of January 15, 2019. This information is current as Immunotherapy CMV-Specific TCR-Transgenic T Cells for Hammerschmidt and Andreas Moosmann Andrea Schub, Ingrid G. Schuster, Wolfgang http://www.jimmunol.org/content/183/10/6819 doi: 10.4049/jimmunol.0902233 October 2009; 2009; 183:6819-6830; Prepublished online 28 J Immunol References http://www.jimmunol.org/content/183/10/6819.full#ref-list-1 , 33 of which you can access for free at: cites 54 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2009 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on January 15, 2019 http://www.jimmunol.org/ Downloaded from by guest on January 15, 2019 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of January 15, 2019.This information is current as
ImmunotherapyCMV-Specific TCR-Transgenic T Cells for
Hammerschmidt and Andreas MoosmannAndrea Schub, Ingrid G. Schuster, Wolfgang
http://www.jimmunol.org/content/183/10/6819doi: 10.4049/jimmunol.0902233October 2009;
2009; 183:6819-6830; Prepublished online 28J Immunol
Referenceshttp://www.jimmunol.org/content/183/10/6819.full#ref-list-1
, 33 of which you can access for free at: cites 54 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 15, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

CMV-Specific TCR-Transgenic T Cells for Immunotherapy1
Andrea Schub,* Ingrid G. Schuster,† Wolfgang Hammerschmidt,* and Andreas Moosmann2*‡
Reactivation of CMV can cause severe disease after allogeneic hemopoietic stem cell transplantation. Adoptive T cell therapy wassuccessfully used for patients who had received transplants from CMV-positive donors. However, patients with transplants fromCMV-negative donors are at highest risk, and an adoptive therapy is missing because CMV-specific T cells are not available fromsuch donors. To address this problem, we used retroviral transfer of CMV-specific TCR genes. We generated CMV-specific T cellclones of several HLA restrictions recognizing the endogenously processed Ag pp65. The genes of four TCRs were cloned andtransferred to primary T cells from CMV-negative donors. These CMV-TCR-transgenic T cells displayed a broad spectrum ofimportant effector functions (secretion of IFN-� and IL-2, cytotoxicity, proliferation) in response to endogenously processed pp65and could be enriched and expanded by strictly Ag-specific stimulation. Expansion of engineered T cells was accompanied by anincrease in specific effector functions, indicating that the transferred specificity is stable and fully functional. Hence, we expectthese CMV-TCR-transgenic T cells to be effective in controlling acute CMV disease and establishing an antiviral memory. TheJournal of Immunology, 2009, 183: 6819–6830.
R eactivation of CMV is an important cause of disease anddeath after allogeneic hemopoietic stem cell transplanta-tion (allo-HSCT).3 Antiviral medication is accompanied
by severe side effects and is often ineffective due to the develop-ment of resistant virus strains. Furthermore, antiviral drugs oftenonly delay CMV-associated disease (1). The development ofCMV-related disease is strongly correlated with absent or delayedreconstitution of CMV-specific T cells (1–5). Thus, current ther-apeutic concepts focus on the reconstitution of the CMV-specificcellular immune response. Adoptive transfer of CMV-specific cy-totoxic T cell clones (6) and T cell lines (7–9) has been veryeffective in treating and preventing CMV-related disease (10). Inthese studies, the transferred T cells had been isolated from CMV-positive HSCT donors. With CMV-seronegative donors, isolationand enrichment of CMV-specific T cells are difficult, if not im-possible, because naive CMV-specific T cells are too rare to bedetected or directly accessed (11, 12). As a consequence, CMV-seropositive recipients of stem cell grafts from CMV-negative do-nors are at particular risk to develop severe CMV-related diseasebecause CMV-specific T cell reconstitution is highly deficient (5,13). To prepare T cells with specificities absent from the donor’srepertoire, TCR gene transfer is a promising option (14–17). Re-
cently, clinical application of TCR-transgenic T cells in malignantmelanoma patients resulted in tumor remissions and was well tol-erated (18). In contrast, TCR-transgenic T cells for antiviral ther-apy have not yet been used in the clinic. Adoptive transfer ofCMV-TCR-transgenic T cells should be especially suitable forclinical application, because CMV-specific TCRs, which were se-lected in vivo for antiviral function and maintenance, are readilyaccessible from the T cell memory of CMV-seropositive donors.Healthy individuals with persistent CMV infection carry large pro-portions of CMV-specific CD8� and CD4� memory T cells (19,20). Many epitopes derived from the immunodominant CMV Agspp65, IE-1, and others, which bind to various HLA molecules,have been characterized (19, 21, 22). Several CMV-specific TCRchain sequences, some of which are shared by various donors(public TCRs), are already known (23–31).
Previously, we showed that Ag-specific T cell lines and clonescan be efficiently generated, expanded, and tested using B cell-based Ag presentation systems (22, 32, 33). In this study, we usedB cells to establish CMV-specific CD8� T cell clones of variousHLA restrictions which recognize endogenously processed CMVAg. Their TCRs were cloned and transferred to primary T cellsfrom CMV-negative donors, and the resulting TCR-transgenic Tcells were analyzed for various effector functions. We tested theircapacity to selectively expand in response to endogenously pro-cessed Ag, and investigated their reactivity spectrum after repeatedAg contact to ensure the stability of their Ag-specific function.Taken together, we intend to show that CMV-TCR-transgenic Tcells are promising tools for adoptive T cell therapy.
Materials and MethodsThe standard cell culture medium was RPMI 1640 (Invitrogen) supple-mented with 10% FCS (PAA Laboratories), 100 U/ml penicillin/100 �g/mlstreptomycin (Invitrogen), and 100 nM sodium selenite (ICN Biochemi-cals). 293T cells were cultivated in DMEM (Invitrogen) with the samesupplements.
Cells and cell lines
PBMCs from healthy donors (Table I) were collected with donors’ in-formed consent following the requirements of the local ethical board andthe principles expressed in the Helsinki Declaration. PBMCs were obtainedby centrifugation on Ficoll/Hypaque (Biochrom). HLA typing was per-formed by PCR-based methods (IMGM). Stably pp65-expressing mini-lymphoblastoid cell lines (mLCL) and control (pp65-negative) mLCLs
*Department of Gene Vectors, Helmholtz Zentrum Munchen, Munich, Germany;†Institute of Molecular Immunology, Helmholtz Zentrum Munchen, Munich, Ger-many; and ‡Clinical Cooperation Group Molecular Oncology, Helmholtz ZentrumMunchen and Ludwig-Maximilians-Universitat Munchen, Munich, Germany
Received for publication July 13, 2009. Accepted for publication September 11, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Deutsche Forschungsgemeinschaft Grants SFB-Tran-sregio 36 and SFB 455 and by the Helmholtz Alliance on Immunotherapy of Cancerfunded by the Initiative and Networking Fund of the Helmholtz Association.2 Address correspondence and reprint requests to Dr. Andreas Moosmann, ClinicalCooperation Group Molecular Oncology, Helmholtz Zentrum Munchen, Marchionin-istrasse 25, 81377 Munich, Germany. E-mail address: [email protected] Abbreviations used in this paper: allo-HSCT, allogeneic hemopoietic stem celltransplantation; mLCL, mini-lymphoblastoid cell line; BBL, CD40-activated B blast;J76, Jurkat 76; J76CD8, Jurkat 76 stably expressing human CD8�; MLV, murineleukemia virus; D, donor; R, recipient; CD62L, L-selectin.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902233
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
were generated by infection of PBMCs with B cell-transforming mini-EBVvectors (32). CD40-activated B-blast (BBL) cultures were established asdescribed (33) and maintained by weekly replating PBMCs on irradiated(140 Gy) murine fibroblasts, stably expressing the human CD40L, in thepresence of 2 ng/ml rIL-4 (R&D Systems). The TCR��-deficient T celllines Jurkat76 (J76) (34) and J76 stably expressing the human CD8� chain(J76CD8; kindly provided by W. Uckert, Max-Delbruck-Center, Berlin,Germany) were used as recipient cells for TCR transfer studies. 293T cellswere used for packaging of the retroviral vectors.
Generation of CMV-specific T cell clones
We generated T cells against the following HLA class I-restricted epitopesfrom pp65: NLVPMVATV, aa 495–503, HLA-A*0201-restricted (abbre-viated NLV); IPSINVHHY, aa 123–131, HLA-B*3501-restricted (IPS);YSEHPTFTSQY, aa 363–373, HLA-A*0101-restricted (YSE).
YSE-specific T cells from donor 7 (Table I) were reactivated withinPBMCs by stimulation with the irradiated (50 Gy) autologous pp65-ex-pressing mLCLs (32). Per well of a 12-well plate, 6 � 106 PBMCs and1.5 � 105 pp65-expressing mLCLs were cocultivated in 3 ml of medium.On day 10 and then every 7 days, cells were pooled, counted using trypanblue staining, and replated at 3 � 106 cells/3 ml of medium per well,adding freshly irradiated (50 Gy) pp65 mLCL as stimulators at an effector-stimulator ratio of 3:1 and 100 U/ml rIL-2 (Proleukin S; Novartis). At day50 of culture, T cell clones were generated by limiting dilution.
For single-cell cloning, 0.5 or 3 T cells/well were seeded into 96-wellround-bottom plates. 2 � 104/well irradiated (50 Gy) autologous pp65-expressing mLCL, 3 � 105/well of a mixture of irradiated (50 Gy) allo-geneic PBMCs from three different donors, and 1000 U/ml IL-2 wereadded. Restimulations of T cell clones were performed every 2 wk, andoutgrowing clones were expanded in 96-well round-bottom plates underthe same conditions.
For reactivation of NLV- or IPS-specific T cells from donor 6 or 8(Table I), PBMCs were pulsed with 5 �g/ml of the relevant CMV peptide(JPT), and plated at 8 � 106 cells/4 ml of medium per well in a 12-wellplate. After 24 h at 37°C and 5% CO2, the cells were washed three timeswith PBS, and replated in the 12-well plate, and supplemented with 10U/ml IL-2. On day 8, the rIL-2 concentration was raised to 50 U/ml. TheT cell culture was expanded according to cell proliferation. At day 14, Tcell clones were generated as described above.
TCR analysis
TCR-PCR analysis of CMV-specific T cell clones was performed as pre-viously described (35). Total RNA from T cell clones was extracted usingthe RNeasy Mini Kit (Qiagen). cDNA was synthesized using murine leu-kemia virus (MLV)-reverse transcriptase and oligodeoxythymidylateprimer (MBI-Fermentas). Subfamily-specific PCR of the TCR �-chain wasperformed with 34 different V� primers (36). The TCR �-chain was ana-lyzed by PCR with degenerated V� primers as previously described (37).Specifically amplified gene products were analyzed by DNA sequencing(Sequiserve). TCR nomenclature according to IMGT was used (38).
