Received 13 May 2003 Accepted 25 June 2003 Published online 30 September 2003 Climate variation and regional gradients in population dynamics of two hole-nesting passerines Bernt-Erik Sæther 1 * , Steinar Engen 2 , Anders Pape Møller 3 , Erik Matthysen 4 , Frank Adriaensen 4 , Wolfgang Fiedler 5 , Agu Leivits 6 , Marcel M. Lambrechts 7 , Marcel E. Visser 8 , Tycho Anker-Nilssen 9 , Christiaan Both 8 , Andre ´ A. Dhondt 4,10 , Robin H. McCleery 11 , John McMeeking 8 , Jamie Potti 12 , Ole Wiggo Røstad 13 and David Thomson 8 1 Department of Biology, and 2 Department of Mathematical Sciences, Norwegian University of Science and Technology, N-7491 Trondheim, Norway 3 Laboratoire de Parasitologie E ´ volutive, CNRS UMR 7103, Universite ´ Pierre et Marie Curie, Ba ˆ timent A, 7e `me e ´tage, 7 quai St Bernard, Case 237, FR-75252 Paris Cedex 05, France 4 Laboratory of Animal Ecology, Department of Biology, University of Antwerp, B-2610 Antwerp, Belgium 5 Max-Planck-Forschungsstelle fu ¨r Ornithologie, Andechs und Radolfzell, Vogelwarte Radolfzell, Schloss Moeggingen, D-78315 Radolfzell, Germany 6 Nigula Nature Reserve Administration, Aia 22-18, Kilingi-No ˜mme 86301, Estonia 7 Laboratoire Centre d’Ecologie Fonctionnelle et Evolutive, Centre National de la Recherche Scientifique, 1919 Route de Mende, F-34293 Montpellier Cedex 5, France 8 Netherlands Institute of Ecology, PO Box 40, 6666 ZG Heteren, The Netherlands 9 Norwegian Institute for Nature Research, Tungasletta 2, N-7485 Trondheim, Norway 10 Laboratory of Ornithology, Cornell University, 159 Sapsucker Road, Ithaca, NY 14850, USA 11 Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK 12 Department of Biologia Animal, Facility of Biology, University of Alcala, E-28871 Alcala de Henares, Spain 13 Agricultural University of Norway, Department of Nature Conservation, PO Box 5014, NO-1432 A Ê s, Norway Latitudinal gradients in population dynamics can arise through regional variation in the deterministic components of the population dynamics and the stochastic factors. Here, we demonstrate an increase with latitude in the contribution of a large-scale climate pattern, the North Atlantic Oscillation (NAO), to the fluctuations in size of populations of two European hole-nesting passerine species. However, this influence of climate induced different latitudinal gradients in the population dynamics of the two species. In the great tit the proportion of the variability in the population fluctuations explained by the NAO increased with latitude, showing a larger impact of climate on the population fluctuations of this species at higher latitudes. In contrast, no latitudinal gradient was found in the relative contribution of climate to the variability of the pied flycatcher populations because the total environmental stochasticity increased with latitude. This shows that the population ecological consequences of an expected climate change will depend on how climate affects the environmental stochasticity in the population process. In both species, the effects will be larger in those parts of Europe where large changes in climate are expected. Keywords: climate change; density dependence; great tit; North Atlantic Oscillation; pied flycatcher; stochastic population dynamics 1. INTRODUCTION The relative importance of density dependence and stoch- astic factors for variation in size of natural populations has, since the publication of the influential books by Andrewar- tha & Birch (1954) and Lack (1954), been one of the most central problems in ecology (reviewed in Turchin 1995). In recent years this question has again received increased attention because of the need to predict the ecological impact of the expected changes in climate. In spite of this increased focus, few hypotheses have been presented under which conditions stochastic variation in the * Author for correspondence ([email protected]). Proc. R. Soc. Lond. B (2003) 270, 2397–2404 2397 Ó 2003 The Royal Society DOI 10.1098/rspb.2003.2499 environment is expected to have large influence on the dynamics of natural populations. One reason for why so few general hypotheses have been developed for predicting variation in the effects on the population fluctuations of stochastic variation in the environment may be that two types of stochasticity affect the population dynamics, which is difficult to separate by using traditional time-series analysis. Demographic stoch- asticity is caused by random variation among individuals in their contribution to fitness, whereas environmental stochasticity arises from random effects that affect the whole or parts of the population at the same time (Lande et al. 2003). These stochastic effects affect the variance in population size differentially: demographic variance s 2 d is most important at smaller population sizes, whereas the effects of the environmental variance s 2 e increase with the

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Received 13 May 2003Accepted 25 June 2003
Published online 30 September 2003
Climate variation and regional gradients in populationdynamics of two hole-nesting passerinesBernt-Erik Saeligther1 Steinar Engen2 Anders Pape Moslashller3 Erik Matthysen4Frank Adriaensen4 Wolfgang Fiedler5 Agu Leivits6 Marcel M Lambrechts7Marcel E Visser8 Tycho Anker-Nilssen9 Christiaan Both8Andre A Dhondt410 Robin H McCleery11 John McMeeking8 Jamie Potti12Ole Wiggo Roslashstad13 and David Thomson8
1Department of Biology and 2Department of Mathematical Sciences Norwegian University of Science and TechnologyN-7491 Trondheim Norway3Laboratoire de Parasitologie Evolutive CNRS UMR 7103 Universite Pierre et Marie Curie Batiment A 7eme etage7 quai St Bernard Case 237 FR-75252 Paris Cedex 05 France4Laboratory of Animal Ecology Department of Biology University of Antwerp B-2610 Antwerp Belgium5Max-Planck-Forschungsstelle fur Ornithologie Andechs und Radolfzell Vogelwarte Radolfzell Schloss MoeggingenD-78315 Radolfzell Germany6Nigula Nature Reserve Administration Aia 22-18 Kilingi-Nomme 86301 Estonia7Laboratoire Centre drsquoEcologie Fonctionnelle et Evolutive Centre National de la Recherche Scientifique1919 Route de Mende F-34293 Montpellier Cedex 5 France8Netherlands Institute of Ecology PO Box 40 6666 ZG Heteren The Netherlands9Norwegian Institute for Nature Research Tungasletta 2 N-7485 Trondheim Norway1 0Laboratory of Ornithology Cornell University 159 Sapsucker Road Ithaca NY 14850 USA1 1Department of Zoology University of Oxford South Parks Road Oxford OX1 3PS UK1 2Department of Biologia Animal Facility of Biology University of Alcala E-28871 Alcala de Henares Spain1 3Agricultural University of Norway Department of Nature Conservation PO Box 5014 NO-1432 AEcirc s Norway
Latitudinal gradients in population dynamics can arise through regional variation in the deterministiccomponents of the population dynamics and the stochastic factors Here we demonstrate an increasewith latitude in the contribution of a large-scale climate pattern the North Atlantic Oscillation (NAO)to the fluctuations in size of populations of two European hole-nesting passerine species However thisinfluence of climate induced different latitudinal gradients in the population dynamics of the two speciesIn the great tit the proportion of the variability in the population fluctuations explained by the NAOincreased with latitude showing a larger impact of climate on the population fluctuations of this speciesat higher latitudes In contrast no latitudinal gradient was found in the relative contribution of climateto the variability of the pied flycatcher populations because the total environmental stochasticity increasedwith latitude This shows that the population ecological consequences of an expected climate change willdepend on how climate affects the environmental stochasticity in the population process In both speciesthe effects will be larger in those parts of Europe where large changes in climate are expected
Keywords climate change density dependence great tit North Atlantic Oscillation pied flycatcherstochastic population dynamics
1 INTRODUCTION
The relative importance of density dependence and stoch-astic factors for variation in size of natural populations hassince the publication of the influential books by Andrewar-tha amp Birch (1954) and Lack (1954) been one of the mostcentral problems in ecology (reviewed in Turchin 1995)In recent years this question has again received increasedattention because of the need to predict the ecologicalimpact of the expected changes in climate In spite of thisincreased focus few hypotheses have been presentedunder which conditions stochastic variation in the
