting with the longer calcanei of
modern cats and earlier machairodonts, such asMachairodus giganteus and M. coloradensis (Pilgrim,1931; Harrison, 1983). The astragalus is typicallyshallow and short-necked. In the cuboid, articular facetsfor the calcaneus and metatarsals are parallel as in thecheetah, rather than diverging as in pantherines. In thenavicular, the astragalar facet has a plantar extensionresembling the modern cats rather than Smilodon.
4.6. Metapodia
The metapodia of Homotherium (Fig. 5) do not differmarkedly in relative gracility from those of modernlions. However, the distal width of MCII in Homother-
ium latidens from Incarcal and Seneze is consistentlynarrower than the same measurement in the sample ofPleistocene lions from Jaurens (Table 2).The dorsal surface of MT II is markedly convex,
resembling the cheetah more than the pantherines, anadditional indication that the metatarsals as a wholewere tightly pressed together rather than splayed.
4.7. Phalanges
The second phalanges (Fig. 6A) are much shorterrelative to width than in pantherines, with less lateralprojection of the distal end, thus resembling the cheetah.However, the middle phalanx of the second digitexhibits considerable inclination of the articular distalsurface as described for modern felids by Bryant et al.(1996). The same features can be seen in the middlephalanges from Hundsheim (Freudenberg, 1914). Mostof the ungual phalanges in the sample are relatively verysmall, except for two specimens likely to representdewclaws (Fig. 6B), which agree in size with the Senezespecimen assigned by Ballesio (1963) to the second digit.That assignation was a reasonable one based oncomparison with a modern lion, but given the smallsize of the great majority of the Homotherium clawsform Incarcal it now seems more likely that the claw
ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011292
from Seneze was a dewclaw as well. The ungualphalanges apparently had reduced hoods comparedwith pantherines, but they resemble the latter in theirstrong lateral compression, unlike the claws of dogs andhyenas. The presence of small claws in digits II–V and avery large claw in digit I in homotherines is confirmedby the discovery of a nearly complete articulatedskeleton of a new species of Pliocene homotherine from
Fig. 5. Composite partial articulated hind foot of Homotherium
latidens from Incarcal (left) compared with the same element of a
modern lion (right): (1) calcaneus; (2) astragalus; (3) navicular; (4)
cuboid; and (5) metatarsal IV. Scale bar 5 cm.
Table 2
Measurements of metacarpal II in Homotherium latidens from Incarcal and
Incarcal Seneze Jaurens
IN-I 266 IN-I 415
Length (L) 105.2 100 110 99
Distal W 19.9 19 23 22
L/W 5.3 5.3 4.8 4.5
All measurements in mm. Symbols as for Table 2.
the Kenyan site of Lothagam (Anton, 2003; Werdelin,2003). The Kenyan skeleton includes an articulatedhand with the phalanges in place, confirming thedisposition of the claws.
5. Functional implications
5.1. Trunk
Homotherium has short lumbars with laterally pro-jecting transverse processes, and thus departs from theprimitive felid model seen in early machairodontinessuch as Nimravides (MacDonald, 1948) and in all livingfelines, where lumbar centra are long and transverseprocesses extend in more marked anterio-inferior direc-tion. Similar changes are seen in bears and hyenas; andof course in a more pronounced manner in ungulates,and make the back of an animal more rigid and strong(Hildebrand, 1989). Two benefits can be inferred fromthis. First, the short, rigid back acts as a passivetransmitter of the propulsive forces generated duringlocomotion by the hindlimbs, saving considerableenergy, and is adopted by many large cursorial
Seneze, compared with Pleistocene lions from Jaurens
99.5 98.5 98 116 118 116
21.5 22 21.5 27 27 25
4.6 4.5 4.6 4.3 4.4 4.6
Fig. 6. Photographs of representative phalanges of Homotherium
latidens from Incarcal (top row), compared with the same elements of a
modern cheetah (middle row) and modern lion (bottom row): (A)
second phalanges of digits II–V of the right fore feet, arranged from
left to right; and (B) selected distal phalanges representing the dew
claw (left) and an undetermined small claw (right). Scale bar 1 cm.

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1293
mammals (Hildebrand, 1989). Secondly, in animals thatput lateral stress on their backs, such as carnivores thatdig or turn large stones, a stiff back serves better tostabilise the trunk during such activities. Bears andbadgers offer examples of such adaptations (Gambar-yan, 1977). Typical felids, in contrast, gain accelerationwith the active flexion and extension of the back in thesaggital plane, an action that adds to bound length andabsolute speed, but which is energetically costly. Theseanimals, called ‘‘active limbers’’ by Hildebrand (1989),quickly achieve high maximum speeds but lack endur-ance (Gambaryan, 1977).For sabre-tooths, an important advantage of lumbar
stiffening would be the added strength to resist thelateral loads caused by the violent movements ofstruggling prey held by the forelimbs. Some adaptationsof the hindlimb are probably related to this stage of thehunt as well, as discussed below.
5.2. Forelimb
The slender humerus of Homotherium indicatescursorial adaptation. Elevation of the greater tuberositygives mechanical advantage to the propulsive action ofthe m. supraspinatus (Gambaryan, 1977; Spoor andBadoux, 1986), while muscles relevant for grapplingprey, such as the deltoids and extensors of the digits,have only moderately developed crests for insertion.However, evidence suggests that the action of at leastone of these muscles was important for prey procure-ment. The right humerus of the Seneze skeleton shows apathological bony growth similar to those found inSmilodon humeri from Rancho la Brea, extending fromthe deltoid crest in a posterolateral direction (Ballesio,1963). This suggests repeated overstressing of the deltoidduring flexion of the arm over the scapula, as wouldhappen when the animal tried to control struggling prey(Turner and Anton, 1997). On the other hand, thelateral (as in pantherines) rather than central (as in thecheetah) position of the greater tuberosity indicates thatthe combined abductor function of the infraspinatus,teres minor and supraspinatus muscles was still veryrelevant. The relatively deep olecranon fossa indicatesthe habitually extended position of the elbow articula-tion, a feature typical of the more cursorial andterrestrial felids such as the cheetah and lion, as opposedto the more flexed elbow in arboreal or scansorialspecies. Furthermore, the vertical orientation of thefossa falls closer to that of cheetah than that inpantherine cats, indicating more cursorial locomotion(Gonyea, 1978).The gracility of the radius is made more evident by its
narrow distal end, part of a series of cursorialadaptations at the wrist joint (Ballesio, 1963; RawnSchatzinger, 1992) that conflict with the necessity tograpple large prey.