Cloning of retroviral vectors containing the CMV-specific TCRchain genes
TCR �- and �-chain genes were separately inserted into the myeloprolif-erative sarcoma virus-derived vector MP71Gpre (39) as previously de-scribed (40). Briefly, specific TCR �- and �-chain genes were amplifiedfrom cDNA isolated from CMV-specific T cell clones by specific primerscontaining 5�-NotI and 3�-EcoRI restriction sites, and cloned as single TCR
chain genes into the vector MP71Gpre (MP71-TCR� and MP71-TCR�)replacing the GFP gene. The GFP-encoding MP71Gpre vector was used ascontrol to evaluate infection efficiencies. All TCR cassettes used in thisstudy were verified by DNA sequencing (Sequiserve).
Retroviral TCR transfer
To produce amphotropic MLV-pseudotyped retroviruses, 293T cells werecotransfected by calcium phosphate precipitation with expression plasmidsencoding the Moloney MLV gag/pol genes (pcDNA3.1MLVg/p) and theMLV-10A1 env gene (pALF-10A1) together 1) with the respective TCR-encoding retroviral vector plasmids MP71-TCR� and MP71-TCR�, or 2)with the GFP-encoding plasmid MP71Gpre (transfection and transductioncontrol), or 3) without retroviral vector plasmids (mock control) (41). For-ty-eight hours after transfection, the retroviral supernatant was harvested,filtered (0.45 �m pore size), and used directly for infection of PBMCs, J76cells, and J76CD8 cells. PBMCs (106 in 1 ml), which had been activatedwith 50 U/ml rIL-2 and 50 ng/ml anti-CD3 Ab (OKT-3, kindly provided byE. Kremmer, Helmholtz Zentrum, Munich, Germany) 2 days earlier, weretransduced in 24-well plates precoated with 5 �g/well RetroNectin(Takara) in the presence of 4 �g/ml protamine sulfate (MP Biomedicals)and 100 U/ml rIL-2. After addition of 1 ml of retrovirus-containing super-natant, the plates were spinoculated for 2 h at 800 � g and 32°C. J76 andJ76CD8 cells (105 in 1 ml) were transduced as described above but withoutadding rIL-2. Medium was replaced after 24 h.
Restimulation of TCR-transduced cells
Weekly restimulation of transduced PBMCs was started 6 days after TCRtransfer. For Ag-specific stimulation, transduced T cells (1.5 � 106/1.5ml/well) were cocultivated with irradiated (50 Gy) autologous pp65-ex-pressing mLCL at a ratio of 4:1 with 100 U/ml rIL-2 in 24-well plates. Fornonspecific stimulation, transduced T cells (1.5 � 106/1.5 ml/well) werecocultivated with 1.5 � 106 per well of a mixture of irradiated (50 Gy)allogeneic PBMCs from three different donors and 1.5 � 105/well irradi-ated (50 Gy) autologous pp65-negative mLCL with 100 U/ml rIL-2 and 30ng/ml OKT-3 in 24-well plates.
Flow cytometry
Multimer staining was performed by incubating the T cells for 10 min atroom temperature with PE-labeled HLA/peptide tetramer or unlabeledHLA/peptide pentamer. The cells were counterstained on ice for 15 minwith anti-CD4-FITC, anti-CD3-PE-Cy5, anti-CD8-APC Abs (all BDPharmingen), and, in the case of unlabeled pentamers, with Pro5 FluorotagR-PE (Proimmune). Directly after staining, the cells were fixed by 1.6%formaldehyde (Carl Roth). As control, T cells were stained as explainedabove but without adding the respective multimer. PE-labeled NLV/A*0201 tetramer was purchased from Beckman Coulter; unlabeled IPS/B*3501 and YSE/A*0101 pentamers were purchased from Proimmune.Cells were analyzed on a BD Biosciences FACSCalibur flow cytometer.Data analysis was performed using FlowJo 8.8.4 software (Tree Star). Foranalysis, viable lymphocytes were gated in a forward-sideward scatter dotplot. All shown dot plots span, on both coordinates, a range from 1 to10,000 arbitrary units of fluorescence intensity in a logarithmic scale.
T cell effector assays
CMV-specific T cell clones and TCR-transduced PBMCs were analyzedfor cytokine secretion by ELISA. Effector cells (104) were cocultivatedovernight with target cells (2 � 104) in 200 �l/well of a 96 V-well plate at37°C and 5% CO2. Then supernatants were harvested, and IFN-� and IL-2
Table I. HLA types and virus carrier state of donors
Donor HLA-A HLA-B HLA-C CMV Serostatus EBV Serostatus
1 *0201, *2502 *1503, *51 *1203, *1402 Nega Pos2 *0201, – *44, *51 *02, *05 Neg Pos3 *0101, *2601 *3501, *5701 *0401, *0602 Neg Neg4 *03, *24 *3501, *4002 *02, *04 Neg Pos5 *01, *0201 *1501, *37 *03, *06 Neg Pos6 *0201, – *3503, *5701 *04, *06 Pos Pos7 *01, *11 *08, *1501 *03, *07 Pos Pos8 *02, *03 *35, *4001 *03, *15 Pos Pos9 *02, *26 *07, *38 *07, *12 N.D. N.D.10 *02, *34 *14, *44 *04, *08 N.D. N.D.
Neg, Negative; Pos, positive; N.D., not determined.
6820 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

ELISAs were performed according to the manufacturer’s recommendations(Mabtech).
Cytotoxicity of TCR-transduced PBMCs was analyzed by calcein re-lease assay as described previously (33).
ResultsGeneration of CMV pp65-specific T cell clones
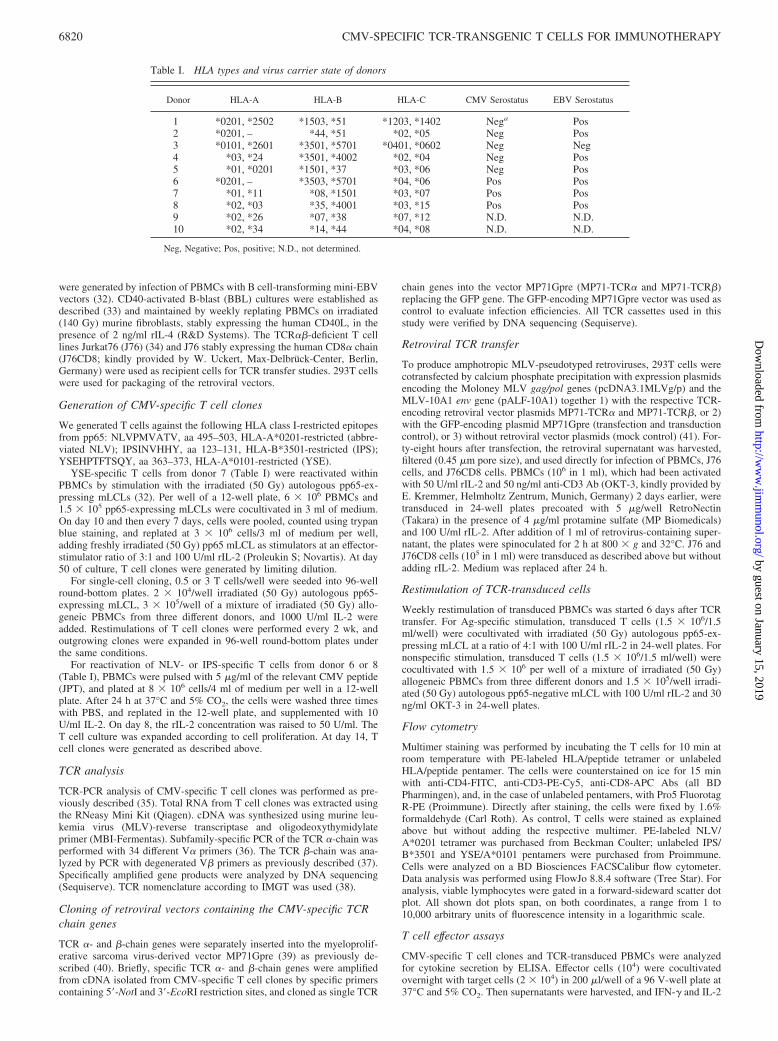
We generated CMV-specific CD8� T cell clones against theepitopes YSE (HLA-A*0101), NLV (HLA-A*0201), and IPS(HLA-B*3501) derived from pp65. CMV-specific T cells wereenriched by Ag-specific stimulation of primary T cells from threedifferent CMV-seropositive donors. NLV- and IPS-specific T cellswere obtained by stimulating PBMCs with the epitope peptide, asshown for NLV-specific T cells in Fig. 1A. YSE-specific T cellswere enriched by repeated stimulation with the autologous pp65-expressing mLCL (Fig. 1B). Subsequently, T cell clones were gen-erated by limiting dilution.
To test whether the T cell clones recognize pp65 Ag when in-tracellularly expressed, processed and presented, we analyzed theirreactivity to pp65-expressing mLCLs, control (pp65-negative)mLCLs, and CD40-activated BBLs from various donors. For char-acterization of their TCR, we selected T cell clones which stronglyand exclusively recognized autologous or HLA-matched pp65-ex-pressing mLCLs, as shown for NLV- and IPS-specific clones (Fig.1, D and E). All T cell clones with this reaction pattern showeda clear staining with the corresponding HLA/peptide multimer(Fig. 1C).
CMV-specific TCRs of these T cell clones were characterizedby TCR subfamily-specific PCR (Table II). The NLV-specific Tcell clones derived from donor 6 used different TRAV and TRBVgenes. The TCR NLV2 from T cell clones JG-9 and JG-24 is apublic TCR and was also found in NLV-specific T cell clones ofanother donor not included in this study. In addition, other groupsdescribed NLV-specific T cells from various donors which usedthe same TCR �- or �-chain or closely related variants (25, 26, 30,31). In contrast, the TCRs NLV3, YSE, and IPS have not beendescribed thus far. We found the CDR3� region of the NLV3-TCRto be extremely short; this TCR might recognize Ag by adoptingan unusual structure, meriting further investigation.