Author for correspondence (bernt-eriksatherbiontnuno)
Proc R Soc Lond B (2003) 270 2397ndash2404 2397 Oacute 2003 The Royal SocietyDOI 101098rspb20032499
environment is expected to have large influence on thedynamics of natural populations
One reason for why so few general hypotheses havebeen developed for predicting variation in the effects onthe population fluctuations of stochastic variation in theenvironment may be that two types of stochasticity affectthe population dynamics which is difficult to separate byusing traditional time-series analysis Demographic stoch-asticity is caused by random variation among individualsin their contribution to fitness whereas environmentalstochasticity arises from random effects that affect thewhole or parts of the population at the same time (Landeet al 2003) These stochastic effects affect the variance inpopulation size differentially demographic variance s 2
d ismost important at smaller population sizes whereas theeffects of the environmental variance s 2
e increase with the
2398 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
square of population size and thus are important in largepopulations as well (Lande 1998) In addition small vari-ations in parameters specifying the deterministic compo-nent of the population dynamics eg the specific growthrate may also affect the population characteristics (May1976 Lande et al 2003) Hence a proper understandingof the effects of environmental variation on populationfluctuations must include estimates of parameters describ-ing the deterministic as well as the stochastic componentsof the dynamics
Basically two alternative hypotheses can be derivedunder which conditions stochastic variation in an environ-mental variable is expected to have a greater influence onthe dynamics of a population One hypothesis may be thata large effect of variation in the environment may beexpected in populations where the stochastic componentsof the population dynamics are large Alternativelyenvironmental stochasticity may more strongly affect thefluctuations of populations with a small stochastic compo-nent because in highly variable populations the effects ofclimate variability will be cancelled out by other factors(see Pimm (1991) for a similar line of argument)
Many important features of the environment relate tovariation in climate The North Atlantic Oscillation(NAO) is one such large-scale climate phenomenon thatis correlated to annual variation in local climate over largeareas (Hurrell 1995 Mysterud et al 2002) The NAO rep-resents changes in the subtropic atmospheric high-press-ure zone centred over the Azores and the low-pressurezone over Iceland and may also be interrelated with otherlarge-scale climate patterns (Visbeck et al 2001) In gen-eral high values of the NAO-index during winter are asso-ciated with strong wind circulation in the North Atlanticcausing an increase in temperatures and precipitation innorthern Europe but dry weather in the Mediterraneanregion (Hurrell 1995) By contrast negative values of theNAO are associated with a decrease in temperature andprecipitation in northwestern Europe
Several studies have documented that variation in cli-mate may affect demographic characteristics of small pass-erines (McCarty 2001 Stenseth et al 2002 Walther et al2002) For instance an advance in laying date withincrease in spring temperature has occurred in manypopulations of the great tit (Parus major L (Winkel ampHudde 1997 McCleery amp Perrins 1998 Visser et al2003)) and the pied flycatcher (Ficedula hypoleuca L(Winkel amp Hudde 1997 Both amp Visser 2001 Sanz et al2003)) which also may be explained by variation in theNAO (Forchammer et al 1998 Sanz 2002 2003) How-ever population dynamical responses to changes in cli-mate have been more difficult to document (but seeSaeligther et al 2000)
In this paper we will using a newly developed theoreti-cal framework (Lande et al 2003) relate fluctuations inthe size of great tit and pied flycatcher populations distrib-uted across Europe to variation in the NAO We will thentry to derive latitudinal gradients in the populationdynamical characteristics of the two species especiallyfocusing on how the NAO affects the noise structure ofthe population process Our aim will be to examinewhether these effects will be similar in the sedentary greattit and the migratory pied flycatcher suggesting generalrelationships for how a large-scale climate phenomenon
Proc R Soc Lond B (2003)
such as the NAO affects the population dynamics withinthe distributional range of a species
2 POPULATION MODEL
Let N be the population size in a given year and N 1DN the population size the following year The stochas-ticity in the population dynamics is then given byvar(DN|N) = N 2s 2
e 1 Ns 2d where s 2
e and s 2d denote the
environmental and demographic variance respectivelyThe logistic model (May 1981) E(DN) = rN(1 2 N K )where r is the mean specific population growth rate andK the carrying capacity has been found to describe thedensity regulation in many bird species quite well(Saeligther amp Engen 2002) Thus for small and moderatefluctuations in population size we find writing X = lnNand using the first-order approximation DX lt DN N thatthe expected change in log population size is
E(DX|X) = s 1 aeX 2 12 s 2
de2X (21)
where the stochastic population growth rate s = r 212s 2e
and a = 2 rK We then partition the environmental variance s 2
e into acomponent Z due to linear effects of the NAO and aresidual component s 2
re s (Engen et al 1998 Saeligther et al2000) so that the environmental stochastic componentof the change in log population size becomesUse = bN A O Z 1 Vsre s where Z is the NAO-index U andV are standardized variables with zero mean and unit vari-ance and bN A O is the regression coefficient Hence thetotal environmental variance is
s 2e = b2
N A O varZ 1 s 2re s = s 2
N A O 1 s 2re s (22)
Using Z as a known covariate we then obtain the model
E(DX|X) = s 2 12 s 2
de2X 1 aeX 1 bN A O Z (23a)
and
var(DX|X) = s 2re s 1 s 2
de2X (23b)
3 METHODS
(a) Population dataWe compiled long-term (10 years or more) time-series of
population fluctuations of the great tit and pied flycatcher fromdifferent parts of Europe (figure 1) Only study sites with arti-ficial nest-boxes were included resulting in more precise popu-lation estimates than are usually recorded in studies of naturalpopulations To avoid problems with N = 0 we include onlypopulations where the estimate of K was larger than 10 Forestimates of the population parameters see Appendix A
(b) Climate dataThe NAO is a global climate phenomenon (Visbeck et al
2001) and is expressed by an index based on the difference ofnormalized sea level pressures between Ponta Delgada AzoresPortugal and Stykkisholmur Iceland from 1864 through to2002 for the winter period December through to March(httpwwwcgducareducasclimind)
(c) Estimation of parametersThe demographic variance s 2
d was estimated from data onindividual variation among females in their fitness contributions
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2399
Figure 1 Location of the study areas Triangles represent pied flycatcher study sites squares represent study sites with greattit populations and circles represent study sites where time-series of both species were present Some adjacent study sites arerepresented only by one symbol
to the following generations (Lande et al 2003) The total con-tribution of a female i in year t (Ri) is the number of femaleoffspring born during the year that survive for at least 1 yearplus 1 if the female survives to the next year (Saeligther et al 1998)The demographic variance was estimated (Lande et al 2003)as the weighted mean across years of s 2
d(t) = E(1a21)S (Ri 2 R)2 where R is the mean contribution of the individualsand a is the number of recorded contributions in year tEstimates of s 2
d were obtained for 10 great tit populations andthree pied flycatcher populations (see Appendix A) When noindividual data were available the species-specific mean of s 2
d
was usedThe other parameters in equations (23) were estimated by
maximum likelihood using the full-likelihood function for theprocess obtained by assuming that DX conditioned on X is nor-mally distributed Uncertainties in the estimates were found byparametric bootstrapping simulating new sets of data from thefitted model (Efron amp Tibshirani 1995) To test the hypothesisthat b = 0 we first carried out the estimation for the reducedmodel with b = 0 and performed bootstrap simulations using theparameters estimated from this model and b = 0 The p-valuefor the test is then obtained as the fractions of the simulationsgiving an estimate of b larger than b The results of the testsare presented in Appendix A