The morphological similarity between the short,square pisiform of Homotherium and that of cheetah,in contrast with the long pisiform of pantherines,suggests a similarity in function as well. This in turnwould be related to the action of the flexor carpi-ulnarismuscle and to the role of carpal extension in cursoriallocomotion.The morphology of the middle phalanges with their
reduced asymmetry suggests a degree of claw retractionsmaller than in pantherines and similar to that of themodern cheetah (Bryant et al., 1996). However, thesecond digit seemingly retained an enhanced graspingfunction, as evidenced by the greater ulnar projection ofthe distal head of the medial phalanx and by thepronounced inclination of its distal articular surface, anadaptation to spread the claw relative to that of thethird digit during protraction (Bryant et al., 1996).The great reduction in ungual phalanx size in digits
II–V in Incarcal Homotherium suggests the claws had anincreased locomotory function, improving traction asthey do in Acinonyx. As in the cheetah, the primarygrasping function relied on the dewclaw. Nonetheless,the laterally compressed and evidently recurved shape ofthe claws in Homotherium suggest that they could stillplay a role in prey holding, in spite of their small size.
5.3. Hindlimb
The shortening of the calcaneus and the shallowtrochlea of the astragalus are features usually associatedwith plantigrade locomotion in Carnivora (Ginsburg,1961), and Homotherium has sometimes been recon-structed with plantigrade hindfeet. However, the meta-tarsals were tightly bound together, as indicated by thedorsal convexity of Mt II among other features, insteadof splaying as in bears and other plantigrade carnivores(Ginsburg, 1961). Other features of the hindlimbosteology, including the shape of articular facets in thecuboid and navicular, indicate a completely digitigradestance and an enhanced mobility in the parasagital plane(Ballesio, 1963; Rawn-Schatzinger, 1992).The relatively shallow trochlea of the astragalus may
be an adaptation to wrestling with struggling prey.When dealing with a large animal, modern cats try tokeep their hindfeet on the ground, often in a plantigradeposition (Leyhausen, 1979), to gain greater control if theprey makes violent movements. In sabertooths, whichneeded to keep large prey almost completely immobi-lised, the tibio-astragalar joint was probably subject tolarge lateral forces, but the feet had to remain firmlyplanted on the ground. It would then be advantageousto provide the joint with some additional degree oflateral mobility, rather than risk being knocked offbalance when a rigid foot lost hold on the ground, andshortening of the calcaneus and metatarsals wasprobably related to this need for stability.

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011294
5.4. Locomotory behaviour
The above observations of the material from Incarcalcombined with data from other sites, and mostespecially from Seneze and Hundsheim, allow us topresent an accurate reconstruction of the skeleton ofHomotherium latidens. This reconstruction is in turncompared with a restored skeleton of a Pleistocene lionbased on an associated specimen from Carjac, in France(Boule, 1906). The differences in size and proportionsare easily seen both in the skeletal and in the lifecomparisons (Fig. 7), showing that Homotherium was arelatively lightly built animal with a slightly sloping backsomewhat resembling that of a spotted hyaena.The hyaena-like proportions of Homotherium suggest
similarities in the locomotory abilities of both animals, apoint already noticed by Ballesio (pers. com.). Therelatively short and rigid back would have contributedless than that of pantherines to sudden acceleration butwould probably provide more endurance, although lessthan in hyaenas. As noted by Ballesio (1963), theshortened tibia and hindfeet imply a lesser jumpingability than in pantherines, but the elongated forelimbs,including a high scapula (Ballesio, 1963; Rawn-Schat-zinger, 1992), would allow Homotherium to run with aneasy canter. This is a type of gallop mainly powered bythe forelimbs and which is more energy-efficient thanother gallops when it comes to long-distance travel(Guthrie, 1990; Kurten and Werdelin, 1990).Rawn-Schatzinger (1992) suggested that the reduced
claws of Homotherium serum would be related to acheetah-like type of sprinting locomotion, because thecheetah is the only modern cat showing a similar degreeof claw reduction. However, since the body proportionsof homotheres were not well adapted to sprinting(especially because of the shortening of the back andhindlimbs), it seems more likely that the claw reduction
Fig. 7. Reconstruction of skeleton and body outline of Homotherium
latidens (top) and Pleistocene lion (bottom). The schematic drawing of
the skeleton shows the functionally relevant regions and features in
whichHomotherium differs from the lion: (1) shorter lumbar vertebrae;
(2) slender humerus; (3) distally narrow radius; (4) short and robust
pisiform; (5) less asymmetrical second phalanges; (6) reduced claws on
digits II–V and very large dew claw; (7) shortened calcaneum; and (8)
shallow and short-necked astragalus. Scale bar 25 cm.
was part of an adaptation to improve traction duringlong-distance, medium-speed travel, as in canids andhyaenids. Such locomotory abilities strongly point to anopen habitat preference for Homotherium, and alsosuggest that it would have foraged over considerabledistances.
5.5. Predatory behaviour and size of prey
Several lines of evidence suggest relatively large preysize for Homotherium. The morphology of the dentition,head and neck are well adapted to cause rapid andextensive bleeding in animals of similar or larger sizethan the predator, particularly with a carefully posi-tioned bite at the throat of restrained prey as discussedin detail by Anton and Galobart (1999). This inter-pretation is supported by adaptations for stability in theback and hindlimbs that suggest an ability to resist greatlateral forces when holding prey, and stress-inducedpathologies in the forelimb indicate violent struggleswith large prey held by the forepaws. Yet the adapta-tions for moderately cursorial locomotion made theanimal comparatively less powerful than a modern lionor tiger, and much less than the dirk-toothed cat,Smilodon, and with a less grasping forelimb. Tocompensate for such loss of individual muscularstrength and grasping ability, it seems likely thatHomotherium would practise group hunting, as dis-cussed below. It is also conceivable that the loss ofprehensile ability of the forepaws was compensated forby the large and projecting incisor battery as an aid inholding prey, as is the case in dogs and hyaenas(Bicknevicius et al., 1996).The initial approach to prey would either be a typical
feline stalk or a more open walk, as often seen incheetahs, dogs and hyaenas. Prey would likely be sloweddown with the aid of the forelimb claws, especially thoseof the first and second digits, and perhaps by biting withthe protracted incisors and lower canines in the areaswhere the skin of prey is loose (for instance betweenbelly and thigh). One or several cats would probablypull the animal to the ground, with backs tensed andhindfeet firmly planted to resist any struggle while thekilling bite was applied to the throat. It is possible that ifmore than one cat were hunting some could bite theabdomen while another individual was biting the throat.Such behaviour has been observed in modern lions,often leading to death by blood loss and eviscerationbefore the killing bite to the throat has had full effect(Schaller, 1972). However, it is always necessary for onelion to control the head of the prey, and solitary catshave never been recorded killing by evisceration to ourknowledge.The ‘‘hit and run’’ hypothesis for sabertooth preda-