Retroviral transfer of CMV-specific TCRs into TCR��-deficientT cell lines
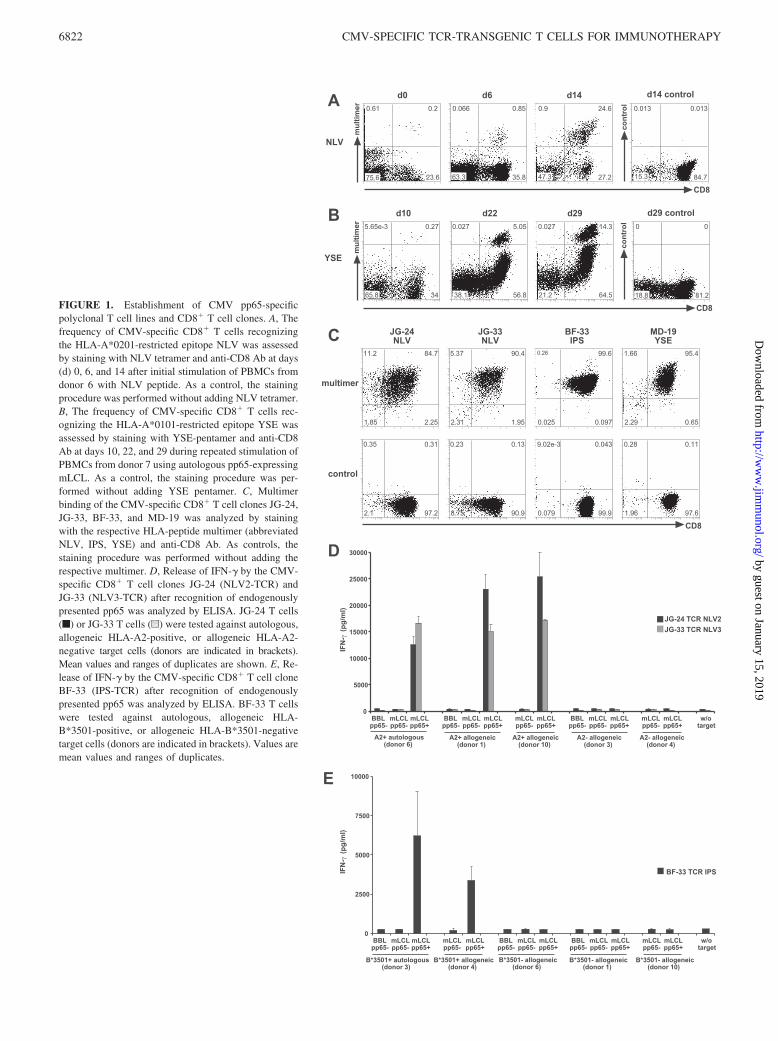
The TCR �- and �-chain genes derived from these four CMV-specific T cell clones were individually inserted into the retroviralvector plasmid pMP71Gpre, and infectious retroviral supernatantswere produced. To investigate transgenic TCR expression in theabsence of an endogenous TCR, we transferred the CMV-specificTCRs to TCR��-deficient J76 cells and to J76CD8 cells stablyexpressing the human CD8� chain (Fig. 2A). All TCRs were ex-pressed on J76CD8 cells and were strongly stained with the re-spective HLA/peptide multimer. Staining levels on J76 cells with-out CD8� were reduced, but three of the four TCRs could bedetected on these cells, except for the IPS-TCR. We conclude thatthe TCRs NLV2, NLV3, and YSE did not strictly require CD8 tobind their target HLA/peptide complex.
Transfer of CMV-specific TCRs into primary T cells ofCMV-seronegative donors
To confer CMV-specific reactivity on primary T cells from CMV-negative donors, we performed retroviral transfer of our CMV-specific TCRs. Four days after transfer, all TCRs were expressedon PBMCs and could be stained with peptide/HLA multimers,whereas mock-transduced cells were multimer negative (Fig. 2, Band C). Multimer-staining levels of the individual TCRs were
comparable between different donors with different HLA back-grounds (Table I), but the levels varied among the four differentTCRs (Fig. 2C). Proportions of multimer-positive cells were sim-ilar within different T cell subsets (CD3�, CD8�, CD4�) exceptfor the IPS-TCR. This TCR was not detectable on CD4�CD8� Tcells, suggesting CD8 dependency, which is consistent with ourprevious observation that this TCR could not be detected on J76cells (Fig. 2A). The general transduction efficiency, as determinedwith GFP-expressing retrovirus, was comparable for all donors(45–60%). To check whether the TCR-transgenic T cells had ac-quired reactivity against pp65-expressing cells already at this earlystage, we cocultivated the transduced PBMCs with autologous,allogeneic HLA-matched and HLA-mismatched target cells,namely pp65-expressing and pp65-negative mLCLs and BBLs, atday 6 after TCR transfer, and determined the amount of releasedIFN-� and IL-2. NLV2-TCR-transduced as well as NLV3-TCR-transduced PBMCs specifically produced considerable amounts ofIFN-� (Fig. 2D) and some IL-2 (Fig. 2E) after coculture withHLA-A2-positive pp65-expressing mLCLs. Cocultivation withBBLs (pp65 and EBV negative) did not induce IFN-�, but lowamounts of IFN-� were released when the TCR-transduced cellswere tested against pp65-negative mLCLs, indicating the presenceof some EBV-specific memory T cells, which was expected be-cause the donor (donor 1) was EBV positive (Table I).
These results show that CMV-specific T cells recognizing en-dogenously processed Ag can be rapidly produced by TCR transferin a simple 8-day procedure from PBMCs from seronegativedonors.
Simultaneous expansion and enrichment of TCR-transgenic Tcells by Ag-specific stimulation
For successful T cell therapy, TCR-transgenic T cells must be ableto recognize the endogenously processed Ag and proliferate in anAg-specific manner in vivo to achieve sufficient function andmaintenance after therapeutic T cell transfer. Therefore, we inves-tigated whether CMV-TCR-transduced T cells could be expandedand enriched by Ag-specific stimulation. As Ag-specific stimula-tors, we used the autologous pp65-expressing mLCL, which pre-sents endogenously processed pp65 epitopes on class I and II HLAmolecules (22, 32). For comparison, we performed nonspecificstimulation with anti-CD3 Ab, a mix of allogeneic PBMCs derivedfrom three unrelated donors, and the autologous mLCL withoutpp65 expression. The stimulation protocol is schematically shownin Fig. 3A.
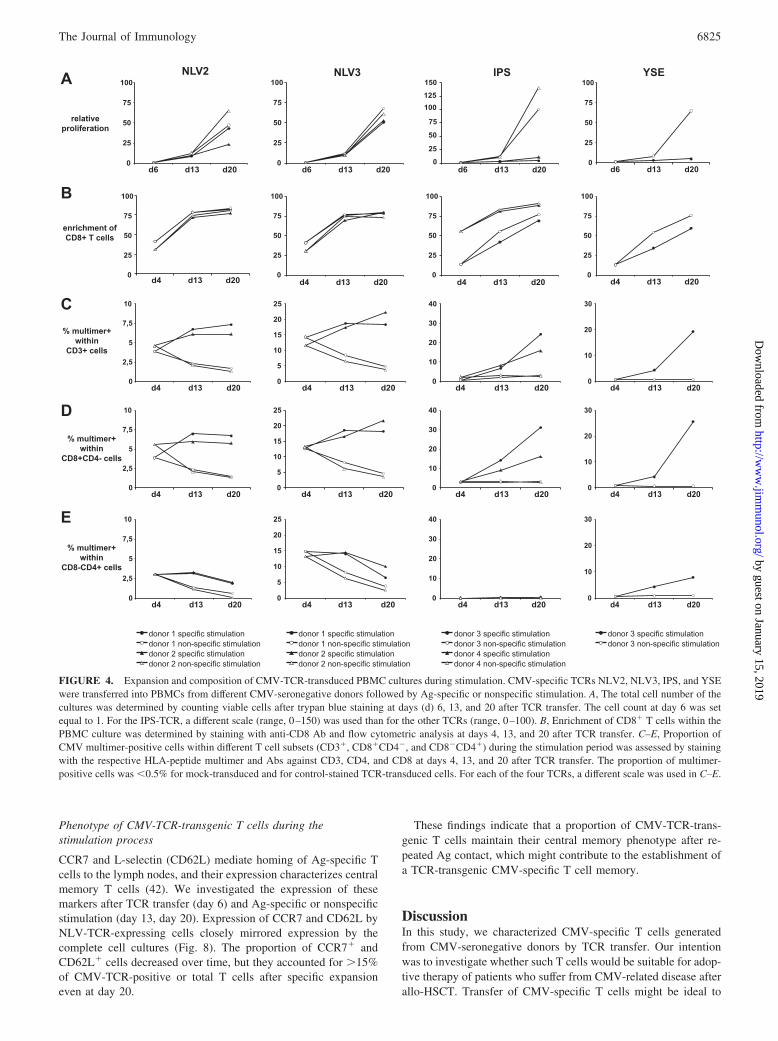
For all four CMV-TCRs, CMV-TCR-transgenic CD3� T cellswere specifically enriched by stimulation with the endogenouslyprocessed CMV Ag pp65 (Fig. 4C). The same was true if onlyCMV-TCR-transgenic CD8� T cells were evaluated (Fig. 4D).Examples of multimer stainings are shown in Fig. 3B. As before,for a given TCR proportions of TCR-transgenic T cells were sim-ilar among different PBMC donors (Fig. 4, C and D). Ag-specificstimulation led to an expansion of each TCR-transduced T cellculture in terms of absolute cell numbers (Fig. 4A). For the twoNLV-TCRs, specific and nonspecific stimulation were comparablein terms of total cell expansion (Fig. 4A). For the IPS- and YSE-TCRs, nonspecific stimulation led to the strongest overall cell ex-pansion (Fig. 4A), but only specific stimulation raised the propor-tion of TCR-transgenic T cells (Fig. 4, C and D). Both stimulationprotocols preferentially expanded CD8� T cells (Fig. 4B).
From these data, we determined the increase in absolute num-bers of TCR-transgenic CD8� T cells (expansion) as well as the
6821The Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
γγ
FIGURE 1. Establishment of CMV pp65-specificpolyclonal T cell lines and CD8� T cell clones. A, Thefrequency of CMV-specific CD8� T cells recognizingthe HLA-A*0201-restricted epitope NLV was assessedby staining with NLV tetramer and anti-CD8 Ab at days(d) 0, 6, and 14 after initial stimulation of PBMCs fromdonor 6 with NLV peptide. As a control, the stainingprocedure was performed without adding NLV tetramer.B, The frequency of CMV-specific CD8� T cells rec-ognizing the HLA-A*0101-restricted epitope YSE wasassessed by staining with YSE-pentamer and anti-CD8Ab at days 10, 22, and 29 during repeated stimulation ofPBMCs from donor 7 using autologous pp65-expressingmLCL. As a control, the staining procedure was per-formed without adding YSE pentamer. C, Multimerbinding of the CMV-specific CD8� T cell clones JG-24,JG-33, BF-33, and MD-19 was analyzed by stainingwith the respective HLA-peptide multimer (abbreviatedNLV, IPS, YSE) and anti-CD8 Ab. As controls, thestaining procedure was performed without adding therespective multimer. D, Release of IFN-� by the CMV-specific CD8� T cell clones JG-24 (NLV2-TCR) andJG-33 (NLV3-TCR) after recognition of endogenouslypresented pp65 was analyzed by ELISA. JG-24 T cells(f) or JG-33 T cells (u) were tested against autologous,allogeneic HLA-A2-positive, or allogeneic HLA-A2-negative target cells (donors are indicated in brackets).Mean values and ranges of duplicates are shown. E, Re-lease of IFN-� by the CMV-specific CD8� T cell cloneBF-33 (IPS-TCR) after recognition of endogenouslypresented pp65 was analyzed by ELISA. BF-33 T cellswere tested against autologous, allogeneic HLA-B*3501-positive, or allogeneic HLA-B*3501-negativetarget cells (donors are indicated in brackets). Values aremean values and ranges of duplicates.