The diffusion approximation to the quasi-stationary distri-bution of population sizes
Proc R Soc Lond B (2003)
f (N N0) = G (N N0)Y E `
1
G (N N0)du (31)
can be derived following Lande et al (2003) Here G(N N0) isthe Green function or the sojourn time starting from an initialsize N0 The Green function expresses the expected cumulativetime spent at each population size before extinction (Karlin ampTaylor 1981) We first computed the variance in the quasi-stationary distribution using N 0 = K that can be explained byvariation in the NAO VNAO using s 2
NAO and then repeated thecomputation with s 2
res to get the residual variance Vres The rela-tive contribution to the variance in the quasi-stationary distri-bution of variation in the NAO then becomes VNAO(VNAO 1
Vres) To facilitate the comparison among populations all vari-ances were standardized in relation to K
4 RESULTS
The stochastic population growth rate s (defined byequation (21)) differed significantly between the twospecies (table 1 F = 2127 df = 167 p 0001) with asmaller mean value of s in the pied flycatcher than in thegreat tit populations No significant ( p 01) interspe-cific variation was present in K or in any components ofthe environmental variance s 2
e
2400 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
Table 1 The mean (x plusmn sd) of the estimated population parameters for the two species(s is the stochastic specific growth rate s 2
d the demographic variance s 2e the environmental variance s 2
NAO the component ofs 2
e due to variation in the NAO-index s 2res the residual component not explained by the NAO and n the number of study
populations)
great tit pied flycatcher
x plusmn sd range n x plusmn sd range n
s 089 plusmn 038 010ndash158 40 049 plusmn 031 014ndash089 28K 56 plusmn 49 11ndash216 40 60 plusmn 58 13ndash290 28s2
d 048 plusmn 015 030ndash077 10 040 plusmn 014 031ndash056 3s2
e 0070 plusmn 0049 0002ndash0250 40 0052 plusmn 0047 0010ndash0211 28s2
NAO 0009 plusmn 0013 000ndash0042 40 0006 plusmn 0008 0000ndash0035 28s2
res 0061 plusmn 0046 0002ndash0209 40 0045 plusmn 0041 0007ndash0176 28
012
008
004
0
012
008
004
0
40 45 50 55
latitude (cN)
abs
(
NA
O)
60 65 40 45 50 55
latitude (cN)
60 65 70 75
b
(a) (b)
abs
(
NA
O)
b
Figure 2 Latitudinal gradients in the absolute value of the regression coefficient of the NAO effect bNAO in (a) great tit and(b) pied flycatcher populations across Europe Filled circles denote positive values and open circles denote negative values ofbNAO
Intraspecific variation in s 2e or in its two components
was not significantly ( p 01) related to s in any of thespecies This is in contrast to previous cross-species com-parisons in birds where a close relationship between para-meters describing the deterministic characteristics and theenvironmental stochasticity have been documented(Saeligther amp Engen 2002 Saeligther et al 2002)
We then examined variation in the component of theenvironmental stochasticity due to fluctuations in theNAO Variation in the NAO was able to explain a signifi-cant proportion of the variance in population size in 20(n = 40) of the great tit and in 21 (n = 29) of the piedflycatcher populations A latitudinal gradient was presentin the relative contribution of the NAO to the populationfluctuations of both species because there was an increasein the absolute value of the regression coefficient bN A O
with latitude (figure 2 correlation coefficient = 0463p = 0003 n = 40 and correlation coefficient = 0378p = 0048 n = 29 in the great tit and pied flycatcherrespectively) However the sign of the effects of the NAOdiffered in both species over relatively short distances(figure 2) in some populations a positive relationship waspresent whereas in others N decreased with increasingNAO Hence the correlation coefficient with latitude afterincluding the sign of bN A O was insignificant (correlationcoefficient = 20153 n = 40 and correlation coefficient =0038 n = 29 p 034 in the great tit and pied fly-catcher respectively)
Proc R Soc Lond B (2003)
These effects of the NAO were associated with latitudi-nal gradients in the effects of environmental stochasticityon the population dynamics In both the great tit and piedflycatcher s 2
N A O increased with latitude (figure 3ab corre-lation coefficient = 0439 p = 0005 n = 40 and corre-lation coefficient = 0487 p = 0007 n = 29 in the great titand pied flycatcher respectively) In the pied flycatcherthe residual component remaining after accounting for theNAO s2
re s increased with latitude (figure 3d correlationcoefficient = 0454 p = 0013 n = 29) resulting in a lati-tudinal gradient in environmental stochasticity in thisspecies (figure 3 f correlation coefficient = 0478p = 0009 n = 29) By contrast s 2
re s and s 2e were inde-
pendent of latitude in the great tit (figure 3ce correlationcoefficient = 20003 and correlation coefficient = 0114p 048 n = 40 for s2
re s and s2e respectively) Conse-
quently the relative contribution of variation in the NAOto s 2
e of the great tit increased with latitude (figure 3gcorrelation coefficient = 0334 p = 0035 n = 40)whereas no such significant relationship was present in thepied flycatcher (figure 3h correlation coefficient = 20179 p 035 n = 29) As a consequence the contri-bution of the NAO to the variance in the quasi-stationarydistribution of population sizes (equation (31)) increasedwith latitude in the great tit (correlationcoefficient = 0326 p = 0034 n = 40) Finally the vari-ance in the population size explained by variation in theNAO increased in both species with the residual compo-
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2401
004N
AO
(a)
003
002
001
040 45 50 55 60 65
004 (b)
003
002
001
040 45 50 55 60 65 70 75
020
res
(c)
015
010
005
040 45 50 55 60 65
020 (d )
015
010
005
040 45 50 55 60 65 70 75
025
2
(e)
020
015
010
005
040 45 50 55 60 65
020
( f )
015
010
005
040 45 50 55 60 65 70 75
006
rela
tive
con
trib
utio
nof
NA
O to
e
005
(g)
004
003
002
001
0
40 45 50
latitude (cN)
55 60 65
(h)08
06
04
02
40 45 50
latitude (cN)
55 60 65 70 75
ss
ss
22
2e
NA
Ore
s 2
rela
tive
con
trib
utio
nof
NA
O to
e
ss
ss
22
2e
Figure 3 Latitudinal gradients in the component of the environmental stochasticity explained by (ab) variation in the NAOs 2
NAO (cd) in residual component of the environmental stochasticity s 2res (e f ) in total environmental variance s 2
e and (gh) inproportion of s 2
e explained by variation in the NAO in great tit (aceg) and pied flycatcher (bd fh) populations across Europe
nent of the environmental variance in population size(correlation coefficient = 0475 p = 0002 n = 40 and cor-relation coefficient = 0636 n = 29 p 0001 in the greattit and pied flycatcher respectively) This shows that con-
Proc R Soc Lond B (2003)
tributions of large-scale climate patterns to variation inpopulation size increased with increasing influence ofenvironmental stochasticity on the population fluctu-ations
2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs

2398 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
square of population size and thus are important in largepopulations as well (Lande 1998) In addition small vari-ations in parameters specifying the deterministic compo-nent of the population dynamics eg the specific growthrate may also affect the population characteristics (May1976 Lande et al 2003) Hence a proper understandingof the effects of environmental variation on populationfluctuations must include estimates of parameters describ-ing the deterministic as well as the stochastic componentsof the dynamics
Basically two alternative hypotheses can be derivedunder which conditions stochastic variation in an environ-mental variable is expected to have a greater influence onthe dynamics of a population One hypothesis may be thata large effect of variation in the environment may beexpected in populations where the stochastic componentsof the population dynamics are large Alternativelyenvironmental stochasticity may more strongly affect thefluctuations of populations with a small stochastic compo-nent because in highly variable populations the effects ofclimate variability will be cancelled out by other factors(see Pimm (1991) for a similar line of argument)