tion proposed by some authors (Akersten, 1985; Rawn-Schatzinger, 1992), involving bites at the belly, retreat to

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1295
avoid retaliation and later tracking of the injured andweakened animal, seems to us an unlikely strategyinvolving too high a risk of losing prey to competingpredators and scavengers. Wounded prey running looseis an invitation to lunch for any watching eyes, and inrelatively open woodland environments there would bemany. All modern large mammalian predators areunwilling to let go of their prey once contacted, and itseems only likely that the same was true for thehomotheres, which could easily lose their catch to clansof giant hyaenas, lion prides, or, just as bad, to othermembers of their own species.It has often been suggested that Homotherium was a
specialist killer of proboscideans, based largely on theassociation of the American species Homotherium serum
with young mammoths at Friesenhahn Cave (Meade,1961) and with an adult mastodon in Gassaway Fissure(Kurten and Anderson, 1980; Rawn Schatzinger, 1992).The association of Homotherium with partially articu-lated skeletons of an adult elephant and a hippopotamusat Incarcal makes it tempting to infer a predator–preyrelationship, but it is more likely that the predators wereattracted to injured or dead animals to scavenge, or eventhat the carcasses of carnivores and herbivores werebrought together by water. The Gassaway Fissurematerial can also be interpreted in terms of scavenging,while the Friesenhahn Cave association, which is themost convincing example of homothere predation onproboscideans, may reflect a local phenomenon wheremost of the animals taken seem to have been around theage of 2 years. Modern lions in parts of Africa areknown to take elephants of similar ages with morefrequency than previously believed (Joubert, 1994;Joubert and Joubert, 1997), largely because they arestarting to leave strong adult protection but have not yetreached the size needed to repel an attack.Two factors seem to be relevant for the taking of large
prey by big cats: individual strength and group action.The muscle power of an individual homothere wasprobably less than a modern lion of comparableshoulder height, as suggested by the relative gracilityof the forelimb, so even if we assume group huntingthere is little reason to infer that the upper limit of whatthey could kill was significantly above that of lions. Agroup of homotheres would probably be able to bringdown animals the size of a young elephant or an adultlarge bovine, but adult rhinos and elephants wouldprobably be virtually immune to predation in normalcircumstances, as they are today.Once prey was brought down and the killing bite
applied, loss of blood would weaken the animal in a fewminutes, giving homotheres a considerable advantageover pantherine cats. While modern lions can finish anyprey smaller than a zebra with a relatively quick bite,killing of buffalo or young elephants can be ratherinefficient because the blunt canine teeth cannot
penetrate their thick skin or exert enough pressure onthe windpipe to cause suffocation. An adult buffalo willoften rise to its feet and escape from a whole lion prideafter long harassment, often leaving injured lions andalways with considerable waste of pride energy. Thusthe key advantage of the scimitar-tooth adaptation wasprobably not so much the access to prey significantlylarger than lions can take but the higher efficiency in thekilling of animals within the upper range of the lion’sprey spectrum.
6. Ecology and social behaviour
6.1. European dispersion of Homotherium, and the
Villafranchian large carnivore guild
Initial dispersion of Homotherium in Europe isprobably linked with changes in the environmentrecorded at around 3.2Ma, when the climate becamecooler and drier and vegetational cover decreased. Therewas an increase in seasonality and the Mediterraneanpattern of summer drought began at that time (Agustıand Anton, 2002), culminating in the well-established2.4Ma event that saw the beginning of northernhemisphere glaciations. The dominant large cat of theEuropean early Pliocene had been the ‘‘false sabre-tooth’’ Dinofelis, an animal with the body proportionsof a forest dweller, but after the 3.2Ma event it wasreplaced by Homotherium latidens and Megantereon
cultridens. The latter was somewhat smaller thanDinofelis but had teeth and forelimbs well adapted totaking moderate-sized prey.What is the meaning of this turnover among the
Pliocene large felids? The answer must lie in a change inavailable resources, both in the form of vegetationalcover and of prey animals. As the vegetation shiftedfrom a more or less uniform forest cover to a mosaic ofgrassland or open woodland punctuated by riverineforest, it is likely that the ungulate guild would havedivided into herds of migrating grazers, including equidand bovid species, and a mass of resident prey species,including both grazers and browsers. The Homother-
ium–Megantereon pairing would be well suited to exploitsuch diversity, with the scimitar-tooth taking largerprey, including migratory species which it might evenfollow, while the smaller dirk-tooth would remain tiedto its territories along the riverine forests and otherwooded areas, taking prey from the resident popula-tions.This evidently worked well for more than 2 million
years, until a further cold and dry event at around1.0Ma. The further reduction in forest cover wasprobably too hard a blow for European populationsofMegantereon, which had to face a reduced availabilityof resident prey and an increasing frequency of

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011296
dangerous encounters with larger predators in thehigher-visibility environments (Turner and Anton,1999). Recent studies (Domınguez-Rodrigo, 2001) pro-vide growing evidence that riparian habitats are andwere places of lower interspecific competition forcarnivores and scavengers than open habitats, reinfor-cing the notion that reduction in the prevalence of suchwoodlands would put increased pressure on the morevulnerable members of the guild.Most recently, Hemmer (2002) has attributed the
extinction of Megantereon in Europe to a combinationof climatic factors and the pressure from hominins ofthe species Homo heidelbergensis. However, evidence ofhominin occupation in Europe by the time of theextinction ofMegantereon is reduced at best to scatteredassemblages of primitive, Oldowan stone tools (Turner,1999a; Roebroeks, 2001; Dennell, 2003), probably bestattributed to an earlier species of hominin, Homo
antecessor, which is likely to have had a lesser impactas a competitor for the larger predators than the MiddlePleistoceneH. heidelbergensis. But the solitary, relativelysmall Megantereon was probably more vulnerable toany pressure than its larger cousins, and the impact ofhominins is not to be lightly dismissed as a contributingfactor to its final extinction. Homotherium in turn,managed to survive for at least another half millionyears.