6822 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
increase in the proportion of TCR-transgenic CD8� T cells (en-richment), after both Ag-specific and nonspecific stimulation (Ta-ble III). For each TCR, both stimulation conditions led to an im-pressive expansion of CD8� T cells expressing the transgenic
TCR (36- to 672-fold; Table III). With the exception of the IPS-TCR-transgenic T cells, Ag-specific stimulation resulted in a su-perior expansion compared with nonspecific stimulation. For allfour TCRs, enrichment of TCR-transgenic CD8� T cells was
FIGURE 2. Transfer of four different CMV-specific TCRs into T cell lines and PBMCs. A, The CMV pp65-specific TCRs NLV2, NLV3, IPS, and YSEwere transferred into J76 cells (top) and J76CD8 cells (bottom). Expression of the introduced TCR was analyzed by staining with the respective HLA/peptide-multimer (thick line) at day 4 after transfer. As control, mock-transduced cells were analyzed by staining with the same multimer (thin line).Numbers indicate the percentage of multimer-positive cells. B, PBMCs from CMV-seronegative donors were transduced with the CMV pp65-specific TCRsNLV2 (donor 1), NLV3 (donor 1), IPS (donor 3), and YSE (donor 3; bottom), or mock-transduced (top). Expression of the introduced TCR was analyzedby staining with the respective HLA-peptide multimer at day 4 after transfer. C, The TCRs NLV2, NLV3, IPS, and YSE were transferred into PBMCs fromdifferent CMV-seronegative donors, and the frequency of multimer-positive cells within different T cell subsets, CD3� cells (black bars), CD8�CD4� cells(dark gray bars), and CD8�CD4� cells (light gray bars), was determined by costaining with the respective HLA/peptide-multimer and Abs against CD3,CD4, and CD8 at day 4 after TCR transfer. D and E, CMV-specific reactivity of TCR-transgenic T cells against cells endogenously presenting the targetAg pp65 was analyzed by IFN-� ELISA (D) and IL-2 ELISA (E) at day 6 after TCR transfer. NLV2 (black bars), NLV3 (dark gray bars), or mock-transduced (light gray bars) PBMCs from donor 1 were tested against autologous, allogeneic HLA-A2-positive, or allogeneic HLA-A2-negative target cells(donors are indicated in brackets). Mean values and ranges of duplicates are shown. The percentage of multimer-positive cells at day 4 after transfer isindicated.
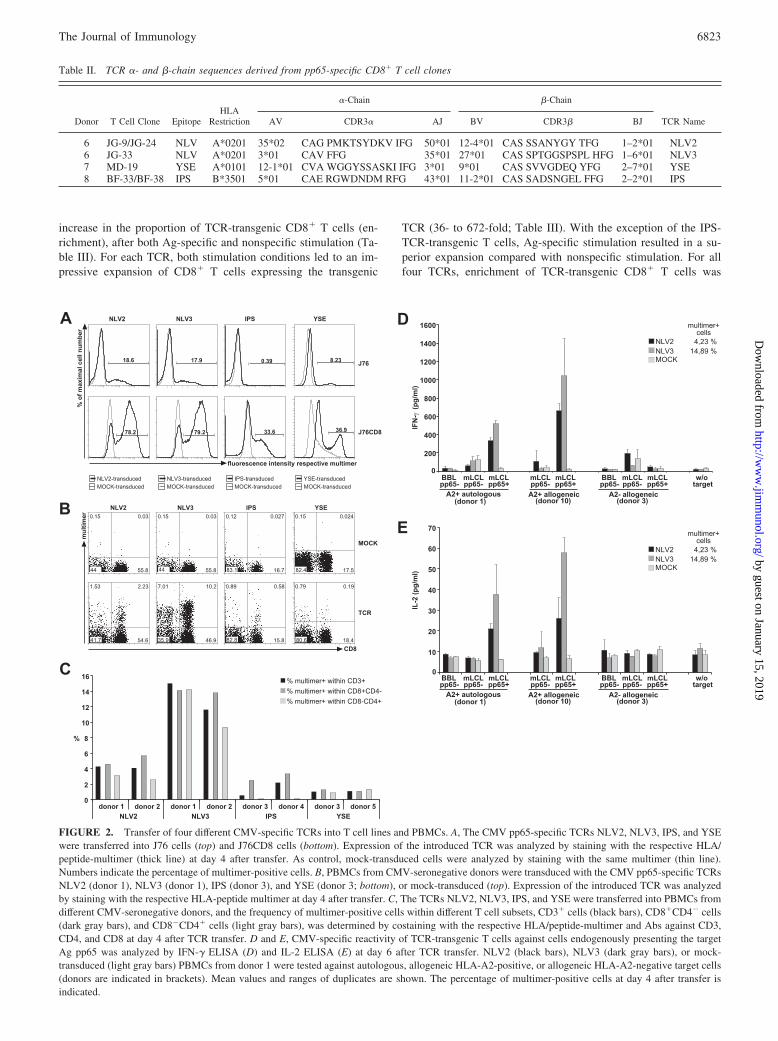
Table II. TCR �- and �-chain sequences derived from pp65-specific CD8� T cell clones
Donor T Cell Clone EpitopeHLA
Restriction
�-Chain �-Chain
TCR NameAV CDR3� AJ BV CDR3� BJ
6 JG-9/JG-24 NLV A*0201 35*02 CAG PMKTSYDKV IFG 50*01 12-4*01 CAS SSANYGY TFG 1–2*01 NLV26 JG-33 NLV A*0201 3*01 CAV FFG 35*01 27*01 CAS SPTGGSPSPL HFG 1–6*01 NLV37 MD-19 YSE A*0101 12-1*01 CVA WGGYSSASKI IFG 3*01 9*01 CAS SVVGDEQ YFG 2–7*01 YSE8 BF-33/BF-38 IPS B*3501 5*01 CAE RGWDNDM RFG 43*01 11-2*01 CAS SADSNGEL FFG 2–2*01 IPS
6823The Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
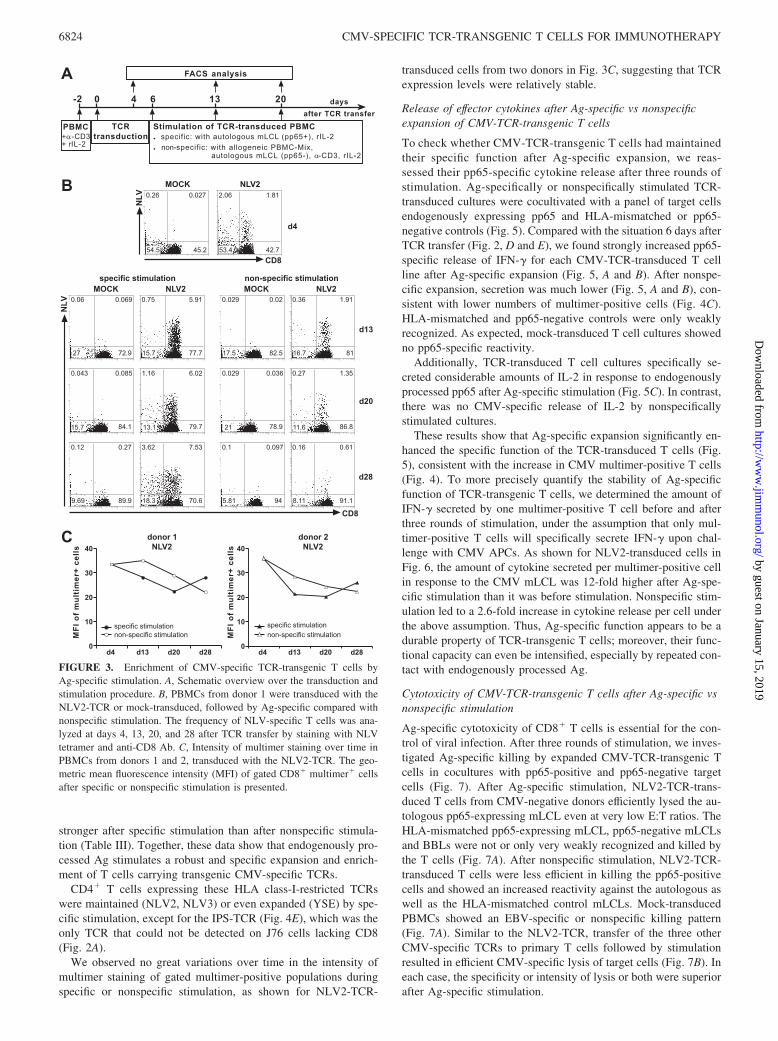
stronger after specific stimulation than after nonspecific stimula-tion (Table III). Together, these data show that endogenously pro-cessed Ag stimulates a robust and specific expansion and enrich-ment of T cells carrying transgenic CMV-specific TCRs.
CD4� T cells expressing these HLA class-I-restricted TCRswere maintained (NLV2, NLV3) or even expanded (YSE) by spe-cific stimulation, except for the IPS-TCR (Fig. 4E), which was theonly TCR that could not be detected on J76 cells lacking CD8(Fig. 2A).
We observed no great variations over time in the intensity ofmultimer staining of gated multimer-positive populations duringspecific or nonspecific stimulation, as shown for NLV2-TCR-
transduced cells from two donors in Fig. 3C, suggesting that TCRexpression levels were relatively stable.
Release of effector cytokines after Ag-specific vs nonspecificexpansion of CMV-TCR-transgenic T cells
To check whether CMV-TCR-transgenic T cells had maintainedtheir specific function after Ag-specific expansion, we reas-sessed their pp65-specific cytokine release after three rounds ofstimulation. Ag-specifically or nonspecifically stimulated TCR-transduced cultures were cocultivated with a panel of target cellsendogenously expressing pp65 and HLA-mismatched or pp65-negative controls (Fig. 5). Compared with the situation 6 days afterTCR transfer (Fig. 2, D and E), we found strongly increased pp65-specific release of IFN-� for each CMV-TCR-transduced T cellline after Ag-specific expansion (Fig. 5, A and B). After nonspe-cific expansion, secretion was much lower (Fig. 5, A and B), con-sistent with lower numbers of multimer-positive cells (Fig. 4C).HLA-mismatched and pp65-negative controls were only weaklyrecognized. As expected, mock-transduced T cell cultures showedno pp65-specific reactivity.
Additionally, TCR-transduced T cell cultures specifically se-creted considerable amounts of IL-2 in response to endogenouslyprocessed pp65 after Ag-specific stimulation (Fig. 5C). In contrast,there was no CMV-specific release of IL-2 by nonspecificallystimulated cultures.
These results show that Ag-specific expansion significantly en-hanced the specific function of the TCR-transduced T cells (Fig.5), consistent with the increase in CMV multimer-positive T cells(Fig. 4). To more precisely quantify the stability of Ag-specificfunction of TCR-transgenic T cells, we determined the amount ofIFN-� secreted by one multimer-positive T cell before and afterthree rounds of stimulation, under the assumption that only mul-timer-positive T cells will specifically secrete IFN-� upon chal-lenge with CMV APCs. As shown for NLV2-transduced cells inFig. 6, the amount of cytokine secreted per multimer-positive cellin response to the CMV mLCL was 12-fold higher after Ag-spe-cific stimulation than it was before stimulation. Nonspecific stim-ulation led to a 2.6-fold increase in cytokine release per cell underthe above assumption. Thus, Ag-specific function appears to be adurable property of TCR-transgenic T cells; moreover, their func-tional capacity can even be intensified, especially by repeated con-tact with endogenously processed Ag.