Many important features of the environment relate tovariation in climate The North Atlantic Oscillation(NAO) is one such large-scale climate phenomenon thatis correlated to annual variation in local climate over largeareas (Hurrell 1995 Mysterud et al 2002) The NAO rep-resents changes in the subtropic atmospheric high-press-ure zone centred over the Azores and the low-pressurezone over Iceland and may also be interrelated with otherlarge-scale climate patterns (Visbeck et al 2001) In gen-eral high values of the NAO-index during winter are asso-ciated with strong wind circulation in the North Atlanticcausing an increase in temperatures and precipitation innorthern Europe but dry weather in the Mediterraneanregion (Hurrell 1995) By contrast negative values of theNAO are associated with a decrease in temperature andprecipitation in northwestern Europe
Several studies have documented that variation in cli-mate may affect demographic characteristics of small pass-erines (McCarty 2001 Stenseth et al 2002 Walther et al2002) For instance an advance in laying date withincrease in spring temperature has occurred in manypopulations of the great tit (Parus major L (Winkel ampHudde 1997 McCleery amp Perrins 1998 Visser et al2003)) and the pied flycatcher (Ficedula hypoleuca L(Winkel amp Hudde 1997 Both amp Visser 2001 Sanz et al2003)) which also may be explained by variation in theNAO (Forchammer et al 1998 Sanz 2002 2003) How-ever population dynamical responses to changes in cli-mate have been more difficult to document (but seeSaeligther et al 2000)
In this paper we will using a newly developed theoreti-cal framework (Lande et al 2003) relate fluctuations inthe size of great tit and pied flycatcher populations distrib-uted across Europe to variation in the NAO We will thentry to derive latitudinal gradients in the populationdynamical characteristics of the two species especiallyfocusing on how the NAO affects the noise structure ofthe population process Our aim will be to examinewhether these effects will be similar in the sedentary greattit and the migratory pied flycatcher suggesting generalrelationships for how a large-scale climate phenomenon
Proc R Soc Lond B (2003)
such as the NAO affects the population dynamics withinthe distributional range of a species
2 POPULATION MODEL
Let N be the population size in a given year and N 1DN the population size the following year The stochas-ticity in the population dynamics is then given byvar(DN|N) = N 2s 2
e 1 Ns 2d where s 2
e and s 2d denote the
environmental and demographic variance respectivelyThe logistic model (May 1981) E(DN) = rN(1 2 N K )where r is the mean specific population growth rate andK the carrying capacity has been found to describe thedensity regulation in many bird species quite well(Saeligther amp Engen 2002) Thus for small and moderatefluctuations in population size we find writing X = lnNand using the first-order approximation DX lt DN N thatthe expected change in log population size is
E(DX|X) = s 1 aeX 2 12 s 2
de2X (21)
where the stochastic population growth rate s = r 212s 2e
and a = 2 rK We then partition the environmental variance s 2
e into acomponent Z due to linear effects of the NAO and aresidual component s 2
re s (Engen et al 1998 Saeligther et al2000) so that the environmental stochastic componentof the change in log population size becomesUse = bN A O Z 1 Vsre s where Z is the NAO-index U andV are standardized variables with zero mean and unit vari-ance and bN A O is the regression coefficient Hence thetotal environmental variance is
s 2e = b2
N A O varZ 1 s 2re s = s 2
N A O 1 s 2re s (22)
Using Z as a known covariate we then obtain the model
E(DX|X) = s 2 12 s 2
de2X 1 aeX 1 bN A O Z (23a)
and
var(DX|X) = s 2re s 1 s 2
de2X (23b)
3 METHODS
(a) Population dataWe compiled long-term (10 years or more) time-series of
population fluctuations of the great tit and pied flycatcher fromdifferent parts of Europe (figure 1) Only study sites with arti-ficial nest-boxes were included resulting in more precise popu-lation estimates than are usually recorded in studies of naturalpopulations To avoid problems with N = 0 we include onlypopulations where the estimate of K was larger than 10 Forestimates of the population parameters see Appendix A
(b) Climate dataThe NAO is a global climate phenomenon (Visbeck et al
2001) and is expressed by an index based on the difference ofnormalized sea level pressures between Ponta Delgada AzoresPortugal and Stykkisholmur Iceland from 1864 through to2002 for the winter period December through to March(httpwwwcgducareducasclimind)
(c) Estimation of parametersThe demographic variance s 2
d was estimated from data onindividual variation among females in their fitness contributions
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2399
Figure 1 Location of the study areas Triangles represent pied flycatcher study sites squares represent study sites with greattit populations and circles represent study sites where time-series of both species were present Some adjacent study sites arerepresented only by one symbol
to the following generations (Lande et al 2003) The total con-tribution of a female i in year t (Ri) is the number of femaleoffspring born during the year that survive for at least 1 yearplus 1 if the female survives to the next year (Saeligther et al 1998)The demographic variance was estimated (Lande et al 2003)as the weighted mean across years of s 2
d(t) = E(1a21)S (Ri 2 R)2 where R is the mean contribution of the individualsand a is the number of recorded contributions in year tEstimates of s 2
d were obtained for 10 great tit populations andthree pied flycatcher populations (see Appendix A) When noindividual data were available the species-specific mean of s 2
d
was usedThe other parameters in equations (23) were estimated by
maximum likelihood using the full-likelihood function for theprocess obtained by assuming that DX conditioned on X is nor-mally distributed Uncertainties in the estimates were found byparametric bootstrapping simulating new sets of data from thefitted model (Efron amp Tibshirani 1995) To test the hypothesisthat b = 0 we first carried out the estimation for the reducedmodel with b = 0 and performed bootstrap simulations using theparameters estimated from this model and b = 0 The p-valuefor the test is then obtained as the fractions of the simulationsgiving an estimate of b larger than b The results of the testsare presented in Appendix A
The diffusion approximation to the quasi-stationary distri-bution of population sizes
Proc R Soc Lond B (2003)
f (N N0) = G (N N0)Y E `
1
G (N N0)du (31)
can be derived following Lande et al (2003) Here G(N N0) isthe Green function or the sojourn time starting from an initialsize N0 The Green function expresses the expected cumulativetime spent at each population size before extinction (Karlin ampTaylor 1981) We first computed the variance in the quasi-stationary distribution using N 0 = K that can be explained byvariation in the NAO VNAO using s 2
NAO and then repeated thecomputation with s 2
res to get the residual variance Vres The rela-tive contribution to the variance in the quasi-stationary distri-bution of variation in the NAO then becomes VNAO(VNAO 1
Vres) To facilitate the comparison among populations all vari-ances were standardized in relation to K
4 RESULTS
The stochastic population growth rate s (defined byequation (21)) differed significantly between the twospecies (table 1 F = 2127 df = 167 p 0001) with asmaller mean value of s in the pied flycatcher than in thegreat tit populations No significant ( p 01) interspe-cific variation was present in K or in any components ofthe environmental variance s 2
e
2400 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
Table 1 The mean (x plusmn sd) of the estimated population parameters for the two species(s is the stochastic specific growth rate s 2
d the demographic variance s 2e the environmental variance s 2
NAO the component ofs 2
e due to variation in the NAO-index s 2res the residual component not explained by the NAO and n the number of study
populations)
great tit pied flycatcher
x plusmn sd range n x plusmn sd range n
s 089 plusmn 038 010ndash158 40 049 plusmn 031 014ndash089 28K 56 plusmn 49 11ndash216 40 60 plusmn 58 13ndash290 28s2
d 048 plusmn 015 030ndash077 10 040 plusmn 014 031ndash056 3s2
e 0070 plusmn 0049 0002ndash0250 40 0052 plusmn 0047 0010ndash0211 28s2
NAO 0009 plusmn 0013 000ndash0042 40 0006 plusmn 0008 0000ndash0035 28s2
res 0061 plusmn 0046 0002ndash0209 40 0045 plusmn 0041 0007ndash0176 28
012
008
004
0
012
008
004
0
40 45 50 55
latitude (cN)
abs
(
NA
O)
60 65 40 45 50 55
latitude (cN)
60 65 70 75
b
(a) (b)
abs
(
NA
O)
b