6.2. Social groupings and habitat preferences
While there is every reason to believe that Meganter-
eon was a solitary ambush predator, there are severallines of evidence suggesting social groupings in Homo-
therium. Its body proportions would make it lesscapable of sudden acceleration and less strong than alion or tiger, a combination of features that made thetypical felid ambush hunting more difficult, and wouldcall for group action if large prey was to be taken. Thereare strong indications that in certain environmentalcircumstances lion-sized cats would find it not onlyadvantageous but also almost necessary to becomesocial (Packer, 1986; Sunquist and Sunquist, 1989). Inopen environments, where carcasses are easily detectedfrom a distance, large cats find advantages in groupingin order to defend their kills and their young fromcompeting carnivores, including other members of thesame species. In the Early Pleistocene, the giant hyaena,Pachycrocuta brevirostris was an important threat, as itslarge size was probably linked with aggressive scaven-ging habits (Turner and Anton, 1996). An example ofthe enormous size of P. brevirostris is the presence inIncarcal of a complete humerus that is about the samelength as a Homotherium humerus from the same site(Galobart et al., 2003).Furthermore, there would be advantages in group
hunting for larger cats living in more open environ-
ments. Although some authors have questioned theadvantages of cooperative hunting, seeing no real gainin terms of per capita foraging success for group-huntingcats compared with solitary ones (Packer, 1986; Caro,1994), studies of lion prides in Etosha National Park(Namibia) show that lions can overcome the problem ofbeing detected by prey in the absence of cover by co-ordinating their activities (Stander and Albon, 1993).While group size is not solely dependent on per capita
foraging returns in Etosha lions (Caro, 1994), it remainstrue that there is a real advantage in group foraging atleast during the dry season when available cover is at itsminimum. It is thus feasible that, as vegetational coverdecreased as a result of Plio-Pleistocene climaticchanges, ancestral homotheres would face the need tocope with increasingly easy detection by their prey. Evenif the problem were only exacerbated in the dry season,the advantage gained during that period could havebeen an added reason for becoming social.The ecological model emerging from this analysis
suggests that Homotherium would not favour woodlandor gallery forests more than modern lions do; if anythingit would favour them less. However, the presence ofHomotherium in a fossil site does not exclude woodland,and those where both Homotherium and Megantereon
are found are likely to reflect the presence of a mosaic ofwoodlands and grasslands, with the smaller cat retreat-ing to forested areas to avoid confrontation withHomotherium. A similar pattern is seen today in Africa,where a mosaic of forest and grassland is essential toallow the bigger lion and the smaller leopard to coexist(Schaller, 1972; Turner and Anton, 1999, 2004). In Asia,the presence of vegetational cover is essential for thecoexistence of tigers and leopards (Seidensticker, 1976;Seidensticker and McDougal, 1993).
6.3. Co-existence of equal-sized large felids in past and
present ecosystems
The presence of both Homotherium latidens and thelion is recorded in several European sites of MiddlePleistocene age, including Mosbach (Koenigswald andHeinrich, 1999), Verteszollos (Janossy, 1990), Westbury(Turner, 1999b), Mauer (Koenigswald and Heinrich,1999) and Atapuerca TD10 (Garcıa, 2002). While theoccurrence of two lion-sized felids in the same site mayat first appear unusual, the fact is that there are severalexamples of such coexistence in the fossil record, as wellas in modern and historical times.In Africa between 3.0 and 1.5Ma, the large carnivore
guild included about twice as many species as today,including both Homotherium and the lion (Turner andAnton, 1999, 2004). In the South African caves inparticular, co-existence of the two species is documentedin several hominin-bearing deposits, including members4 and 5 of Sterkfontein (Turner, 1997) and Kromdraai

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1297
A (Brain, 1981). Extinctions of archaic carnivores inAfrica occurred during the Early Pleistocene (Turnerand Anton, 2004) with several of the more specialisedtaxa disappearing, and it has been said that in Africa,‘‘for carnivores, the last million years has been the age ofthe generalists’’ (Werdelin and Lewis, 2001).In the North American Pleistocene, Homotherium
serum and the American lion, Panthera atrox, werejoined by a third lion-sized felid, the dirk-toothSmilodon fatalis (Kurten and Anderson, 1980). In SouthAmerica, a late Pleistocene immigration of northerntaxa resulted in the co-existence of Panthera atrox andSmilodon fatalis west of the Andes (Kurten andWerdelin, 1990). In China, both Homotherium latidens
and the tiger, Panthera tigris, are recorded in the MiddlePleistocene localities 9 and 13 of Zhoukoudian (Teilhardde Chardin, 1936).In historic times, the range of the Asian lion, Panthera
leo persica, overlapped considerably with that of thetiger in northern India (Pocock, 1939). In present-dayCentral and South America the ranges of the jaguar(Panthera onca) and the puma (Puma concolor) overlapextensively, with both cats often competing for similarfood resources (Nunez et al., 2000). In India in historicaltimes, and in present day Africa, the similarly sizedcheetah (Acinonyx jubatus) and leopard (Panthera
pardus) display a clear range overlap.What are the conditions that allow the coexistence of
such competing predators in their respective ecosystems?In the first place, while there is considerable overlap insize, there are indeed differences in body mass betweenmost of the pairings or groupings of felids mentioned.The Pleistocene lions of Europe averaged considerablylarger than their modern counterparts (Anyonge, 1993;Turner and Anton, 1997), but a recent estimate of bodymass for Homotherium yields a surprising range of210–400 kg (Hemmer, 2002). This is well above theaverage for modern lions, but our own comparisonsclearly show that the postcranial bones of Homotherium
were similar to or more gracile than in modern lions,pointing to a similar, or even slightly smaller body mass(Fig. 7). This underlines the problems of estimatingabsolute values for body mass in extinct felids. Amongthe possible methods for making such estimates,inferences based on the dimensions of individual teethappear to be least reliable (Turner and O’Regan, 2002).In the absence of more solid criteria for estimates ofabsolute mass in machairodontines it appears better torely on comparative approaches, and in the case ofHomotherium a comparison of size and morphology ofthe skull and appendicular skeleton with that of modernlions is likely to provide the best working estimates. But,since the lions with which Homotherium co-existed in thePleistocene were in general larger and more robust thantheir modern relatives, perhaps the most useful compar-ison is that between the reconstructed skeleton and body
shape of Homotherium and a Pleistocene lion shown inFig. 7. Accurate reconstructions based on skeletalmeasurements and on careful consideration of thedisposition of muscular masses can be a useful tool toassist in body size comparisons, and one that can help incorrecting gross misinterpretations, like those sometimesderived from the use of tooth measurements for bodymass estimates.A considerable overlap in body weight apparently
occurred between Smilodon fatalis and the enormousAmerican lion (Anyonge, 1993), while the tigers andlions of India are remarkably similar in size. Jaguarsoverlap with pumas, and leopards overlap with chee-tahs, but in both pairings the first species averagesheavier than the second (Kitchener, 1991).While differences in body mass can help different
carnivore species to partition the available resources, itis also evident that, in spite of such differences, mostlarge carnivores in an ecosystem will tend to prey on themost abundant suitably sized ungulate species. InKruger National Park in South Africa, lions, leopards,cheetahs, spotted hyaenas and wild dogs all have theabundant impala as their main prey, although theantelope may seem a little small for the lions and a littlebig for the cheetah (Mills and Biggs, 1993). In BandipurReserve, India, tigers, leopards and dholes (Cuon
alpinus) all have the chital deer as their main prey.Larger sambar deer, seemingly a more suitable prey forthe tiger, are also present, but while they are selected thesheer abundance of the chital makes it the most frequentprey of the tiger (Johnsingh, 1992). It can thus beexpected that in Pleistocene ecosystems, as in the presentday, all large carnivores would compete for a fewabundant prey species. With this in mind, the complextheoretical models of ecological hierarchy among fossilfelids, such as that proposed by Hemmer (2002) for theEarly Pleistocene German site of Untermassfeld, may beoffering too neat an image of prey apportionment andunderestimating the real extent of competition.Other important means by which large carnivores can
minimise direct competition and confrontation is byhunting in different sections of the ecosystem, and atdifferent times of day. Thus, cheetahs and wild dogs inKruger tend to hunt more during the day, therebyavoiding the more nocturnal lions, leopards and hyaenas(Mills and Biggs, 1993). But perhaps the most importantfactor allowing the coexistence of several sympatricspecies of large carnivores is the vegetation. As wementioned, the presence of a mosaic of open and closedvegetation is essential for the co-existence of tigers andlions in India and of lions and leopards in Africa, andriparian woodlands are generally seen as places ofrelatively low interspecific competition between preda-tors (Domınguez-Rodrigo, 2001). An especially inter-esting example is the case of the African wild dog, acursorial carnivore associated in the popular mind with

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011298
the open spaces of the Serengeti grassland, but whichactually becomes especially vulnerable to competitionfrom spotted hyaenas and lions in such open environ-ments. As a consequence, it actually fares better in areaswith more cover, such as the Selous Game Reserve inTanzania or the Okavango Delta in Botswana, while inthe Serengeti it has come to the verge of extinction(Creel and Creel, 1996, 2002; Gorman et al., 1998). It isthus possible that in the European Pleistocene we wouldsee the paradox of the more cursorial carnivore (thescimitar-tooth) seeking the refuge of wooded areas toescape harassment from the more robust carnivore (thelion).