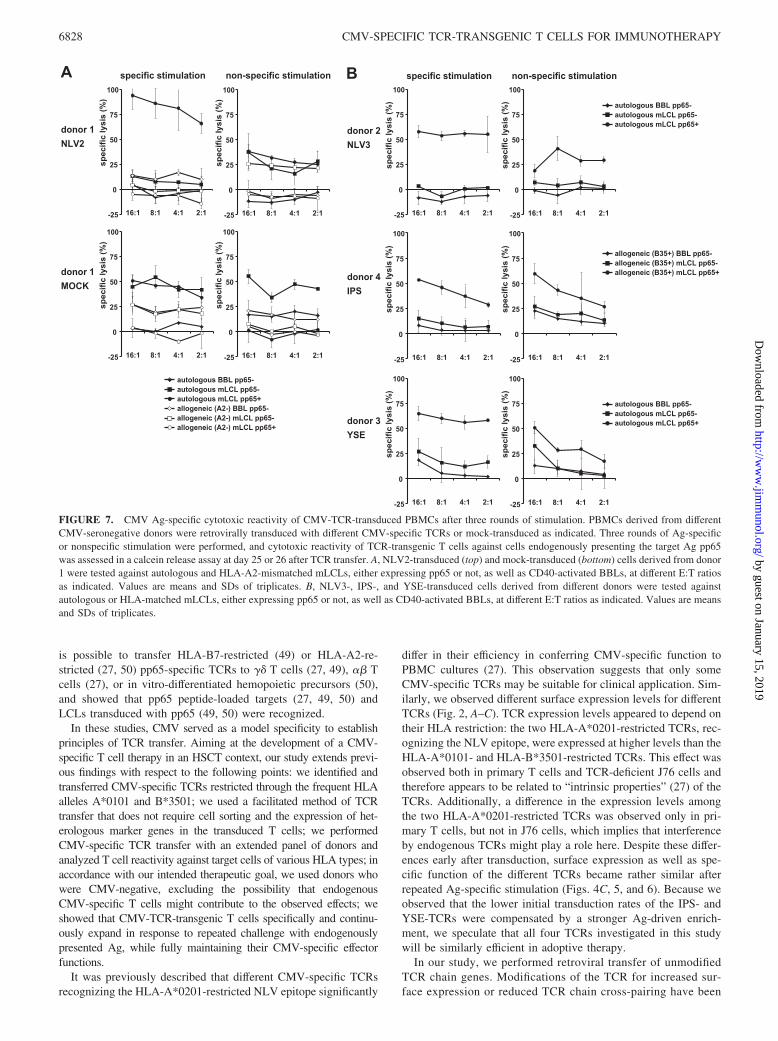
Cytotoxicity of CMV-TCR-transgenic T cells after Ag-specific vsnonspecific stimulation
Ag-specific cytotoxicity of CD8� T cells is essential for the con-trol of viral infection. After three rounds of stimulation, we inves-tigated Ag-specific killing by expanded CMV-TCR-transgenic Tcells in cocultures with pp65-positive and pp65-negative targetcells (Fig. 7). After Ag-specific stimulation, NLV2-TCR-trans-duced T cells from CMV-negative donors efficiently lysed the au-tologous pp65-expressing mLCL even at very low E:T ratios. TheHLA-mismatched pp65-expressing mLCL, pp65-negative mLCLsand BBLs were not or only very weakly recognized and killed bythe T cells (Fig. 7A). After nonspecific stimulation, NLV2-TCR-transduced T cells were less efficient in killing the pp65-positivecells and showed an increased reactivity against the autologous aswell as the HLA-mismatched control mLCLs. Mock-transducedPBMCs showed an EBV-specific or nonspecific killing pattern(Fig. 7A). Similar to the NLV2-TCR, transfer of the three otherCMV-specific TCRs to primary T cells followed by stimulationresulted in efficient CMV-specific lysis of target cells (Fig. 7B). Ineach case, the specificity or intensity of lysis or both were superiorafter Ag-specific stimulation.
FIGURE 3. Enrichment of CMV-specific TCR-transgenic T cells byAg-specific stimulation. A, Schematic overview over the transduction andstimulation procedure. B, PBMCs from donor 1 were transduced with theNLV2-TCR or mock-transduced, followed by Ag-specific compared withnonspecific stimulation. The frequency of NLV-specific T cells was ana-lyzed at days 4, 13, 20, and 28 after TCR transfer by staining with NLVtetramer and anti-CD8 Ab. C, Intensity of multimer staining over time inPBMCs from donors 1 and 2, transduced with the NLV2-TCR. The geo-metric mean fluorescence intensity (MFI) of gated CD8� multimer� cellsafter specific or nonspecific stimulation is presented.
6824 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
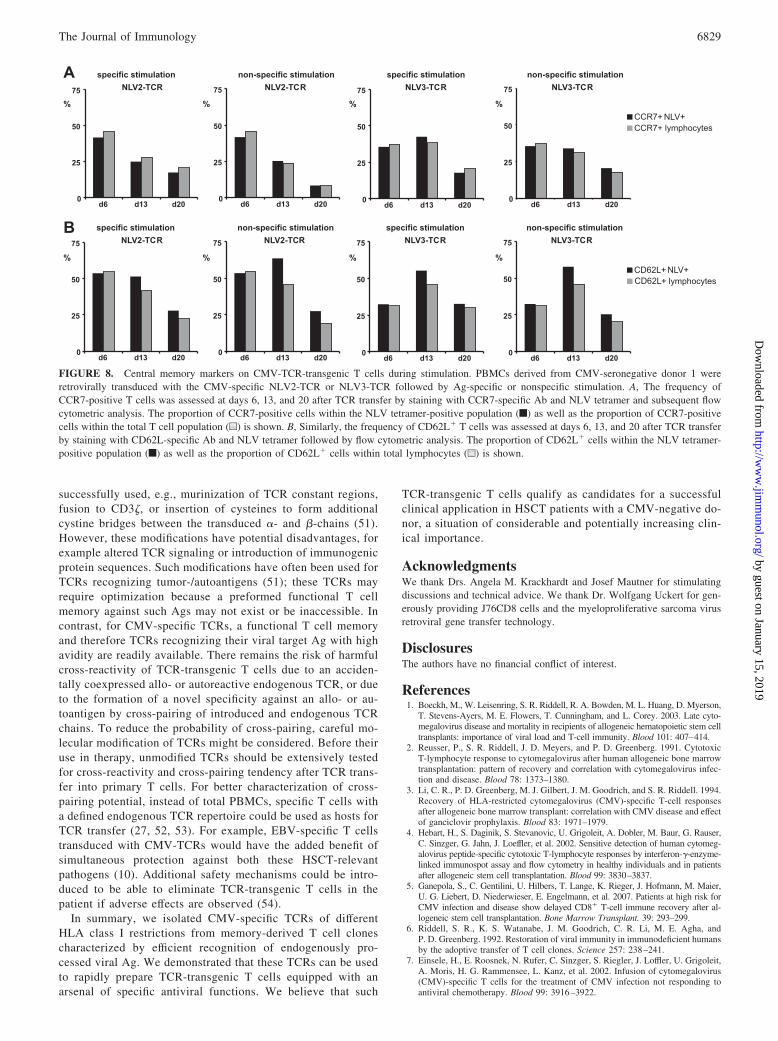
Phenotype of CMV-TCR-transgenic T cells during thestimulation process
CCR7 and L-selectin (CD62L) mediate homing of Ag-specific Tcells to the lymph nodes, and their expression characterizes centralmemory T cells (42). We investigated the expression of thesemarkers after TCR transfer (day 6) and Ag-specific or nonspecificstimulation (day 13, day 20). Expression of CCR7 and CD62L byNLV-TCR-expressing cells closely mirrored expression by thecomplete cell cultures (Fig. 8). The proportion of CCR7� andCD62L� cells decreased over time, but they accounted for �15%of CMV-TCR-positive or total T cells after specific expansioneven at day 20.
These findings indicate that a proportion of CMV-TCR-trans-genic T cells maintain their central memory phenotype after re-peated Ag contact, which might contribute to the establishment ofa TCR-transgenic CMV-specific T cell memory.
DiscussionIn this study, we characterized CMV-specific T cells generatedfrom CMV-seronegative donors by TCR transfer. Our intentionwas to investigate whether such T cells would be suitable for adop-tive therapy of patients who suffer from CMV-related disease afterallo-HSCT. Transfer of CMV-specific T cells might be ideal to
FIGURE 4. Expansion and composition of CMV-TCR-transduced PBMC cultures during stimulation. CMV-specific TCRs NLV2, NLV3, IPS, and YSEwere transferred into PBMCs from different CMV-seronegative donors followed by Ag-specific or nonspecific stimulation. A, The total cell number of thecultures was determined by counting viable cells after trypan blue staining at days (d) 6, 13, and 20 after TCR transfer. The cell count at day 6 was setequal to 1. For the IPS-TCR, a different scale (range, 0–150) was used than for the other TCRs (range, 0–100). B, Enrichment of CD8� T cells within thePBMC culture was determined by staining with anti-CD8 Ab and flow cytometric analysis at days 4, 13, and 20 after TCR transfer. C–E, Proportion ofCMV multimer-positive cells within different T cell subsets (CD3�, CD8�CD4�, and CD8�CD4�) during the stimulation period was assessed by stainingwith the respective HLA-peptide multimer and Abs against CD3, CD4, and CD8 at days 4, 13, and 20 after TCR transfer. The proportion of multimer-positive cells was �0.5% for mock-transduced and for control-stained TCR-transduced cells. For each of the four TCRs, a different scale was used in C–E.
6825The Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

prevent and cure CMV disease, but for the combination CMV-negative donor/CMV-positive recipient (D�/R�) such cells areusually not available. However, for these patients, the risk of CMVdisease is highest. D�/R� is usually the most frequent situation inallo-HSCT (5), but with decreasing CMV prevalence in the pop-ulation, D�/R� HSCTs will occur with increasing frequency.
The hypothesis underlying this study was that suitable CMV-specific TCRs are readily available within the memory T cell rep-ertoire of CMV-seropositive donors. It can be expected that theTCR repertoire of CMV-specific memory T cells is shaped, at leastin part, by the necessity to control CMV infection and to protectfrom disease. To select appropriate candidate T cell clones to
FIGURE 5. CMV Ag-specific release of effector cytokines by CMV-TCR-transduced PBMCs after three rounds of stimulation. PBMCs derived fromCMV-seronegative donors (A, donor 2; B and C, donor 3) were retrovirally transduced with different CMV-specific TCRs, or mock-transduced as indicated.Three rounds of Ag-specific (f) compared with nonspecific (u) stimulation were performed, and CMV-specific reactivity of TCR-transgenic T cells againstcells endogenously presenting the target Ag pp65 was analyzed by IFN-� ELISA (A and B) and IL-2 ELISA (C) at day 25 after TCR transfer. As targetcells, autologous and allogeneic (matched or mismatched to the HLA restriction of the transgenic TCRs) pp65-expressing mLCLs, control mLCL, orCD40-activated BBLs were used. w/o, Without. Mean values and range of duplicates are shown.