Figure 2 Latitudinal gradients in the absolute value of the regression coefficient of the NAO effect bNAO in (a) great tit and(b) pied flycatcher populations across Europe Filled circles denote positive values and open circles denote negative values ofbNAO
Intraspecific variation in s 2e or in its two components
was not significantly ( p 01) related to s in any of thespecies This is in contrast to previous cross-species com-parisons in birds where a close relationship between para-meters describing the deterministic characteristics and theenvironmental stochasticity have been documented(Saeligther amp Engen 2002 Saeligther et al 2002)
We then examined variation in the component of theenvironmental stochasticity due to fluctuations in theNAO Variation in the NAO was able to explain a signifi-cant proportion of the variance in population size in 20(n = 40) of the great tit and in 21 (n = 29) of the piedflycatcher populations A latitudinal gradient was presentin the relative contribution of the NAO to the populationfluctuations of both species because there was an increasein the absolute value of the regression coefficient bN A O
with latitude (figure 2 correlation coefficient = 0463p = 0003 n = 40 and correlation coefficient = 0378p = 0048 n = 29 in the great tit and pied flycatcherrespectively) However the sign of the effects of the NAOdiffered in both species over relatively short distances(figure 2) in some populations a positive relationship waspresent whereas in others N decreased with increasingNAO Hence the correlation coefficient with latitude afterincluding the sign of bN A O was insignificant (correlationcoefficient = 20153 n = 40 and correlation coefficient =0038 n = 29 p 034 in the great tit and pied fly-catcher respectively)
Proc R Soc Lond B (2003)
These effects of the NAO were associated with latitudi-nal gradients in the effects of environmental stochasticityon the population dynamics In both the great tit and piedflycatcher s 2
N A O increased with latitude (figure 3ab corre-lation coefficient = 0439 p = 0005 n = 40 and corre-lation coefficient = 0487 p = 0007 n = 29 in the great titand pied flycatcher respectively) In the pied flycatcherthe residual component remaining after accounting for theNAO s2
re s increased with latitude (figure 3d correlationcoefficient = 0454 p = 0013 n = 29) resulting in a lati-tudinal gradient in environmental stochasticity in thisspecies (figure 3 f correlation coefficient = 0478p = 0009 n = 29) By contrast s 2
re s and s 2e were inde-
pendent of latitude in the great tit (figure 3ce correlationcoefficient = 20003 and correlation coefficient = 0114p 048 n = 40 for s2
re s and s2e respectively) Conse-
quently the relative contribution of variation in the NAOto s 2
e of the great tit increased with latitude (figure 3gcorrelation coefficient = 0334 p = 0035 n = 40)whereas no such significant relationship was present in thepied flycatcher (figure 3h correlation coefficient = 20179 p 035 n = 29) As a consequence the contri-bution of the NAO to the variance in the quasi-stationarydistribution of population sizes (equation (31)) increasedwith latitude in the great tit (correlationcoefficient = 0326 p = 0034 n = 40) Finally the vari-ance in the population size explained by variation in theNAO increased in both species with the residual compo-
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2401
004N
AO
(a)
003
002
001
040 45 50 55 60 65
004 (b)
003
002
001
040 45 50 55 60 65 70 75
020
res
(c)
015
010
005
040 45 50 55 60 65
020 (d )
015
010
005
040 45 50 55 60 65 70 75
025
2
(e)
020
015
010
005
040 45 50 55 60 65
020
( f )
015
010
005
040 45 50 55 60 65 70 75
006
rela
tive
con
trib
utio
nof
NA
O to
e
005
(g)
004
003
002
001
0
40 45 50
latitude (cN)
55 60 65
(h)08
06
04
02
40 45 50
latitude (cN)
55 60 65 70 75
ss
ss
22
2e
NA
Ore
s 2
rela
tive
con
trib
utio
nof
NA
O to
e
ss
ss
22
2e
Figure 3 Latitudinal gradients in the component of the environmental stochasticity explained by (ab) variation in the NAOs 2
NAO (cd) in residual component of the environmental stochasticity s 2res (e f ) in total environmental variance s 2
e and (gh) inproportion of s 2
e explained by variation in the NAO in great tit (aceg) and pied flycatcher (bd fh) populations across Europe
nent of the environmental variance in population size(correlation coefficient = 0475 p = 0002 n = 40 and cor-relation coefficient = 0636 n = 29 p 0001 in the greattit and pied flycatcher respectively) This shows that con-
Proc R Soc Lond B (2003)
tributions of large-scale climate patterns to variation inpopulation size increased with increasing influence ofenvironmental stochasticity on the population fluctu-ations
2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2399
Figure 1 Location of the study areas Triangles represent pied flycatcher study sites squares represent study sites with greattit populations and circles represent study sites where time-series of both species were present Some adjacent study sites arerepresented only by one symbol
to the following generations (Lande et al 2003) The total con-tribution of a female i in year t (Ri) is the number of femaleoffspring born during the year that survive for at least 1 yearplus 1 if the female survives to the next year (Saeligther et al 1998)The demographic variance was estimated (Lande et al 2003)as the weighted mean across years of s 2
d(t) = E(1a21)S (Ri 2 R)2 where R is the mean contribution of the individualsand a is the number of recorded contributions in year tEstimates of s 2
d were obtained for 10 great tit populations andthree pied flycatcher populations (see Appendix A) When noindividual data were available the species-specific mean of s 2
d
was usedThe other parameters in equations (23) were estimated by
maximum likelihood using the full-likelihood function for theprocess obtained by assuming that DX conditioned on X is nor-mally distributed Uncertainties in the estimates were found byparametric bootstrapping simulating new sets of data from thefitted model (Efron amp Tibshirani 1995) To test the hypothesisthat b = 0 we first carried out the estimation for the reducedmodel with b = 0 and performed bootstrap simulations using theparameters estimated from this model and b = 0 The p-valuefor the test is then obtained as the fractions of the simulationsgiving an estimate of b larger than b The results of the testsare presented in Appendix A
The diffusion approximation to the quasi-stationary distri-bution of population sizes
Proc R Soc Lond B (2003)
f (N N0) = G (N N0)Y E `
1
G (N N0)du (31)
can be derived following Lande et al (2003) Here G(N N0) isthe Green function or the sojourn time starting from an initialsize N0 The Green function expresses the expected cumulativetime spent at each population size before extinction (Karlin ampTaylor 1981) We first computed the variance in the quasi-stationary distribution using N 0 = K that can be explained byvariation in the NAO VNAO using s 2
NAO and then repeated thecomputation with s 2
res to get the residual variance Vres The rela-tive contribution to the variance in the quasi-stationary distri-bution of variation in the NAO then becomes VNAO(VNAO 1
Vres) To facilitate the comparison among populations all vari-ances were standardized in relation to K
4 RESULTS
The stochastic population growth rate s (defined byequation (21)) differed significantly between the twospecies (table 1 F = 2127 df = 167 p 0001) with asmaller mean value of s in the pied flycatcher than in thegreat tit populations No significant ( p 01) interspe-cific variation was present in K or in any components ofthe environmental variance s 2
e
2400 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
Table 1 The mean (x plusmn sd) of the estimated population parameters for the two species(s is the stochastic specific growth rate s 2
d the demographic variance s 2e the environmental variance s 2
NAO the component ofs 2
e due to variation in the NAO-index s 2res the residual component not explained by the NAO and n the number of study
populations)
great tit pied flycatcher
x plusmn sd range n x plusmn sd range n
s 089 plusmn 038 010ndash158 40 049 plusmn 031 014ndash089 28K 56 plusmn 49 11ndash216 40 60 plusmn 58 13ndash290 28s2
d 048 plusmn 015 030ndash077 10 040 plusmn 014 031ndash056 3s2
e 0070 plusmn 0049 0002ndash0250 40 0052 plusmn 0047 0010ndash0211 28s2
NAO 0009 plusmn 0013 000ndash0042 40 0006 plusmn 0008 0000ndash0035 28s2
res 0061 plusmn 0046 0002ndash0209 40 0045 plusmn 0041 0007ndash0176 28
012
008
004
0
012
008
004
0
40 45 50 55
latitude (cN)
abs
(
NA
O)
60 65 40 45 50 55
latitude (cN)
60 65 70 75
b
(a) (b)
abs
(
NA
O)
b