6.4. Extinction
We may finally consider what our model of homo-therine behaviour and ecology tells us about the likelycauses for its extinction. If Homotherium was not only asemi-cursorial animal adapted to open environments butalso had some form of social structure, then simplistictheories linking its extinction with the spreading ofgrassland and decline of forest must be wrong. So toomust the traditional assumption that the evolution offaster ungulates brought about sabertooth extinctionsand the triumph of pantherines. It seems possiblehowever, that some extremely arid episodes during thePleistocene would have brought about an extraordinarydecrease in the mass of resident ungulates in any givenarea. That factor, coupled with decreased cover andincreased visibility, would likely have exacerbated thecompetition between different species of large carnivoresand pushed the most specialised ones toward extinction,as discussed in detail by Turner and Anton (1999).Besides that, it is likely that under the rapidly oscillatingclimatic conditions of the Pleistocene the versatilemorphology and behaviour of pantherines gave theman edge over the more specialised sabre-tooths. Homo-
therium in particular is likely to have found it difficult toadopt a solitary lifestyle taking a high proportion ofrelatively small prey in forest environments during someof the warmer climatic stages in Europe, something thatthe lions apparently managed to do as inferred formtheir continued presence in all phases of the EuropeanPleistocene (Turner, 1995b).The recent discovery of a mandible of Homotherium
dated to about 28,000 yr BP in the North Sea (Reumeret al., 2003) deserves special comment in any discussionof extinctions. Until recently, a terminal date of around400,000–500,000 years for Homotherium in Europe waswidely accepted (Turner and Anton, 1999). If, as seemslikely, the new date is correct, then two quite differentpossibilities are raised. The first would establish thecontinued presence of Homotherium in Europe andstretch the co-existence with lions there for a further350,000 years or so. The second would see the find as
evidence for an immigration of North Americanrepresentatives via Beringia. Many other large mammalshave such a late Pleistocene Holarctic distribution(Kurten and Anderson, 1980), and it is worth pointingout that although the mandible is attributed to the OldWorld species H. latidens is also morphologicallyindistinguishable from the American species H. serum.At present we cannot choose between these twoalternatives, but it is worth pointing out that in eitherevent the animal must have led a very quiet existence,for it managed to escape incorporation in other depositsover that time. This lack of other confirmed records forthe latter part of the Middle Pleistocene and into theLate Pleistocene would strongly suggest that the lionwas the dominant large felid in most if not all parts ofEurope.What happens if a further intelligent predator, one
equipped mentally and technologically for group action,is then added to that mix?
7. Machairodonts and hominins in the European Middle
Pleistocene
The importance of machairodontines as providers ofresources for such scavengers as hyaenids and homininshas been a matter of long debate. The idea thatmachairodonts were unable to clean the bones of meatbecause of the shape of their teeth was largely falsifiedby the study of Marean and Ehrhardt (1995) of aHomotherium den in Friesenhahn Cave, Texas, wheretooth marks on elephant bones showed that homothereswere able to use their protracted incisor battery, ascould anyway be inferred from the simple observation ofhomotherine dental morphology. It remains true, none-theless, that Homotherium and all machairodontineswere ill-adapted to break bones, and were likely to leavebehind most of the in-bone nutrients of their kills. Thatwould be an attractive resource to both hyaenids andhominins.Were the remains of the kills of Homotherium
therefore an important resource for hominins? Marean(1989) postulated three key factors bearing on that issue:
1.
The large size of prey, allegedly larger than that ofmodern lions.2.
The inability of Homotherium to consume bone, thusleaving in-bone nutrients behind.3.