Table III. Total expansion and enrichment of CD8� T cells expressing transgenic TCRs by Ag-specific or nonspecific stimulation
CD8�Multimer� Cells
NLV2 NLV3 IPSYSE
Donor 3Donor 1 Donor 2 Donor 1 Donor 2 Donor 3 Donor 4
Expansiona
Ag-specific stimulation 148 60 141 224 228 88 672Nonspecific stimulation 36 40 47 40 640 183 301
Enrichmentb
Ag-specific stimulation 1.74 1.02 1.44 1.64 11.15 4.78 31.6Nonspecific stimulation 0.37 0.23 0.23 0.27 1.13 0.79 0.79
a Expansion of CMV-TCR-transgenic CD8� T cells was calculated by dividing the absolute number of multimer�CD8� T cells on day 20 by the absolute number ofmultimer�CD8� T cells on day 4 (see also Fig. 4). Ag-specific or nonspecific stimulation was performed on days 6 and 13 (see Fig. 3A).
b Enrichment of CMV-TCR-transgenic CD8� T cells was calculated by dividing the proportions of multimer�CD8� T cells within total cells on day 20 by the proportionsof multimer�CD8� T cells within total cells on day 4 (see Fig. 4D).
6826 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

derive TCRs for transfer, we used a screening procedure that se-lected T cells specifically reacting to endogenously processedCMV Ag. Furthermore, we assumed that CMV Ag-specific T cellscan be engineered by CMV-TCR transfer independent of the HLArestriction of the TCRs, although up to now mainly transfer ofHLA-A*0201-restricted tumor-specific TCRs has been studied(17). To verify these assumptions, we isolated, characterized, andcloned several CMV-specific TCRs that recognize differentepitopes from pp65 restricted through various HLA allotypes. Oneof these TCRs, NLV2, was present in both HLA-A*0201-positivedonors investigated, confirming the earlier description of this TCRor its �-chain as public (25, 26, 30, 31). A second TCR, NLV3,recognizing the same epitope, has not been described previouslyand was identified in only one of the donors. Because this TCR hasan unusually short CDR3� region (Table II), it might be interestingto investigate the structure of the corresponding TCR-peptide-MHC complex. Additionally, we characterized two previously un-described TCRs with different HLA restrictions, one recognizingthe HLA-A*0101-restricted epitope YSE and one specific for theHLA-B*3501-restricted epitope IPS. We found that retroviraltransfer of all these CMV-specific TCRs resulted in TCR-trans-genic T cells which exhibited a wide spectrum of desirable func-tions in response to endogenously processed CMV Ag. By stim-ulation with endogenously processed Ag, TCR-transgenic T cellscould be specifically expanded and enriched in relative and abso-lute terms (Fig. 4 and Table III), indicating a strong proliferativepotential of these cells after Ag contact. This feature will be im-portant for controlling CMV infection or reactivation in vivo. Theincrease in CMV-TCR-expressing T cells was accompanied by anincrease in Ag-specific effector functions (cytotoxicity and cyto-kine secretion) of similar magnitude (Figs. 5 and 7), showing thatthe functionality of these TCR-transgenic T cells is fully main-
tained after Ag contact and Ag-specific proliferation in vitro. Inline with these observations, staining intensity of HLA-peptidemultimer-positive populations was stable (Fig. 3C), and the func-tional capacity of multimer-positive populations even appeared toincrease over time (Fig. 6). Furthermore, no stimulation of TCR-transduced T cells, cell sorting or depletion was required to obtainT cells displaying Ag-specific function at very convincing levels asearly as 6 days after TCR transfer (Fig. 2).
These results prompt us to suggest that a simple protocol con-sisting of TCR transfer to PBMCs, without further expansion orselection steps, will produce CMV-TCR-transgenic T cell popula-tions suitable for immediate use in T cell transfer therapy. Careshould be taken to keep the total number of allogeneic T cellstransferred to a patient low enough to minimize the probability ofgraft-vs-host disease, for example, �106 cells/kg for HLA-matched donor-recipient pairs (43), because alloreactive or origi-nally tolerant self-reactive T cells might be present in the TCR-transduced T cell preparation. Such potentially harmful T cellsmight be activated by the anti-CD3 treatment preceding transduc-tion, or by means of a transgenic CMV-specific TCR coexpressedon the same cell and recognizing its target Ag in the patient. How-ever, as long as T cell therapy is performed early enough in apre-emptive situation, low numbers of CMV Ag-specific T cellsmay be sufficient to avert disease (9). We consider it likely thatAg-driven T cell expansion in the CMV-infected HSCT patient,possibly favored by lymphopenia (1), will be at least as effective inproducing sufficient numbers of specific effector T cells than anyfurther expansion in vitro. The considerable proportions of CMV-TCR-transgenic T cells positive for the central memory markersCCR7 and CD62L (Fig. 8), which were largely retained after Agcontact in vitro, further suggest that TCR-transgenic cells will beable to form an effective CMV-specific memory in vivo.
Different strategies to obtain CMV-specific T cells for adoptivetherapy after D�/R� HSCT might be taken into consideration.CMV-specific T cells could be obtained directly from a third un-related CMV-seropositive donor. In this context, third-party-de-rived EBV-specific T cells had very good therapeutic effects insolid organ transplant recipients (44). However, allogeneic T cellsmight be rapidly rejected, hampering long-term antiviral protec-tion. Therefore, T cells derived from the HSCT donor are to bepreferred for adoptive therapy.
Another alternative to CMV-TCR transfer would be in vitropriming and expansion of rare naive CMV-specific T cells fromthe donor. Remarkably, with a small proportion of CMV-seroneg-ative donors it has been possible to generate CMV-specific T cellsby in vitro priming (45–47). However, we expect it to be verydifficult to translate these observations into feasible clinical pro-cedures, because the precursor frequency of naive CMV-specificCD8� T cells in CMV-seronegative individuals, estimated fromgeneral considerations on TCR diversity (48), is unlikely to beabove 1 in 106 naive T cells. Therefore, transfer of CMV-specificTCRs to primary T cells will be the easiest and most efficientmethod for adoptive therapy of CMV-related complications afterHSCT with CMV-seronegative donors.
By using CMV-specific TCRs of various HLA restrictions, popu-lation coverage can be optimized. With the selected HLA-A*0101-,HLA-A*0201-, and HLA-B*3501-restricted CMV-specific TCRs, atleast 70% of Europeans can be covered, HLA-A*0201 being the mostfrequent and HLA-A*0101 the second most frequent HLA allo-type. Obviously, further extension of this TCR repertoire to coveradditional HLA allotypes is highly desirable.
CMV-specific TCR genes have previously been cloned andtransferred to T cells (27, 49, 50). These studies established that it
FIGURE 6. CMV Ag-specific cytokine release per multimer-positivecell before and after stimulation of CMV-TCR-transduced PBMC. PBMCsfrom CMV-seronegative donor 1 were transduced with the NLV2-TCR,and IFN-� secretion in reaction to target cells was analyzed before (day 6)and after (day 25) three rounds of Ag-specific (left) or nonspecific (right)stimulation. Target cells included autologous and HLA-mismatched pp65-expressing mLCLs and control mLCLs. Cytokine release per multimer-positive cell was calculated by dividing the total amount of cytokine de-tected in the reaction supernatant by the number of total T cells per reactionmultiplied by the proportion of multimer-positive cells assessed at the near-est available time point: 4.23% on day 4 (before stimulation); 11.2% onday 28 (after specific stimulation); 0.79% on day 28 (after nonspecificstimulation).
6827The Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
is possible to transfer HLA-B7-restricted (49) or HLA-A2-re-stricted (27, 50) pp65-specific TCRs to �� T cells (27, 49), �� Tcells (27), or in vitro-differentiated hemopoietic precursors (50),and showed that pp65 peptide-loaded targets (27, 49, 50) andLCLs transduced with pp65 (49, 50) were recognized.
In these studies, CMV served as a model specificity to establishprinciples of TCR transfer. Aiming at the development of a CMV-specific T cell therapy in an HSCT context, our study extends previ-ous findings with respect to the following points: we identified andtransferred CMV-specific TCRs restricted through the frequent HLAalleles A*0101 and B*3501; we used a facilitated method of TCRtransfer that does not require cell sorting and the expression of het-erologous marker genes in the transduced T cells; we performedCMV-specific TCR transfer with an extended panel of donors andanalyzed T cell reactivity against target cells of various HLA types; inaccordance with our intended therapeutic goal, we used donors whowere CMV-negative, excluding the possibility that endogenousCMV-specific T cells might contribute to the observed effects; weshowed that CMV-TCR-transgenic T cells specifically and continu-ously expand in response to repeated challenge with endogenouslypresented Ag, while fully maintaining their CMV-specific effectorfunctions.
It was previously described that different CMV-specific TCRsrecognizing the HLA-A*0201-restricted NLV epitope significantly
differ in their efficiency in conferring CMV-specific function toPBMC cultures (27). This observation suggests that only someCMV-specific TCRs may be suitable for clinical application. Sim-ilarly, we observed different surface expression levels for differentTCRs (Fig. 2, A–C). TCR expression levels appeared to depend ontheir HLA restriction: the two HLA-A*0201-restricted TCRs, rec-ognizing the NLV epitope, were expressed at higher levels than theHLA-A*0101- and HLA-B*3501-restricted TCRs. This effect wasobserved both in primary T cells and TCR-deficient J76 cells andtherefore appears to be related to “intrinsic properties” (27) of theTCRs. Additionally, a difference in the expression levels amongthe two HLA-A*0201-restricted TCRs was observed only in pri-mary T cells, but not in J76 cells, which implies that interferenceby endogenous TCRs might play a role here. Despite these differ-ences early after transduction, surface expression as well as spe-cific function of the different TCRs became rather similar afterrepeated Ag-specific stimulation (Figs. 4C, 5, and 6). Because weobserved that the lower initial transduction rates of the IPS- andYSE-TCRs were compensated by a stronger Ag-driven enrich-ment, we speculate that all four TCRs investigated in this studywill be similarly efficient in adoptive therapy.