Figure 2 Latitudinal gradients in the absolute value of the regression coefficient of the NAO effect bNAO in (a) great tit and(b) pied flycatcher populations across Europe Filled circles denote positive values and open circles denote negative values ofbNAO
Intraspecific variation in s 2e or in its two components
was not significantly ( p 01) related to s in any of thespecies This is in contrast to previous cross-species com-parisons in birds where a close relationship between para-meters describing the deterministic characteristics and theenvironmental stochasticity have been documented(Saeligther amp Engen 2002 Saeligther et al 2002)
We then examined variation in the component of theenvironmental stochasticity due to fluctuations in theNAO Variation in the NAO was able to explain a signifi-cant proportion of the variance in population size in 20(n = 40) of the great tit and in 21 (n = 29) of the piedflycatcher populations A latitudinal gradient was presentin the relative contribution of the NAO to the populationfluctuations of both species because there was an increasein the absolute value of the regression coefficient bN A O
with latitude (figure 2 correlation coefficient = 0463p = 0003 n = 40 and correlation coefficient = 0378p = 0048 n = 29 in the great tit and pied flycatcherrespectively) However the sign of the effects of the NAOdiffered in both species over relatively short distances(figure 2) in some populations a positive relationship waspresent whereas in others N decreased with increasingNAO Hence the correlation coefficient with latitude afterincluding the sign of bN A O was insignificant (correlationcoefficient = 20153 n = 40 and correlation coefficient =0038 n = 29 p 034 in the great tit and pied fly-catcher respectively)
Proc R Soc Lond B (2003)
These effects of the NAO were associated with latitudi-nal gradients in the effects of environmental stochasticityon the population dynamics In both the great tit and piedflycatcher s 2
N A O increased with latitude (figure 3ab corre-lation coefficient = 0439 p = 0005 n = 40 and corre-lation coefficient = 0487 p = 0007 n = 29 in the great titand pied flycatcher respectively) In the pied flycatcherthe residual component remaining after accounting for theNAO s2
re s increased with latitude (figure 3d correlationcoefficient = 0454 p = 0013 n = 29) resulting in a lati-tudinal gradient in environmental stochasticity in thisspecies (figure 3 f correlation coefficient = 0478p = 0009 n = 29) By contrast s 2
re s and s 2e were inde-
pendent of latitude in the great tit (figure 3ce correlationcoefficient = 20003 and correlation coefficient = 0114p 048 n = 40 for s2
re s and s2e respectively) Conse-
quently the relative contribution of variation in the NAOto s 2
e of the great tit increased with latitude (figure 3gcorrelation coefficient = 0334 p = 0035 n = 40)whereas no such significant relationship was present in thepied flycatcher (figure 3h correlation coefficient = 20179 p 035 n = 29) As a consequence the contri-bution of the NAO to the variance in the quasi-stationarydistribution of population sizes (equation (31)) increasedwith latitude in the great tit (correlationcoefficient = 0326 p = 0034 n = 40) Finally the vari-ance in the population size explained by variation in theNAO increased in both species with the residual compo-
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2401
004N
AO
(a)
003
002
001
040 45 50 55 60 65
004 (b)
003
002
001
040 45 50 55 60 65 70 75
020
res
(c)
015
010
005
040 45 50 55 60 65
020 (d )
015
010
005
040 45 50 55 60 65 70 75
025
2
(e)
020
015
010
005
040 45 50 55 60 65
020
( f )
015
010
005
040 45 50 55 60 65 70 75
006
rela
tive
con
trib
utio
nof
NA
O to
e
005
(g)
004
003
002
001
0
40 45 50
latitude (cN)
55 60 65
(h)08
06
04
02
40 45 50
latitude (cN)
55 60 65 70 75
ss
ss
22
2e
NA
Ore
s 2
rela
tive
con
trib
utio
nof
NA
O to
e
ss
ss
22
2e
Figure 3 Latitudinal gradients in the component of the environmental stochasticity explained by (ab) variation in the NAOs 2
NAO (cd) in residual component of the environmental stochasticity s 2res (e f ) in total environmental variance s 2
e and (gh) inproportion of s 2
e explained by variation in the NAO in great tit (aceg) and pied flycatcher (bd fh) populations across Europe
nent of the environmental variance in population size(correlation coefficient = 0475 p = 0002 n = 40 and cor-relation coefficient = 0636 n = 29 p 0001 in the greattit and pied flycatcher respectively) This shows that con-
Proc R Soc Lond B (2003)
tributions of large-scale climate patterns to variation inpopulation size increased with increasing influence ofenvironmental stochasticity on the population fluctu-ations
2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs
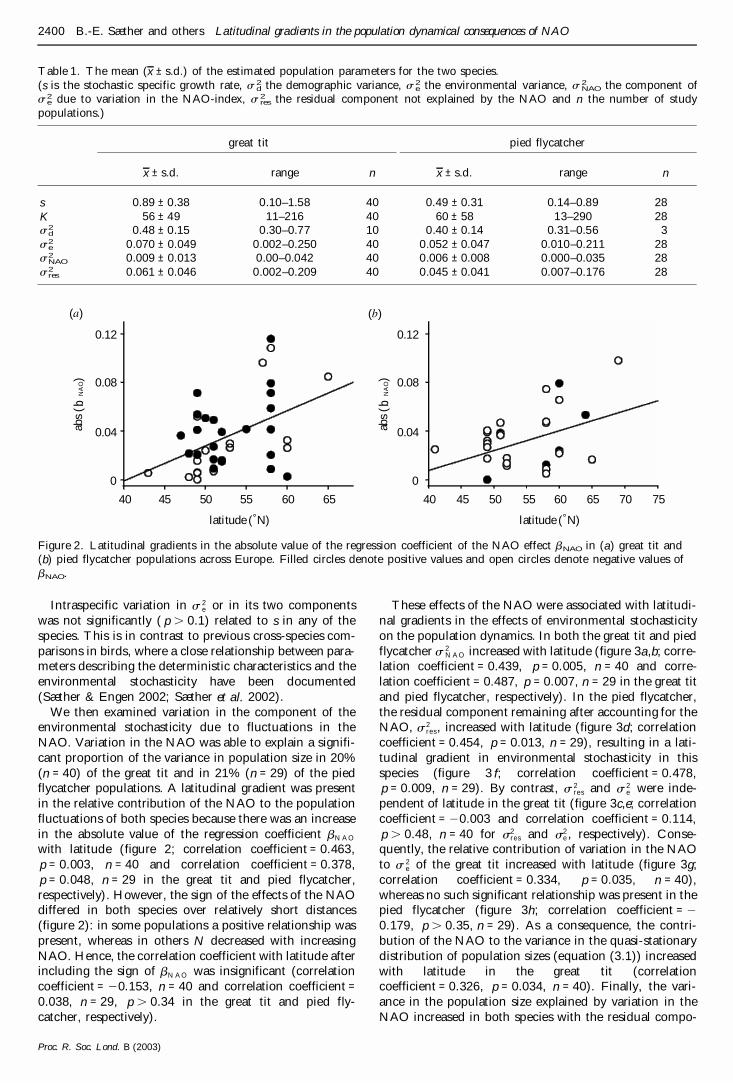
2400 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
Table 1 The mean (x plusmn sd) of the estimated population parameters for the two species(s is the stochastic specific growth rate s 2
d the demographic variance s 2e the environmental variance s 2
NAO the component ofs 2
e due to variation in the NAO-index s 2res the residual component not explained by the NAO and n the number of study
populations)
great tit pied flycatcher
x plusmn sd range n x plusmn sd range n
s 089 plusmn 038 010ndash158 40 049 plusmn 031 014ndash089 28K 56 plusmn 49 11ndash216 40 60 plusmn 58 13ndash290 28s2
d 048 plusmn 015 030ndash077 10 040 plusmn 014 031ndash056 3s2
e 0070 plusmn 0049 0002ndash0250 40 0052 plusmn 0047 0010ndash0211 28s2
NAO 0009 plusmn 0013 000ndash0042 40 0006 plusmn 0008 0000ndash0035 28s2
res 0061 plusmn 0046 0002ndash0209 40 0045 plusmn 0041 0007ndash0176 28
012
008
004
0
012
008
004
0
40 45 50 55
latitude (cN)
abs
(
NA
O)
60 65 40 45 50 55
latitude (cN)
60 65 70 75
b
(a) (b)
abs
(
NA
O)
b
Figure 2 Latitudinal gradients in the absolute value of the regression coefficient of the NAO effect bNAO in (a) great tit and(b) pied flycatcher populations across Europe Filled circles denote positive values and open circles denote negative values ofbNAO
Intraspecific variation in s 2e or in its two components
was not significantly ( p 01) related to s in any of thespecies This is in contrast to previous cross-species com-parisons in birds where a close relationship between para-meters describing the deterministic characteristics and theenvironmental stochasticity have been documented(Saeligther amp Engen 2002 Saeligther et al 2002)
We then examined variation in the component of theenvironmental stochasticity due to fluctuations in theNAO Variation in the NAO was able to explain a signifi-cant proportion of the variance in population size in 20(n = 40) of the great tit and in 21 (n = 29) of the piedflycatcher populations A latitudinal gradient was presentin the relative contribution of the NAO to the populationfluctuations of both species because there was an increasein the absolute value of the regression coefficient bN A O
with latitude (figure 2 correlation coefficient = 0463p = 0003 n = 40 and correlation coefficient = 0378p = 0048 n = 29 in the great tit and pied flycatcherrespectively) However the sign of the effects of the NAOdiffered in both species over relatively short distances(figure 2) in some populations a positive relationship waspresent whereas in others N decreased with increasingNAO Hence the correlation coefficient with latitude afterincluding the sign of bN A O was insignificant (correlationcoefficient = 20153 n = 40 and correlation coefficient =0038 n = 29 p 034 in the great tit and pied fly-catcher respectively)
Proc R Soc Lond B (2003)
These effects of the NAO were associated with latitudi-nal gradients in the effects of environmental stochasticityon the population dynamics In both the great tit and piedflycatcher s 2
N A O increased with latitude (figure 3ab corre-lation coefficient = 0439 p = 0005 n = 40 and corre-lation coefficient = 0487 p = 0007 n = 29 in the great titand pied flycatcher respectively) In the pied flycatcherthe residual component remaining after accounting for theNAO s2
re s increased with latitude (figure 3d correlationcoefficient = 0454 p = 0013 n = 29) resulting in a lati-tudinal gradient in environmental stochasticity in thisspecies (figure 3 f correlation coefficient = 0478p = 0009 n = 29) By contrast s 2
re s and s 2e were inde-
pendent of latitude in the great tit (figure 3ce correlationcoefficient = 20003 and correlation coefficient = 0114p 048 n = 40 for s2
re s and s2e respectively) Conse-
quently the relative contribution of variation in the NAOto s 2