The possibility that kills were made in riparianwoodlands, where detection by non-hominin scaven-gers would take longer.The first is not supported by the results of this study,since we argue that Homotherium was individually nostronger than a lion and that the key to understandingthe scimitar-tooth adaptation was not so much the

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1299
killing of very large prey but a higher efficiency in killinganimals within the higher range of the lion’s prey-sizespectrum. For the second, homotheres would indeed belikely to leave most in-bone nutrients behind, but in thatthey did not differ greatly from modern lions, whichhave very sectorial cheek teeth and rarely, if ever, breaklong bones for marrow. The third factor is notsupported by the results of this study because the post-cranial morphology of Homotherium points strongly toa semi-cursorial adaptation, which in turn is suggestiveof more open-habitat preferences. This is in agreementwith independent studies by Lewis (1997) and Anyonge(1996).Thus it seems likely that while Homotherium may not
have been a more prolific producer of carcasses or ofcarcasses with significantly more meat for a scavengerthan other large cats, those carcasses are likely tohave lain in more open terrain and been quite accessibleto a scavenger. As such, they may therefore haverepresented an attractive target for a range of otherspecies, including humans. But a further point toremember is that social cats are more effective atdefending their kills than solitary cats. If our argumentsfor group living in homotheres are valid, then thechances for a troop of early African hominins gainingaccess to a Homotherium kill through confrontationalscavenging seems remote. Homotherium would be aboutas good—or as bad—a provider of carcasses for earlyaustralopithecine, or australopithecine-like, hominins asmodern lions.However, the situation would have been different in
Middle Pleistocene Europe, where Homo heidelbergensis
was more organised and wielded a more impressiveweaponry and tool kit. Pre-Neanderthals would notonly have been more capable of displacing Homotherium
from its kills than earlier hominins, but they would alsohave had a greater impact on resource availability intheir role as active hunters. As the archaeological recordimproves it seems likely that the origins of effectivehunting will be pushed back further and further(Roebroeks, 2001). This in itself should not surpriseus; while previous arguments for scavenging as a majorprocurement strategy (Turner, 1992) remain strong inour view, it is clear that the environmental pressures onhuman occupants of Middle Pleistocene Europe wouldstrongly favour the development of any advantages thatcould be gained as an intelligent, technologicallyequipped predator. If Homotherium was under pressurefrom changes in its optimum habitat and approachingthe brink around 400,000 years ago, then the activities ofhumans added to those of lions may have been sufficientto push it over the edge, as the bulk of the fossil recordsuggests. If on the other hand it did manage to surviveor recolonise Europe, as the North Sea find wouldsuggest, it would be a sad irony if it managed to survivefor an additional 350,000 years only to disappear soon
after the Late Pleistocene arrival of the first anatomi-cally modern humans in the continent.
8. Conclusion
The comparison of elements of the post-cranialskeleton of Homotherium latidens from Incarcal withthe same elements in modern big cats and in other sabre-toothed taxa reveals the presence of moderate cursorialadaptations. Homotherium latidens had shortened lum-bar vertebrae; a slender humerus with an elevatedgreater tuberosity, a narrow distal end and a verticaland deep olecranon fossa; a straight radius with anarrow distal end; a shortened robust pisiform; smallclaws on digits II–V with reduced retraction, a largedew-claw; a shortened calcaneus and a flattened, short-necked astragalus (Fig. 7). This set of morphologicalfeatures indicates an adaptation for efficient locomotionat moderate speeds, with less ability for jumping andsudden acceleration than in pantherine cats, and withrelatively less power to subdue large prey individually.Combined with the evidence from cranio-cervicalanatomy of adaptations to kill large prey, the post-cranial evidence suggests that Homotherium wouldconcentrate on prey animals up to the size of an adultbovine or a young proboscidean, and that the largerprey animals would probably be hunted by means ofgroup action.The hunting technique inferred for Homotherium
would work best in relatively open environments, butfaced with the competition from the large Pleistocenelions it would benefit from the protection provided bythe presence of some vegetational cover. The MiddlePleistocene extinction of Homotherium in Europe afterabout 200,000 years of co-existence with lions, assuggested by the bulk of the fossil evidence, may beattributed to a combination of climatic-induced vegeta-tional changes, exacerbated competition among thelarge carnivores and increased pressure from Homo
heidelbergensis as a new member of the predatory guild.If extinction took place during the last glaciation assuggested by the recent find, then it would appear thatHomotherium survived the overwhelming dominance oflions for an additional 300,000 years, only to succumbshortly after the arrival of the first modern humans.
Acknowledgements
We thank the curators of the Museu ArqueologicComarcal de Banyoles, Josep Tarrus, and the Museu deZoologia de Barcelona, Eulalia Garcia, for access totheir collections and facilities for study. This work hasbeen supported by project BOS2001-1004 of theMinisterio de Ciencia y Tecnologia of Spain.

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–13011300
References
Agustı, J., Anton, M., 2002. Mammoths, Sabertooths and Hominids.
Columbia University Press, New York.
Akersten, W., 1985. Canine function in Smilodon (Mammalia, Felidae,
Machairodontinae). Los Angeles County Museum Contributions
in Science 356, 1–22.
Altuna, J., 1972. Fauna de mamıferos de los yacimientos prehistoricos
de Guipuzcoa. Munibe 24 (1–4), 1–464.
Anton, M., 2003. Notes on the reconstructions of fossil vertebrates
form Lothagam. In: Leakey, M.G., Harris, J. (Eds.), Lothagam:
Dawn of Humanity in East Africa. Columbia University Press,
New York, pp. 661–665.
Anton, M., Galobart, A., 1999. Neck function and predatory behavior
in the scimitar toothed cat Homotherium latidens (Owen). Journal
of Vertebrate Paleontology 19 (4), 771–784.
Anyonge, W., 1993. Body mass in extinct and extant carnivores.
Journal of Zoology of London 231, 339–350.
Anyonge, W., 1996. Locomotor behaviour in Plio-Pleistocene sabre-
tooth cats: a biomechanical analysis. Journal of Zoology, London
238, 395–413.
Ballesio, R., 1963. Monographie d’un Machairodus du gisement
villafranchien de Seneze: Homotherium crenatidens Fabrini. Tra-
vaux du Laboratoire de Geologie de la Faculte de Sciences de
Lyon, Nouveau Series 9, 1–129.
Barone, R., 1967. La myologie du lion (Panthera leo). Mammalia 31,
459–514.
Barone, R., 1989. Anatomie Comparee des Mammiferes Domestiques,
Tome 1 (Osteologie) Tome 2 (Arthrologie et myologie). Vigot,
Paris.
Bicknevicius, A.R., Van Valkenburgh, B., Walker, J., 1996. Incisor size
and shape: implications for feeding behaviors in saber-toothed
‘‘cats’’. Journal of Vertebrate Paleontology 16 (3), 510–521.
Boule, M., 1906. Les grands chats des cavernes. Annales de
Paleontologie Tome 1 (fascicle 1 and 2).
Brain, C.K., 1981. The Hunters or the Hunted? University of Chicago
Press, Chicago.
Bryant, H.N., Russell, A.P., 1992. The role of phylogenetic analysis in
the inference of unpreserved attributes of extinct taxa. Philosophi-
cal Transactions of the Royal Society of London B 337, 405–418.
Bryant, H.N., Seymour, K., 1990. Observations and comments on the
reliability of muscle reconstruction in fossil vertebrates. Journal of
Morphology 206, 109–117.
Bryant, H.N., Russell, A.P., Laroyia, B., Powell, G.L., 1996. Claw
retraction and protraction in the Carnivora: skeletal microvaria-
tion in the phalanges of the Felidae. Journal of Morphology 229,
289–308.
Caro, T.M., 1994. Cheetahs of the Serengeti Plains: Group Living in
an a-Social Species. University of Chicago Press, Chicago.