In our study, we performed retroviral transfer of unmodifiedTCR chain genes. Modifications of the TCR for increased sur-face expression or reduced TCR chain cross-pairing have been
FIGURE 7. CMV Ag-specific cytotoxic reactivity of CMV-TCR-transduced PBMCs after three rounds of stimulation. PBMCs derived from differentCMV-seronegative donors were retrovirally transduced with different CMV-specific TCRs or mock-transduced as indicated. Three rounds of Ag-specificor nonspecific stimulation were performed, and cytotoxic reactivity of TCR-transgenic T cells against cells endogenously presenting the target Ag pp65was assessed in a calcein release assay at day 25 or 26 after TCR transfer. A, NLV2-transduced (top) and mock-transduced (bottom) cells derived from donor1 were tested against autologous and HLA-A2-mismatched mLCLs, either expressing pp65 or not, as well as CD40-activated BBLs, at different E:T ratiosas indicated. Values are means and SDs of triplicates. B, NLV3-, IPS-, and YSE-transduced cells derived from different donors were tested againstautologous or HLA-matched mLCLs, either expressing pp65 or not, as well as CD40-activated BBLs, at different E:T ratios as indicated. Values are meansand SDs of triplicates.
6828 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
successfully used, e.g., murinization of TCR constant regions,fusion to CD3�, or insertion of cysteines to form additionalcystine bridges between the transduced �- and �-chains (51).However, these modifications have potential disadvantages, forexample altered TCR signaling or introduction of immunogenicprotein sequences. Such modifications have often been used forTCRs recognizing tumor-/autoantigens (51); these TCRs mayrequire optimization because a preformed functional T cellmemory against such Ags may not exist or be inaccessible. Incontrast, for CMV-specific TCRs, a functional T cell memoryand therefore TCRs recognizing their viral target Ag with highavidity are readily available. There remains the risk of harmfulcross-reactivity of TCR-transgenic T cells due to an acciden-tally coexpressed allo- or autoreactive endogenous TCR, or dueto the formation of a novel specificity against an allo- or au-toantigen by cross-pairing of introduced and endogenous TCRchains. To reduce the probability of cross-pairing, careful mo-lecular modification of TCRs might be considered. Before theiruse in therapy, unmodified TCRs should be extensively testedfor cross-reactivity and cross-pairing tendency after TCR trans-fer into primary T cells. For better characterization of cross-pairing potential, instead of total PBMCs, specific T cells witha defined endogenous TCR repertoire could be used as hosts forTCR transfer (27, 52, 53). For example, EBV-specific T cellstransduced with CMV-TCRs would have the added benefit ofsimultaneous protection against both these HSCT-relevantpathogens (10). Additional safety mechanisms could be intro-duced to be able to eliminate TCR-transgenic T cells in thepatient if adverse effects are observed (54).
In summary, we isolated CMV-specific TCRs of differentHLA class I restrictions from memory-derived T cell clonescharacterized by efficient recognition of endogenously pro-cessed viral Ag. We demonstrated that these TCRs can be usedto rapidly prepare TCR-transgenic T cells equipped with anarsenal of specific antiviral functions. We believe that such
TCR-transgenic T cells qualify as candidates for a successfulclinical application in HSCT patients with a CMV-negative do-nor, a situation of considerable and potentially increasing clin-ical importance.
AcknowledgmentsWe thank Drs. Angela M. Krackhardt and Josef Mautner for stimulatingdiscussions and technical advice. We thank Dr. Wolfgang Uckert for gen-erously providing J76CD8 cells and the myeloproliferative sarcoma virusretroviral gene transfer technology.
DisclosuresThe authors have no financial conflict of interest.
References1. Boeckh, M., W. Leisenring, S. R. Riddell, R. A. Bowden, M. L. Huang, D. Myerson,
T. Stevens-Ayers, M. E. Flowers, T. Cunningham, and L. Corey. 2003. Late cyto-megalovirus disease and mortality in recipients of allogeneic hematopoietic stem celltransplants: importance of viral load and T-cell immunity. Blood 101: 407–414.
2. Reusser, P., S. R. Riddell, J. D. Meyers, and P. D. Greenberg. 1991. CytotoxicT-lymphocyte response to cytomegalovirus after human allogeneic bone marrowtransplantation: pattern of recovery and correlation with cytomegalovirus infec-tion and disease. Blood 78: 1373–1380.
3. Li, C. R., P. D. Greenberg, M. J. Gilbert, J. M. Goodrich, and S. R. Riddell. 1994.Recovery of HLA-restricted cytomegalovirus (CMV)-specific T-cell responsesafter allogeneic bone marrow transplant: correlation with CMV disease and effectof ganciclovir prophylaxis. Blood 83: 1971–1979.
4. Hebart, H., S. Daginik, S. Stevanovic, U. Grigoleit, A. Dobler, M. Baur, G. Rauser,C. Sinzger, G. Jahn, J. Loeffler, et al. 2002. Sensitive detection of human cytomeg-alovirus peptide-specific cytotoxic T-lymphocyte responses by interferon-�-enzyme-linked immunospot assay and flow cytometry in healthy individuals and in patientsafter allogeneic stem cell transplantation. Blood 99: 3830–3837.
5. Ganepola, S., C. Gentilini, U. Hilbers, T. Lange, K. Rieger, J. Hofmann, M. Maier,U. G. Liebert, D. Niederwieser, E. Engelmann, et al. 2007. Patients at high risk forCMV infection and disease show delayed CD8� T-cell immune recovery after al-logeneic stem cell transplantation. Bone Marrow Transplant. 39: 293–299.
6. Riddell, S. R., K. S. Watanabe, J. M. Goodrich, C. R. Li, M. E. Agha, andP. D. Greenberg. 1992. Restoration of viral immunity in immunodeficient humansby the adoptive transfer of T cell clones. Science 257: 238–241.
7. Einsele, H., E. Roosnek, N. Rufer, C. Sinzger, S. Riegler, J. Loffler, U. Grigoleit,A. Moris, H. G. Rammensee, L. Kanz, et al. 2002. Infusion of cytomegalovirus(CMV)-specific T cells for the treatment of CMV infection not responding toantiviral chemotherapy. Blood 99: 3916–3922.
FIGURE 8. Central memory markers on CMV-TCR-transgenic T cells during stimulation. PBMCs derived from CMV-seronegative donor 1 wereretrovirally transduced with the CMV-specific NLV2-TCR or NLV3-TCR followed by Ag-specific or nonspecific stimulation. A, The frequency ofCCR7-positive T cells was assessed at days 6, 13, and 20 after TCR transfer by staining with CCR7-specific Ab and NLV tetramer and subsequent flowcytometric analysis. The proportion of CCR7-positive cells within the NLV tetramer-positive population (f) as well as the proportion of CCR7-positivecells within the total T cell population (u) is shown. B, Similarly, the frequency of CD62L� T cells was assessed at days 6, 13, and 20 after TCR transferby staining with CD62L-specific Ab and NLV tetramer followed by flow cytometric analysis. The proportion of CD62L� cells within the NLV tetramer-positive population (f) as well as the proportion of CD62L� cells within total lymphocytes (u) is shown.
6829The Journal of Immunology
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

8. Peggs, K. S., S. Verfuerth, A. Pizzey, N. Khan, M. Guiver, P. A. Moss, andS. Mackinnon. 2003. Adoptive cellular therapy for early cytomegalovirus infec-tion after allogeneic stem-cell transplantation with virus-specific T-cell lines.Lancet 362: 1375–1377.
9. Cobbold, M., N. Khan, B. Pourgheysari, S. Tauro, D. McDonald, H. Osman,M. Assenmacher, L. Billingham, C. Steward, C. Crawley, et al. 2005. Adoptivetransfer of cytomegalovirus-specific CTL to stem cell transplant patients afterselection by HLA-peptide tetramers. J. Exp. Med. 202: 379–386.
10. Moss, P., and A. Rickinson. 2005. Cellular immunotherapy for viral infectionafter HSC transplantation. Nat. Rev. Immunol. 5: 9–20.
11. Borysiewicz, L. K., S. Graham, J. K. Hickling, P. D. Mason, and J. G. Sissons.1988. Human cytomegalovirus-specific cytotoxic T cells: their precursor fre-quency and stage specificity. Eur. J. Immunol. 18: 269–275.
12. Gillespie, G. M., M. R. Wills, V. Appay, C. O’Callaghan, M. Murphy, N. Smith,P. Sissons, S. Rowland-Jones, J. I. Bell, and P. A. Moss. 2000. Functional hetero-geneity and high frequencies of cytomegalovirus-specific CD8� T lymphocytes inhealthy seropositive donors. J. Virol. 74: 8140–8150.
13. Boeckh, M., and W. G. Nichols. 2004. The impact of cytomegalovirus serostatusof donor and recipient before hematopoietic stem cell transplantation in the era ofantiviral prophylaxis and preemptive therapy. Blood 103: 2003–2008.
14. Dembic, Z., W. Haas, S. Weiss, J. McCubrey, H. Kiefer, H. von Boehmer, andM. Steinmetz. 1986. Transfer of specificity by murine � and � T-cell receptorgenes. Nature 320: 232–238.
15. Clay, T. M., M. C. Custer, J. Sachs, P. Hwu, S. A. Rosenberg, and M. I. Nishimura.1999. Efficient transfer of a tumor antigen-reactive TCR to human peripheral bloodlymphocytes confers anti-tumor reactivity. J. Immunol. 163: 507–513.
16. Schumacher, T. N. 2002. T-cell-receptor gene therapy. Nat. Rev. Immunol. 2:512–519.
17. Stauss, H. J., M. Cesco-Gaspere, S. Thomas, D. P. Hart, S. A. Xue, A. Holler,G. Wright, M. Perro, A. M. Little, C. Pospori, J. King, and E. C. Morris. 2007.Monoclonal T-cell receptors: new reagents for cancer therapy. Mol. Ther. 15:1744–1750.
18. Morgan, R. A., M. E. Dudley, J. R. Wunderlich, M. S. Hughes, J. C. Yang,R. M. Sherry, R. E. Royal, S. L. Topalian, U. S. Kammula, N. P. Restifo, et al.2006. Cancer regression in patients after transfer of genetically engineered lym-phocytes. Science 314: 126–129.
19. Wills, M. R., A. J. Carmichael, K. Mynard, X. Jin, M. P. Weekes, B. Plachter, andJ. G. Sissons. 1996. The human cytotoxic T-lymphocyte (CTL) response to cy-tomegalovirus is dominated by structural protein pp65: frequency, specificity, andT-cell receptor usage of pp65-specific CTL. J. Virol. 70: 7569–7579.
20. Sylwester, A. W., B. L. Mitchell, J. B. Edgar, C. Taormina, C. Pelte, F. Ruchti,P. R. Sleath, K. H. Grabstein, N. A. Hosken, F. Kern, J. A. Nelson, and L. J. Picker.2005. Broadly targeted human cytomegalovirus-specific CD4� and CD8� T cellsdominate the memory compartments of exposed subjects. J. Exp. Med. 202:673–685.
21. Elkington, R., S. Walker, T. Crough, M. Menzies, J. Tellam, M. Bharadwaj, andR. Khanna. 2003. Ex vivo profiling of CD8�-T-cell responses to human cyto-megalovirus reveals broad and multispecific reactivities in healthy virus carriers.J. Virol. 77: 5226–5240.
22. Wiesner, M., C. Zentz, M. H. Hammer, M. Cobbold, F. Kern, H. J. Kolb,W. Hammerschmidt, R. Zeidler, and A. Moosmann. 2005. Selection of CMV-specific CD8� and CD4� T cells by mini-EBV-transformed B cell lines. Eur.J. Immunol. 35: 2110–2121.