e of the great tit increased with latitude (figure 3gcorrelation coefficient = 0334 p = 0035 n = 40)whereas no such significant relationship was present in thepied flycatcher (figure 3h correlation coefficient = 20179 p 035 n = 29) As a consequence the contri-bution of the NAO to the variance in the quasi-stationarydistribution of population sizes (equation (31)) increasedwith latitude in the great tit (correlationcoefficient = 0326 p = 0034 n = 40) Finally the vari-ance in the population size explained by variation in theNAO increased in both species with the residual compo-
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2401
004N
AO
(a)
003
002
001
040 45 50 55 60 65
004 (b)
003
002
001
040 45 50 55 60 65 70 75
020
res
(c)
015
010
005
040 45 50 55 60 65
020 (d )
015
010
005
040 45 50 55 60 65 70 75
025
2
(e)
020
015
010
005
040 45 50 55 60 65
020
( f )
015
010
005
040 45 50 55 60 65 70 75
006
rela
tive
con
trib
utio
nof
NA
O to
e
005
(g)
004
003
002
001
0
40 45 50
latitude (cN)
55 60 65
(h)08
06
04
02
40 45 50
latitude (cN)
55 60 65 70 75
ss
ss
22
2e
NA
Ore
s 2
rela
tive
con
trib
utio
nof
NA
O to
e
ss
ss
22
2e
Figure 3 Latitudinal gradients in the component of the environmental stochasticity explained by (ab) variation in the NAOs 2
NAO (cd) in residual component of the environmental stochasticity s 2res (e f ) in total environmental variance s 2
e and (gh) inproportion of s 2
e explained by variation in the NAO in great tit (aceg) and pied flycatcher (bd fh) populations across Europe
nent of the environmental variance in population size(correlation coefficient = 0475 p = 0002 n = 40 and cor-relation coefficient = 0636 n = 29 p 0001 in the greattit and pied flycatcher respectively) This shows that con-
Proc R Soc Lond B (2003)
tributions of large-scale climate patterns to variation inpopulation size increased with increasing influence ofenvironmental stochasticity on the population fluctu-ations
2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs

Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2401
004N
AO
(a)
003
002
001
040 45 50 55 60 65
004 (b)
003
002
001
040 45 50 55 60 65 70 75
020
res
(c)
015
010
005
040 45 50 55 60 65
020 (d )
015
010
005
040 45 50 55 60 65 70 75
025
2
(e)
020
015
010
005
040 45 50 55 60 65
020
( f )
015
010
005
040 45 50 55 60 65 70 75
006
rela
tive
con
trib
utio
nof
NA
O to
e
005
(g)
004
003
002
001
0
40 45 50
latitude (cN)
55 60 65
(h)08
06
04
02
40 45 50
latitude (cN)
55 60 65 70 75
ss
ss
22
2e
NA
Ore
s 2
rela
tive
con
trib
utio
nof
NA
O to
e
ss
ss
22
2e
Figure 3 Latitudinal gradients in the component of the environmental stochasticity explained by (ab) variation in the NAOs 2
NAO (cd) in residual component of the environmental stochasticity s 2res (e f ) in total environmental variance s 2
e and (gh) inproportion of s 2
e explained by variation in the NAO in great tit (aceg) and pied flycatcher (bd fh) populations across Europe
nent of the environmental variance in population size(correlation coefficient = 0475 p = 0002 n = 40 and cor-relation coefficient = 0636 n = 29 p 0001 in the greattit and pied flycatcher respectively) This shows that con-
Proc R Soc Lond B (2003)
tributions of large-scale climate patterns to variation inpopulation size increased with increasing influence ofenvironmental stochasticity on the population fluctu-ations
2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs

2402 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
5 DISCUSSION
Our results show that in both the great tit and pied fly-catcher the NAO affects the population dynamics (figure 2ab)and that this influence increases with latitude (figure 3ab)However the regional variation in the contribution of theNAO to the population fluctuations differed between the twospecies In the great tit the proportion of the annual variationin population change explained by the NAO increased withlatitude (figure 3g) By contrast in the pied flycatcher boths 2
N AO and s 2res were large at high latitudes (figure 3bd)
resulting in a latitudinal increase in the stochastic componentof the population dynamics (figure 3e)
The European pied flycatcher is a trans-Saharanmigrant that winters in western Africa (Wernham et al2002) whereas the great tit is a stationary species overmost of its breeding range In spite of these differencesin migratory behaviour a large-scale climate phenomenonacting during winter affected the population dynamics ofboth species One reason for this may be that winteringconditions for the pied flycatcher in West Africa arerelated to variation in the NAO Some evidence does infact suggest a tele-connection between several large-scaleclimate patterns (Hoerling et al 2001) However even insedentary tit species it has been difficult to relate fluctu-ations in population size to winter climate (Slagsvold1975a) Another explanation is that the NAO affects localwinter climate which in turn influences the phenologicaldevelopment of the environment Thus several repro-ductive traits of both pied flycatcher and great tit areinfluenced by annual variation in the phenological devel-opment of the environment (Slagsvold 1975b 1976)Accordingly it has been suggested that the effects ofspring warming will be greater in populations experiencinga short period of peak in the food supply for the nestlingsthan in the habitats with greater variance around the meandate of peak food supply (Visser et al 2003)
Although the contribution of the NAO to the fluctu-ations in population size increased with latitude (figure 3)the sign of the regression coefficient of change in popu-
APPENDIX A
The estimates of the parameters characterizing the population dynamics of the great tit (GT) and the pied flycatcher(PF) at the different study sites
(For locations see figure 1 GER Germany SE Sweden NO Norway BE Belgium HU Hungary UK UnitedKingdom NL The Netherlands EST Estonia FI Finland DK Denmark FR France ES Spain)
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Allmenwald GE GT 49 19 0801 51 mdash 002 2 acute 1027 002Allmenwald GE PF 49 20 0764 47 mdash 0045 0005 005Amarnas SE PF 65 57 0264 141 mdash 0036 0001 0038Amseldell GER GT 49 21 1262 22 mdash 012 0006 0126Amseldell GER PF 49 21 0319 13 mdash 0049 0003 0052Antonihof GER GT 49 21 132 27 mdash 0064 0015 0079Averoslashya NO PF 60 6 0224 24 031 0083 0019 0102Averoslashya NO GT 60 6 081 13 035 0022 0005 0027Bad Kreuznach GER GT 49 50 059 27 mdash 0134 2 acute 1024 0134Bad Kreuznach GER PF 49 50 0616 20 mdash 0074 2 acute 1029 0074Boswachter BE GT 51 16 0817 49 041 0075 0003 0079Budapest HU GT 47 32 0429 35 mdash 0067 0005 0072
(Continued)
Proc R Soc Lond B (2003)
lation size on the NAO differed over relatively short dis-tances at the same latitude (figure 2) This suggests thatmicrogeographical variation in how large-scale climatephenomena are related to fluctuations in local weathermay generate differences in local population dynamicsover short distances Accordingly the relationshipbetween the NAO and different reproductive traits showregional variation in both species (Sanz 2002 2003)Similarly Mysterud et al (2000) showed that the relation-ship between snow depth in March that affected the sexratio in red deer (Cervus elaphus L) and the NAO differedover short distances dependent on local topographicalconditions
Cross-species comparisons of avian population dynam-ics have shown that the effects of environmental stochas-ticity on the population fluctuations are related toparameters (eg specific growth rate form of densitydependence) characterizing the deterministic componentof the population dynamics (Saeligther amp Engen 2002Saeligther et al 2002) No such relationship was howeverpresent among populations of either the great tit or thepied flycatcher One reason for this may be that largeuncertainties in r (table 1) mask any such relationshipbecause of the relatively short time-series used in theanalysis
Modelling has shown large regional variation in themagnitude of the expected changes in climate that areexpected in the future (Houghton et al 2001) Some ofthe largest changes are expected to occur at high latitudesOur analyses (figures 2 and 3) suggest a particularly stronginfluence of climate on the population dynamics of greattit and pied flycatcher in those areas likely to be moststrongly affected by an expected climate change
We are grateful to Erik Borgstrom Hans Kallander ErikNyholm Per Gustav Thingstad and Wolfgang Winkel for pro-viding unpublished data and Ivor Herfindal for producing themap The study was financed by grants from the EuropeanCommission (project METABIRD) and the Research Councilof Norway (Klimaeffekt-programmet) This is publication3208 from N100-KNAW
Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs

Latitudinal gradients in the population dynamical consequences of NAO B-E Saeligther and others 2403
locality species degN 9N s K s2d s2
re s s2N A O s 2
e
Buhlbronn GER GT 48 51 0801 132 mdash 0029 0002 0031Finsterbrunnertal GER GT 49 21 0715 24 mdash 0116 0001 0118Forest of Dean UK PF 51 40 0264 62 mdash 0011 0004 0016Frankfurt GER GT 50 21 1168 183 mdash 0042 0007 0049Ghent BE GT 51 0 0567 36 048 0048 5 acute 1024 0048Harthausen GER GT 49 18 104 85 mdash 0007 1024 0007Harthausen GER PF 49 18 044 60 mdash 0031 0001 0032Hemsedal NO PF 60 52 0138 30 mdash 0007 0003 001Hoge Veleuwe NL GT 52 5 0937 116 039 0117 0001 0118Hoge Veleuwe NL PF 52 5 0399 90 033 0027 0002 0028Hollenbach GER GT 49 44 0599 101 mdash 0018 001 0028Illu EST GT 58 31 0565 15 mdash 0071 0012 0083Illu EST PF 58 31 0257 110 mdash 0017 5 acute 1024 0017Kilingi EST GT 58 9 1152 97 mdash 0028 0001 003Kilksama EST GT 58 28 1571 13 mdash 0032 0022 0054Kilksama EST PF 58 28 1448 21 mdash 0041 0009 005Kilpisjarvi FI PF 69 3 0592 30 mdash 0176 0035 0211Kunnametsa EST GT 58 35 1345 11 mdash 0017 3 acute 1024 0017Kunnametsa EST PF 58 35 063 42 mdash 003 1024 003Kaas DK GT 57 17 0152 64 mdash 0052 0041 0093La Hiruela ES PF 41 4 0560 75 056 0004 0004 0007Langdell GER GT 49 20 0827 18 mdash 0118 0002 012Langdell GER PF 49 20 0256 23 mdash 006 0007 0067Lauvsjoslashen NO PF 64 20 022 17 mdash 0138 0016 0154Leipzig GER PF 51 17 0863 50 mdash 0027 0005 0032Lemsjoslashholm FI GT 60 5 1579 19 mdash 0124 0002 0126Lemsjoslashholm FI PF 60 5 0886 39 mdash 0013 0002 0015Liesbos NL GT 51 35 1043 40 0345 0064 3 acute 1024 0065Lingen GER GT 52 3 0795 99 mdash 0108 0001 0109Lingen GER PF 52 3 0408 92 mdash 004 6 acute 1024 004Linnebejer SE GT 55 44 1212 47 0593 0104 001 0114Matapere EST GT 58 20 1036 31 mdash 0025 0006 0031Matapere EST PF 58 20 0596 73 mdash 0022 4 acute 1024 0022Neubruch GE PF 49 20 0513 13 mdash 001 0006 0016Niidu EST GT 58 23 1447 67 mdash 0026 0018 0044Niidu EST PF 58 23 0093 114 mdash 0014 0008 0022Osterhout NL GT 51 55 0476 29 0634 0094 0013 0107Oti EST PF 58 28 0345 27 mdash 0083 2 acute 1024 0083Oulu FI GT 65 0 0963 23 mdash 009 0033 0123Pfullingen GER GT 48 26 1183 51 mdash 0003 2 acute 1025 0003Pillnitz GER PF 51 1 0118 14 mdash 0005 0009 0014Riitskulli EST GT 58 10 1686 14 mdash 0 0051 0051Riitskulli EST PF 58 10 0973 115 mdash 001 6 acute 1024 0011Rothense GE GT 49 20 0955 14 mdash 002 0001 0021Rothense GE PF 49 20 0586 18 mdash 0059 0004 0063Rouviere FR GT 43 40 0767 35 mdash 0068 2 acute 1024 0068RaEcirc de SE GT 60 0 0994 91 mdash 0016 4 acute 1025 0016Sanga EST GT 58 24 01 23 mdash 0209 0042 025Sanga EST PF 58 24 0727 53 mdash 0104 0021 0125Simmertal GER GT 49 49 0819 35 mdash 0029 003 0059Siuntio FI PF 60 15 0425 62 mdash 003 0015 0045Theresienstein GER GT 50 16 1012 16 mdash 0059 0003 0062Tresswell UK GT 53 18 0493 35 0300 002 0003 0024Vlieland NL GT 53 15 0153 134 0769 0084 0005 0089Wolfsburg GER GT 52 27 0651 100 mdash 0019 0008 0027Wolfsburg GER PF 52 27 0313 290 mdash 003 9 acute 1024 0031Wytham Wood UK GT 51 46 0719 216 0565 007 0002 0072
Proc R Soc Lond B (2003)
2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs

2404 B-E Saeligther and others Latitudinal gradients in the population dynamical consequences of NAO
REFERENCES
Andrewartha H G amp Birch L C 1954 The distribution andabundance of animals University of Chicago Press
Both C amp Visser M E 2001 Adjustment to climate changeis constrained by arrival data in a long-distance migrant birdNature 411 296ndash298
Efron B amp Tibshirani R J 1995 An introduction to the boot-strap New York Chapman amp Hall
Engen S Bakke Oslash amp Islam A 1998 Demographic andenvironmental stochasticity concepts and definitions Bio-metrics 54 840ndash846
Forchammer M C Post E amp Stenseth N C 1998 Breed-ing phenology and climate Nature 391 29ndash30
Hoerling M P Hurrell J W amp Xu T Y 2001 Tropical ori-gins for recent North Atlantic climate change Science 29290ndash92
Houghton J T Ding Y Griggs D J Noguer M Van derLinden P J amp Xiasou D 2001 Climate change 2001 Cam-bridge University Press
Hurrell J W 1995 Decadal trends in the North Atlantic Oscil-lation regional temperatures and precipitation Science 269676ndash679
Karlin S amp Taylor H M 1981 A second course in stochasticprocesses New York Academic
Lack D 1954 The natural regulation of animal numbers OxfordUniversity Press
Lande R 1998 Demographic stochasticity and Allee effect ona scale with isotrophic noise Oikos 83 353ndash358
Lande R Engen S amp Saeligther B-E 2003 Stochastic popu-lation dynamics in ecology and conservation Oxford Univer-sity Press
McCarty J P 2001 Ecological consequences of recent climatechange Conserv Biol 15 320ndash331
McCleery R H amp Perrins C M 1998 Temperature and egg-laying trends Nature 391 30ndash31
May R M 1976 Simple mathematical models with very com-plicated dynamics Nature 261 459ndash467
May R M 1981 Models for single populations In Theoreticalecology 2nd edn (ed R M May) pp 5ndash29 LondonBlackwell Science
Mysterud A Yoccoz N G Stenseth N C amp Langvatn R2000 Relationships between sex ratio climate and densityin red deer the importance of spatial scale J Anim Ecol69 959ndash974
Mysterud A Stenseth N C Yoccoz N G Ottersen G ampLangvatn R 2002 The response of terrestrial ecosystems toclimate variability associated with the North Atlantic Oscil-lation In The North Atlantic Oscillation climatic significanceand environmental impact Geographical monographs series vol132 (ed J W Hurrell Y Kushnir G Ottersen amp MVisbeck) pp 235ndash262 Washington DC American Geo-physical Union
Pimm S L 1991 The balance of nature University ofChicago Press
Saeligther B-E amp Engen S 2002 Pattern of variation in avianpopulation growth rates Phil Trans R Soc Lond B 3571185ndash1196 (DOI 101098rstb20021119)
Proc R Soc Lond B (2003)
Saeligther B-E Engen S Islam A McCleery R H amp Per-rins C 1998 Environmental stochasticity and extinction riskin a population of a small songbird the great tit Am Nat151 441ndash450
Saeligther B-E Tufto J Engen S Jerstad K RoslashstadO W amp SkaEcirc tan J E 2000 Population dynamical conse-quences of climate change for a small temperate songbirdScience 287 854ndash856
Saeligther B-E Engen S amp Matthysen E 2002 Demographiccharacteristics and population dynamical patterns of solitarybirds Science 295 2070ndash2073
Sanz J J 2002 Climate change and breeding parameters ofgreat and blue tits throughout the western Palearctic GlobalChange Biol 8 1ndash14
Sanz J J 2003 Large-scale effect of climate change on breed-ing parameters of pied flycatchers in Western EuropeEcography 26 45ndash50
Sanz J J Potti J Moreno J Merino S amp Frotilde as O 2003Climate change and fitness components of a migratory birdbreeding in the Mediterranean region Global Change Biol9 1ndash12
Slagsvold T 1975a Critical period for regulation of great tit(Parus major L) and blue tit (Parus caeruleus L) populationsNorw J Zool 23 67ndash88
Slagsvold T 1975b Competition between the great tit Parusmajor and the pied flycatcher Ficedula hypoleuca in the breed-ing season Ornis Scand 6 179ndash190
Slagsvold T 1976 Annual and geographical variation in thetime of breeding of the great tit Parus major and the piedflycatcher Ficedula hypoleuca in relation to environmentalphenology and spring temperature Ornis Scand 7 125ndash145
Stenseth N C Mysterud A Ottersen G Hurrell J WChan K-S amp Lima M 2002 Ecological effects of climatefluctuations Science 297 1292ndash1296
Turchin P W 1995 Population regulation old arguments anda new synthesis In Population dynamics new approaches andsynthesis (ed N Cappucino amp P W Price) pp 19ndash40 SanDiego CA Academic
Visbeck M H Hurrell J W Polvani L amp Cullen H M2001 The North Atlantic Oscillation past present andfuture Proc Natl Acad Sci USA 98 12 876ndash12 877
Visser M E (and 13 others) 2003 Variable responses to large-scale climate change in European Parus populations Proc RSoc Lond B 270 367ndash372 DOI 101098rspb20022244)
Walther G R Post E Convey P Menzel A ParmesanC Beebee T J C Fromentin J M Hoegh-GuldbergO amp Bairlein F 2002 Ecological responses to recent climatechange Nature 416 389ndash395
Wernham C Toms M Marchant J Clark J SiriwardenaG amp Baillie S 2002 Migration atlas movements of the birdsof Britain and Ireland London T amp AD Poyser
Winkel W amp Hudde H 1997 Long-term trends in repro-ductive traits of tits (Parus major P caeruleus) and pied fly-catchers Ficedula hypoleuca J Avian Biol 28 187ndash190
As this paper exceeds the maximum length normally permitted theauthors have agreed to contribute to production costs
Related Documents