Colomer, F., Gaete, R., Galobart, A., Maroto, J., Menendez, E., Ros,
X., 1990. El jaciment paleontologic d’Incarcal (Crespia, Pla de
l’Estany). El Medi Natural del Valles (III Colloquis del Natur-
alistes Vallesans) 3, 26–32.
Creel, S., Creel, N.M., 1996. Limitation of African wild dogs by
competition with larger carnivores. Conservation Biology 10,
526–538.
Creel, S., Creel, N.M., 2002. The African Wild Dog: Behavior,
Ecology and Conservation. Princeton University Press, Princeton.
Dennell, R., 2003. Dispersal and colonisation, long and short
chronologies: how continuous is the Early Pleistocene record for
hominids outside East Africa? Journal of Human Evolution 45,
421–440.
Dominguez-Rodrigo, M., 2001. A study of carnivore competition in
riparian and open habitats of modern savannas and its implications
for hominid behavioral modelling. Journal of Human Evolution 40
(2), 77–98.
Evans, H.E., Christensen, C., 1976. Miller’s Anatomy of the Dog,
second ed. W.B. Saunders Co., Philadelphia.
Freudenberg, W., 1914. Die Saugetiere des alteren quartars von
mitteleuropa. Geologische und Palaeontologische Abhandlungen,
Neueus Folge 12, 455–671.
Galobart, A., 1996. Estudi de la fauna de mamıfers dels jaciments del
pleistoce inferior d’Incarcal (Crespia, Pla de l’Estany): sistematica,
tafonomia i paleoecologia. Ph.D. Dissertation, Universitat Auton-
oma de Barcelona, Barcelona.
Galobart, A., 2003. Origin de las acumulaciones, diversidad y
reconstruccion de los yacimientos de Incarcal I e Incarcal V.
Paleontologia I Evolucio 34, 249–260.
Galobart, A., Maroto, J., Menendez, E., Ros, X., Gaete, R., Colomer,
F., 1990. El yacimiento del Pleistoceno Inferior de Incarcal
(Crespia, Girona). Comunicaciones Reunion de TafonomIa y
Fosilizacion, Madrid, 161–167.
Galobart, A., Maroto, J., Ros, X., 1996. La faunas cuaternarias de
mamiferos de la Cuenca Banyoles-Besalu (Girona). Revista
Espanola de Paleontologia. Extraordinario, 248–255.
Galobart, A., Anton, M., Maroto, J., 2003. Hienidos y canidos de los
yacimientos de Incarcal (Girona, NE Peninsula Iberica). Una
aproximacion a la paleobiologia del Pleistoceno inferior. Paleon-
tologia I Evolucio 34, 69–88.
Gambaryan, P.P., 1977. How Mammals Run: Anatomical Adapta-
tions. Halsted Press, London.
Garcıa, N., 2002. Los carnıvoros de los yacimientos Pleistocenos de la
Sierra de Atapuerca. Ph.D. Dissertation, Departamento de
Biologıa Animal I, Facultad de Ciencias Biologicas, Universidad
Complutense de Madrid.
Ginsburg, L., 1961. Plantigradie et digitigradie chez les carnivores
fissipedes. Mammalia 25, 1–21.
Gonyea, W.J., 1978. Functional implications of felid forelimb
anatomy. Acta Anatomica 102, 111–121.
Gorman, M.L., Mills, M., Raath, J.P., Speakman, J.R., 1998. High
hunting costs make African wild dogs vulnerable to kleptoparasit-
ism by hyaenas. Nature 391, 479–481.
Guthrie, R.D., 1990. Frozen Fauna of the Mammoth Steppe: The
Story of Blue Babe. University of Chicago Press, Chicago.
Harrison, J.A., 1983. The Carnivora of the Edson local fauna (Late
Hemphilian), Kansas. Smithsonian Contributions in Paleobiology
54, 1–42.
Hemmer, H., 2002. Die Feliden aus dem Epivillafranchium von
Untermassfeld. In: Kahlke, R.-D. (Ed.), Das Pleistozan von
Untermassfeld Bei Meiningen (Thuringen). Romisch-Germa-
nisches Zentralmuseum Bande 40(3), 699–782.
Hildebrand, M., 1989. Analysis of Vertebrate Structure. Wiley, New
York.
Janossy, D., 1990. Vertebrate fauna of site II. Vertesszollos man site
and culture. Akademiai Kiado, Budapest.
Johnsingh, A.J.T., 1992. Prey selection in three large sympatric
carnivores in Bandipur. Mammalia 56 (4), 517–526.
Joubert, D., 1994. Lions of darkness. National Geographic Magazine
186 (2), 35–53.
Joubert, D., Joubert, B., 1997. The Lions of Savuti: Hunting with the
Moon. National Geographic Society, Washington, DC.
Kitchener, A., 1991. The Natural History of the Wild Cats. New York,
Cornell University.
Koenigswald, W.V., Heinrich, W.-D., 1999. Mittelpleistozane Sauge-
tierfaunen aus Mitteleuropa-der Versuch einer biostratigrafischen
Zuordnung. Kaupia 9, 53–112.
Kurten, B., 1952. The Chinese Hipparion fauna: a quantitative survey
with comments on the ecology of the machairodonts and hyaenids
and the taxonomy of the gazelles. Commentationes Biologiaes
Societas Scientificas Fennica 13 (4), 1–82.
Kurten, B., Anderson, E., 1980. Pleistocene Mammals of North
America. Columbia University Press, New York.

ARTICLE IN PRESSM. Anton et al. / Quaternary Science Reviews 24 (2005) 1287–1301 1301
Kurten, B., Werdelin, L., 1990. Relationships between North and
South American Smilodon. Journal of Vertebrate Paleontology 10
(2), 158–169.
Leyhausen, P., 1979. Cat Behavior. Garland STPM Press, New York.
Lewis, M.E., 1997. Carnivoran paleoguilds of Africa: implications for
hominid food procurement strategies. Journal of Human Evolution
32, 257–288.
MacDonald, J.R., 1948. A new species of Pseudaelurus from the Lower
Pliocene of Nebraska. University of California Publications
Bulletin of the Department of Geological Sciences 28 (2), 45–52.
Marean, C.W., 1989. Sabertooth cats and their relevance for early
hominid diet and evolution. Journal of Human Evolution 18,
559–582.
Marean, C.W., Ehrhardt, C.L., 1995. Paleoanthropological and
paleological implications of the taphonomy of a sabretooth’s den.
Journal of Human Evolution 29, 515–547.
Martin, L.D., 1980. Functional morphology and the evolution of cats.
Transactions of the Nebraska Academy of Sciences 8, 141–154.
Martin, L.D., 1989. Fossil history of the terrestrial carnivora. In:
Gittleman, J. (Ed.), Carnivore Behavior, Ecology and Evolution.