23. Khan, N., M. Cobbold, R. Keenan, and P. A. Moss. 2002. Comparative analysisof CD8� T cell responses against human cytomegalovirus proteins pp65 andimmediate early 1 shows similarities in precursor frequency, oligoclonality, andphenotype. J. Infect. Dis. 185: 1025–1034.
24. Gandhi, M. K., M. R. Wills, G. Okecha, E. K. Day, R. Hicks, R. E. Marcus,J. G. Sissons, and A. J. Carmichael. 2003. Late diversification in the clonalcomposition of human cytomegalovirus-specific CD8� T cells following alloge-neic hemopoietic stem cell transplantation. Blood 102: 3427–3438.
25. Trautmann, L., M. Rimbert, K. Echasserieau, X. Saulquin, B. Neveu, J. Dechanet,V. Cerundolo, and M. Bonneville. 2005. Selection of T cell clones expressinghigh-affinity public TCRs within human cytomegalovirus-specific CD8 T cellresponses. J. Immunol. 175: 6123–6132.
26. Price, D. A., J. M. Brenchley, L. E. Ruff, M. R. Betts, B. J. Hill, M. Roederer,R. A. Koup, S. A. Migueles, E. Gostick, L. Wooldridge, et al. 2005. Avidity forantigen shapes clonal dominance in CD8� T cell populations specific for per-sistent DNA viruses. J. Exp. Med. 202: 1349–1361.
27. Heemskerk, M. H., R. S. Hagedoorn, M. A. van der Hoorn, L. T. van der Veken,M. Hoogeboom, M. G. Kester, R. Willemze, and J. H. Falkenburg. 2007. Efficiencyof T-cell receptor expression in dual-specific T cells is controlled by the intrinsicqualities of the TCR chains within the TCR-CD3 complex. Blood 109: 235–243.
28. Day, E. K., A. J. Carmichael, I. J. ten Berge, E. C. Waller, J. G. Sissons, andM. R. Wills. 2007. Rapid CD8� T cell repertoire focusing and selection of high-affinity clones into memory following primary infection with a persistent humanvirus: human cytomegalovirus. J. Immunol. 179: 3203–3213.
29. Brennan, R. M., J. J. Miles, S. L. Silins, M. J. Bell, J. M. Burrows, andS. R. Burrows. 2007. Predictable �� T-cell receptor selection toward an HLA-B*3501-restricted human cytomegalovirus epitope. J. Virol. 81: 7269–7273.
30. Venturi, V., H. Y. Chin, T. E. Asher, K. Ladell, P. Scheinberg, E. Bornstein,D. van Bockel, A. D. Kelleher, D. C. Douek, D. A. Price, and M. P. Davenport.2008. TCR �-chain sharing in human CD8� T cell responses to cytomegalovirusand EBV. J. Immunol. 181: 7853–7862.
31. Weekes, M. P., M. R. Wills, K. Mynard, A. J. Carmichael, and J. G. Sissons.1999. The memory cytotoxic T-lymphocyte (CTL) response to human cytomeg-alovirus infection contains individual peptide-specific CTL clones that have un-dergone extensive expansion in vivo. J. Virol. 73: 2099–2108.
32. Moosmann, A., N. Khan, M. Cobbold, C. Zentz, H. J. Delecluse, G. Hollweck,A. D. Hislop, N. W. Blake, D. Croom-Carter, B. Wollenberg, et al. 2002. B cellsimmortalized by a mini-Epstein-Barr virus encoding a foreign antigen efficientlyreactivate specific cytotoxic T cells. Blood 100: 1755–1764.
33. Wiesner, M., C. Zentz, C. Mayr, R. Wimmer, W. Hammerschmidt, R. Zeidler,and A. Moosmann. 2008. Conditional immortalization of human B cells by CD40ligation. PLoS ONE 3: e1464.
34. Heemskerk, M. H., M. Hoogeboom, R. A. de Paus, M. G. Kester,M. A. van der Hoorn, E. Goulmy, R. Willemze, and J. H. Falkenburg. 2003.Redirection of antileukemic reactivity of peripheral T lymphocytes using genetransfer of minor histocompatibility antigen HA-2-specific T-cell receptor com-plexes expressing a conserved � joining region. Blood 102: 3530–3540.
35. Roth, M. E., M. J. Lacy, L. K. McNeil, and D. M. Kranz. 1989. Analysis of T cellreceptor transcripts using the polymerase chain reaction. BioTechniques 7: 746–754.
36. Steinle, A., C. Reinhardt, P. Jantzer, and D. J. Schendel. 1995. In vivo expansionof HLA-B35 alloreactive T cells sharing homologous T cell receptors: evidencefor maintenance of an oligoclonally dominated allospecificity by persistent stim-ulation with an autologous MHC/peptide complex. J. Exp. Med. 181: 503–513.
37. Zhou, D., R. Srivastava, V. Grummel, S. Cepok, H. P. Hartung, and B. Hemmer.2006. High throughput analysis of TCR-beta rearrangement and gene expressionin single T cells. Lab. Invest. 86: 314–321.
38. Lefranc, M. P. 2003. IMGT databases, web resources and tools for immunoglobulinand T cell receptor sequence analysis: http://imgt.cines.fr. Leukemia 17: 260–266.
39. Engels, B., H. Cam, T. Schuler, S. Indraccolo, M. Gladow, C. Baum,T. Blankenstein, and W. Uckert. 2003. Retroviral vectors for high-level transgeneexpression in T lymphocytes. Hum. Gene Ther. 14: 1155–1168.
40. Sommermeyer, D., J. Neudorfer, M. Weinhold, M. Leisegang, B. Engels,E. Noessner, M. H. Heemskerk, J. Charo, D. J. Schendel, T. Blankenstein,H. Bernhard, and W. Uckert. 2006. Designer T cells by T cell receptor replace-ment. Eur. J. Immunol. 36: 3052–3059.
41. Leisegang, M., B. Engels, P. Meyerhuber, E. Kieback, D. Sommermeyer,S. A. Xue, S. Reuss, H. Stauss, and W. Uckert. 2008. Enhanced functionality ofT cell receptor-redirected T cells is defined by the transgene cassette. J. Mol.Med. 86: 573–583.
42. Sallusto, F., D. Lenig, R. Forster, M. Lipp, and A. Lanzavecchia. 1999. Twosubsets of memory T lymphocytes with distinct homing potentials and effectorfunctions. Nature 401: 708–712.
43. Rizzieri, D. A., P. Dev, G. D. Long, C. Gasparetto, K. M. Sullivan, M. Horwitz,J. Chute, and N. J. Chao. 2009. Response and toxicity of donor lymphocyteinfusions following T-cell depleted non-myeloablative allogeneic hematopoieticSCT from 3-6/6 HLA matched donors. Bone Marrow Transplant. 43: 327–333.
44. Haque, T., G. M. Wilkie, M. M. Jones, C. D. Higgins, G. Urquhart, P. Wingate,D. Burns, K. McAulay, M. Turner, C. Bellamy, et al. 2007. Allogeneic cytotoxicT-cell therapy for EBV-positive posttransplantation lymphoproliferative disease:results of a phase 2 multicenter clinical trial. Blood 110: 1123–1131.
45. Kleihauer, A., U. Grigoleit, H. Hebart, A. Moris, P. Brossart, A. Muhm,S. Stevanovic, H. G. Rammensee, C. Sinzger, S. Riegler, et al. 2001. Ex vivogeneration of human cytomegalovirus-specific cytotoxic T cells by peptide-pulsed dendritic cells. Br. J. Haematol. 113: 231–239.
46. Szmania, S., A. Galloway, M. Bruorton, P. Musk, G. Aubert, A. Arthur, H. Pyle,N. Hensel, N. Ta, L. Lamb, Jr., et al. 2001. Isolation and expansion of cytomeg-alovirus-specific cytotoxic T lymphocytes to clinical scale from a single blooddraw using dendritic cells and HLA-tetramers. Blood 98: 505–512.
47. Kondo, E., Y. Akatsuka, K. Kuzushima, K. Tsujimura, S. Asakura, K. Tajima,Y. Kagami, Y. Kodera, M. Tanimoto, Y. Morishima, and T. Takahashi. 2004.Identification of novel CTL epitopes of CMV-pp65 presented by a variety ofHLA alleles. Blood 103: 630–638.
48. Arstila, T. P., A. Casrouge, V. Baron, J. Even, J. Kanellopoulos, and P. Kourilsky.1999. A direct estimate of the human �� T cell receptor diversity. Science 286:958–961.
49. van der Veken, L. T., R. S. Hagedoorn, M. M. van Loenen, R. Willemze,J. H. Falkenburg, and M. H. Heemskerk. 2006. �� T-cell receptor engineered ��T cells mediate effective antileukemic reactivity. Cancer Res. 66: 3331–3337.
50. Van Lent, A. U., M. Nagasawa, M. M. van Loenen, R. Schotte,T. N. Schumacher, M. H. Heemskerk, H. Spits, and N. Legrand. 2007. Functionalhuman antigen-specific T cells produced in vitro using retroviral T cell receptortransfer into hematopoietic progenitors. J. Immunol. 179: 4959–4968.
51. Uckert, W., and T. N. Schumacher. 2009. TCR transgenes and transgene cassettes forTCR gene therapy: status in 2008. Cancer Immunol. Immunother. 58: 809–822.
52. Heemskerk, M. H., M. Hoogeboom, R. Hagedoorn, M. G. Kester, R. Willemze, andJ. H. Falkenburg. 2004. Reprogramming of virus-specific T cells into leukemia-re-active T cells using T cell receptor gene transfer. J. Exp. Med. 199: 885–894.
53. van Loenen, M. M., R. S. Hagedoorn, M. G. Kester, M. Hoogeboom,R. Willemze, J. H. Falkenburg, and M. H. Heemskerk. 2009. Kinetic preservationof dual specificity of coprogrammed minor histocompatibility antigen-reactivevirus-specific T cells. Cancer Res. 69: 2034–2041.
54. Kieback, E., J. Charo, D. Sommermeyer, T. Blankenstein, and W. Uckert. 2008.A safeguard eliminates T cell receptor gene-modified autoreactive T cells afteradoptive transfer. Proc. Natl. Acad. Sci. USA 105: 623–628.
6830 CMV-SPECIFIC TCR-TRANSGENIC T CELLS FOR IMMUNOTHERAPY
by guest on January 15, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
Related Documents











![IMMUNOGLOBULINE E T CELL RECEPTOR T. Strachan e A.P. … · B cell antigen receptor tetramero [ IgH 2 + IgL 2 (Ig oppure Ig )] T cell receptor (TCR) eterodimero TCR /TCR TCR /TCR](https://static.cupdf.com/doc/110x72/5c017b5c09d3f26f1e8cc6a0/immunoglobuline-e-t-cell-receptor-t-strachan-e-ap-b-cell-antigen-receptor.jpg)