Chapman & Hall, London, pp. 535–568.
Meade, G.E., 1961. The saber toothed cat, Dinoblastis serus. Texas
Memorial Museum 3, 23–60.
Merriam, J.C., Stock, C., 1932. The Felidae of Rancho La Brea.
Carnegie Institution of Washington Publications 442, 1–231.
Mills, M.G.L., Biggs, H.C., 1993. Prey apportionment and related
ecological relationships between large carnivores in Kruger
National Park. Symposium of the Zoological Society of London
65, 253–268.
Nunez, R., Miller, B., Lindzey, F., 2000. Food habits of jaguars and
pumas in Jalisco, Mexico. Journal of Zoology 252, 373–379.
Packer, C., 1986. The ecology of sociability in felids. In: Rubenstein,
D.I., Wranghan, R.W. (Eds.), Ecological Aspects of Social
Evolution: Birds and Mammals. Princeton University Press,
Princeton, NJ, pp. 429–451.
Pilgrim, G.E., 1931. Catalogue of the Pontian Carnivora of Europe in
the Departament of Geology. British Museum of Natural History,
London.
Pocock, R.I., 1939. The Fauna of British India. Mammalia, vol. I.
Junk, The Hague.
Pratt, A.E., 1990. Taphonomy of the large vertebrate fauna from the
Thomas Farm locality (Miocene, Hemingfordian), Gilchrist
County, Florida. Bulletin of the Florida Museum of Natural
History, Biological Sciences 35 (2), 35–130.
Rawn-Schatzinger, V., 1992. The scimitar cat Homotherium serum
Cope. Osteology, functional morphology, and predator behavior.
Illinois State Museum Reports of Investigation 47, 1–80.
Reumer, J.F.W., Rook, L., Van der Borg, K., Post, K., Moll, D., De
Vos, J., 2003. Late Pleistocene survival of the saber-toothed cat
Homotherium in Northwestern Europe. Journal of Vertebrate
Paleontology 23 (1), 260–262.
Roebroeks, W., 2001. Hominid behaviour and the earliest occupation
of Europe: an exploration. Journal of Human Evolution 41,
437–461.
Ros, X., Palomar, J., Galobart, A., Maroto, J., 2003. Contexto
geografico y geologico regional y local de los yacimientos del plio-
pleistoceno de Incarcal (Girona, NE Peninsual Iberica). Paleonto-
logia I Evolucio 34, 5–14.
Schaller, G.B., 1972. The Serengeti Lion. University of Chicago Press,
Chicago.
Schaub, S., 1925. Uber die osteologie von Machaerodus cultridens
Cuvier. Ecologae Geologica Helvetiae 19 (1), 255–266.
Seidensticker, J.C., 1976. On the ecological separation between tigers
and leopards. Biotropica 8, 225–234.
Seidensticker, J., McDougal, C., 1993. Tiger predatory behaviour,
ecology and conservation. Symposia of the Zoological Society of
London 65, 105–125.
Spoor, C.F., Badoux, D.M., 1986. Descriptive and functional myology
of the neck and forelimb of the striped hyena (Hyaena hyaena, L.
1758). Anatomischer Anzeiger, Jena 161, 375–387.
Stander, P.E., Albon, S.D., 1993. Hunting success of lions in a semi-
arid environment. Symposia of the Zoological Society of London
65, 127–148.
Sunquist, M.E., Sunquist, F.C., 1989. Ecological constraints on
predation by large felids. In: Gitlemann, J.L. (Ed.), Carnivore
Behavior, Ecology and Evolution. Chapman & Hall, London,
pp. 220–245.
Teilhard de Chardin, P., 1936. Fossil Mammals from Locality 9 of
Choukoutien. Palaeontologia Sinica Series C, vol. VII (fascicle 4),
pp. 5–71.
Turner, A., 1992. Large carnivores and earliest European hominids:
changing determinants of resource availability during the Lower
and Middle Pleistocene. Journal of Human Evolution 22, 109–126.
Turner, A., 1995a. The Villafranchian large carnivore guild: geo-
graphic distribution and structural evolution. Il Quaternario 8,
349–356.
Turner, A., 1995b. Evidence for Pleistocene contact between the
British Isles and the European continent based on distributions of
larger carnivores. Geological Society Special Publication 96,
141–149.
Turner, A., 1997. Further remains of Carnivora (Mammalia) from the
Sterkfontein hominid site. Paloentologia Africana 34, 115–126.
Turner, A., 1999a. Assessing earliest human settlement of Eurasia:
Late Pliocene dispersions from Africa. Antiquity 73, 563–570.
Turner, A., 1999b. Larger carnivores (Mammalia, Carnivora) from
Westbury-sub-Mendip, Somerset. In: Andrews, P., Cook, J.,
Currant, A., Stringer, C. (Eds.), Westbury Cave: The Natural
History Museum Excavations 1976–1984. Western Academic &
Specialist Press, Bristol, pp. 175–193.
Turner, A., Anton, M., 1996. The giant Hyaena Pachycrocuta
brevirostris (Mammalia, Carnivora, Hyaenidae). Geobios 29,
455–468.
Turner, A., Anton, M., 1997. The Big Cats and their Fossil Relatives.
Columbia University Press, New York.
Turner, A., Anton, M., 1999. Climate and evolution: implications of
some extinction patterns in African and European machairodon-
tine cats of the Plio-Pleistocene. Estudios Geologicos 54, 209–230.
Turner, A., Anton, M., 2004. Evolving Eden. An Illustrated Guide to
the Evolution of the African Large Mammal Fauna. Columbia
University Press, New York.
Turner, A., O’Regan, H., 2002. The assessment of size in fossil Felidae.
Estudios Geologicos 58, 45–54.
Werdelin, L., 1996. Carnivoran ecomorphology: a phylogenetic
perspective. In: Gittleman, J. (Ed.), Carnivore Behavior, Ecology
and Evolution, vol. 2. Cornell University Press, Ithaca, NY,
pp. 582–624.
Werdelin, L., 2003. Mio-Pliocene Carnivora from Lothagam, Kenya.
In: Leakey, M.G., Harris, J. (Eds.), Lothagam: Dawn of Humanity
in East Africa. Columbia University Press, New York,
pp. 261–328.
Werdelin, L., Lewis, M.E., 2001. Diversity and turnover in Eastern
African Plio-Pleistocene Carnivora. Journal of Vertebrate Paleon-
tology 21 (3), 112A (Abstract).
Witmer, L.M., 1995. The extant phylogenetic bracket and the
importance of reconstructing soft tissues in fossils. In: Thomason,
J.J. (Ed.), Functional Morphology in Vertebrate Paleontology.
University of Cambridge Press, Cambridge, pp. 19–33.
Related Documents











